Submitted:
18 April 2025
Posted:
21 April 2025
You are already at the latest version
Abstract
The non-coding RNAs (ncRNAs) have been reported in numerous studies, and their significant roles in several diseases have been verified subsequently. tRNA-derived fragments (tRFs) are a newly discovered class of small ncRNAs which are produced by mature or precursor tRNAs. In light of the development of RNA sequencing, the evidences have shown that tRFs are widely involved in the generation and progression of diseases by series mechanisms including RNA silencing, translational regulation, epigenetic regulation, reverse-transcriptional regulation, cellular apoptosis. Several researches have identified tRFs participate varied aspects in several cancers. Furthermore, these novel tRFs may hold significant potential as both diagnostic biomarkers and therapeutic targets in clinical applications. In this review, we discuss the biogenesis and classification of tRFs, illustrate the fundamental functions of tRFs, and summarize the most recent discoveries of tRFs in cancers.
Keywords:
non-coding RNA
; tRNA-derived fragments
; biomarker
; cancer
1. Introduction
Non-coding RNAs (ncRNAs) have been the focus of many studies over the last few decades, including long non-coding RNAs (lncRNAs) and small non-coding RNAs (sncRNAs), in various biological processes [1]. There is increasing evidence that sncRNAs play critical roles in multiple regulatory processes, including transcription, post-transcription, and translation. The involvement of sncRNAs in complex mechanisms is vital to the development and progression of cancers and their potential value as biomarkers and/or therapeutic targets has gaining heightened focus [2,3].
With the development of high-throughput sequencing technology and improvements in bioinformatics analysis, in addition to endogenous siRNAs (endosiRNAs), microRNAs (miRNAs), and Piwi-interacting RNAs (piRNAs) [4], researchers have uncovered the existence of a new type of sncRNAs derived from tRNAs, called tRNA-derived fragments (tRFs). tRFs, 14 to 32 nt long single-stranded RNA, are generated from mature tRNAs or precursor tRNAs at different sites under the control of a set of highly conservative and precise site-specific cutting mechanisms [1,5,6]. Emerging evidence indicates that tRFs may participate in cell proliferation, priming of viral reverse transcriptases, regulation of gene expression, RNA processing, modulation of the DNA damage response, tumor suppression, and neurodegeneration [7]. Moreover, tRFs have emerged as key contributors to the development and progression of multiple human pathologies, including cancer [8,9], inflammation [10], metabolic disorder [11], infection [12,13,14], and neurodegenerative disease [15,16]. Here, we will review the biogenesis and classification of tRFs and demonstrate parts of the major biological functions of tRFs as well as recent reports on the roles of tRFs in different types of cancer.
2. Biogenesis and Classification
tRNAs experience comprehensive processing and a series of chemical modifications throughout their lifecycle. In eukaryocyte, pre-tRNAs undergo ribonuclease P and ribonuclease Z cleavage of the 5’ leader and 3’ tail sequences respectively [17,18]. And the addition of the CCA tail is catalyzed by CCA-adding enzyme (tRNA nucleotidyltransferase) [19]. This is accompanied by further modifications, ultimately leading to the formation of mature tRNA. The structure of mature tRNA is highly conserved and characterized by a distinctive configuration comprising four arms (stem and loop): the D-arm, anticodon arm, TψC arm, acceptor arm, and variable arm at the connection of the anticodon arm and the TΨC arm [20,21]. Mature tRNAs or pre-tRNAs can be cleaved at specific positions by corresponding ribonucleases, thereby generating a diverse array of tRFs [22,23,24]. As classified, tRNAs can generate 5 subtypes of tRFs: tRF-5 (5’-tRF), tRF-3(3’-tRF), i-tRF, tRNA halves and tRF-1. The former four types of tRFs are derived from mature tRNAs, and tRF-1s come from pre-tRNA.
tRF-5s are produced by cleavage of the 5’ end in D-arm and tRF-3s are produced through cleavage of the 3’ end in TψC arm, respectively [25,26]. tRF-5s can be classified into three specific subtypes for diverse length: i) type a (14–16 nt), ii) type b (22–24 nt), and iii) type c (28–30 nt), and the cleavage sites for type a, b, and c locates in the D loop, D stem, and the 5’ half of the anticodon stem [5,27]. The tRF-3s can be cleaved into two subcategories: tRF-3a and tRF-3b. The diversity between these two subtypes is that the members of tRF-3b family are ~4 nt longer than the tRF-3a family, which the length of tRF-3a is 18 nt and tRF-3b is 22 nt long approximately [5,28]. Generally, tRF-5s and tRF-3s are generated from Dicer-dependent fashion [14,28]; however, now evidences from subsequent investigations have revealed that angiogenin (ANG) and other members of the ribonuclease A superfamily are actively involved in the biogenesis of these tRFs [28,29].
i-tRFs are a new type of tRF that originate from the internal zone of mature tRNAs. Different starting positions of cleavage define the different i-tRF subtypes. Based on the location of the 5’terminus along the mature tRNA, one research divided i-tRFs into six subtypes [30]. In addition, there are other classification methodologies. The generation of i-tRFs may be associated with some specific conditions such as hypoxia [31]. The biogenesis and generation of i-tRFs are demonstrated incompletely and still require further research.
tRNA halves, also defined as tRNA-derived stress-induced RNAs (tiRNAs), are generated through the cleavage of mature tRNAs at the anticodon arm by ANG, divided into 5’-half and 3’-half [32]. 5’-halves have 30–35 nts ranging from the 5’ end to the anticodon loop, while 3’- halves have 40–50 nts from the anticodon loop to the 3’ end [6,26]. As a stress-inducible ribonuclease, the biological function of ANG exhibits a strong correlation with the cellular status. Under various stress conditions including heat shock, ultraviolet irradiation, hypoxia, arsenite exposure, lack of amino acids, and viral infection, the specific cleavage of tRNAs by ANG is typically induced, leading to the generation of tRNA halves [26,33,34,35]. One study found that ANG is not the only RNase that produces tRNA halves, as other RNases may also be involved in this procession [36]. Whereas, tRNA halves are often produced disproportionately, leading to a much more amount of 5’-halves than 3’-halves [37].
Distinct from other tRFs, tRF-1s are produced from tRNA maturation, which is originated from the 3’ trailer fragment of tRNA before the addition of CCA tail trimmed by the endonuclease Z (RNaseZ/ELAC2) [23,38]. With 16–48 nt in length, tRF-1s begin the downstream of the 3′ end in the pre-tRNA and end with a polyuridine sequence (UUUUU, UUCUU, AUCUU, or GUCUU) [23,39]. However, tRF-1s are almost cytoplasmic, which is considered to undergo translocation following their biogenesis in the nucleus [40].
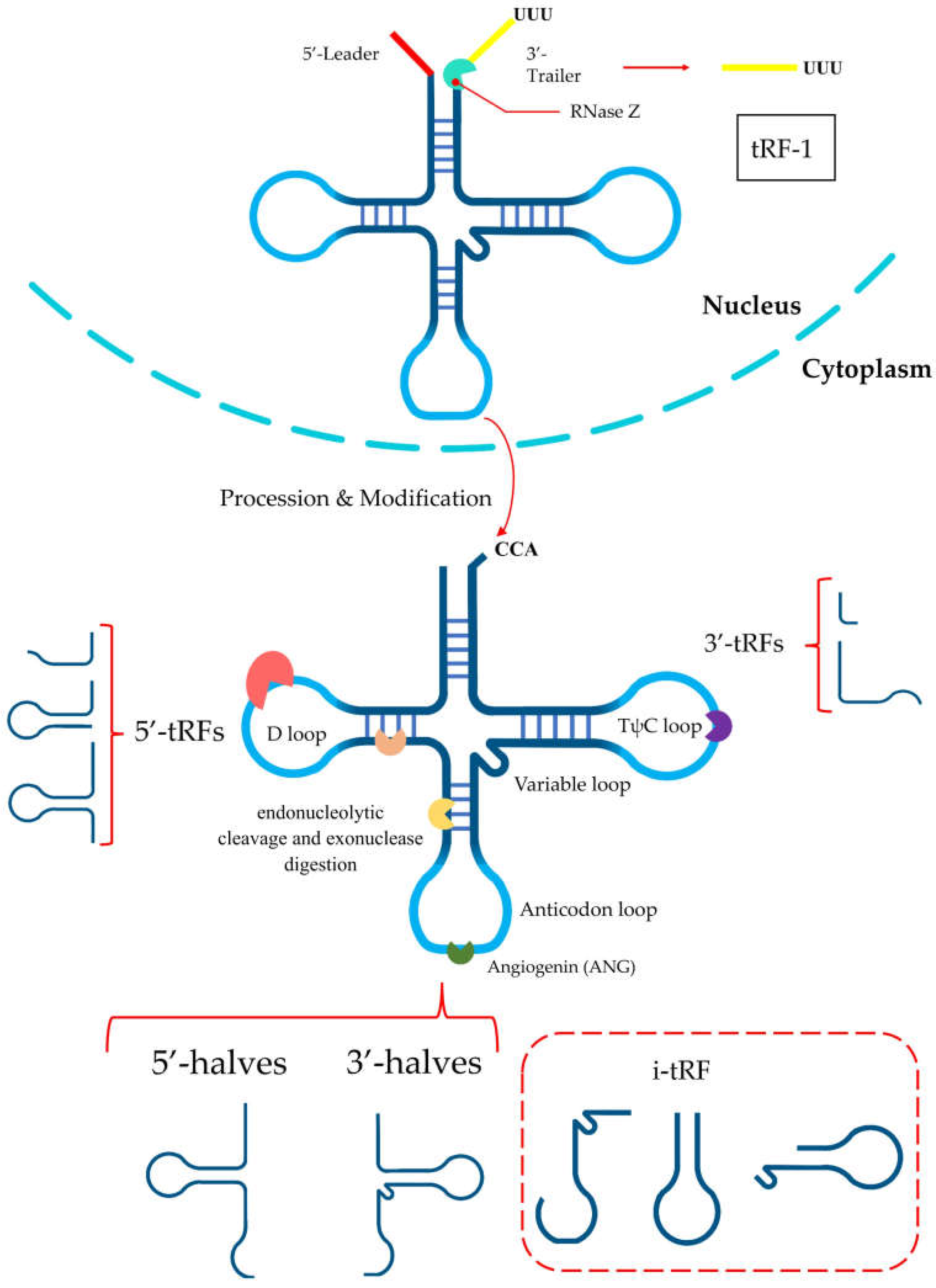
Figure 1.
Different types of tRNA-derived RNA fragments. tRF-5s are produced by cleavage of the 5’ end in D-arm and tRF-3s are produced through cleavage of the 3’ end in TψC arm. tRNA halves are generated through the cleavage of mature tRNAs at the anticodon arm by ANG. i-tRFs are a new type of tRF that originate from the internal zone of mature tRNAs. tRF-1s are produced from tRNA maturation.
Figure 1.
Different types of tRNA-derived RNA fragments. tRF-5s are produced by cleavage of the 5’ end in D-arm and tRF-3s are produced through cleavage of the 3’ end in TψC arm. tRNA halves are generated through the cleavage of mature tRNAs at the anticodon arm by ANG. i-tRFs are a new type of tRF that originate from the internal zone of mature tRNAs. tRF-1s are produced from tRNA maturation.
3. Biological Functions of tRFs
Increasing depth of RNA sequencing has enabled the discovery of the Biogenesis and the clarification of Categories of tRFs, which gives us an opportunity to Explore the fundamental functions of tRFs. Although the biological functions of tRFs are complicated and necessitate additional investigation, our current knowledge of their function has been summarized into five categories as follows, including RNA silencing, translational regulation, epigenetic regulation, reverse-transcriptional regulation, cellular apoptosis.
3.1. RNA Silencing
Recent researches cast more light on tRFs’ important roles in RNA interference (RNAi)-mediated silencing. Several studies have indicated that disparate tRFs show distinct affinities with various AGO subtypes. Meta-analysis of PAR-CLIP data reveals that tRF-5s and tRF-3s are associated with AGO1, 3 and 4 rather than AGO2, and analysis of positional T to C mutational frequency indicates the pattern of the association between tRFs and Argonautes is similar to that of miRNAs [5]. For example, Green’s group illustrated that a specific tRF namely tRF-3003a,produced by the cleavage of tRNA-CysGCA confers gene silencing of Janus Kinase 3 (JAK3) via AGO/RISC formation in osteoarthritis chondrocytes [41]. In contrast to microRNAs, tRFs exhibit a distinct subcellular localization predominantly within the cytoplasmic compartment and demonstrate selective binding to Argonautes, while notably lacking association with the MOV10. Moreover, tRF-3s exhibits canonical microRNA-/siRNA-like trans-silencing capacity, while tRF-1 does not exhibit apparent trans-silencing activity [38]. Further study shows that Dicer-dependent tRFs promote gene silencing through a mechanism distinct from PTGS and TGS which can lead to downregulation of target genes by targeting introns via nascent RNA silencing (NRS) in nuclei [42]. In addition to AGO-dependent mechanism for gene silencing by tRFs, AGO-independent mechanisms have been confirmed to exist, including competitive binding of target proteins with mRNAs. Hee et al. demonstrated RF-U3-1 inhibits HCV IRES-mediated translation by sequestering the limiting amounts of La/SSB available in the cytoplasm via La/SSB knockout Huh7 cell lines [43]. Hence, tRFs might be involved in multiple pathways to regulate RNA silencing which is urgingly needed to be discovered.
3.2. Translational Regulation
tRFs can modulate translational processes through the regulation of translation initiation, elongation, and ribosome biogenesis, thereby exerting an influence on the overall synthesis of proteins. The involvement of tRFs in the regulation of translation was initially discovered in stress-related conditions [32]. It has identified that transfection of natural 5’-halves, but not 3’-halves, results in the suppression of global translation in U2OS cells [32]. As previously mentioned, cellular stress leads to the production of tRNA halves by ANG. These specific 5’-tiRNAs such as 5’-tiRNAAla and 5’-tiRNACys can possess the ability to displace the mRNA cap-binding protein eIF4E [44]. Further research found this restraint is achieved by the aggregation of 5’-tiRNAs into tetrameric G-quadruplexes (G4) mediated through the 5’ terminal oligoguanine (5’ TOG) motif [45,46]. G4-tiRNAs, confirmed to be necessary for tRFs in the regulation of mRNA translation, can directly binding to the HEAT1 domain of eIF4G [45,47]. And this displacement can induce the phospho-eIF2α independent assembly of stress granules (SGs) and leading to the failure of the translation initiation scanning process, consequently suppressing the global translation [48].
Besides tRNA halves, the tRF-5s can also affect translation. A study demonstrated in embryonic stem cells, the Ψ modification at U8 mediated by PUS7 can motivate TOG -containing tRF-5s to bind to polyadenylate-binding protein 1 (PABPC1), a protein in charge of eIF4G/A interaction and initiation of cap-dependent translation. The mTOG-PABPC1 complex can intercept PABPC1 recruitment to eIF4F, thus blocking translation [49]. And a general conserved “GG” dinucleotide in tRF-5s is also revealed to engage the process of protein translation [50].
A considerable fact in the production of tsRNAs is the 5-methylcytosine (m5C) modification. NSun2 is a specific cytosine-5 RNA methyltransferase of proteins that methylates the cytidine residues of most tRNAs. When in abstraction of NSun2, the loss of cytosine-5 RNA methylation increases the cleavage of tRNA resulting an accumulation of 5’tRFs. These abnormal processes reduce protein translation rates eventually [37].
Besides, tRFs can also promote protein generation through strengthening translation-induced ribosome biogenesis. Binding at least two ribosomal protein (RPS28 and RPS15) mRNAs, a kind of 3’-tRF from tRNALeu (CAG) can enhance translation [51].
3.3. Epigenetic Regulation
The regulation of genetic expression is mediated through an integrated interplay between genomic DNA sequences and epigenetic regulatory mechanisms. Epigenetics regulate gene expression mainly through DNA methylation, histone modification, nucleosome remodeling, and ncRNA regulation [52]. Previous studies have highlighted the evidence suggesting that Epigenetic mis-regulation can culminate in human diseases, such as cancer [53]. Here we will summarize potential epigenetic mechanisms of tRFs into two basic parts: PIWI-independent epigenetic regulation and PIWI-dependent epigenetic regulation.
Transposons are genetic sequences that can translocate their sites within a genome [54], which together with their repetitive sequences contribute to the formation and function of chromosomes, and induce epigenetic regulation of specific genes. Because of the high mobility and mutation of transposons, eukaryotic cells have evolved retiform epigenetic mechanisms in order to preserve the genetic integrity [55]. Furthermore, Substantial evidence has shown that various cancers are significantly correlated with the transcriptional activity of transposons which is concordant with above statement. Recently, Andrea et al. discovered that tRF-3s (18 and 22 nt in length) containing the 3′-terminal CCA sequence of mature tRNAs could inhibit long terminal repeat (LTR)-retrotransposons (also known as endogenous retroviruses; ERVs) by different mechanisms involving sequence complementarity with the PBS sequence [56]. Besides, Sharma et al. identified that 5′ tRF-Gly-GCC (tRF-GG) affects chromatin accessibility in MERVL elements and throughout heterochromatin in embryonic stem (ES) cells and preimplantation embryos [57]. Sharma’s discovery suggests that the underlying mechanisms of interactions between tRFs and chromatin remain to be elucidated, thereby offering novel insights for future investigations.
Next, we will briefly introduce PIWI-dependent epigenetic regulation of tRFs. Couvillion et al. revealed the potential connection between tRF-3s, 18–22 nt length tsRNA with the predominant 5’ end in the TΨC loop and the 3’ end at the mature tRNA 3’ terminus, and the Tetrahymena thermophila Ago/Piwi protein Twi12 and their essential roles in epigenetic regulation [58]. They discovered that tRF-3 is implicated in nuclear translocation of Twi12, while Twi12 plays essential roles in ribosomal RNA processing by assembling with Xrn2 and Tan1 proteins [58]. Another research elucidated that IL-4 potently decreases the biogenesis of a PIWI-interacting RNAs (piRNAs), tRNA-Glu–derived piRNAs [td-piR(Glu)]. Further, Xue Zhang et al. revealed that the td-piR(Glu)/PIWIL4 complex recruits SETDB1, SUV39H1, and heterochromatin protein 1b to the CD1A promoter region and facilitates H3K9 methylation, leading to the significant inhibition of transcription of CD1A [59]. These results suggest that td-piR(Glu) participates in chromatin remodeling in immune cells.
3.4. Reverse-Transcriptional Regulation
A part of roles of tRFs in reverse transcription have also been demonstrated by some studies including gene expression and oncology [60]. A study conducted in mice has identified that 3’-tRFs are crucial regulators of the retrotransposons in the cell [61]. Transposable elements (TEs) actuate transcription, and in mice, the most active transposons are long terminal repeat (LTR)-retrotransposons, also called as endogenous retroviruses (ERVs). There is a large amount of 3’-tRFs attacking ERVs in mouse stem cell lines. The 18 nt 3’-tRFs specifically restrain ERVs’ activity by competing with tRNAs for the highly conserved primer binding site (PBS) of ERVs, resulting the blockade of reverse transcription. Meanwhile, it is noteworthy that the 22 nt 3’-tRFs can also bind to the PBS sequence, which affect transposon expression by post-transcriptionally silencing of ERV mRNA [61]. Both of these two 3’-tRFs inhibit ERVs activity by different mechanisms but both depending on base complementarity with PBS sequence.
And in viral, especially in retrovirus infection, tRFs can affect the reverse transcription. Human T cell lymphotroic virus Type I (HTLV-1) is a human retrovirus first discovered [62]. In infected CD4 cells, tRF-3s are considerably more abundant than other tRFs [63]. Among the tRF-3s, tRF-3019 corresponds to the 3′ end of tRNA-Pro, the tRNA considered to be the primer for HTLV-1 reverse transcriptase [64]. The portion of tRNA-Pro corresponding to tRF-3019 is complementary to the HTLV-1 PBS, which may reveal that the tRF would be fully sufficient as a primer for reverse transcription. As a reverse transcription primer, this special kind of tRF-3 can induce HTLV-1 to initiate reverse transcription and accumulate the replication of virus [60,63].
3.5. Cellular Apoptosis
Recent studies have indicated the potential role of tRFs in regulating cell apoptosis. Previous reports demonstrated that apoptosome formation in vitro was inhibited by the addition of in vitro-synthesized full-length tRNAs [65,66,67]. Subsequent research has disclosed that angiogenin-induced accumulation of tRNA halves (or tiRNAs) is associated with increased survival in hyper-osmotically stressed mouse embryonic fibroblasts by inhibiting cell apoptosis. The underlying mechanism is that Cyt c released from the mitochondria during hyperosmotic stress forms RNPs with tRNAs, which disturb the formation of the apoptosome by interfering the interaction of apoptotic protease activating factor-1 (APAF1) and Cyt c. Saikia et al. revealed a novel signaling pathway that interferes with Cyt c-mediated caspase activation, a novel function of tiRNAs as inhibitors of apoptosome formation and function, and a novel antiapoptotic mechanism involving ANG-mediated tRNA cleavage. However, how cytochrome c recognizes specific tRNA targets needs further research [68].
Another study discovered a novel tRFs, tRF-3022b, which shows an increasing trend in CRC tissues compared to adjacent normal tissues, utilizing whole-transcriptome sequencing technology. tRF-3022b plays a vital role in apoptosis inhibition and CRC progression through the regulatory pathway by binding to galectin 1 (LGALS1) and macrophage migration inhibitory factor (MIF) in CRC cells [69]. But tRFs are not always play the bad guy in cancer. Ling Pan et al. identified an inflammatory cytokine–regulated transfer RNA–derived (tRNA-derived) fragment, tRF-21-VBY9PYKHD (tRF-21), which suppresses the progression of pancreatic ductal adenocarcinoma (PDAC) by enhancing apoptosis. Further research revealed that tRF-21 knockdown promotes the phosphorylation of heterogeneous nuclear ribonucleoprotein L (hnRNP L) and the formation of hnRNP L and dead-box helicase 17 (DDX17) complexes. These complexes play crucial roles in splicing Caspase 9 into Caspase 9b (with anti-apoptotic specificity) and mH2A1.2 (with pro-invasive specificity), while upregulation of tRF-21 exerts the opposite effect [70].
4. tRFs in Cancer
Through deep sequencing and microarrays, more tRFs have been detected and promoted the development of several tRF-related databases [51,71]. Thus, the function of tRFs in specific disease is clear gradually. The occurrence and progression of cancers are closely associated with abnormal gene expression. More empirical data substantiates the involvement of tRFs in a multitude of biological mechanisms that are intricately linked to the oncogenesis and advancement of malignancies. One possible explanation is rapid proliferation in tumor cells often results in deficiency of blood supply leading to hypoxic stress and other tumor microenvironment (TME). To adapt the stressful environments, tumors adopt several unique mechanisms which may promote the production of tRFs for responses to stress. Given the distinct spatiotemporal characteristics of various cancer cell types, the interactions between different cancer cells and their respective internal tRFs exhibit significant heterogeneity.
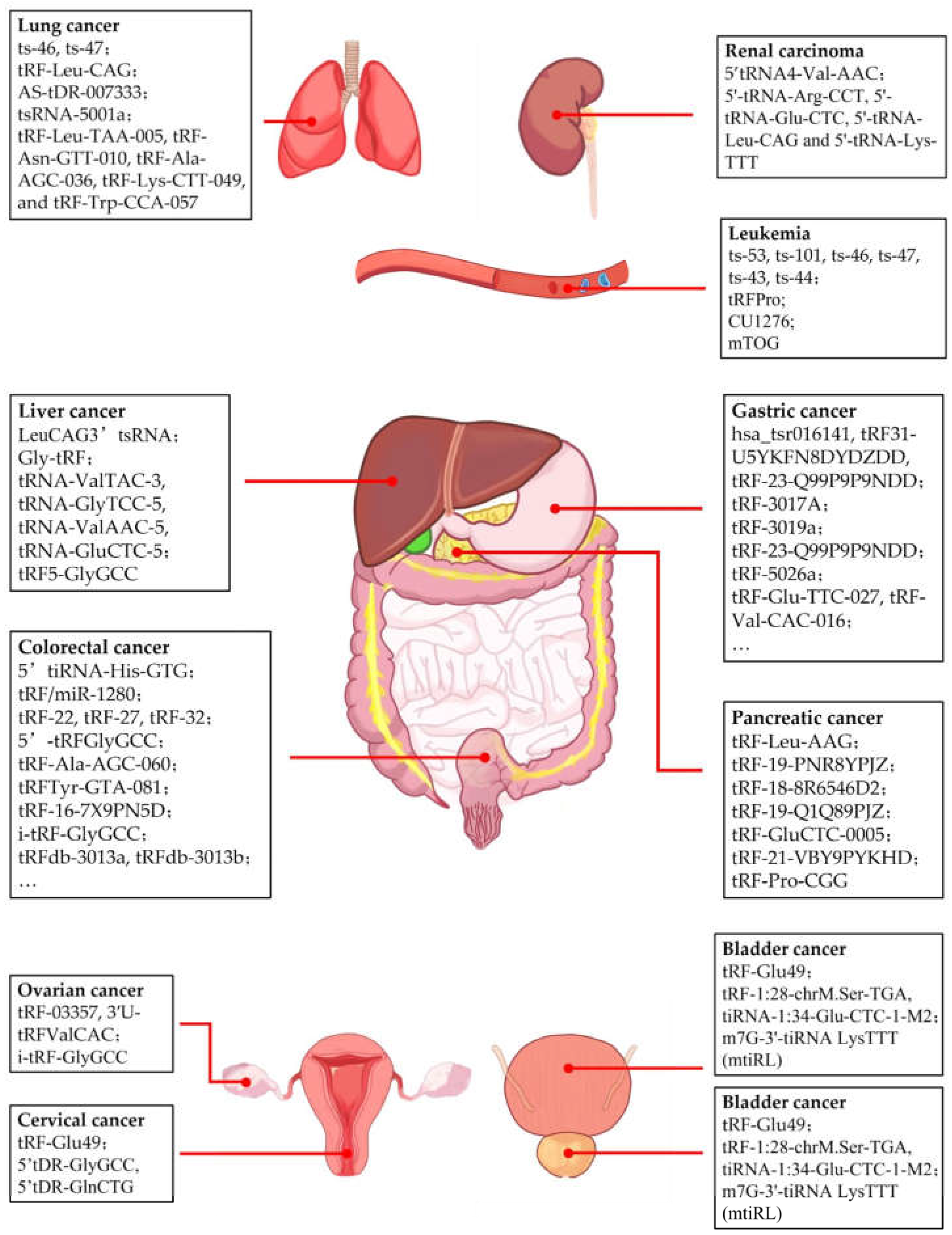
Figure 2.
Fuctions of tRFs in different kinds of cancer. tRFs are associated with many types of cancer including breast cancer, prostate cancer, pancreatic cancer, liver cancer, gastric cancer, colorectal cancer, leukemia, lung cancer, and other cancers. These tRFs play differing roles in different types of cancer by various mechanisms.
Figure 2.
Fuctions of tRFs in different kinds of cancer. tRFs are associated with many types of cancer including breast cancer, prostate cancer, pancreatic cancer, liver cancer, gastric cancer, colorectal cancer, leukemia, lung cancer, and other cancers. These tRFs play differing roles in different types of cancer by various mechanisms.
4.1. Breast Cancer
Breast cancers (BC) is the most common cancer in tRFs studies. The role of tRFs in BC has received increasing attention over the past few years. tRFs is serving as promising candidates for BC diagnostic and prognostic biomarkers and targets of possible therapy. A few tRFs have been identified as differentially expressed in BC [72]. High-throughput sequencing is applied to identifying differentially expressed tsRNAs in plasma samples of patients with BC, and a study has found some specific tsRNAs in plasma could play a role as diagnostic and prognostic biomarkers [73]. The circulating tRF-ArgCCT-017, tRF-Gly-CCC-001 and tiRNA-Phe-GAA-003 are upregulated in plasma samples of BC patients, indicating their potential as biomarkers [73]. By using custom tsRNA microarray chip, it has found a signature of 34 tsRNAs able to distinguish the normal cells with BC cells in comparation [72]. And the expression of the same tRFs also varies in two cell lines from different stages of BC, suggesting that tRF may play a regulatory role during tumor development.
Different kinds of tRFs have various functions in BC tumorigenesis and progression. A few tRFs have been found to act as enhancing factors in BC. Through regulation of ribosomal protein, tRF-19-W4PU732S from the mature tRNA-Ser-AGA can affect the cancer [74]. Targeting inhibition of RPL27A (ribosomal protein-L27A), tRF-19-W4PU732S promotes BC cell proliferation, malignant activity, invasion, EMT and CSC phenotypes, and the suppression of apoptosis. Sex hormone-dependent tRNA-derived RNAs (SHOT-RNAs) are specifically expressed in ER-positive BC cell lines, and 5′-SHOT-RNA can also accelerate cell proliferation in BC cells similarly [75]. Farina et al. demonstrated ts-112 as an oncogenic tRF which was selectively inhibited by tumor suppressor RUNX1 to prevent overactive expression in breast epithelial cells [76]. Recently it was discovered that tRF-33 from mature tRNA-LysTTT, was significantly upregulated in human epidermal receptor 2 (HER2)-negative BC cells and tissue [77]. By directly interacting with the 3’-UTR of IGF1, tRF-33 was identified to lead to the downregulation of IGF1 mRNA expression and protein synthesis, eventually disrupting mitochondrial homeostasis and contributing to the progression of BC pathogenesis.
Some tRFs that act as tumor suppressors have also been identified. Under hypoxic stress, a new class of tRFs is produced from tRNAGlu, tRNAAsp, tRNAGly, and tRNATyr [31]. This class of tRFs has a same motif that corresponds to the YBX1 implicated in cancer progression and shown to promote cancer metastasis [78,79]. These tRFs can inhibit the development of BC metastasis by binding YBX1 and displacing multiple oncogenic transcripts such as EIF4EBP1 and AKT1. There is a first study to show that 5′-tiRNAVal as a new tumor-suppressor can directly target the 3′-UTR sequence in frizzled class receptor 3 (FZD3), inhibit the FZD3-mediated Wnt/β-Catenin signaling pathway in BC cells [80]. And its downregulation in serum was positively related to BC pathological stage and lymph node metastasis. Coming from the 3’-end of the mature tRNAGluTTC, tRF3E is specifically expressed in healthy mammary glands but not in BC. This tRF specifically interacts with nucleolin (NCL), and causes the release of p53 mRNA promoting its translation by competing for NCL. Through NCL-mediated mechanism, tRF3E proposes inhibition of BC cell proliferation [81].
Triple-negative breast cancer (TNBC) is the most aggressive and malignant type of BC with restricted treatment options, chemoresistance and poor prognosis. By analysis of the expression profiles of tRFs in cancer stem cells (CSCs) isolated from TNBC and non-TNBC cell lines, research found the tDR-000620 acted as a novel candidate biomarker for the early detection of recurrence in TNBC patients independently, with the continuously lower expression level in TNBC CSCs and serum samples [82]. tRFLys-CTT-010 is significantly increased in TNBC and promotes TNBC proliferation and migration. tRFLys-CTT-010 interacted with glucose6-phosphatase catalytic subunit (G6PC) to regulate lactate production and glycogen consumption, leading to cell survival and proliferation [83]. It reminds us that the adjustment of glucose metabolism and tRFLys-CTT-010/G6PC axis may provide innovative therapeutic targets for TNBC treatment.
In treatment of BC, the modulation of the above tRFs may represent a promising approach to suppress tumor growth and progression. Several studies have demonstrated that these novel targets could lead to improved therapeutic efficacy. However, it should be noted that the drug resistance during treatment may also be associated with tRFs. tDR-0009 and tDR-7336 from hypoxia were found involved in maintenance of stem cell population and cellular response to IL-6, thereby facilitating the doxorubicin resistance in TNBC [84]. Besides, in HER2-positive BC patients, a specific monoclonal antibody trastuzumab has immensely prolongated the survival of these patients [85]. Overexpression of tRF-27 competitively binding to Ras GTPase-activating protein-binding proteins 1 and 2 (G3BPs) and activated mechanistic target of rapamycin complex 1(MTORC1) to enhance cell proliferation and entice the resistance of HER2-positive BC against trastuzumab [86]. Meanwhile, the potential of tRFs to alleviate chemoradiotherapy resistance is still noteworthy. It has discovered that the silencing of 3’tRF-AlaAGC enhanced the sensitivity of BC cells to Adriamycin through the NF-κb signaling pathway [87].
4.2. Prostate Cancer
Prostate cancer is the third leading cause of cancer-related death among men worldwide [88]. Previous study reported the discovery and differential expression of tRFs in clinical samples of PCa [89]. Michael et al. analyzed the expression of tRFs in fresh frozen patient samples derived from normal adjacent prostate and different stages of PCa by RNA-sequencing and found that the tRF-5s comprise the most abundant class of tRFs in general and represent the major class among upregulated tRFs. To the contrary, the tRF-3s types are dominant among downregulated tRFs in PCa [90]. Analysis of tRFs profiles from the Prostate Cancer (PRAD) datasets of The Cancer Genome Atlas (TCGA) found that tRFs have extensive correlations with messenger RNAs (mRNAs). These correlations are disrupted in PRAD, which suggests disruptions of the regulatory network in the disease state [91]. Researches above indicate that tRFs might play an important role in the pathogenesis of cancer.
Yong Sun Lee et al. discovered a specific tRF-1 called tRF-1001, generated in the cytoplasm by a prostate cancer susceptibility gene tRNA 39-endonuclease ELAC2, is necessary for cell proliferation in PCa [23]. In addition, the expression of 5′-SHOT-RNA also has significant functional involvement in cell proliferation. Sex HOrmone-dependent TRNA-derived RNAs (SHOT-RNAs) are specifically and abundantly expressed in androgen receptor (AR)-positive prostate cancer cell lines, which produced from amino-acylated mature tRNAs by angiogenin-mediated anticodon cleavage, promoted by sex hormones and their receptors [75].
As for the treatment, cisplatin is a standard treatment for prostate cancer. Changwon Yang et al. confirmed that tRF-315 derived from tRNALys prevents cisplatin-induced apoptosis and alleviated cisplatin-induced mitochondrial dysfunction in prostate cancer cells (LNCaP and DU145) by targeting the tumor suppressor gene GADD45A. Thus, tRF-315 diminishes therapeutic effect of cisplatin in prostate cancer [92].
Moreover, newly growing evidence highlights that METTL1 depletion causes the loss of m7G tRNA methylation and promotes the biogenesis of 5′terminal oligoguanine-containing tRNA fragments (5′TOGs), which enhances the synthesis of key regulators of tumor growth suppression, interferon pathway, and immune effectors, providing novel insights into prostate cancer therapeutic strategies [93].
4.3. Pancreatic Cancer
Extensive research has gradually revealed the pivotal role and clinical potential of tRFs in pancreatic cancer. The expression of tRF-Leu-AAG is observably upregulated in pancreatic cancer (PC) cells as compared to adjacent normal tissues detected by high-throughput second-generation sequencing techniques, positively correlated with cell proliferation, migration, and invasion. Studies have shown that UPF1, a downstream target gene of tRF-Leu-AAG, promotes the development of pancreatic cancer when downregulated [94]. Similarly, tRF-19-PNR8YPJZ promotes pancreatic cancer migration and invasion by acting through the AXIN2 axis [95]. Moreover, tRF-18-8R6546D2, a novel oncogenic factor, promotes PC malignancy partly by directly silencing ASCL2 and further regulating its downstream genes such as MYC and CASP3 [96]. To the contrary, the downregulation of tRF-19-Q1Q89PJZ in pancreatic cancer promotes cell proliferation and metastasis by inhibiting HK1 expression [97]. As for pancreatic ductal adenocarcinoma (PDAC), the upregulation of tRF-GluCTC-0005 enhances the mRNA stability of WDR1, thereby promoting cancer cell proliferation, migration, and invasion [98], and also plays a crucial role in PDAC liver metastasis [99]. On the other hand, tRF-21-VBY9PYKHD (tRF-21), a tumor suppressor in PDAC, promotes cancer cell proliferation, migration, and invasion when downregulated. The downregulation provokes hnRNP L-DDX17 activity in order to form the Caspase 9b and mH2A1.2, thereby promoting PDAC cell malignant phenotypes [70]. Study also highlights the role of tRFs as potential biomarkers for PDAC prognosis and therapy. Jun Li et al. revealed that the significant downregulation of tRF-Pro-CGG is associated with an advanced TNM stage (P=0.000) and the N stage (P=0.000) of patients. More importantly, low tRF-Pro-CGG expression predicts poor survival in PDAC patients (P=0.003) [100].
4.4. Liver Cancer
At present, research on the role of tRFs in liver cancer is still lacking, especially related to their molecular mechanisms. A specific tRF-3 derived from the Leu-CAG tRNA (LeuCAG3’tsRNA) was reported in 2017. Inhibition of LeuCAG3′tsRNA induced apoptosis in HeLa and HCT-116 cells in vitro and in a patient-derived orthotopic hepatocellular carcinoma model in mice. Behind molecular mechanism is that LeuCAG3′tsRNA interacts with RPS28 and RPS15, two ribosomal protein mRNAs, to enhance their translation and increase tumor cell viability [51]. Recent research reported that glycine tRNA-derived fragment (Gly-tRF), generated from the 5′-arm of glycine tRNA, was highly expressed in HCC cell lines and tumor tissues. Gly-tRF increased the liver cancer stem cell (LCSC) subpopulation proportion and LCSC-like cell properties, and promoted Epithelial-mesenchymal transition (EMT). Mechanistically, Gly-tRF decreased the level of Nedd4 family interacting protein 2 (NDFIP2) mRNA by binding to the NDFIP2 mRNA 3′ UTR and also activate the AKT signaling pathway [101]. Another research confirmed that Carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) is direct target of Gly-tRF. Overexpression of Gly-tRF enhances growth and migration of HCC cells in vivo [102]. These findings suggests that Gly-tRF could act as a tumor promoter and provide new targets for the prognostic evaluation and treatment of HCC. Accumulating evidence demonstrates that tRFs could also have potential clinical values, no matter in diagnosis, prognosis or treatment of HCC. Yi Zuo et al. built a random forests (RF) diagnostic model based on tsRNA profiling of paired tumor and adjacent normal samples. This model can be reliably utilized in the diagnosis of liver cancer and high- low risk classification of patients based upon tsRNA characterization [103]. Lei Zhu et al. compared exosomal tsRNA levels between liver cancer patients and healthy donors, revealed that levels of four tsRNAs (tRNA-ValTAC-3, tRNA-GlyTCC-5, tRNA-ValAAC-5 and tRNA-GluCTC-5) are significantly higher in liver cancer patients [104]. This finding demonstrates that plasma exosomal tsRNA could serve as a novel diagnostic biomarker for HCC. A growing number of studies have revealed that radiotherapy can not only killing tumor cells directly, but also enhance NK cell cytotoxicity in tumor cells by upregulating NKG2D ligands [105,106]. However, radiotherapy also suppresses the antitumor immunity of NK cells, which limits therapeutic efficacy [107,108]. Yihang Gong et al. revealed that glycocholic acid (GCA)/tRNA-derived fragment 5 (tRF5)-GlyGCC signaling axis is activated in mouse HCC model after radiotherapy. tRF5-GlyGCC epigenetically upregulates Runx2 by interacting with KDM6B and then transcriptionally activates ITGBL1 and S100A9 expression in HCC cells, which further reduces NK cell cytotoxicity directly and attracts myeloid-derived suppressor cell (MDSC) to inhibit NK cell function indirectly, respectively [109]. This research highlights the possibility that radiotherapy combined with tRF5-GlyGCC inhibitor in order to potentiate NK cell antitumor immunity and prevent HCC recurrence postresection.
4.5. Gastric Cancer
Increasing numbers of tRFs involved in Gastric cancer (GC) have been revealed. In GC tissues and serum, several researches have been demonstrated respectively that the overexpression of hsa_tsr016141, tRF31-U5YKFN8DYDZDD and tRF-23-Q99P9P9NDD were closely related to tumor grade, lymph node metastasis and invasion [110,111,112]. Derived from tRNA-Val-TAC, tRF-3017A regulated the invasion and migration of GC cells through forming the RNA-induced silencing complex (RISC) with Ago2 to target silencing tumor suppressor gene NELL2 [9]. Besides targeting the gene, another study showed tRF-3019a, a kind of 3’-tRF, bound with mRNA 3’UTR of tumor suppressor gene F-box protein 47 (FBXO47), leading to GC cell proliferation, migration and invasion [113]. Concurrently, certain upregulated tRFs can modulate metabolic processes within tumor cells. The overexpressed tRF-23-Q99P9P9NDD could bind to the 3’UTR site of acyl-coenzyme A dehydrogenase short/branched chain (ACADSB) thereby affecting GC lipid metabolism and ferroptosis and promote progression of tumor [114].
Additionally, a significant number of downregulated tRFs have been identified in GC. Some special tRFs, including tRF-33-P4R8YP9LON4VDP and tRF-193L7L73JD, disrupt the cell cycle of GC cells at G0/G1 phases, thereby suppressing their growth and proliferation [115,116]. Certain tRFs exert impacts on GC through their regulatory effects on signal transduction pathways. tRF-5026a inhibited the occurrence and development of GC through PI3K/AKT signaling pathway, however, it was observed in a low expression in GC tissues and cells [117]. And it also found that tRF-5026a can change cell cycle progression in GC. The MAPK signaling pathway is also discovered to be involved in the regulatory network orchestrated by GC-associated tRFs. tRF-Glu-TTC-027 showed the capacity in regulation of MAPK pathway probably targeting the sites of 3’ UTR in transforming growth factor beta 2 (TGFB2) [118]. Similarly, tRF-Val-CAC-016 directly targeted the Calcium Voltage-Gated Channel Subunit Alpha1 D (CACNA1d) to regulate MAPK signaling pathway in GC progression [119]. Meanwhile, tRF-Tyr bound directly to the hnRNPD protein and competitively displaced the binding of hnRNPD to the c-Myc 3’UTR, regulating the c-Myc/Bcl2/Bax pathway and ultimately inhibiting the progression of GC [120].
Now the utilization of these tRFs in GC detection is currently under exploration. A kind of 3’tRF tRF-Val directly bound to the EEF1A1, mediated its transport into the nucleus and promoted its interaction with MDM2, a specific p53 E3 ubiquitin ligase, thus promoting proliferation and invasion, and inhibited apoptosis in GC cells [121]. This finding may provide a new potential therapeutic target for GC and a new explanation for the occurrence of GC. And effective therapies may be achieved through modulation of the tRFs above which functions within the signal transduction pathway.
4.6. Colorectal Cancer
Colorectal cancer (CRC) has been the third most commonly diagnosed cancer worldwide and is the second leading cause of cancer death [122]. The mechanisms of tRFs in CRC remain under investigation. Some of tRFs can affect signaling pathways to act carcinogenic or anti-carcinogenic effects. A specific 5’-half, 5’tiRNA-His-GTG can induce cell apoptosis under the regulation of HIF1α/angiogenin (ANG) axis. It reduces the expression of LATS2, a core kinase within the Hippo signal pathway, and subsequently promotes CRC cells multiplication and tumor growth by inhibition of the Hippo pathway [35]. Research has found that tRF/miR-1280 targeted a notch ligand JAG2 to activate the Notch signal pathway and enhances CRC tumor stem cell activity, promoting proliferation and metastasis of CRC [123]. tRFs can also affect the progression of CRC by regulating transcription process. Specifically targeting to the SPIB, 5’tiRNA-Gly-GCC regulates the promoter region of STAT6 and stimulates STAT6 transcription, and positively modulates the cJAK1/STAT6 signaling pathway to contribute to the advancement of CRC collectively [124]. Influenced by tumor-associated mesenchymal cells within the TME, Tumor cells should undergo epithelial-mesenchymal transition (EMT) to gain the ability of metastasis, and the elevated expression of Claudin will accelerate EMT formation [125]. The anti-CRC mechanism of tRF-20M0NK5Y93 is to suppress Claudin-1 and attenuate the invasion, migration and metastasis of CRC [123].
Given the differential expression of tRFs in CRC patients, a growing body of research has been dedicated to investigating their potential biomarkers utility and developing new diagnostic methods. One study chose tRF-22/27/32 as plasma markers for CRC diagnose. By analyzing the RNA results of plasma from CRC patients and healthy individuals, these tRFs showed higher diagnostic values than traditional tumor markers [126]. And the combination of tRFs and traditional markers may get a more effective diagnostic result. When combined with CEA and CA199, the sensitivity and specificity of 5’-tRFGlyGCC in distinguishing CRC patients are 86% and 84% respectively, revealing clinical application potential [127]. Through the upregulated tRF-Ala-AGC-060 and downregulated tRFTyr-GTA-081 and combining with CEA, the diagnostic efficacy for CRC screening is better compared to individual diagnostic methods [128].
In therapy, radiotherapy and chemotherapy always serve as critical strategies in CRC treatment. Substantial downregulation of tRF-16-7X9PN5D was found in radioresistant CRC cells. It presents a link between tRNA and cancer radiation resistance that tRF-16-7X9PN5D could target MKNK1 and enhance the phosphorylation of eIF4E, contributing to the radio resistance in CRC cells eventually [129]. As a natural compounds, quercetin is recognized for its anti-cancer ability against CRC [130]. It found that quercetin changed the sensitivity of CRC to conventional chemotherapeutic agent 5-FU through tRFs and enhanced the effect of 5-FU in CRC cells. Moreover, quercetin could regulate the expression of specific tRFs and elevate the apoptotic effect of quercetin in tumor cells [131]. And one notable thing is the significant association between intestinal microbiota including Escherichia coli and CRC [132]. Coming from non-pathogenic Escherichia coli, 5’-tRF-Leu (CAA) showed the ability of suppressing CRC cells [133].
The functions of tRFs also reflect in the prediction of prognosis in CRC patients. The level of i-tRF-GlyGCC in CRC tissues was significantly lower than normal tissues, and patients with relatively higher i-tRF-GlyGCC have shorter disease-free survival (DFS) and overall survival (OS) periods, and an increased risk of recurrence [134]. It also identified this influence to adverse prognosis was evident among patients at the TNM stage II. A recent study has found that the tRFdb-3013a and tRFdb-3013b were significantly decreased in colon and rectum adenocarcinomas, especially in patients with lymphatic or vascular invasion present. This reduction of the two tRFs are significantly associated with a worse clinical survival rate and a shortened OS period in CRC patients, showing the potential as novel biomarkers for prognosis in CRC [135].
4.7. Leukemia
Studies have also begun to uncover tRF signatures in leukemia. Previously Veronica Balatti et al. demonstrated that dysregulation of the T-cell leukemia/lymphoma 1 (TCL1) oncogene is a critical contributing event in the pathogenesis of B-cell chronic lymphocytic leukemia (CLL). Furthermore, they found that ts-53 targets the 3′UTR of TCL1, a key oncogene in the development of aggressive CLL and that its down-regulation in leukemic cells is inversely correlated with TCL1 expression, contributing to CLL progression [136]. Together with ts-53, ts-101, ts-46, and ts-47 are also downregulated in CLL [72]. Similarly, another report elucidated that compared with normal B cells, both ts-43 and ts-44 derived from distinct transcripts of pre-tRNAHis, as well as tRF-5 from tRNAHis, are downregulated in CLL [137]. Researches above indicate that dysregulations of tRFs are ubiquitous in CLL and related to tumorigenesis.
Apart from CLL, tRFs also play a vital role in other types of leukemia. Human T-cell leukemia virus type 1 (HTLV-1) is the causative agent of adult T-cell leukemia/lymphoma (ATLL). Reverse transcription (RT) is considered as a key event in the retroviral life cycle. A recent study determined that a 3′18 nt tRNAPro-derived fragment (tRFPro), packaged into HTLV-1 particles, serves as an RT primer in vitro [138]. These findings indicate tRFPro could represent a novel target for therapies aimed at controlling HTLV-1 infection [63]. Roy L. Maute et al. discovered a novel tRFs, CU1276 is abundant in normal germinal center B cells but absent in germinal center-derived lymphomas, suggesting a role in the pathogenesis of this disease. Furthermore, CU1276 suppresses proliferation and modulates the molecular response to DNA damage due to repression of endogenous RPA1, an essential gene involved in many aspects of DNA dynamics [139]. Researchers also found that pseudouridylation (Ψ) of a stem cell-enriched tRF subtype, mini tRFs containing a 5′ terminal oligoguanine (mTOG), selectively inhibits aberrant protein synthesis programs, through disturbing the recruitment of translational co-activator PABPC1-interacting protein 1 (PAIP1), thereby strongly repressing the translation. Dysregulation of mTOG is clinically associated with leukemic transformation and reduced patient survival. These findings define a critical role for tRFs and Ψ in difficult-to-treat subsets of myelodysplastic syndrome (MDS) characterized by high risk of progression to acute myeloid leukemia (AML) [140].
4.8. Lung Cancer
Lung cancer was the most frequently diagnosed cancer in 2022, responsible for almost 2.5 million new cases, or one in eight cancers worldwide (12.4% of all cancers globally) [122]. However, further research is needed on the role of tRFs in lung cancer, especially their underlying molecular mechanisms. Previous study indicated that ts-53, ts-101, ts-46, and ts-47 are not only downregulated in CLL, but also in lung cancer. Overexpression of ts-46 and ts-47 in H1299 and A549 lung cancer cell lines leads to a strong inhibition of colony formation compared with untreated cells, confirming that tsRNAs affect cell growth and survival in lung cancer [136]. Yang Shao et al. reported a novel type of tRNA halves, tRF-Leu-CAG, which is upregulated in NSCLC tumor tissues and NSCLC cell lines, and serum concentration of tRF-Leu-CAG is significantly associated with NSCLC stage progression. They also found that inhibition of tRF-Leu-CAG suppressed cell proliferation and impeded cell cycle in H1299 cells. All these findings reveal the potential therapeutic value of tRF-Leu-CAG [141]. Another tRF, a 28 nt long tRF-5 cleaved at site 1 to 28 on the 5′ end of tRNA-Gly-GCC, AS-tDR-007333 were identified in pre- and post-operation plasma from patients with NSCLC. AS-tDR-007333 is significantly upregulated in NSCLC tissues, plasma, and cells and clinically associated with poorer prognosis. Further study demonstrated that overexpression of AS-tDR-007333 enhanced proliferation and migration of NSCLC cells by activating MED29 through two different pathways. Firstly, interacting with HSPB1, AS-tDR-007333 enhances H3K4me1 and H3K27ac in MED29 promoter, thereby activating MED29 expression. Secondly, AS-tDR-007333 increases the transcription of MED29 promoter by stimulating the expression of transcription factor ELK4 [142]. Fang Hu et al. illustrated that tsRNA-5001a is significantly upregulated in lung adenocarcinoma tissues and positively correlated to the risk of postoperative recurrences and poor prognosis in patients [143]. Recent studies have cast light on diagnostic and therapeutic potential of tRFs in lung cancer. Gu et al. developed a TRY-RNA signature composed of tRFs, rRNA-derived small RNAs, and YRNA-derived small RNAs from human peripheral blood mononuclear cells, which precisely discriminates between healthy control, lung cancer and pulmonary tuberculosis [144]. Baibing Zheng et al. discovered that the expression levels of exosomal tRF-Leu-TAA-005, tRF-Asn-GTT-010, tRF-Ala-AGC-036, tRF-Lys-CTT-049, and tRF-Trp-CCA-057 were significantly downregulated in NSCLC patients. This significant difference suggests that these five exosomal tRFs may be promising diagnostic biomarkers for NSCLC [145].
4.9. Other Cancers
In addition, tRFs have also been reported in other cancers including ovarian cancer, cervical cancer, bladder cancer, and renal cancer.
For ovarian cancer, differentially expressed tRFs were verified in high-grade serous ovarian cancer (HGSOC), involved in the functions of protein phosphorylation, transcription and cell migration and the pathway of cancer, and the MAPK and Wnt signaling pathways. Importantly, tRF-03357 promoted SK-OV-3 cell proliferation, migration and invasion by targeting and downregulating HMBOX1 [146]. Similarly, 3′U-tRFValCAC, derived from pre-tRNAValCAC, also promotes SK-OV-3 cell growth and migration [147]. Another research focused on clinical benefit of tRFs in ovarian cancer and highlighted the significant association of i-tRF-GlyGCC with advanced FIGO stages, suboptimal debulking and most importantly, with early progression and poor overall survival of EOC patients [148].
For cervical cancer, Yang Wang et al. identified a potential tumor suppressor gene tRF-Glu49 in cervical carcinoma, inhibiting cervical cell proliferation, migration and invasion processes by directly regulating the oncogene, fibrinogen-like protein-1 (FGL1) [149]. On the contrary, two 5’-tRFs, 5’tDR-GlyGCC and 5’tDR-GlnCTG, were revealed to enhance tumor progression of cervical cancer by promoting ribosome assembly and preventing cell apoptosis [150]. Further research is needed to reveal the underlying mechanism of tRFs in pathogenesis and progression of cervical cancer.
For bladder cancer, diagnosis of bladder cancer is primarily by invasive tests now, however, more researches are devoting to the exploration of tRFs as screening and diagnostic biomarkers for bladder cancer. A 5’-tRF, 5’-tRF-LysCTT, was associated with aggressive tumor phenotype, early progression and poor survival, and the integration of 5’-tRF-LysCTT with the clinically available markers could achieve superior specificity and improve prediction of disease progression for bladder cancer [151]. The circulating tRF-1:28-chrM.Ser-TGA and tiRNA-1:34-Glu-CTC-1-M2 were specifically expressed by bladder cancer cells and were positively associated with the degree of malignancy, which are not only applied as new biomarkers but also expected to provide novel targets in the therapy of bladder cancer [152]. tRFs have also demonstrated potential in treatment of bladder cancer. A novel m7G-modified tRF, m7G-3’-tiRNA LysTTT (mtiRL), has been defined as a critical molecule to promote bladder cancer malignancy in vitro and in vivo and provided an efficient way to treat bladder cancer by targeting this tRF [153].
To date, studies investigating the role of tRFs in renal carcinoma remains relatively limited, but certain tRFs have been implicated in modulating the tumorigenesis and progression of renal carcer. In clear cell renal cell carcinoma (ccRCC), a study has identified the 5’tRNA4-Val-AAC expression had an inverse correlation with the stage and grade of tumor [154]. And another research found four kinds of 5’-halves were downregulated in the serum and tissue of ccRCC. Their lower expression not only indicated a potential role as tumor suppressors, but alco suggesting the potential of these 5’-halves as novel ccRCC biomarkers [155].

Table 1.
Functional tRFs in different types of cancer.
| Cancer type | tRF name | Role | Function | Ref |
| Breast cancer (BC) | tRF-ArgCCT-017, tRF-Gly-CCC-001, tiRNA-Phe-GAA-003 | upregulate in BC plasma samples | potential biomarker of BC | [73] |
| tRF-19-W4PU732S | oncogenic factor | promote proliferation and malignance of BC and suppress apoptosis by inhibiting RPL27A | [74] | |
| 5′-SHOT-RNA | oncogenic factor | promote proliferation of BC | [75] | |
| ts-112 | oncogenic factor | promote proliferation of BC | [76] | |
| tRF-33 | oncogenic factor | disrupt mitochondrial homeostasis and promote progression of BC by interacting with IGF1 | [77] | |
| tRFGlu, tRFAsp, tRFGly, tRFTyr | tumor suppressor | inhibit metastasis of BC via YBX1 displacement | [78,79] | |
| 5′-tiRNAVal | tumor suppressor | inhibit the FZD3-mediated Wnt/β-Catenin signaling pathway in BC | [80] | |
| tRF3E | tumor suppressor | inhibit proliferation of BC through NCL-mediated mechanism | [81] | |
| tDR-000620 | downregulate in TNBC CSCs and serum sample | potential biomarker of TNBC | [82] | |
| tRFLys-CTT-010 | oncogenic factor | promote proliferation of TNBC by regulating glucose metabolism via tRFLys-CTT-010/G6PC axis | [83] | |
| tDR-0009 and tDR-7336 | drug-resistance driver | facilitate doxorubicin resistance in TNBC | [84] | |
| tRF-27 | oncogenic factordrug-resistance driver | facilitate trastuzumab resistance and promote cell proliferation in BC | [86] | |
| 3’tRF-AlaAGC | drug-resistance driver | enhance the Adriamycin sensitivity in BC via NF-κb signaling pathway by silencing 3’tRF-AlaAGC | [87] | |
| Prostate cancer (PCa) | tRF-1001 | oncogenic factor | promote proliferation of PCa | [23] |
| 5′-SHOT-RNA | oncogenic factor | promote proliferation of PCa | [75] | |
| tRF-315 | drug-resistance driver | facilitate cisplatin resistance in PCa | [92] | |
| 5′TOGs | tumor suppressor | inhibit proliferation of PCa | [93] | |
| Pancreatic cancer (PC) | tRF-Leu-AAG | oncogenic factor | promote proliferation, migration, and invasion of PC by downregulating UPF1 | [94] |
| tRF-19-PNR8YPJZ | oncogenic factor | promote migration and invasion of PC via AXIN2 axis | [95] | |
| tRF-18-8R6546D2 | oncogenic factor | promote malignancy of PC by silencing ASCL2 and regulating MYC and CASP3 | [96] | |
| tRF-19-Q1Q89PJZ | tumor suppressor | inhibit proliferation and metastasis of PC by suppressing HK1 | [97] | |
| expression | [97] | |||
| tRF-GluCTC-0005 | oncogenic factor | promote proliferation, migration and invasion of PDAC and liver metastasis | [98,99] | |
| tRF-21-VBY9PYKHD | tumor suppressor | promote proliferation, migration, and invasion of PDCA when downregulated | [70] | |
| tRF-Pro-CGG | downregulate in advanced PC | potential biomarker of PC | [100] | |
| Liver cancer | LeuCAG3’tsRNA | oncogenic factor | increase viability of HCT-116 cell by interacting with RPS28 and RPS15 | [51] |
| Gly-tRF | oncogenic factor | increase LCSC subpopulation proportion and promote EMT in liver cancer | [101,102] | |
| tRNA-ValTAC-3, tRNA-GlyTCC-5, tRNA-ValAAC-5, tRNA-GluCTC-5 | upregulate in liver cancer | potential biomarker of liver cancer | [104] | |
| tRF5-GlyGCC | radiotherapy inhibitor | reduce NK cell cytotoxicity and limit radiotherapeutic efficacy | [109] | |
| Gastric cancer (GC) | hsa_tsr016141, tRF31-U5YKFN8DYDZDD, tRF-23-Q99P9P9NDD | oncogenic factor | upregulation is associated with tumor grade, lymph node metastasis and invasion | [110,111.112] |
| tRF-3017A | oncogenic factor | promote migration and invasion of GC by targeting NELL2 | [9] | |
| tRF-3019a | oncogenic factor | promote proliferation, migration and invasion of GC | [113] | |
| tRF-23-Q99P9P9NDD | oncogenic factor | promote progression of GC by affecting lipid metabolism and ferroptosis via targeting ACADSB | [114] | |
| tRF-33-P4R8YP9LON4VDP, tRF-193L7L73JD | tumor suppressor | inhibit proliferation of GC by disrupting cell cycle | [115,116] | |
| tRF-5026a | tumor suppressor | inhibit progression of GC via PI3K/AKT signaling pathway and regulate cell cycle | [117] | |
| tRF-Glu-TTC-027, tRF-Val-CAC-016 | tumor suppressor | inhibit malignance of GC via MAPK pathway | [118,119] | |
| tRF-Tyr | tumor suppressor | inhibit progression of GC via c-Myc/Bcl2/Bax pathway | [120] | |
| tRF-Val | oncogenic factor | promote proliferation and invasion and inhibit cell apoptosis of GC | [121] | |
| Colorectal cancer (CRC) | 5’tiRNA-His-GTG | oncogenic factor | promote proliferation and inhibit cell apoptosis of CRC | [122] |
| tRF/miR-1280 | oncogenic factor | promoting proliferation and metastasis of CRC via Notch | [123] | |
| signal pathway | [123] | |||
| 5’tiRNA-Gly-GCC | drug-resistance driver | facilitate 5-FU resistance in CRC | [124] | |
| tRF-20M0NK5Y93 | tumor suppressor | inhibit metastasis of CRC by suppressing Claudin-1 | [125] | |
| tRF-22, tRF-27, tRF-32 | upregulate in CRC tissue and plasma samples | potential biomarker of CRC | [126] | |
| 5’-tRFGlyGCC | upregulate in CRC plasma samples | potential biomarker of CRC | [127] | |
| tRF-Ala-AGC-060 | upregulate in CRC tissue samples | potential biomarker of CRC | [128] | |
| tRFTyr-GTA-081 | downregulate in CRC tissue samples | potential biomarker of CRC | [128] | |
| tRF-16-7X9PN5D | radiosensitizer | promote proliferation, migration, invasion and radio resistance when downregulated | [129] | |
| 5’-tRF-Leu (CAA) | tumor suppressor | inhibit progression of CRC | [133] | |
| i-tRF-GlyGCC | downregulate in CRC tissue samples | potential biomarker of CRC | [134] | |
| tRFdb-3013a, tRFdb-3013b | downregulate in colon and rectum adenocarcinomas | potential biomarker of CRC | [135] | |
| Leukemia | ts-53, ts-101, ts-46, and ts-47, ts-43, ts-44 | tumor suppressor | promote progression of CLL | [72,136,137] |
| tRFPro | RT primer of HTLV-1 | potential target for preventing ATLL | [138] | |
| CU1276 | tumor suppressor | inhibit proliferation and molecularly modulate DNA damage repair | [139] | |
| mTOG | dysregulation associated with leukemic transformation | potential biomarker for MDS transformation to AML | [140] | |
| Lung cancer | ts-46, ts-47 | tumor suppressor | inhibit proliferation of lung cancer cell lines when upregulated | [136] |
| tRF-Leu-CAG | oncogenic factorupregulated in NSCLC tissues and cell lines | inhibit proliferation and impede cell cycle of NSCLCpotential biomarker of NSCLC | [141] | |
| AS-tDR-007333 | oncogenic factorupregulated in NSCLC tissues and plasma | promote proliferation and migration of NSCLCpotential biomarker of NSCLC | [142] | |
| tsRNA-5001a | upregulated in lung adenocarcinoma tissues | potential biomarker for poor prognosis | [143] | |
| tRF-Leu-TAA-005, tRF-Asn-GTT-010, tRF-Ala-AGC-036, tRF-Lys-CTT-049, and tRF-Trp-CCA-057 | downregulated in NSCLC tissues | potential biomarker of NSCLC | [145] | |
| ovarian cancer | tRF-03357, 3′U-tRFValCAC | oncogenic factor | promote proliferation, migration and invasion of SK-OV-3 cell | [146,147] |
| i-tRF-GlyGCC | upregulation is associated with advanced FIGO stages | potential biomarker of ovarian cancer | [148] | |
| cervical cancer | tRF-Glu49 | tumor suppressor | inhibit proliferation of cervical carcinoma by regulating FGL1 | [149] |
| 5’tDR-GlyGCC, 5’tDR-GlnCTG | oncogenic factor | promote progression of cervical cancer | [150] | |
| bladder cancer | 5’-tRF-LysCTT | upregulation is associated with poor prognosis | potential biomarker of bladder cancer | [151] |
| tRF-1:28-chrM.Ser-TGA, tiRNA-1:34-Glu-CTC-1-M2 | oncogenic factor | promote malignancy of bladder cancer | [152] | |
| m7G-3’-tiRNA LysTTT (mtiRL) | oncogenic factor | promote malignancy of bladder cancer | [153] | |
| renal carcinoma | 5’tRNA4-Val-AAC | downregulation is associated with advanced stage | potential biomarker of ccRCC | [154] |
| 5’-tRNA-Arg-CCT, 5’-tRNA-Glu-CTC, 5’-tRNA-Leu-CAG and 5’-tRNA-Lys-TTT | downregulated in tumor tissues and serum samples | potential biomarker of ccRCC | [155] |
5. Conclusions and Perspective
With the application of novel technologies such as high-throughput transcriptome sequencing, the researches of tRFs are growing tremendously, while their diversity and functional roles have also been progressively elucidated. Numerous tRFs have been collated and collected into several databases, facilitating further discovery and functional researches. Related to the situation of cells, the production of tRFs may be influenced by the stress conditions. The tRFs can be primarily divided into tRNA halves and tRFs, and other special fragments such as i-tRFs, tRF-1s. Meanwhile, various modifications of tRNAs regulate tRF biogenesis, and influence the stability and function of tRFs. We review the key roles of tRFs including nascent RNA silencing, translation regulation, epigenetic gene silencing, and reverse transcription inhibition.
Increasing evidence has manifested that tRFs are associated with many aspects of various kinds of cancer, which are involved in the generation, procession and migration of tumors. Here, we summarized the related tRFs relevant with several cancer types such as breast cancer, gastric cancer, liver cancer, colorectal cancer and pancreatic cancer, prostatic cancer and so on. Whereas, the underlying mechanisms of how tRFs regulate tumors are still understood incompletely. Meanwhile, the biological function of the same tRFs seem different in diverse types of cancer. Furthermore, the characteristics of distinct types of cancers, including growth patterns, metastatic behaviors and tumor microenvironment heterogeneity, drive the production of different tRFs that exert varied functional roles [156]. For example, the hypoxia-induced tRFs play important roles in BC [31]. And according to the different expression levels in related cancer, some specific tRFs can be applied as as novel non-invasive biomarkers for the diagnosis and detection of cancer, and therapeutic targets for clinical application.
There are still some limitations in the research of tRF. The nomenclature of tRF is debated, leading to the inconsistency in publications and confusion of readers. Some tRFs are named by the order of discovery, and this difference may hinder the searching and publication [72,157]. And although several tRFs dysregulated in cancer have been characterized, their underlying mechanisms of action require further investigation to verify, particularly the determination of causal relationships. There is a long process to utilize tRF in clinic as biomarkers for cancer. Although some researches emphasized the potential of tRF for cancer diagnosis, the detection of tRFs mainly relies on high throughput sequencing and other innovative methods, which are relative expensive or complicated for large scale detection clinically. So more attempts are compulsory for developing appropriate tRF testing methods.
tRFs and their functional roles are being progressively elucidated. Accumulating evidence from series studies has demonstrated significant associations between tRF dysregulation and cancer progression. Further investigations remain imperative to comprehensively identify the mechanistic contributions of tRFs in oncogenesis and to utilize as clinically actionable biomarkers and innovative therapeutic targets for cancer diagnosis and prognosis.
References
- Yu M, Lu B, Zhang J, Ding J, Liu P, Lu Y. tRNA-derived RNA fragments in cancer: current status and future perspectives. J Hematol Oncol. 2020 Sep 4;13(1):121. [CrossRef]
- Zhang Z, Zhang J, Diao L, Han L. Small non-coding RNAs in human cancer: function, clinical utility, and characterization. Oncogene. 2021 Mar;40(9):1570-1577. [CrossRef]
- Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018 Jan;18(1):5-18.
- Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014 Mar 27;157(1):77-94. [CrossRef]
- Kumar P, Anaya J, Mudunuri SB, Dutta A. Meta-analysis of tRNA derived RNA fragments reveals that they are evolutionarily conserved and associate with AGO proteins to recognize specific RNA targets. BMC Biol. 2014 Oct 1;12:78.
- Shen Y, Yu X, Zhu L, Li T, Yan Z, Guo J. Transfer RNA-derived fragments and tRNA halves: biogenesis, biological functions and their roles in diseases. J Mol Med (Berl). 2018 Nov;96(11):1167-1176. [CrossRef]
- Kumar P, Kuscu C, Dutta A. Biogenesis and Function of Transfer RNA-Related Fragments (tRFs). Trends Biochem Sci. 2016 Aug;41(8):679-689. [CrossRef]
- Pan Y, Ying X, Zhang X, Jiang H, Yan J, Duan S. The role of tRNA-Derived small RNAs (tsRNAs) in pancreatic cancer and acute pancreatitis. Noncoding RNA Res. 2024 Dec 30;11:200-208. [CrossRef]
- Tong L, Zhang W, Qu B, Zhang F, Wu Z, Shi J, Chen X, Song Y, Wang Z. The tRNA-Derived Fragment-3017A Promotes Metastasis by Inhibiting NELL2 in Human Gastric Cancer. Front Oncol. 2021 Feb 16;10:570916. [CrossRef]
- Qiu P, Jiang Q, Song H. Unveiling the hidden world of transfer RNA-derived small RNAs in inflammation. J Inflamm (Lond). 2024 Nov 12;21(1):46. [CrossRef]
- Weng Q, Wang Y, Xie Y, Yu X, Zhang S, Ge J, Li Z, Ye G, Guo J. Extracellular vesicles-associated tRNA-derived fragments (tRFs): biogenesis, biological functions, and their role as potential biomarkers in human diseases. J Mol Med (Berl). 2022 May;100(5):679-695. [CrossRef]
- Garcia-Silva MR, Cabrera-Cabrera F, das Neves RF, Souto-Padrón T, de Souza W, Cayota A. Gene expression changes induced by Trypanosoma cruzi shed microvesicles in mammalian host cells: relevance of tRNA-derived halves. Biomed Res Int. 2014;2014:305239. [CrossRef]
- Deng J, Ptashkin RN, Chen Y, Cheng Z, Liu G, Phan T, Deng X, Zhou J, Lee I, Lee YS, Bao X. Respiratory Syncytial Virus Utilizes a tRNA Fragment to Suppress Antiviral Responses Through a Novel Targeting Mechanism. Mol Ther. 2015 Oct;23(10):1622-9. [CrossRef]
- Li Z, Ender C, Meister G, Moore PS, Chang Y, John B. Extensive terminal and asymmetric processing of small RNAs from rRNAs, snoRNAs, snRNAs, and tRNAs. Nucleic Acids Res. 2012 Aug;40(14):6787-99. [CrossRef]
- Schaffer AE, Eggens VR, Caglayan AO, Reuter MS, Scott E, Coufal NG, Silhavy JL, Xue Y, Kayserili H, Yasuno K, Rosti RO, Abdellateef M, Caglar C, Kasher PR, Cazemier JL, Weterman MA, Cantagrel V, Cai N, Zweier C, Altunoglu U, Satkin NB, Aktar F, Tuysuz B, Yalcinkaya C, Caksen H, Bilguvar K, Fu XD, Trotta CR, Gabriel S, Reis A, Gunel M, Baas F, Gleeson JG. CLP1 founder mutation links tRNA splicing and maturation to cerebellar development and neurodegeneration. Cell. 2014 Apr 24;157(3):651-63. [CrossRef]
- Tian H, Hu Z, Wang C. The Therapeutic Potential of tRNA-derived Small RNAs in Neurodegenerative Disorders. Aging Dis. 2022 Apr 1;13(2):389-401. [CrossRef]
- Phan HD, Lai LB, Zahurancik WJ, Gopalan V. The many faces of RNA-based RNase P, an RNA-world relic. Trends Biochem Sci. 2021 Dec;46(12):976-991. [CrossRef]
- Maraia RJ, Lamichhane TN. 3’ processing of eukaryotic precursor tRNAs. Wiley Interdiscip Rev RNA. 2011 May-Jun;2(3):362-75.
- Xiong Y, Steitz TA. A story with a good ending: tRNA 3’-end maturation by CCA-adding enzymes. Curr Opin Struct Biol. 2006 Feb;16(1):12-7. [CrossRef]
- Holley RW, Apgar J, Everett GA, Madison JT, Marquisee M, Merrill SH, et al. STRUCTURE OF A RIBONUCLEIC ACID. Science. 1965 Mar 19;147(3664):1462-5.
- Suzuki T. The expanding world of tRNA modifications and their disease relevance. Nat Rev Mol Cell Biol. 2021 Jun;22(6):375-392. [CrossRef]
- Yang M, Mo Y, Ren D, Liu S, Zeng Z, Xiong W. Transfer RNA-derived small RNAs in tumor microenvironment. Mol Cancer. 2023 Feb 16;22(1):32. [CrossRef]
- Lee YS, Shibata Y, Malhotra A, Dutta A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009 Nov 15;23(22):2639-49. [CrossRef]
- Fu BF, Xu CY. Transfer RNA-Derived Small RNAs: Novel Regulators and Biomarkers of Cancers. Front Oncol. 2022 Apr 28;12:843598. [CrossRef]
- Kumar P, Mudunuri SB, Anaya J, Dutta A. tRFdb: a database for transfer RNA fragments. Nucleic Acids Res. 2015 Jan;43(Database issue):D141-5. [CrossRef]
- Tao EW, Cheng WY, Li WL, Yu J, Gao QY. tiRNAs: A novel class of small noncoding RNAs that helps cells respond to stressors and plays roles in cancer progression. J Cell Physiol. 2020 Feb;235(2):683-690. [CrossRef]
- Lu J, Zhu P, Zhang X, Zeng L, Xu B, Zhou P. tRNA-derived fragments: Unveiling new roles and molecular mechanisms in cancer progression. Int J Cancer. 2024 Oct 15;155(8):1347-1360. [CrossRef]
- Xie Y, Yao L, Yu X, Ruan Y, Li Z, Guo J. Action mechanisms and research methods of tRNA-derived small RNAs. Signal Transduct Target Ther. 2020 Jun 30;5(1):109. [CrossRef]
- Cole C, Sobala A, Lu C, Thatcher SR, Bowman A, Brown JW, Green PJ, Barton GJ, Hutvagner G. Filtering of deep sequencing data reveals the existence of abundant Dicer-dependent small RNAs derived from tRNAs. RNA. 2009 Dec;15(12):2147-60.
- Telonis AG, Loher P, Magee R, Pliatsika V, Londin E, Kirino Y, Rigoutsos I. tRNA Fragments Show Intertwining with mRNAs of Specific Repeat Content and Have Links to Disparities. Cancer Res. 2019 Jun 15;79(12):3034-3049.
- Goodarzi H, Liu X, Nguyen HC, Zhang S, Fish L, Tavazoie SF. Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement. Cell. 2015 May 7;161(4):790-802. [CrossRef]
- Yamasaki S, Ivanov P, Hu GF, Anderson P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J Cell Biol. 2009 Apr 6;185(1):35-42. [CrossRef]
- Thompson DM, Parker R. Stressing out over tRNA cleavage. Cell. 2009 Jul 23;138(2):215-9. [CrossRef]
- Liu S, Chen Y, Ren Y, Zhou J, Ren J, Lee I, Bao X. A tRNA-derived RNA Fragment Plays an Important Role in the Mechanism of Arsenite -induced Cellular Responses. Sci Rep. 2018 Nov 15;8(1):16838. [CrossRef]
- Tao EW, Wang HL, Cheng WY, Liu QQ, Chen YX, Gao QY. A specific tRNA half, 5’tiRNA-His-GTG, responds to hypoxia via the HIF1α/ANG axis and promotes colorectal cancer progression by regulating LATS2. J Exp Clin Cancer Res. 2021 Feb 15;40(1):67.
- Su Z, Kuscu C, Malik A, Shibata E, Dutta A. Angiogenin generates specific stress-induced tRNA halves and is not involved in tRF-3-mediated gene silencing. J Biol Chem. 2019 Nov 8;294(45):16930-16941. [CrossRef]
- Blanco S, Dietmann S, Flores JV, Hussain S, Kutter C, Humphreys P, et al. Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders. EMBO J. 2014 Sep 17;33(18):2020-39. [CrossRef]
- Haussecker D, Huang Y, Lau A, Parameswaran P, Fire AZ, Kay MA. Human tRNA-derived small RNAs in the global regulation of RNA silencing. RNA. 2010 Apr;16(4):673-95. [CrossRef]
- Kim HK, Yeom JH, Kay MA. Transfer RNA-Derived Small RNAs: Another Layer of Gene Regulation and Novel Targets for Disease Therapeutics. Mol Ther. 2020 Nov 4;28(11):2340-2357. [CrossRef]
- Sobala A, Hutvagner G. Transfer RNA-derived fragments: origins, processing, and functions. Wiley Interdiscip Rev RNA. 2011 Nov-Dec;2(6):853-62.
- Green JA, Ansari MY, Ball HC, Haqqi TM. tRNA-derived fragments (tRFs) regulate post-transcriptional gene expression via AGO-dependent mechanism in IL-1β stimulated chondrocytes. Osteoarthritis Cartilage. 2020 Aug;28(8):1102-1110. [CrossRef]
- Di Fazio A, Schlackow M, Pong SK, Alagia A, Gullerova M. Dicer dependent tRNA derived small RNAs promote nascent RNA silencing. Nucleic Acids Res. 2022 Feb 22;50(3):1734-1752. [CrossRef]
- Cho H, Lee W, Kim GW, Lee SH, Moon JS, Kim M, Kim HS, Oh JW. Regulation of La/SSB-dependent viral gene expression by pre-tRNA 3’ trailer-derived tRNA fragments. Nucleic Acids Res. 2019 Oct 10;47(18):9888-9901. [CrossRef]
- Ivanov P, Emara MM, Villen J, Gygi SP, Anderson P. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol Cell. 2011 Aug 19;43(4):613-23. [CrossRef]
- Lyons SM, Gudanis D, Coyne SM, Gdaniec Z, Ivanov P. Identification of functional tetramolecular RNA G-quadruplexes derived from transfer RNAs. Nat Commun. 2017 Oct 24;8(1):1127. Erratum in: Nat Commun. 2017 Dec 5;8(1):2020. [CrossRef]
- Ivanov P, O’Day E, Emara MM, Wagner G, Lieberman J, Anderson P. G-quadruplex structures contribute to the neuroprotective effects of angiogenin-induced tRNA fragments. Proc Natl Acad Sci U S A. 2014 Dec 23;111(51):18201-6. [CrossRef]
- Lyons SM, Kharel P, Akiyama Y, Ojha S, Dave D, Tsvetkov V, Merrick W, Ivanov P, Anderson P. eIF4G has intrinsic G-quadruplex binding activity that is required for tiRNA function. Nucleic Acids Res. 2020 Jun 19;48(11):6223-6233. [CrossRef]
- Emara MM, Ivanov P, Hickman T, Dawra N, Tisdale S, Kedersha N, Hu GF, Anderson P. Angiogenin-induced tRNA-derived stress-induced RNAs promote stress-induced stress granule assembly. J Biol Chem. 2010 Apr 2;285(14):10959-68. [CrossRef]
- Guzzi N, Cieśla M, Ngoc PCT, Lang S, Arora S, Dimitriou M, et al. Pseudouridylation of tRNA-Derived Fragments Steers Translational Control in Stem Cells. Cell. 2018 May 17;173(5):1204-1216.e26. [CrossRef]
- Sobala A, Hutvagner G. Small RNAs derived from the 5’ end of tRNA can inhibit protein translation in human cells. RNA Biol. 2013 Apr;10(4):553-63. [CrossRef]
- Kim HK, Fuchs G, Wang S, Wei W, Zhang Y, Park H, Roy-Chaudhuri B, Li P, Xu J, Chu K, Zhang F, Chua MS, So S, Zhang QC, Sarnow P, Kay MA. A transfer-RNA-derived small RNA regulates ribosome biogenesis. Nature. 2017 Dec 7;552(7683):57-62. [CrossRef]
- Henikoff S, Greally JM. Epigenetics, cellular memory and gene regulation. Curr Biol. 2016 Jul 25;26(14):R644-8. [CrossRef]
- Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012 Jul 6;150(1):12-27.
- Yu T, Huang X, Dou S, Tang X, Luo S, Theurkauf WE, Lu J, Weng Z. A benchmark and an algorithm for detecting germline transposon insertions and measuring de novo transposon insertion frequencies. Nucleic Acids Res. 2021 May 7;49(8):e44. [CrossRef]
- Slotkin RK, Martienssen R. Transposable elements and the epigenetic regulation of the genome. Nat Rev Genet. 2007 Apr;8(4):272-85. [CrossRef]
- Watanabe T, Tomizawa S, Mitsuya K, Totoki Y, Yamamoto Y, Kuramochi-Miyagawa S, Iida N, Hoki Y, Murphy PJ, Toyoda A, Gotoh K, Hiura H, Arima T, Fujiyama A, Sado T, Shibata T, Nakano T, Lin H, Ichiyanagi K, Soloway PD, Sasaki H. Role for piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science. 2011 May 13;332(6031):848-52. [CrossRef]
- Fields BD, Kennedy S. Chromatin Compaction by Small RNAs and the Nuclear RNAi Machinery in C. elegans. Sci Rep. 2019 Jun 21;9(1):9030. [CrossRef]
- Couvillion MT, Bounova G, Purdom E, Speed TP, Collins K. A Tetrahymena Piwi bound to mature tRNA 3’ fragments activates the exonuclease Xrn2 for RNA processing in the nucleus. Mol Cell. 2012 Nov 30;48(4):509-20. [CrossRef]
- Zhang X, He X, Liu C, Liu J, Hu Q, Pan T, Duan X, Liu B, Zhang Y, Chen J, Ma X, Zhang X, Luo H, Zhang H. IL-4 Inhibits the Biogenesis of an Epigenetically Suppressive PIWI-Interacting RNA To Upregulate CD1a Molecules on Monocytes/Dendritic Cells. J Immunol. 2016 Feb 15;196(4):1591-603. [CrossRef]
- Zhang B, Pan Y, Li Z, Hu K. tRNA-derived small RNAs: their role in the mechanisms, biomarkers, and therapeutic strategies of colorectal cancer. J Transl Med. 2025 Jan 13;23(1):51. [CrossRef]
- Schorn AJ, Gutbrod MJ, LeBlanc C, Martienssen R. LTR-Retrotransposon Control by tRNA-Derived Small RNAs. Cell. 2017 Jun 29;170(1):61-71.e11. [CrossRef]
- Manivannan AC, Devaraju V, Velmurugan P, Sathiamoorthi T, Sivakumar S, Subbiah SK, Ravi AV. Tumorigenesis and diagnostic practice applied in two oncogenic viruses: Epstein Barr virus and T-cell lymphotropic virus-1-Mini review. Biomed Pharmacother. 2021 Oct;142:111974. [CrossRef]
- Ruggero K, Guffanti A, Corradin A, Sharma VK, De Bellis G, Corti G, Grassi A, Zanovello P, Bronte V, Ciminale V, D’Agostino DM. Small noncoding RNAs in cells transformed by human T-cell leukemia virus type 1: a role for a tRNA fragment as a primer for reverse transcriptase. J Virol. 2014 Apr;88(7):3612-22. [CrossRef]
- Seiki M, Hattori S, Yoshida M. Human adult T-cell leukemia virus: molecular cloning of the provirus DNA and the unique terminal structure. Proc Natl Acad Sci U S A. 1982 Nov;79(22):6899-902. [CrossRef]
- Mei Y, Yong J, Liu H, Shi Y, Meinkoth J, Dreyfuss G, Yang X. tRNA binds to cytochrome c and inhibits caspase activation. Mol Cell. 2010 Mar 12;37(5):668-78. [CrossRef]
- Mei Y, Stonestrom A, Hou YM, Yang X. Apoptotic regulation and tRNA. Protein Cell. 2010 Sep;1(9):795-801. [CrossRef]
- Hou YM, Yang X. Regulation of cell death by transfer RNA. Antioxid Redox Signal. 2013 Aug 20;19(6):583-94. [CrossRef]
- Saikia M, Jobava R, Parisien M, Putnam A, Krokowski D, Gao XH, Guan BJ, Yuan Y, Jankowsky E, Feng Z, Hu GF, Pusztai-Carey M, Gorla M, Sepuri NB, Pan T, Hatzoglou M. Angiogenin-cleaved tRNA halves interact with cytochrome c, protecting cells from apoptosis during osmotic stress. Mol Cell Biol. 2014 Jul;34(13):2450-63.
- Lu S, Wei X, Tao L, Dong D, Hu W, Zhang Q, Tao Y, Yu C, Sun D, Cheng H. A novel tRNA-derived fragment tRF-3022b modulates cell apoptosis and M2 macrophage polarization via binding to cytokines in colorectal cancer. J Hematol Oncol. 2022 Dec 16;15(1):176. [CrossRef]
- Pan L, Huang X, Liu ZX, Ye Y, Li R, Zhang J, Wu G, Bai R, Zhuang L, Wei L, Li M, Zheng Y, Su J, Deng J, Deng S, Zeng L, Zhang S, Wu C, Che X, Wang C, Chen R, Lin D, Zheng J. Inflammatory cytokine-regulated tRNA-derived fragment tRF-21 suppresses pancreatic ductal adenocarcinoma progression. J Clin Invest. 2021 Nov 15;131(22):e148130. [CrossRef]
- [bz]Pekarsky Y, Balatti V, Palamarchuk A, Rizzotto L, Veneziano D, Nigita G, Rassenti LZ, Pass HI, Kipps TJ, Liu CG, Croce CM. Dysregulation of a family of short noncoding RNAs, tsRNAs, in human cancer. Proc Natl Acad Sci U S A. 2016 May 3;113(18):5071-6. [CrossRef]
- Balatti V, Nigita G, Veneziano D, Drusco A, Stein GS, Messier TL, et al. tsRNA signatures in cancer. Proc Natl Acad Sci U S A. 2017 Jul 25;114(30):8071-8076.
- Wang J, Ma G, Ge H, Han X, Mao X, Wang X, Veeramootoo JS, Xia T, Liu X, Wang S. Circulating tRNA-derived small RNAs (tsRNAs) signature for the diagnosis and prognosis of breast cancer. NPJ Breast Cancer. 2021 Jan 5;7(1):4. [CrossRef]
- Zhang Z, Liu Z, Zhao W, Zhao X, Tao Y. tRF-19-W4PU732S promotes breast cancer cell malignant activity by targeting inhibition of RPL27A (ribosomal protein-L27A). Bioengineered. 2022 Feb;13(2):2087-2098. [CrossRef]
- Honda S, Loher P, Shigematsu M, Palazzo JP, Suzuki R, Imoto I, Rigoutsos I, Kirino Y. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc Natl Acad Sci U S A. 2015 Jul 21;112(29):E3816-25. [CrossRef]
- Farina NH, Scalia S, Adams CE, Hong D, Fritz AJ, Messier TL, Balatti V, Veneziano D, Lian JB, Croce CM, Stein GS, Stein JL. Identification of tRNA-derived small RNA (tsRNA) responsive to the tumor suppressor, RUNX1, in breast cancer. J Cell Physiol. 2020 Jun;235(6):5318-5327.
- Lou Y, Fu B, Liu L, Song J, Zhu M, Xu C. The tRF-33/IGF1 axis dysregulates mitochondrial homeostasis in HER2-negative breast cancer. Am J Physiol Cell Physiol. 2025 Feb 1;328(2):C627-C638. [CrossRef]
- Jürchott K, Kuban RJ, Krech T, Blüthgen N, Stein U, Walther W, Friese C, Kiełbasa SM, Ungethüm U, Lund P, et al. Identification of Y-box binding protein 1 as a core regulator of MEK/ERK pathway-dependent gene signatures in colorectal cancer cells. PLoS Genet. 2010 Dec 2;6(12):e1001231. [CrossRef]
- Uchiumi T, Fotovati A, Sasaguri T, Shibahara K, Shimada T, Fukuda T, Nakamura T, Izumi H, Tsuzuki T, Kuwano M, Kohno K. YB-1 is important for an early stage embryonic development: neural tube formation and cell proliferation. J Biol Chem. 2006 Dec 29;281(52):40440-9.
- Mo D, Jiang P, Yang Y, Mao X, Tan X, Tang X, Wei D, Li B, Wang X, Tang L, Yan F. A tRNA fragment, 5’-tiRNAVal, suppresses the Wnt/β-catenin signaling pathway by targeting FZD3 in breast cancer. Cancer Lett. 2019 Aug 10;457:60-73.
- Falconi M, Giangrossi M, Zabaleta ME, Wang J, Gambini V, Tilio M, Bencardino D, Occhipinti S, Belletti B, Laudadio E, Galeazzi R, Marchini C, Amici A. A novel 3’-tRNAGlu-derived fragment acts as a tumor suppressor in breast cancer by targeting nucleolin. FASEB J. 2019 Dec;33(12):13228-13240.
- Feng W, Li Y, Chu J, Li J, Zhang Y, Ding X, Fu Z, Li W, Huang X, Yin Y. Identification of tRNA-derived small noncoding RNAs as potential biomarkers for prediction of recurrence in triple-negative breast cancer. Cancer Med. 2018 Oct;7(10):5130-5144. [CrossRef]
- Zhu P, Lu J, Zhi X, Zhou Y, Wang X, Wang C, Gao Y, Zhang X, Yu J, Sun Y, Zhou P. tRNA-derived fragment tRFLys-CTT-010 promotes triple-negative breast cancer progression by regulating glucose metabolism via G6PC. Carcinogenesis. 2021 Oct 5;42(9):1196-1207. Erratum in: Carcinogenesis. 2022 Sep 19;43(8):813. [CrossRef]
- Cui Y, Huang Y, Wu X, Zheng M, Xia Y, Fu Z, Ge H, Wang S, Xie H. Hypoxia-induced tRNA-derived fragments, novel regulatory factor for doxorubicin resistance in triple-negative breast cancer. J Cell Physiol. 2019 Jun;234(6):8740-8751.
- Clynes RA, Towers TL, Presta LG, Ravetch JV. Inhibitory Fc receptors modulate in vivo cytotoxicity against tumor targets. Nat Med. 2000 Apr;6(4):443-6. [CrossRef]
- He Y, Liu Y, Gong J, Yang F, Sun C, Yan X, Duan N, Hua Y, Zeng T, Fu Z, Liang Y, Li W, Huang X, Tang J, Yin Y. tRF-27 competitively Binds to G3BPs and Activates MTORC1 to Enhance HER2 Positive Breast Cancer Trastuzumab Tolerance. Int J Biol Sci. 2024 Jul 15;20(10):3923-3941. [CrossRef]
- Mo D, Tang X, Ma Y, Chen D, Xu W, Jiang N, Zheng J, Yan F. tRNA-derived fragment 3’tRF-AlaAGC modulates cell chemoresistance and M2 macrophage polarization via binding to TRADD in breast cancer. J Transl Med. 2024 Jul 30;22(1):706.
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011 Mar-Apr;61(2):69-90. [CrossRef]
- Martens-Uzunova ES, Jalava SE, Dits NF, van Leenders GJ, Møller S, Trapman J, Bangma CH, Litman T, Visakorpi T, Jenster G. Diagnostic and prognostic signatures from the small non-coding RNA transcriptome in prostate cancer. Oncogene. 2012 Feb 23;31(8):978-91. [CrossRef]
- Olvedy M, Scaravilli M, Hoogstrate Y, Visakorpi T, Jenster G, Martens-Uzunova ES. A comprehensive repertoire of tRNA-derived fragments in prostate cancer. Oncotarget. 2016 Apr 26;7(17):24766-77. [CrossRef]
- Magee RG, Telonis AG, Loher P, Londin E, Rigoutsos I. Profiles of miRNA Isoforms and tRNA Fragments in Prostate Cancer. Sci Rep. 2018 Mar 28;8(1):5314. [CrossRef]
- Yang C, Lee M, Song G, Lim W. tRNALys-Derived Fragment Alleviates Cisplatin-Induced Apoptosis in Prostate Cancer Cells. Pharmaceutics. 2021 Jan 4;13(1):55. [CrossRef]
- García-Vílchez R, Añazco-Guenkova AM, Dietmann S, López J, Morón-Calvente V, D’Ambrosi S, Nombela P, Zamacola K, Mendizabal I, García-Longarte S, Zabala-Letona A, Astobiza I, Fernández S, Paniagua A, Miguel-López B, Marchand V, Alonso-López D, Merkel A, García-Tuñón I, Ugalde-Olano A, Loizaga-Iriarte A, Lacasa-Viscasillas I, Unda M, Azkargorta M, Elortza F, Bárcena L, Gonzalez-Lopez M, Aransay AM, Di Domenico T, Sánchez-Martín MA, De Las Rivas J, Guil S, Motorin Y, Helm M, Pandolfi PP, Carracedo A, Blanco S. METTL1 promotes tumorigenesis through tRNA-derived fragment biogenesis in prostate cancer. Mol Cancer. 2023 Jul 29;22(1):119. [CrossRef]
- Sui S, Wang Z, Cui X, Jin L, Zhu C. The biological behavior of tRNA-derived fragment tRF-Leu-AAG in pancreatic cancer cells. Bioengineered. 2022 Apr;13(4):10617-10628. [CrossRef]
- Cao W, Dai S, Ruan W, Long T, Zeng Z, Lei S. Pancreatic stellate cell-derived exosomal tRF-19-PNR8YPJZ promotes proliferation and mobility of pancreatic cancer through AXIN2. J Cell Mol Med. 2023 Sep;27(17):2533-2546. [CrossRef]
- Lan S, Liu S, Wang K, Chen W, Zheng D, Zhuang Y, Zhang S. tRNA-derived RNA fragment, tRF-18-8R6546D2, promotes pancreatic adenocarcinoma progression by directly targeting ASCL2. Gene. 2024 Nov 15;927:148739. [CrossRef]
- Cao W, Zeng Z, Lei S. 5’-tRF-19-Q1Q89PJZ Suppresses the Proliferation and Metastasis of Pancreatic Cancer Cells via Regulating Hexokinase 1-Mediated Glycolysis. Biomolecules. 2023 Oct 12;13(10):1513. [CrossRef]
- Li H, Liu X, Jiang S, Zhou X, Yao L, Di Y, Jiang Y, Gu J, Mao Y, Li J, Jin C, Yang P, Fu D. WD repeat-containing protein 1 maintains β-Catenin activity to promote pancreatic cancer aggressiveness. Br J Cancer. 2020 Sep;123(6):1012-1023. [CrossRef]
- Chen W, Peng W, Wang R, Bai S, Cao M, Xiong S, Li Y, Yang Y, Liang J, Liu L, Yazdani HO, Zhao Y, Cheng B. Exosome-derived tRNA fragments tRF-GluCTC-0005 promotes pancreatic cancer liver metastasis by activating hepatic stellate cells. Cell Death Dis. 2024 Jan 30;15(1):102. [CrossRef]
- Li J, Jin L, Gao Y, Gao P, Ma L, Zhu B, Yin X, Sui S, Chen S, Jiang Z, Zhu C. Low expression of tRF-Pro-CGG predicts poor prognosis in pancreatic ductal adenocarcinoma. J Clin Lab Anal. 2021 May;35(5):e23742. [CrossRef]
- Zhou Y, Hu J, Liu L, Yan M, Zhang Q, Song X, Lin Y, Zhu D, Wei Y, Fu Z, Hu L, Chen Y, Li X. Gly-tRF enhances LCSC-like properties and promotes HCC cells migration by targeting NDFIP2. Cancer Cell Int. 2021 Sep 18;21(1):502. [CrossRef]
- Liu D, Wu C, Wang J, Zhang L, Sun Z, Chen S, Ding Y, Wang W. Transfer RNA-derived fragment 5’tRF-Gly promotes the development of hepatocellular carcinoma by direct targeting of carcinoembryonic antigen-related cell adhesion molecule 1. Cancer Sci. 2022 Oct;113(10):3476-3488. [CrossRef]
- Zuo Y, Chen S, Yan L, Hu L, Bowler S, Zitello E, Huang G, Deng Y. Development of a tRNA-derived small RNA diagnostic and prognostic signature in liver cancer. Genes Dis. 2021 Jan 28;9(2):393-400. [CrossRef]
- Zhu L, Li J, Gong Y, Wu Q, Tan S, Sun D, Xu X, Zuo Y, Zhao Y, Wei YQ, Wei XW, Peng Y. Exosomal tRNA-derived small RNA as a promising biomarker for cancer diagnosis. Mol Cancer. 2019 Apr 2;18(1):74. [CrossRef]
- Herrera FG, Ronet C, Ochoa de Olza M, Barras D, Crespo I, Andreatta M, Corria-Osorio J, Spill A, Benedetti F, Genolet R, Orcurto A, Imbimbo M, Ghisoni E, Navarro Rodrigo B, Berthold DR, Sarivalasis A, Zaman K, Duran R, Dromain C, Prior J, Schaefer N, Bourhis J, Dimopoulou G, Tsourti Z, Messemaker M, Smith T, Warren SE, Foukas P, Rusakiewicz S, Pittet MJ, Zimmermann S, Sempoux C, Dafni U, Harari A, Kandalaft LE, Carmona SJ, Dangaj Laniti D, Irving M, Coukos G. Low-Dose Radiotherapy Reverses Tumor Immune Desertification and Resistance to Immunotherapy. Cancer Discov. 2022 Jan;12(1):108-133. [CrossRef]
- Weiss T, Schneider H, Silginer M, Steinle A, Pruschy M, Polić B, Weller M, Roth P. NKG2D-Dependent Antitumor Effects of Chemotherapy and Radiotherapy against Glioblastoma. Clin Cancer Res. 2018 Feb 15;24(4):882-895.
- Donlon NE, Power R, Hayes C, Reynolds JV, Lysaght J. Radiotherapy, immunotherapy, and the tumour microenvironment: Turning an immunosuppressive milieu into a therapeutic opportunity. Cancer Lett. 2021 Apr 1;502:84-96. [CrossRef]
- Ji J, Ding K, Cheng B, Zhang X, Luo T, Huang B, Yu H, Chen Y, Xu X, Lin H, Zhou J, Wang T, Jin M, Liu A, Yan D, Liu F, Wang C, Chen J, Yan F, Wang L, Zhang J, Yan S, Wang J, Li X, Chen G. Radiotherapy-Induced Astrocyte Senescence Promotes an Immunosuppressive Microenvironment in Glioblastoma to Facilitate Tumor Regrowth. Adv Sci (Weinh). 2024 Apr;11(15):e2304609. [CrossRef]
- Gong Y, Zeng F, Zhang F, Liu X, Li Z, Chen W, Liu H, Li X, Cheng Y, Zhang J, Feng Y, Wu T, Zhou W, Zhang T. Radiotherapy plus a self-gelation powder encapsulating tRF5-GlyGCC inhibitor potentiates natural kill cell immunity to prevent hepatocellular carcinoma recurrence. J Nanobiotechnology. 2025 Feb 10;23(1):100. [CrossRef]
- Gu X, Ma S, Liang B, Ju S. Serum hsa_tsr016141 as a Kind of tRNA-Derived Fragments Is a Novel Biomarker in Gastric Cancer. Front Oncol. 2021 May 13;11:679366. [CrossRef]
- Zhang Y, Gu X, Qin X, Huang Y, Ju S. Evaluation of serum tRF-23-Q99P9P9NDD as a potential biomarker for the clinical diagnosis of gastric cancer. Mol Med. 2022 Jun 11;28(1):63. [CrossRef]
- Huang Y, Zhang H, Gu X, Qin S, Zheng M, Shi X, Peng C, Ju S. Elucidating the Role of Serum tRF-31-U5YKFN8DYDZDD as a Novel Diagnostic Biomarker in Gastric Cancer (GC). Front Oncol. 2021 Aug 23;11:723753. [CrossRef]
- Zhang F, Shi J, Wu Z, Gao P, Zhang W, Qu B, Wang X, Song Y, Wang Z. A 3’-tRNA-derived fragment enhances cell proliferation, migration and invasion in gastric cancer by targeting FBXO47. Arch Biochem Biophys. 2020 Sep 15;690:108467.
- Zhang Y, Gu X, Li Y, Li X, Huang Y, Ju S. Transfer RNA-derived fragment tRF-23-Q99P9P9NDD promotes progression of gastric cancer by targeting ACADSB. J Zhejiang Univ Sci B. 2024 May 15;25(5):438-450. [CrossRef]
- Shen Y, Yu X, Ruan Y, Li Z, Xie Y, Yan Z, Guo J. Global profile of tRNA-derived small RNAs in gastric cancer patient plasma and identification of tRF-33-P4R8YP9LON4VDP as a new tumor suppressor. Int J Med Sci. 2021 Feb 4;18(7):1570-1579. [CrossRef]
- Shen Y, Xie Y, Yu X, Zhang S, Wen Q, Ye G, Guo J. Clinical diagnostic values of transfer RNA-derived fragment tRF-19-3L7L73JD and its effects on the growth of gastric cancer cells. J Cancer. 2021 Apr 2;12(11):3230-3238. [CrossRef]
- Zhu L, Li Z, Yu X, Ruan Y, Shen Y, Shao Y, Zhang X, Ye G, Guo J. The tRNA-derived fragment 5026a inhibits the proliferation of gastric cancer cells by regulating the PTEN/PI3K/AKT signaling pathway. Stem Cell Res Ther. 2021 Jul 22;12(1):418. [CrossRef]
- Xu W, Zhou B, Wang J, Tang L, Hu Q, Wang J, Chen H, Zheng J, Yan F, Chen H. tRNA-Derived Fragment tRF-Glu-TTC-027 Regulates the Progression of Gastric Carcinoma via MAPK Signaling Pathway. Front Oncol. 2021 Aug 23;11:733763. [CrossRef]
- Xu W, Zheng J, Wang X, Zhou B, Chen H, Li G, Yan F. tRF-Val-CAC-016 modulates the transduction of CACNA1d-mediated MAPK signaling pathways to suppress the proliferation of gastric carcinoma. Cell Commun Signal. 2022 May 19;20(1):68. [CrossRef]
- Cui H, Liu Z, Peng L, Liu L, Xie X, Zhang Y, Gao Z, Zhang C, Yu X, Hu Y, Liu J, Shang L, Li L. A novel 5’tRNA-derived fragment tRF-Tyr inhibits tumor progression by targeting hnRNPD in gastric cancer. Cell Commun Signal. 2025 Feb 14;23(1):88. [CrossRef]
- Cui H, Li H, Wu H, Du F, Xie X, Zeng S, Zhang Z, Dong K, Shang L, Jing C, Li L. A novel 3’tRNA-derived fragment tRF-Val promotes proliferation and inhibits apoptosis by targeting EEF1A1 in gastric cancer. Cell Death Dis. 2022 May 18;13(5):471. [CrossRef]
- Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I, Jemal A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024 May-Jun;74(3):229-263.
- Huang B, Yang H, Cheng X, Wang D, Fu S, Shen W, Zhang Q, Zhang L, Xue Z, Li Y, Da Y, Yang Q, Li Z, Liu L, Qiao L, Kong Y, Yao Z, Zhao P, Li M, Zhang R. tRF/miR-1280 Suppresses Stem Cell-like Cells and Metastasis in Colorectal Cancer. Cancer Res. 2017 Jun 15;77(12):3194-3206. [CrossRef]
- Xu R, Du A, Deng X, Du W, Zhang K, Li J, Lu Y, Wei X, Yang Q, Tang H. tsRNA-GlyGCC promotes colorectal cancer progression and 5-FU resistance by regulating SPIB. J Exp Clin Cancer Res. 2024 Aug 17;43(1):230.
- Primeaux M, Liu X, Gowrikumar S, Fatima I, Fisher KW, Bastola D, Vecchio AJ, Singh AB, Dhawan P. Claudin-1 interacts with EPHA2 to promote cancer stemness and chemoresistance in colorectal cancer. Cancer Lett. 2023 Nov 28;579:216479. [CrossRef]
- Ye C, Cheng F, Huang L, Wang K, Zhong L, Lu Y, Ouyang M. New plasma diagnostic markers for colorectal cancer: transporter fragments of glutamate tRNA origin. J Cancer. 2024 Jan 12;15(5):1299-1313. [CrossRef]
- Wu Y, Yang X, Jiang G, Zhang H, Ge L, Chen F, Li J, Liu H, Wang H. 5’-tRF-GlyGCC: a tRNA-derived small RNA as a novel biomarker for colorectal cancer diagnosis. Genome Med. 2021 Feb 9;13(1):20.
- Dang Y, Dai L, Xu J, Zhou W, Xu Y, Ji G. tRNA-derived fragments are promising biomarkers for screening of early colorectal cancer. MedComm (2020). 2023 May 3;4(3):e227. [CrossRef]
- Huang T, Chen C, Du J, Zheng Z, Ye S, Fang S, Liu K. A tRF-5a fragment that regulates radiation resistance of colorectal cancer cells by targeting MKNK1. J Cell Mol Med. 2023 Dec;27(24):4021-4033. [CrossRef]
- Neamtu AA, Maghiar TA, Alaya A, Olah NK, Turcus V, Pelea D, Totolici BD, Neamtu C, Maghiar AM, Mathe E. A Comprehensive View on the Quercetin Impact on Colorectal Cancer. Molecules. 2022 Mar 14;27(6):1873. [CrossRef]
- Yang C, Song J, Park S, Ham J, Park W, Park H, An G, Hong T, Kim HS, Song G, Lim W. Targeting Thymidylate Synthase and tRNA-Derived Non-Coding RNAs Improves Therapeutic Sensitivity in Colorectal Cancer. Antioxidants (Basel). 2022 Oct 31;11(11):2158. [CrossRef]
- Wong CC, Yu J. Gut microbiota in colorectal cancer development and therapy. Nat Rev Clin Oncol. 2023 Jul;20(7):429-452. [CrossRef]
- Cao KY, Pan Y, Yan TM, Tao P, Xiao Y, Jiang ZH. Antitumor Activities of tRNA-Derived Fragments and tRNA Halves from Non-pathogenic Escherichia coli Strains on Colorectal Cancer and Their Structure-Activity Relationship. mSystems. 2022 Apr 26;7(2):e0016422. [CrossRef]
- Christodoulou S, Katsaraki K, Vassiliu P, Danias N, Michalopoulos N, Tzikos G, Sideris DC, Arkadopoulos N. High Intratumoral i-tRF-GlyGCC Expression Predicts Short-Term Relapse and Poor Overall Survival of Colorectal Cancer Patients, Independent of the TNM Stage. Biomedicines. 2023 Jul 8;11(7):1945.
- Tan L, Wu X, Tang Z, Chen H, Cao W, Wen C, Zou G, Zou H. The tsRNAs (tRFdb-3013a/b) serve as novel biomarkers for colon adenocarcinomas. Aging (Albany NY). 2024 Mar 6;16(5):4299-4326. [CrossRef]
- Balatti V, Rizzotto L, Miller C, Palamarchuk A, Fadda P, Pandolfo R, Rassenti LZ, Hertlein E, Ruppert AS, Lozanski A, Lozanski G, Kipps TJ, Byrd JC, Croce CM, Pekarsky Y. TCL1 targeting miR-3676 is codeleted with tumor protein p53 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2015 Feb 17;112(7):2169-74. [CrossRef]
- Veneziano D, Tomasello L, Balatti V, Palamarchuk A, Rassenti LZ, Kipps TJ, Pekarsky Y, Croce CM. Dysregulation of different classes of tRNA fragments in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2019 Nov 26;116(48):24252-24258. [CrossRef]
- Syu YC, Hatterschide J, Budding CR, Tang Y, Musier-Forsyth K. Human T-cell leukemia virus type 1 uses a specific tRNAPro isodecoder to prime reverse transcription. RNA. 2024 Jul 16;30(8):967-976. [CrossRef]
- Maute RL, Schneider C, Sumazin P, Holmes A, Califano A, Basso K, Dalla-Favera R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc Natl Acad Sci U S A. 2013 Jan 22;110(4):1404-9. [CrossRef]
- Guzzi N, Muthukumar S, Cieśla M, Todisco G, Ngoc PCT, Madej M, Munita R, Fazio S, Ekström S, Mortera-Blanco T, Jansson M, Nannya Y, Cazzola M, Ogawa S, Malcovati L, Hellström-Lindberg E, Dimitriou M, Bellodi C. Pseudouridine-modified tRNA fragments repress aberrant protein synthesis and predict leukaemic progression in myelodysplastic syndrome. Nat Cell Biol. 2022 Mar;24(3):299-306. [CrossRef]
- Shao Y, Sun Q, Liu X, Wang P, Wu R, Ma Z. tRF-Leu-CAG promotes cell proliferation and cell cycle in non-small cell lung cancer. Chem Biol Drug Des. 2017 Nov;90(5):730-738.
- Yang W, Gao K, Qian Y, Huang Y, Xiang Q, Chen C, Chen Q, Wang Y, Fang F, He Q, Chen S, Xiong J, Chen Y, Xie N, Zheng D, Zhai R. A novel tRNA-derived fragment AS-tDR-007333 promotes the malignancy of NSCLC via the HSPB1/MED29 and ELK4/MED29 axes. J Hematol Oncol. 2022 May 7;15(1):53.
- Hu F, Niu Y, Mao X, Cui J, Wu X, Simone CB 2nd, Kang HS, Qin W, Jiang L. tsRNA-5001a promotes proliferation of lung adenocarcinoma cells and is associated with postoperative recurrence in lung adenocarcinoma patients. Transl Lung Cancer Res. 2021 Oct;10(10):3957-3972.
- Gu W, Shi J, Liu H, Zhang X, Zhou JJ, Li M, Zhou D, Li R, Lv J, Wen G, Zhu S, Qi T, Li W, Wang X, Wang Z, Zhu H, Zhou C, Knox KS, Wang T, Chen Q, Qian Z, Zhou T. Peripheral blood non-canonical small non-coding RNAs as novel biomarkers in lung cancer. Mol Cancer. 2020 Nov 12;19(1):159. [CrossRef]
- Zheng B, Song X, Wang L, Zhang Y, Tang Y, Wang S, Li L, Wu Y, Song X, Xie L. Plasma exosomal tRNA-derived fragments as diagnostic biomarkers in non-small cell lung cancer. Front Oncol. 2022 Oct 31;12:1037523. [CrossRef]
- Zhang M, Li F, Wang J, He W, Li Y, Li H, Wei Z, Cao Y. tRNA-derived fragment tRF-03357 promotes cell proliferation, migration and invasion in high-grade serous ovarian cancer. Onco Targets Ther. 2019 Aug 16;12:6371-6383. [CrossRef]
- Panoutsopoulou K, Magkou P, Dreyer T, Dorn J, Obermayr E, Mahner S, van Gorp T, Braicu I, Magdolen V, Zeillinger R, Avgeris M, Scorilas A. tRNA-derived small RNA 3’U-tRFValCAC promotes tumour migration and early progression in ovarian cancer. Eur J Cancer. 2023 Feb;180:134-145. [CrossRef]
- Panoutsopoulou K, Dreyer T, Dorn J, Obermayr E, Mahner S, Gorp TV, Braicu I, Zeillinger R, Magdolen V, Avgeris M, Scorilas A. tRNAGlyGCC-Derived Internal Fragment (i-tRF-GlyGCC) in Ovarian Cancer Treatment Outcome and Progression. Cancers (Basel). 2021 Dec 22;14(1):24.
- Wang Y, Xia W, Shen F, Zhou J, Gu Y, Chen Y. tRNA-derived fragment tRF-Glu49 inhibits cell proliferation, migration and invasion in cervical cancer by targeting FGL1. Oncol Lett. 2022 Aug 9;24(4):334.
- Chen Z, Qi M, Shen B, Luo G, Wu Y, Li J, Lu Z, Zheng Z, Dai Q, Wang H. Transfer RNA demethylase ALKBH3 promotes cancer progression via induction of tRNA-derived small RNAs. Nucleic Acids Res. 2019 Mar 18;47(5):2533-2545. [CrossRef]
- Papadimitriou MA, Avgeris M, Levis P, Papasotiriou EC, Kotronopoulos G, Stravodimos K, Scorilas A. tRNA-Derived Fragments (tRFs) in Bladder Cancer: Increased 5’-tRF-LysCTT Results in Disease Early Progression and Patients’ Poor Treatment Outcome. Cancers (Basel). 2020 Dec 6;12(12):3661. [CrossRef]
- Xu X, Chen J, Bai M, Liu T, Zhan S, Li J, Ma Y, Zhang Y, Wu L, Zhao Z, Liu S, Chen X, Fang F, Guo H, Sun Y, Yang R. Plasma tsRNA Signatures Serve as a Novel Biomarker for Bladder Cancer. Cancer Sci. 2025 Feb 13. [CrossRef]
- Ying X, Hu W, Huang Y, Lv Y, Ji D, Chen C, Yang B, Zhang C, Liang Y, Zhang H, Liu M, Yuan G, Wu W, Ji W. A Novel tsRNA, m7G-3’ tiRNA LysTTT, Promotes Bladder Cancer Malignancy Via Regulating ANXA2 Phosphorylation. Adv Sci (Weinh). 2024 Aug;11(31):e2400115.
- Nientiedt M, Deng M, Schmidt D, Perner S, Müller SC, Ellinger J. Identification of aberrant tRNA-halves expression patterns in clear cell renal cell carcinoma. Sci Rep. 2016 Nov 24;6:37158. [CrossRef]
- Zhao C, Tolkach Y, Schmidt D, Kristiansen G, Müller SC, Ellinger J. 5’-tRNA Halves are Dysregulated in Clear Cell Renal Cell Carcinoma. J Urol. 2018 Feb;199(2):378-383. [CrossRef]
- Yang M, Mo Y, Ren D, Liu S, Zeng Z, Xiong W. Transfer RNA-derived small RNAs in tumor microenvironment. Mol Cancer. 2023 Feb 16;22(1):32. [CrossRef]
- Lee YS, Shibata Y, Malhotra A, Dutta A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009 Nov 15;23(22):2639-49. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



