Submitted:
17 April 2025
Posted:
19 April 2025
You are already at the latest version
Abstract
The metalloid tellurium (Te) is toxic to bacteria; however, the element is also extremely rare. Thus, most bacteria will never encounter Te in their environment. Nonetheless significant research has been performed on bacterial Te resistance, due to the medical applications of the element. The so-called “tellurium resistance (TeR) genes” were first described on plasmids isolated from clinically relevant Enterobacteriaceae. With time, it has become apparent that, given the rarity of Te on the planet, the primary functions of these genes are distinct from their originally described TeR activity. Nonetheless, the description of these genes as “tellurium resistance genes” has persisted. In this review, we examine the history and discovery of the TeR genes. An analysis of 137,000 high-quality (meta)genomes revealed terZABCDF, telA, and tehAB to be relatively common among bacterial genome annotations, and they frequently described as “tellurium resistance genes”. We synthesized the literature to describe the functions of these ubiquitous genes beyond TeR. These genes have functions in diverse cellular processes including phage resistance, antibiotic resistance, cell cycle regulation, manganese resistance, and metalation of exoenzymes. Considering this analysis, we propose that it is time to rethink the primary functions of the so-called “tellurium resistance genes”.
Keywords:
metalloids
; metals
; tellurium
; antibiotic resistance
; bacteriology
; microbiology
1. Introduction
The metalloid tellurium (Te) has a fascinating history within the field of microbiology. On average, the element is extremely rare in Earth’s crust, comparable to the precious metal platinum (Pt) [1]. While there are Te hotspots, for example around hydrothermal vents or associated with gold mine tailings [1], most microorganisms will never encounter sufficiently high enough Te concentrations in their local environment to experience Te-induced stress. However, in 1912, Conradi and Touch began using the Te oxyanion tellurite as a selective medium component to isolate and identify strains of the human pathogen Corynebacterium diphtheriae [2]. This ultimately led to the development of tellurite blood agar, which remains in usage today for isolating and differentiating C. diphtheriae strains [3]. Tellurite is also part of a modified MacConkey medium [4] for selection of verocytotoxigenic Escherichia coli. Historically, tellurite saw usage as an antimicrobial to treat patients with diphtheria and syphilis [5] Thus, was born the microbiology field’s interest in both this rare element’s mechanism of toxicity and microorganisms’ mechanisms of resistance.
In 1977, Summers and Jacoby described plasmids from E. coli and Pseudomonas aeruginosa that conferred increased resistance to tellurite, with this resistance phenotype called “TeR” [6]. Additional plasmids associated with TeR were identified in various members of the Enterobacteriaceae [7,8,9]. Soon after, the genes responsible for the plasmid-encoded TeR phenotype were sequenced, leading to the identification of the terABCDE operon in 1988 by Jobling and Ritchie [10,11] (later expanded to terZABCDEF [12]), followed by the tehAB [13] in 1992 by Walter and Taylor, and telAB genes in 1991 by Walter, Taylor, and colleagues [14]. While these genes were originally considered to be plasmid-encoded resistance determinants, homologs have been identified throughout the chromosomes of diverse microorganisms [15]. Additional TeR genes were described on the chromosome of Rhodobacter sphaeroides 2.4.1., referred to as trgAB [16].
The mechanisms of TeR for these genes are enigmatic. Recent research of tellurium stress in microorganisms has shifted focus towards the basal metabolic and stress response genes important to the cellular response to Te exposure [17]. Since the initial reports of the TeR genes, others have described alternative functions that range from general stress responses to essential metal homeostasis. However, the original annotations (as TeR genes) have been retained within the literature and in databases [18,19], obscuring the reality that these genes are unlikely to have primary functions in TeR given the extreme rarity of the element. While we contend that these genes have primary functions unrelated to tellurium resistance, for the sake of brevity, we will collectively refer to them as “TeR genes” in this review.
2. Ubiquity of Ter Genes in Genome Annotations
We examined the prevalence of the TeR across bacterial genome annotations by analyzing all bacterial (meta)genomes available within JGI’s IMG/M database as of July 29th, 2024 (137,572 in total). We searched these annotations for the names of each TeR gene products. We searched by annotation text rather than homology since researchers often rely upon annotations to predict the functions of the protein products within novel (meta)genomes (Table 1). The most common encoded TeR proteins were TerC (annotated in 82.1% of genomes), TerB (14.3%) and TerD (14.0%) (Table 1). For comparison, gene products annotated as CzcA (the colbalt/zinc/cadmium efflux pump) are found in 24.9% of these same genomes.
From the TerC counts alone, we concluded that at least eight out of ten sequenced bacterial genomes will have one of the TeR genes named in their annotations. Concerningly, most of the annotations not only give the gene product name (e.g. “TerC”) but also explicitly link the protein to tellurite/tellurium resistance (e.g., “tellurite resistance protein TerC”), which would lead researchers unfamiliar with these genes to conclude that their functions are primarily in Te resistance. Due to the relative ubiquity of many of these genes in bacterial genomes, there is a need to describe their functions beyond TeR. For the purposes of this review, we have chosen to focus on the TeR genes most common in bacterial (meta)genome annotations: terZABCDF, telA, and tehAB.
3. Primary Functions of the TER Genes
TehA is a multidrug efflux pump involved in virulence while TehB is a methyltransferase.
The tellurite resistance determinant tehAB operon was discovered near the terminus of the E. coli K-12 chromosome [20]. tehA encodes an inner membrane protein whereas tehB encodes a methyltransferase. When overexpressed, both tehA and tehB increased resistance of E. coli K-12 to tellurite [21,22]. Despite TehA being a membrane-associated protein, Turner et al. (1995) found that it has no impact on tellurite import or efflux [23]. Deletion of tehB in Haemophilus influenzae increased the sensitivity of the strain of tellurite relative to the wild-type strain [24]. Purified E. coli K-12 TehB has methylation activity with tellurite, selenite, selenium dioxide, and selenate [25] (Figure 1).
TehA—as determined from the crystal structure of a H. influenzae homolog—is a homotrimer with each monomer consisting of ten transmembrane helices. [26] In contrast, TehB is a soluble cytoplasmic protein. The crystal structure of E. coli K-12 TehB revealed an α/β/α protein with seven β strand and six α helices [25]. TehB has three conserved motifs that are involved in the binding of the cofactor S-adenosyl-l-methionine (SAM) [25], consistent with the observation that TehB-catalyzed detoxification of tellurite in E. coli cells is SAM-dependent [22].
The biochemical mechanism of TehAB-mediated tellurite resistance is unclear [20,27,28]. When natively expressed in E. coli K-12, tehAB are phenotypically “silent” with respect to tellurite resistance. Increased tellurite resistance is only observed when these genes are overexpressed on multicopy plasmids [29]. In E. coli K-12 other phenotypes have been associated with the overexpression of the tehAB genes. TehA is homologous to small multidrug resistance family transporters, which are involved in the efflux of quaternary ammonium compounds and dyes [30,31,32]. Overexpression of tehA altered E. coli resistance to lipophilic quaternary ammonium compounds and intercalating dyes [21]. tehA overexpression increased sensitivity to dequalinium chloride and methyl viologen (compounds with two quaternary cations) and resistance to tetraphenylarsonium, ethidium bromide, crystal violet, and proflavine (compounds with a single quaternary cation) [21]. Interestingly, the simultaneous overexpression of tehB and tehA eliminated the ethidium bromide resistance phenotype conferred by overexpression of tehA alone by inhibiting the transport of ethidium bromide [21]. We speculate that TehB may act as a regulatory element through its methyltransferase activity that either directly or indirectly regulates the activity of TehA [33]. While the above-described phenotypes were dependent upon overexpression of tehA and/or tehB, the deletion of a chromosomal region in E. coli containing tehAB reduced the rate of ethidium efflux, supporting a physiological function for TehA in the efflux of these compounds [21] (Figure 1).
Similar to other multi-drug efflux pumps, TehA may have more generalizable stress response functions that aid in the colonization of a host environment [34,35,36]. During the process of infection, reactive oxygen species (ROS) and reactive nitrogen species (RNS) are important host defense mechanisms. In E. coli, a binding site for the nitrosative stress negative regulator YjeB is present in the tehA promoter [37], and nitric oxide (NO) increases expression of the tehAB operon [38]. Other multi-drug efflux pumps have been implicated in ROS and RNS stress responses [36,39,40,41,42]. Similar to these other efflux pumps, TehA may export toxic, nitrosated (or oxidized) biomolecules produced during periods of oxidative or nitrosative stress [4,34,36,39] (Figure 1). TehB has also been linked to resistance to chemically diverse oxidants. A H. influenzae tehB mutant was sensitive to hydrogen peroxide, tert-butyl peroxide, and cumene hydroperoxide [24] This tehB mutant was also impaired in its ability to cause infection in rats [24]. TehB may play a role in resistance to oxidative damage as a response regulator via its methyltransferase activity. H. influenzae has a TehA homolog that, while not included in this study, could be one target of TehB [24].
Multi-drug efflux pumps, including TehA homologs, are also important for the virulence of pathogens [36]. In Vibrio cholerae C6706, a TehA homolog plays a role in chloramphenicol resistance and intestinal colonization [44]. Relative to the parental strain, ∆tehA mutants are impaired in the colonization of mice. However, unlike the E. coli K-12 TehA [38], the V. cholerae TehA had no impact on nitrosative stress resistance [44]. Instead, the V. cholerae ∆tehA mutants are significantly impaired in adherence to mouse intestinal tissue [44]. Multi-drug efflux pumps have been linked to pathogen virulence through multiple mechanisms independent of the transport of antimicrobial compounds out of the cell. For example, autoinducers (AIs) are substrates for both the Pseudomonas aeruginosa MexAB-OprM efflux system [45] and the Burkholdaria pseudomallei BpeAB-OprB efflux system [46], with mutants lacking these systems deficient in quorum sensing. In the plant pathogens Burkholdaria gluma [47] and Pseudomonas syringae [48], multidrug efflux pumps are linked to the export of virulence-related toxins.
Reconciling the tellurite resistance phenotype of cells overexpressing tehA with the other functions described above for TehA may, at first, pose a challenge, considering that the E. coli TehA does not have tellurite efflux activity when overexpressed [23] despite transporting diverse xenobiotic substrates [21]. We can speculate on two possible explanations. Our first hypothesis is that TehA transports cytoplasmic biomolecules oxidized during tellurite exposure to prevent their accumulation inside the cell. Intracellular accumulation of lipid peroxidation-derived aldehydes such as butanaldehyde, propanaldehyde, acrolein, and malondialdehyde occurs following tellurite exposure [49,50]. These lipid peroxidation products can cause damaging protein modifications such as carbonylation [51] (Figure 1). However, we also considered that the attribution of tellurite resistance to tehA may be an instance of “guilt-by-association” with its neighboring tehB, which has a clearer role in tellurite resistance when overexpressed. Substitution of key cysteine residues in TehB (expressed from a high-copy number plasmid in E. coli K-12) with alanine significantly decreases tellurite resistance compared to the overexpressed unaltered protein. These cysteine residues are responsible for the binding of TehB to tellurite [29]. TehB has SAM-dependent methyltransferase activity and—when either overexpressed or in its purified form in vivo—will methylate non-physiological targets such as tellurite [22]. This methylation activity appears to be promiscuous as TehB also methylates selenite, selenium dioxide, and selenate in vivo [22]. We suggest that TehB has a primary function in the regulation—either directly or indirectly—of the activity of TehA as a multi-drug efflux pump under stress conditons [21]. In contrast, there is no strong evidence that tehA confers increased tellurite resistance. In the initial report of the chromosomal E. coli tehAB operon, tellurite resistance was determined by expressing the complete operon on a high-copy number plasmid [20]. Follow-up studies continued to characterize tellurite resistance associated with the complete tehAB operon cloned into high-copy number plasmids [22,29,52]. Deletion of tehA in V. cholerae C6706 had no impact on tellurite sensitivity [44]. Thus, a role for tehA in tellurite resistance, apart from its genetic linkage to tehB, waits to be unambiguously demonstrated, making its annotation as a “tellurium resistance gene” (Table 1) concerning.
TelA confers resistance to cell envelope-targeting antimicrobial compounds
The telA gene was first identified on IncPα plasmids from Gram-negative bacteria [8,53]. The IncPα plasmid RK2 carries a cryptic TeR determinant associated with the three gene kilA operon: kilA, telA, and telB (also referred to as klaA, klaB, and klaC) [54]. This operon is implicated in fertility inhibition of IncW plasmids [55], conditional host lethality [55,56], and TeR [53,57]. However, this plasmid-borne TeR operon only confers increased tellurite resistance after a point mutation occurs in the telB (klaC) [54]. While TelB/KlaC is poorly characterized, the protein (UniProt: Q52329) does contain a TrbC/VirB2 domain. These domains have functions in conjugal transfer systems [58,59], suggestive of a direct role for KlaC/TelB in plasmid maintenance, replication, and transfer rather than TeR. In R. sphaeroides, a chromosomal TeR locus encodes TelA, which shares 65% similarity with the IncPα-encoded TelA but lacks the other two genes found in IncPα plasmid TeR locus. Inactivation of R. sphaeroides telA leads to a significant decrease in tellurite resistance [16]. In the Gram-positive Bacillus subtilis and Bacillus anthracis, the TelA homolog YceH is encoded by a gene that is part of the chromosomal yceCDEFGH operon encoding the TeR protein homologs YceC (a TerE/D/Z homolog), YceD (TerD/E homolog), YceE (TerD/E homolog), and TceF (TerC homolog) [60,61]. These Ter homologs are discussed in later sections of this review. A B. anthracis yceH deletion strain had increased susceptibility to tellurite.
telA encodes a hydrophilic protein 42 kDa in size and 396 amino acids in length [54,62]. TelA is a cytoplasmic protein, predicted to form an α-helical structure with at least 10 conserved helices. telA is genetically linked with terC and terD as well as two genes encoding YceG and XpaC family proteins [63]. These two proteins are discussed in greater detail in the section on TerD. Briefly, YceG is linked to stress responses and membrane integrity, while XpaC is a 5-bromo-4-chloroindolyl phosphate hydrolysis protein [64]. A bioinformatic analysis performed by Anantharaman et al., (2012) predicted that, based on genomic proximity, TelA is a part of a membrane-associated stress response complex with other TeR proteins (TerC, TerD, and TerB) controlling diverse cellular processes such as phosphorylation-dependent signal transduction, RNA-mediated regulation, the biosynthesis of nucleoside-like metabolites, and DNA processing [63]. However, a later proteomic study of the uropathogenic E. coli KL53 TerC interactome found no evidence for interaction between TelA and TerC [65]. Thus, the interaction of TelA with other TeR proteins requires further investigation.
Multiple studies have identified a role for TelA in resistance to antimicrobial compounds targeting the cell envelope. A TelA homolog is required for nisin resistance in Listeria monocytogenes [66]. The telA locus was identified by screening a mariner transposon mutant library of L. monocytogenes for mutants susceptible to nisin. The role of telA in nisin resistance was further validated in a targeted deletion mutant (ΔtelA). ΔtelA had greater susceptibility to nisin as well as gallidermin, cefuroxime, cefotaxime, and bacitracin—all of which target the cell envelope [67]. Similarly, the TelA homolog in B. subtilis, YceH, was required for nisin resistance [61]. In Enterococcus faecalis, both telA and the genetically linked xpaC were connected to daptomycin resistance [64]. However, the mechanism through which TelA confers resistance to these cell envelope-targeting antimicrobials remains an open area for investigation. In B. subtilis, the TelA homolog YceH is expressed as part of the stress-responsive alternative sigma factor SigB regulon, which is responsive to diverse environmental stressors [68]. Thus, TelA homologs may be part of the general stress response systems of bacteria [69]. Given the phenotypes described here, these proteins may mediate the cellular response to membrane stressors, including certain antibiotics and tellurite (Figure 1).
TerD domain-containing proteins have undergone significant functional diversification
The proteins TerD, TerA, TerZ, TerE, and TerF share a TerD domain [63]. terD, terA, terZ, terE, and terF were common among the bacterial genome annotations that we analyzed (Table 1), with prevalences ranging from 1 to 15%. This was consistent with a prior study using the NCBI non-redundant (nr) database, which found that TerD homologs occur across most bacterial lineages and infrequently in some families of archaea and eukaryotes [63]. The terABCDE operon was first observed by Jobling and Ritchie on the plasmid pMER610 isolated from an Alcaligenes strain, which conferred resistance to Te and mercury [10,11]. When transferred to E. coli K-12, pMER610 increased tellurite resistance by 100-fold [11]. A similar operon, terZABCDEF, was observed in a region conferring TeR, phage resistance, and colicin resistance within the IncHI2 plasmid R478 from a clinical strain of Serratia marcescens [12,70]. terB and terC will be discussed in detail in the later sections of this review as they are structurally distinct from the other members of this operon. Cloning of this multi-resistance region into E. coli K-12 followed by transposon mutagenesis confirmed that terZ and terD confer TeR. Disruption of terA, terE, and terF had no impact on TeR [12]. An earlier study of the pMER610 ter operon expressed in E. coli K-12 reported that an insertion disrupting a region including terA and terB (discussed below) resulted in a tellurite hyposensitive phenotype, suggestive of a possible regulatory role for one or both of these two proteins [10,11]. When the pMER610 terD C-terminus region and the entirety of terE were deleted, cells became extremely tellurite sensitive [10,11].
TerD, TerA, TerZ, TerE, and TerF share a TerD domain [63], which consists of a β-sandwich fold formed by two β-sheets of five β-strands each and six short helices [71]. An NMR study of the Klebsiella pneumoniae TerD revealed two calcium ion (Ca2+) binding sites within the TerD domain [71]. An analysis of the polypeptide sequences retrieved from NCBI revealed that the S. marcescens R478-encoded TerD (NP_941153), TerZ (NP_941149.1), and TerE (NP_941154.1) are all 191-192 amino acid residues in length. A BLASTp [72] alignment of these three sequences revealed that TerE and TerZ have 65% and 40% identity to TerD and 37% identity to each other across >90% of the protein length. Using InterProScan [73], all three are predicted to contain a single TerD domain. These findings are consistent with the early report that the pMER610-encoded terE and terD are functionally redundant with respect to tellurite resistance [10]. TerA and TerF both contain a TerD domain fused to additional domains. TerA (NP_941150.1) is 386 amino acid residues in length and includes two fused TerD domains while TerF (NP_941155.1) is 413 amino acid residues in length and includes an N-terminus TerD domain fused to a C-terminus von Willebrand factor type A (vWA) domain. vWA domains are Rossmann folds consisting of a β-sheet sandwiched by α helices [74]. Bacterial vWA domains are poorly characterized; however, the majority are predicted to bind to a divalent cation [74]. Fusions of TerD with other domains have been reported including those involved in DNA processing, RNA binding, and peptide cleavage [63].
The roles of most of the TerD domain-containing Ter proteins in both tellurium resistance and other cellular functions remain poorly described. As detailed above, Anantharaman and colleagues (2012) performed a bioinformatic analysis of the genomic contexts and domain architecture of TeR genes including the TerD domain-containing Ter proteins [63] . Several major patterns of terD genomic neighborhoods were observed. Across most major bacterial clades, genetic linkage is observed between tandem arrays of terD paralogs, terC (described below), telA (described above), terF (encoding the TerD-vWR fusion), and genes encoding the poorly characterized proteins YceG, XpaC, and Aim24 [63]. In eukaryotes, Aim24 family proteins are involved in mitochondrial biogenesis [76,77,78]. XpaC is annotated as a 5-bromo-4-chloroindolyl phosphate hydrolysis protein; however, in Enterococcus faecium, xpaC was implicated in daptomycin resistance [64,79]. yceG was previously connected to multiple stress-response phenotypes in Bacillus anthracis including tellurite resistance; hydrogen peroxide and hypochlorite resistance; chloramphenicol, tetracycline, and penicillin resistance; and survival in a mammalian model of infection [60]. In Actinobacteria, Firmicutes, and Gammaproteobacteria, the paralogous terD genes commonly co-occur with terC, terB, and a biosynthetic module containing genes encoding an ATP-grasp amide bond synthetase, a TIM barrel enzyme, phosphoribosyltransferases, and a HAD superfamily phosphatase [63]. In these genomes, the ter operon and the biosynthetic module rarely occur independently of each other, suggestive of linked functions. This biosynthetic module may be involved in the production of a pyrimidine-derived ribonucleoside [63]. In some bacteria, terD and terB are combined in a conserved operon with genes involved in DNA-processing previously found in a phage restriction operon [63,80].
These genomic data, when combined with the observation that TerD domains are commonly fused to other domains of diverse functions, paint a picture of TerD domain-containing proteins as flexible modules that have been “mixed-and-matched” with a variety of cellular processes. From this bioinformatic analysis, it was proposed that TerD interacts at the membrane with a complex of other TeR proteins, namely TerF, TelA, TerB, and the membrane-bound TerC [63]. Two protein interaction studies have confirmed interactions between TerB and TerC [65,81] and TerB and TerD [81]. Additionally, TerE interacts with TerD and TerB and homoprotein complexes of both TerE and TerD were observed [81]. However, both studies failed to show TerD (or the homologous TerE) interaction with TerC. An analysis of the localization of heterologously expressed TerD and TerE found both proteins exclusively in the cytoplasm of E. coli K-12 while recombinant TerB and TerC were associated with the cytoplasmic and membrane fractions and the membrane fraction, respectively [81].
There is experimental evidence to suggest a function for some TerD domain proteins in the cellular response to membrane stress (Figure 1). Transposon disruption of the R478 terZ or terD resulted in significantly decreased resistance to colicins B, A, and K and somewhat decreased resistance to T5 phage infection [12]. T5 is a lytic phage [82], and colicins B, A, and K are toxins produced by certain strains of E. coli that form pores in bacterial cell membranes [83,84,85]. Disruption of terA somewhat decreased resistance to T5 phage infection while having no impact on colicin resistance. Disruption of terE or terF had no impact on either resistance to T5 phage infection or colicin resistance [12]. It is possible that the homologous TerE and TerD may be functionally redundant while TerF—with its vWA domain fusion to the TerD domain [63]—may have functionally diverged.
The TerD domain-containing proteins of the soil bacterium Streptomyces coelicolor have received considerable attention due to their role in cellular development and environmental stress responses [86,87]. S. coelicolor are multicellular mycelial bacteria with a complex morphological development process. The S. coelicolor genome contains 17 TerD domain-encoding genes. These proteins range in length from 190 amino acid residues (the length of the S. marcescens R478 TerD, TerZ, and TerE) to almost 700 residues [86]. The proteins longer than 200 amino acid residues likely represent fusions between a TerD domain and another domain [63]. Tdd8 (191 amino acid residues in length) is one of the most abundant proteins of the S. coelicolor proteome [88] and secretome [89] and has 58-69% identity with the S. marcescens TerE and TerD. However, neither deletion nor over-expression of tdd8 changes tellurite resistance in S. coelicolor. Instead, this protein has been linked to diverse cellular processes [86] including the ethanol stress response [90], the nitrogen limitation response [91,92], and plant-microbe interactions [89]. Tdd8 was also enriched in the cytoplasm of S. coelicolor during programmed cell death [93], a key step in the morphological development of S. coelicolor [94]. Subsequent work found that a tdd8 deletion strain grows more rapidly in liquid medium, has altered colony morphology on agar plates, and increased production of spores relative to the wild-type strain [86]. Increased expression of several genes involved in morphological differentiation and sporulation were also observed in this Dtdd8 strain [87]. In S. coelicolor, cellular differentiation and stress responses are interrelated [95,96], consistent with these observation that Tdd8 plays a role in both stress response and morphological development in the bacterium.
We speculate that a unifying feature of the functionally diverse TerD domain-containing proteins is their Ca2+-binding activity [63,71] (Figure 1). The K. pneumoniae TerD NMR structure revealed two Ca2+ binding sites that are highly conserved among TerD domain-containing proteins [71]. It is important to note that some TerD domain-containing proteins have poor conservation at one or both Ca2+ binding sites, suggestive of further functional divergence [63]. However, our discussion here will focus on those that retained both or one of these Ca2+ binding sites. A role for TerD domain-containing proteins in calcium homeostasis has been described in S. coelicolor [87]. Like the K. pneumoniae TerD, the S. coelicolor Tdd8 binds two Ca2+ ions. Transcriptome analysis of the tdd8 deletion strain revealed increased expression of genes involved in calcium-dependent antibiotic production as well as the calcium-binding protein CabC, which is implicated in the regulation spore germination and morphological development [87,97]. Intracellular calcium concentrations are significantly lower in the tdd8 deletion strain, leading to downstream impacts on redox stress response regulons responsive to cytoplasmic Ca2+ concentrations [87]. Calcium signaling is a form of intracellular communication widely used by eukaryotic organisms, regulating a range of cellular processes [98]. Calcium homeostasis systems are needed to maintain low intracellular Ca2+ concentrations relative to the external environment so that high transient intracellular Ca2+ concentrations in response to a stimulus can be exploited to transmit information [99]. Calcium is also believed to act as a signaling molecule in bacteria, controlling processes such as cell cycle and differentiation, motility, virulence, and stress resistance [100]. Like in eukaryotes, free intracellular concentrations of Ca2+ are low but transiently increase in response to external stimuli [101,102,103,104,105,106]. For example, in Bacillus subtilis, free intracellular [Ca2+] increased with hydrogen peroxide stress [107].
Calcium homeostasis in bacteria depends on the interplay between Ca2+ transporters/channels and intracellular Ca2+ binding proteins that are responsive to free Ca2+ [100]. We speculate that TerD domains are mediators of calcium signaling in bacteria through their Ca2+-binding activity. This likely occurs through one of two non-mutually exclusive mechanisms: (1) TerD domain-containing proteins may regulate free intracellular Ca2+ concentrations [87] or (2) TerD domains act as sensors that detect and initiate the cellular response to an influx cytoplasmic Ca2+ triggered by an external stimulus [108]. Calcium may be central in mediating the cellular response to stressors like tellurite, phage infection, and antibiotics as well as regulating stress-linked differentiation processes in certain bacteria.
TerB is a poorly characterized stress response protein genetically linked to TerD and TerC
TerB is present in the core ter operon originally described in the plasmids pMER610 and R478 10– [12,109]. A transposon insertion just upstream of the initiation methionine codon of the R478 terB led to decreased tellurite resistance in E. coli K-12 (the host strain of the cloned R478 ter operon region), similar to terZ, terD, terE, and terC [12]. Like the other R478 ter genes, this disruption of the terB upstream region also led to decreased resistance to T5 phage infection and pore-forming colicins B, A, and K [12]. As described above, an earlier study of the pMER610 ter operon expressed in E. coli K-12 reported that an insertion disrupting a region including terA and terB resulted in a tellurite hyposensitive phenotype [10,11]. However, it is unclear whether this phenotype is due to disruption of terA alone, terB alone, or both in combination. Additionally, Peng et al demonstrated that the terB gene of the highly tellurite-resistant Pseudomonas citronellolis SJTE-3 had no impact on tellurite resistance when expressed in E. coli K-12 [110].
An NMR structure for the K. pneumoniae TerB (KP-TerB) showed that the protein consists of 151 amino acids, forming seven α-helices and a 310 helix [111]. A crystal structure of the Nostoc punctiforme PCC73102 TerB (NP-TerB) suggested that this protein forms a homodimeric complex [112]. This finding is further supported by an in vitro pull-down assay demonstrating homotypic interactions for recombinant TerB from the uropathogenic E. coli KL53 [81]. A computational analysis of the KP-TerB and NP-TerB [111,112] uncovered a common internal repeat of two tetra-helical units that are closely stacked together to form an eight-helical structure. These tetra-helical units contain a DsxhxxxE motif (s = small amino acid; h = hydrophobic amino acid) in the hairpin loop linking the two central helices. These conserved acidic residues create a metal-binding site occupied by Zn²⁺ in the NP-TerB structure [63]. Moreover, the crystal structure shows that NP-TerB binds the soluble molecule indole-3-carbaldehyde in a pocket on the opposite the metal-binding site [63,112], suggesting that this pocket could be the binding pocket for TerB’s physiological substrate [63].
In the bioinformatic analysis of the TeR genes by Anantharaman et al., several genomic contexts for terB were described. terB is frequently genomically linked to terC and terD in the context of the biosynthetic module described in the previous section on terD [63]. Protein-protein interaction studies confirmed physical interaction of TerB with both TerC and TerD [65,81] (Figure 1). Based on the observation that NP-TerB [112] has a binding pocket for soluble metabolites, it was speculated that TerB may help link TerD and TerC to the enzymes encoded by the biosynthetic module [63]. As described in the previous section, terD and terB are also linked in a conserved operons containing genes for various DNA-processing enzymes, some of which are also present in a phage restriction operon [63,76]. Anantharaman et al. also discovered several new combinations of TerB domain fusions. Most of these fusions appear to involve domains with lipid-binding activity or that are membrane-associated [63]. These proteins fall into one of five major groupings of domain architecture: (1) TerB combined with transmembrane helices and a C-terminal DnaJ domain, which may help recruit Hsp70 chaperones in response to stress conditions [113]. (2) TerB combined with either a dynamin-like GTPase or a TRAFAC clade GTPase, both of which also have TM regions [63]. (3) TerB fused with globular domains, including novel domains possibly or one or more N-terminal Tim44 domains, which have lipid-binding functions are often associated with the inner leaflet of membrane bilayers [114]. (4) A TerB fusion with a fatty acyl-CoA oxidase module [115], which plays a role in lipid oxidation. (5) TerB domains fused to a Coq4 domain, which is localized to the inner leaflet of membranes and involved in membrane protein organization [116]. Overall, these findings suggest that to the TerB domains may have diverse membrane-related roles across a range of bacterial species. Experimental work is needed to confirm these putative functions.
Experimental evidence for a TerB function is sparse. While TerB has been linked to phage infection and colicins resistance, the biochemical mechanism underlying this phenotype has not been described [12]. TerB has been experimentally linked to a few additional bacterial phenotypes; however, the mechanistic details behind these observations are also lacking. In the enteropathogenic Yersinia pseudotuberculosis, terB expression is down-regulated during growth at body-temperature [117]. While in the multi-stressor resistant bacterium Deinococcus radiodurans, the terB gene is linked to UVC irradiation resistance [118], vacuum stress resistance [118], hydrogen peroxide resistance [119] , and gamma irradiation resistance [120,121,122]. Due to the limited experimental studies on this protein, it is challenging to speculate on a unifying mechanism that ties together these distinct phenotypes. Given the association (both genomically [63] and through protein-protein interaction studies [65,81]) between TerB and both TerD and TerC, it is apparent that further work is called for on this ubiquitous protein that likely plays a central role in linking disparate environmental stress response systems.
TerC is a manganese efflux pump with a role in metalation of secreted proteins.
In our earlier analysis, we found that terC is common in bacterial genome annotations (Table 1). Like the other ter genes, terC was first reported on plasmids conferring the TeR phenotype isolated from members of the Enterobacteriaceae [11,12]. Transposon-mediated disruption of terC on the S. marcescens R478 plasmid expressed in E. coli K-12 resulted in a loss of the TeR in the host strain [12]. Similar results were reported by two separate studies using transposon mutagenesis to characterize terC cloned from the clinical KL53 strain of E. coli and expressed in E. coli K-12 [123] as well as a chromosomally encoded terC in Proteus mirabilis [124]. However, the mechanism by which terC confers TeR was never established.
TerC belongs to the lysine exporter (LysE) superfamily of transporters [125,126]. LysE superfamily members are transmembrane transporters with characterized functions in the export of amino acids, lipids, and metal ions [126]. Most members within this superfamily are similarly sized (~200 amino acid residues in length) and include multiple transmembrane a-helical segments [126]. In TerC homologs, there are multiple intramembrane metal-binding sites [63]. However, despite being an integral membrane protein with homology to metal transporters, there has never been any evidence to suggest that TerC is a tellurite transporter.
The physiological functions of TerC homologs are well-described thanks to series of recent mechanistic studies performed in several different bacteria. Three recent studies have linked TerC to manganese efflux. In E. coli K-12, a gene encoding a TerC homolog, called Alx, is under the control of the manganese ion (Mn2+) and alkaline pH-responsive yybP-ykoY riboswitch [127]. alx is both Mn2+-responsive—with increasing expression at increasingly higher Mn2+ concentrations [127]—and highly induced under alkaline conditions [128]. While Sharma and Mishanina [129] confirmed these earlier findings, deletion of alx had no impact on the growth rate of E. coli K-12 at alkaline pH. Additionally, there was no difference in the cytoplasmic pH between the parental and Dalx strains over a range of external pH conditions. Thus, it is unlikely that Alx has a direct function in maintaining pH homeostasis [129]. Rather, alkalization of the cytoplasm causes increased intracellular [Mn2+], and Alx exports this excess Mn2+. Increased expression of alx resulted in decreased intracellular [Mn] only under alkaline (pH 8.4) conditions. The mechanism underlying the increased [Mn2+] under alkaline conditions is unclear; however, some induction of a katG (a bifunctional catalase-peroxidase) transcriptional reporter was observed at pH 8.4 [129]. In E. coli K-12, Mn2+ import is a critical part of the oxidative stress response [130]. Mn2+ can replace Fe2+ as a cofactor for enzymes like superoxide dismutase (SOD) during periods of oxidative stress [131,132] and, under these conditions, cells also express Mn-dependent isoforms of enzymes like NrdEF ribonucleotide reductase [133] and the heme biosynthetic enzyme HemF (coproporphyrinogen III) [134] During periods of acute oxidative stress where cytoplasmic [Mn2+] increases, Alx may fine-tune the intracellular Mn2+ concentrations [129] to prevent over-import and cellular toxicity [135].
Similar to E. coli, in the avian pathogen Riemerella anatipestifer, a DterC mutant strain had increased sensitivity to Mn2+ alongside increased intracellular accumulation of Mn2+, supporting its function in Mn2+ efflux [136]. The expression of this TerC was induced by high concentrations of Mn2+ and repressed under iron-limited conditions, with Mn2+ efflux previously linked to iron homeostasis in R. anatipestifer [137]. In R. anatipestifer, a functional TerC was essential for sodium hypochlorite-induced oxidative stress resistance [136] as Mn2+ accumulation sensitizes cells to sodium hypochlorite-induced oxidative stress [138]. The R. anatipestifer ∆terC strain also had reduced virulence and colonization in a duckling model of infection; however, this phenotype was decoupled from the intracellular accumulation of Mn2+ [136], suggestive of additional functions of TerC in cellular physiology, possibly similar to what is described below for B. subtilis.
In B. subtilis, TerC has been linked to both manganese efflux and metalation of secreted proteins [139] (Figure 1). Similar functions were confirmed for homologs in L. monocytogenes, and B. anthracis [139]. MneP and MneS are cation diffusion facilitator proteins that are the primary Mn2+ efflux pumps in B. subtilis [140]. mnePS double mutants are extremely sensitive to Mn2+ [140]. However, this growth defect can be partially rescued by overexpression of the TerC homolog YceF. A paralogous protein, YkoY, is partially redundant in function with YceF. However, the contributions of YceG and YkoY to Mn2+ efflux in B. subtilis are minor when compared to MneP and MneS. In a subsequent study, it was found that both YceF (renamed to MeeF for metalation of exoenzymes) and YkoY (renamed to MeeY) are involved in the metalation of exoenzymes with Mn either during or after translocation via the SecYEG translocon139,141. A meeFY double mutant has a generalized defect in protein secretion, including native enzymes like proteases and the heterologously expressed α-amylase from B. amyloliquefaciens (AmyQ). This generalized impairment of protein secretion is due to nascent un-metalated metalloproteins jamming the SecYEG translocon. FtsH protease, which selectively degrades jammed partially translocated proteins in the SecYEG translocon, is required for the viability of the meeFY double mutant [139]. MeeF and MeeY co-immunoprecipitate with proteins of the Sec translocon (SecDF, SecY, YrbF) and quality control proteases (FrsH, PsrA, HtpX) [139]. Similarly, the uropathogenic E. coli KL53 TerC was previously found to co-immunoprecipitate with proteins involved in protein synthesis, folding, export (including SecY, SecF, and SecD), and degradation (including FtsH) [65], suggesting a conserved function for TerC in protein secretion. In addition to metalating actively translocating proteins, the B. subtilis MeeF and MeeY also export Mn to generate high localized concentrations of Mn at the cell surface for metalation of metalloproteins post-translocation. For example, MeeF and MeeY contribute to the post-translocational metalation of LtaS, an integral membrane protein with lipoteichoic acid synthase activity [141]. Other metalloenzyme targets of MeeF and MeeY remain to be identified.
TerC has a primary function in Mn2+ export, possibly for both the metalation of secreted proteins and prevention of manganese intoxication. While these activities appear to be independent of TeR, nonetheless the disruption of terC in clinical strains of E. coli and close relatives leads to increased susceptibility to tellurite. How, then, do we reconcile these observations? Acidic residues in the transmembrane helices of both E. coli Alx and R. anatipestifer TerC are responsible for Mn2+ transport, making it unlikely that this transporter would also efflux the oxyanion tellurite129,136. TerC, like the E. coli homolog Alx, may fine-tune cytoplastic Mn2+ concentrations during tellurite stress [142]. Reduction of cytoplasmic tellurite by cellular enzymes and low molecular weight thiols generates superoxide [143,144], leading to increased expression of Mn-utilizing antioxidant enzymes [17,143,145]. This may result in transient increases of cytoplasmic Mn2+ [131] (Figure 1). Another possibility is that all TerC homologs have a primary function in the metalation of secreted proteins like the B. subtilis MeeF and MeeY. The lipoteichoic acid synthase LtaS is metalated by MeeY and MeeF-secreted Mn2+. In the Actinobacterium Micromonospora, tellurite exposure led to qualitative shifts in the chemical structure of the polysaccharide content of the cell wall, including lipoteichoic acid (LTA). LTA synthesis could be part of the tellurite stress response in Gram-positive organisms [146]. However, this would not explain the function of TerC in tellurite resistance in Gram-negative bacteria such as E. coli. In these organisms, TerC may metalate other secreted enzymes that mediate the cellular response to envelope stress (Figure 1). This model also explains the observation that TerC confers resistance to both phage infection and pore-forming colicins [12].
4. Future Directions
In the era of low-cost (meta)genome sequencing, we are increasingly reliant on database annotations to provide hints towards the functioning of diverse microorganisms in the world around us. While powerful resources, these annotations often miss important functional information available in the literature for genes and the proteins they encode. For example, the BacMet Antibacterial Biocide & Metal Resistance Genes Database [18] includes entries for, terABCDEZ as tellurium resistance genes while neglecting to mention that many of the ter genes have been linked to colicin resistance [12] and that terC has a function in manganese resistance [129,147]. We searched for papers citing the original BacMet publication that included the terms “tellurium” or “tellurite”, identifying 106 in total. These numbers have steadily increased over time (Figure 2). However, this problem is not limited to the BacMet database, the JGI [148] and NCBI [149] standard annotation pipelines still largely refer to these genes as tellurium resistance genes. The COG (Clusters of Orthologous Genes) database [19] contains entries for “tellurite resistance protein TehB” (COG3615), “tellurite resistance protein TehA” (COG1275), “tellurite resistance protein TerB” (COG3794, COG4103), “tellurite resistance membrane protein TerC” (COG0861), and “tellurium resistance protein TerA/TerD” (COG4110). We recommend that annotations in human-curated databases, where relevant, be updated to reflect these additional functions of the TeR genes.
Many questions remain with respect to the functions of these genes. Key areas of research outlined in this review include:
- (i)
- Confirming if TehA has a function in tellurium resistance.
- (ii)
- Determining if and how TehB directly or indirectly regulates the activity of TehA.
- (iii)
- Determining the physiological substrate of TehA. Is it antimicrobial compounds? Or does it export oxidatively damaged biomolecules?
- (iv)
- A mechanism by which TelA confers resistance to membrane-targeting antimicrobials.
- (v)
- Describing the mechanism(s) by which TerD domains participate in calcium signaling. How do cells use calcium as a signal of extracellular stress and how do TerD domains integrate these signals?
- (vi)
- Confirming if the function of TerC in exoenzyme metalation is universal. While this function is conserved between B. subtilis, L. monocytogenes, and B. anthracis, does this hold true beyond the Firmicutes phylum?
- (vii)
- Identifying additional metalloenzyme targets of TerC in B. subtilis and other bacteria.
We also see several new and exciting research directions for the TeR proteins with respect to their evolution. For example, ter [63] and telA [150] homologs are observed in eukaryotes and archaea. Are these the result of horizontal gene transfer from bacteria? Have the proteins they encode functionally diverged from their bacterial homologs? Another interesting question concerns the relationship between the plasmid-borne and chromosomally encoded versions of the TeR genes: are these proteins functionally identical, or have they diverged and specialized? Finally, we found many reports of bacteriophage genomes encoding homologs of TeR genes [151,152,153,154,155,156,157,158,159,160]. Given that terB, terC, and terD confer phage resistance in E. coli K-12 [12], it is tempting to speculate that their viral-encoded counterparts may be involved in phage superinfection exclusion in the host [161,162]. While considerable future work is needed to address these questions, we believe that doing so will reveal that the TeR genes play key roles in generalized stress responses, and possibly other fundamental cellular functions, across diverse organisms. Given the linkages of many of these genes to bacterial antibiotic resistance, phage resistance, and pathogen virulence, efforts to better characterize their functioning at a mechanistic level may reveal novel strategies for antimicrobial development.
Funding
Funding for this work was provided by start-up funds for JLG from the Department of Chemistry at SUNY College of Environmental Science and Forestry.
References
- Missen, O. P.; Ram, R.; Mills, S. J.; Etschmann, B.; Reith, F.; Shuster, J.; Smith, D. J.; Brugger, J. Love Is in the Earth: A Review of Tellurium (Bio)Geochemistry in Surface Environments. Earth-Sci. Rev. 2020, 204, 103150. [Google Scholar] [CrossRef]
- Conradi, H.; Troch, P. Ein Verfahren Zum Nachweis Der Diphtheriebazillen. Muench Med Wochenschr 1912, 59, 1652–1653. [Google Scholar]
- Glass, V. The Isolation and Typing of C. Diphtheriæ on Tellurite Blood Agar. J. Pathol. Bacteriol. 1937, 44, 235–246. [Google Scholar] [CrossRef]
- Zadik, P. M.; Chapman, P. A.; Siddons, C. A. Use of Tellurite for the Selection of Verocytotoxigenic Escherichia Coli O157. J. Med. Microbiol. 1993, 39, 155–158. [Google Scholar] [CrossRef]
- Presentato, A.; Turner, R. J.; Vasquez, C. C.; Yurkov, V.; Zannoni, D. Tellurite-Dependent Blackening of Bacteria Emerges from the Dark Ages. Environ. Chem. 2019, 16, 266–288. [Google Scholar] [CrossRef]
- Summers, A.; Jacoby, G. Plasmid-Determined Resistance to Tellurium Compounds. J. Bacteriol. 1977, 129, 276–281. [Google Scholar] [CrossRef]
- Hou, Y.; Taylor, D. E. Incidence of Tellurite Resistance Determinants among Plasmids of Different Incompatibility Groups. Plasmid 1994, 32, 306–311. [Google Scholar] [CrossRef]
- Bradley, D. E. Detection of Tellurite-Resistance Determinants in IncP Plasmids. Microbiology 1985, 131, 3135–3137. [Google Scholar] [CrossRef]
- Taylor, D. E.; Summers, A. O. Association of Tellurium Resistance and Bacteriophage Inhibition Conferred by R Plasmids. J. Bacteriol. 1979, 137, 1430–1433. [Google Scholar] [CrossRef]
- Jobling, M. G.; Ritchie, D. A. The Nucleotide Sequence of a Plasmid Determinant for Resistance to Tellurium Anions. Gene 1988, 66, 245–258. [Google Scholar] [CrossRef]
- Jobling, M. G.; Ritchie, D. A. Genetic and Physical Analysis of Plasmid Genes Expressing Inducible Resistance of Tellurite in Escherichia Coli. Mol. Gen. Genet. MGG 1987, 208, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K. F.; Colleran, E.; Taylor, D. E. Phage Inhibition, Colicin Resistance, and Tellurite Resistance Are Encoded by a Single Cluster of Genes on the IncHI2 Plasmid R478. J. Bacteriol. 1995, 177, 5016–5027. [Google Scholar] [CrossRef] [PubMed]
- Walter, E. G.; Taylor, D. E. Plasmid-Mediated Resistance to Tellurite: Expressed and Cryptic. Plasmid 1992, 27, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Walter, E. G.; Thomas, C. M.; Ibbotson, J. P.; Taylor, D. E. Transcriptional Analysis, Translational Analysis, and Sequence of the kilA-Tellurite Resistance Region of Plasmid RK2Ter. J. Bacteriol. 1991, 173, 1111–1119. [Google Scholar] [CrossRef]
- Taylor, D. E. Bacterial Tellurite Resistance. Trends Microbiol. 1999, 7, 111–115. [Google Scholar] [CrossRef]
- O’Gara, J. P.; Gomelsky, M.; Kaplan, S. Identification and Molecular Genetic Analysis of Multiple Loci Contributing to High-Level Tellurite Resistance in Rhodobacter Sphaeroides 2.4.1. Appl. Environ. Microbiol. 1997, 63, 4713–4720. [Google Scholar] [CrossRef]
- Chasteen, T. G.; Fuentes, D. E.; Tantaleán, J. C.; Vásquez, C. C. Tellurite: History, Oxidative Stress, and Molecular Mechanisms of Resistance. FEMS Microbiol. Rev. 2009, 33, 820–832. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D. G. BacMet: Antibacterial Biocide and Metal Resistance Genes Database. Nucleic Acids Res. 2014, 42, D737–43. [Google Scholar] [CrossRef]
- Galperin, M. Y.; Wolf, Y. I.; Makarova, K. S.; Vera Alvarez, R.; Landsman, D.; Koonin, E. V. COG Database Update: Focus on Microbial Diversity, Model Organisms, and Widespread Pathogens. Nucleic Acids Res. 2021, 49, D274–D281. [Google Scholar] [CrossRef]
- Taylor, D. E.; Hou, Y.; Turner, R. J.; Weiner, J. H. Location of a Potassium Tellurite Resistance Operon (tehA tehB) within the Terminus of Escherichia Coli K-12. J. Bacteriol. 1994, 176, 2740–2742. [Google Scholar] [CrossRef]
- Turner, R. J.; Taylor, D. E.; Weiner, J. H. Expression of Escherichia Coli TehA Gives Resistance to Antiseptics and Disinfectants Similar to That Conferred by Multidrug Resistance Efflux Pumps. Antimicrob. Agents Chemother. 1997, 41, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Liu Mingfu; Turner Raymond J. ; Winstone Tara L.; Saetre Andrea; Dyllick-Brenzinger Melanie; Jickling Glen; Tari Leslie W.; Weiner Joel H.; Taylor Diane E. Escherichia Coli TehB RequiresS-Adenosylmethionine as a Cofactor To Mediate Tellurite Resistance. J. Bacteriol. 2000, 182, 6509–6513. [Google Scholar] [CrossRef] [PubMed]
- Turner, R. J.; Weiner, J. H.; Taylor, D. E. Neither Reduced Uptake nor Increased Efflux Is Encoded by Tellurite Resistance Determinants Expressed in Escherichia Coli. Can. J. Microbiol. 1995, 41, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Whitby, P. W.; Seale, T. W.; Morton, D. J.; VanWagoner, T. M.; Stull, T. L. Characterization of the Haemophilus Influenzae tehB Gene and Its Role in Virulence. Microbiology 2010, 156, 1188–1200. [Google Scholar] [CrossRef]
- Choudhury, H. G.; Cameron, A. D.; Iwata, S.; Beis, K. Structure and Mechanism of the Chalcogen-Detoxifying Protein TehB from Escherichia Coli. Biochem. J. 2011, 435, 85–91. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Hu, L.; Punta, M.; Bruni, R.; Hillerich, B.; Kloss, B.; Rost, B.; Love, J.; Siegelbaum, S. A.; Hendrickson, W. A. Homologue Structure of the SLAC1 Anion Channel for Closing Stomata in Leaves. Nature 2010, 467, 1074–1080. [Google Scholar] [CrossRef]
- Taylor, D. E.; Walter, E. G.; Sherburne, R.; Bazett-Jones, D. P. Structure and Location of Tellurium Deposited in Escherichia Coli Cells Harbouring Tellurite Resistance Plasmids. J. Ultrastruct. Mol. Struct. Res. 1988, 99, 18–26. [Google Scholar] [CrossRef]
- Tomás, J. M.; Kay, W. W. Tellurite Susceptibility and Non-Plasmid-Mediated Resistance in Escherichia Coli. Antimicrob. Agents Chemother. 1986, 30, 127–131. [Google Scholar] [CrossRef]
- Dyllick-Brenzinger, M.; Liu, M.; Winstone, T. L.; Taylor, D. E.; Turner, R. J. The Role of Cysteine Residues in Tellurite Resistance Mediated by the TehAB Determinant. Biochem. Biophys. Res. Commun. 2000, 277, 394–400. [Google Scholar] [CrossRef]
- Gambino, L.; Gracheck, S. J.; Miller, P. F. Overexpression of the MarA Positive Regulator Is Sufficient to Confer Multiple Antibiotic Resistance in Escherichia Coli. J. Bacteriol. 1993, 175, 2888–2894. [Google Scholar] [CrossRef]
- Seoane, A. S.; Levy, S. B. Identification of New Genes Regulated by the marRAB Operon in Escherichia Coli. J. Bacteriol. 1995, 177, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I. T.; Skurray, R. A.; Tam, R.; Saier Jr, M. H.; Turner, R. J.; Weiner, J. H.; Goldberg, E. B.; Grinius, L. L. The SMR Family: A Novel Family of Multidrug Efflux Proteins Involved with the Efflux of Lipophilic Drugs. Mol. Microbiol. 1996, 19, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Huang, M.; Wei, Y. Diversity of the Reaction Mechanisms of SAM-Dependent Enzymes. Acta Pharm. Sin. B 2021, 11, 632–650. [Google Scholar] [CrossRef]
- Poole, K. Bacterial Multidrug Efflux Pumps Serve Other Functions. Microbe-Am. Soc. Microbiol. 2008, 3, 179. [Google Scholar] [CrossRef]
- Piddock, L. J. V. Multidrug-Resistance Efflux Pumps ? Not Just for Resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef]
- Fernando, D. M.; Kumar, A. Resistance-Nodulation-Division Multidrug Efflux Pumps in Gram-Negative Bacteria: Role in Virulence. Antibiotics 2013, 2, 163–181. [Google Scholar] [CrossRef]
- Bodenmiller, D. M.; Spiro, S. The yjeB (nsrR) Gene of Escherichia Coli Encodes a Nitric Oxide-Sensitive Transcriptional Regulator. J. Bacteriol. 2006, 188, 874–881. [Google Scholar] [CrossRef]
- Justino, M. C.; Vicente, J. B.; Teixeira, M.; Saraiva, L. M. New Genes Implicated in the Protection of Anaerobically Grown Escherichia Coli against Nitric Oxide*. J. Biol. Chem. 2005, 280, 2636–2643. [Google Scholar] [CrossRef]
- Fetar, H.; Gilmour, C.; Klinoski, R.; Daigle, D. M.; Dean, C. R.; Poole, K. mexEF-oprN Multidrug Efflux Operon of Pseudomonas Aeruginosa: Regulation by the MexT Activator in Response to Nitrosative Stress and Chloramphenicol. Antimicrob. Agents Chemother. 2011, 55, 508–514. [Google Scholar] [CrossRef]
- Ma, D.; Cook, D. N.; Alberti, M.; Pon, N. G.; Nikaido, H.; Hearst, J. E. Genes acrA and acrB Encode a Stress-induced Efflux System of Escherichia Coli. Mol. Microbiol. 1995, 16, 45–55. [Google Scholar] [CrossRef]
- Fraud, S.; Poole, K. Oxidative Stress Induction of the MexXY Multidrug Efflux Genes and Promotion of Aminoglycoside Resistance Development in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Zhang, Q.; Ryu, S.; Jeon, B. Transcriptional Regulation of the CmeABC Multidrug Efflux Pump and the KatA Catalase by CosR in Campylobacter Jejuni. J. Bacteriol. 2012, 194, 6883–6891. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Sobel, M. L.; Poole, K. Antibiotic Inducibility of the MexXY Multidrug Efflux System of Pseudomonas Aeruginosa: Involvement of the Antibiotic-Inducible PA5471 Gene Product. J. Bacteriol. 2006, 188, 1847–1855. [Google Scholar] [CrossRef]
- Pei, B.; Wang, Y.; Katzianer, D.; Wang, H.; Wu, H.; Zhong, Z.; Zhu, J. Role of a TehA Homolog in Vibrio Cholerae C6706 Antibiotic Resistance and Intestinal Colonization. Can. J. Microbiol. 2013, 59, 136–139. [Google Scholar] [CrossRef]
- Evans, K.; Passador, L.; Srikumar, R.; Tsang, E.; Nezezon, J.; Poole, K. Influence of the MexAB-OprM Multidrug Efflux System on Quorum Sensing in Pseudomonas Aeruginosa. J. Bacteriol. 1998, 180, 5443–5447. [Google Scholar] [CrossRef]
- Chan, Y. Y.; Chua, K. L. The Burkholderia Pseudomallei BpeAB-OprB Efflux Pump: Expression and Impact on Quorum Sensing and Virulence. J. Bacteriol. 2005, 187, 4707–4719. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.-G.; Kang, Y.; Jang, J. Y.; Jog, G. J.; Lim, J. Y.; Kim, S.; Suga, H.; Nagamatsu, T.; Hwang, I. Quorum Sensing and the LysR-Type Transcriptional Activator ToxR Regulate Toxoflavin Biosynthesis and Transport in Burkholderia Glumae. Mol. Microbiol. 2004, 54, 921–934. [Google Scholar] [CrossRef]
- Kang, H.; Gross, D. C. Characterization of a Resistance-Nodulation-Cell Division Transporter System Associated with the Syr-Syp Genomic Island of Pseudomonas Syringae Pv. Syringae. Appl. Environ. Microbiol. 2005, 71, 5056–5065. [Google Scholar] [CrossRef]
- Pérez, J. M.; Arenas, F. A.; Pradenas, G. A.; Sandoval, J. M.; Vásquez, C. C. Escherichia Coli YqhD Exhibits Aldehyde Reductase Activity and Protects from the Harmful Effect of Lipid Peroxidation-Derived Aldehydes. J. Biol. Chem. 2008, 283, 7346–7353. [Google Scholar] [CrossRef]
- Pradenas, G. A.; Díaz-Vásquez, W. A.; Pérez-Donoso, J. M.; Vásquez, C. C. Monounsaturated Fatty Acids Are Substrates for Aldehyde Generation in Tellurite-Exposed Escherichia Coli. BioMed Res. Int. 2013, 2013, 563756. [Google Scholar] [CrossRef]
- Refsgaard, H. H. F.; Tsai, L.; Stadtman, E. R. Modifications of Proteins by Polyunsaturated Fatty Acid Peroxidation Products. Proc. Natl. Acad. Sci. 2000, 97, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Turner, R. J.; Weiner, J. H.; Taylor, D. E. The Tellurite-Resistance Determinants tehAtehB and klaAklaBtelB Have Different Biochemical Requirements. Microbiology 1995, 141, 3133–3140. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D. E. Bacterial Tellurite Resistance. Trends Microbiol. 1999, 7, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Turner, R. J.; Weiner, J. H.; Taylor, D. E. In Vivo Complementation and Site-Specific Mutagenesis of the Tellurite Resistance Determinant kilAtelAB from IncP Alpha Plasmid RK2Ter. Microbiol. Read. Engl. 1994, 140 ( Pt 6) Pt 6, 1319–1326. [Google Scholar] [CrossRef]
- Goncharoff, P.; Saadi, S.; Chang, C. H.; Saltman, L. H.; Figurski, D. H. Structural, Molecular, and Genetic Analysis of the kilA Operon of Broad-Host-Range Plasmid RK2. J. Bacteriol. 1991, 173, 3463–3477. [Google Scholar] [CrossRef]
- Holčík, M.; Iyer, V. M. Conditionally Lethal Genes Associated with Bacterial Plasmids. Microbiology 1997, 143, 3403–3416. [Google Scholar] [CrossRef]
- Pansegrau, W.; Lanka, E.; Barth, P. T.; Figurski, D. H.; Guiney, D. G.; Haas, D.; Helinski, D. R.; Schwab, H.; Stanisich, V. A.; Thomas, C. M. Complete Nucleotide Sequence of Birmingham IncPα Plasmids: Compilation and Comparative Analysis. J. Mol. Biol. 1994, 239, 623–663. [Google Scholar] [CrossRef]
- Schulein, R.; Dehio, C. The VirB/VirD4 Type IV Secretion System of Bartonella Is Essential for Establishing Intraerythrocytic Infection. Mol. Microbiol. 2002, 46, 1053–1067. [Google Scholar] [CrossRef]
- Kishida, K.; Li, Y. G.; Ogawa-Kishida, N.; Khara, P.; Mamun, A. A. M. A.; Bosserman, R. E.; Christie, P. J. Chimeric Systems Composed of Swapped Tra Subunits between Distantly-Related F Plasmids Reveal Striking Plasticity among Type IV Secretion Machines. PLOS Genet. 2024, 20, e1011088. [Google Scholar] [CrossRef]
- Franks, S. E.; Ebrahimi, C.; Hollands, A.; Okumura, C. Y.; Aroian, R. V.; Nizet, V.; McGillivray, S. M. Novel Role for the yceGH Tellurite Resistance Genes in the Pathogenesis of Bacillus Anthracis. Infect. Immun. 2014, 82, 1132–1140. [Google Scholar] [CrossRef]
- Kingston, A. W.; Liao, X.; Helmann, J. D. Contributions of the σ, σ and σ Regulons to the Lantibiotic Resistome of Acillus Subtilis. Mol. Microbiol. 2013, 90, 502–518. [Google Scholar] [CrossRef]
- Walter, E. G.; Thomas, C. M.; Ibbotson, J. P.; Taylor, D. E. Transcriptional Analysis, Translational Analysis, and Sequence of the kilA-Tellurite Resistance Region of Plasmid RK2Ter. J. Bacteriol. 1991, 173, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Anantharaman, V.; M. Iyer, L.; Aravind, L. Ter-Dependent Stress Response Systems: Novel Pathways Related to Metal Sensing, Production of a Nucleoside-like Metabolite, and DNA-Processing. Mol. Biosyst. 2012, 8, 3142–3165. [Google Scholar] [CrossRef] [PubMed]
- Tran, T. T.; Panesso, D.; Diaz, L.; Rios, R.; Arias, C. A. 601. TelA and XpaC Are Novel Mediators of Daptomycin Resistance in Enterococcus Faecium. Open Forum Infect. Dis. 2019, 6 (Supplement_2). [Google Scholar] [CrossRef]
- Turkovicova, L.; Smidak, R.; Jung, G.; Turna, J.; Lubec, G.; Aradska, J. Proteomic Analysis of the TerC Interactome: Novel Links to Tellurite Resistance and Pathogenicity. J. Proteomics 2016, 136, 167–173. [Google Scholar] [CrossRef]
- Collins, B.; Joyce, S.; Hill, C.; Cotter, P. D.; Ross, R. P. TelA Contributes to the Innate Resistance of Listeria Monocytogenes to Nisin and Other Cell Wall-Acting Antibiotics. Antimicrob Agents Chemother 2010, 54, 4658–4663. [Google Scholar] [CrossRef]
- Collins, B.; Joyce, S.; Hill, C.; Cotter, P. D.; Ross, R. P. TelA Contributes to the Innate Resistance of Listeria Monocytogenes to Nisin and Other Cell Wall-Acting Antibiotics. Antimicrob. Agents Chemother. 2010, 54, 4658–4663. [Google Scholar] [CrossRef]
- Petersohn, A.; Brigulla, M.; Haas, S.; Hoheisel, J. D.; Völker, U.; Hecker, M. Global Analysis of the General Stress Response ofBacillus Subtilis. J. Bacteriol. 2001, 183, 5617–5631. [Google Scholar] [CrossRef]
- Rodriguez Ayala, F.; Bartolini, M.; Grau, R. The Stress-Responsive Alternative Sigma Factor SigB of Bacillus Subtilis and Its Relatives: An Old Friend With New Functions. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Hedges, R. W.; Rodriguez-Lemoine, V.; Datta, N. R Factors from Serratia Marcescens. Microbiology 1975, 86, 88–92. [Google Scholar] [CrossRef]
- Pan, Y.-R.; Lou, Y.-C.; Seven, A. B.; Rizo, J.; Chen, C. NMR Structure and Calcium-Binding Properties of the Tellurite Resistance Protein TerD from Klebsiella Pneumoniae. J. Mol. Biol. 2011, 405, 1188–1201. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T. L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36 (suppl_2), W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Zdobnov, E. M.; Apweiler, R. InterProScan – an Integration Platform for the Signature-Recognition Methods in InterPro. Bioinformatics 2001, 17, 847–848. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, C. A.; Hynes, R. O. Distribution and Evolution of von Willebrand/Integrin A Domains: Widely Dispersed Domains with Roles in Cell Adhesion and Elsewhere. Mol. Biol. Cell 2002, 13, 3369–3387. [Google Scholar] [CrossRef]
- Anantharaman, V.; Iyer, L. M.; Aravind, L. Ter-Dependent Stress Response Systems: Novel Pathways Related to Metal Sensing, Production of a Nucleoside-like Metabolite, and DNA-Processing. Mol. Biosyst. 2012, 8, 3142–3165. [Google Scholar] [CrossRef]
- Makarova, K. S.; Wolf, Y. I.; Koonin, E. V. Towards Functional Characterization of Archaeal Genomic Dark Matter. Biochem. Soc. Trans. 2019, 47, 389–398. [Google Scholar] [CrossRef]
- Hess, D. C.; Myers, C. L.; Huttenhower, C.; Hibbs, M. A.; Hayes, A. P.; Paw, J.; Clore, J. J.; Mendoza, R. M.; Luis, B. S.; Nislow, C.; Giaever, G.; Costanzo, M.; Troyanskaya, O. G.; Caudy, A. A. Computationally Driven, Quantitative Experiments Discover Genes Required for Mitochondrial Biogenesis. PLOS Genet. 2009, 5, e1000407. [Google Scholar] [CrossRef]
- Deckers, M.; Balleininger, M.; Vukotic, M.; Römpler, K.; Bareth, B.; Juris, L.; Dudek, J. Aim24 Stabilizes Respiratory Chain Supercomplexes and Is Required for Efficient Respiration. FEBS Lett. 2014, 588, 2985–2992. [Google Scholar] [CrossRef]
- Tran, T. T.; Panesso, D.; Gao, H.; Roh, J. H.; Munita, J. M.; Reyes, J.; Diaz, L.; Lobos, E. A.; Shamoo, Y.; Mishra, N. N.; Bayer, A. S.; Murray, B. E.; Weinstock, G. M.; Arias, C. A. Whole-Genome Analysis of a Daptomycin-Susceptible Enterococcus Faecium Strain and Its Daptomycin-Resistant Variant Arising during Therapy. Antimicrob. Agents Chemother. 2013, 57, 261–268. [Google Scholar] [CrossRef]
- Sumby, P.; Smith, M. C. M. Genetics of the Phage Growth Limitation (Pgl) System of Streptomyces Coelicolor A3(2). Mol. Microbiol. 2002, 44, 489–500. [Google Scholar] [CrossRef]
- Valkovicova, L.; Vavrova, S. M.; Mravec, J.; Grones, J.; Turna, J. Protein–Protein Association and Cellular Localization of Four Essential Gene Products Encoded by Tellurite Resistance-Conferring Cluster “Ter” from Pathogenic Escherichia Coli. Antonie Van Leeuwenhoek 2013, 104, 899–911. [Google Scholar] [CrossRef] [PubMed]
- McCorquodale, D. J.; Warner, H. R. Bacteriophage T5 and Related Phages. In The Bacteriophages; Calendar, R., Ed.; Springer US: Boston, MA, 1988. [Google Scholar] [CrossRef]
- Gillor, O.; Kirkup, B. C.; Riley, M. A. Colicins and Microcins: The next Generation Antimicrobials. Adv. Appl. Microbiol. 2004, 54, 129–146. [Google Scholar] [PubMed]
- Hilsenbeck, J. L.; Park, H.; Chen, G.; Youn, B.; Postle, K.; Kang, C. Crystal Structure of the Cytotoxic Bacterial Protein Colicin B at 2.5 Å Resolution. Mol. Microbiol. 2004, 51, 711–720. [Google Scholar] [CrossRef]
- Pilsl, H.; Braun, V. Strong Function-Related Homology between the Pore-Forming Colicins K and 5. J. Bacteriol. 1995, 177, 6973–6977. [Google Scholar] [CrossRef]
- Sanssouci, É.; Lerat, S.; Grondin, G.; Shareck, F.; Beaulieu, C. Tdd8: A TerD Domain-Encoding Gene Involved in Streptomyces Coelicolor Differentiation. Antonie Van Leeuwenhoek 2011, 100, 385–398. [Google Scholar] [CrossRef]
- Daigle, F.; Lerat, S.; Bucca, G.; Sanssouci, É.; Smith, C. P.; Malouin, F.; Beaulieu, C. A terD Domain-Encoding Gene (SCO2368) Is Involved in Calcium Homeostasis and Participates in Calcium Regulation of a DosR-Like Regulon in Streptomyces Coelicolor. J. Bacteriol. 2015, 197, 913–923. [Google Scholar] [CrossRef]
- Thomas, L.; Hodgson, D. A.; Wentzel, A.; Nieselt, K.; Ellingsen, T. E.; Moore, J.; Morrissey, E. R.; Legaie, R.; Consortium, T. S.; Wohlleben, W.; Rodríguez-García, A.; Martín, J. F.; Burroughs, N. J.; Wellington, E. M. H.; Smith, M. C. M. Metabolic Switches and Adaptations Deduced from the Proteomes of Streptomyces Coelicolor Wild Type and phoP Mutant Grown in Batch Culture *. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef]
- Langlois, P.; Bourassa, S.; Poirier, G. G.; Beaulieu, C. Identification of Streptomyces Coelicolor Proteins That Are Differentially Expressed in the Presence of Plant Material. Appl. Environ. Microbiol. 2003, 69, 1884–1889. [Google Scholar] [CrossRef]
- Novotna, J.; Vohradsky, J.; Berndt, P.; Gramajo, H.; Langen, H.; Li, X.-M.; Minas, W.; Orsaria, L.; Roeder, D.; Thompson, C. J. Proteomic Studies of Diauxic Lag in the Differentiating Prokaryote Streptomyces Coelicolor Reveal a Regulatory Network of Stress-Induced Proteins and Central Metabolic Enzymes. Mol. Microbiol. 2003, 48, 1289–1303. [Google Scholar] [CrossRef]
- Tiffert, Y.; Franz-Wachtel, M.; Fladerer, C.; Nordheim, A.; Reuther, J.; Wohlleben, W.; Mast, Y. Proteomic Analysis of the GlnR-Mediated Response to Nitrogen Limitation in Streptomyces Coelicolor M145. Appl. Microbiol. Biotechnol. 2011, 89, 1149–1159. [Google Scholar] [CrossRef]
- Novotna, J.; Vohradsky, J.; Berndt, P.; Gramajo, H.; Langen, H.; Li, X.-M.; Minas, W.; Orsaria, L.; Roeder, D.; Thompson, C. J. Proteomic Studies of Diauxic Lag in the Differentiating Prokaryote Streptomyces Coelicolor Reveal a Regulatory Network of Stress-Induced Proteins and Central Metabolic Enzymes. Mol. Microbiol. 2003, 48, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Manteca, A.; Mäder, U.; Connolly, B. A.; Sanchez, J. A Proteomic Analysis of Programmed Cell Death. PROTEOMICS 2006, 6, 6008–6022. [Google Scholar] [CrossRef] [PubMed]
- Tenconi, E.; Traxler, M. F.; Hoebreck, C.; van Wezel, G. P.; Rigali, S. Production of Prodiginines Is Part of a Programmed Cell Death Process in Streptomyces Coelicolor. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Sevciková, B.; Benada, O.; Kofronova, O.; Kormanec, J. Stress-Response Sigma Factor σH Is Essential for Morphological Differentiation of Streptomyces Coelicolor A3(2). Arch. Microbiol. 2001, 177, 98–106. [Google Scholar] [CrossRef]
- Kelemen, G. H.; Viollier, P. H.; Tenor, J.; Marri, L.; Buttner, M. J.; Thompson, C. J. A Connection between Stress and Development in the Multicellular Prokaryote Streptomyces Coelicolor A3(2). Mol. Microbiol. 2001, 40, 804–814. [Google Scholar] [CrossRef]
- Wang, S.-L.; Fan, K.-Q.; Yang, X.; Lin, Z.-X.; Xu, X.-P.; Yang, K.-Q. CabC, an EF-Hand Calcium-Binding Protein, Is Involved in Ca2+-Mediated Regulation of Spore Germination and Aerial Hypha Formation in Streptomyces Coelicolor. J. Bacteriol. 2008, 190, 4061–4068. [Google Scholar] [CrossRef]
- Clapham, D. E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Carafoli, E. Intracellular Calcium Homeostasis. Annu. Rev. Biochem. 1987, 56, 395–433. [Google Scholar] [CrossRef]
- Domínguez, D. C.; Guragain, M.; Patrauchan, M. Calcium Binding Proteins and Calcium Signaling in Prokaryotes. Cell Calcium 2015, 57, 151–165. [Google Scholar] [CrossRef]
- Ramstad, S.; Futsaether, C. M.; Johnsson, A. Porphyrin Sensitization and Intracellular Calcium Changes in the Prokaryote Propionibacterium Acnes. J. Photochem. Photobiol. B 1997, 40, 141–148. [Google Scholar] [CrossRef]
- Werthén, M.; Lundgren, T. Intracellular Ca2+ Mobilization and Kinase Activity during Acylated Homoserine Lactone-Dependent Quorum Sensing in Serratia Liquefaciens *. J. Biol. Chem. 2001, 276, 6468–6472. [Google Scholar] [CrossRef] [PubMed]
- Torrecilla, I.; Leganés, F.; Bonilla, I.; Fernández-Piñas, F. Use of Recombinant Aequorin to Study Calcium Homeostasis and Monitor Calcium Transients in Response to Heat and Cold Shock in Cyanobacteria1. Plant Physiol. 2000, 123, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Watkins, N. J.; Knight, M. R.; Trewavas, A. J.; Campbell, A. K. Free Calcium Transients in Chemotactic and Non-Chemotactic Strains of Escherichia Coli Determined by Using Recombinant Aequorin. Biochem. J. 1995, 306, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A. K.; Naseem, R.; Wann, K.; Holland, I. B.; Matthews, S. B. Fermentation Product Butane 2,3-Diol Induces Ca2+ Transients in E. Coli through Activation of Lanthanum-Sensitive Ca2+ Channels. Cell Calcium 2007, 41, 97–106. [Google Scholar] [CrossRef]
- Campbell, A. K.; Naseem, R.; Holland, I. B.; Matthews, S. B.; Wann, K. T. Methylglyoxal and Other Carbohydrate Metabolites Induce Lanthanum-Sensitive Ca2+ Transients and Inhibit Growth in E. Coli. Arch. Biochem. Biophys. 2007, 468, 107–113. [Google Scholar] [CrossRef]
- Herbaud, M.-L.; Guiseppi, A.; Denizot, F.; Haiech, J.; Kilhoffer, M.-C. Calcium Signalling in Bacillus Subtilis. Biochim. Biophys. Acta BBA - Mol. Cell Res. 1998, 1448, 212–226. [Google Scholar] [CrossRef]
- Luan, S.; Wang, C. Calcium Signaling Mechanisms Across Kingdoms. Annu. Rev. Cell Dev. Biol. 2021, 37, 311–340. [Google Scholar] [CrossRef]
- Farias, P.; Francisco, R.; Morais, P. V. Potential of Tellurite Resistance in Heterotrophic Bacteria from Mining Environments. iScience 2022, 25, 104566. [Google Scholar] [CrossRef]
- Peng, W.; Wang, Y.; Fu, Y.; Deng, Z.; Lin, S.; Liang, R. Characterization of the Tellurite-Resistance Properties and Identification of the Core Function Genes for Tellurite Resistance in Pseudomonas Citronellolis SJTE-3. Microorganisms 2022, 10, 95. [Google Scholar] [CrossRef]
- Chiang, S.-K.; Lou, Y.-C.; Chen, C. NMR Solution Structure of KP-TerB, a Tellurite-Resistance Protein from Klebsiella Pneumoniae. Protein Sci. 2008, 17, 785–789. [Google Scholar] [CrossRef]
- Joint Center for Structural Genomics (JCSG). Crystal Structure of Tellurite Resistance Protein of COG3793 (ZP_0010 9916.1) from Nostoc Punctiforme PCC 73102 at 1.85 A Resolution, 2007. [CrossRef]
- Walsh, P.; Bursać, D.; Law, Y. C.; Cyr, D.; Lithgow, T. The J-protein Family: Modulating Protein Assembly, Disassembly and Translocation. EMBO Rep. 2004, 5, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; van der Laan, M.; Mehnert, C. S.; Guiard, B.; Mick, D. U.; Hutu, D. P.; Truscott, K. N.; Wiedemann, N.; Meisinger, C.; Pfanner, N.; Rehling, P. Distinct Forms of Mitochondrial TOM-TIM Supercomplexes Define Signal-Dependent States of Preprotein Sorting. Mol. Cell. Biol. 2010, 30, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Dym, O.; Eisenberg, D. Sequence-Structure Analysis of FAD-Containing Proteins. Protein Sci. 2001, 10, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Marbois, B.; Gin, P.; Gulmezian, M.; Clarke, C. F. The Yeast Coq4 Polypeptide Organizes a Mitochondrial Protein Complex Essential for Coenzyme Q Biosynthesis. Biochim. Biophys. Acta BBA - Mol. Cell Biol. Lipids 2009, 1791, 69–75. [Google Scholar] [CrossRef]
- Rosso, M.-L.; Chauvaux, S.; Dessein, R.; Laurans, C.; Frangeul, L.; Lacroix, C.; Schiavo, A.; Dillies, M.-A.; Foulon, J.; Coppée, J.-Y.; Médigue, C.; Carniel, E.; Simonet, M.; Marceau, M. Growth of Yersinia Pseudotuberculosis in Human Plasma: Impacts on Virulence and Metabolic Gene Expression. BMC Microbiol. 2008, 8, 211. [Google Scholar] [CrossRef]
- Ott, E.; Kawaguchi, Y.; Kölbl, D.; Chaturvedi, P.; Nakagawa, K.; Yamagishi, A.; Weckwerth, W.; Milojevic, T. Proteometabolomic Response of Deinococcus Radiodurans Exposed to UVC and Vacuum Conditions: Initial Studies Prior to the Tanpopo Space Mission. PLoS ONE 2017, 12, e0189381. [Google Scholar] [CrossRef]
- Yang, S.; Xu, H.; Wang, J.; Liu, C.; Lu, H.; Liu, M.; Zhao, Y.; Tian, B.; Wang, L.; Hua, Y. Cyclic AMP Receptor Protein Acts as a Transcription Regulator in Response to Stresses in Deinococcus Radiodurans. PLoS ONE 2016, 11, e0155010. [Google Scholar] [CrossRef]
- Appukuttan, D.; Seo, H. S.; Jeong, S.; Im, S.; Joe, M.; Song, D.; Choi, J.; Lim, S. Expression and Mutational Analysis of DinB-Like Protein DR0053 in Deinococcus Radiodurans. PLoS ONE 2015, 10, e0118275. [Google Scholar] [CrossRef]
- Basu, B.; Apte, S. K. Gamma Radiation-Induced Proteome of Deinococcus Radiodurans Primarily Targets DNA Repair and Oxidative Stress Alleviation. Mol. Cell. Proteomics MCP 2012, 11, M111.011734. [Google Scholar] [CrossRef]
- Tanaka, M.; Earl, A. M.; Howell, H. A.; Park, M.-J.; Eisen, J. A.; Peterson, S. N.; Battista, J. R. Analysis of Deinococcus Radiodurans’s Transcriptional Response to Ionizing Radiation and Desiccation Reveals Novel Proteins That Contribute to Extreme Radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef]
- Kormutakova, R.; Klucar, L.; Turna, J. DNA Sequence Analysis of the Tellurite-Resistance Determinant from Clinical Strain of Escherichia Coli and Identification of Essential Genes.
- Toptchieva, A.; Sisson, G.; Bryden, L. J.; Taylor, D. E.; Hoffman, P. S. An Inducible Tellurite-Resistance Operon in Proteus Mirabilis. Microbiology 2003, 149, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Zeinert, R.; Martinez, E.; Schmitz, J.; Senn, K.; Usman, B.; Anantharaman, V.; Aravind, L.; Waters, L. S. Structure–Function Analysis of Manganese Exporter Proteins across Bacteria. J. Biol. Chem. 2018, 293, 5715–5730. [Google Scholar] [CrossRef] [PubMed]
- Tsu, B. V.; Saier, M. H. The LysE Superfamily of Transport Proteins Involved in Cell Physiology and Pathogenesis. PLoS ONE 2015, 10, e0137184. [Google Scholar] [CrossRef] [PubMed]
- Dambach, M.; Sandoval, M.; Updegrove, T. B.; Anantharaman, V.; Aravind, L.; Waters, L. S.; Storz, G. The Ubiquitous yybP-ykoY Riboswitch Is a Manganese-Responsive Regulatory Element. Mol. Cell 2015, 57, 1099–1109. [Google Scholar] [CrossRef]
- Bingham, R. J.; Hall, K. S.; Slonczewski, J. L. Alkaline Induction of a Novel Gene Locus, Alx, in Escherichia Coli. J. Bacteriol. 1990, 172, 2184–2186. [Google Scholar] [CrossRef]
- Sharma, R.; Mishanina, T. V. A Riboswitch-Controlled TerC Family Transporter Alx Tunes Intracellular Manganese Concentration in Escherichia Coli at Alkaline pH. J. Bacteriol. 2024, 206, e00168–24. [Google Scholar] [CrossRef]
- Anjem, A.; Varghese, S.; Imlay, J. A. Manganese Import Is a Key Element of the OxyR Response to Hydrogen Peroxide in Escherichia Coli. Mol. Microbiol. 2009, 72, 844–858. [Google Scholar] [CrossRef]
- Aguirre, J. D.; Culotta, V. C. Battles with Iron: Manganese in Oxidative Stress Protection *. J. Biol. Chem. 2012, 287, 13541–13548. [Google Scholar] [CrossRef]
- Imlay, J. A. The Mismetallation of Enzymes during Oxidative Stress *. J. Biol. Chem. 2014, 289, 28121–28128. [Google Scholar] [CrossRef]
- Martin, J. E.; Imlay, J. A. The Alternative Aerobic Ribonucleotide Reductase of Escherichia Coli, NrdEF, Is a Manganese-Dependent Enzyme That Enables Cell Replication during Periods of Iron Starvation. Mol. Microbiol. 2011, 80, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Breckau, D.; Mahlitz, E.; Sauerwald, A.; Layer, G.; Jahn, D. Oxygen-Dependent Coproporphyrinogen III Oxidase (HemF) from Escherichia Coli Is Stimulated by Manganese *. J. Biol. Chem. 2003, 278, 46625–46631. [Google Scholar] [CrossRef] [PubMed]
- Martin, J. E.; Waters, L. S.; Storz, G.; Imlay, J. A. The Escherichia Coli Small Protein MntS and Exporter MntP Optimize the Intracellular Concentration of Manganese. PLOS Genet. 2015, 11, e1004977. [Google Scholar] [CrossRef]
- Chen Qinyuan; Guo Fang; Huang Li; Wang Mengying; Shi Chunfeng; Zhang Shutong; Yao Yizhou; Wang Mingshu; Zhu Dekang; Jia Renyong; Chen Shun; Zhao Xinxin; Yang Qiao; Wu Ying; Zhang Shaqiu; Tian Bin; Huang Juan; Ou Xumin; Gao Qun; Sun Di; Zhang Ling; Yu Yanling; He Yu; Wu Zhen; Götz Friedrich; Cheng Anchun; Liu Mafeng. Functional Characterization of a TerC Family Protein of Riemerella Anatipestifer in Manganese Detoxification and Virulence. Appl. Environ. Microbiol. 2023, 90, e01350–23. [Google Scholar] [CrossRef]
- Guo Fang; Wang Mengying; Huang Mi; Jiang Yin; Gao Qun; Zhu Dekang; Wang Mingshu; Jia Renyong; Chen Shun; Zhao Xinxin; Yang Qiao; Wu Ying; Zhang Shaqiu; Huang Juan; Tian Bin; Ou Xumin; Mao Sai; Sun Di; Cheng Anchun; Liu Mafeng. Manganese Efflux Achieved by MetA and MetB Affects Oxidative Stress Resistance and Iron Homeostasis in Riemerella Anatipestifer. Appl. Environ. Microbiol. 2023, 89, e01835–22. [Google Scholar] [CrossRef]
- Grunenwald, C. M.; Choby, J. E.; Juttukonda, L. J.; Beavers, W. N.; Weiss, A.; Torres, V. J.; Skaar, E. P. Manganese Detoxification by MntE Is Critical for Resistance to Oxidative Stress and Virulence of Staphylococcus Aureus. MBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- He, B.; Sachla, A. J.; Helmann, J. D. TerC Proteins Function during Protein Secretion to Metalate Exoenzymes. Nat. Commun. 2023, 14, 6186. [Google Scholar] [CrossRef]
- Huang, X.; Shin, J.; Pinochet-Barros, A.; Su, T. T.; Helmann, J. D. Bacillus Subtilis MntR Coordinates the Transcriptional Regulation of Manganese Uptake and Efflux Systems. Mol. Microbiol. 2017, 103, 253–268. [Google Scholar] [CrossRef]
- He, B.; Helmann, J. D. Metalation of Extracytoplasmic Proteins and Bacterial Cell Envelope Homeostasis. Annual Review of Microbiology, 2024, 78, 83–102. [Google Scholar] [CrossRef]
- Grunenwald, C. M.; Choby, J. E.; Juttukonda, L. J.; Beavers, W. N.; Weiss, A.; Torres, V. J.; Skaar, E. P. Manganese Detoxification by MntE Is Critical for Resistance to Oxidative Stress and Virulence of Staphylococcus Aureus. mBio 2019, 10, 10.1128–mbio.02915. [Google Scholar] [CrossRef]
- Díaz-Vásquez, W. A.; Abarca-Lagunas, M. J.; Arenas, F. A.; Pinto, C. A.; Cornejo, F. A.; Wansapura, P. T.; Appuhamillage, G. A.; Chasteen, T. G.; Vásquez, C. C. Tellurite Reduction by Escherichia Coli NDH-II Dehydrogenase Results in Superoxide Production in Membranes of Toxicant-Exposed Cells. BioMetals 2014, 27, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Goff, J. L.; Boyanov, M. I.; Kemner, K. M.; Yee, N. The Role of Cysteine in Tellurate Reduction and Toxicity. BioMetals 2021, 34, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Borsetti, F.; Tremaroli, V.; Michelacci, F.; Borghese, R.; Winterstein, C.; Daldal, F.; Zannoni, D. Tellurite Effects on Rhodobacter Capsulatus Cell Viability and Superoxide Dismutase Activity under Oxidative Stress Conditions. Res. Microbiol. 2005, 156, 807–813. [Google Scholar] [CrossRef]
- Piacenza, E.; Campora, S.; Carfì Pavia, F.; Chillura Martino, D. F.; Laudicina, V. A.; Alduina, R.; Turner, R. J.; Zannoni, D.; Presentato, A. Tolerance, Adaptation, and Cell Response Elicited by Micromonospora Sp. Facing Tellurite Toxicity: A Biological and Physical-Chemical Characterization. Int. J. Mol. Sci. 2022, 23, 12631. [Google Scholar] [CrossRef] [PubMed]
- Srinand Paruthiyil; Azul Pinochet-Barros; Xiaojuan Huang; John D. Helmann; Tina M. Henkin. Bacillus Subtilis TerC Family Proteins Help Prevent Manganese Intoxication. J. Bacteriol. 2020, 202, e00624–19. [Google Scholar] [CrossRef]
- Chen, I.-M. A.; Chu, K.; Palaniappan, K.; Ratner, A.; Huang, J.; Huntemann, M.; Hajek, P.; Ritter, S. J.; Webb, C.; Wu, D.; Varghese, N. J.; Reddy, T. B. K.; Mukherjee, S.; Ovchinnikova, G.; Nolan, M.; Seshadri, R.; Roux, S.; Visel, A.; Woyke, T.; Eloe-Fadrosh, E. A.; Kyrpides, N. C.; Ivanova, N. N. The IMG/M Data Management and Analysis System v.7: Content Updates and New Features. Nucleic Acids Res. 2023, 51, D723–D732. [Google Scholar] [CrossRef]
- Li, W.; O’Neill, K. R.; Haft, D. H.; DiCuccio, M.; Chetvernin, V.; Badretdin, A.; Coulouris, G.; Chitsaz, F.; Derbyshire, M. K.; Durkin, A. S.; Gonzales, N. R.; Gwadz, M.; Lanczycki, C. J.; Song, J. S.; Thanki, N.; Wang, J.; Yamashita, R. A.; Yang, M.; Zheng, C.; Marchler-Bauer, A.; Thibaud-Nissen, F. RefSeq: Expanding the Prokaryotic Genome Annotation Pipeline Reach with Protein Family Model Curation. Nucleic Acids Res. 2021, 49, D1020–D1028. [Google Scholar] [CrossRef]
- Suprenant, K. A.; Bloom, N.; Fang, J.; Lushington, G. The Major Vault Protein Is Related to the Toxic Anion Resistance Protein(TelA) Family. J. Exp. Biol. 2007, 210, 946–955. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, J.; Zeng, W.; Xiao, Q.; Zhu, Y.; Kong, W.; Zou, J.; Liu, T.; Yin, H. Dissecting the Metal Resistance Genes Contributed by Virome from Mining-Affected Metal Contaminated Soils. Front. Environ. Sci. 2023, 11. [Google Scholar] [CrossRef]
- Goff, J. L.; Lui, L. M.; Nielsen, T. N.; Poole, F. L., II; Smith, H. J.; Walker, K. F.; Hazen, T. C.; Fields, M. W.; Arkin, A. P.; Adams, M. W. W. Mixed Waste Contamination Selects for a Mobile Genetic Element Population Enriched in Multiple Heavy Metal Resistance Genes. ISME Commun. 2024, ycae064. [Google Scholar] [CrossRef]
- Moore, C. J.; Bornemann, T. L. V.; Figueroa-Gonzalez, P. A.; Esser, S. P.; Moraru, C.; Soares, A. R.; Hinzke, T.; Trautwein-Schult, A.; Maaß, S.; Becher, D.; Starke, J.; Plewka, J.; Rothe, L.; Probst, A. J. Time-Series Metaproteogenomics of a High-CO2 Aquifer Reveals Active Viruses with Fluctuating Abundances and Broad Host Ranges. microLife 2024, 5, uqae011. [Google Scholar] [CrossRef] [PubMed]
- Decewicz, P.; Dziewit, L.; Golec, P.; Kozlowska, P.; Bartosik, D.; Radlinska, M. Characterization of the Virome of Paracoccus Spp. (Alphaproteobacteria) by Combined in Silico and in Vivo Approaches. Sci. Rep. 2019, 9, 7899. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Peng, D.; Fu, C.; Luo, X.; Guo, S.; Li, L.; Yin, H. Pan-Metagenome Reveals the Abiotic Stress Resistome of Cigar Tobacco Phyllosphere Microbiome. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Labonté, J. M.; Pachiadaki, M.; Fergusson, E.; McNichol, J.; Grosche, A.; Gulmann, L. K.; Vetriani, C.; Sievert, S. M.; Stepanauskas, R. Single Cell Genomics-Based Analysis of Gene Content and Expression of Prophages in a Diffuse-Flow Deep-Sea Hydrothermal System. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Sazinas, P.; Smith, C.; Suhaimi, A.; Hobman, J. L.; Dodd, C. E. R.; Millard, A. D. Draft Genome Sequence of the Bacteriophage vB_Eco_slurp01. Genome Announc. 2016, 4, 10.1128–genomea.01111. [Google Scholar] [CrossRef]
- Estrada Bonilla, B.; Costa, A. R.; Van Den Berg, D. F.; Van Rossum, T.; Hagedoorn, S.; Walinga, H.; Xiao, M.; Song, W.; Haas, P.-J.; Nobrega, F. L. Genomic Characterization of Four Novel Bacteriophages Infecting the Clinical Pathogen Klebsiella Pneumoniae. DNA Res. 2021, 28, dsab013. [Google Scholar] [CrossRef]
- Rahlff, J.; Esser, S. P.; Plewka, J.; Heinrichs, M. E.; Soares, A.; Scarchilli, C.; Grigioni, P.; Wex, H.; Giebel, H.-A.; Probst, A. J. Marine Viruses Disperse Bidirectionally along the Natural Water Cycle. Nat. Commun. 2023, 14, 6354. [Google Scholar] [CrossRef]
- Takeuchi, N.; Hamada-Zhu, S.; Suzuki, H. Prophages and Plasmids Can Display Opposite Trends in the Types of Accessory Genes They Carry. Proc. R. Soc. B Biol. Sci. 2023, 290, 20231088. [Google Scholar] [CrossRef]
- Bucher, M. J.; Czyż, D. M. Phage against the Machine: The SIE-Ence of Superinfection Exclusion. Viruses 2024, 16, 1348. [Google Scholar] [CrossRef]
- Bondy-Denomy, J.; Qian, J.; Westra, E. R.; Buckling, A.; Guttman, D. S.; Davidson, A. R.; Maxwell, K. L. Prophages Mediate Defense against Phage Infection through Diverse Mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef]
- Dyllick-Brenzinger, M.; Liu, M.; Winstone, T. L.; Taylor, D. E.; Turner, R. J. The Role of Cysteine Residues in Tellurite Resistance Mediated by the TehAB Determinant. Biochem. Biophys. Res. Commun. 2000, 277, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Chasteen, T. G.; Bentley, R. Biomethylation of Selenium and Tellurium: Microorganisms and Plants. Chem. Rev. 2003, 103, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Piacenza, E.; Campora, S.; Carfì Pavia, F.; Chillura Martino, D. F.; Laudicina, V. A.; Alduina, R.; Turner, R. J.; Zannoni, D.; Presentato, A. Tolerance, Adaptation, and Cell Response Elicited by Micromonospora Sp. Facing Tellurite Toxicity: A Biological and Physical-Chemical Characterization. Int. J. Mol. Sci. 2022, 23, 12631. [Google Scholar] [CrossRef]
- Molina-Quiroz, R. C.; Loyola, D. E.; Muñoz-Villagrán, C. M.; Quatrini, R.; Vásquez, C. C.; Pérez-Donoso, J. M. DNA, Cell Wall and General Oxidative Damage Underlie the Tellurite/Cefotaxime Synergistic Effect in Escherichia Coli. PLOS ONE 2013, 8, e79499. [Google Scholar] [CrossRef]
- Pérez, J. M.; Calderón, I. L.; Arenas, F. A.; Fuentes, D. E.; Pradenas, G. A.; Fuentes, E. L.; Sandoval, J. M.; Castro, M. E.; Elías, A. O.; Vásquez, C. C. Bacterial Toxicity of Potassium Tellurite: Unveiling an Ancient Enigma. PloS One 2007, 2, e211. [Google Scholar] [CrossRef]
- Turkovicova, L.; Smidak, R.; Jung, G.; Turna, J.; Lubec, G.; Aradska, J. Proteomic Analysis of the TerC Interactome: Novel Links to Tellurite Resistance and Pathogenicity. J. Proteomics 2016, 136, 167–173. [Google Scholar] [CrossRef]
- Ross, D. E.; Ruebush, S. S.; Brantley, S. L.; Hartshorne, R. S.; Clarke, T. A.; Richardson, D. J.; Tien, M. Characterization of Protein-Protein Interactions Involved in Iron Reduction by Shewanella Oneidensis MR-1. Appl. Environ. Microbiol. 2007, 73, 5797–5808. [Google Scholar] [CrossRef]
- Turner, R. J.; Taylor, D. E.; Weiner, J. H. Expression of Escherichia Coli TehA Gives Resistance to Antiseptics and Disinfectants Similar to That Conferred by Multidrug Resistance Efflux Pumps. Antimicrob. Agents Chemother. 1997, 41, 440–444. [Google Scholar] [CrossRef]
Figure 1.
Proposed model for the functioning of the TeR proteins during tellurite stress (left) and alternative functions (right). Activities with strong experimental evidence are indicated by solid arrows while more speculative activities are indicated by dashed arrows. Cell stressors are indicated by red text. [1] TehA may export lipid peroxidation-derived aldehydes that accumulate during tellurite exposure [50]. [2] The activity of TehA may be regulated directly or indirectly (not shown) by the methyltransferase enzymes TehB [163]. TehB is a SAM-dependent methyltransferase that also non-specifically methylated tellurite methylated tellurides [22], which can then diffuse out of the cell [164] (note that SAM = S=adenosyl-L-methionine and AdoHcy = S-adenosyl-L-homocysteine). [3] The role of TelA in the cellular response to tellurite stress is enigmatic; however, it may be involved in the cellular response to tellurite-induced cell envelope damage [165,166,167]. [4] TerC may function to fine-tune intracellular manganese concentrations during tellurite stress [129]. Tellurite exposure increases cytoplasmic reactive oxygen species (ROS), leading to increased expression of superoxide dismutase (SOD) [167]. TerC may balance the influx of Mn2+ ions imported by a manganese importer such as MntH [131]. [5] Alternatively, TerC may metalate Mn-utilizing exometalloenzymes that are critical to the cellular response to tellurite stress [139]. These enzymes may be metalated during or after their export via the SecYEG translocon. In Gram-positive bacteria, this could include LtaS, involved in lipoteichoic acid biosynthesis or other, yet-uncharacterized, enzymes. [6] TerD domain-containing proteins are likely involved in mediating Ca2+-based cell signaling [63,87]. An influx of Ca2+ may be triggered my tellurite exposure, as a stress signal. [7] The role of TerB in the cellular response to tellurite stress is enigmatic; however the protein interacts with both TerD and TerC and may help coordinate the responses of the two enzymes to tellurite stress [168,169]. [8] TehA is a multi-drug efflux pump that exports quaternary ammonium compounds such as ethidium bromide [170]. However, given the essentiality of TehA in the virulence of human pathogens [44], it is likely that the pump has alternative functions, like other multi-drug efflux pumps. For example, TehA promotes host colonization, thus it may export compounds like autoinducers and virulence-related toxins [34,35]. [9] TehA is also involved in the cellular response to reactive nitrogen stress (RNS) [37,38], possibly exporting toxic nitrosated biomolecules [34]. [10] TehA may be regulated directly or indirectly by TehB through its SAM-dependent methyltransferase activity [24,163]. [11] TelA is involved in resistance to cell envelope-targeting antimicrobials. However, the mechanism through which this occurs is not known. [12] Under certain conditions, such as alkaline pH stress, TerC protects cells against manganese intoxication by acting as a Mn2+ efflux pump [129,147]. [13] TerC interacts with the SecYEG translocon to metalate proteins while they are being secreted. For certain exoenzymes, TerC helps generate sufficiently high local Mn2+ to facilitate their post-translocational metalation [139]. [14] TerD domains are integrated into diverse cellular processes, including numerous stress response systems, via their Ca2+ binding activity. External stressors such as antibiotics, other chemicals, and phage infection may trigger transient increases in cytoplasmic Ca2+. [15] A mechanism of activity is not well-described for TerB. However, it is known to interact with both TerC and TerD [81,168] and has been implicated into the cellular response to diverse stressors including UV irradiation [118], oxidative stress [119], and phage infection [12]. Thus, TerC, TerD, and TerB may all have functions in generalized stress responses.
Figure 1.
Proposed model for the functioning of the TeR proteins during tellurite stress (left) and alternative functions (right). Activities with strong experimental evidence are indicated by solid arrows while more speculative activities are indicated by dashed arrows. Cell stressors are indicated by red text. [1] TehA may export lipid peroxidation-derived aldehydes that accumulate during tellurite exposure [50]. [2] The activity of TehA may be regulated directly or indirectly (not shown) by the methyltransferase enzymes TehB [163]. TehB is a SAM-dependent methyltransferase that also non-specifically methylated tellurite methylated tellurides [22], which can then diffuse out of the cell [164] (note that SAM = S=adenosyl-L-methionine and AdoHcy = S-adenosyl-L-homocysteine). [3] The role of TelA in the cellular response to tellurite stress is enigmatic; however, it may be involved in the cellular response to tellurite-induced cell envelope damage [165,166,167]. [4] TerC may function to fine-tune intracellular manganese concentrations during tellurite stress [129]. Tellurite exposure increases cytoplasmic reactive oxygen species (ROS), leading to increased expression of superoxide dismutase (SOD) [167]. TerC may balance the influx of Mn2+ ions imported by a manganese importer such as MntH [131]. [5] Alternatively, TerC may metalate Mn-utilizing exometalloenzymes that are critical to the cellular response to tellurite stress [139]. These enzymes may be metalated during or after their export via the SecYEG translocon. In Gram-positive bacteria, this could include LtaS, involved in lipoteichoic acid biosynthesis or other, yet-uncharacterized, enzymes. [6] TerD domain-containing proteins are likely involved in mediating Ca2+-based cell signaling [63,87]. An influx of Ca2+ may be triggered my tellurite exposure, as a stress signal. [7] The role of TerB in the cellular response to tellurite stress is enigmatic; however the protein interacts with both TerD and TerC and may help coordinate the responses of the two enzymes to tellurite stress [168,169]. [8] TehA is a multi-drug efflux pump that exports quaternary ammonium compounds such as ethidium bromide [170]. However, given the essentiality of TehA in the virulence of human pathogens [44], it is likely that the pump has alternative functions, like other multi-drug efflux pumps. For example, TehA promotes host colonization, thus it may export compounds like autoinducers and virulence-related toxins [34,35]. [9] TehA is also involved in the cellular response to reactive nitrogen stress (RNS) [37,38], possibly exporting toxic nitrosated biomolecules [34]. [10] TehA may be regulated directly or indirectly by TehB through its SAM-dependent methyltransferase activity [24,163]. [11] TelA is involved in resistance to cell envelope-targeting antimicrobials. However, the mechanism through which this occurs is not known. [12] Under certain conditions, such as alkaline pH stress, TerC protects cells against manganese intoxication by acting as a Mn2+ efflux pump [129,147]. [13] TerC interacts with the SecYEG translocon to metalate proteins while they are being secreted. For certain exoenzymes, TerC helps generate sufficiently high local Mn2+ to facilitate their post-translocational metalation [139]. [14] TerD domains are integrated into diverse cellular processes, including numerous stress response systems, via their Ca2+ binding activity. External stressors such as antibiotics, other chemicals, and phage infection may trigger transient increases in cytoplasmic Ca2+. [15] A mechanism of activity is not well-described for TerB. However, it is known to interact with both TerC and TerD [81,168] and has been implicated into the cellular response to diverse stressors including UV irradiation [118], oxidative stress [119], and phage infection [12]. Thus, TerC, TerD, and TerB may all have functions in generalized stress responses.
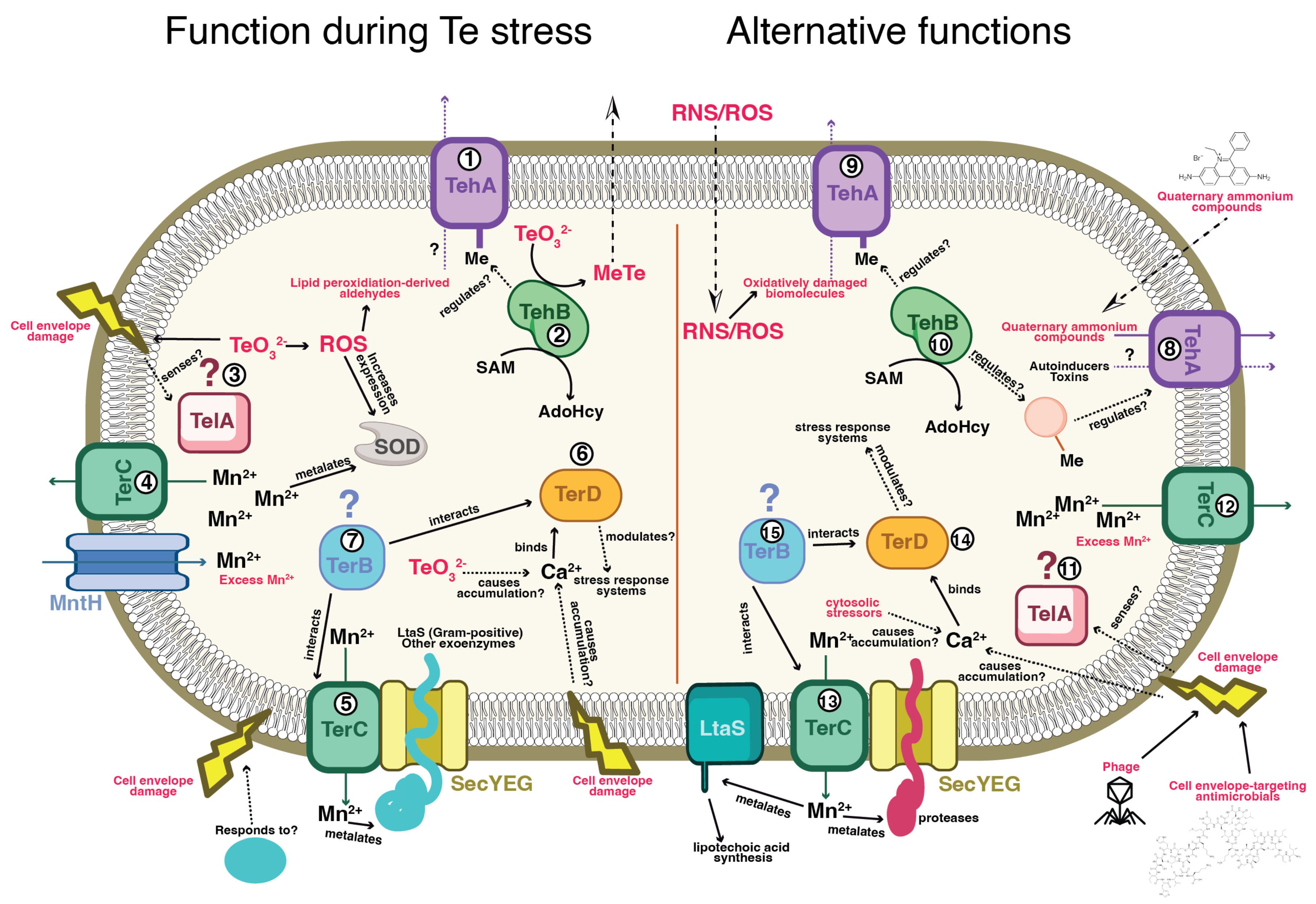
Figure 2.
Publications citing the BacMet database containing the terms “tellurium” or “tellurite”.
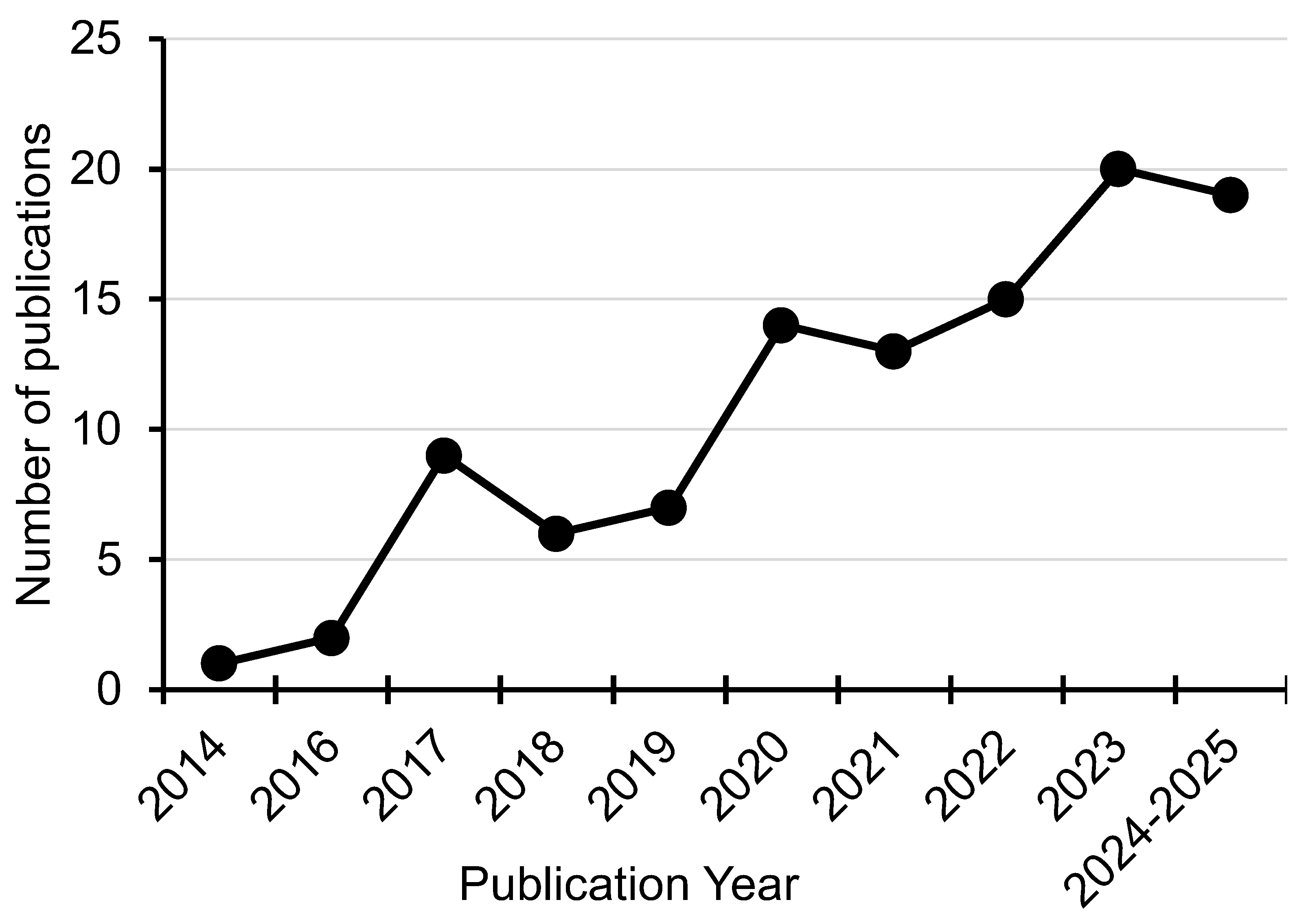

Table 1.
Analysis of annotated genomes in the JGI IMG/M database. 137,572 total genomes and metagenomes were analyzed. The data in the second column of the table shows the number of genomes that contain at least one annotation for the ter, teh, tel, or trg gene products. The third column shows the number of genomes that contain at least one annotation that links the ter, teh, tel, or trg gene products with the terms “tellurite” or “tellurium”.
Table 1.
Analysis of annotated genomes in the JGI IMG/M database. 137,572 total genomes and metagenomes were analyzed. The data in the second column of the table shows the number of genomes that contain at least one annotation for the ter, teh, tel, or trg gene products. The third column shows the number of genomes that contain at least one annotation that links the ter, teh, tel, or trg gene products with the terms “tellurite” or “tellurium”.
| Genome Hits: “protein name” (% total) | Genome Hits: “protein name” + “tellurite”/“tellurium” (% total) | |
|---|---|---|
| ter gene products | ||
| TerZ | 10,506 (7.6%) | 10,347 (7.5%) |
| TerA | 11,237 (8.2%) | 11,221 (8.2%) |
| TerB | 19,633 (14.3%) | 17,809 (12.9%) |
| TerC | 112,963 (82.1%) | 91,432 (66.5%) |
| TerD | 19,266 (14.0%) | 17,415 (12.7%) |
| TerE | 45 (0.03%) | 28 (0.02%) |
| TerF | 1,895 (1.4%) | 7 (0.005%) |
| teh gene products | ||
| TehA | 11,682 (8.5%) | 11,679 (8.5%) |
| TehB | 2,495 (1.8%) | 2,493 (1.8%) |
| tel gene products | ||
| TelA | 557 (0.4%) | 11 (0.008%) |
| TelB | No hits | No hits |
| trg gene products | ||
| TrgA | 8 (0.006%) | 5 (0.004%) |
| TrgB | 11 (0.008%) | 8 (0.006%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



