
Submitted:
27 January 2025
Posted:
28 January 2025
You are already at the latest version
Abstract
Background and Objective: Mammea siamensis (MS) is a Thai herb used in traditional medicine. Previous studies have reported the antiproliferative effects of its constituents in various cancer cell lines. However, the effects of MS extract on cytotoxicity and mo-lecular mechanisms of apoptosis induction in HCT116 colon cancer cells have not been fully explored. Methods and Results: The cytotoxic effect of MS extract on HCT116 cells was assessed using the MTT assay. MS extract increased cytotoxicity in a concentra-tion-dependent manner. It also induced nuclear morphological changes and disrupted the mitochondrial membrane potential (ΔΨm), as assessed by Hoechst 33342 and JC-1 staining, respectively. These findings indicated that MS extract induced apoptosis, which was further confirmed by flow cytometry showing an increase in the sub-G1 phase. To investigate the expression of signaling proteins, western blot analysis was conducted. The results showed that MS extract activated caspase activity (caspase-8, -9, and -7) and inhibited PARP activity. Additionally, MS extract upregulated pro-apoptotic proteins (tBid, Bak, and cytochrome c) while downregulating anti-apoptotic proteins (Bcl-2 and Bcl-xL). Mechanistic studies revealed that MS extract activated MAPK pathways while inactivating the PI3K/Akt/NF-κB and GSK-3β/β-catenin pathways. Notably, MS extract also inhibited V-ATPases, as evaluated by acridine orange staining and western blot analysis. Conclusions: Our findings suggest that MS extract induces apoptosis via the activation of both intrinsic and extrinsic pathways associated with the key signaling pathways. Therefore, MS extract shows potential as a therapeutic agent for colon cancer.
Keywords:
Mammea siamensis
; apoptosis
; colon cancer
; MAPK
; PI3K/Akt/ NF-κB
; GSK-3β/β-catenin
; V-ATPases
1. Introduction
Colon cancer (CRC) is the second leading cause of cancer-related mortality and the third most common type of cancer globally. Recent statistics indicate that approximately 1.9 million people were diagnosed with CRC in 2020 [1]. Most diagnoses occur in individuals over the age of 50, making CRC a significant public health concern in many countries worldwide [2]. Various cancer treatments, including surgery, chemotherapy, targeted therapy, immunotherapy, and radiation, are available. However, these treatments are often prohibitively expensive for low-income patients and can also cause side effects to healthy cells in the body [3,4]. Therefore, there is an urgent need to search for new agents for cancer treatment.
Mammea siamensis (Miq.) T. Anders, known locally in Thai as Sarapi or Saraphi, belongs to the Calophyllaceae family. M. siamensis is a Thai herb used in traditional medicine, such as a heart tonic, to reduce fever and to increase appetite [5]. Previous chemical studies of the flowers of M. siamensis have reported the isolation of various bioactive compounds, including several coumarins, xanthone, flavonoids, and other such bioactive compounds [5,6]. This plant and its constituents reportedly possess anti-malarial activity [7,8], antioxidant activity [9], anti-microbial activities [10], and anti-proliferative [6]. It has shown anti-proliferative effects against a variety of cancer cells, such as human gastric carcinoma cells (HSC-2, HSC-4, and MKN-45) [5] and human prostate carcinoma cells (LNCaP) [6], including colon cancer cells (HCT116) [11]. Kayeassamin A was isolated from the M. siamensis flower has been shown to have anti-cancer activity in human leukemia HL-60 cells via apoptosis induction [12]. Interestingly, M. siamensis is utilized both in Ayurveda and by indigenous communities in Northeast Thailand for cancer prevention and healing purposes [11]. The "Poh-Pu" remedy, a Traditional Thai Medicine (TTM) formulation used in cancer treatment, includes M. siamensis as one of its components. This remedy has been prescribed for over thirty years at the Jitmeatta Mercy Foundation for Cancer Patients (JFCT) in Phetchaburi province, Thailand [11,13], which indicates that this plant has shown potential for anti-cancer effects. However, the molecular mechanism of its extract on HCT116 colon cancer cells has not been explained.
Recently, apoptosis inducers have been utilized in cancer treatment to help control or potentially halt the uncontrolled growth of cancer cells, aiming to inhibit their development and progression [14,15,16]. Apoptosis is a programmed cell death that follows two primary pathways: the extrinsic and intrinsic pathways. The intrinsic, or mitochondrial pathway, is triggered by internal signals, such as DNA damage, ischemia, and oxidative stress, while external signals activate the extrinsic pathway. The intrinsic apoptosis pathway is started with cytochrome c releasing from mitochondria, then caspase-9 is activated, which subsequently activates caspases-3 or -7, leading to cell death. Moreover, the Bcl-2 family proteins also play a key role in the intrinsic pathway. Conversely, the extrinsic pathway is associated with ligand binding to death receptors on the cell surface, resulting in the activation of caspase-8. Activated caspase-8 activates caspases-3 or -7, then cleaves Bid (pro-apoptotic protein in the Bcl-2 family) into the truncated form of Bid (tBid). Then, tBid promotes the release of cytochrome c from mitochondria [16,17,18]. The abnormal apoptosis is associated with a variety of diseases, including cancer. Dysregulated apoptosis results in the accumulation of damaged cells, ultimately leading to tumorigenesis, as well as resistance to cancer therapy. Additionally, interference with cell signaling pathways, including the glycogen synthase kinase-3β (GSK-3β)/β-catenin, phosphoinositide 3-kinase (PI3K)/protein kinase B (PKB, Akt), and mitogen-activated protein kinase (MAPK) pathway, play an important role in cancer development and progression [19,20,21].
Vacuolar ATPase (V-ATPase) is an ATP-driven H⁺ pump that transports protons across both intracellular and plasma membranes, regulating the pH levels inside and outside of cells. This regulation is essential for various cellular functions, including cancer cell growth, invasion, metastasis, and proliferation [22]. Recently, V-ATPases have emerged as a promising target for cancer therapy [23]. Research has shown that V-ATPases are overexpressed in various cancers, where they play a role in promoting cancer cell proliferation and metastasis [24,25]. Notably, inhibiting V-ATPase activity has been found to trigger apoptosis [23], suggesting that targeting V-ATPases could be a valuable strategy for developing new anti-cancer treatments.
Therefore, this study explored the impact of MS extract on cytotoxicity and apoptosis induction, as well as the molecular mechanisms involved in HCT116 human colon cancer cells. The findings provide new insights into the potential of MS extract as an anticancer agent for colon cancer and its possible therapeutic applications and its possible use in synergy with cancer treatments in the future.
2. Results
2.1. 1H NMR Analysis of MS Methanolic Extract
1H NMR spectroscopic analysis of the MS methanolic extract revealed that the extract contained two C-glycosyl flavones, as shown in Figure 1. The structure of these compounds in the extract was elucidated by analysis of their spectroscopic data and by comparison with literature values [26,27]. The presence of two singlets at 13.17 and 13.56 ppm, characteristic signals of 5-OH proton of flavonoid skeleton, were observed. Two singlets at 6.29 and 6.54 were ascribed to H-6 and H-8a on ring A. Four doublets with ortho-coupling (J =10 Hz) at 6.90, 6.93, 7.93, and 8.02 ppm were attributed to H-3' and H-5', H-3a' and H-5a', H-2a' and H-6a', and H-2' and H6', respectively, on aromatic ring B which are typical pattern of the asymmetrical p-disubstituted benzene ring. In addition, from gCOSY experiment, the signal at 6.90 was correlated to the signal at 8.02, and the signal at 6.93 was correlated to the signal at 7.93 ppm. Two singlets at 6.77 and 6.78 ppm were ascribed to H-3 and H-3a' on ring C. Therefore, it could be concluded that the NMR spectral data corresponded to those previously reported for the mixture of vitexin and isovitexin detected in M. siamensis T. Anders [27].
2.2. MS Extract Increased Cell Toxicity in HCT116 Cells
The MTT assay was used to evaluate cytotoxicity in HCT116 cells. We treated the cells with various concentrations of MS extract for 24 h. The viability of the cells in response to the treatment decreased in a dose-dependent manner compared with the untreated control, with an IC₅₀ of 79.57±0.25 µg/mL (Figure 3), indicating increased cell toxicity. While an IC50 of 301.3±1.27 µg/mL was observed in normal Vero cells (Figure S1). This result suggests the potential of MS extract on cell proliferation inhibition in HCT116 cells.
2.3. MS Extract Induced Nuclear Morphological Change in HCT116 Cells
Nuclear condensation is a characteristic of nuclear morphological changes in apoptotic cells [17,28], which can be evaluated by Hoechst 33342 staining and visualized under a fluorescence microscope. As illustrated in Figure 4, the number of condensed HCT116 cells and apoptotic bodies (indicated by red arrows) increased with higher doses of MS extract. This suggests that MS extract-induced apoptosis may play a role in the decreased viability of HCT116 cells.
2.4. MS Extract Reduced Mitochondrial Membrane Potential (ΔΨm) in HCT116 Cells
Mitochondrial membrane potential plays a critical role in regulating intrinsic apoptosis, as investigated using JC-1 staining [29]. The findings showed a significant reduction in red fluorescence intensity of JC-1 aggregates in the treated cells compared to the control group, indicating a decrease in ΔΨm (Figure 5). This suggests that the MS extract may induce apoptosis in HCT116 cells via induced mitochondrial dysfunction.
2.5. MS Extract Increased the Population of Sub-G1 Apoptotic HCT116 Cells
Apoptotic cells are identified by their decreased DNA content, which can be measured using flow cytometry through the sub-G1 DNA content [28]. Therefore, HCT116-treated cells were stained with Guava Cell Cycle® reagent, allowing us to measure DNA content by flow cytometry. After 24 hours of treatment with MS extract, a significant increase in the percentage of the sub-G1 population was observed, correlating with increased dose levels (Figure 6).
2.6. MS Extract Stimulated Apoptosis Pathways in HCT116 Cells
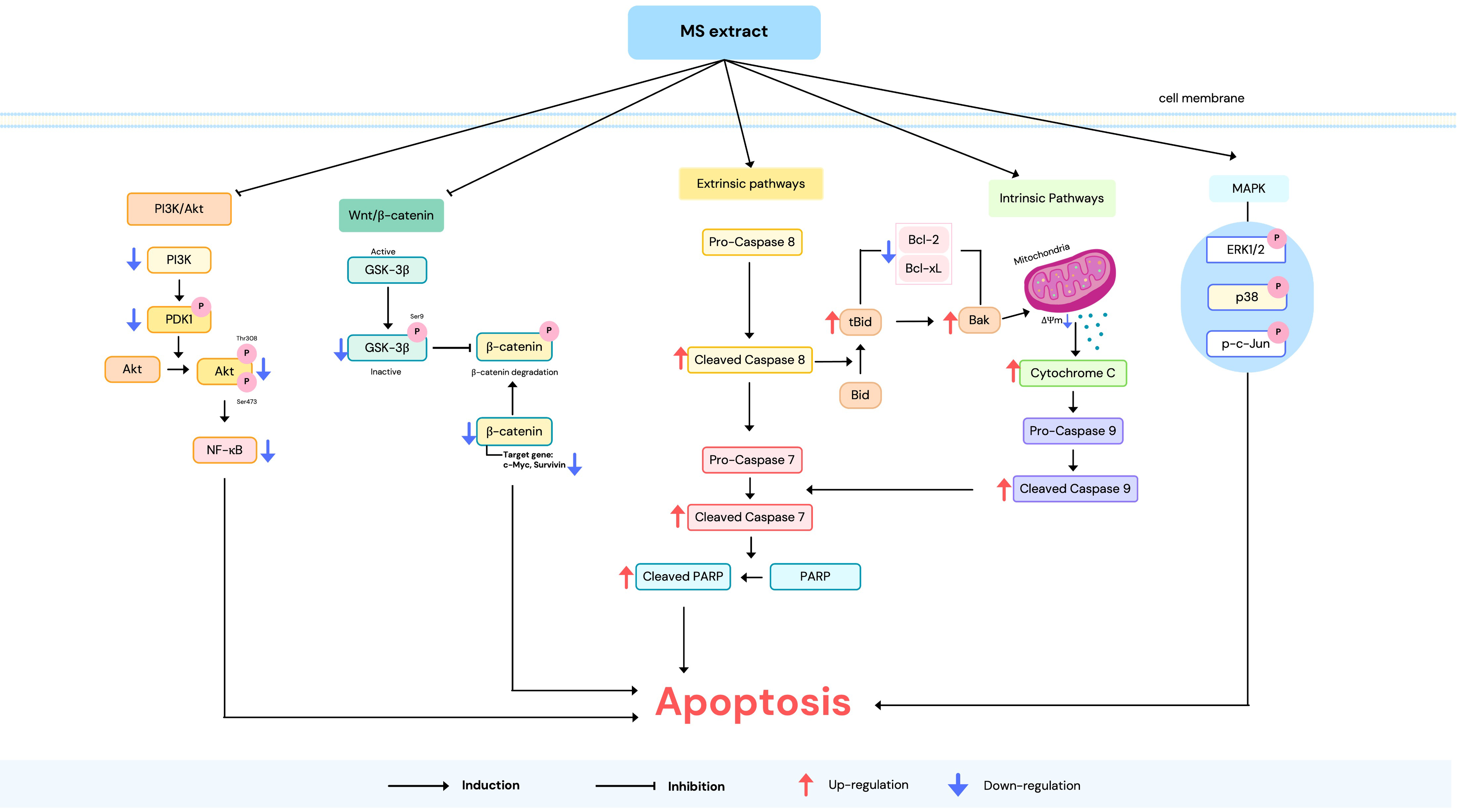
The molecular mechanisms of apoptosis induction are triggered through two main pathways: the extrinsic and intrinsic pathways [17]. Therefore, we investigated protein expression in these pathways to elucidate the molecular mechanisms of the MS extract. As depicted in Figure 7, western blot analysis revealed that the expression of the anti-apoptotic protein Bcl-2 decreased in comparison to the pro-apoptotic Bax protein, as well as cytochrome c, which was enhanced. The expression of cleaved caspase-9 and caspase-7, as well as cleaved PARP, also increased following MS extract treatment. These results suggest the activation of intrinsic pathways. Additionally, the MS extract increased the levels of cleaved caspase-8, indicating that the extrinsic pathways are involved. This resulted in the increased truncation of Bid (tBid) in the MS-treated cells, suggesting that there is a cross-talk between the extrinsic and intrinsic pathways.
2.7. MS Extract Inhibited V-ATPase Activity in HCT116 Cells
Previous studies have reported that inhibitors of V-ATPases can induce apoptosis [30]. In our current study, MS extract shows the potential to induce apoptosis. Thus, we employed AO staining and western blotting to explore whether MS extract could inhibit V-ATPase activity in HCT116 cells. The results showed that MS extract can inhibit V-ATPase activity by suppressing the orange or red fluorescence of AO dye compared to the control group (Figure 8A). The results of the Western blot showed that the protein level of the B2 subunit of H+-ATPase (ATP6V1B2) was decreased (Figure 8B). These results are consistent with those obtained using Bafilomycin A1 (BMA), a specific V-ATPase inhibitor. Therefore, these findings demonstrate that MS extract is a potent V-ATPase inhibitor for the treatment of HCT116 cells.
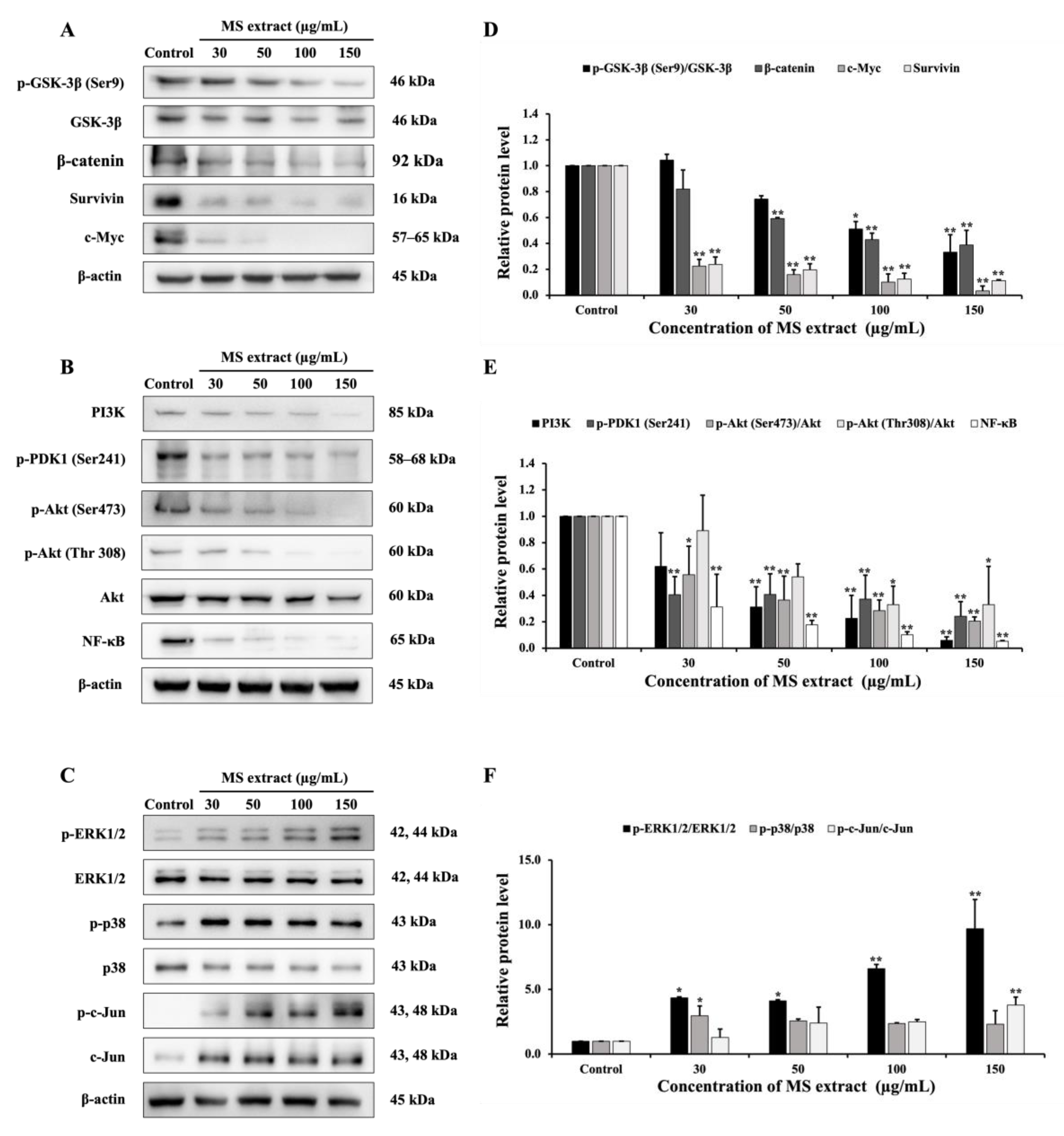
2.8. MS Extract Relegated the GSK-3β/β-catenin, PI3K/Akt/NF-κB, and MAPK Signaling Pathways in HCT116 Cells
There are many cellular signaling pathways involved in cancer. Therefore, a western blot assay was performed to examine the expression of proteins related to the GSK-3β/β-catenin, PI3K/Akt, and MAPK signaling pathways in order to further investigate the mechanisms of the MS extract in HCT116 cells. First, results showed that MS extract down-regulated p-GSK-3β, β-catenin, c-Myc, and survivin (Figure 9A). Next, the expression of PI3K, p-PDK1, p-Akt (Ser473), p-Akt (Thr308), and NF-κB were also down-regulated (Figure 9B). Finally, MS extract up-regulated the phosphorylated forms of ERK (p-ERK), p38 (p-p38), and c-Jun (p-c-Jun), as shown in Figure 9C.
3. Discussion
Herbal plants have been traditionally used in folk medicine across many countries worldwide. Natural extracts from the fruits, seeds, leaves, bark, flowers, and roots of medicinal plants offer numerous benefits, including skin care, food enhancement, and nutritional improvement [31]. In Thailand, the flower of M. siamensis is particularly notable for its use in cancer prevention, treatment, and regeneration in traditional medicine [11,13]. However, the molecular mechanism of M. siamensis extract remains unclear. Several prior studies suggest that M. siamensis and its components demonstrate anti-cancer activity by inhibiting cell proliferation in various cancer cell types. Specific coumarin compounds isolated from M. siamensis flowers have shown anti-proliferative effects in LNCaP prostate cancer cells [6]. Additionally, the human cancer cell lines MDA-MB-231 (breast cancer), U-251 (brain tumor), HCT-116 (colon cancer), and CCRF-CEM (leukemia) have all been significantly affected by the cytotoxic activity of coumarins such as mammea A/AA, deacetylmammea E/BA, and deacetylmammea E/BB, which are extracted from M. siamensis flowers [11]. Overall, these studies suggest that the chemical constituents of M. siamensis play a key role in inhibiting cancer cell proliferation.
In our study, we conducted a chemical analysis of M. siamensis flowers using 1H NMR. As shown in Figure 1, we identified vitexin and isovitexin in the methanolic extract, which may contribute to the cytotoxic effects observed in HCT116 cells. Previous investigations have reported that M. siamensis contains high levels of vitexin and isovitexin, well-known bioactive flavonoid compounds recognized for their anti-cancer and anti-tumor properties [27,32]. Our findings align with these reports, demonstrating that MS extract significantly decreases cell proliferation in HCT116 cells. However, further studies are necessary to fully evaluate the anti-cancer activity of the isolated compounds.
In this investigation, we showed that the MS extract inhibits HCT116 cell proliferation through apoptosis induction. Apoptosis was characterized by typical morphological changes, including cell shrinkage, DNA condensation, fragmentation, and the formation of apoptotic bodies [17]. Our results showed that MS extract induced nuclear condensation, apoptotic bodies, and decreased ΔΨm, indicators of apoptosis, as determined by Hoechst 33342 and JC-1 staining. Additionally, flow cytometric analysis confirmed apoptosis in HCT116 cells by revealing an increased sub-G1 population. These characteristics, along with specific molecular events, are associated with the activation of apoptosis signaling pathways.
We investigated the expression of apoptosis-related proteins to understand the molecular mechanisms through which MS extract triggers apoptosis. The primary regulators of the intrinsic apoptotic process are Bcl-2 family proteins, which play a role in the disruption of ΔΨm [18]. When mitochondrial integrity is compromised, pro-apoptotic proteins such as cytochrome c are released, triggering the activation of caspase proteins. This suggests that MS extract induces apoptosis in HCT116 cells via the intrinsic apoptosis pathway. Additionally, we observed caspase-8 activation, an initiator of the extrinsic apoptosis pathway, and enhanced levels of tBid. These findings indicate potential cross-talk between the intrinsic and extrinsic pathways [16,33]. DNA fragmentation, a key indicator of apoptosis, was evidenced by the cleavage of PARP [34]. The loss of PARP activity prevents DNA repair, allowing endonucleases like CAD (caspase-activated DNase) to fragment DNA [35]. In summary, our findings demonstrate that MS extract induces apoptosis in HCT116 cells through both intrinsic and extrinsic pathways.
Similar results have been reported for mammeasin A and surangin B, compounds containing a geranyl group. These compounds, isolated from the methanol extract of M. siamensis flowers, exhibited strong activity against HSC-4 cells, involving apoptotic cell death [5]. Additionally, Kayeassamin A (KA), isolated from M. siamensis flowers, induces apoptosis in HL-60 human leukemia cells, as evidenced by chromatin condensation, DNA fragmentation, and an increase in sub-G1 phase DNA content. KA also strongly activates caspase-3 and -8, weakly activates caspase-9, and triggers PARP cleavage [12,36].
V-ATPases represent a promising target for cancer treatment due to their minimal impact on normal cells. Inhibiting V-ATPase activity has been associated with growth inhibition and apoptosis induction [22]. In our study, we found that MS extract inhibited V-ATPase activity, similar to bafilomycin A1, a well-known potent V-ATPase inhibitor. Previous research demonstrated that bafilomycin A1 induces apoptosis in pediatric B-cell acute lymphoblastic leukemia by targeting mitochondria and activating a caspase-independent pathway. The anticancer effects of bafilomycin A1 are linked to its inhibition of V-ATPase activity [37]. Furthermore, a study on C. cochinchinense extracts found that they inhibited V-ATPase activity and induced apoptosis in several cancer cell types, including MDA-MB-231, A431, and A375 cells [38]. Therefore, MS extract could be further developed as an anti-cancer drug with V-ATPase inhibitory properties. However, the detailed mechanisms by which V-ATPase inhibition induces apoptosis in HCT116 cells require further investigation. Notably, inhibition of V-ATPase activity by bafilomycin also blocks the Wnt/β-catenin pathway [39]. Consequently, we next investigated the effect of MS extract on the GSK-3β/β-catenin pathway.
The Wnt/β-catenin pathway regulates various cellular functions, including proliferation, differentiation, migration, and apoptosis. It suppresses apoptosis by blocking the release of cytochrome c. In most colorectal cancer (CRC) patients, the Wnt/β-catenin signaling pathway is abnormally activated. This pathway is typically initiated by the binding of TCF/LEF transcription factors with β-catenin, which activates downstream target genes, such as c-Myc and other proto-oncogenes. This promotes cell proliferation and supports tumor development and progression [40]. Consequently, downregulating the Wnt/β-catenin pathway is associated with inhibited tumor cell growth in CRC cell lines [41]. Our data support these findings, suggesting that the MS extract inhibits the growth of HCT116 cells, likely through the downregulation of β-catenin and its downstream targets. A previous study demonstrated that apigenin, a widely known flavonoid, blocked the nuclear translocation of β-catenin, thereby decreasing the expression of Wnt pathway target genes and preventing colorectal cancer cells from proliferating, migrating, invading, and forming organoids [42]. Additionally, the MS extract-treated cells showed a reduction in GSK3β, which is the first known substrate of Akt [43].
Our results also indicate that MS extract decreased the expression of PI3K, p-PDK1, p-Akt (Ser473), and p-Akt (Thr308), suggesting that the PI3K/Akt pathway was inhibited. The PI3K pathway and its downstream effector Akt are key survival proteins and major contributors to chemoresistance in cancer therapy across various tumor types. When Akt is activated, it stimulates PAK1, which then phosphorylates Bad, leading to Bad's release from the mitochondrial membrane into the cytoplasm. At the same time, Akt inhibits the translocation of Bax from the cytoplasm to the mitochondria. As a result, the overactivation of Akt promotes cell survival by enhancing Bcl-2 activity and inhibiting Bax, both of which play crucial roles in cancer cell resistance [44].
Our mechanistic study highlights the anti-cancer properties of vitexin, a flavonoid compound derived from natural sources. It has been reported that vitexin inhibits cell growth and promotes cell apoptosis by inducing G2/M cell cycle arrest, primarily through the suppression of the Akt/mTOR pathway in human glioblastoma cells [45]. Similarly, in human non-small cell lung cancer A549 cells, vitexin induces apoptosis by inhibiting the PI3K/Akt/mTOR pathway [46], which supports the findings of this study. Additionally, numerous protein targets that regulate cell motility, proliferation, and survival are phosphorylated by Akt. According to a previous study, Akt also controls NF-κB transcriptional activity [47]. Our findings show that MS extract reduced the expression of NF-κB, a transcription factor known for its role in preventing apoptosis. A prior study demonstrated that NF-κB inhibition promotes cell apoptosis [48]. Flavonoids, including quercetin, are recognized as inhibitors of NF-κB signaling, with evidence showing that quercetin suppresses NF-κB activity by inhibiting IκBα and p65 phosphorylation, thereby blocking NF-κB nuclear translocation [49]. Moreover, previous research has shown that the dietary phytochemical quercetin causes apoptosis in human cervical carcinoma (HeLa) cells via a mechanism dependent on the suppression of p53 and NF-κB [50].
Furthermore, MAPK pathways control a range of biological functions through various cellular mechanisms. Depending on the type of cell and the specific stimulation, MAPK can act as either activators or inhibitors of these processes, including apoptosis [51]. In hepatocellular carcinoma cell lines, vitexin exerted cytotoxic effects in vitro by inducing apoptosis through the activation of the JNK MAPK pathway, as evidenced by the upregulation of p-JNK levels. The apoptosis induced by vitexin was reduced by co-treatment with the JNK inhibitor SP60012 [52]. Additionally, anthocyanins, a type of flavonoid, induce apoptotic cell death in HCT-116 cells through p38-MAPK activation [53]. These observations suggest that apoptosis induction is linked to the activation of both p38-MAPK and JNK-MAPK pathways. In our study, we demonstrated that MS extract enhanced the expression of p-p38, p-c-Jun (downstream targets of the JNK cascade), and p-ERK1/2. While the activation of p38 and JNK is typically associated with pro-apoptotic effects, ERK activation is generally linked to anti-apoptotic effects. However, there are instances where ERK1/2 signaling can also promote apoptosis [51]. ERK activity can stimulate caspase-8 activation or mitochondrial cytochrome c release, which can enhance both intrinsic and extrinsic apoptotic pathways [54]. For example, morusin, a naturally occurring prenylated flavonoid, exhibited pro-apoptotic effects on A549 and NCI-H292 cells by activating both the JNK and ERK pathways, as evidenced by changes in the ratio of phosphorylated to total protein levels. The use of a MEK/ERK inhibitor (U0126) and a JNK inhibitor (SP600125) confirmed the involvement of these pathways in apoptosis induced by morusin [55].
4. Materials and Methods
4.1. Materials
The reagents used in this study include MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) and acridine orange dye, both purchased from Sigma-Aldrich (St. Louis, MO, USA). Hoechst 33342 dye [2′-(4-Ethoxyphenyl)-6-(4-methyl-1-piperazinyl)-1H,3’H-2,5’-bibenzimidazole] was obtained from Thermo Fisher Scientific (Invitrogen™, Thermo Fisher Scientific Inc., Waltham, MA, USA). Fetal bovine serum (FBS), trypsin-EDTA, Roswell Park Memorial Institute (RPMI 1640) medium, and penicillin-streptomycin were acquired from HiMedia Laboratories (Mumbai, India), while JC-1 dye (5,5’,6,6’-Tetrachloro-1,1’,3,3-tetraethylbenzimidazolylcarbocyanine iodide) was sourced from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Lastly, the Immobilon™ Western Chemiluminescent HRP Substrate and Guava Cell Cycle® reagent were purchased from Merck Millipore (Merck Millipore Corp., Darmstadt, Germany).
4.2. Plant Extraction
The dried flowers of Mammea siamensis were purchased from the Thai Lanna Herbal Industry, Chiang Mai province, Thailand, in September 2018. The extraction method used was as described in our previous study [56]. Briefly, the powder was macerated in methanol solvents at a plant sample 50 g/500 mL solvent ratio (1:10 w/v) for 3 days. The extractant was filtered using a filter cloth and Whatman® Qualitative filter paper No. 4 after three days, and it was then evaporated using a rotary vacuum evaporator. The MS extract was concentrated using a vacuum desiccator for a day, resulting in 9.94 g (19.88%) of extract, which was subsequently stored at -20 °C for use in experiments. The DMSO was used to dissolve the MS extract.
4.3. 1H NMR Analysis
Proton nuclear magnetic resonance (1H NMR) and gradient correlation spectroscopy (gCOSY) were recorded on a 500 MHz Bruker Avance NMR spectrometer in DMSO-d6 as the solvent and referenced to the solvent peak at 2.50 ppm.
4.4. Cells Culture
The human colon cancer HCT116 cell line was obtained from the American Type Culture Collection (ATCC, Manassas, VA). Monolayer cells were cultured in RPMI 1640 media supplemented with 10% fetal bovine serum (FBS), 100 U/mL of penicillin, and 100 µg/mL of streptomycin. A CO₂ incubator was used to cultivate the cells at 37°C, 5% CO2, and 95% relative humidity. Trypsin-EDTA at a concentration of 0.25% was used for subtraction.
4.5. Cytotoxicity Assay
The effect of MS extract on the cytotoxicity was determined by MTT assay. HCT116 cells were plated overnight before being exposed to different concentrations of MS extract (30, 50, 100, and 150 µg/mL) for 24 h, while 0.3% DMSO was administered to the control. Following a 2-hour incubation with MTT solution (0.5 mg/mL), the formazan crystal was dissolved using DMSO. Colored formazan product was measured at 570 nm by using a Multiskan Sky microplate reader (Thermo Fisher Scientific, Waltham, MA, USA). The results were presented as percentages of cell viability, and GraphPad Prism 9 (GraphPad Prism Software, Inc., San Diego, CA, USA) was used to calculate the IC50.
4.6. Hoechst33342 Staining
Nuclear morphological changes were observed by Hoechst33342 staining. HCT116 cells were treated with various concentrations of MS extract (30, 50, 100, and 150 µg/mL) for 24 h, while 0.3% DMSO was administered to the control group. Next, the cells were treated with Hoechst 33342 dye and allowed to incubate for 30 minutes. Following staining, cells were observed using a fluorescence microscope (DP73+IX71 Olympus, Tokyo, Japan).
4.7. JC-1 Staining
Mitochondrial Membrane Potential (ΔΨm) was examined by JC-1 staining. HCT116 cells were treated with various concentrations of MS extract (30, 50, 100, and 150 µg/mL) for 6 h, while 0.3% DMSO was administered to the control group. After staining with JC-1 dye, the cells were photographed with a fluorescence microscope (DP73+IX71 Olympus, Tokyo, Japan).
4.8. Cell Cycle Analysis
To investigate cell populations in different phases of the cell cycle, we used flow cytometry. After 24 hours of treatment, the cells were harvested and washed with PBS, then fixed by ice-cold 70% ethanol. Following that, the cells were stained with Guava Cell Cycle® reagent. Subsequently, the DNA content was analyzed using the Guava EasyCyteTM flow cytometer and GuavaSoftTM software version 3.2 (Merck Millipore Corp., Merck KGaA, Darmstadt, Germany).
4.9. Acridine Orange (AO) Staining
To determine the effect of MS extract on V-ATPase activity, we performed AO staining. HCT116 cells were plated in 24-well culture plates and grown overnight. The following day, the cells were treated with MS extract at various concentrations (30, 50, 100, and 150 µg/mL) or with 50 nM bafilomycin A1 (BMA), a V-ATPase inhibitor, and 0.3% DMSO was administered to the control group for 6 h. After treatment, the cells were stained with AO fluorescence dye (1 µg/mL), followed by washing with PBS, and were then observed under a fluorescence microscope (DP73+IX71, Olympus, Tokyo, Japan).
4.10. Western Blot Analysis
The Western blot examination revealed the protein expression findings described in our previous study [56]. Briefly, the protein extracted was separated using SDS-PAGE and subsequently transferred to PVDF membranes. These membranes were blocked with 5% BSA and incubated overnight with the primary antibody, followed by a secondary antibody conjugated with HRP (Cell Signaling Technology, Beverly, MA.) for 1 h, respectively. β-actin served as the internal control (Merck Millipore Corp., Darmstadt, Germany).
4.11. Statistical Analysis
Statistical analysis was examined by one-way analysis variance (ANOVA), followed by Tukey’s post hoc test, using SPSS software version 20.0 (IBM Crop., Albany, NY, USA). The statistical significance was accepted at p < 0.05 and p < 0.01. All the data are shown as mean ± standard deviation (SD).
5. Conclusions
In conclusion, this study suggests that the methanol extract of Mammea siamensis (MS) can inhibit cancer cell growth in HCT116 cells by inducing apoptosis through both intrinsic and extrinsic pathways. Mechanistic analysis revealed that the MS extract activates the MAPK pathway while inhibiting the PI3K/Akt, NF-κB, and GSK-3β/β-catenin pathways. Notably, MS extract was identified as a potent V-ATPase inhibitor. Based on these findings, MS extract demonstrates significant anti-cancer activity. However, further studies are needed to clarify the interactions between these signaling pathways. Future research should also focus on purifying the active compounds from the MS extract, which could pave the way for their potential use as therapeutic agents for colon cancer.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Effect of MS extract on cell viability in Vero cells by MTT assay.
Author Contributions
Conceptualization, W.P. and R.W.; methodology, P.S.; W.P.; A.K. and S.S.; validation, P.S.; S.I. and R.W.; formal analysis, P.S. and S.S.; investigation, P.S.; resources, R.W.; data curation R.W.; P.S., writing—original draft preparation, P.S.; writing—review and editing, S.I. and R.W.; visualization, R.W.; supervision, R.W.; project administration, R.W.; funding acquisition, R.W. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research project is supported by the National Research Council of Thailand (NRCT): NRCT5-RGJ63017-152.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
We would like to thank the Graduate School of Srinakharinwirot University and the Faculty of Medicine, Srinakharinwirot University, Bangkok, Thailand for the instruments and laboratory rooms.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R. L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F., Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71 (3), 209-249. [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A., Epidemiology of colorectal cancer: incidence, mortality, survival, and risk factors. Prz Gastroenterol 2019, 14 (2), 89-103. [CrossRef]
- Kuppusamy, P.; Yusoff, M. M.; Maniam, G. P.; Ichwan, S. J.; Soundharrajan, I.; Govindan, N., Nutraceuticals as potential therapeutic agents for colon cancer: a review. Acta Pharm Sin B 2014, 4 (3), 173-81. [CrossRef]
- Badavenkatappa Gari, S.; Nelson, V. K.; Peraman, R., Tinospora sinensis (Lour.) Merr alkaloid rich extract induces colon cancer cell death via ROS mediated, mTOR dependent apoptosis pathway: "an in-vitro study". BMC Complement Med Ther 2023, 23 (1), 33. [CrossRef]
- Luo, F.; Sugita, H.; Muraki, K.; Saeki, S.; Chaipech, S.; Pongpiriyadacha, Y.; Muraoka, O.; Morikawa, T., Anti-proliferative activities of coumarins from the Thai medicinal plant Mammea siamensis (Miq.) T. Anders. against human digestive tract carcinoma cell lines. Fitoterapia 2021, 148, 104780. [CrossRef]
- Luo, F.; Manse, Y.; Chaipech, S.; Pongpiriyadacha, Y.; Muraoka, O.; Morikawa, T., Structures of Mammeasins P and Q, Coumarin-Related Polysubstituted Benzofurans, from the Thai Medicinal Plant Mammea siamensis (Miq.) T. Anders.: Anti-Proliferative Activity of Coumarin Constituents against Human Prostate Carcinoma Cell Line LNCaP. Pharmaceuticals (Basel) 2023, 16 (2). [CrossRef]
- Chaniad, P.; Chukaew, A.; Payaka, A.; Phuwajaroanpong, A.; Techarang, T.; Plirat, W.; Punsawad, C., Antimalarial potential of compounds isolated from Mammea siamensis T. Anders. flowers: in vitro and molecular docking studies. BMC Complementary Medicine and Therapies 2022, 22 (1), 266. [CrossRef]
- Chaniad, P.; Chukaew, A.; Na-Ek, P.; Yusakul, G.; Chuaboon, L.; Phuwajaroanpong, A.; Plirat, W.; Konyanee, A.; Septama, A. W.; Punsawad, C., In vivo antimalarial effect of 1-hydroxy-5,6,7-trimethoxyxanthone isolated from Mammea siamensis T. Anders. flowers: pharmacokinetic and acute toxicity studies. BMC Complement Med Ther 2024, 24 (1), 129. [CrossRef]
- Steinrut, L.; Itharat, A.; Ruangnoo, S., Free radical scavenging and lipid peroxidation of Thai medicinal plants used for diabetic treatment. J Med Assoc Thai 2011, 94 Suppl 7, S178-82.
- Sukkasem, K.; Panthong, S.; Itharat, A., Antimicrobial Activities of Thai Traditional Remedy “Kheaw-Hom” and Its Plant Ingredients for Skin Infection Treatment in Chickenpox. J Med Assoc Thai 2016, 99 suupl 4, 116-23.
- Noysang, C.; Mahringer, A.; Zeino, M.; Saeed, M.; Luanratana, O.; Fricker, G.; Bauer, R.; Efferth, T., Cytotoxicity and inhibition of P-glycoprotein by selected medicinal plants from Thailand. Journal of Ethnopharmacology 2014, 155 (1), 633-641. [CrossRef]
- Uto, T.; Tung, N. H.; Thongjankaew, P.; Lhieochaiphant, S.; Shoyama, Y., Kayeassamin A Isolated from the Flower of Mammea siamensis Triggers Apoptosis by Activating Caspase-3/-8 in HL-60 Human Leukemia Cells. Pharmacognosy Res 2016, 8 (4), 244-248. [CrossRef]
- Maki, P.; Itharat, A.; Thongdeeying, P.; Tuy-on, T.; Kuropakornpong, P.; Pipatrattanaseree, W.; Mingmalairak, C.; Davies, N. M., Ethnopharmacological nexus between the traditional Thai medicine theory and biologically based cancer treatment. Journal of Ethnopharmacology 2022, 287, 114932. [CrossRef]
- Pfeffer, C. M.; Singh, A. T. K., Apoptosis: A Target for Anticancer Therapy. International Journal of Molecular Sciences 2018, 19 (2), 448. [CrossRef]
- Hu, W.; Kavanagh, J. J., Anticancer therapy targeting the apoptotic pathway. Lancet Oncol 2003, 4 (12), 721-9. [CrossRef]
- Park, C.; Park, S.; Chung, Y. H.; Kim, G.-Y.; Choi, Y. W.; Kim, B. W.; Choi, Y. H., Induction of apoptosis by a hexane extract of aged black garlic in the human leukemic U937 cells. Nutr Res Pract 2014, 8 (2), 132-137. [CrossRef]
- Elmore, S., Apoptosis: a review of programmed cell death. Toxicol Pathol 2007, 35 (4), 495-516. [CrossRef]
- Jin, Z.; El-Deiry, W. S., Overview of cell death signaling pathways. Cancer Biol Ther 2005, 4 (2), 139-63. [CrossRef]
- Zhang, Y.; Wang, X., Targeting the Wnt/β-catenin signaling pathway in cancer. Journal of Hematology & Oncology 2020, 13 (1), 165. [CrossRef]
- Glaviano, A.; Foo, A. S. C.; Lam, H. Y.; Yap, K. C. H.; Jacot, W.; Jones, R. H.; Eng, H.; Nair, M. G.; Makvandi, P.; Geoerger, B.; Kulke, M. H.; Baird, R. D.; Prabhu, J. S.; Carbone, D.; Pecoraro, C.; Teh, D. B. L.; Sethi, G.; Cavalieri, V.; Lin, K. H.; Javidi-Sharifi, N. R.; Toska, E.; Davids, M. S.; Brown, J. R.; Diana, P.; Stebbing, J.; Fruman, D. A.; Kumar, A. P., PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Molecular Cancer 2023, 22 (1), 138. [CrossRef]
- Dhillon, A. S.; Hagan, S.; Rath, O.; Kolch, W., MAP kinase signalling pathways in cancer. Oncogene 2007, 26 (22), 3279-3290. [CrossRef]
- Whitton, B.; Okamoto, H.; Packham, G.; Crabb, S. J., Vacuolar ATPase as a potential therapeutic target and mediator of treatment resistance in cancer. Cancer Medicine 2018, 7 (8), 3800-3811. [CrossRef]
- Cotter, K.; Stransky, L.; McGuire, C.; Forgac, M., Recent Insights into the Structure, Regulation, and Function of the V-ATPases. Trends Biochem Sci 2015, 40 (10), 611-622. [CrossRef]
- Chen, F.; Kang, R.; Liu, J.; Tang, D., The V-ATPases in cancer and cell death. Cancer Gene Therapy 2022, 29 (11), 1529-1541. [CrossRef]
- Sennoune, S. R.; Martinez-Zaguilan, R., Plasmalemmal vacuolar H+-ATPases in angiogenesis, diabetes and cancer. Journal of Bioenergetics and Biomembranes 2007, 39 (5), 427-433. [CrossRef]
- Joshi, K. R.; Devkota, H. P.; Nakamura, T.; Watanabe, T.; Yahara, S., Chemical constituents and their DPPH radical scavenging activity of Nepalese crude drug Begonia picta. Records of Natural Products 2015, 9 (3), 446.
- Kuerban, M.; Ma, F.; Shan, L.; Wang, Y.; Zhou, G., Comparative discriminant analysis of Mesua ferrea L. and its adulterants. Heliyon 2024, 10 (7), e28459. [CrossRef]
- Plesca, D.; Mazumder, S.; Almasan, A., DNA damage response and apoptosis. Methods Enzymol 2008, 446, 107-22. [CrossRef]
- Yu, Y.; Fei, Z.; Qin, L., Anticancer effects of α-mangostin in OVACAR-3 human ovarian carcinoma cells are mediated via involvement of reactive oxygen species, mitochondrial -mediated apoptosis, suppression of cell migration and invasion and m-TOR/PI3K/AKT signaling pathway. J buon 2020, 25 (5), 2293-2300.
- Yokomakura, A.; Hong, J.; Ohuchi, K.; Oh, S. E.; Lee, J. Y.; Mano, N.; Takahashi, T.; Hwang, G. W.; Naganuma, A., Increased production of reactive oxygen species by the vacuolar-type (H(+))-ATPase inhibitors bafilomycin A1 and concanamycin A in RAW 264 cells. J Toxicol Sci 2012, 37 (5), 1045-8. [CrossRef]
- Ribeiro, A. S.; Estanqueiro, M.; Oliveira, M. B.; Sousa Lobo, J. M., Main Benefits and Applicability of Plant Extracts in Skin Care Products. Cosmetics 2015, 2 (2), 48-65. [CrossRef]
- Wang, Q.; Zhang, J.; Ye, J.; Guo, J., Vitexin exerts anti-tumor and anti-angiogensis effects on cervical cancer through VEGFA/VEGFR2 pathway. European Journal of Gynaecological Oncology 2022, 43 (4), 86-91. [CrossRef]
- Zou, S.; Wu, Y.; Wen, M.; Liu, J.; Chen, M.; Yuan, J.; Zhou, B. Potential Molecular Mechanism of Illicium simonsii Maxim Petroleum Ether Fraction in the Treatment of Hepatocellular Carcinoma Pharmaceuticals [Online], 2024. [CrossRef]
- Giordano, F.; Comità, S.; Venneri, G.; Rago, V.; Naimo, G. D.; De Amicis, F.; De Bartolo, A.; Tundis, R.; Mauro, L.; Panno, M. L., Poncirus trifoliata (L.) Raf. Seed Extract Induces Cell Cycle Arrest and Apoptosis in the Androgen Receptor Positive LNCaP Prostate Cancer Cells. Int J Mol Sci 2023, 24 (22). [CrossRef]
- Prokhorova, E. A.; Zamaraev, A. V.; Kopeina, G. S.; Zhivotovsky, B.; Lavrik, I. N., Role of the nucleus in apoptosis: signaling and execution. Cell Mol Life Sci 2015, 72 (23), 4593-612. [CrossRef]
- Tung, N. H.; Uto, T.; Sakamoto, A.; Hayashida, Y.; Hidaka, Y.; Morinaga, O.; Lhieochaiphant, S.; Shoyama, Y., Antiproliferative and apoptotic effects of compounds from the flower of Mammea siamensis (Miq.) T. Anders. on human cancer cell lines. Bioorganic & Medicinal Chemistry Letters 2013, 23 (1), 158-162. [CrossRef]
- Yuan, N.; Song, L.; Zhang, S.; Lin, W.; Cao, Y.; Xu, F.; Fang, Y.; Wang, Z.; Zhang, H.; Li, X.; Wang, Z.; Cai, J.; Wang, J.; Zhang, Y.; Mao, X.; Zhao, W.; Hu, S.; Chen, S.; Wang, J., Bafilomycin A1 targets both autophagy and apoptosis pathways in pediatric B-cell acute lymphoblastic leukemia. Haematologica 2015, 100 (3), 345-56. [CrossRef]
- Watanapokasin, R.; Innajak, S.; Nilwarangoon, S., PO-230 Effect of C. cochinchinense extract on V-ATPase inhibition triggering apoptosis in human cancer cell lines. ESMO Open 2018, 3, A110. [CrossRef]
- Lu, D.; Choi, M. Y.; Yu, J.; Castro, J. E.; Kipps, T. J.; Carson, D. A., Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocytic leukemia cells. Proceedings of the National Academy of Sciences 2011, 108 (32), 13253-13257. [CrossRef]
- Chen, Y.; Chen, M.; Deng, K., Blocking the Wnt/β-catenin signaling pathway to treat colorectal cancer: Strategies to improve current therapies (Review). Int J Oncol 2023, 62 (2), 24. [CrossRef]
- Chen, S.; Guttridge, D. C.; You, Z.; Zhang, Z.; Fribley, A.; Mayo, M. W.; Kitajewski, J.; Wang, C. Y., Wnt-1 signaling inhibits apoptosis by activating beta-catenin/T cell factor-mediated transcription. J Cell Biol 2001, 152 (1), 87-96. [CrossRef]
- Xu, M.; Wang, S.; Song, Y.; Yao, J.; Huang, K.; Zhu, X., Apigenin suppresses colorectal cancer cell proliferation, migration and invasion via inhibition of the Wnt/β-catenin signaling pathway. Oncol Lett 2016, 11 (5), 3075-3080. [CrossRef]
- He, Y.; Sun, M. M.; Zhang, G. G.; Yang, J.; Chen, K. S.; Xu, W. W.; Li, B., Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduction and Targeted Therapy 2021, 6 (1), 425. [CrossRef]
- Rascio, F.; Spadaccino, F.; Rocchetti, M. T.; Castellano, G.; Stallone, G.; Netti, G. S.; Ranieri, E., The Pathogenic Role of PI3K/AKT Pathway in Cancer Onset and Drug Resistance: An Updated Review. Cancers (Basel) 2021, 13 (16). [CrossRef]
- Zhang, G.; Li, D.; Chen, H.; Zhang, J.; Jin, X., Vitexin induces G2/M-phase arrest and apoptosis via Akt/mTOR signaling pathway in human glioblastoma cells. Mol Med Rep 2018, 17 (3), 4599-4604. [CrossRef]
- Liu, X.; Jiang, Q.; Liu, H.; Luo, S., Vitexin induces apoptosis through mitochondrial pathway and PI3K/Akt/mTOR signaling in human non-small cell lung cancer A549 cells. Biological Research 2019, 52 (1), 7. [CrossRef]
- Bai, D.; Ueno, L.; Vogt, P. K., Akt-mediated regulation of NFkappaB and the essentialness of NFkappaB for the oncogenicity of PI3K and Akt. Int J Cancer 2009, 125 (12), 2863-70. [CrossRef]
- Xia, Z. B.; Meng, F. R.; Fang, Y. X.; Wu, X.; Zhang, C. W.; Liu, Y.; Liu, D.; Li, G. Q.; Feng, F. B.; Qiu, H. Y., Inhibition of NF-κB signaling pathway induces apoptosis and suppresses proliferation and angiogenesis of human fibroblast-like synovial cells in rheumatoid arthritis. Medicine (Baltimore) 2018, 97 (23), e10920. [CrossRef]
- Ye, Y.; Zhou, J., The protective activity of natural flavonoids against osteoarthritis by targeting NF-κB signaling pathway. Frontiers in Endocrinology 2023, 14. [CrossRef]
- Vidya Priyadarsini, R.; Senthil Murugan, R.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S., The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. European Journal of Pharmacology 2010, 649 (1), 84-91. [CrossRef]
- Yue, J.; López, J. M., Understanding MAPK Signaling Pathways in Apoptosis. International Journal of Molecular Sciences 2020, 21 (7), 2346. [CrossRef]
- He, J. D.; Wang, Z.; Li, S. P.; Xu, Y. J.; Yu, Y.; Ding, Y. J.; Yu, W. L.; Zhang, R. X.; Zhang, H. M.; Du, H. Y., Vitexin suppresses autophagy to induce apoptosis in hepatocellular carcinoma via activation of the JNK signaling pathway. Oncotarget 2016, 7 (51), 84520-84532. [CrossRef]
- Shin, D. Y.; Lee, W. S.; Lu, J. N.; Kang, M. H.; Ryu, C. H.; Kim, G. Y.; Kang, H. S.; Shin, S. C.; Choi, Y. H., Induction of apoptosis in human colon cancer HCT-116 cells by anthocyanins through suppression of Akt and activation of p38-MAPK. Int J Oncol 2009, 35 (6), 1499-1504. [CrossRef]
- Cagnol, S.; Chambard, J.-C., ERK and cell death: Mechanisms of ERK-induced cell death – apoptosis, autophagy and senescence. The FEBS Journal 2010, 277 (1), 2-21. [CrossRef]
- Wang, J.; Liu, X.; Zheng, H.; Liu, Q.; Zhang, H.; Wang, X.; Shen, T.; Wang, S.; Ren, D., Morusin induces apoptosis and autophagy via JNK, ERK and PI3K/Akt signaling in human lung carcinoma cells. Chemico-Biological Interactions 2020, 331, 109279. [CrossRef]
- Sitthisuk, P.; Innajak, S.; Poorahong, W.; Samosorn, S.; Dolsophon, K.; Watanapokasin, R., Effect of Acacia concinna Extract on Apoptosis Induction Associated with Endoplasmic Reticulum Stress and Modulated Intracellular Signaling Pathway in Human Colon HCT116 Cancer Cells. Nutrients 2024, 16 (21), 3764. [CrossRef]
Figure 1.
1H-NMR spectrum with expansion of MS methanolic extract in DMSO-d6.
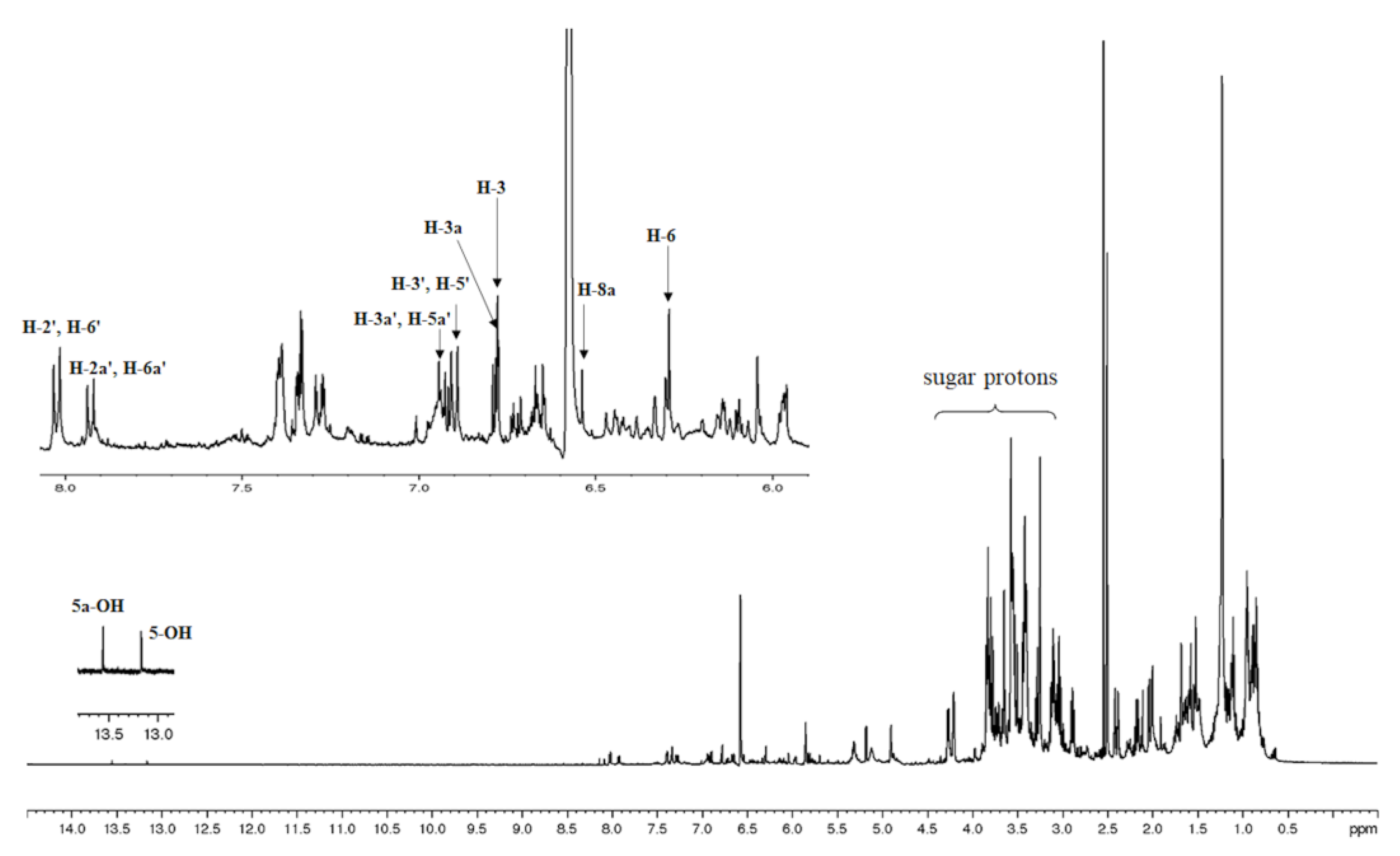
Figure 2.
Chemical structures of vitexin and isovitexin.
Figure 3.
Effects of MS extracts on cytotoxicity in HCT116 cells. Cells were treated with various concentrations of MS extract for 24 h and examined by using the MTT assay. The percentage of cell viability was shown as mean values ± SD of three independent experiments. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 3.
Effects of MS extracts on cytotoxicity in HCT116 cells. Cells were treated with various concentrations of MS extract for 24 h and examined by using the MTT assay. The percentage of cell viability was shown as mean values ± SD of three independent experiments. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
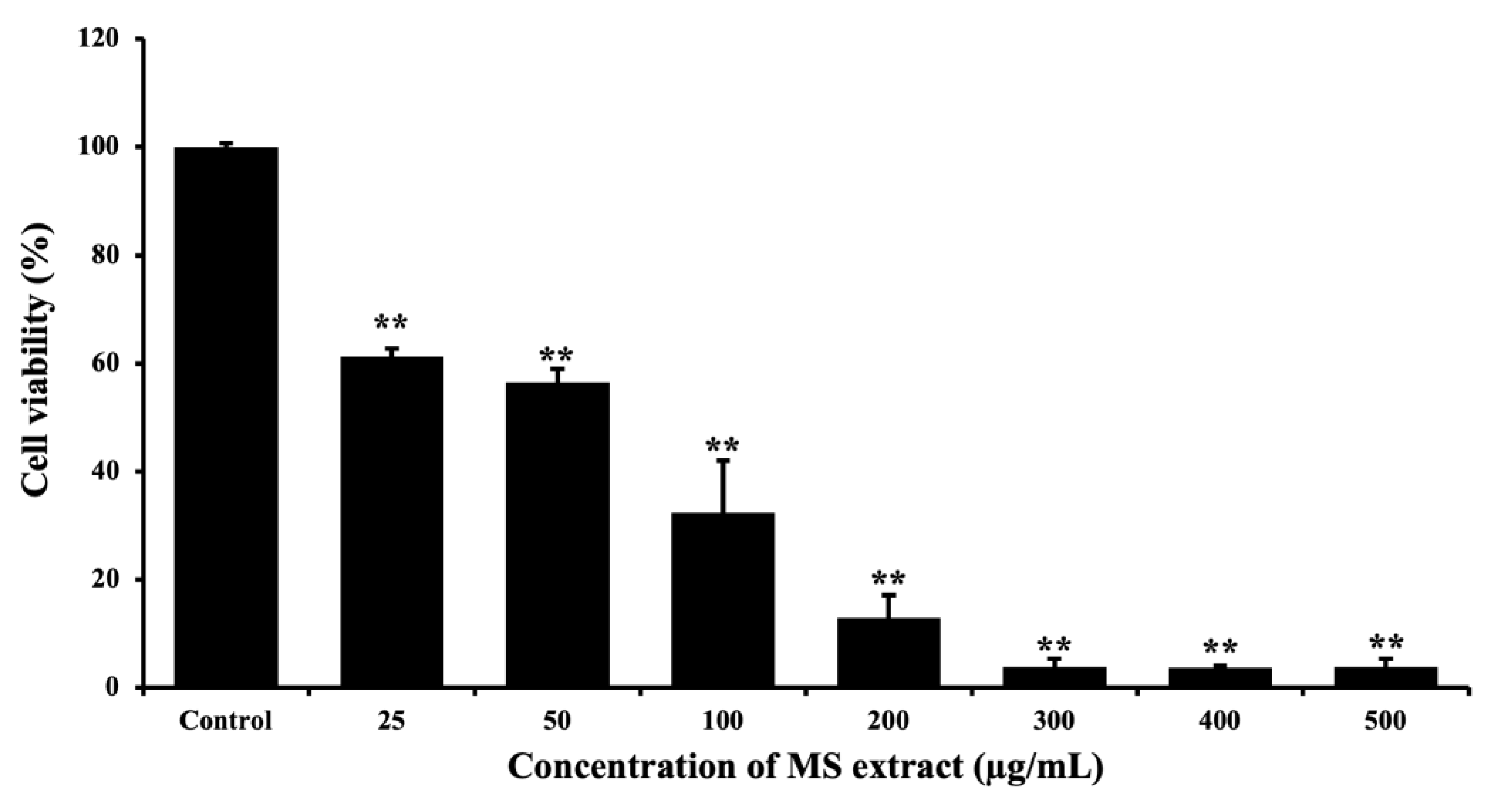
Figure 4.
Effects of MS extracts on nuclear morphological change in HCT116 cells. (A) Fluorescence pictures of Hoechst 33342 staining captured with a fluorescence microscope (20x). Red arrows indicate condensed HCT116 cells and the presence of apoptotic bodies. (B) Histogram showing the proportion of nuclear-condensed cells compared to control cells. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 4.
Effects of MS extracts on nuclear morphological change in HCT116 cells. (A) Fluorescence pictures of Hoechst 33342 staining captured with a fluorescence microscope (20x). Red arrows indicate condensed HCT116 cells and the presence of apoptotic bodies. (B) Histogram showing the proportion of nuclear-condensed cells compared to control cells. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
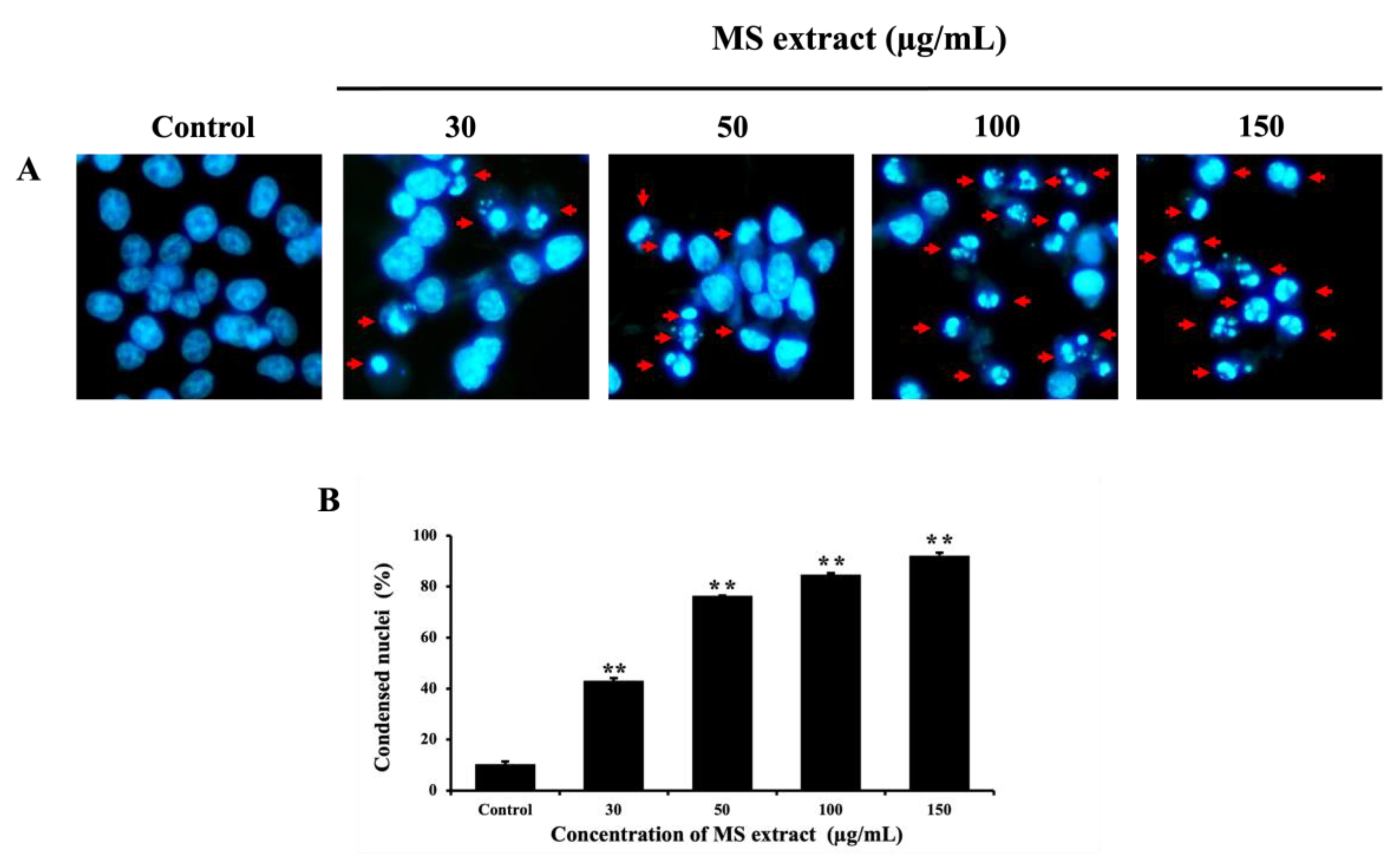
Figure 5.
Effects of MS extracts on ΔΨm in HCT116 cells. (A) Fluorescence pictures of JC-1 staining captured with a fluorescence microscope (20x). An increase in green fluorescence signifies a reduction in ΔΨm. (B) Histogram showing the relative red fluorescence intensity compared to control cells. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 5.
Effects of MS extracts on ΔΨm in HCT116 cells. (A) Fluorescence pictures of JC-1 staining captured with a fluorescence microscope (20x). An increase in green fluorescence signifies a reduction in ΔΨm. (B) Histogram showing the relative red fluorescence intensity compared to control cells. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
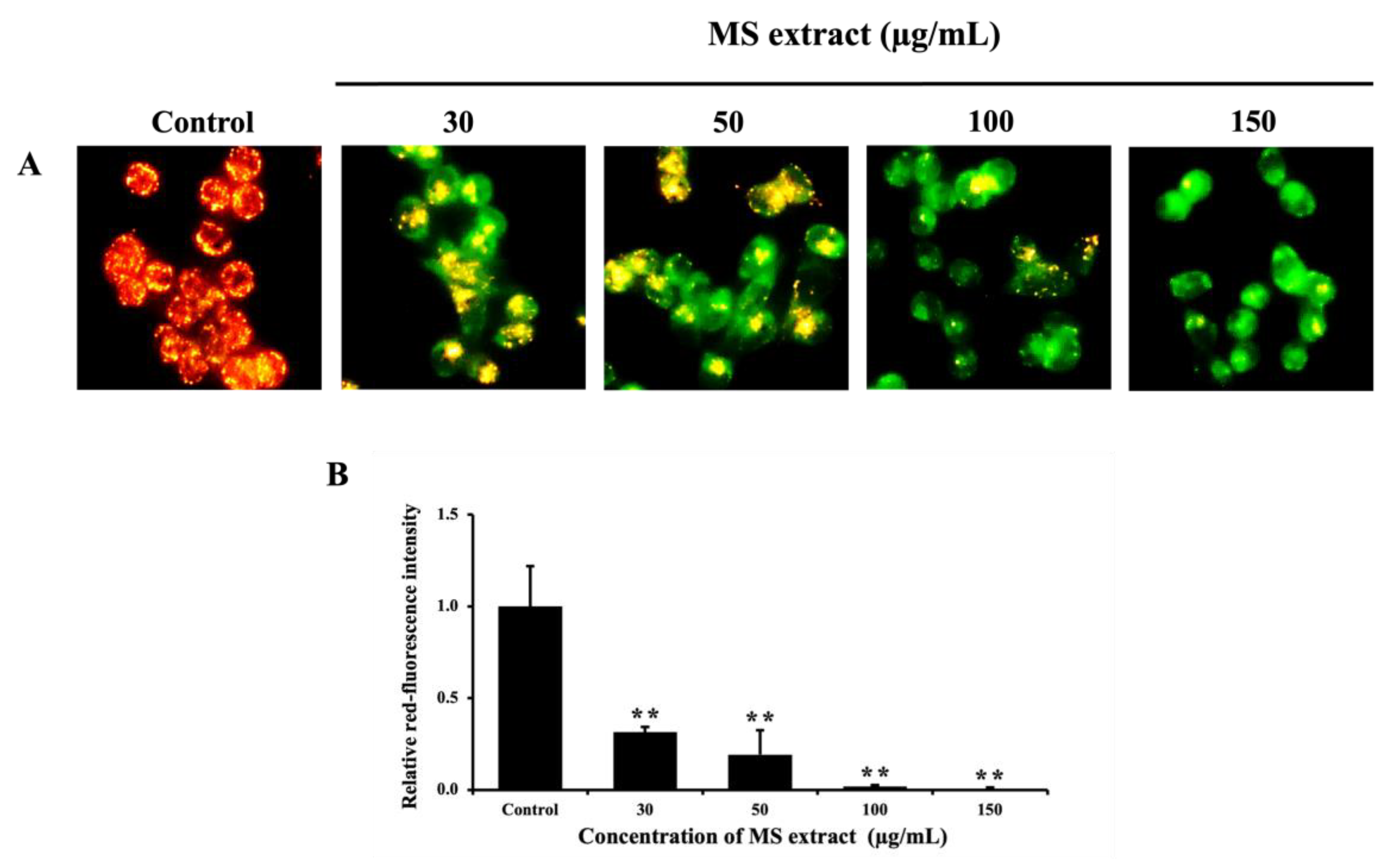
Figure 6.
Effects of MS extracts on the sub-G1 population in HCT116 cells. (A) Representative histograms analyzed by flow cytometry. (B) Percentage of cells in the sub-G1 phase relative to control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 6.
Effects of MS extracts on the sub-G1 population in HCT116 cells. (A) Representative histograms analyzed by flow cytometry. (B) Percentage of cells in the sub-G1 phase relative to control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
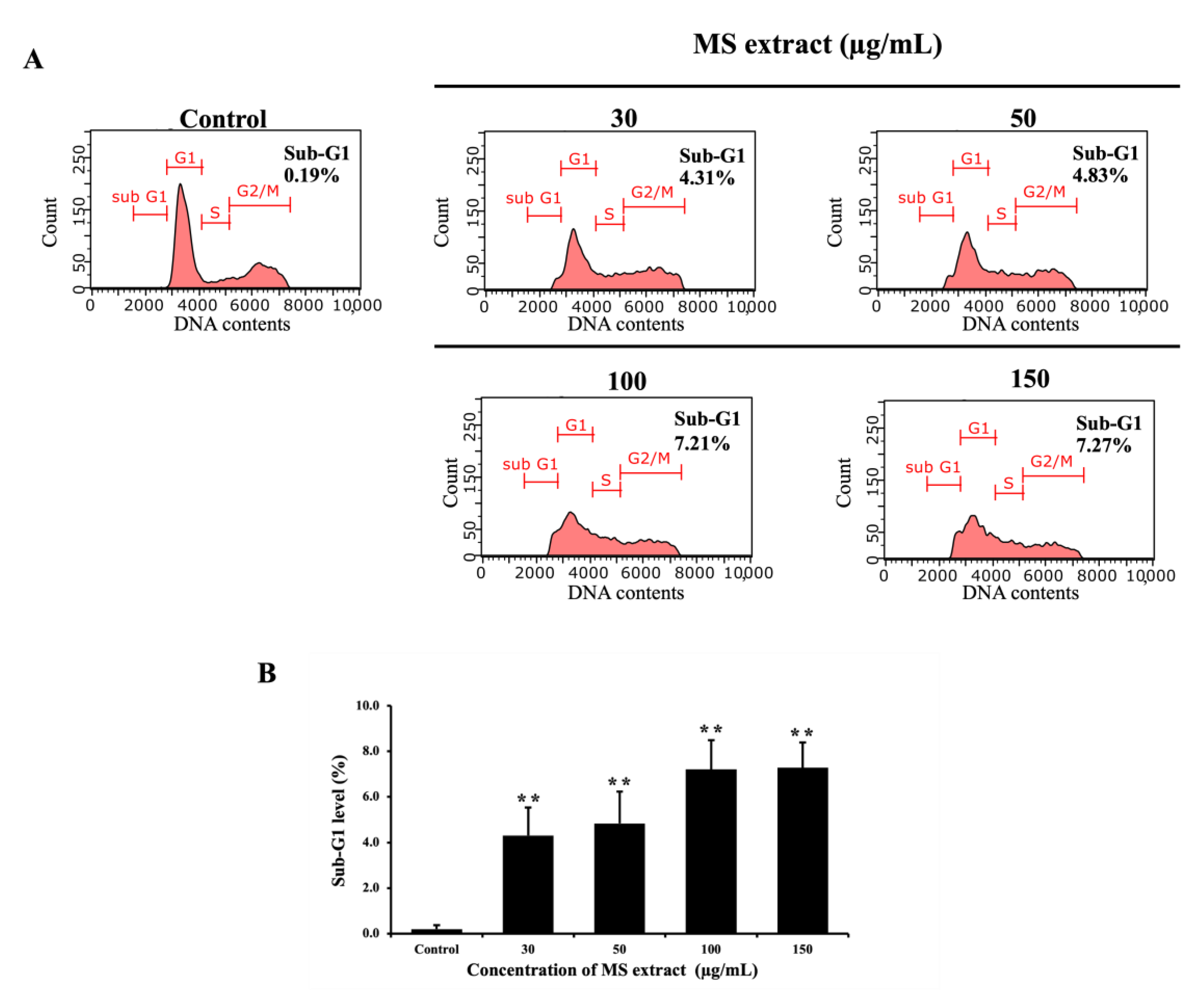
Figure 7.
The effects of MS extracts on the protein expression of (A) Bcl-2 family proteins, (B) caspase, and PARP in HCT116 cells were detected by western blot analysis. (C,D) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 7.
The effects of MS extracts on the protein expression of (A) Bcl-2 family proteins, (B) caspase, and PARP in HCT116 cells were detected by western blot analysis. (C,D) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
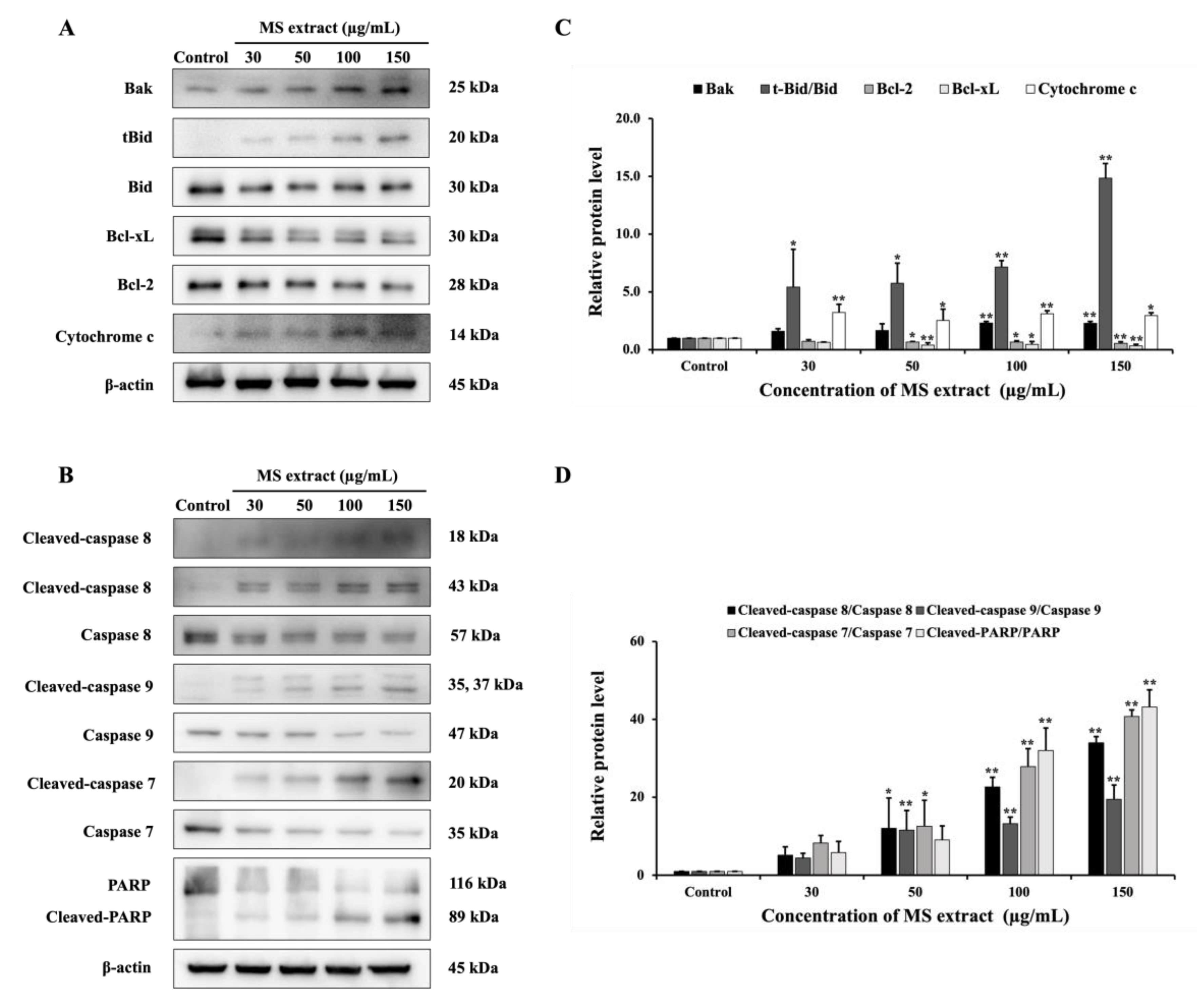
Figure 8.
Effects of MS extracts on V-ATPase activity in HCT116 cells. (A) Fluorescence microscopy images of AO-stained cells (20X). The orange or red fluorescence indicates acidic vesicular organelles (AVOs), which might be generated by V-ATPase. BMA was used as a positive control. (B) The protein expression of ATP6V1B2 was detected by western blot analysis. (C) The histogram represents the relative red fluorescence intensity of AO staining compared to the control. (D) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 8.
Effects of MS extracts on V-ATPase activity in HCT116 cells. (A) Fluorescence microscopy images of AO-stained cells (20X). The orange or red fluorescence indicates acidic vesicular organelles (AVOs), which might be generated by V-ATPase. BMA was used as a positive control. (B) The protein expression of ATP6V1B2 was detected by western blot analysis. (C) The histogram represents the relative red fluorescence intensity of AO staining compared to the control. (D) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
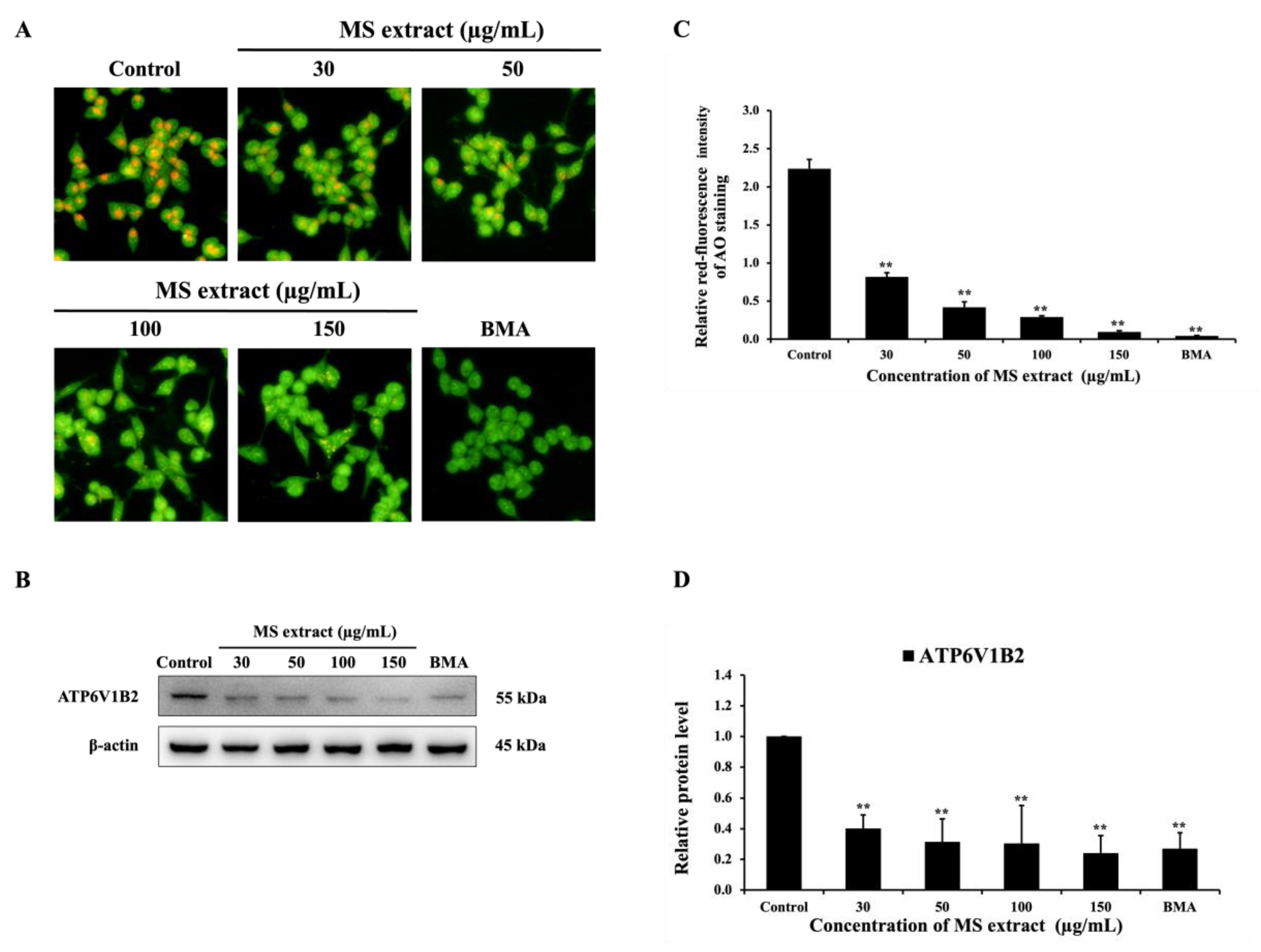
Figure 9.
Effects of MS extract on protein expression involved (A) the GSK-3β/β-catenin pathway, (B) the PI3K/Akt/ NF-κB pathway, and (C) the MAPK pathway in HCT116 cells were detected by western blot analysis. (D,E,F) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Figure 9.
Effects of MS extract on protein expression involved (A) the GSK-3β/β-catenin pathway, (B) the PI3K/Akt/ NF-κB pathway, and (C) the MAPK pathway in HCT116 cells were detected by western blot analysis. (D,E,F) The intensity of the bands in relation to the control. *p < 0.05 and **p < 0.01 suggests a statistically significant difference relative to the control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



