
Submitted:
27 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
The accelerated spread of bacterial resistance has been demonstrated to reduce the effectiveness of antibiotic treatments for infections, resulting in higher morbidity and mortality rates, as well as increased costs for livestock producers. It is expected that the majority of future antimicrobial use will be in animal production. The management of AMR in the livestock sector poses significant challenges due to the multifaceted nature of the problem. In order to identify appropriate solutions to the rise of antimicrobial resistance, it is imperative to have a comprehensive understanding of the disease dynamics underpinning the ways in which antimicrobial resistance is transmitted between humans and animals. Furthermore, in consideration of the anticipated requirement to satisfy the global demand for food, it is imperative to guarantee that resistance is not transmitted or propagated during the treatment and disposal of animal waste, particularly from intensive farming. It is also crucial to formulate a research agenda to investigate how antibiotic resistance in animal feces from livestock farming is affected by intensified farming activities. The review analyses the environment's role in the transmission resistance chain and reviews methodologies for disrupting the link. A particular focus is placed on the limitations of the applied methodologies to reduce antimicrobial resistance in global animal production.
Keywords:
intensive livestock farming
; trasmission of multidrug resistance
; antimicrobial resistance policies
1. Introduction
The 20th century witnessed the discovery of molecules capable of combating infection, thus engendering optimism regarding the development of cures [1]. The utilisation of antibiotics as a pharmaceutical intervention for the management of bacterial infections in humans and animals can be traced back to the early and middle 20th century, respectively [2]. However, Fleming's predictions were soon corroborated, as within a few decades, microbes had developed resistance to the drugs, thereby rendering the majority of them ineffective for treating diseases [3]. In less than a hundred years, the world has witnessed a global health crisis of unprecedent proportions, with multidrug resistance significantly impacting global socio-economic development [4]. Studies have shown that antibiotic resistance is an inherent feature of microorganisms, with evidence found in samples from glacial waters over two millennia old, and thus in environments unaffected by humans [5]. Nevertheless, the emergence of bacterial resistance is a direct consequence of the excessive utilisation of antibiotics. It is evident that, despite the inevitability of this phenomenon due to the process of natural selection in which bacteria adapt in order to evade the effects of pharmaceutical drugs, the repercussions of this occurrence are, nevertheless, intensified by the excessive utilisation of antibiotics. This practice is particularly prevalent within the sphere of animal husbandry [6,7] . This phenomenon carries profound implications for global public health, resulting in elevated healthcare costs, prolonged hospital stays and, most significantly, increased mortality for those infected by a drug-resistant pathogen [8]. In order to comprehend the significance of this phenomenon, it is crucial to recognise that in 2019, the number of deaths attributable to this issue exceeded 1.2 million, a figure that is projected to escalate to 10 million by the year 2050 [9,10]. Within the European Union, the mortality rate due to antibiotic resistance in 2020 exceeded 35,000, with Italy accounting for nearly one-third of these casualties [11]. The term 'silent pandemic' has thus been coined to denote the gravity of this situation [12]. The increased mutation and mobilisation of DNA at low antibiotic concentrations has also been demonstrated to result in antimicrobial resistance, thereby allowing microorganisms to adapt and evolve [3]. These genes evolve in a competitive environment, countering other strains in order to ensure their own survival. Chromosomal DNA has the capacity to convey resistance genes via mutations; however, these genes are predominantly located on mobile elements, such as transposons and plasmids, which can facilitate the spread of resistance genes [13]. Once a pathogen has acquired these genes, eradication becomes challenging, particularly if the gene does not impose a significant metabolic burden [14]. It is important to note that a limit exists for the cost associated with the maintenance of these resistance genes. Therefore, they may not become firmly established within the bacterial population, unless there is a significant environmental selective pressure for their maintenance [15]. It is therefore evident that the development and expansion of new resistances is contingent on the environment, where selective pressure encourages gene transfer and maintenance [16]. The utilisation of antibiotics in healthcare and agriculture, specifically in intensive livestock farming, is problematic as it can result in the emergence of novel resistance due to misuse and overuse of these antimicrobials [8]. A number of factors have been identified that, by applying a form of selective pressure, favor the transfer of resistant pathogenic bacteria and/or resistance genes, affecting various sectors across the board [17]. Industrial waste, especially in the pharmaceutical industry, leads to the release of significant quantities of molecules with antibiotic activity into the environment [18]. The inadequate management of wastewater, including that from agricultural activities and residential areas, plays a central role in the contamination of the ecosystem and the escalating prevalence of the resistome [19]. Moreover, soil and surface water play a significant role in the spread of antimicrobial resistance and resistant pathogenic bacteria, acting as a reservoir and a pathway for these organisms [20]. Contaminated water from sewage treatment plants is frequently used for irrigation in agricultural settings, further contributing to the dissemination of these bacteria [21]. The expansion of antimicrobial resistance is also influenced by globalisation, which has resulted in the international marketing of agricultural products, livestock and/or their products and derivatives [22]. In addressing the emergence of the MDR threat, it is imperative to recognise the multifactorial nature of the problem, in which the health of humans, animals and the entire ecosystem are inextricably linked.
2. The Use of Antibiotics in the Veterinary Sector and in the Livestock Industry
The overuse and/or misuse of antibiotics is the primary cause of AMR. This misuse occurs in all three principal areas: human, veterinary and environmental [23]. The solution to this problem is straightforward: usage must be reduced and reserved solely for essential applications in all three areas. The veterinary sector has frequently been cited as a significant contributor to the phenomenon of antibiotic resistance [24]. It has been estimated that 'more than 50% of antibiotics used globally are used in veterinary medicine [25,26]. Antibiotic-resistant bacteria, including Escherichia coli, Salmonella, Campylobacter and enterococci, have demonstrated the ability to cross the species barrier and infect humans, often through direct contact, via the food chain or by the transfer of resistance genes (plasmids) from animal bacteria to common human pathogens, particularly those residing in the intestinal tract [27,28]. The emergence of resistance to antibiotics is an inherent property of bacterial organisms. Nevertheless, the temporal interval between the introduction of ampicillin (one of the most prevalent penicillins) into the human therapeutic market and the initial manifestation of resistance by the genus Salmonella to this antibiotic was too brief (a mere 12 months) for the phenomenon to be ascribed exclusively to the prescription practices of medical professionals [29]. To this end, a comprehensive investigation was conducted on a multitude of Salmonella strains, obtained from over 30 countries and diverse origins (humans, farm animals, feed and food), using whole genome sequencing. This investigation revealed that the ampicillin resistance gene (blaTEM-1) emerged in humans in the 1950s, prior to the introduction of the antibiotic in the pharmaceutical market [29]. It can thus be deduced that the addition of penicillin to animal feed in the decade leading up to its introduction for human use may have contributed to the spread of resistance [3]. These findings suggest that antibiotic residues in soil, wastewater and livestock manure may have a significantly greater impact on the spread of resistance than was previously thought. The utilisation of antibiotics in veterinary medicine is subject to stringent regulation in numerous countries, frequently exceeding the regulatory standards observed in human medicine [30]. However, the utilization of antibiotics in veterinary medicine initially followed the same trajectory as in human medicine [31]. Examples of antibiotics used in both human medicine and veterinary medicine include tetracyclines and penicillin [25]. In the USA, oxytetracycline is still used, but only for therapeutic use in pig feed, with concentrations ranging from 200 to 800 mg/kg [32]. When employed for growth-ptomoting purposes, concentrations range from 50 to 200 mg/kg. Table 1 shows the most common types of antibiotics used in veterinary medicine.
Conversely, other antibiotics, not employed in a medicinal context, are specifically utilised in animal feed for their effect on growth (e.g. avilamycin, avoparcin, flavomycin) [33]. In contrast, Europe and numerous other countries have regulations that prohibit the use of antibiotics registered for therapeutic use in humans and animals as growth promoters [34].
3. The Key Reasons for Using Antibiotics in the Field of Animal Husbandry
The use of antibiotics in the field of veterinary medicine is principally driven by three factors: firstly, the treatment of specific infectious diseases; the second is control or metaphylactic use - the treatment of a group of animals after disease has been diagnosed in part of the group. The objective of this intervention is to treat animals that are manifesting clinical signs of illness, with the aim of preventing the spread of disease to animals in close proximity that may already be infected. The third category, preventative or prophylactic, involves the treatment of an animal or group of animals before clinical signs of disease occur [35,36]. Growth-promoting antibiotics are extensively employed in the farming of pigs, chickens and calves. A comprehensive understanding of the mode of action of such antibiotics can be obtained from the observation that the growth of germ-free animals is 20% higher than the growth of animals that have been left free and not received growth-promoting antibiotics in their diet [3].
The population has exhibited steady growth, with the economy, particularly in developing countries, demonstrating concomitant expansion, resulting in augmented buying capacity. This has resulted in an increased demand for food, especially animal proteins, compared to previous decades [37]. In response to this mounting demand, there has been a transition towards more expansive agricultural production methods. This transition has been primarily characterised by intensive farming systems, particularly for cattle, poultry, and pigs [38]. The underlying objective of this transition is to reduce costs by rendering this category of products suitable for large-scale consumption. The evolution of animal production systems has resulted in two key objectives: the optimisation of efficiency and the implementation of stringent disease prevention measures. Concurrently, there has been an increase in the utilisation of antibiotics, employed to prevent the occurrence of outbreaks on the farm, thereby averting substantial economic losses [7]. Antimicrobials have played a pivotal role in the prevention, treatment and control of food-animal diseases caused by pathogens such as E. coli, S. aureus, Campylobacter spp. and Salmonella spp [39]. Nevertheless, the widespread use of veterinary antimicrobials has favoured the diffusion of antimicrobial resistance, with important consequences for animal and potentially human health. Indeed, the utilisation of antimicrobials in food-producing animals has a long history, dating back to 1951 when the Food and Drug Administration authorised their use as additives in animal feed in the USA, without the need for a veterinary prescription [40]. Since then, antimicrobials have been employed for many decades, not only for the treatment of animal infections, but also for other applications. It is estimated that over 70% of global antimicrobials are used in food-producing animals, particularly in nations with substantial livestock industries, including China, Brazil and the USA [40,41]. [42]. The therapeutic use of antibiotics in animals refers to the administration of antimicrobials for the treatment of animals with clinically diagnosed infectious diseases [25]. A wide range of antibiotics have been approved for this purpose, including for respiratory diseases, enteric diseases and mastitis, with many benefits for both animal health and food safety [25]. While the treatment of sick animals should not be delayed or avoided, the use of antibiotics for this purpose is often inappropriate. Broad spectrum antimicrobials are frequently used before or instead of obtaining a confirmed diagnosis and antimicrobial susceptibility testing. These antimicrobials are then administered to all individuals in the group, even those without any signs of infection, solely on the basis of economic considerations [43]. The term 'disease prevention' is employed to denote the administration of antibiotics for the purpose of prophylaxis or metaphylaxis [34]. Metaphylaxis, on the other hand, involves the administration of antimicrobials to animals with subclinical infections. The aim of metaphylaxis is to reduce the risk of the infection becoming clinically apparent, spreading to other tissue or organs, or being transmitted to other individuals [44]. Prophylaxis, in contrast, is directed at healthy animals with the objective of averting the onset of an infectious disease in instances where the introduction of a pathogen is anticipated on the basis of history, clinical judgement or epidemiological knowledge [45]. Appropriate prophylaxis should be prescribed by a veterinarian, at a therapeutic dosage and for a limited time, with the aim of reducing morbidity and mortality, and potentially the overall use of antimicrobials, should the need for longer therapeutic antimicrobial treatments arise [46]. Moreover, the discovery of the growth-promoting properties of antimicrobials in farm animals has led to the practice of feeding sub-therapeutic doses of antimicrobials for prolonged periods [22]. This has subsequently become a component of the production systems developed in industrialized animal husbandry in numerous countries. The impact of low-dose administration of antimicrobials on the growth rate, feed conversion, litter size and milk yield in dairy cows has been demonstrated to be a significant benefit to animal production [46]. However, it is important to note that repeated exposure to sub-therapeutic doses of antimicrobials has also been shown to cause alterations in the structure and function of the intestinal microbiota. This, in turn, has been demonstrated to promote the growth of resistant microbes and the dissemination of antimicrobial genes [47]. These genes have been observed to be capable of acting against not only the antimicrobial agent used, but also against other antimicrobials. Another significant concern regarding the utilisation of antimicrobials in farm animals pertains to potential implications for human health. A considerable proportion of antimicrobials authorised for veterinary use are classified as antimicrobial classes that are also employed in human medicine [48]. Examples include glycopeptides, polypeptides, macrolides, tetracyclines, beta-lactams, sulfonamides and aminoglycosides. Despite the dearth of studies that have directly addressed this issue, there is an evident consensus in the scientific literature that there exist pathways for AMR to disseminate in both directions, i.e. between bacterial populations of humans and food-producing animals [49,50]. The most commonly cited route of transmission is through the consumption of food contaminated with AMR bacteria. These bacteria may be commensal in animals but pathogens in humans, or vice versa, or may be commensal in both species but subsequently confer resistance to critical pathogens for both humans and animals [51]. The evidence base concerning the transmission of antibiotic resistance between humans and animals is limited and remains challenging to quantify, but concrete evidence exists in some cases. For instance, the rise in fluoroquinolone resistance observed in animal Salmonella isolates has been associated with a subsequent increase in human infections through the consumption of contaminated eggs. Another significant concern regarding the utilisation of antimicrobials in farm animals pertains to the question of potential implications for human health [52]. Furthermore, a robust correlation has been demonstrated between the utilisation of fluoroquinolones in food-producing animals and the emergence of resistance to ciprofloxacin in both animal and human isolates [48]. In the context of E. coli infections, of particular significance is the rapid global emergence of ESBL-producing strains, which has been observed not only in human infections but also in bacteria isolated from food-producing animals, including cows and chickens [53]. The use of avoparcin (a glycopetide analogous to vancomycin) as an antimicrobial growth promoter in poultry has been associated with the emergence of vancomycin-resistant Enterococci [54]. The existence of this resistance has been documented in both food animals and in the human population. A comparable association has been documented between the European Union countries' prohibition of avoparcin in 1997 and a consequent decrease in resistance. [55]. In view of the possible relationship between the use of antibiotics in animal husbandry and its consequences for human health, the Global Action Plan (GAP) was initiated in 2015 as a joint undertaking between the World Health Organisation (WHO), the Food and Agriculture Organisation (FAO), and the Office International des Epizooties (OIE) [56]. GAP's objective is to implement strategies to combat antimicrobial-resistant bacteria. This includes awareness-raising, optimising application and reinforcing surveillance and research [57]. However, policies for antimicrobials in animal production vary between countries. Whilst the use of antimicrobials as growth promoters is prohibited in certain countries, they are utilised extensively in other regions [37]. The use of antibiotics in farm animals is a multifaceted issue that requires a balanced, responsible approach to preserve their effectiveness for humans and animals [58]. It is therefore essential that sustainable, responsible farming practices, such as the implementation of prevention measures (e.g. vaccines) and the improvement of farming conditions, are employed in order to reduce unnecessary antimicrobials.
4. How Do Antibiotics in Livestock Impact Antibiotic-Resistant Human Infections?
The use of antibiotics in intensive animal farming for food production first emerged in the 1940s in the United States of America [42]. A decade later, the marketing of antibiotics for addition to animal feed, with the aim of promoting growth, became globalised. The first molecules to be used were streptomycins and penicillins. Further observations showed that the implementation of this practice in certain regions correlated with an increase in antimicrobial resistance in both pathogens (Salmonella spp., Campylobacter spp.) and commensals (E. coli, Enterococcus spp.) involved in animal-to-human infections [59,60]. The WHO Foodborne Diseases Epidemiology Reference Group (WHO FERG) estimated that foodborne diseases cause 600 million infections, 420,000 deaths and 33 million DALYs each year. Children under five are particularly at risk, with a mortality rate of 125,000 each year (one third) accounting for 40% of cases [61]. These data provide a comprehensive understanding of the global challenges posed by food safety. At present, foodborne and/or animal-borne diseases are considered to be among the principal causes of morbidity and mortality on a global scale [62]. In Europe, the first signs of potential dangers to human health from the use of antibiotics in animal husbandry were observed in 1960, with particular concern expressed in the United Kingdom [63].
The initial evidence of the impact of antibiotic use in livestock farming was presented by the WHO in 1997, following extensive data analysis [64]. The analysis yielded the following findings: 1) The use of antibiotics for growth promotion and animal health on livestock farms led to the emergence of bacteria resistant to multiple antibiotic classes, including those of clinical importance to humans; 2) These resistant bacteria and their associated genes can be transmitted to humans through direct contact or via derived foodstuffs [65,66] (Figure 1).
In 2005, the Food and Agriculture Organization (FAO), the World Organization for Animal Health (OIE) and the World Health Organization (WHO) established the importance of drawing up a list of Critically Important Antimicrobials (CIA), which was subsequently renamed Medically Important Antimicrobials (MIA) and subdivided according to their critical importance for humans [67]. This initiative was conceived as a mechanism to facilitate medical decision-making concerning antibiotic selection, thereby indirectly contributing to risk management and preserving the efficacy of these medications. The primary objective of this strategy was to contain the spread of MDR isolated from animals intended for food production [68]. In the MIA list, the various classes of antibiotics are subdivided into those exclusively designated for human use, those exclusively designated for animal use, and those designated for shared use between humans and animals [69]. A further subdivision, within the field of antibiotics whose use is shared between animals and humans, is characterized by three categories in descending order of importance: critically important, highly important, and important. It was decided that three antibiotic classes should be given top priority: fluoroquinolones, 3rd and 4th generation cephalosporins and polymyxins [69]. The emergence of resistance to new antibiotics is unsurprising when considering that these medications are administered in farming contexts prior to their approval for human use, thereby compromising their efficacy. This occurrence can be attributed to the intricate regulations that govern the evaluation of any potential adverse effects that these novel pharmaceutical agents are subject to prior to their approval for human use [3,10]. Use in an agricultural or animal setting is a quicker method of recouping the costs incurred in research, development and production of the drug. A relevant example in the literature is the FDA's approval of ceftiofur for animals before its approval for humans [40]. This led to a rapid acquisition of resistance to the cephalosporin class in human infections, which restricted its use. Antibiotic usage is a pivotal factor in this regard, with farming settings exhibiting higher usage than clinical settings. The duration of exposure also differs, with farming settings having a greater exposure time. This can be exacerbated by inadequate supervision by veterinarians or by the use of antibiotics that have been banned for this purpose [10,43,66].
5. Mechanisms of Antibiotic Resistance
The main way AMR spreads is through horizontal transmission of resistance genes, which involves: a) conjugation (the transfer of DNA via mobile genetic elements); b) transduction (bacteriophage-mediated gene transfer); and c) transformation (the retrieval of naked DNA from the environment and its incorporation into chromosomes via recombination) (Figure 1) [70].
The activation of these mechanisms enables microorganisms to respond rapidly to the pressure exerted by the suboptimal utilisation of antibiotics. Moreover, this results in the activation of internal microbiota-based signaling, leading to additional mutations that counterbalance the cost of maintained acquired resistance genes, while preserving their persistence (bet hedging) [8,70]. Gene transfer often involves replacing whole gene cassettes with multiple antibiotic resistances, a process called "cross-resistance" [71]. This can result in a form of resistance selection, where exposure to the antibiotic in the cassette leads to resistance against various other antibiotics from different classes, which are then transferred together [71]. This contributes to the spread of multidrug resistance (MDR). Horizontal gene transfer (HGT) is a process that occurs most rapidly under conditions of high microbial density. This phenomenon is particularly evident in the case of the gut microbiota, both in the animal and human environments [72]. The use of sub-optimal amounts of antibiotics in livestock farming creates a selective pressure that favours the exchange of mobile elements between gut bacteria at a rate 25 times faster than that observed in the soil [73]. The microbiota is made up of many species of anaerobic bacteria that, despite their resistance, do not pose a threat. However, facultative anaerobic microorganisms in the Proteobacteria phylum could be pathogenic. Under selective pressure, these microorganisms may acquire new resistance genes from other bacteria in the microbiota [74]. Animals play a key role in the transmission chain, as shown by the presence of antibiotics and antibiotic-resistant bacteria in the environment, linked to faecal and urinary elimination [74]. A recent study has estimated that between 30% and 90% of the administered antibiotics are excreted with faeces, often in the form of unmetabolised compounds [75]. Intensive livestock farms are characterised by overcrowded conditions, frequently resulting in elevated animal population densities. In such environments, animals share space and resources, including water and food, and their waste, both solid and liquid, is accumulated in septic tanks or litter in the case of poultry [64]. Several antibiotics have been found in poultry litter, including virginiamycin, salinomycin, penicillin and bacitracin [76]. Residues of antibiotics are also found in waste, including residues from feed, food and water used to clean the animals. This production process generates a large amount of antibiotic waste, which plays a significant role in the transfer of antibiotics to the environment [77,78]. The use of these materials as fertiliser is a well-established practice, but production sometimes exceeds demand, leading to surplus being sent to disposal processes. This transfer of microbiological and genetic material can alter the existing microbiota. Studies have shown that diseases can be spread through food from crops infected with AMR micro-organisms and resistance genes, due to manure from fertilisation or irrigation with surface water contaminated by runoff from animal waste [79]. In particular, manure has been shown to have a significantly negative impact on the environment (Figure 1). The issue is further exacerbated by the fact that less than 8% of the manure produced in Europe is treated, with significant regional variations being observed. In addition, there is a paucity of evidence that traditional treatments are efficacious in reducing pathogen load [78]. Employing an array of analytical methods facilitated the differentiation of distinct transmission modes at each stage of the chain, from breeding through to sale. The transfer of bacteria and resistance genes from animals to humans can occur via several pathways [51]. Farm workers, who have direct contact with animals and their excreta, are at the forefront of direct transmission (Figure 1). Several studies have documented that these workers are the first to be exposed to AMR pathogens. Furthermore, in the absence of adequate personal hygiene and protective equipment, the clothing or surfaces worn or used by these workers may act as a vector for the spread of AMR bacteria within the home environment, with the risk of compromising the health of the resident community [11]. In conclusion, it is imperative to emphasize that intensive livestock farming facilities are deficient in biosecurity measures. This inadequacy enables unregulated ingress and egress of insects and diminutive animals, encompassing rodents and avian species (Figure 1). These creatures can function as reservoirs and/or carriers for the transmission of antimicrobial resistance (AMR) over extensive distances, thus underscoring the absence of effective barriers to contain the proliferation of AMR [80]. The transportation, slaughter and consumption of animal products and their by-products (e.g. milk, eggs, cheese) constitute additional pivotal phases in this sequence (Figure 1) [7]. Slaughter is carried out in facilities where a high volume of animals is slaughtered and processed on a daily basis. This process involves the movement of animals from one location to another while they are still alive, and even after they have been slaughtered, thereby facilitating the cross-transfer of resistant microbes [10]. Additionally, the waste produced during processing can be disposed of in canals, ultimately entering the environment (Figure 1). The dissemination of antimicrobial resistant bacteria, particularly in under-resourced nations, has been attributed to the release of effluent from industrial facilities [19]. Whilst live microorganisms can be eradicated through standard wastewater treatment processes, the presence of DNA fragments, including AMR genes, remains a persistent concern [20,81]. Despite the development of innovative methodologies that address both pathogens and resistance genes, these approaches are not always integrated within conventional waste management practices [82]. The international trade of live animals has been identified as a potential contributing factor to the dissemination of antibiotic resistance genes. Exports of poultry, beef and pig products have been identified as a significant source of these genes, with quantities so substantial that effective biological control becomes challenging without risking product spoilage [83]. This assertion is substantiated by a Danish study which revealed the prevalence of resistance to extended-spectrum cephalosporins in E. coli strains isolated from a chicken farm. The chickens in question had been imported from a farm in Scotland where the use of these molecules was permitted prior to 2012, and the resistance genes were thus introduced to Denmark via the chicken farm, despite the export ban [84].
The transmission of antimicrobial resistance (AMR) bacteria from animals to humans has been well-documented, particularly in the case of infections caused by non-typhoid Salmonella spp, Campylobacter spp, S. aureus [85]. A recent study has observed that 75% of human infections in recent years are of zoonotic origin [86]. Salmonellosis, a foodborne infection, has been observed worldwide, often resulting from the ingestion of infected meat (cattle, pigs, turkeys, chicken), eggs, and milk [87]. EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control) have reported that, in 2021, out of a total of 2,201 cases, 590 (approximately 28%) were attributable to Salmonella spp. [85]. According to data from the United States-based FoodNet project (2022), the incidence of Salmonella and Campylobacter infections per 100,000 individuals was documented to be, respectively, 14.4 and 17.2. The elevated incidence was predominantly observed among children under the age of five [88,89]. Furthermore, the European Food Safety Authority (EFSA) has reported that a significant proportion of Salmonella spp. isolates from human infections in 2023 demonstrated resistance to ampicillin (21.3%), sulphonamides (20.8%) and tetracyclines (21.8%) [85]. Concurrently, Salmonella spp. strains were isolated from farm animals in the majority of European countries, exhibiting an equivalent percentage of resistance to the same antibiotic classes [85]. Furthermore, while a decrease in the percentage of resistance to ampicillin and tetracyclines was observed in human isolates during the period 2014-2023, the percentage of resistance to ampicillin in isolates derived from broilers showed an increasing trend throughout Europe [85]. Only 1.6% of Salmonella spp were resistant to the third-generation cephalosporin cefotaxime, and only 1.3% to ceftazidime in human infections. These findings align with the low levels of resistance in animals: 0.2% in laying hens, 0.8% in pigs and 1.4% in young cattle [60,85,90].
In 2022, a high prevalence of fluoroquinolone resistance was observed among Salmonella spp isolated from broiler chickens (55.5% ciprofloxacin). In 2023, 21.8% of Salmonella strains isolated from humans demonstrated resistance to ciprofloxacin. From 2014 to 2023, there was a statistically significant increase in resistance to ciprofloxacin in broiler and laying hens. Such increasing trends were also observed in Salmonella spp. isolates from humans in at least nine countries [85]. Minimal resistance to colistin has been recorded in Salmonella spp. isolates from farm animals and poultry carcasses [85]. According to EFSA data, in the 2022-2023 period, the resistance levels in Campylobacter jejuni isolates of human and animal origin to fluoroquinolones were sufficiently high to exclude this class of antibiotics from the recommended therapeutic treatment for humans [85]. An analysis of data from 2014 to 2023 revealed a significant increase in ciprofloxacin resistance among C. jejuni isolates from humans in 11 European countries. In 2022, 78.1% of C. jejuni isolates from turkeys and 70.9% from broilers exhibited resistance to ciprofloxacin [85]. The evaluation of MRSA resistance in S. aureus species, both in food and in animals, is inadequate due to its voluntary nature, and consequently, not all countries have adopted it. The LA-MRSA clone CC398, despite its low pathogenicity to humans, has the potential to serve as a reservoir for antimicrobial resistance genes that could be transmissible to virulent human S. aureus strains [91]. Levels of MRSA were observed to vary between species and countries, with isolation rates ranging from 0% in pigs in Norway to 80% in pigs in Belgium [85]. In addition, all isolates with MRSA resistance carried the mecA gene. Germany and Denmark have reported increased isolation of LA-MRSA strains that can cause human infections. Transmission is via infected meat. However, CA and/or HA-MRSA lineages associated with human infections have been detected in farmed animals, suggesting an incidental exchange of strains between animals and humans [92]. There is little data on E. faecalis and E. faecium. The latter is more resistant to vancomycin than the former, but the difference is minimal in young cattle (age < 1 year) [85].
6. Policies and Strategies for Tackling Antimicrobial Resistance from Livestock to Humans
As previously stated, AMR exerts a considerable influence on public health, primarily due to the transmission of antibiotic-resistant bacteria from livestock to humans and vice versa [10]. Consequently, numerous surveillance networks have been implemented in recent decades to systematically collect data on antibiotic usage and antibiotic resistance in livestock (Figure 2).
Antimicrobial quantities (in tonnes) intended for use in animals, as well as the number of countries reporting use or non-use of growth promoters in 2022, were extracted from the WOAH Annual Report 2023 (https://amu.woah.org/amu-system-portal/amu-data);. Furthermore, regions exhibiting elevated antimicrobial use intensity (i.e. 'hotspots' [37]) are delineated on the map. These regions include:
- -
- Asia: eastern China, southern India, Indonesia, central Thailand, the eastern coastline of Vietnam, western South Korea, eastern India and Bangladesh, Pakistan and north-west Iran
- -
- Europe: northern Italy, northern Germany and central Poland
- -
- The Americas: south of Brazil and the Midwest of the USA
- -
- Africa: Nile delta and peri-urban areas of Johannesburg
These networks are vital for government interventions as they allow continuous monitoring of antibiotic usage, enabling the assessment of resistance trends across different animals [93]. The first country to establish a monitoring and research programme on antimicrobial resistance was Denmark (DANMAP) in 1995 [94]. The objective of DANMAP was to collect data on the sales of antibiotics for veterinary use. Reports from DANMAP highlighted a potential link between the widespread use of avoparcin in poultry and the contamination of meat with vancomycin-resistant enterococci. Subsequent to this, Denmark witnessed a surge in antibiotic usage, which exhibited a direct correlation with the emergence of ESBL-producing E. coli isolates within the livestock population. Government interventions culminated in a decline in antibiotic consumption, concomitant with a concurrent decline in ESBL-producing E. coli cases in pigs. In 1996, US agencies-initiated monitoring of antimicrobial resistance in bacteria from humans, animals and meat. The surveillance programme revealed the presence of fluoroquinolone resistance in Campylobacter samples from poultry. This finding was of such significance that it resulted in a reduction in the utilisation of enrofloxacin [95]. Furthermore, in 1997, the French National Observatory for Epidemiology of Antibiotic Resistance (RESAPATH) was established for the purpose of gathering data on antibiotic consumption in animals. The research revealed a 27.9% increase in antibiotic usage between 1999 and 2009, concomitant with a surge in cases of multi-resistant E. coli [96]. In 2002, Canada launched the Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS), a comprehensive surveillance system that integrates data on antibiotic use across both human and animal populations, with the objective of assessing its impact on public health. A notable finding from CIPARS was the identification of an increase in multidrug-resistant Salmonella in humans, which was found to be associated with the use of ceftiofur in poultry in specific regions of Canada [97]. This finding underscores the close correlation between antimicrobial resistance in humans and livestock, highlighting the need for interdisciplinary research and policy interventions to address these pressing public health concerns. China is a major producer and consumer of antibiotics, with an estimated production of 248,000 tons in 2013, 52% of which was allocated to livestock farming, approximately 2.8 times the amount used in the United States [98] (Figure 2). In 2019, China initiated measures to combat antimicrobial resistance by prohibiting the production, importation, and consumption of eight antibiotics. The following substances are proscribed: guaiacolin, methylene salicylic acid, bacitracin, bacitracin zinc, chlortetracycline, aflatoxin, virginia, and quinenone. These substances were previously employed as growth promoters and for the prevention of infection in livestock [99,100]. Japan was the first nation in Asia to establish a reporting system, the Japanese Veterinary Antimicrobial Monitoring System (JVARM), which commenced data collection in 2000 [101]. Since 2005, the European Surveillance of Veterinary Consumption (ESVAC) group of the European Medicines Agency has been responsible for the collection of data on the utilisation of veterinary antibiotics, with the initial findings published in 2011. This inaugural publication utilised sales data from eight countries and was expanded in 2017 to encompass all European countries [102] (Figure 2). These measures have included the prohibition of the use of certain antibiotics for non-therapeutic purposes and the development of stricter veterinary prescriptions to prevent overuse.
6.1. Alternative Strategies to Tackle Antimicrobial Resistance
Further investigation is necessary to determine the effectiveness of probiotics and prebiotics in reducing drug administration and to explore the potential application of veterinary vaccines [83]. In the future, should technological advancements render this a possibility, genetic modifications could be used to develop species that are resistant to infection. The development of transgenic chickens that do not transmit avian influenza can be cited as an example of this phenomenon [103]. Alternative strategies (not antibiotics) are needed to promote animal health and prevent diseases (Figure 3).
A number of key actions must be taken in order to reduce the occurrence of antimicrobial resistance, and to curtail the emergence and subsequent propagation of resistance in food and agricultural sectors worldwide. The aim of these efforts must be to ensure the preservation of the efficacy of antimicrobials in the treatment of both animal and human infections.
They should be part of a comprehensive health management programme, not used separately. Vaccines are a promising alternative to antibiotics. They can reduce the need for antimicrobials, preventing bacterial disease or viral infections. They can also lower antibiotic use by reducing the risk of misdiagnosis and preventing secondary bacterial infections. Their mode of action is to trigger an immune response without causing the harmful clinical consequences associated with the disease [104,105]. However, many currently available veterinary vaccines have limitations, reducing their usefulness. For example, the vaccine may not match the circulating field strains for Streptococcus suis, swine influenza virus, Haemophilus parasuis and Eimeria [106,107,108,109]. In other cases, protection after vaccination may be short-lived and require frequent booster vaccinations, as for Clostridium perfringens and bovine respiratory syncytial virus [110].
Antibiotics have long been used to promote growth, but we don't know much about the metabolic pathways they affect. The current leading theory is related to the gut microbiota and how it relates to productivity. Research has shown that low-dose antibiotics, without killing the beneficial bacteria needed for efficient nutrient absorption, can change the microbiota [111]. Studies have also shown that not all antibiotics stimulate growth equally. For example, virginiamycin enhances growth, while tetracycline has low yield. This suggests that they interact differently with the microbiota. Virginiamycin appears to reduce the microbiota's nutrient needs by changing the abundance of its component genera [112]. It has also been observed that different races respond differently to antibiotic administration, affecting nutrient uptake for body mass [113,114]. It is crucial to note that the absence of illness is fundamental. To illustrate this point, consider a case of a banal intestinal infection in a farm chicken, which has a growth cycle of 40-45 days. In such a scenario, a loss of 1-2 days of nutritional intake due to diarrhoea can result in weight loss, which may have significant economic consequences. Many alternative substances are being investigated as replacements for antibiotics as in-feed growth promoters. Natural dietary supplements such as in-feed enzymes, probiotics and prebiotics show promise (Figure 3). Enzymes help the animals break down and digest plant materials that they otherwise cannot utilize [115]. Broiler chickens have improved nutrient uptake, enhanced weight gain and better feed conversion ratios when supplemented with enzymes. However, the response to food enzymes is influenced by factors like feed humidity, pH and length of interaction [116]. A significant effect of probiotics is their capacity to compete with pathogens for adhesion sites along the intestinal mucosa, a phenomenon referred to as competitive exclusion. By establishing themselves in high numbers, probiotics limit the space available for harmful microorganisms, preventing colonization and infection [117]. Probiotics boost the immune system, stimulating immune cell activity to improve surveillance and accelerate reactions against potential threats. Probiotics also influence the intestinal metabolic functions, promoting bioactive compounds that can combat pathogens [118]. A variety of microbial strains are employed in probiotic formulations. These include Lactobacillus and Streptococcus spp., which have been the focus of substantial research due to their recognised beneficial effects on gut health. Prebiotics, such as fructooligosaccharides (FOS), galactooligosaccharides (GOS), inulin and mannan-oligosaccharides (MOS), are non-digestible fibres that stimulate the growth of beneficial microbes, such as Bifidobacterium and Lactobacillus, while preventing the presence of pathogens that cause gastrointestinal diseases, including C. perfringens [119]. Conversely, alternatives to antibiotics, such as antimicrobial peptides, phytochemicals and organic acids, when incorporated into animal diets, have the potential to diminish reliance on antimicrobials and thereby contribute to the reduction of antimicrobial resistance [120]. Unlike in the clinical setting where there are systems to control infections and health workers, there are no regulations about risks of pathogens for workers living with animals. There is no collection of data on exposures and diseases to determine the risk and disease burden. Biosecurity and management practices are important in disease prevention, improving animal health and reducing the risk of pathogens [121]. The following elements are of particular significance in the context of biosecurity on farms and in livestock production:
- The implementation of controlled access measures for people, vehicles, and animals that may potentially carry pathogens.
- The maintenance of adequate farm fencing to prevent contact with stray and wild animals.
- The adherence to stringent hygiene standards, including regular handwashing, boot disinfection, and thorough cleaning of equipment, to minimise the spread of pathogens within farm environments. Effective sanitation protocols are essential for disinfecting surfaces, tools and equipment to minimise the risk of disease transmission (Figure 3). Regular monitoring of animal health and behaviour allows for the early identification of potential illnesses, enabling timely interventions to prevent infection [122]. Maintaining good ventilation and not overcrowding animals are important strategies to improve air quality and support animal health. Regular monitoring allows farmers to quickly report any issues to the relevant authorities. This helps to contain outbreaks. Surveillance and research are vital for guiding stakeholders on how to slow the spread of antimicrobial resistance. They provide reliable data on antimicrobial-resistant microorganisms, antimicrobial use and the presence of antimicrobial residues in food and feed [8]. Meat slaughter plants are another vulnerable sector. A study of a chicken processing plant revealed that biofilms can diminish the efficacy of cleaning procedures involving various chemicals [123]. Moreover, while urban plants are subject to regulatory standards for pathogens in wastewater, there are no restrictions on the treatment of waste from production animal farms. Storage and composting separate solid waste from liquid waste. But they don't reduce pathogens or pollutants. Calcium oxide stabilises solid waste before using it as fertiliser. The composition of wastewater renders conventional treatment methods ineffective, as evidenced by the frequent presence of elevated concentrations of antibiotics in post-treated European samples. Recent advancements in wastewater treatment technologies have been demonstrated by the adoption of adsorption methods utilising wheat straw, which have achieved 90-98% removal of ciprofloxacin [124]. A range of alternative techniques is also being reviewed, including membrane separation, microbial electrolysis, photocatalytic degradation and advanced oxidation. However, implementation of these methods requires efficiency per cycle, safety during use, storage and transportation, and treatment of secondary polluting products, which would lead to increased costs [124]. In recent years, there has been an increasing focus among researchers on the development of new strategies for tackling antibiotic resistance in animal husbandry. The objectives of these strategies are twofold: firstly, to improve animal health, and secondly, to reduce reliance on conventional antibiotics. These strategies include the use of peptides and nanoparticles with antimicrobial activity [125]. Antimicrobial peptides (AMPs) are a class of small molecules composed of fewer than 100 amino acid residues, which are naturally produced by most living organisms. These peptides have demonstrated broad-spectrum antimicrobial activity against bacteria, fungi, and protozoa. The mechanisms through which they exert their effects are diverse, targeting both intracellular and extracellular components, thereby strengthening the host's immune defences against pathogens [126]. Research has shown that AMPs are effective at preventing infections. They have also been found to enhance the effectiveness of antibiotics, reducing the development of drug resistance [127]. In some cases, dietary supplementation with AMPs has also been observed to promote animal growth. For example, microcin J25 (MccJ25), a peptide produced by E. coli and found in animal faeces, has been shown to positively influence animal growth [128]. Another innovative and promising technology under investigation involves the synthesis of nanoparticles (NPs) [129]. The synthesis of NPs can be achieved through a variety of methodologies, including physical, chemical, or green synthesis methods. The classification of these particles can be categorised into two broad categories: organic and inorganic. The organic category includes liposomes, polymeric, and lipid nanoparticles, while the inorganic group includes quantum dots and metal/metal oxide nanoparticles. These can be used as animal feed additives, as disinfectants or vaccine adjuvants to boost immunity. Moreover, they can be utilised as standalone antimicrobial agents [130]. Several studies have demonstrated that NPs improve antimicrobial activity against antibiotic-resistant bacteria and inhibit biofilm formation [131]. However, numerous aspects related to the properties, applications, mechanisms of action, and optimal dosages of NPs remain to be elucidated and require further investigation. To date, no nanoparticle-based product has been approved for veterinary use [132].
7. Conclusion And Future Directions
The extensive use of antibiotics in animal husbandry has been shown to contribute to the selection of bacterial populations that are increasingly resistant to the drugs used to treat animal and human diseases. Furthermore, it has been definitively established that there are multiple transmission routes through which these resistant bacteria can be transferred to humans. The issue is further compounded by the presence of antibiotic residues in foodstuffs. It has been demonstrated that the regular ingestion of low-dose antibiotics in conjunction with foodstuffs engenders a selective pressure on intestinal bacteria, thereby promoting the proliferation of antibiotic-resistant bacteria. This genetic information is then transferred to other bacteria, including pathogenic ones. In an effort to contain the development of antibiotic resistance, the scientific community has reached a consensus that the use of antibiotics should be reserved exclusively for the treatment of diseases, following a rigorous diagnosis. Furthermore, the implementation of mass treatments for both metaphylactic and, most significantly, prophylactic purposes should be avoided or strictly limited. However, it has been observed that these pharmaceuticals are frequently administered to all animals, whether diluted in water or mixed in food, a practice that is likely to result in unintended consequences, including the increased prevalence of antibiotic resistance. The mass administration of antibiotics to all animals, even those only a few days old, in the absence of any evidence of specific pathologies, is driven by the objective of maintaining maximum animal survival. Unfortunately, even where there are regulations prohibiting this use of antibiotics, such as European legislation banning preventive mass treatments on farms, data show that the majority of antibiotics sold for use in livestock in several countries are still used for mass treatments. Nevertheless, the issue poses a threat to global public health, as the proliferation of antibiotic-resistant bacteria transcends the geographical boundaries of nation states. It is therefore imperative to acknowledge the global nature of this problem and to devise equally global solutions. In order to comprehend the veracity of this assertion, one merely needs to consider the recent pandemic. Despite the temporal and geographical distance of the primary occurrence, owing to the phenomenon of globalization, its consequences have nevertheless had a universal resonance. From this standpoint, the global transportation of animals and goods is widespread and consequently, a phenomenon (e.g. a resistant bacterium) originating in Asia or Oceania can reach Europe within a remarkably brief timeframe. Moreover, the paucity of data regarding veterinary antibiotics poses a significant challenge in the estimation of the amount of antibiotics utilised in the context of livestock farming. The absence of a standardised approach to the collection and sharing of data has the effect of making it difficult to apply these partial data findings to the global context. These limitations compromise the precision of estimates, hindering effective measures to address antimicrobial resistance. It is evident that a reduction in the use of antibiotics in food animals will not be achieved without a multifaceted approach. Despite the ongoing utilisation of antibiotics in numerous developing countries with the aim of enhancing productivity, there are positive indications that this practice is being phased out. For instance, all UN policies on sustainable development include the objective of reducing antibiotic use from a farm-to-fork perspective It is imperative to acknowledge that the prospective unfavourable consequences of antibiotic reduction in animal husbandry can be mitigated. In fact, there exists a plethora of alternative measures capable of maintaining high productivity whilst simultaneously ensuring animal health. The primary strategy entails the formulation and execution of a comprehensive plan to enhance environmental and nutritional conditions, biosecurity measures, hygiene practices, and disease surveillance mechanisms within extensive livestock farms. The second strategy encompasses the advancement of novel technologies capable of facilitating the production of novel and more efficacious vaccines against both viral and bacterial diseases afflicting intensively farmed animals. The development of vaccines tailored to specific pathogens, with the aim of preventing disease outbreaks, is an effective strategy for reducing the reliance on antibiotic use. This approach has been demonstrated to engender two principal benefits: firstly, the reduction of healthcare costs and secondly, the enhancement of public health outcomes.
Author Contributions
Conceptualization, M.T., S.De.G., E.S. C.B.; figure and preparation of the original draft, M.T., S.De.G, E.S., A.M., G.M., S.S. and T.C.; proofreading and editing, M.T., S.De.G., E.S., and C.B. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mancuso, G.; De Gaetano, S.; Midiri, A.; Zummo, S.; Biondo, C. The Challenge of Overcoming Antibiotic Resistance in Carbapenem-Resistant Gram-Negative Bacteria: "Attack on Titan". Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, W.A. The Treasure Called Antibiotics. Annals of Ibadan postgraduate medicine 2016, 14, 56–57. [Google Scholar] [PubMed]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Selvarajan, R.; Obize, C.; Sibanda, T.; Abia, A.L.K.; Long, H. Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance. Antibiotics 2022, 12. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.F. Antibiotic resistance in the environment. Nature reviews. Microbiology 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Wisniewski, P.; Trymers, M.; Chajecka-Wierzchowska, W.; Tkacz, K.; Zadernowska, A.; Modzelewska-Kapitula, M. Antimicrobial Resistance in the Context of Animal Production and Meat Products in Poland-A Critical Review and Future Perspective. Pathogens 2024, 13. [Google Scholar] [CrossRef]
- Oliveira, M.; Antunes, W.; Mota, S.; Madureira-Carvalho, A.; Dinis-Oliveira, R.J.; Dias da Silva, D. An Overview of the Recent Advances in Antimicrobial Resistance. Microorganisms 2024, 12. [Google Scholar] [CrossRef]
- Antimicrobial Resistance, C. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11. [Google Scholar] [CrossRef]
- Ponzo, E.D.G., S.; Midiri, A.; Mancuso, G.; Giovanna, P.; Giuliana, D.; Zummo, S.; Biondo, C. The Antimicrobial Resistance Pandemic Is Here: Implementation Challenges and the Need for the One Health Approach. Hygiene 2024, 297–316. [CrossRef]
- Sirota, M. Should we stop referring to the pandemic of antimicrobial resistance as silent? JAC-antimicrobial resistance 2024, 6, dlae018. [Google Scholar] [CrossRef] [PubMed]
- Porse, A.; Jahn, L.J.; Ellabaan, M.M.H.; Sommer, M.O.A. Dominant resistance and negative epistasis can limit the co-selection of de novo resistance mutations and antibiotic resistance genes. Nature communications 2020, 11, 1199. [Google Scholar] [CrossRef] [PubMed]
- Sirwan Khalid Ahmed, S.H., Karzan Qurbani, Radhwan Hussein Ibrahim, Abdulmalik Fareeq, Kochr Ali Mahmood, Mona Gamal Mohamed,. Antimicrobial resistance: Impacts, challenges, and future prospects. Journal of Medicine, Surgery, and Public Health 2024, 2. [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS microbiology reviews 2018, 42. [Google Scholar] [CrossRef]
- Iwu, C.D.; Korsten, L.; Okoh, A.I. The incidence of antibiotic resistance within and beyond the agricultural ecosystem: A concern for public health. MicrobiologyOpen 2020, 9, e1035. [Google Scholar] [CrossRef]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic Resistance: One Health One World Outlook. Frontiers in cellular and infection microbiology 2021, 11, 771510. [Google Scholar] [CrossRef]
- Pramod Barathe, K.K., Sagar Reddy, Varsha Shriram, Vinay Kumar. Antibiotic pollution and associated antimicrobial resistance in the environment. Journal of Hazardous Materials Letters 2024, 5. [CrossRef]
- Sambaza, S.S.; Naicker, N. Contribution of wastewater to antimicrobial resistance: A review article. Journal of global antimicrobial resistance 2023, 34, 23–29. [Google Scholar] [CrossRef]
- La Rosa, M.C.; Maugeri, A.; Favara, G.; La Mastra, C.; Magnano San Lio, R.; Barchitta, M.; Agodi, A. The Impact of Wastewater on Antimicrobial Resistance: A Scoping Review of Transmission Pathways and Contributing Factors. Antibiotics 2025, 14. [Google Scholar] [CrossRef]
- Ali, A.S.; Gari, S.R.; Goodson, M.L.; Walsh, C.L.; Dessie, B.K.; Ambelu, A. Fecal Contamination in the Wastewater Irrigation System and its Health Threat to Wastewater-Based Farming Households in Addis Ababa, Ethiopia. Environmental health insights 2023, 17, 11786302231181307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Nickerson, R.; Zhang, W.; Peng, X.; Shang, Y.; Zhou, Y.; Luo, Q.; Wen, G.; Cheng, Z. The impacts of animal agriculture on One Health-Bacterial zoonosis, antimicrobial resistance, and beyond. One health 2024, 18, 100748. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Vercelli, C.; Gambino, G.; Amadori, M.; Re, G. Implications of Veterinary Medicine in the comprehension and stewardship of antimicrobial resistance phenomenon. From the origin till nowadays. Veterinary and animal science 2022, 16, 100249. [Google Scholar] [CrossRef]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Teuber, M. Veterinary use and antibiotic resistance. Current opinion in microbiology 2001, 4, 493–499. [Google Scholar] [CrossRef]
- Shang, K.; Kim, J.H.; Park, J.Y.; Choi, Y.R.; Kim, S.W.; Cha, S.Y.; Jang, H.K.; Wei, B.; Kang, M. Comparative Studies of Antimicrobial Resistance in Escherichia coli, Salmonella, and Campylobacter Isolates from Broiler Chickens with and without Use of Enrofloxacin. Foods 2023, 12. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. International journal of environmental research and public health 2013, 10, 2643–2669. [Google Scholar] [CrossRef]
- Tran-Dien, A.; Le Hello, S.; Bouchier, C.; Weill, F.X. Early transmissible ampicillin resistance in zoonotic Salmonella enterica serotype Typhimurium in the late 1950s: A retrospective, whole-genome sequencing study. The Lancet. Infectious diseases 2018, 18, 207–214. [Google Scholar] [CrossRef]
- Schmerold, I.; van Geijlswijk, I.; Gehring, R. European regulations on the use of antibiotics in veterinary medicine. European journal of pharmaceutical sciences: Official journal of the European Federation for Pharmaceutical Sciences 2023, 189, 106473. [Google Scholar] [CrossRef]
- Moulin, G.; Cavalie, P.; Pellanne, I.; Chevance, A.; Laval, A.; Millemann, Y.; Colin, P.; Chauvin, C.; Antimicrobial Resistance ad hoc Group of the French Food Safety, A. A comparison of antimicrobial usage in human and veterinary medicine in France from 1999 to 2005. The Journal of antimicrobial chemotherapy 2008, 62, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Granados-Chinchilla, F.; Rodriguez, C. Tetracyclines in Food and Feedingstuffs: From Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. Journal of analytical methods in chemistry 2017, 2017, 1315497. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well known antibiotics on gram-positive bacteria. Clinical microbiology reviews 2003, 16, 175–188. [Google Scholar] [CrossRef]
- Simjee, S.; Ippolito, G. European regulations on prevention use of antimicrobials from january 2022. Brazilian journal of veterinary medicine 2022, 44, e000822. [Google Scholar] [CrossRef] [PubMed]
- Ardakani, Z.; Canali, M.; Aragrande, M.; Tomassone, L.; Simoes, M.; Balzani, A.; Beber, C.L. Evaluating the contribution of antimicrobial use in farmed animals to global antimicrobial resistance in humans. One health 2023, 17, 100647. [Google Scholar] [CrossRef]
- Allen, H.K.; Levine, U.Y.; Looft, T.; Bandrick, M.; Casey, T.A. Treatment, promotion, commotion: Antibiotic alternatives in food-producing animals. Trends in microbiology 2013, 21, 114–119. [Google Scholar] [CrossRef]
- Mulchandani, R.; Wang, Y.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in food-producing animals: 2020 to 2030. PLOS global public health 2023, 3, e0001305. [Google Scholar] [CrossRef]
- Komarek, A.M.; Dunston, S.; Enahoro, D.; Godfray, H.C.J.; Herrero, M.; Mason-D'Croz, D.; Rich, K.M.; Scarborough, P.; Springmann, M.; Sulser, T.B.; et al. Income, consumer preferences, and the future of livestock-derived food demand. Global environmental change: Human and policy dimensions 2021, 70, 102343. [Google Scholar] [CrossRef]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on Major Food-Borne Zoonotic Bacterial Pathogens. Journal of tropical medicine 2020, 2020, 4674235. [Google Scholar] [CrossRef]
- Lees, P.; Pelligand, L.; Giraud, E.; Toutain, P.L. A history of antimicrobial drugs in animals: Evolution and revolution. Journal of veterinary pharmacology and therapeutics 2021, 44, 137–171. [Google Scholar] [CrossRef]
- Hosain, M.Z.; Kabir, S.M.L.; Kamal, M.M. Antimicrobial uses for livestock production in developing countries. Veterinary world 2021, 14, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.J.; Wellington, M.; Shah, R.M.; Ferreira, M.J. Antibiotic Stewardship in Food-producing Animals: Challenges, Progress, and Opportunities. Clinical therapeutics 2020, 42, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L.; Sweeney, M.T.; Lubbers, B.V. Antimicrobial Susceptibility Testing of Bacteria of Veterinary Origin. Microbiology spectrum 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Gens, K.D.; Singer, R.S.; Dilworth, T.J.; Heil, E.L.; Beaudoin, A.L. Antimicrobials in Animal Agriculture in the United States: A Multidisciplinary Overview of Regulation and Utilization to Foster Collaboration: On Behalf Of the Society of Infectious Diseases Pharmacists. Open forum infectious diseases 2022, 9, ofac542. [Google Scholar] [CrossRef]
- Amenu, K.; McIntyre, K.M.; Moje, N.; Knight-Jones, T.; Rushton, J.; Grace, D. Approaches for disease prioritization and decision-making in animal health, 2000-2021: A structured scoping review. Frontiers in veterinary science 2023, 10, 1231711. [Google Scholar] [CrossRef]
- Odey, T.O.J.; Tanimowo, W.O.; Afolabi, K.O.; Jahid, I.K.; Reuben, R.C. Antimicrobial use and resistance in food animal production: Food safety and associated concerns in Sub-Saharan Africa. International microbiology: The official journal of the Spanish Society for Microbiology 2024, 27, 1–23. [Google Scholar] [CrossRef]
- Pokharel, S.; Shrestha, P.; Adhikari, B. Antimicrobial use in food animals and human health: Time to implement 'One Health' approach. Antimicrobial resistance and infection control 2020, 9, 181. [Google Scholar] [CrossRef]
- Theodoridou Oxinou, D.; Lamnisos, D.; Filippou, C.; Spernovasilis, N.; Tsioutis, C. Antimicrobial Use and Antimicrobial Resistance in Food-Producing Animals: Cross-Sectional Study on Knowledge, Attitudes, and Practices Among Veterinarians and Operators of Establishments in the Republic of Cyprus. Antibiotics 2025, 14. [Google Scholar] [CrossRef]
- Kimera, Z.I.; Mshana, S.E.; Rweyemamu, M.M.; Mboera, L.E.G.; Matee, M.I.N. Antimicrobial use and resistance in food-producing animals and the environment: An African perspective. Antimicrobial resistance and infection control 2020, 9, 37. [Google Scholar] [CrossRef]
- Graham, D.W.; Bergeron, G.; Bourassa, M.W.; Dickson, J.; Gomes, F.; Howe, A.; Kahn, L.H.; Morley, P.S.; Scott, H.M.; Simjee, S.; et al. Complexities in understanding antimicrobial resistance across domesticated animal, human, and environmental systems. Annals of the New York Academy of Sciences 2019, 1441, 17–30. [Google Scholar] [CrossRef]
- Argudin, M.A.; Deplano, A.; Meghraoui, A.; Dodemont, M.; Heinrichs, A.; Denis, O.; Nonhoff, C.; Roisin, S. Bacteria from Animals as a Pool of Antimicrobial Resistance Genes. Antibiotics 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Marimon, J.M.; Gomariz, M.; Zigorraga, C.; Cilla, G.; Perez-Trallero, E. Increasing prevalence of quinolone resistance in human nontyphoid Salmonella enterica isolates obtained in Spain from 1981 to 2003. Antimicrobial agents and chemotherapy 2004, 48, 3789–3793. [Google Scholar] [CrossRef] [PubMed]
- Mandujano-Hernandez, A.; Martinez-Vazquez, A.V.; Paz-Gonzalez, A.D.; Herrera-Mayorga, V.; Sanchez-Sanchez, M.; Lara-Ramirez, E.E.; Vazquez, K.; de Jesus de Luna-Santillana, E.; Bocanegra-Garcia, V.; Rivera, G. The Global Rise of ESBL-Producing Escherichia coli in the Livestock Sector: A Five-Year Overview. Animals: An open access journal from MDPI 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Sorum, M.; Holstad, G.; Lillehaug, A.; Kruse, H. Prevalence of vancomycin resistant enterococci on poultry farms established after the ban of avoparcin. Avian diseases 2004, 48, 823–828. [Google Scholar] [CrossRef]
- Simm, R.; Slettemeas, J.S.; Norstrom, M.; Dean, K.R.; Kaldhusdal, M.; Urdahl, A.M. Significant reduction of vancomycin resistant E. faecium in the Norwegian broiler population coincided with measures taken by the broiler industry to reduce antimicrobial resistant bacteria. PLoS ONE 2019, 14, e0226101. [Google Scholar] [CrossRef]
- Wernli, D.; Harbarth, S.; Levrat, N.; Pittet, D. A 'whole of United Nations approach' to tackle antimicrobial resistance? A mapping of the mandate and activities of international organisations. BMJ global health 2022, 7. [Google Scholar] [CrossRef]
- Majumder, M.A.A.; Rahman, S.; Cohall, D.; Bharatha, A.; Singh, K.; Haque, M.; Gittens-St Hilaire, M. Antimicrobial Stewardship: Fighting Antimicrobial Resistance and Protecting Global Public Health. Infection and drug resistance 2020, 13, 4713–4738. [Google Scholar] [CrossRef]
- Rahman, M.M.; Alam Tumpa, M.A.; Zehravi, M.; Sarker, M.T.; Yamin, M.; Islam, M.R.; Harun-Or-Rashid, M.; Ahmed, M.; Ramproshad, S.; Mondal, B.; et al. An Overview of Antimicrobial Stewardship Optimization: The Use of Antibiotics in Humans and Animals to Prevent Resistance. Antibiotics 2022, 11. [Google Scholar] [CrossRef]
- Barata, R.; Saavedra, M.J.; Almeida, G. A Decade of Antimicrobial Resistance in Human and Animal Campylobacter spp. Isolates. Antibiotics 2024, 13. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, V.; Dapkevicius, M.L.E.; Canica, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria Among Food-Producing Animals: Health Implications of Extended Spectrum beta-lactamase (ESBL) Production. Animals: An open access journal from MDPI 2020, 10. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS medicine 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed]
- Elbehiry, A.; Abalkhail, A.; Marzouk, E.; Elmanssury, A.E.; Almuzaini, A.M.; Alfheeaid, H.; Alshahrani, M.T.; Huraysh, N.; Ibrahem, M.; Alzaben, F.; et al. An Overview of the Public Health Challenges in Diagnosing and Controlling Human Foodborne Pathogens. Vaccines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic use in food animals worldwide, with a focus on Africa: Pluses and minuses. Journal of global antimicrobial resistance 2020, 20, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Pandey, S.; Doo, H.; Keum, G.B.; Kim, E.S.; Kwak, J.; Ryu, S.; Choi, Y.; Kang, J.; Kim, S.; Lee, N.R.; et al. Antibiotic resistance in livestock, environment and humans: One Health perspective. Journal of animal science and technology 2024, 66, 266–278. [Google Scholar] [CrossRef]
- Bava, R.; Castagna, F.; Lupia, C.; Poerio, G.; Liguori, G.; Lombardi, R.; Naturale, M.D.; Mercuri, C.; Bulotta, R.M.; Britti, D.; et al. Antimicrobial Resistance in Livestock: A Serious Threat to Public Health. Antibiotics 2024, 13. [Google Scholar] [CrossRef]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J.; Group, W.H.O.G.D. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrobial resistance and infection control 2018, 7, 7. [Google Scholar] [CrossRef]
- Berman, T.S.; Barnett-Itzhaki, Z.; Berman, T.; Marom, E. Antimicrobial resistance in food-producing animals: Towards implementing a one health based national action plan in Israel. Israel journal of health policy research 2023, 12, 18. [Google Scholar] [CrossRef]
- Scott, H.M.; Acuff, G.; Bergeron, G.; Bourassa, M.W.; Gill, J.; Graham, D.W.; Kahn, L.H.; Morley, P.S.; Salois, M.J.; Simjee, S.; et al. Critically important antibiotics: Criteria and approaches for measuring and reducing their use in food animal agriculture. Annals of the New York Academy of Sciences 2019, 1441, 8–16. [Google Scholar] [CrossRef]
- Michaelis, C.; Grohmann, E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Galgano, M.; Pellegrini, F.; Catalano, E.; Capozzi, L.; Del Sambro, L.; Sposato, A.; Lucente, M.S.; Vasinioti, V.I.; Catella, C.; Odigie, A.E.; et al. Acquired Bacterial Resistance to Antibiotics and Resistance Genes: From Past to Future. Antibiotics 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T.; Aminov, R. Potential Effects of Horizontal Gene Exchange in the Human Gut. Frontiers in immunology 2017, 8, 1630. [Google Scholar] [CrossRef] [PubMed]
- Ghimpeteanu, O.M.; Pogurschi, E.N.; Popa, D.C.; Dragomir, N.; Dragotoiu, T.; Mihai, O.D.; Petcu, C.D. Antibiotic Use in Livestock and Residues in Food-A Public Health Threat: A Review. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiao, L.; Zhou, M.; Zhang, H. The microbiota: A crucial mediator in gut homeostasis and colonization resistance. Frontiers in microbiology 2024, 15, 1417864. [Google Scholar] [CrossRef]
- Berendsen, B.J.; Wegh, R.S.; Memelink, J.; Zuidema, T.; Stolker, L.A. The analysis of animal faeces as a tool to monitor antibiotic usage. Talanta 2015, 132, 258–268. [Google Scholar] [CrossRef]
- Tripathi, A.; Jaiswal, A.; Kumar, D.; Pandit, R.; Blake, D.; Tomley, F.; Joshi, M.; Joshi, C.G.; Dubey, S.K. Whole genome sequencing revealed high occurrence of antimicrobial resistance genes in bacteria isolated from poultry manure. International journal of antimicrobial agents 2025, 65, 107452. [Google Scholar] [CrossRef]
- Cortes, P.; Pokrant, E.; Yevenes, K.; Maddaleno, A.; Flores, A.; Vargas, M.B.; Trincado, L.; Maturana, M.; Lapierre, L.; Cornejo, J. Antimicrobial Residues in Poultry Litter: Assessing the Association of Antimicrobial Persistence with Resistant Escherichia coli Strains. Antibiotics 2025, 14. [Google Scholar] [CrossRef]
- Yevenes, K.; Pokrant, E.; Trincado, L.; Lapierre, L.; Galarce, N.; Martin, B.S.; Maddaleno, A.; Hidalgo, H.; Cornejo, J. Detection of Antimicrobial Residues in Poultry Litter: Monitoring a Risk through a Selective and Sensitive HPLC-MS/MS Method. Animals: An open access journal from MDPI 2021, 11. [Google Scholar] [CrossRef]
- Hazards, E.P.o.B.; Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; et al. Role played by the environment in the emergence and spread of antimicrobial resistance (AMR) through the food chain. EFSA journal. European Food Safety Authority 2021, 19, e06651. [Google Scholar] [CrossRef]
- Gwenzi, W.; Chaukura, N.; Muisa-Zikali, N.; Teta, C.; Musvuugwa, T.; Rzymski, P.; Abia, A.L.K. Insects, Rodents, and Pets as Reservoirs, Vectors, and Sentinels of Antimicrobial Resistance. Antibiotics 2021, 10. [Google Scholar] [CrossRef]
- Fouz, N.; Pangesti, K.N.A.; Yasir, M.; Al-Malki, A.L.; Azhar, E.I.; Hill-Cawthorne, G.A.; Abd El Ghany, M. The Contribution of Wastewater to the Transmission of Antimicrobial Resistance in the Environment: Implications of Mass Gathering Settings. Tropical medicine and infectious disease 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.E.; Laing, G.; McMahon, B.J.; Fanning, S.; Stekel, D.J.; Pahl, O.; Coyne, L.; Latham, S.M.; McIntyre, K.M. The need for One Health systems-thinking approaches to understand multiscale dissemination of antimicrobial resistance. The Lancet. Planetary health 2024, 8, e124–e133. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Kong, L.; Gao, H.; Cheng, X.; Wang, X. A Review of Current Bacterial Resistance to Antibiotics in Food Animals. Frontiers in microbiology 2022, 13, 822689. [Google Scholar] [CrossRef] [PubMed]
- Che, M.; Birk, T.; Hansen, L.T. Prevalence and Transmission of Extended-Spectrum Cephalosporin (ESC) Resistance Genes in Escherichia coli Isolated from Poultry Production Systems and Slaughterhouses in Denmark. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- European Food Safety, A.; European Centre for Disease, P.; Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2021-2022. EFSA journal. European Food Safety Authority 2024, 22, e8583. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiology spectrum 2018, 6. [Google Scholar] [CrossRef]
- Teklemariam, A.D.; Al-Hindi, R.R.; Albiheyri, R.S.; Alharbi, M.G.; Alghamdi, M.A.; Filimban, A.A.R.; Al Mutiri, A.S.; Al-Alyani, A.M.; Alseghayer, M.S.; Almaneea, A.M.; et al. Human Salmonellosis: A Continuous Global Threat in the Farm-to-Fork Food Safety Continuum. Foods 2023, 12. [Google Scholar] [CrossRef]
- Delahoy, M.J.; Shah, H.J.; Weller, D.L.; Ray, L.C.; Smith, K.; McGuire, S.; Trevejo, R.T.; Scallan Walter, E.; Wymore, K.; Rissman, T.; et al. Preliminary Incidence and Trends of Infections Caused by Pathogens Transmitted Commonly Through Food - Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2022. MMWR. Morbidity and mortality weekly report 2023, 72, 701–706. [Google Scholar] [CrossRef]
- Pourzand, F.; Kim, A.H.M.; Chambers, T.; Grout, L.; Baker, M.G.; Hales, S. Examining campylobacteriosis disease notification rates: Association with water supply characteristics. Environmental research 2025, 271, 121064. [Google Scholar] [CrossRef]
- Pietsch, M.; Simon, S.; Meinen, A.; Trost, E.; Banerji, S.; Pfeifer, Y.; Flieger, A. Third generation cephalosporin resistance in clinical non-typhoidal Salmonella enterica in Germany and emergence of bla(CTX-M)-harbouring pESI plasmids. Microbial genomics 2021, 7. [Google Scholar] [CrossRef]
- Matuszewska, M.; Murray, G.G.R.; Ba, X.; Wood, R.; Holmes, M.A.; Weinert, L.A. Stable antibiotic resistance and rapid human adaptation in livestock-associated MRSA. eLife 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Cuny, C.; Wieler, L.H.; Witte, W. Livestock-Associated MRSA: The Impact on Humans. Antibiotics 2015, 4, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Harant, A.; Fernandes, G.; Mwamelo, A.J.; Hein, W.; Dekker, D.; Sridhar, D. Measuring the global response to antimicrobial resistance, 2020–2021: A systematic governance analysis of 114 countries. The Lancet. Infectious diseases 2023, 23, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Hammerum, A.M.; Heuer, O.E.; Emborg, H.D.; Bagger-Skjot, L.; Jensen, V.F.; Rogues, A.M.; Skov, R.L.; Agerso, Y.; Brandt, C.T.; Seyfarth, A.M.; et al. Danish integrated antimicrobial resistance monitoring and research program. Emerging infectious diseases 2007, 13, 1632–1639. [Google Scholar] [CrossRef]
- Yang, Y.; Feye, K.M.; Shi, Z.; Pavlidis, H.O.; Kogut, M.; A, J.A.; Ricke, S.C. A Historical Review on Antibiotic Resistance of Foodborne Campylobacter. Frontiers in microbiology 2019, 10, 1509. [CrossRef]
- Mader, R.; Jarrige, N.; Haenni, M.; Bourely, C.; Madec, J.Y.; Amat, J.P.; Eu, J. OASIS evaluation of the French surveillance network for antimicrobial resistance in diseased animals (RESAPATH): Success factors underpinning a well-performing voluntary system. Epidemiology and infection 2021, 149, e104. [Google Scholar] [CrossRef]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.M.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur resistance in Salmonella enterica serovar Heidelberg from chicken meat and humans, Canada. Emerging infectious diseases 2010, 16, 48–54. [Google Scholar] [CrossRef]
- Yang, H.; Paruch, L.; Chen, X.; van Eerde, A.; Skomedal, H.; Wang, Y.; Liu, D.; Liu Clarke, J. Antibiotic Application and Resistance in Swine Production in China: Current Situation and Future Perspectives. Frontiers in veterinary science 2019, 6, 136. [Google Scholar] [CrossRef]
- Wen, R.; Li, C.; Zhao, M.; Wang, H.; Tang, Y. Withdrawal of antibiotic growth promoters in China and its impact on the foodborne pathogen Campylobacter coli of swine origin. Frontiers in microbiology 2022, 13, 1004725. [Google Scholar] [CrossRef]
- Song, Y.; Han, Z.; Song, K.; Zhen, T. Antibiotic Consumption Trends in China: Evidence From Six-Year Surveillance Sales Records in Shandong Province. Frontiers in pharmacology 2020, 11, 491. [Google Scholar] [CrossRef]
- Tiseo, K.; Huber, L.; Gilbert, M.; Robinson, T.P.; Van Boeckel, T.P. Global Trends in Antimicrobial Use in Food Animals from 2017 to 2030. Antibiotics 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Grave, K.; Torren-Edo, J.; Muller, A.; Greko, C.; Moulin, G.; Mackay, D.; Group, E. Variations in the sales and sales patterns of veterinary antimicrobial agents in 25 European countries. The Journal of antimicrobial chemotherapy 2014, 69, 2284–2291. [Google Scholar] [CrossRef] [PubMed]
- Idoko-Akoh, A.; Goldhill, D.H.; Sheppard, C.M.; Bialy, D.; Quantrill, J.L.; Sukhova, K.; Brown, J.C.; Richardson, S.; Campbell, C.; Taylor, L.; et al. Creating resistance to avian influenza infection through genome editing of the ANP32 gene family. Nature communications 2023, 14, 6136. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, V.; Roviello, G.N. The Potential Role of Vaccines in Preventing Antimicrobial Resistance (AMR): An Update and Future Perspectives. Vaccines 2023, 11. [Google Scholar] [CrossRef]
- Callaway, T.R.; Lillehoj, H.; Chuanchuen, R.; Gay, C.G. Alternatives to Antibiotics: A Symposium on the Challenges and Solutions for Animal Health and Production. Antibiotics 2021, 10. [Google Scholar] [CrossRef]
- Jeffery, A.; Gilbert, M.; Corsaut, L.; Gaudreau, A.; Obradovic, M.R.; Cloutier, S.; Frenette, M.C.; Surprenant, C.; Lacouture, S.; Arnal, J.L.; et al. Immune response induced by a Streptococcus suis multi-serotype autogenous vaccine used in sows to protect post-weaned piglets. Veterinary research 2024, 55, 57. [Google Scholar] [CrossRef]
- Duan, Y.; Hao, Y.; Feng, H.; Shu, J.; He, Y. Research progress on Haemophilus parasuis vaccines. Frontiers in veterinary science 2025, 12, 1492144. [Google Scholar] [CrossRef]
- Blake, D.P.; Vrba, V.; Xia, D.; Jatau, I.D.; Spiro, S.; Nolan, M.J.; Underwood, G.; Tomley, F.M. Genetic and biological characterisation of three cryptic Eimeria operational taxonomic units that infect chickens (Gallus gallus domesticus). International journal for parasitology 2021, 51, 621–634. [Google Scholar] [CrossRef]
- Petro-Turnquist, E.; Pekarek, M.J.; Weaver, E.A. Swine influenza A virus: Challenges and novel vaccine strategies. Frontiers in cellular and infection microbiology 2024, 14, 1336013. [Google Scholar] [CrossRef]
- Hoelzer, K.; Bielke, L.; Blake, D.P.; Cox, E.; Cutting, S.M.; Devriendt, B.; Erlacher-Vindel, E.; Goossens, E.; Karaca, K.; Lemiere, S.; et al. Vaccines as alternatives to antibiotics for food producing animals. Part 2: New approaches and potential solutions. Veterinary research 2018, 49, 70. [Google Scholar] [CrossRef]
- Yan, H.; Yu, B.; Degroote, J.; Spranghers, T.; Van Noten, N.; Majdeddin, M.; Van Poucke, M.; Peelman, L.; De Vrieze, J.; Boon, N.; et al. Antibiotic affects the gut microbiota composition and expression of genes related to lipid metabolism and myofiber types in skeletal muscle of piglets. BMC veterinary research 2020, 16, 392. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.P.; Lisboa, J.A.N.; Bessegatto, J.A.; Montemor, C.H.; Paulino, L.R.; Alfieri, A.A.; Weese, J.S.; Costa, M.C. Impact of virginiamycin on the ruminal microbiota of feedlot cattle. Canadian journal of veterinary research = Revue canadienne de recherche veterinaire 2024, 88, 114–122. [Google Scholar]
- El-Fateh, M.; Bilal, M.; Zhao, X. Effect of antibiotic growth promoters (AGPs) on feed conversion ratio (FCR) of broiler chickens: A meta-analysis. Poultry science 2024, 103, 104472. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ghanbari, M.; May, A.; Abeel, T. Effects of antibiotic growth promoter and its natural alternative on poultry cecum ecosystem: An integrated analysis of gut microbiota and host expression. Frontiers in microbiology 2024, 15, 1492270. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.E.; Ledesma-Amaro, R. The microbial food revolution. Nature communications 2023, 14, 2231. [Google Scholar] [CrossRef]
- Radhi, K.S.; Arif, M.; Rehman, A.U.; Faizan, M.; Almohmadi, N.H.; Youssef, I.M.; Swelum, A.A.; Suliman, G.M.; Tharwat, M.; Ebrahim, A.; et al. Growth performance of broiler chickens fed diets supplemented with amylase and protease enzymes individually or combined. Open veterinary journal 2023, 13, 1425–1435. [Google Scholar] [CrossRef]
- Latif, A.; Shehzad, A.; Niazi, S.; Zahid, A.; Ashraf, W.; Iqbal, M.W.; Rehman, A.; Riaz, T.; Aadil, R.M.; Khan, I.M.; et al. Probiotics: Mechanism of action, health benefits and their application in food industries. Frontiers in microbiology 2023, 14, 1216674. [Google Scholar] [CrossRef]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8. [Google Scholar] [CrossRef]
- Gupta, R.; Sharma, S. Role of alternatives to antibiotics in mitigating the antimicrobial resistance crisis. The Indian journal of medical research 2022, 156, 464–477. [Google Scholar] [CrossRef]
- Klous, G.; Huss, A.; Heederik, D.J.J.; Coutinho, R.A. Human-livestock contacts and their relationship to transmission of zoonotic pathogens, a systematic review of literature. One health 2016, 2, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Makovska, I.; Chantziaras, I.; Caekebeke, N.; Dhaka, P.; Dewulf, J. Assessment of Cleaning and Disinfection Practices on Pig Farms across Ten European Countries. Animals: An open access journal from MDPI 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Reiche, T.; Hageskal, G.; Mares, M.; Hoel, S.; Tondervik, A.; Heggeset, T.M.B.; Haugen, T.; Sperstad, S.B.; Troen, H.H.; Bjorkoy, S.; et al. Shifts in surface microbiota after cleaning and disinfection in broiler processing plants: Incomplete biofilm eradication revealed by robotic high-throughput screening. Applied and environmental microbiology 2025, 91, e0240124. [Google Scholar] [CrossRef]
- Alawa, B.; Singh, S.; Chakma, S.; Kishor, R.; Stalsby Lundborg, C.; Diwan, V. Development of novel biochar adsorbent using agricultural waste biomass for enhanced removal of ciprofloxacin from water: Insights into the isotherm, kinetics, and thermodynamic analysis. Chemosphere 2025, 375, 144252. [Google Scholar] [CrossRef] [PubMed]
- Qadeer, A.; Khan, A.; Khan, N.M.; Wajid, A.; Ullah, K.; Skalickova, S.; Chilala, P.; Slama, P.; Horky, P.; Alqahtani, M.S.; et al. Use of nanotechnology-based nanomaterial as a substitute for antibiotics in monogastric animals. Heliyon 2024, 10, e31728. [Google Scholar] [CrossRef]
- Abreu, R.; Semedo-Lemsaddek, T.; Cunha, E.; Tavares, L.; Oliveira, M. Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Mhlongo, J.T.; Waddad, A.Y.; Albericio, F.; de la Torre, B.G. Antimicrobial Peptide Synergies for Fighting Infectious Diseases. Advanced science 2023, 10, e2300472. [Google Scholar] [CrossRef]
- Wang, G.; Song, Q.; Huang, S.; Wang, Y.; Cai, S.; Yu, H.; Ding, X.; Zeng, X.; Zhang, J. Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella. Animals: An open access journal from MDPI 2020, 10. [Google Scholar] [CrossRef]
- Mancuso, G.; Trinchera, M.; Midiri, A.; Zummo, S.; Vitale, G.; Biondo, C. Novel Antimicrobial Approaches to Combat Bacterial Biofilms Associated with Urinary Tract Infections. Antibiotics 2024, 13. [Google Scholar] [CrossRef]
- Burlec, A.F.; Corciova, A.; Boev, M.; Batir-Marin, D.; Mircea, C.; Cioanca, O.; Danila, G.; Danila, M.; Bucur, A.F.; Hancianu, M. Current Overview of Metal Nanoparticles' Synthesis, Characterization, and Biomedical Applications, with a Focus on Silver and Gold Nanoparticles. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef]
- Moradi, F.; Ghaedi, A.; Fooladfar, Z.; Bazrgar, A. Recent advance on nanoparticles or nanomaterials with anti-multidrug resistant bacteria and anti-bacterial biofilm properties: A systematic review. Heliyon 2023, 9, e22105. [Google Scholar] [CrossRef]
- Danchuk, O.; Levchenko, A.; da Silva Mesquita, R.; Danchuk, V.; Cengiz, S.; Cengiz, M.; Grafov, A. Meeting Contemporary Challenges: Development of Nanomaterials for Veterinary Medicine. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
Figure 1.
Pathways of antibiotic-resistant bacteria. Antibiotics used in veterinary medicine have the potential to contribute to the emergence of antimicrobial resistance (AMR) if not regulated effectively. The mechanisms by which resistance genes are transferred are more rapid in the gut tract of animals due to prolonged contact between different species, the greater number of microorganisms, and their close proximity. The following mechanisms have been identified: conjugation, transduction and transformation. The subsequent transmission of these genes to humans can occur through diverse routes, including but not limited to: direct contact with animals by farm workers, indirect contact with family members and, consequently, the community, vectors such as mice, insects and birds, inadequately treated organic waste used as fertiliser in agriculture or disposed of in the environment, contaminated raw or undercooked food and export of live animals.
Figure 1.
Pathways of antibiotic-resistant bacteria. Antibiotics used in veterinary medicine have the potential to contribute to the emergence of antimicrobial resistance (AMR) if not regulated effectively. The mechanisms by which resistance genes are transferred are more rapid in the gut tract of animals due to prolonged contact between different species, the greater number of microorganisms, and their close proximity. The following mechanisms have been identified: conjugation, transduction and transformation. The subsequent transmission of these genes to humans can occur through diverse routes, including but not limited to: direct contact with animals by farm workers, indirect contact with family members and, consequently, the community, vectors such as mice, insects and birds, inadequately treated organic waste used as fertiliser in agriculture or disposed of in the environment, contaminated raw or undercooked food and export of live animals.
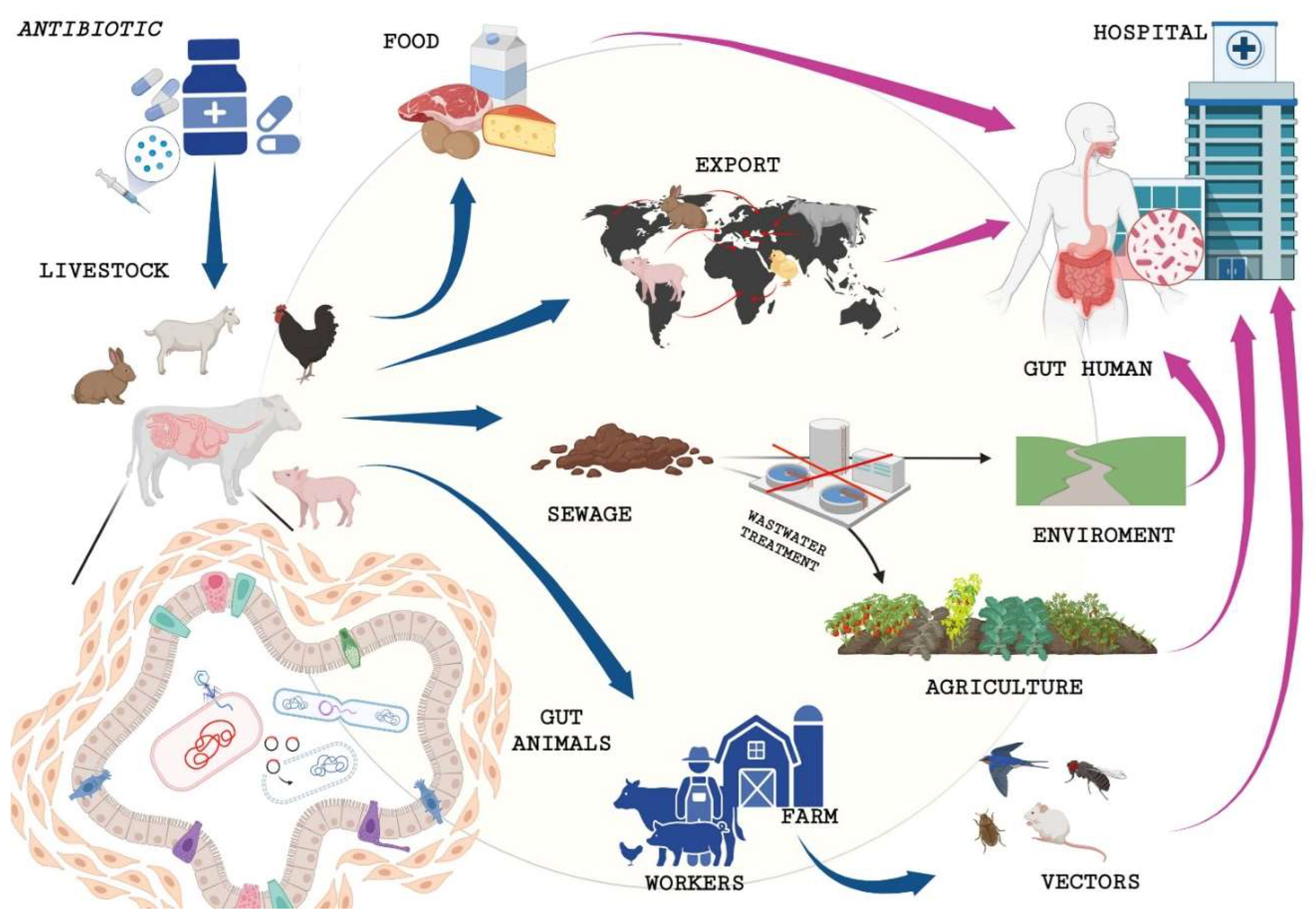
Figure 2.
Global map integrating data on antimicrobial use (AMU) in food-producing animals. The most recent years of recorded AMU values were obtained from the WOAH AMU Data Portal. The 2022 data are available for the following countries: Finland, Ireland, Montenegro, the United Kingdom and Senegal. The 2023 data set is from the following countries: Albania, Belgium, Bosnia and Herzegovina, Bulgaria, Croatia, Denmark, Estonia, France, Iceland, Italy, Latvia, Moldova, Norway, Romania, Slovakia, Slovenia, Spain, Sweden, Switzerland, the Netherlands, Myanmar, Sri Lanka, Bolivia, Canada, Costa Rica, Cuba, St. Vincent and the Grenadines, Cape Verde, the Republic of the Congo, Egypt, Gabon, Kenya and Togo. Countries that use antimicrobials as growth promoters are highlighted, showing if they regulate this use (as for Myanmar, yellow icon) or not (as for Bolivia, Canada and Costa Rica, red icon).
Figure 2.
Global map integrating data on antimicrobial use (AMU) in food-producing animals. The most recent years of recorded AMU values were obtained from the WOAH AMU Data Portal. The 2022 data are available for the following countries: Finland, Ireland, Montenegro, the United Kingdom and Senegal. The 2023 data set is from the following countries: Albania, Belgium, Bosnia and Herzegovina, Bulgaria, Croatia, Denmark, Estonia, France, Iceland, Italy, Latvia, Moldova, Norway, Romania, Slovakia, Slovenia, Spain, Sweden, Switzerland, the Netherlands, Myanmar, Sri Lanka, Bolivia, Canada, Costa Rica, Cuba, St. Vincent and the Grenadines, Cape Verde, the Republic of the Congo, Egypt, Gabon, Kenya and Togo. Countries that use antimicrobials as growth promoters are highlighted, showing if they regulate this use (as for Myanmar, yellow icon) or not (as for Bolivia, Canada and Costa Rica, red icon).
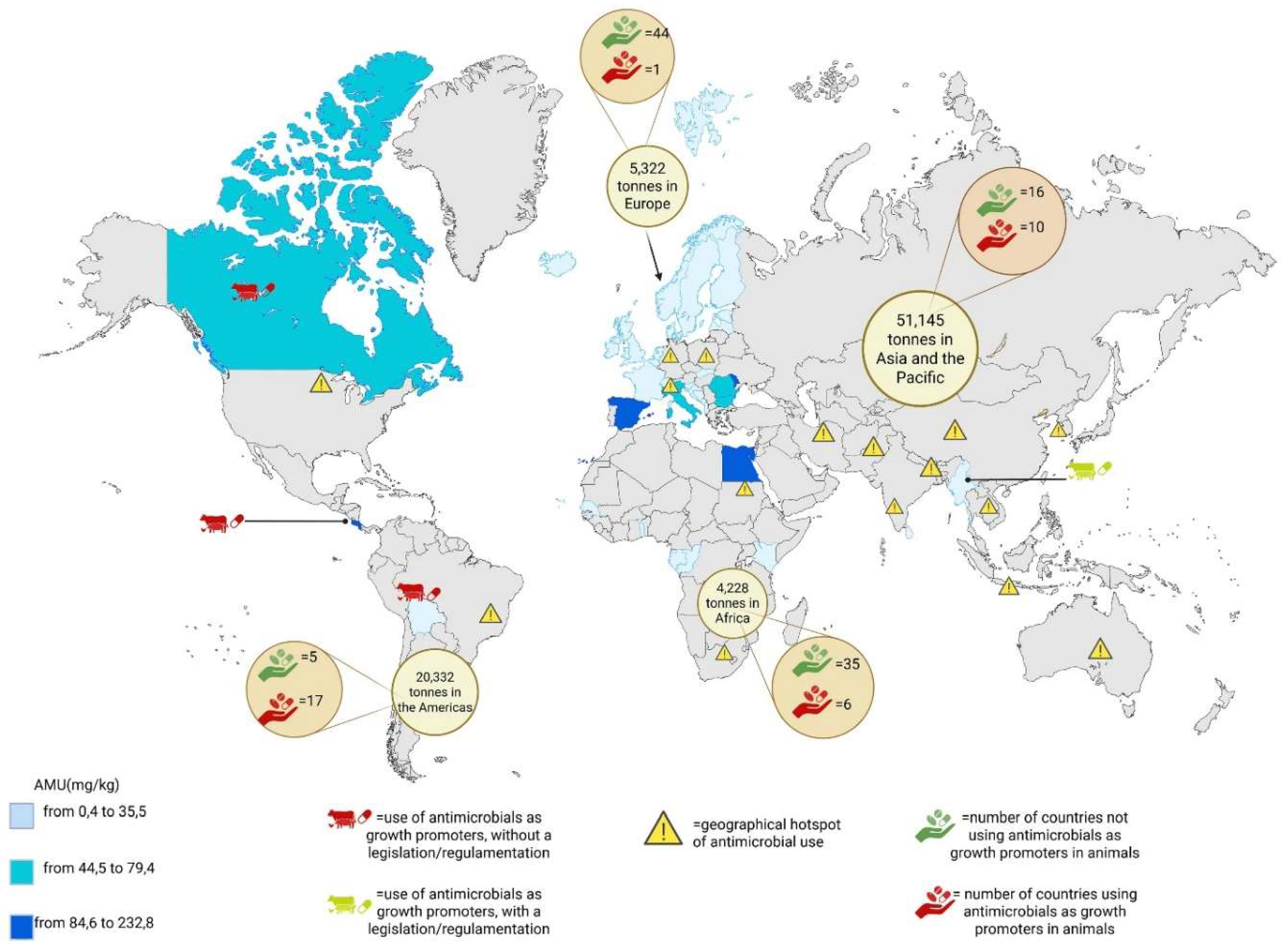
Figure 3.
Strategies for combatting antimicrobial resistance.
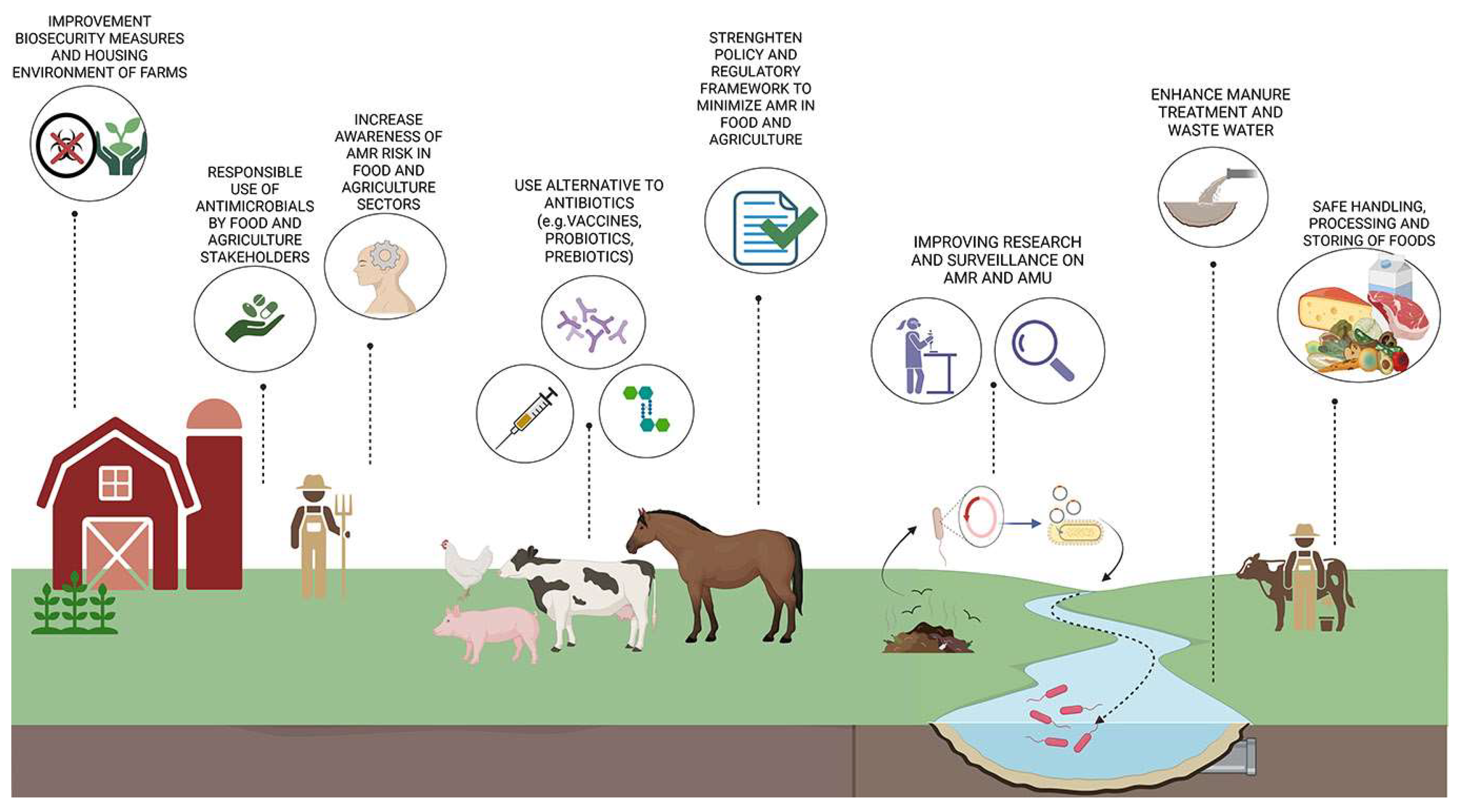

Table 1.
Classes of antibiotics used in veterinary medicine.
| ANTIMICROBIAL AGENTS | SPECIES | USE | MODE OF ACTION | BACTERIAL RESISTANCE |
|---|---|---|---|---|
|
AMINOCOUMARIN Novobiocin |
Avian, Bovine, Caprine, Ovine, Fish |
Novobiocin is for treating mastitis. It is currently only used on animals. |
Novobiocin is a narrow-spectrum antimicrobial. It is bacteriostatic, but may be bactericidal at higher concentrations. It is active mostly against gram-positive bacteria, but also against a few gram-negative bacteria. Can be used on association, for synergistic effect, with tetracyclines. |
Many species of bacteria can develop resistance to novobiocin. |
|
AMINOCYCLITOL Spectinomycin |
Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine |
Used for respiratory infections in cattle and enteric infections in multiple species. |
Spectinomycin belongs to the class of aminocyclic antibiotics. It has been demonstrated to inhibit bacterial protein synthesis by binding to the 30S subunit of the ribosome, thereby causing incorrect reading of the genetic code by tRNA. Spectinomycin exerts bacteriostatic activity. | The resistance of the antibiotic spectinomycin has been linked to the aadA gene, and this association was found to be plasmid-mediated. Its precise location was determined to be within a variant of the integrative and conjugative element ICE Mh1, which encodes an enzyme termed aminoglycoside 3'-adenylate transferase. |
|
AMINOGLYCOSIDES Dihydrostreptomycin Streptomycin |
Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine Bee, Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine |
The broad spectrum of applications renders aminoglycosides a pivotal component of veterinary medicine. Aminoglycosides have a broad spectrum of applications in veterinary medicine, including the treatment of: -septicaemias -digestive infections -respiratory infections -urinary diseases Gentamicin is indicated for Pseudomonas aeruginosa infections, with few alternatives. Apramycin and Fortimycin are currently only used in animals. |
Aminoglycosides have been demonstrated to be effective against rapidly multiplying organisms. The amino groups contribute to the cationic nature of this class of antimicrobials, which is important in mediating the transport across the cytoplasmic membrane and binding with specific anionic sites on LPS. The internalization process is followed by irreversible binding to multiple sites of the 30S ribosomal subunit, as well as the 50S ribosomal subunit, thereby inhibiting protein synthesis. Nevertheless, at low concentrations, all aminoglycosides may be bacteriostatic activity. |
The mechanisms of resistance encompass the process of enzymatic modification of aminoglycosides, which may be either plasmid-encoded or chromosomally mediated. Enzymes have been observed in both gram-negative and gram-positive bacteria. To date, 50 enzymes have been identified, which can be categorized into three major classes: • Acetyltransferases • Nucleotidyltransferases • Phosphotransferases Chemical modification stabilizes the drug, decreasing susceptibility to enzymatic destruction. For instance, amikacin is produced through the chemical modification of kanamycin, which exhibits enhanced resistance to enzymatic hydrolysis. Other resistance mechanisms include higher levels of calcium and magnesium ions, which stop cationic drugs from binding to bacteria. |
|
AMINOGLYCOSIDES + 2 DEOXYSTREPTAMINE Amikacin Apramycin Fortimycin *Framycetin Gentamicin Kanamycin *Neomycin Paromomycin Tobramycin |
Equine Avian, Bovine, Rabbit, Ovine, Swine Bovine, Rabbit, Ovine, Swine Bovine, Caprine, Ovine Avian, Bovine, Camel, Caprine, Equine, Rabbit, Ovine, Swine Avian, Bovine, Equine, Fish, Swine Bee, Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine Avian, Bovine, Caprine, Ovine, Rabbit, Swine Equine |
|||
|
AMPHENICOLS Florfenicol Thiamphenicol |
Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Avian, Bovine, Caprine, Ovine, Fish, Swine |
Phenicol drugs have a wide range of uses, making them very important in veterinary medicine, especially for treating fish diseases. They are also used to treat respiratory infections in cattle, pigs and poultry, with florfenicol used to treat pasteurellosis in these animals. |
This antibiotic is bacteriostatic. It binds to the 50S ribosomal subunit, stopping protein production in bacteria, and may be bactericidal against S. pseudopneumoniae |
Research has identified florfenicol resistance on mobile genetic elements. Florfenicol, a derivative of chloramphenicol, has been found to be resistant to certain chloramphenicol resistance genes. The primary mechanism of resistance to chloramphenicol is enzymatic inactivation. |
|
ANSAMYCIN – RIFAMYCINS Rifampicin Rifaximin |
Equine Bovine, Caprine, Equine, Rabbit, Ovine, Swine |
This antimicrobial class is authorized in a limited number of countries and for a restricted number of indications (mastitis), and there are few alternatives. Rifampicin is employed in the treatment of Rhodococcus equi infections in foals. |
The antibiotic is classed as an extended-spectrum antibiotic. Rifamycins inhibit the synthesis of RNA in microrganisms by binding to DNA-dependent RNA polymerase subunits. They effectively penetrate tissues and cells, making them particularly efficacious in the treatment of intracellular organisms. |
The development of antimicrobial resistance, even when administered in combination with other antimicrobials, is a key concern. |
|
BICYCLOMYCIN Bicozamycin |
Bovine, Fish, Swine |
Bicyclomycin has been included in the list of pharmaceuticals used in the treatment of digestive and respiratory diseases in cattle. | NA | NA |
|
CEPHALOSPORINS FIRST GENERATION Cefacetrile Cefalexin Cefalonium Cefalotin Cefapyrin Cefazolin |
Bovine Avian, Bovine, Caprine, Equine, Ovine, Swine Bovine, Caprine, Ovine Equine Bovine Bovine, Caprine, Ovine |
Cephalosporins are employed in the treatment of the following conditions: • Septicemia • Respiratory infections • Mastitis |
Cephalosporins are categorized within the class of beta-lactam antibiotics. These antibiotics disrupt the process of bacterial cell wall formation by interacting with a group of proteins known as the penicillin-binding proteins (PBPs). These transpeptidase enzymes are responsible for the formation of cross-links between peptidoglycan strands. It is noteworthy that the antibiotic activity of beta-lactams is confined to organisms in the log phase of growth or during active multiplication. Consequently, static bacteria are unaffected and may persist. However, at appropriate concentrations, they are bactericidal to most bacteria. Cephalosporins are weak acids that are derived from 7-minocephalosporanic acid. Modifications at this nucleus, in addition to substitutions on the side chains, result in differences among cephalosporins with respect to their antibacterial spectra, beta-lactamase sensitivities and pharmacokinetics. |
The mechanisms of antimicrobial resistance are multifactorial and common to all beta-lactam antibiotics. These mechanisms may comprise one or more combinations of different types of resistance, including: • modification of the drug target, via alterations to the PBPs (of which there are at least nine different types on the cell wall, with differences in PBP targets); • downregulation of porins, with reduction in cell permeability; • increased expression of efflux pumps; • production of degrading enzymes (cephalosporinase). The beta-lactam nucleus of the cephalosporins is susceptible to hydrolysis by beta-lactamase enzymes. This reaction, in turn, is known to result in the cleavage of the beta-lactam nucleus. This, in turn, leads to the inactivation of the beta-lactam, which, ultimately, results in the failure of the drug to bind to their target PBPs. It is estimated that more than 800 beta-lactamases have been documented, a figure which reflects the mounting pressure resulting from the increased use of beta-lactam. Inhibitors of beta-lactamases, such as clavulanic acid and sulbactam, have been introduced to inhibit beta-lactam degradation. |
|
CEPHALOSPORINS SECOND GENERATION Cefuroxime |
Bovine |
|||
|
CEPHALOSPORINS THIRD GENERATION Cefoperazone Ceftiofur Ceftriaxone |
Bovine, Caprine, Ovine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine Bovine, Ovine, Swine |
The wide range of applications make cephalosporin third and fourth generation extremely important for veterinary medicine. These cephalosporins are employed in the management of: -septicaemias -respiratory infections -mastitis Alternatives are characterized by limited efficacy, either due to inadequate spectrum or the presence of antimicrobial resistance. |
||
|
CEPHALOSPORINS FOURTH GENERATION Cefquinome |
Bovine, Caprine, Equine, Rabbit, Ovine, Swine |
|||
|
FUSIDANE Fusidic acid |
Bovine, Equine |
Fusidic acid is employed in the treatment of ophthalmic diseases in cattle and horses. |
Fusidic acid has been demonstrated to interfere with the function of elongation factor and to inhibit protein synthesis at the 50S subunit of the ribosome. It is noteworthy that the action of fusidic acid is bacteriostatic against gram-positive organisms, and bactericidal against Staphylococcus aureus. |
Fusidic acid resistance can develop quickly. |
|
IONOPHORES Lasalocid Maduramycin Monensin Narasin Salinomycin Semduramicin |
Avian, Bovine, Rabbit, Ovine Avian Bee, Avian, Bovine, Caprine Avian, Bovine Avian, Rabbit, Bovine, Swine Avian |
Ionophores are of critical importance to the health of animals, particularly in the context of controlling intestinal parasitic coccidiosis (Eimeria spp.), a condition for which there are limited treatment options when alternative remedies are unavailable. In the context of poultry, ionophores play a pivotal role. It is important to note that the use of ionophores is currently limited to animal husbandry. |
Ionophores are lipid-soluble molecules that can bind and transport ions across cell membranes. Inside the cell, ionophores change the concentration of different types of ions, including calcium, potassium, hydrogen and sodium. This can block protein transport, reduce metabolic activity and kill the organism. |
Resistance to ionophores is probably adaptive, not due to mutation or gene acquisition. |
|
LINCOSAMIDES Lincomycin Pirlimycin |
Bee, Avian, Bovine, Caprine, Ovine, Fish, Swine Bovine, Swine |
In the context of treating swine, lincosamides have been found to be essential for treating the following conditions: • Mycoplasmal pneumonia • infectious arthritis • haemorrhagic enteritis |
Lincosamides are antibiotics that exclusively bind to the 50S subunit of bacterial ribosomes, suppressing protein synthesis via inhibition of peptidyl transferases. The antibacterial potency of lincosamides varies by concentration, being bacteriostatic at lower concentrations and bactericidal at higher concentrations. Lincosamides are also contingent upon bacterial load and species. |
Resistance to lincosamides develops gradually, due to plasmid or chromosomal methylation of the ribosomal subunit. Other mechanisms include increased efflux pump activation and drug destruction. |
|
MACROLIDES 14- MEMBERED RING Erythromycin Oleandomycin |
Bee, Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Bovine |
The extensive range of applications of macrolides renders them of significant importance in the domain of veterinary medicine. The following conditions are treated with macrolides: -Mycoplasma infections in pigs and poultry; -haemorrhagic digestive disease in pigs (Lawsonia intracellularis); -liver abscesses (Fusobacterium necrophorum) in cattle. In all these conditions, macrolides represent the only effective treatment available. This class of antibiotics is also used for respiratory infections in cattle. |
There are various classes of macrolides distinguished by the size of their ring. All classes of macrolides share a common mechanism that inhibits protein synthesis by binding to the 50S ribosomal subunit, which is similar to how phenicol antibiotics work. These antibiotics block the translocation process that helps the peptide chain grow, which is essential for life. Macrolides are classified as bacteriostatic drugs, but at high concentrations they can be bactericidal. | Macrolide resistance falls into three main categories: intrinsic, acquired, or constitutive/inducible. Intrinsic resistance is linked to the cell wall of gram-negative bacteria, which is impermeable to water. Gram-positive bacteria can demonstrate resistance through alterations in ribosomal structure (e.g. through target site methylation or mutation). Post-translational methylation can result in cross-resistance to lincosamides and streptogramins (macrolide-lincosamide-streptogramin B, or MLSB, resistance). A further mechanism of resistance is efflux from cells. Finally, although rare, the inactivation of the drug itself can be a factor. The development of resistance may be rapid or gradual and resistance may also be cross-resistant between macrolides. |
|
MACROLIDES 15- MEMBERED RING Gamithromycin Tulathromycin |
Bovine Bovine, Swine |
|||
|
MACROLIDES 16- MEMBERED RING Carbomycin Josamycin Kitasamycin Mirosamycin Spiramycin Terdecamycin Tildipirosin Tilmicosin Tylosin Tylvalosin |
Avian Fish, Swine Avian, Swine, Fish Bee, Avian, Swine, Fish Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Swine Bovine, Swine Avian, Bovine, Caprine, Rabbit, Ovine, Swine Bee, Avian, Bovine, Caprine, Rabbit, Ovine, Swine Avian, Swine |
|||
|
MACROLIDES C17 Sedecamycin |
Swine |
|||
|
ORTHOSOMYCINS Avilamycin |
Avian, Rabbit, Swine |
Avilamycin is employed in the treatment of enteric diseases affecting poultry, swine and rabbits. Presently, this class of compounds is exclusively employed in veterinary medicine. |
Avilamycin is an antibiotic that stops proteins being built by blocking the A-tRNA site of ribosomal RNA 50S. It works against gram-positive bacteria but not gram-negative ones. This is probably because gram-negative bacteria have different ways of defending themselves. |
The molecular mechanisms underlying the resistance of avilamycin appear to be associated with mutations in single nucleotides at positions H89 and H91 of the ribosomal RNA 50S helices. |
|
NATURAL PENICILLINS (including esters and salts) Benethamine penicillin Benzylpenicillin Benzylpenicillin procaine / Benzathine penicillin Penethamate (hydroiodide) |
Bovine Avian, Bovine, Caprine, Camel, Equine, Rabbit, Ovine, Swine Avian, Bovine, Camel, Caprine, Equine, Ovine, Swine Bovine |
Penicillins are of significant importance in the field of veterinary medicine due to their wide range of applications. Penethamate (hydroiodide) is currently only used in vet medicine. This group of antibiotics has been proven effective in treating sepsis, respiratory infections, and urinary tract infections. Few economical alternatives are available. |
Penicillins of this class have been demonstrated to be effective against a broad spectrum of microorganisms. These include many types of gram-positive and aerobic bacteria, as well as a limited number of gram-negative bacteria, such as Haemophilus and Neisseria species, and certain strains of Bacteroides, with the exception of B. fragilis. Broad-spectrum penicillins are derived semisynthetically and demonstrate efficacy against a wide range of gram-positive and gram-negative bacteria. Semisynthetic penicillins with extended spectra have been demonstrated to be efficacious against Pseudomonas aeruginosa, some Proteus spp and, in a limited number of cases, even strains of Klebsiella, Shigella and Enterobacter spp.. |
Organisms devoid of a cell wall, such as Mycoplasma, possess an inherent resistance to beta-lactam antimicrobials. Penicillins are susceptible to hydrolysis by beta-lactamases, also known as penicillinases. Broad-spectrum penicillins are classified as sensitive beta-lactamases. The combination of broad-spectrum penicillins and beta-lactamase inhibitors has been demonstrated to enhance the efficacy of treatment against both gram-positive and gram-negative pathogens. For instance, the combination of clavulanate-associated amoxicillin has been shown to enhance its efficacy. Semisynthetic penicillins with extended spectra are frequently characterised by a certain degree of beta-lactamase resistance and are typically effective against one or more characteristic penicillin-resistant organisms. |
|
AMDINOPENICILLINS Mecillinam |
Bovine, Swine |
|||
|
AMINOPENICILLINS Amoxicillin Ampicillin Hetacillin |
Avian, Bovine, Caprine, Equine, Ovine, Fish, Swine Avian, Bovine, Caprine, Equine, Ovine, Fish, Swine Bovine |
|||
|
AMINOPENICILLIN + BETALACTAMASE INHIBITOR Amoxicillin + Clavulanic Acid Ampicillin + Sulbactam |
Avian, Bovine, Caprine, Equine, Ovine, Swine Bovine, Swine |
|||
|
CARBOXYPENICILLINS Ticarcillin Tobicillin |
Equine Fish |
|||
|
UREIDOPENICILLIN Aspoxicillin |
Bovine, Swine |
|||
|
PHENOXYPENICILLINS Phenethicillin Phenoxymethylpenicillin |
Equine Avian, Swine |
|||
|
ANTISTAPHYLOCOCCAL PENICILLINS Cloxacillin Dicloxacillin Nafcillin Oxacillin |
Bovine, Caprine, Equine, Ovine, Swine Bovine, Caprine, Ovine, Avian, Swine Bovine, Caprine, Ovine Bovine, Caprine, Equine, Ovine, Swine |
|||
|
PHOSPHONIC ACID DERIVATIVES Fosfomycin |
Avian, Bovine, Fish, Swine |
Fosfomycin is a medication that plays a crucial role in the treatment of some fish infections. However, the limited availability of this pharmaceutical product in numerous countries leads to its classification as a Very Hard to Import (VHIA) medication. | Fosfomycin functions as a competitive inhibitor of the substrate phosphoenolpyruvate (PEP) binding to the enzyme UDP-GlcNAc enolpyruvyl transferase (MurA). MurA catalyses the first committed step in bacterial peptidoglycan biosynthesis, which is thereby inhibited by the binding of fosfomycin via an irreversible covalent bond. | The following mechanisms are representative of resistance: • Drug inactivating enzymes. Several enzymes have been detected capable of modifying fosfomycin, including FosA, FosB, FosC and FosX. Of these, FosA has been found to be the most prevalent. • Reduced absorption, associated with mutation in genes involved in the GlpT and UhpT transport systems. In contrast, mutations in chromosomal genes or the over-expression of the target protein manifest less frequently. |
|
PLEUROMUTILINS Tiamulin Valnemulin |
Avian, Caprine, Rabbit, Ovine, Swine Swine |
The class of pleuromutilins is imperative in combating respiratory infections in both pigs and poultry. Moreover, this class of compounds has been demonstrated to be efficacious in the treatment of swine dysentery. However, the limited availability of these medications in certain countries has led to their classification as VHIA. |
The pleuromutilin antibiotics tiamulin and valnemulin target the bacterial cell wall. These antibiotics are active against gram-positive bacteria, mycoplasmas and anaerobes. They inhibit protein synthesis at the bacterial ribosome, preventing the polypeptide chain from growing. In therapeutic concentrations, pleuromutilin exerts a bacteriostatic effect. |
The development of resistance to pleuromutilins occurs through mutations to chromosomal targets. These occur in the 23S rRNA and rplC genes linked to bacterial ribosomes. Mutations are not transferred horizontally and take time to appear. |
|
POLYPEPTIDES Bacitracin Enramycin Gramicidin |
Avian, Bovine, Rabbit, Swine, Ovine Avian, Swine Equine |
Bacitracin is employed in the treatment of necrotic enteritis in poultry. The following diseases and conditions are treated with this class of antibiotics: -septicaemias -colibacillosis -salmonellosis -urinary infections |
The mechanism of action of this antibiotic involves interference with cell membrane function, suppression of cell wall formation via prevention of peptidoglycan strand formation, and inhibition of protein synthesis. It has been demonstrated to possess bactericidal activity, and the presence of divalent cations, such as zinc, is a prerequisite for its activity. |
Rare resistance cases reported. |
|
POLYMYXINS Polymixin B *Polymixin E (colistin) |
Bovine, Caprine, Equine, Rabbit, Ovine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine |
Polymyxin E (colistin) is employed in the treatment of Gram-negative enteric infections. |
Polymyxins are a group of antibiotics that have bactericidal properties. Their efficacy is attributable to their capacity to disrupt the phospholipid composition of bacterial cell membranes, thereby markedly compromising their permeability and functionality. The polymyxins have been shown to be more effective against gram-negative bacteria than gram-positive bacteria. |
The occurrence of resistance to antibiotics at the polymyxis is an exceptionally rare phenomenon, and it was hypothesised that this was dependent on chromosome mutations. This would limit the transmission to vertical dissemination. However, subsequent research has invalidated this hypothesis. This is due to the discovery of a novel and highly conjugable plasmid-mediated gene, designated mcr-1, which confers colistin resistance. |
|
QUINOLONES FIRST GENERATION Flumequin Miloxacin Nalidixic acid Oxolinic acid |
Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Fish Bovine Avian, Bovine, Rabbit, Fish, Swine, Ovine |
The following conditions are indicated for treatment with quinolones of the 1st generation: • Septicaemias • Colibacillosis. |
Quinolones are a class of antibiotics that demonstrate bactericidal properties. These antibiotics possess a multitude of structurally analogous ring frameworks and demonstrate several shared characteristics. The quinolones inhibit bacterial enzyme topoisomerases, in particular: - Topoisomerase II, also known as DNA gyrase, which facilitates single-strand nicks in the DNA that underpin coiling and uncoiling - Topoisomerase IV, which plays a role in unravelling DNA as chromosomes separate. Subsequently, the reduction in inhibition results in a reduction of supercoiling. This, in turn, leads to the disruption of the spatial arrangement of DNA molecules and the failure of DNA repair mechanisms. |
The classic chromosomal resistance mechanism exhibited by bacteria involves specific target mutations in GyrA/GyrB (DNA gyrase, characteristic of gram-negative bacteria) and ParC/ParE (topoisomerase IV, characteristic of gram-positive bacteria). An alternative mechanism of resistance is the combined effect of increased expression of efflux pumps and decreased expression of porins (AcrAB and MexAB gene mutations), leading to decreased intracellular concentrations. In addition to these mechanisms, the presence of fluoroquinolone-resistant proteins (qnrA, qnrB, qnrS) encoded by transmissible plasmids has been confirmed. Qnr encodes proteins capable of protecting DNA gyrase and topoisomerase IV from these antimicrobials. Other resistance mechanisms have been described, including AAC(6')-Ib-cr, which codes an enzyme (acetylase) capable of inactivating antimicrobials of two classes (aminoglycosides and fluoroquinolones). It has been demonstrated that ciprofloxacina e norfloxacina are the only antimicrobials susceptible to inactivation by this enzyme. The qepA gene, located on a plasmid, has been identified as the causative agent of this mechanism and is responsible for the production of an efflux pump capable of only hydrophilic fluoroquinolones, such as norfloxacin and ciprofloxacin. The utilisation of these medications has been demonstrated to foster the emergence of resistance. |
|
QUINOLONES SECOND GENERATION (FLUOROQUINOLONES) Ciprofloxacin Danofloxacin Difloxacin Enrofloxacin Marbofloxacin Norfloxacin Ofloxacin Orbifloxacin Sarafloxacin |
Avian, Bovine, Swine Bovine, Caprine, Rabbit, Ovine, Swine Avian, Bovine, Rabbit, Swine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Bovine, Equine, Rabbit, Swine Avian, Bovine, Caprine, Rabbit, Ovine, Swine Avian, Swine Bovine, Swine Fish |
The wide range of applications make fluoroquinolones extremely important for veterinary medicine. Fluoroquinolones are critically important in the treatment of: • Septicaemias • Respiratory infection • Enteric diseases |
||
|
QUINOXALINES Carbadox Olaquindox |
Swine Swine |
Quinoxalines (carbadox) is employed in the treatment of digestive diseases affecting swine, including swine dysentery. Presently, this class of compounds is exclusively employed in veterinary medicine. |
NA | NA |
|
SULFONAMIDES Phthalylsulfathiazole Sulfacetamide Sulfachlorpyridazine Sulfadiazine Sulfadimethoxazole Sulfadimethoxine Sulfadimidine (Sulfamethazine, Sulfadimerazine) Sulfadoxine Sulfafurazole Sulfaguanidine Sulfamerazine Sulfamethoxine Sulfamonomethoxine Sulfanilamide Sulfapyridine Sulfaquinoxaline |
Swine Avian, Bovine, Ovine Avian, Bovine, Swine Avian, Bovine, Caprine, Ovine, Swine Avian, Bovine, Swine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine Avian, Bovine, Equine, Ovine, Swine Bovine, Fish Avian, Caprine, Ovine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine Avian, Fish, Swine Avian, Fish, Swine Bovine, Caprine, Ovine Bovine, Swine Avian, Bovine, Caprine, Rabbit, Ovine |
Sulfonamides are of significant importance in the field of veterinary medicine due to the wide variety of applications for which they are utilised. These classes alone or in combination are critically important in the treatment of a wide range of infections: • Bacterial • Coccidial • Protozoal |
Sulfonamides are derivatives of sulfanilamide, replacing various functional groups with the amido group. They are structurally similar to PABA (para-aminobenzoic acid). They competitively inhibit dihydropterate synthetase enzyme (DPS), interrupting the synthesis of dihydrofolic acid, precursor of the folic acid. Folic acid is coenzyme implicated in the synthesis of nucleic acids, whose absence leads to blockage of several enzymes present in this metabolic pathway. They have action bacteriostatic, but can be bactericidal at the high concentrations. Diaminopyrimidines, such as trimethoprim, I am able from inhibit dihydrofolate reductase, which is further into the folic acid synthesis pathway. The combination with sulfonamide is synergistic, is enhance the action antibiotic. Diaminopyrimidines are an antibiotics classes able to inhibit dihydrofolate reductase in bacteria and protozoa. However, alone these antibiotics are not effective against bacteria, furthermore can be resistance develops rapidly. For which they are frequently used on combined with sulfonamides, with action bactericidal. |
The phenomenon of resistance to sulphonamides can be categorised into two distinct types: intrinsic, chromosomally mediated, and acquired, plasmid-mediated. The chromosomal mechanism of resistance is attributable to mutations in the genes encoding dihydropterate synthetase. While the resistance plasmid mediated involves mutations in dihydrofolate reductases. The latter causing high-level resistance to trimethoprim. |
|
SULFONAMIDES+ DIAMINOPYRIMIDINES Ormetoprim+ Sulfadimethoxine Sulfamethoxypyridazine Trimethoprim+ Sulfonamide |
Avian, Fish Avian, Bovine, Equine, Swine Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Fish, Swine |
|||
|
DIAMINOPYRIMIDINES Baquiloprim Ormetoprim Trimethoprim |
Bovine, Swine Avian Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine |
|||
|
STREPTOGRAMINS Virginiamycin |
Avian, Bovine, Ovine, Swine |
Virginiamycin is a significant antimicrobial agent that plays a crucial role in the prevention of necrotic enteritis (Clostridium perfringens). |
These antibiotics bind to the 50S subunit of bacterial ribosomes to stop protein production by blocking peptidyl transferases. |
Cross-resistance with macrolides and lincosamides is classified as macrolide-lincosamide-streptogramin B (MLSB) resistance due to their similar mechanisms. |
|
TETRACYCLINES Chlortetracycline Doxycycline *Oxytetracycline Tetracycline |
Avian, Bovine, Caprine, Equine, Rabbit, Ovine, Swine Avian, Bovine, Camel, Caprine, Equine, Rabbit, Ovine, Fish, Swine Bee, Avian, Bovine, Camel, Caprine, Equine, Rabbit, Ovine, Fish, Swine Bee, Avian, Bovine, Camel, Caprine, Equine, Rabbit, Ovine, Fish, Swine |
The broad spectrum of applications renders tetracyclines of primary importance in veterinary medicine. This class of antibiotics is of particular significance in the treatment of chlamydial infections. Moreover, tetracyclines are of critical importance in the treatment of animals against heartwater (Ehrlichia ruminantium) and anaplasmosis (Anaplasma marginale), due to the absence of antimicrobial alternatives. |
Tetracyclines are a class of antibiotic that have the capacity to bind reversibly to the bacterial 30S ribosomal subunit. It has been demonstrated that these antibiotics are capable of impeding the process of ribosomal translation at the aminoacyl-tRNA acceptor (A) site on the mRNA ribosomal complex. While these antibiotics generally act as bacteriostatic agents, at elevated concentrations, they can attain bactericidal properties. It is also notable that doxycycline and minocycline have the capacity to inhibit metalloproteinases. |
The phenomenon of resistance to antibiotics is associated with two distinct mechanisms. The first of these mechanisms is characterised by the acquisition of efflux pumps, plasmid or transposon-mediated. These mechanisms can be transferred either via conjugation or transduction. The second mechanism is attributed to the synthesis of a protective protein, which functions by either preventing binding at the ribosomal target. |
|
THIOSTREPTON Nosiheptide |
Swine |
This class is currently used in the treatment of some dermatological conditions. |
Nosiheptide, an antibiotic, functions by impeding the formation of the 70S initiation complex. It has been demonstrated that Nosiheptide exerts its inhibitory effect on the following processes: -The initiation G protein IF2 -Elongation, by interfering with the G proteins EF-Tu, which is required for the rapid binding of the aminoacyl-tRNA to the ribosome -EF-G, which catalyzes the translocation of the tRNA-mRNA complex from the A and P sites to the P and E sites. |
The mechanism of resistance exhibited by nosiheptide is attributable to target site modification, a process catalysed by an enzyme action of a methyltransferase (NHR) belonging to the class SpoU. This results in resistance to the antibiotic nosiheptide through 2'O-methylation of the 23S rRNA at the nucleotide A1067, where the elongation (F-G) factor performs its function. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



