Submitted:
27 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
The susceptibility of meat products to contamination with Listeria monocytogenes, foodborne pathogen associated with high mortality among consumers, requires the exploration of novel strategies with the use of hurdle technology. Essential oils have been shown to reduce exposure times to heat treatments, while the concentration of their main antimicrobial compounds impacts food safety by reducing their quantities applied in foods. The aim of this study was to evaluate the effect of the sublethal concentrations of Poliomintha longiflora pure oregano essential oil (PEO) and fractionated at 140 °C (FIV) on L. monocytogenes inactivation at 52, 54, 57.5 and 63 °C. The Weibull-Mafart model was suitable to describe the observed inactivation data. The FIV group, characterized by a chemical profile of 60.23 % carvacrol and 21.17 % thymol, resulted in δ-values of δ52 °C = 0.40, δ54 °C = 0.44, δ57.5°C= 0.35 and δ63°C=0.33 min, representing a significant reduction (P < 0.05) in comparison to the control group. The z-values were estimated as 5.75, 5.20 and 5.00 °C for control, PEO and FIV, respectively. Therefore, the application of fractionated oregano essential oil in meat products presents a promising alternative to reducing exposure times to heat treatments, effectively lowering the incidence of L. monocytogenes contamination while preserving product quality.
Keywords:
thermal inactivation
; predictive microbiology
; foodborne pathogens
; food safety
; hurdle technology
; natural antimicrobials
1. Introduction
Meat products have the necessary nutritional composition for the survival and growth of a wide diversity of foodborne pathogens, making them potentially hazardous [1]. Listeria monocytogenes, a gram-positive bacterium found in soil, water, and mammals can cause listeriosis when contaminated foods are consumed [2]. It has been highlighted as one of the food-associated bacteria with the highest mortality [3]. Gastrointestinal symptoms are common in listeriosis, but the infection can spread to the nervous system, leading to severe complications in immunocompromised individuals. In pregnant women, it can result in infections that may cause miscarriage or neonatal sepsis [4].
In the latest report from the European Food Safety Authority (EFSA), it was stated that there was a 5.80 % increase in listeriosis cases in 2023 compared to 2022, with nearly 3,000 reported cases in the European Union. Of these, 96.50 % resulted in hospitalizations with a total of 335 deaths [4]. In North America, the Centers for Disease Control and Prevention (CDC) monitor and control foodborne outbreaks, with L. monocytogenes being one of the major concerns in the United States, being associated with a wide variety of products, including vegetables [5], ice cream [6,7], cheeses [8], and meat products [9]. The CDC reports that L. monocytogenes is responsible for more than 1,500 illnesses and 200 deaths annually in the U.S., with a significant portion of these cases requiring hospitalization [10]. The continued presence of L. monocytogenes in food products not only poses a risk to public health in the U.S., but also has potential implications for neighboring countries, such as Mexico, where similar risks are likely associated with cross-border food trade and consumption patterns [11].
It has been reported that L. monocytogenes has the ability to persist in food environments and survive against preservation treatments generally applied to inhibit or stop the growth of other foodborne pathogens [12]. These preservation techniques in food processing include the use of salt amounts lower than 10 % [13], acidic pH levels within 4 and 6 [14] and low (< 5 °C) or high temperatures (> 60°C) [1,15]. Long-term exposure to these stress conditions has been shown to enhance the adaptive response of L. monocytogenes, leading to alter its lag phase and its growth rate [16,17]. As a result, researchers are increasingly exploring combined treatments to reduce the incidence of this pathogen and its application in food matrices [18]. When implementing thermal treatments, avoiding the misuse of time-temperature parameters has prompted the application of novel sustainable strategies for microbial inactivation in foods. These include the use of natural additives like bacteriocins, organic acids, plant extracts and essential oils (EO) [19,20,21].
Essential oils are natural substances usually classified as “Generally Recognized as Safe'' (GRAS) that interact with the bacterial membrane changing its permeability, decreasing proton force, and causing coagulation of cytoplasm [22]. Oregano and thyme EOs contain high concentrations of monoterpenes, which have been reported to exhibit antilisterial activity in foods, with significant logarithmic reductions of L. monocytogenes observed in pork meat stored at 4 °C [23]. Origanum vulgare, a European species of oregano, has been widely explored for its antimicrobial properties against Gram-positive and Gram-negative bacteria with Minimum Inhibitory Concentrations (MIC) ranging from 0.49 to 1.90 mg/mL, and Minimum Bactericidal Concentrations (MBC) ranging from 0.99 to 7.90 mg/mL, where Gram-positive bacteria has been more resistant than negative [24,25]. Sublethal concentrations (SLC) of O. vulgare (1.25 µL/mL) at 30 °C can alter the growth parameters of L. monocytogenes after 1 h of exposure. This effect may improve the efficacy of antibiotics in medical applications or be combined with other inactivation treatments in the food sector [26]. Thus, the inactivation of L. monocytogenes can be achieved through novel strategies using mild concentrations, aligning with the hurdle technology concept [22].
In developing countries, including Mexico, regulations for the surveillance of L. monocytogenes in foods and infected consumers are often limited [27]. However, its presence in Mexican vegetables [28], dairy products [29], and meat products [11,30] indicate a possible health risk to the population. The EO and extracts of Mexican oregano, Lippia graveolens, are very popular in America due to its antimicrobial activity against Salmonella Typhimurium [31,32]. Poliomintha longiflora is another Mexican species of oregano but it has been scarcely studied in comparison with other oregano species. The chemical composition of a wild species from northern Mexico revealed an increased concentration of carvacrol and thymol [33]. This composition influences its antimicrobial activity as in O. vulgare and L. graveolens. By fractionating the pure EO, the concentration of key terpenes such as carvacrol and thymol can be enhanced, leading to increased antilisterial activity [34]. The investigation of natural sources plays a crucial role in understanding their potential in combating pathogens, fostering innovation in the region. This Mexican species represents a valuable and locally sourced resource, offering a promising alternative for food safety interventions [35]. Moreover, it provides an opportunity to develop sustainable, natural antimicrobial solutions that leverage Mexico’s rich biodiversity [36]. However, when applying these natural compounds, it is essential to assess SLC to optimize processing treatments. Despite being considered safe, EOs can be toxic to health when used under inappropriate conditions or at excessive concentrations [37].
Previous studies have reported that combining physical treatments such as High-Pressure Processing (HPP), freezing, irradiation, or thermal treatments, with sublethal concentrations of EOs can successfully reduce foodborne pathogens such as Salmonella spp., L. monocytogenes, Escherichia coli, Bacillus cereus and Campylobacter jejuni in food matrices [1,38,39]. The use of temperatures below 70 °C with oregano OE (OEO) in low concentrations improves thermal inactivation in food processing [40,41,42]. The microbial reductions yielded from combining EOs with other technologies can be estimated using predictive models [41]. Predictive models are mathematical equations that help elucidate the effects of intrinsic and extrinsic factors (e.g., temperature, pH, water activity) on the microbial responses in foods and are valuable tools for enabling the assessment of interventions to ensure food safety [43,44]. Among these interventions, the use of P. longiflora EO fractions with high concentrations of carvacrol and thymol has garnered attention due to its strong antimicrobial properties [34]. This approach reduces the need for high concentrations of EOs, making it a sustainable and effective option for food safety interventions. Besides, by focusing on local resources like P. longiflora, the potential toxic effects and bacterial resistance can be reduced, while promoting sustainable practices.
The aim of this work was to assess the impact of pure and fractionated P. longiflora EO (PEO and FIV) on the thermal inactivation of L. monocytogenes at mild temperatures (52, 54, 57.5 and 63 °C). This was achieved by determining the pathogen inactivation parameters in an In vitro assay using a simulated meat medium.
2. Materials and Methods
2.1. Bacterial Culture and Growth Conditions
L. monocytogenes PM1, a clinical isolate from the listeriosis outbreak in Andalusia linked to the consumption of ready-to-eat meats in 2019, was provided by the Food Science and Technology Department from the University of Córdoba, Spain. The strain was stored at -80 °C in 20 % v/v glycerol/Brain Heart Infusion broth (BHI; Difco Laboratories, Sparks MD., USA) until use. To obtain working cultures, two successive transfers were performed in BHI broth. Initially, an aliquot of 100 μL of the frozen culture was transferred to a tube containing 10 mL of BHI and incubated at 37 °C for 24 h. After incubation, 0.1 mL of the grown culture was transferred to another 10 mL BHI tube and incubated at the same conditions described above. The inoculum level was then adjusted to an OD 600 = 0.5 (ONDA V-10 Plus spectrophotometer CE) to obtain an initial inoculum of ~108 CFU/mL.
2.2. Essential Oil of P. longiflora
P. longiflora EO and its fraction was provided by the School of Engineering and Sciences from Tecnológico de Monterrey (México) and obtained as follows [34]: the plant was collected during the spring-summer period flowering period in northern Mexico and identified as P. longiflora by the herbarium of the Faculty of Biological Sciences of the Autonomous University of Nuevo Leon (UANL). P. longiflora EO was obtained from the leaves, flowers, and stem (ratio of 90:9:1, respectively) via steam distillation. This oil was referred to as PEO. PEO was treated by fractional distillation at 140 °C to obtain a new concentrated oil identified as FIV. PEO and FIV were characterized by Gas Chromatography (GC) (Perkin Elmer-Clarus 690) coupled to Mass Spectroscopy (MS) analysis (Perkin Elmer-Clarus SQ8T) and characterization results were reported in a previous study performed by the authors [45]. The relative area of each compound was considered as an equivalent of the composition in percentage. The major component was carvacrol with the highest concentration identified in FIV with 60.23 % followed by thymol with 21.17 %. The concentration of carvacrol in PEO was 34.09 % followed by o-Cymene with 21.51 %. However, PEO, showed a greater diversity of compounds in lower concentrations, including eucalyptol, myrcene, terpinene, humulene, and caryophyllene [45].
2.2.1. Preparation of Oil Working Solutions
Due to the hydrophobicity of the OEO, oil working solutions were prepared according to our previous data [45] for the antimicrobial screening tests. Briefly, a 1 % (v/v) of Polysorbate 80 (Tween 80) solution was prepared and sterilized. Subsequently, 400 μL of PEO and the FIV were separately mixed with 600 μL of the 1 % Tween 80 solution and stirred for 2 min to ensure dilution [46]. The resulting oil working solution was stored in the dark at refrigeration until use.
2.3. Antimicrobial Activity: Minimum Bactericidal Concentration and Sublethal Concentrations.
The MBC was determined using the microdilution method from CLSI [47] with slight modifications. A 96-well microplate (Corning, Costar; Cambridge, MA; USA) was used, with each well filled with 190 μL of BHI broth previously inoculated with L. monocytogenes (1 % v/v; 1x106 CFU/mL). Subsequently, 10 μL of PEO and FIV working solution was added to the first column of wells, followed by 10 μL dilutions across the rows. Each experiment was performed in duplicate, with two replicates per treatment (PEO, FIV, and a control without EO). After incubation at 37 °C for 24 h, 10 μL from each well was transferred to TSA plates using the drip method. The plates were incubated under the same conditions, and the MBC was identified as the lowest concentration of PEO or FIV at which no bacterial growth was observed. To estimate the Sublethal Concentrations (SLC), the same microdilution method used for MBC was used but, in this case, the OEO was diluted to concentrations between the MBC and the Minimum Inhibitory Concentration (MIC, results not shown) to achieve a reduction in L. monocytogenes without reaching a lethal effect, ensuring that the concentration remains compatible with minimal processing conditions such as the use of mild temperatures [48]. SLC was considered the maximum concentration where the OEO did not cause a decrease in the bacterial population.
2.4. Thermal Inactivation Treatments
The effect of PEO or FIV in its SLC on the thermal inactivation of L. monocytogenes was evaluated at 52, 54, 57.5 and 63 °C [1,40,49,50] in a modified Simulated Meat Medium (SMM), based on BHI broth [51]. The SMM was prepared in assay tubes containing 4 mL of broth and supplemented with glucose (18 g/L) and yeast extract (3 g/L) without modifying water activity. After sterilization of the SMM, 0.5 mL of OEO (PEO and FIV) solutions were added to reach the SLC and a control with only Tween 80 was considered (each tube represented a sample). Tubes containing SMM and OEO were placed into a hot water bath, with temperature monitored using a mercury thermometer in a control tube. When the internal temperature approached the experimental temperature (± 1.0 °C), 0.5 mL of the adjusted inoculum (108 CFU/mL) of L. monocytogenes was added to obtain a final volume of 5 mL. When the internal temperature reached the experimental temperature, samples from the three treatments (PEO, FIV and control) were taken from the tubes at appropriate intervals, ranging from 0 to 30 min, depending on the temperature evaluated. Samples at time zero did not undergo heat treatment. Samples were treated and analyzed in triplicate and the entire experiment was performed twice.
Serial dilutions of the samples were carried out in saline solution (0.85 %v/v) and aliquots of 100 μL were plated in Oxford agar petri dishes (90 x 14 mm), using a Spiral Plater (Eddy Jet 2W, IUL Instruments SA, Spain) and the plates were incubated at 37 °C for 48 h. The enumeration of the colonies (CFU/mL) per sampling time at different temperatures was performed using an automatic colony counter Flash & Go (Interscience®, IUL Instruments, Spain).
2.4.1. Estimation of the L. monocytogenes Inactivation Parameters
The Weibull-Mafart and Bigelow primary inactivation models, available in the user-friendly software Bioinactivation4 [52], were fitted to the experimental data obtained (log CFU/mL) over time (sec) for each treatment (PEO, FIV and control without EO) and temperature condition (52, 54, 57.5 and 63 °C). The Mafart model describes the time required to reduce a microbial population by one Log10 unit at a constant temperature, represented by the δ-value. This model is often combined with the Weibull model to interpret non-linear regression [53]. In contrast, the Bigelow model represents microbial inactivation at a constant temperature but follows a first-order, linear relationship [54,55]. To assess the goodness-of-fit of the primary models, several statistical indices were calculated to select the best model to fit the obtained results from the thermal inactivation of L. monocytogenes (RMSE, AIC, Bf, and Af, as described below). These indicators were particularly useful for estimating the D-values (δ value for the Mafart model), which represent the time required to achieve 1-log reduction in the L. monocytogenes population. The Bigelow primary model is described by Equation 1 [54].
where the D-value at a constant temperature T is identified as DT; N(t) is the population at time t (CFU/g); and N0 is the initial population (CFU/g).
The Weibull-Mafart model is described by the Equation 2 [53]:
where the time to reduce 90 % of the microbial population at temperature T is represented by δT (scale parameter); and p is the shape parameter (dimensionless). When p > 1, the curve is convex, when p < 1 the curve is concave and, when p = 1 the curve follows a linear trend [56].
For the secondary models, both one-step and two-step approaches were employed to estimate the z-values, which describe the temperature dependence of the inactivation process. These secondary models were fitted to the complete experimental dataset, incorporating all temperature and treatment conditions. The time-temperature data were adjusted using 57.5 °C as the reference temperature (TR), as recommended for similar studies on L. monocytogenes [40,57,58]. The z-values were calculated by fitting the secondary models to the entire dataset, ensuring that temperature variation was adequately accounted for the inactivation curves. For the Mafart model, Equation 3 describes the secondary behavior and evaluates the sensitivity of microorganisms at a certain temperature [55]:
where δR is the δ-value at the TR.
In evaluating the goodness-of-fit of each model, the Root Mean Square Error (RMSE, Equation 4) was calculated. Values lower than 0.50, indicate an acceptable fit between the model and the experimental data [56,59].
where Xobs and Xfit represent the observed and fitted values, respectively; n is the total number of observations; and s is the number of model parameters.
In the two-step inactivation model, the Residual Standard Error (RSE) is used to measure how well the residuals fit in the regression model, accounting for the degrees of freedom (df). It is calculated using the Equation 5 [60], which is closely related to RMSE, in the one-step, but is not identical:
To facilitate a direct comparison on a common scale between the RMSE and RSE in the two-step inactivation model, the RMSE is standardized (RMSEstd) by adjusting for the degrees of freedom (Equation 6). This standardization accounts for the difference in the number of parameters between the one- and two-step models, allowing for a more meaningful comparison of their goodness-of-fit [60].
The Akaike Information Criteria (AIC) related the maximum likelihood (loglik) to the estimated parameters was calculated according to Equation 7 [61].
where n is the total number of experiments, SSE is the sum of squares of errors, and s (or k) is the number of parameters in the model [61,62].
Additionally, the Bias Factor (Bf, Equation 8) and Accuracy Factor (Af, Equation 9) were assessed, with values closer to 1.0 indicating good predictive performance and minimal discrepancy between predicted and observed data [63].
where xpred represent the predicted values; Xobs represents the observed values; and n is the total number of experimental data points [62,64].
2.5. Statistical Analysis
The estimated D- and/or δ-values were analyzed using the statistical program OpenStat, comparing means across different temperatures for the same treatment and across different treatments at the same temperature, with P < 0.05 set as the level of significance.
3. Results and Discussion
Fractionation has been shown to enhance the activity of EOs by exerting bactericidal effects and interfering with microbial resistance and virulence mechanisms, providing a multifaceted approach to microbial control [65]. The MBC was considered as the lowest concentration at which no microbial growth was observed across all replicates and repetitions. The MBC against L. monocytogenes obtained when using PEO was 3.10 ± 0.00 µg/mL, while the MBC with FIV was significantly lower (0.02 ± 0.00 µg/mL, P < 0.05). To allow a more viable result and use of the OEO in the thermal treatments, a SLC test was performed and considered as the lowest concentration of EO where the bacterial growth was not affected. In this way, the SLC of PEO was 0.13 ± 0.00 % and 0.09 ± 0.05 % for FIV.
For setting the concentration for thermal treatments, 0.06 % of PEO and FIV were used because of the rapid inactivation that made colony counting difficult when using the SLC value (data not shown). OEO has promising antibacterial properties, but its effectiveness can be influenced by factors such as bacterial strain and oil composition. Guo et al. [66] determined the Minimum Inhibitory Concentration (MIC) of OEO (does not specify the oregano species used) against L. monocytogenes to be 0.03 % (v/v), then treated L. monocytogenes with approximately 0.05 % OEO for 30 minutes, resulting in around 6-log reductions of the bacterial count. In other investigation, Bakkali et al. [67] assessed the MBC of Origanum compactum (68.99 % of carvacrol) against E. coli, reporting an MBC of 0.63 % (v/v) and a MIC of 0.31 %.
Figure 1 presents the inactivation curves of L. monocytogenes (log N/N0 versus time) at 52, 54, 57.5 and 63 °C in the SMM. A more pronounced initial reduction can be observed in the treatments with OEO compared to the control group. The Weibull-Mafart and Bigelow inactivation models were compared for their suitability for describing the observed data (Table 1). The models were fitted to the inactivation data for the selection of the best model in estimating the D-value. Both models showed low RMSE; however, all the goodness-of-fit indices evaluated indicated better performance for the Weibull-Mafart model. Specifically, RMSE and SER showed lower values while Af and Bf were closer to one. Therefore, the Weibull-Mafart model was chosen in this study as the most suitable for describing the observed data.
The Weibull-Mafart model allows to estimate the delta (ẟ) value which represents the time to achieve one logarithm reduction of the microbial population under a specific temperature, which is equivalent to the D value (min) [53]. Table 2 shows the estimations of ẟ (min) of L. monocytogenes for each group at the four temperatures evaluated. While the temperature increases, it is observed that in the three groups, ẟ decreases. In all cases, ẟ-values using FIV were significantly lower (P < 0.05) than the control. Only at 52 and 63 °C there was a significant difference (P < 0.05) between PEO and control, while at 54 and 57.5 °C significant differences (P < 0.05) were observed between PEO and FIV.
The estimated Weibull-Mafart model p parameter values were lower than 1 (Table 2) for all the treatments, indicating a concave shape of the curves (Figure 1). As inactivation time was extended, a tail appeared in the curve, suggesting an adaptation of the microorganism to the environment (EO and medium) and/or the presence of a more susceptible subpopulation, within the L. monocytogenes population (Figure 1) [68].
Considering the goodness-of-fit indices for both primary and secondary models, the One-Step procedure was selected to fit the model to the experimental data to estimate the z-values (Table 3). The resulting secondary models are shown in Figure 2. This One-Step model fitting were carried out using the bionactivation4 shiny and verified using the negative inverse of the slope of each line in MS Excel. The z-values estimated were 5.75, 5.20 and 5.00 °C for Control, PEO and FIV, respectively, where no significant differences were observed between the groups (P > 0.05). Similar z-values around 5 °C were obtained in previous studies of the use of OEO against L. monocytogenes species in seafood [40,69,70]. These previous results, along with those obtained in this study for the z- and δ-values are consistent with the thermal resistance profile and susceptibility to the applied antimicrobial agents. These results indicate that while L. monocytogenes maintains a consistent sensitivity to temperature, its susceptibility differs depending on the antimicrobial agent applied [39].
Table 4 summarizes the findings from previous studies on thermal inactivation of L. monocytogenes at temperatures ranging from 52 to 65 °C, using EOs, phenolic compounds and plant extracts as natural antimicrobial agents. Notably, at 55 °C, the D value varied depending on the food matrix, highlighting the influence of the matrix on the effectiveness of thermal treatments. The lowest D values were observed with treatments containing isolated EO compounds, such as carvacrol, thymol, and cinnamaldehyde, indicating their strong antimicrobial properties. These results aligned with those obtained in our study using FIV, where the higher concentration of active compounds and reduced interference from EO components with lower antimicrobial activity contributed to more effective bacterial inactivation.
Incorporating FIV and PEO into food processing could provide an effective strategy to control L. monocytogenes, especially in Ready-to-Eat (RTE) foods where the pathogen's ability to grow at refrigeration temperatures poses a challenge [78]. Furthermore, FIV offers the potential to enhance antimicrobial efficacy while reducing the sensory impact on food products, a critical consideration for maintaining consumer acceptability [37]. By leveraging predictive microbiology models, the thermal and antimicrobial effects of FIV can be optimized, allowing for evidence-based decisions that balance food safety and quality. This approach represents an opportunity to minimize heat treatments while ensuring safe food products, particularly in regions with limited regulatory oversight for listeriosis [79]. Moreover, integrating these predictive models into user-friendly tools could facilitate widespread adoption, enabling food processors and regulators to make informed decisions with greater ease and efficiency.
4. Conclusions
P. longiflora fractionated oregano essential oil significantly reduced the time for achieving L. monocytogenes logarithmic reductions under heat treatments at all the mild temperatures applied in this study. This effect is related to the high concentration of carvacrol, and thymol reached by the fractional distillation at 140 °C. The inactivation parameters estimated in our study through model fitting can be used to assess suitable oregano essential oil and mild heat treatments to achieve desired levels of pathogen inactivation. The thermal inactivation experiments of L. monocytogenes in a food matrix using ground meat showed results consistent with those obtained in the meat simulation medium, suggesting that Mexican P. longiflora fractionated essential oil has strong potential for application in this food matrix. Further investigations on the sensory effects of this application are underway.
Author Contributions
Investigation, and writing—original draft preparation M.P-G; Conceptualization and supervision A.V. and S.C; Methodology writing—review and editing, A.P. and E.S-G; Resources, J.R-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), now Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI), grant number 1184786.
Data Availability Statement
Dataset available on request from the authors. The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
To Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), now Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI), for supporting the studies during which this project was carried out (CVU number: 1184786) at the Department of Food Sciences, Universidad Autónoma de Nuevo León, México. We also thank FUNDACIÓN UANL for funding a research stay at the Department of Food Science and Technology, Universidad de Córdoba, Spain, from January to July 2023.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
| °C | Degree Celsius |
| µL | microliter |
| Af | Accuracy factor |
| AIC | Akaike Information Criterion |
| Bf | Bias factor |
| BHI | Brain Heart Infusion |
| CFU/mL | Colony Forming Units per milliliter |
| EO | Essential Oils |
| FIV | Fractionated Oregano Essential Oil |
| g/L | grams per liter |
| GRAS | Generally Recognized As Safe |
| h | hours |
| HPP | High Pressure Processing |
| MBC | Minimum Bactericidal Concentration |
| min | minutes |
| mL | milliliter |
| OD | Optical Density |
| OEO | Oregano Essential Oil |
| PEO | Pure Oregano Essential Oil |
| RMSE | Root Mean Square Error |
| sec | seconds |
| SER | Standard Error of Regression |
| v/v | volume/volume |
References
- Food and Drug Administration. Hazard Analysis and Risk-Based Preventive Controls for Human Food: Draft Guidance for Industry 2024. Available online: https://www.regulations.gov/document/FDA-2016-D-2343-0092 (accessed on 16 April 2025).
- Food and Drug Administration. Bad Bug Book-Foodborne Pathogenic Microorganisms and Natural Toxins-Second Edition 2 Bad Bug Book Handbook of Foodborne Pathogenic Microorganisms and Natural Toxins, Washington DC 2022; pp. 99-102.
- Ricci, A., Allende, A., Bolton, D., Chemaly, M., Davies, R., Fernández Escámez, P. S., Girones, R., Herman, L., Koutsoumanis, K., Nørrung, B., Robertson, L., Ru, G., Sanaa, M., Simmons, M., Skandamis, P., Snary, E., Speybroeck, N., Ter Kuile, B., Threlfall, J., … Lindqvist, R. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU 2018. EFSA Journal, 16(1):5134. [CrossRef]
- Koutsoumanis, K., Allende, A., Bolton, D., Bover-Cid, S., Chemaly, M., De Cesare, A., Herman, L., Hilbert, F., Lindqvist, R., Nauta, M., Nonno, R., Peixe, L., Ru, G., Simmons, M., Skandamis, P., Suffredini, E., Fox, E., Gosling, R., Gil, B. M., Alvarez-Ordóñez, A. Persistence of microbiological hazards in food and feed production and processing environments 2024. EFSA Journal, 22(1):e8521. [CrossRef]
- CDC. Listeria Outbreak Linked to Leafy Greens – February 2023. Available online: https://www.cdc.gov/listeria/outbreaks/monocytogenes-02-23/index.html (accessed on 09 march 2025).
- CDC. Listeria Outbreak Linked to Ice Cream – August 2023. Available online: https://www.cdc.gov/listeria/outbreaks/ice-cream-08-23/index.html (accessed on 09 march 2025).
- CDC. Listeria Outbreak Linked to Queso Fresco and Cotija Cheese – February 2024. Available online: https://www.cdc.gov/listeria/outbreaks/cheese-02-24/index.html (accessed on 09 march 2025).
- CDC. Listeria Outbreak Linked to Ready-To-Eat Meat and Poultry Products. Available online: https://www.cdc.gov/listeria/outbreaks/meat-and-poultry-products-11-24/index.html (accessed on 09 march 2025).
- CDC. More Illnesses and Deaths in Listeria Outbreak Linked to Deli Meats is Reminder to Avoid Recalled Products 2024. Available online: https://www.cdc.gov/media/releases/2024/s0828-listeria-outbreak-deli-meats.html (accessed on 09 march 2025).
- CDC. Outbreak Investigation of Listeria monocytogenes: Frozen Supplemental Shakes (February 2025). Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/outbreak-investigation-listeria-monocytogenes-frozen-supplemental-shakes-february-2025 (accessed on 09 march 2025).
- Guel-García, P., García De León, F. J., Aguilera-Arreola, G., Mandujano, A., Mireles-Martínez, M., Oliva-Hernández, A., Cruz-Hernández, M. A., Vásquez-Villanueva, J., Rivera, G., Bocanegra-García, V., & Martínez-Vázquez, A. V. Prevalence and Antimicrobial Resistance of Listeria monocytogenes in Different Raw Food from Reynosa, Tamaulipas, Mexico, Foods 2024, 13(11). [CrossRef]
- Matle, I., Mbatha, K. R., & Madoroba, E.. A review of listeria monocytogenes from meat and meat products: Epidemiology, virulence factors, antimicrobial resistance, and diagnosis. Onderstepoort Journal of Veterinary Research 2020, 87(1), 1–20. [CrossRef]
- Hong, H., Yang, S. M., Kim, E., Kim, H. J., & Park, S. H. Comprehensive metagenomic analysis of stress-resistant and -sensitive Listeria monocytogenes. Applied Microbiology and Biotechnology 2023, 107(19), 6047–6056. [CrossRef]
- Labidi, S., Jánosity, A., Yakdhane, A., Yakdhane, E., Surányi, B., Mohácsi-Farkas, C., & Kiskó, G. Effects of pH, sodium chloride, and temperature on the growth of Listeria monocytogenes biofilms. Acta Alimentaria 2023, 52(2), 270–280. [CrossRef]
- Mahgoub, S. A., El-Mekkawy, R. M., Abd El-Hack, M. E., El-Ghareeb, W. R., Suliman, G. M., Alowaimer, A. N., & Swelum, A. A. Inactivation of Listeria monocytogenes in ready-to-eat smoked turkey meat by combination with packaging atmosphere, oregano essential oil and cold temperature. AMB Express 2019, 9(1). [CrossRef]
- Magalhães, R., Ferreira, V., Brandão, T. R. S., Palencia, R. C., Almeida, G., & Teixeira, P. Persistent and non-persistent strains of Listeria monocytogenes: A focus on growth kinetics under different temperature, salt, and pH conditions and their sensitivity to sanitizers. Food Microbiology 2016, 57, 103–108. [CrossRef]
- Yue, S., Liu, Y., Wang, X., Xu, D., Qiu, J., Liu, Q., & Dong, Q. Modeling the effects of the preculture temperature on the lag phase of Listeria monocytogenes at 25°c. Journal of Food Protection 2019, 82(12), 2100–2107. [CrossRef]
- Wei, J., Ismael, M., Huang, M., Han, T., & Zhong, Q. The bactericidal effects of combined sterilization methods on Listeria monocytogenes and the application in prepared salads, Elsevier 2025 preprint article: https://ssrn.com/abstract=5102418.
- Ghabraie, M., Vu, K. D., Huq, T., Khan, A., & Lacroix, M.. Antilisterial effects of antibacterial formulations containing essential oils, nisin, nitrite and organic acid salts in a sausage model. Journal of Food Science and Technology 2016. 53, 2625–2633. [CrossRef]
- Mani-López, E., García, H. S., & López-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Research International 2012, 45(2), 713–721. [CrossRef]
- Puvača, N., Milenković, J., Galonja Coghill, T., Bursić, V., Petrović, A., Tanasković, S., Pelić, M., Ljubojević Pelić, D., & Miljković, T. Antimicrobial activity of selected essential oils against selected pathogenic bacteria: In vitro study. Antibiotics 2021. 10(5). [CrossRef]
- Targino de Souza Pedrosa, G., Pimentel, T. C., Gavahian, M., Lucena de Medeiros, L., Pagán, R., & Magnani, M. The combined effect of essential oils and emerging technologies on food safety and quality. LWT 2021. 147. [CrossRef]
- Vidaković Knežević S., Knežević S., Vranešević J., Kravić S., Lakićević B., Kocić-Tanackov S., Karabasil N. Effects of Selected Essential Oils on Listeria monocytogenes in Biofilms and in a Model Food System. Foods 2023. May 1;12(10). [CrossRef]
- Tejada-Muñoz, S., Cortez, D., Rascón, J., Chavez, S. G., Caetano, A. C., Díaz-Manchay, R. J., Sandoval-Bances, J., Huyhua-Gutierrez, S., Gonzales, L., Chenet, S. M., & Tapia-Limonchi, R. Antimicrobial Activity of Origanum vulgare Essential Oil against Staphylococcus aureus and Escherichia coli. Pharmaceuticals 2024, 17(11). [CrossRef]
- Pinto, L., Cervellieri, S., Netti, T., Lippolis, V., & Baruzzi, F. Antibacterial Activity of Oregano (Origanum vulgare L.) Essential Oil Vapors against Microbial Contaminants of Food-Contact Surfaces. Antibiotics 2024, 13(4). [CrossRef]
- Maggio, F., Rossi, C., Chaves-Lópe, C., Valbonetti, L., Desideri, G., Paparella, A., & Serio, A. A single exposure to a sublethal concentration of Origanum vulgare essential oil initiates response against food stressors and restoration of antibiotic susceptibility in Listeria monocytogenes. Food Control 2022, 132. [CrossRef]
- Cortés Sánchez, A. D. J., Guzmán Robles, M. L., Garza Torres, R., Espinosa Chaurand, L. D., & Diaz Ramírez, M. Food Safety, Fish and Listeriosis. Turkish Journal of Agriculture - Food Science and Technology 2019, 7(11), 1908–1916. [CrossRef]
- Cabrera-Díaz, E., Martínez-Chávez, L., Gutiérrez-González, P., Pérez-Montaño, J. A., Rodríguez-García, M. O., & Martínez-Gonzáles, N. E. Effect of storage temperature and time on the behavior of Salmonella, Listeria monocytogenes, and background microbiota on whole fresh avocados (Persea americana var Hass). International Journal of Food Microbiology 2022, 369. [CrossRef]
- Márquez-González, M., Osorio, L. F., Velásquez-Moreno, C. G., & García-Lira, A. G. Thermal Inactivation of Salmonella enterica and Listeria monocytogenes in Quesillo Manufactured from Raw Milk. International Journal of Food Science 2022. [CrossRef]
- Rubio Lozano, S. M., Martínez Bruno, F. J., Hernández Castro, R., Bonilla Contreras, C., Danilo Méndez Medina, R., Núñez Espinosa, F. J., Echeverry, A. Detection of Listeria monocytogenes, Salmonella and Yersinia enterocolitica in beef at points of sale in Mexico. In RES Rev Mex Cienc Pecu 2013. 4(1). pp. 107-115.
- Rubio Ortega A, Guinoiseau E, Poli JP, Quilichini Y, de Rocca Serra D, del Carmen Travieso Novelles M, Espinosa Castaño I., Pino Pérez O., Berti L. The Primary Mode of Action of Lippia graveolens Essential Oil on Salmonella enterica subsp. Enterica Serovar Typhimurium. Microorganisms. 2023 Dec 1;11(12). [CrossRef]
- Ortega-Nieblas M., Robles-Burgueño M., Acedo-Félix E., González-León A., Morales-Trejo A., Vázquez-Moreno L. Chemical Composition and Antimicrobial Activity of Oregano (Lippia palmeri S. Wats) Essential Oil. Rev Fitotec Nex. 2011; 34 (1); 11-17.
- Mora-Zúñiga, A. E., Treviño-Garza, M. Z., Amaya Guerra, C. A., Galindo Rodríguez, S. A., Castillo, S., Martínez-Rojas, E., Rodríguez-Rodríguez, J., & Báez-González, J. G. Comparison of Chemical Composition, Physicochemical Parameters, and Antioxidant and Antibacterial Activity of the Essential Oil of Cultivated and Wild Mexican Oregano Poliomintha longiflora Gray. Plants 2022, 11(14). [CrossRef]
- de Rostro-Alanis, M. J., Báez-González, J., Torres-Alvarez, C., Parra-Saldívar, R., Rodriguez-Rodriguez, J., & Castillo, S. Chemical composition and biological activities of oregano essential oil and its fractions obtained by vacuum distillation. Molecules 2019. 24(10). [CrossRef]
- Levario-Gómez A, Ávila-Sosa R, Gutiérrez-Méndez N, López-Malo A, Nevárez-Moorillón GV. Modeling the Combined Effect of pH, Protein Content, and Mexican Oregano Essential Oil Against Food Spoilage Molds. Front Sustain Food Syst. 2020 4:34. [CrossRef]
- Zapién-Chavarría, K. A., Plascencia-Terrazas, A., Venegas-Ortega, M. G., Varillas-Torres, M., Rivera-Chavira, B. E., Adame-Gallegos, J. R., González-Rangel, M. O., & Nevárez-Moorillón, G. V. Susceptibility of multidrug-resistant and biofilm-forming uropathogens to Mexican oregano essential oil. Antibiotics 2019, 8(4). [CrossRef]
- Ruiz-Hernández, K., Sosa-Morales, M. E., Cerón-García, A., & Gómez-Salazar, J. A. Physical, Chemical and Sensory Changes in Meat and Meat Products Induced by the Addition of Essential Oils: A Concise Review. In Food Reviews International 2021. 39(4), 2027–2056. Taylor and Francis Ltd. [CrossRef]
- Agregán, R., Munekata, P. E. S., Zhang, W., Zhang, J., Pérez-Santaescolástica, C., & Lorenzo, J. M. High-pressure processing in inactivation of Salmonella spp. in food products. In Trends in Food Science and Technology 2021. 107, 31–37. [CrossRef]
- Soni, A., Bremer, P., & Brightwell, G. A Comprehensive Review of Variability in the Thermal Resistance (D-Values) of Food-Borne Pathogens—A Challenge for Thermal Validation Trials. Foods 2022. 11(24). MDPI. [CrossRef]
- Dogruyol, H., Mol, S., & Cosansu, S.. Increased thermal sensitivity of Listeria monocytogenes in sous-vide salmon by oregano essential oil and citric acid. Food Microbiology 2020. 90. [CrossRef]
- Juneja, V. K., Huang, L., & Yan, X. Thermal inactivation of foodborne pathogens and the USDA pathogen modeling program. Journal of Thermal Analysis and Calorimetry 2011, 106(1), 191–198. [CrossRef]
- Siroli, L., Patrignani, F., Gardini, F., & Lanciotti, R. Effects of sub-lethal concentrations of thyme and oregano essential oils, carvacrol, thymol, citral and trans-2-hexenal on membrane fatty acid composition and volatile molecule profile of Listeria monocytogenes, Escherichia coli and Salmonella enteritidis. Food Chemistry 2015, 182, 185–192. [CrossRef]
- Stavropoulou, E., & Bezirtzoglou, E. Predictive modeling of microbial behavior in food. Foods 2019. 8(12). MDPI. [CrossRef]
- Pérez-Rodríguez, F. & Valero, A. Application of Predictive Models in Quantitative Risk Assessment and Risk Management. In: Harter RW, ed. Predictive Microbiology in Foods. Springer New York, Heidelberg, Dordrecht, London 2013. p.87-97.
- Sánchez García, E.; Torres-Alvarez, C.; Morales Sosa, E.G.; Pimentel-González, M.; Villarreal Treviño, L.; Amaya Guerra, C.A.; Castillo, S. & Rodríguez Rodríguez, J. Essential Oil of Fractionated Oregano as Motility Inhibitor of Bacteria Associated with Urinary Tract Infections. Antibiotics 2024, 13(7), 665. [CrossRef]
- Possas A, Posada-Izquierdo GD, Pérez-Rodríguez F, Valero A, García-Gimeno RM, Duarte MCT. Application of predictive models to assess the influence of thyme essential oil on Salmonella Enteritidis behaviour during shelf life of ready-to-eat turkey products. Int J Food Microbiol. 2017 Jan 2;240:40–6. [CrossRef]
- Lazou, T.P. & Chaintoutis, S.C. Comparison of disk diffusion and broth microdilution methods for antimicrobial susceptibility testing of Campylobacter isolates of meat origin. J. Microbiol. Methods 2023, 204, 106649. [CrossRef]
- Castillo, S., Dávila-Aviña, J., Heredia, N., & Garcia, S. Antioxidant activity and influence of Citrus byproduct extracts on adherence and invasion of Campylobacter jejuni and on the relative expression of cadF and ciaB. Food Science and Biotechnology 2017. 26(2), 453–459. [CrossRef]
- Miller, F. A., Gil, M. M., Brandão, T. R. S., Teixeira, P., & Silva, C. L. M. Sigmoidal thermal inactivation kinetics of Listeria innocua in broth: Influence of strain and growth phase. Food Control 2009, 20(12), 1151–1157. [CrossRef]
- Garre, A., González-Tejedor, G. A., Aznar, A., Fernández, P. S., & Egea, J. A. Mathematical modelling of the stress resistance induced in Listeria monocytogenes during dynamic, mild heat treatments. Food Microbiology 2019. 84. [CrossRef]
- Possas, A., Pérez-Rodríguez, F., Valero, A., Rincón, F., & García-Gimeno, R. M. Mathematical approach for the Listeria monocytogenes inactivation during high hydrostatic pressure processing of a simulated meat medium. Innovative Food Science and Emerging Technologies 2018. 47, 271–278. [CrossRef]
- Garre, A. biogrowth & bioinactivation-making predictive microbiology a bit easier IAFP Software Fair Series 2022.
- Albert, I., Mafart, P. A modified Weibull model for bacterial inactivation. International Journal of Food Microbiology 2005. 100, 197-211. [CrossRef]
- Bigelow, W. D. The Logarithmic Nature of Thermal Death Time Curves. In Jour. Infec. Dis. 1921, 29, 528-536.
- Huertas, J.-P., Ros-Chumillas, M., Garre, A., Fernández, P. S., Aznar, A., Iguaz, A., Esnoz, A., & Palop, A. Impact of Heating Rates on Alicyclobacillus acidoterrestris Heat Resistance under Non-Isothermal Treatments and Use of Mathematical Modelling to Optimize Orange Juice Processing. Foods 2021.10, 1496. [CrossRef]
- Buzrul S. The Weibull Model for Microbial Inactivation. Vol. 14, Food Engineering Reviews. Springer; 2022. p. 45–61.
- Valenzuela-Melendres M, Peña-Ramos EA, Juneja VK, Camou JP, Cumplido-Barbeitia G. Effect of grapefruit seed extract on thermal inactivation of Listeria monocytogenes during sous-vide processing of two marinated Mexican meat entrées. J Food Prot. 2016 Jul 1;79(7):1174–80.
- Zakrzewski A, Gajewska J, Chajęcka-Wierzchowska W, Zadernowska A. Effect of sous-vide processing of fish on the virulence and antibiotic resistance of Listeria monocytogenes. NFS Journal. 2023 Jun 1;31:155–61.
- Salazar JK, Fay ML, Fleischman G, Khouja BA, Stewart DS, Ingram DT. Inactivation kinetics of Listeria monocytogenes and Salmonella enterica on specialty mushroom garnishes based on ramen soup broth temperature. Front Microbiol. 2024;15.
- Kutner, M. H. ., Nachtsheim, Chris., Neter, John., & Li, William. Applied linear statistical models; 5th edition; McGraw-Hill Irwin, New York, 2005. pp. 15-23.
- Portet, S. A primer on model selection using the Akaike Information Criterion. Infectious Disease Modelling 2020, 5, 111–128. [CrossRef]
- Tarlak, F. The Use of Predictive Microbiology for the Prediction of the Shelf Life of Food Products. Foods 2023, Vol. 12, Issue 24. [CrossRef]
- Pouillot, R., Kiermeier, A., Guillier, L., Cadavez, V., & Sanaa, M. Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer. Foods 2024, 13(5). [CrossRef]
- Milkievicz, T., Badia, V., Souza, V. B., Longhi, D. A., Galvão, A. C., & da Silva Robazza, W. Development of a general model to describe Salmonella spp. growth in chicken meat subjected to different temperature profiles. Food Control 2020, 112. [CrossRef]
- Dudek-Wicher, R., Paleczny, J., Kowalska-Krochmal, B., Szymczyk-Ziółkowska, P., Pachura, N., Szumny, A., & Brożyna, M. Activity of liquid and volatile fractions of essential oils against biofilm formed by selected reference strains on polystyrene and hydroxyapatite surfaces. Pathogens 2021, 10(5). [CrossRef]
- Guo, P., Li, Z., Cai, T., Guo, D., Yang, B., Zhang, C., Shan, Z., Wang, X., Peng, X., Liu, G., Shi, C., Alharbi, M., & Alasmari, A. F. Inhibitory effect and mechanism of oregano essential oil on Listeria monocytogenes cells, toxins, and biofilms. Microbial Pathogenesis 2024, 194. [CrossRef]
- Bakkali M., Arakrak A., Laglaoui A., D. Evaluation of the Antibacterial Activity of Essential Oils Against E. coli Isolated From Rabbits. In Iraqi Journal of Agricultural Sciences 2022. 2022:53 (4), 802-809.
- Fang, T., Wu, Y., Xie, Y., Sun, L., Qin, X., Liu, Y., Li, H., Dong, Q., & Wang, X. Inactivation and Subsequent Growth Kinetics of Listeria monocytogenes After Various Mild Bactericidal Treatments. Frontiers in Microbiology 2021. 12. [CrossRef]
- Shi Y, Tang J, Yue T, Rasco B, Wang S. Pasteurizing Cold Smoked Salmon (Oncorhynchus nerka): Thermal Inactivation Kinetics of Listeria monocytogenes and Listeria innocua. Journal of Aquatic Food Product Technology. 2015 Oct 3;24(7):712–22.
- Li C, Huang L, Hwang CA. Effect of temperature and salt on thermal inactivation of Listeria monocytogenes in salmon roe. Food Control. 2017 Mar 1;73:406–10.
- Moura-Alves M, Gouveia AR, de Almeida JMMM, Monteiro-Silva F, Silva JA, Saraiva C. Behavior of Listeria monocytogenes in beef Sous vide cooking with Salvia officinalis L. essential oil, during storage at different temperatures. LWT. 2020 Oct 1;132.
- Wang Y, Li X, Lu Y, Wang J, Suo B. Synergistic effect of cinnamaldehyde on the thermal inactivation of Listeria monocytogenes in ground pork. Food Science and Technology International. 2020 Jan 1;26(1):28–37.
- Kamdem SS, Belletti N, Magnani R, Lanciotti R, Gardini F. Effects of carvacrol, (E)-2-hexenal, and citral on the thermal death kinetics of Listeria monocytogenes. J Food Prot. 2011 Dec;74(12):2070–8.
- Guevara L, Antolinos V, Palop A, Periago PM. Impact of Moderate Heat, Carvacrol, and Thymol Treatments on the Viability, Injury, and Stress Response of Listeria monocytogenes. Biomed Res Int. 2015. [CrossRef]
- Arioli S, Montanari C, Magnani M, Tabanelli G, Patrignani F, Lanciotti R, Mora D., Gardini F. Modelling of Listeria monocytogenes Scott A after a mild heat treatment in the presence of thymol and carvacrol: Effects on culturability and viability; Journal of Food Engineering. Elsevier Ltd; 2019. Vol. 240. p. 73–82.
- Essia Ngang JJ, Nyegue MA, Ndoye FC, Tchuenchieu Kamgain AD, Sado Kamdem SL, Lanciotti R, Gardini F., Etoa F-X. Characterization of Mexican coriander (Eryngium foetidum) essential oil and its inactivation of Listeria monocytogenes in vitro and during mild thermal pasteurization of pineapple juice. J Food Prot. 2014 Mar;77(3):435–43.
- Juneja VK, Garcia-Dávila J, Lopez-Romero JC, Pena-Ramos EA, Camou JP, Valenzuela-Melendres M. Modeling the effects of temperature, sodium chloride, and green tea and their interactions on the thermal inactivation of Listeria monocytogenes in Turkey. J Food Prot. 2014 Oct 1;77(10):1696–702.
- Butler, F., Hunt, K., Redmond, G., dOnofrio, F., Barron, U. G., Fernandes, S., Cadavez, V., Iannetti, L., Centorotola, G., Pomilio, F., Diaz, A. V., Rodríguez, F. P., & Luque, O. M. B. Application of novel predictive microbiology techniques to shelf-life studies on Listeria monocytogenes in ready-to-eat foods (ListeriaPredict). EFSA Supporting Publications 2023. 20(12). [CrossRef]
- Contreras-Soto, M., Medrano-Félix, J., Ibarra-Rodríguez, J., Martínez-Urtaza, J., Chaidez, Q., & Castro-del Campo, N. The last 50 years of Salmonella in Mexico: Sources of isolation and factors that influence its prevalence and diversity. Bio Ciencias 2019. 6(SPE). [CrossRef]
Figure 1.
Thermal inactivation curves In vitro of L. monocytogenes at 52, 54, 57.5 and 63 °C exposed to the PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil) and Control groups. The data points represent two independent replicates.
Figure 1.
Thermal inactivation curves In vitro of L. monocytogenes at 52, 54, 57.5 and 63 °C exposed to the PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil) and Control groups. The data points represent two independent replicates.
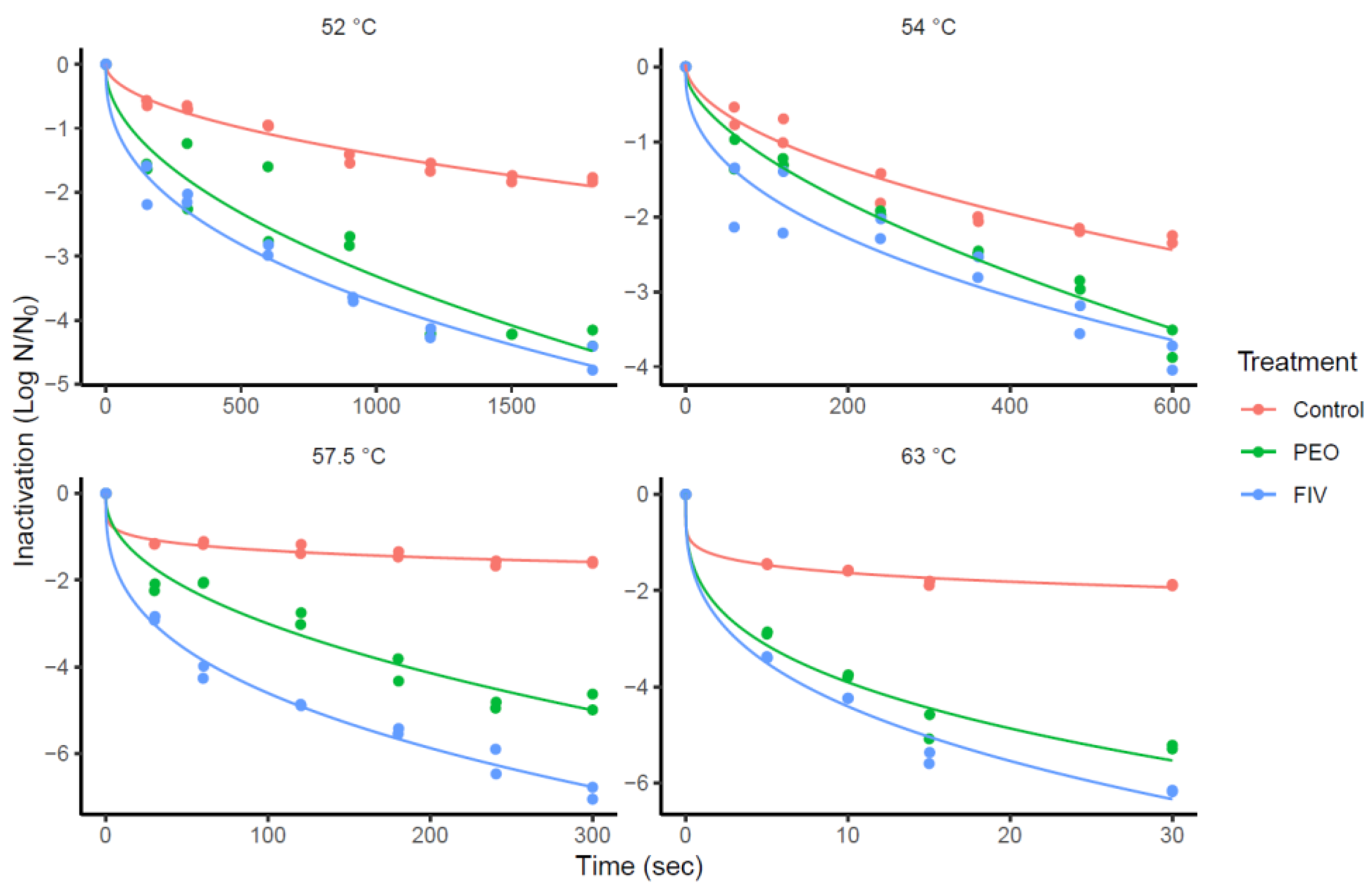
Figure 2.
Relationship between the δ parameter estimated for L. monocytogenes with the mild heat treatment temperature applied for the Control group, PEO (Pure Oregano Essential Oil) and FIV (Fractionated Oregano Essential Oil) groups. Grey zones represent the confidence intervals of parameter estimates.
Figure 2.
Relationship between the δ parameter estimated for L. monocytogenes with the mild heat treatment temperature applied for the Control group, PEO (Pure Oregano Essential Oil) and FIV (Fractionated Oregano Essential Oil) groups. Grey zones represent the confidence intervals of parameter estimates.
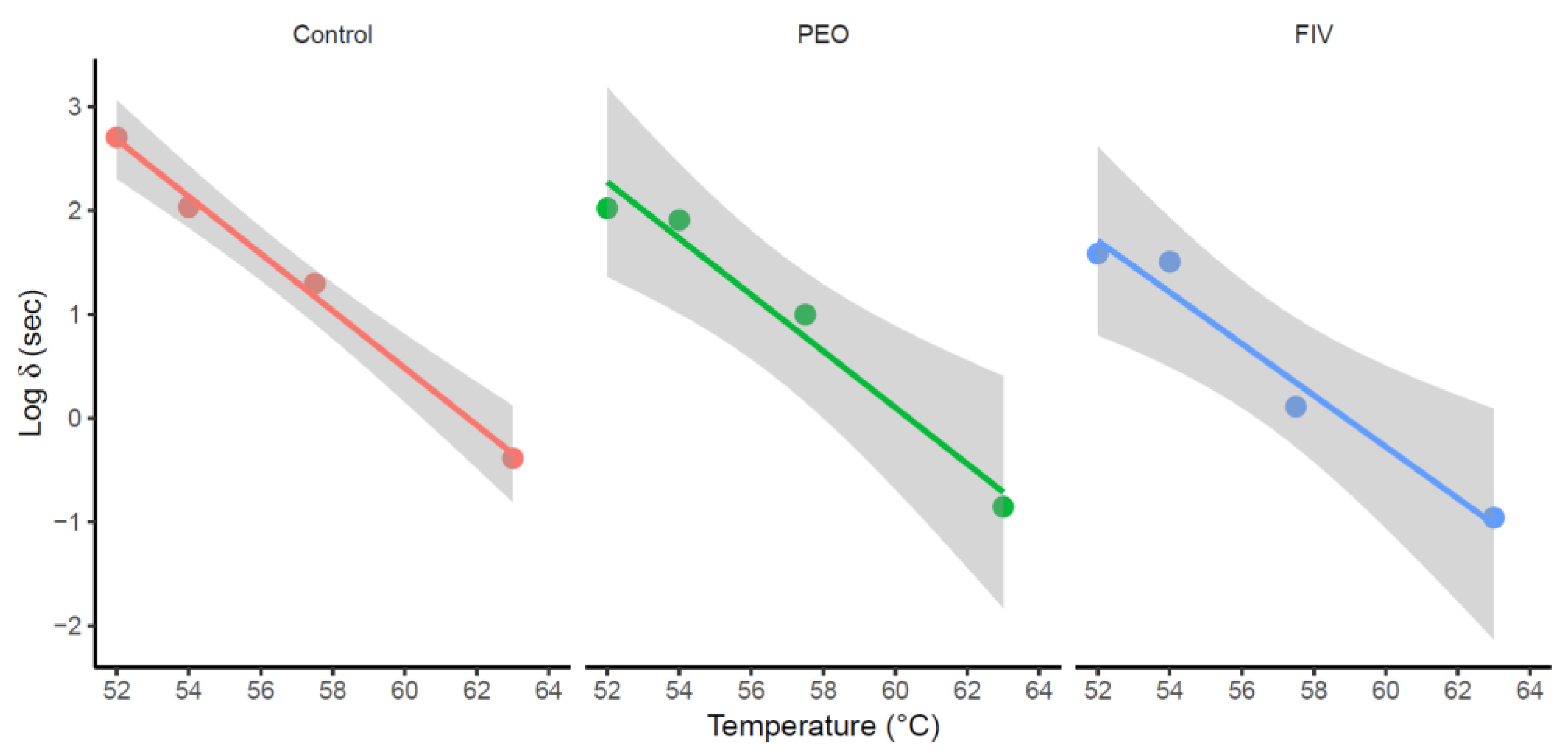

Table 1.
Goodness-of-fit indices estimated by fitting the primary models Weibull-Mafart and Bigelow to the inactivation data: RMSE (Root Mean Square Error), Loglik (Log likelihood), AIC (Akaike Information Criteria), Af (Accuracy factor) and Bf (Bias factor).
Table 1.
Goodness-of-fit indices estimated by fitting the primary models Weibull-Mafart and Bigelow to the inactivation data: RMSE (Root Mean Square Error), Loglik (Log likelihood), AIC (Akaike Information Criteria), Af (Accuracy factor) and Bf (Bias factor).
| Weibull-Mafart | Bigelow | ||||||||
| Group | T (°C) | RMSE | Loglik | AIC | Af /Bf | RMSE | Loglik | AIC | Af/Bf |
| Control | 52 | 0.10 | 42.70 | -81.40 | 1.01/0.99 | 0.20 | 22.36 | -42.72 | 1.02/1.00 |
| 54 | 0.16 | 24.40 | -44.80 | 1.02/1.00 | 0.29 | 13.30 | -24.60 | 1.04/1.00 | |
| 57.5 | 0.08 | 47.44 | -90.88 | 1.01/1.00 | 0.37 | 11.92 | -21.84 | 1.04/1.00 | |
| 63 | 0.06 | 46.21 | -88.42 | 1.01/0.99 | 0.48 | 5.35 | -8.70 | 1.07/1.00 | |
| PEO | 52 | 0.42 | 10.36 | -16.72 | 1.07/1.00 | 0.55 | 7.57 | -13.14 | 1.09/1.00 |
| 54 | 0.18 | 22.73 | -41.46 | 1.02/1.00 | 0.30 | 14.33 | -26.66 | 1.05/1.01 | |
| 57.5 | 0.33 | 10.96 | -17.92 | 1.07/1.00 | 0.62 | 6.47 | -10.94 | 1.45/1.42 | |
| FIV | 63 | 0.34 | 11.06 | -18.12 | 1.06/1.00 | 1.00 | 2.52 | -3.04 | 1.26/0.99 |
| 52 | 0.20 | 19.14 | -34.28 | 1.06/1.03 | 0.61 | 6.22 | -10.44 | 1.12/1.02 | |
| 54 | 0.33 | 11.55 | -19.10 | 1.05/1.01 | 0.51 | 8.35 | -14.70 | 1.06/1.00 | |
| 57.5 | 0.20 | 19.50 | -35.00 | 1.10/0.98 | 1.04 | 4.14 | -6.28 | 1.30/0.94 | |
| 63 | 0.24 | 11.94 | -19.88 | 0.99/1.07 | 1.11 | 2.28 | -2.56 | 1.39/0.95 | |
Table 2.
Weibull-Mafart model parameters estimated by fitting the inactivation data obtained at 52, 54, 57.5, and 63 °C for the PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil), and Control groups.
Table 2.
Weibull-Mafart model parameters estimated by fitting the inactivation data obtained at 52, 54, 57.5, and 63 °C for the PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil), and Control groups.
| Control | PEO | FIV | ||||
| Temp (°C) | δ(min) | p | δ(min) | p | δ(min) | p |
| 52 | 8.470 ± 1.510 | 0.51 | 1.750 ± 1.050 | 0.52 | 0.640 ± 0.240 | 0.40 |
| 54 | 1.800 ± 0.500 | 0.53 | 1.350 ± 0.350 | 0.61 | 0.540 ± 0.350 | 0.44 |
| 57.5 | 0.330 ± 0.170 | 0.17 | 0.170 ± 0.090 | 0.47 | 0.020 ± 0.010 | 0.35 |
| 63 | 0.007 ± 0.004 | 0.15 | 0.002 ± 0.001 | 0.32 | 0.002 ± 0.002 | 0.33 |
Table 3.
Z-values (°C) estimated by the One-Step and Two-Step procedures and the Root Mean Square Error (RMSE), Residual Standard Error (RSE) and Standardized RMSE (RMSEstd) indices, estimated from the thermal inactivation of L. monocytogenes using PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil) and the control group.
Table 3.
Z-values (°C) estimated by the One-Step and Two-Step procedures and the Root Mean Square Error (RMSE), Residual Standard Error (RSE) and Standardized RMSE (RMSEstd) indices, estimated from the thermal inactivation of L. monocytogenes using PEO (Pure Oregano Essential Oil), FIV (Fractionated Oregano Essential Oil) and the control group.
| One-Step | Two-Step | ||||||
| Group | RMSE | RMSEstd | z-value | z-value | RSE | t value | Pr (>|t|) |
| Control | 0.31 | 0.06 | 5.75 ± 0.28 | 3.63 ± 0.19 | 0.12 | 19.29 | 0.003 |
| PEO | 0.43 | 0.08 | 5.20 ± 0.14 | 3.69 ± 0.46 | 0.28 | 8.00 | 0.02 |
| FIV | 0.46 | 0.09 | 5.00 ± 0.13 | 4.03 ± 0.55 | 0.28 | 7.27 | 0.02 |
Table 4.
D or δ- and z-values of L. monocytogenes using mild temperatures and essential oil reported in previous studies.
Table 4.
D or δ- and z-values of L. monocytogenes using mild temperatures and essential oil reported in previous studies.
| Matrix | Treatment | D- or δ-values (min) |
Z Values (°C) |
Reference | ||||||
| D52 | D54 | D55 | D57.5 | D60 | D63 | D65 | ||||
| BHI broth supplemented with glucose and yeast extract | P. longiflora PEO 0.06%: | 1.75 | 1.35 | - | 0.17 | - | 2.00x10-3 | - | 5.20 | Present study |
| P. longiflora FIV 0.06%: | 0.64 | 0.54 | - | 0.02 | - | 2.00x10-3 | - | 5.00 | ||
| Sous-vide salmon | Origanum vulgare EO 1% | - | - | 10.03 | 4.88 | 1.81 | - | - | 5.62 | [40] |
| Sous-vide beef | Salvia officinalis EO 0.6% | - | - | 21.17 | - | - | - | - | - | [71] |
| Ground pork | Cinnamaldehyde 0.5% | - | - | 3.61 | - | 0.63 | - | 0.52 | - | [72] |
| BHI broth | Carvacrol 30 µg/mL | - | - | 8.17 | - | 0.67 | 0.17 | 0.07 | - | [73] |
| 2-Hexenal 65 µg/mL | - | - | 8.03 | - | 0.60 | 0.12 | 0.08 | - | ||
| Citral 50 µg/mL | - | - | 8.42 | - | 0.66 | 0.16 | 0.08 | - | ||
| TSBYE | Thymol | - | - | 0.25 | - | - | - | - | - | [74] |
| Carvacrol | - | - | 0.25 | - | - | - | - | - | ||
| Thymol + Carvacrol | - | - | 0.18 | - | - | - | - | - | ||
| PBS | Thymol | - | - | 1.47 | - | - | - | - | - | [75] |
| Carvacrol | - | - | 1.48 | - | - | - | - | - | ||
| Thymol + Carvacrol | - | - | 0.38 | - | - | - | - | - | ||
| Pineapple juice | Mexican coriander 15 µg/mL | 5.61 | 0.53 | 0.28 | [76] | |||||
| Mexican coriander 60 µg/mL | 5.47 | 0.44 | 0.16 | |||||||
| Beef marinated | Grapefruit seed extract 200 ppm | - | - | 22.17 | 6.11 | 3.69 | - | - | 7.98 | [59] |
| Ground Turkey | Sodiumchloride (1%) and green tea polyphenol extract (1.5%) | - | - | 30.40 | - | 5.50 | - | 0.90 | - | [77] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



