Submitted:
15 April 2025
Posted:
16 April 2025
You are already at the latest version
Abstract
Metformin, a cornerstone treatment for type 2 diabetes, has recently emerged as a multifunctional therapeutic agent with applications extending far beyond glycemic control. This mini-review explores its promising roles within extracellular matrix (ECM)-based three-dimensional (3D) cell culture models, which closely mimic the in vivo cellular microenvironment. Through modulation of key pathways—including AMPK activation, inhibition of TGF-β signaling, suppression of aerobic glycolysis, and regulation of microRNAs—metformin exerts profound effects on ECM remodeling, fibrosis attenuation, and tumor progression. Furthermore, its integration into ECM-inspired drug delivery systems and regenerative scaffolds has shown great potential in applications such as diabetic wound healing, bone repair, and implant integration. Collectively, current findings highlight metformin as a versatile agent capable of modulating ECM biology in disease-specific 3D systems, paving the way for innovative therapeutic strategies in oncology, metabolic disorders, and tissue engineering.
Keywords:
Metformin
; Extracellular matrix
; 3D cultures
; Type 2 diabetes
1. Introduction
Metformin, a drug primarily used to treat type 2 diabetes, has garnered substantial attention in medical research. Beyond its glucose-lowering effects, studies have revealed that metformin can exert therapeutic benefits in various diseases through multiple cellular mechanisms, including metabolic regulation, inflammation reduction, and modulation of ECM properties, as evidenced by recent review articles [1,2,3].
The ECM plays a crucial role in maintaining cell structure, signaling, and tissue homeostasis, and serving as a structural and functional support for cells, plays a pivotal role in physiological and pathological processes [4,5]. Alterations in ECM structure and function can contribute to various diseases, including cancer, fibrosis, and metabolic disorders [6,7,8].
In three-dimensional (3D) cell culture systems, which more accurately mimic the physiological microenvironment compared to traditional 2D cultures, ECM dynamics are essential for cell proliferation, differentiation, and migration [9,10]. Emerging evidence suggests that metformin can alter ECM composition and mechanics, potentially influencing cell-ECM interactions, remodeling processes, and mechanotransduction pathways [11].These findings highlight a novel avenue for metformin’s therapeutic applications in conditions where ECM remodeling is dysregulated, such as tumor microenvironments and fibrotic diseases.
Understanding metformin’s role in ECM regulation could pave the way for novel therapeutic strategies that leverage its pleiotropic effects, particularly in cancer treatment and tissue engineering. This review aims to explore the multifaceted roles of metformin in ECM regulation within 3D cell cultures, emphasizing its potential applications beyond traditional glycemic control
2. Metformin: Its Properties and Applications
Metformin is a widely used medication primarily for managing type 2 diabetes, known for its glucose-lowering effects. Beyond its metabolic actions, metformin also exhibits several non-metabolic effects that are gaining attention. Metformin primarily reduces hepatic glucose production, improving hyperglycemia and insulin sensitivity through both AMPK-dependent and independent pathways [12,13]. It influences mitochondrial respiration, enhancing energy metabolism in the liver and other tissues and also decreases lipid secretion from intestinal cells and enhances fatty acid oxidation in adipose tissue and muscles, contributing to improved metabolic profiles [13,14]. Metformin has favorable effects on cardiovascular health, reducing heart failure incidence and mortality by improving myocardial energy metabolism and reducing cardiac remodeling [15,16].
It exhibits potential anti-cancer effects by inhibiting mitochondrial complex I, causing bioenergetic stress in cancer cells, and affecting cancer cell metabolism [17,18]. Metformin resulted in cell cycle arrest in the sub-G1 phase with G1 and indicate the positive impact in treating human breast cancer [19].Also, Metformin reduces pro-inflammatory markers and regulates adipokines, which are beneficial in conditions like metabolic syndrome and has been linked to increased levels of neurotrophic factors [20,21] (Figure 1).
3. Three-Dimensional(3D) Cell Culture Models of Disease
Three-dimensional (3D) cell culture models that incorporate extracellular matrix (ECM) components are crucial for accurately mimicking the in vivo environment in disease research. These models provide a more physiologically relevant context for studying cellular behavior, disease mechanisms, and drug responses [22,23].
3D cultures are particularly useful in cancer research, where they help in understanding tumor biology, including cell-extracellular matrix interactions and the development of hypoxic microenvironments, which are crucial for evaluating drug responses and resistance mechanisms [23,24,25]. They also play a role in studying neurodegenerative diseases, such as Alzheimer’s, by mimicking the brain’s extracellular matrix [26,27].
4. Extracellular Matrix (ECM)
The ECM is composed of proteins such as collagens, elastin, laminins, and fibronectin, as well as proteoglycans and glycosaminoglycans (GAG) [4,30]. These components form a dynamic network that interacts with cell surface receptors to influence cell behavior [30,31]. It provides structural support, facilitates cell adhesion, and transduces signals that regulate cell survival, growth, migration, and differentiation [30,32].Abnormal ECM remodeling is linked to various diseases, including cancer, fibrosis, and metabolic disorders. It can influence tumor progression and metastasis by modifying the tumor microenvironment [33,34,35].
Creating an extracellular matrix (ECM) for 3D cell cultures involves using various natural and synthetic materials to mimic the in vivo environment and can be categorized based on the presence or absence of scaffolds into two main types: scaffold-based ECM and scaffold-free ECM.
I. scaffold-based ECM
This type involves the use of engineered scaffolds that mimic the natural ECM, providing structural support and biochemical cues for cell attachment, proliferation, and differentiation. These scaffolds can be made from various materials, including synthetic polymers, natural component and decellularized tissues, and are designed to facilitate tissue regeneration in applications such as bone, cartilage, and skin repair [36]. Scaffold-based approaches allow for controlled architecture, which can influence cellular behavior and tissue organization [37].
a-Natural ECM Components
Matrigel: A gelatinous protein mixture derived from mouse sarcoma, commonly used for its ability to support cell growth and differentiation [38].
Hydrogels: These are water-swollen networks that can be made from natural polymers (like alginate or collagen) or synthetic materials. They provide a scaffold for cells, promoting interactions and mimicking tissue properties [39].
b-Synthetic ECM
These are engineered using materials chemistry to replicate the in vivo cell microenvironments. They involve tuning biochemical and structural features to regulate cell fate, using scaffolds that incorporate signaling biomolecules to create bioresponsive environments [40].
c-Decellularized ECM (dECM):
dECM is derived from tissues and used to create physiomimetic 3D tumor models. It closely mimics the native tumor-supporting matrix, providing a robust platform for cancer research [41].
II. scaffold free ECM
In scaffold-free systems, cells are allowed to aggregate and form their own ECM without the aid of external scaffolding materials. This method can lead to the creation of more physiologically relevant tissue constructs, as cells interact directly with each other and produce their own ECM [42]. Scaffold-free techniques are particularly advantageous for high-throughput applications, such as drug screening and disease modeling, although they may face challenges in maintaining cell viability and function over time [43].
5. Effects of Metformin on ECM
Metformin has notable effects on the extracellular matrix (ECM) across various tissues, primarily through its action on cellular signaling pathways. Metformin reduces excessive ECM deposition in white adipose tissue and liver by activating AMP-activated protein kinase (AMPK) and inhibiting the transforming growth factor-beta1 (TGF-β1)/Smad3 signaling pathway. This action decreases collagen deposition and fibrotic gene expression, which is beneficial in conditions like obesity and liver fibrosis [44,45]. In hepatocellular carcinoma (HCC), metformin’s effects are influenced by the stiffness of the ECM. It inhibits the proliferation, migration, and invasion of cancer cells by modulating the PTEN/PI3K/Akt pathway. However, increased matrix stiffness can attenuate these effects, indicating a complex interaction between metformin and the ECM in cancer settings [46]. In aged female mice, metformin alters ECM-related gene expression in the heart [47], although it does not improve cardiac function. In intervertebral disc cells, metformin affects ECM-like structures, potentially leading to adverse effects [48]. Conversely, it enhances the release and quality of extracellular vesicles from mesenchymal stem cells, which may aid in intervertebral disc regeneration [49]. Metformin inhibits ECM-related processes such as vascular calcification and tumor cell migration by affecting pathways like ferroptosis and matrix metalloproteinase-9 (MMP-9) activation, respectively. These actions are mediated through AMPK activation and other signaling pathways [50,51].
6. Key Mechanisms Underlying Metformin’s Effects on the ECM
To elucidate how metformin exerts its diverse effects on ECM remodeling within 3D systems, we outline the core molecular mechanisms underpinning its activity.
I. Modulation of miRNAs
Metformin upregulates miR-33a, which targets and downregulates c-MYC, reducing ECM deposition and fibrosis [52]. It increases DICER expression (critical for miRNA processing), altering energy metabolism-related miRNAs that suppress ECM-related genes [52,53].
In cancer, metformin elevates miR-34a and miR-200, inhibiting epithelial-mesenchymal transition (EMT) and collagen production. It Also, regulates miR-143-3p, to inhibit ECM deposition Metformin [53,54].
II. Inhibition of Aerobic Glycolysis
Metformin suppresses PFKFB3 (a key glycolytic enzyme), reducing lactate production and glucose consumption. This inhibition is mediated via AMPK/mTOR signaling, lowering collagen I/III and α-SMA synthesis [53,55]. Blocking glycolysis with metformin prevents ECM remodeling in fibrotic conditions.
III. AMPK Activation
Mitochondrial complex I inhibition: Metformin reduces ATP, increasing AMP/ATP ratios to activate AMPK, which inhibits mTOR and ECM synthesis [55].
Lysosomal v-ATPase pathway: At clinical doses, metformin binds PEN2, inhibiting v-ATPase and activating AMPK independently of AMP levels [56].AMPK phosphorylates ACC1/2 and suppresses mTOR/4E-BP1, reducing collagen and fibronectin production [56,57].
IV. TGF-β Inhibition
Metformin blocks TGF-β1-induced Smad2/3 phosphorylation, preventing myofibroblast differentiation and ECM accumulation. AMPK activation by metformin disrupts TGF-β/Smad3 signaling, lowering α-SMA and fibronectin expression [44], suppresses TGF-β signaling, a key driver of fibrosis in various diseases [58].
These mechanisms not only highlight metformin’s pleiotropic influence on ECM dynamics but also underpin its therapeutic efficacy in 3D disease models—particularly in cancer, fibrosis, and metabolic dysfunctions—as explored in the following sections (Figure 2).
7. Application of Metformin in 3D Disease Models
7.1. Cancer
Metformin shows significant effects on tumor cell organization and behavior in 3D models. The drug reduces sphere-forming ability and targets cancer stem/progenitor cell populations, though it does not completely eliminate them [59]. In 3D spheroid models, metformin treatment leads to decreased invasive capacity of cancer cells, with treated spheroids showing reduced growth after 37 hours compared to untreated controls [60].
A key mechanism of metformin’s impact on cell behavior involves its effect on cell-ECM adhesion. The drug increases cell adhesion to collagen by approximately 35% [61]. This enhanced adhesion is partly due to metformin’s ability to upregulate integrin β1 expression, which plays a crucial role in maintaining cell-matrix adhesion [62]. In terms of cell migration and invasion, metformin demonstrates significant inhibitory effects. Studies have shown that the drug can reduce cell migration by approximately 63% in the absence of ECM and decrease invasion by about 40% in the presence of ECM [63]. These effects appear to be consistent across different glucose conditions, suggesting that metformin’s impact on invasion is not dependent on glucose levels [60].
In multicellular spheroid models, metformin disrupts large single-cluster formation, causing cells to become more loosely attached to each other [64]. However, when used as a single agent in some contexts, such as pancreatic microtumors with cancer-associated fibroblasts, metformin’s efficacy may be limited, though it can enhance the effectiveness of other treatments like oxaliplatin or photodynamic therapy [65] (Table 1).
7.2. Fibrose
The understanding of fibrosis and its relationship with the extracellular matrix (ECM) is fundamental to studying metformin’s therapeutic potential. Fibrosis is characterized by excessive deposition of ECM proteins, with collagens and fibronectins being the predominant components [66].
At the cellular level, fibroblasts are the primary cells responsible for maintaining and producing various ECM components, including collagen, elastin, and proteopolysaccharides [67].
During fibrotic tissue remodeling, a crucial transformation occurs where fibroblasts become activated and differentiate into myofibroblasts. These activated cells are characterized by their expression of α-smooth muscle actin (αSMA) and increased production of ECM proteins [68].
The myofibroblasts significantly contribute to structural and functional changes in tissues by increasing the deposition of ECM components, particularly collagen types I and III [68,69].
The regulation of ECM production and degradation involves complex molecular mechanisms. Fibroblasts secrete various growth factors, including TGF-β and TNF-α, as well as matrix metalloproteinases (MMPs). These MMPs are metal-dependent proteolytic enzymes that play crucial roles in ECM degradation, cell migration, differentiation, and tissue reconstitution [67].
Metformin demonstrates consistent effects in reducing the production and accumulation of extracellular matrix components across multiple studies. At concentrations of 1-10 mmol/L, metformin significantly decreases the expression of major ECM proteins, including collagen type I (COL1A1) and collagen type III (COL3A1) [3].This reduction extends to other ECM components, such as elastin (ELN) and hyaluronic acid (HA), suggesting a broad impact on matrix composition [3,70].
A key mechanism of metformin’s anti-fibrotic action involves its ability to suppress the transformation of fibroblasts into myofibroblasts, as evidenced by decreased expression of α-smooth muscle actin (α-SMA) [71]. This effect has been demonstrated across multiple tissue types, including cardiac tissue, where metformin treatment significantly reduced fibrotic gene expression and ECM deposition [72,73].
At the molecular level, metformin’s effects are mediated through the suppression of key signaling molecules involved in ECM production. The drug reduces the expression of transforming growth factor-β (TGF-β), platelet-derived growth factor-β (PDGF-β), and downstream signaling via SMAD-2 [3]. Additionally, metformin decreases the production of fibronectin, another major ECM component, particularly in response to PDGF-BB stimulation [45].
A particularly interesting mechanism is metformin’s ability to promote trans differentiation of myofibroblasts. Through AMPK activation, metformin triggers a phenotypic switch from myofibroblasts to lipo-fibroblasts, involving increased bone morphogenetic protein 2 expression and peroxisome proliferator-activated receptor-gamma phosphorylation [74]. This process, also termed fibrosis reversion, represents a promising therapeutic approach, particularly in lung tissue [75].
In cardiac tissue models, metformin administration significantly reduces the expression of collagen types I and III, leading to improved cardiac fibrosis outcomes and reduced histopathological necrotic areas [76].
Studies on orbital fibroblasts (OFs) in thyroid-associated ophthalmopathy demonstrate that metformin inhibits TGF-β1-induced expression of multiple fibrosis-related molecules, including αSMA, various collagen types (COL1A1, COL2A1, COL3A1), and fibronectin (FN1) [77].
In renal tissue models, metformin effectively blocks angiotensin II-induced ECM overproduction in cultured renal fibroblasts. This effect extends to in vivo models, where metformin treatment reduces the expression of fibronectin and collagen I in unilateral ureteral obstruction (UUO) mouse models [78].In bone tissue models, metformin influences ECM composition differently, promoting the formation of mineralized extracellular matrix rich in calcium and phosphorous deposits [79]. The development and validation of more sophisticated 3D culture methods represent a critical next step in understanding metformin’s therapeutic potential. These advanced models are needed to better evaluate drug efficacy, optimal dosage regimens, and administration routes (Table 1).
7.3. Diabetes and Metabolic Diseases
The application of 3D culture techniques in diabetes research has shown promising potential in both regenerative medicine and drug discovery [80].
These models are especially valuable because diabetes affects multiple organs, and 3D systems can more accurately replicate the pathophysiology of diabetic complications through various formats such as spheroids, organoids, and bioprints [80].
Researchers have successfully demonstrated practical applications, such as the formation of insulin-producing cells from human amniotic epithelial cells in 3D spheroid cultures, showing glucose-dependent insulin secretion [81,82].
The response to metformin in 3D models varies depending on glucose concentration and cell type. Ghandour.f et al. using co-culture systems has shown that metformin’s effects on angiogenic factors are glucose-dependent. For instance, in endothelial cells, metformin inhibits pro-angiogenic factors like EMMPRIN and MMP-9 across various glucose concentrations, while its effect on VEGF secretion is only observed under high glucose conditions [83].
Metformin’s effects in 3D culture models have been extensively studied across various tissue types, revealing its diverse therapeutic mechanisms. In intestinal organoids, metformin inhibits cell proliferation through two main pathways: AMPK activation and p53-dependent activation of REDD1, which leads to mTOR inhibition and cell cycle arrest [84,85].
In pancreatic models, metformin shows significant protective effects, particularly in maintaining cell viability under high glucose conditions [86]. The drug protects beta cells from fatty acid-induced apoptosis through AMPK-mediated autophagy activation [87].
The drug’s effects on adipose tissue in 3D models are particularly noteworthy. Metformin impairs adipogenesis by improving stemness in human adipose-derived stem cells through autophagy activation and mTOR signaling inhibition [88]. Interestingly, metformin’s effects on adipogenesis appear to be dose-dependent, with lower concentrations potentially promoting and higher concentrations inhibiting adipogenesis [89,90].
In liver models, metformin demonstrates significant anti-steatotic properties, reducing cellular lipid content and fatty acid consumption [91,92].The drug’s effectiveness in 3D culture environments is enhanced by the improved cell-cell communication and more frequent drug-cell interactions provided by these models [92].
Recent studies using multi-organ models have shown that metformin can rescue mitochondrial dysfunction and improve glucose transport in both liver tissue and organoid islets under high glucose conditions [93]. This demonstrates the drug’s potential to address multiple aspects of metabolic pathology simultaneously when studied in complex 3D systems (Table 1).
8. Other Application of Metformin with ECM
In controlled drug delivery systems, extracellular matrix (ECM) can be utilized as a drug delivery system. Metformin-loaded ECM refers to a biomaterial system that incorporates metformin, a widely used antidiabetic drug, into a scaffold made from ECM components. This approach aims to enhance drug delivery and therapeutic efficacy, particularly in regenerative medicine and cancer treatment.
A key advantage of metformin in tissue regeneration applications is its ability to influence multiple cellular processes simultaneously. The drug shows particular promise in regulating mesenchymal stem cells (MSCs), which are crucial for tissue repair and regeneration due to their ability to differentiate into various cell types including osteoblasts, chondrocytes, and adipocytes. Research has demonstrated that metformin can stimulate osteogenesis of umbilical cord MSCs while also modulating immune responses by promoting anti-inflammatory M2 macrophages and reducing pro-inflammatory M1 macrophages [94].
In the context of wound healing, metformin’s immunomodulatory and anti-inflammatory properties make it particularly valuable. Studies have shown that metformin treatment can enhance wound healing processes, increase angiogenesis, improve epithelialization, and promote both hair follicle formation and collagen deposition [95].
Metformin-loaded scaffolds show significant potential in bone tissue engineering, particularly when combined with mesenchymal stem cells. Studies using calcium phosphate cement scaffolds and polylactic acid/polycaprolactone composites have demonstrated enhanced osteogenic differentiation and bone formation in calvarial defects [96]. Calcium phosphate cement-chitosan composites incorporating metformin have shown promise in dental tissue engineering, particularly for dentin regeneration. These scaffolds support dental pulp cell viability while enhancing odontogenic differentiation [97].
Guided bone regeneration (GBR) membranes containing metformin have been developed specifically for periodontal applications. Polycaprolactone/polyvinyl alcohol membranes with 10 wt% metformin have shown particularly good results in improving osteogenic properties and bone regeneration [98,99].
Metformin-loaded scaffolds have demonstrated effectiveness in treating various cranio-maxillo-facial defects. Clinical outcomes show significant improvement when compared to treatments without metformin [98].
Sol-gel coatings containing metformin have been developed for metallic implants. These functionalized surfaces enhance cell proliferation and metabolic activity of adipose-derived stem cells, showing potential for improved implant integration [100,101].
Gelatin-based scaffolds incorporating metformin (GHMS) show promise for treating chronically infected alveolar defects, with evidence of enhanced osteogenic differentiation and vascularization potential [102].
Metformin incorporation into tissue engineering scaffolds has shown remarkable success in promoting wound healing through multiple mechanisms. Studies have demonstrated that metformin treatment increases wound healing rates and angiogenesis, while also improving epithelialization and promoting both hair follicle formation and collagen deposition [95].
The combination of metformin with other therapeutic agents in composite scaffolds has proven particularly effective for diabetic wound healing. When incorporated into chitosan/gelatin/polycaprolactone and polyvinyl pyrrolidone nanofibrous scaffolds alongside other diabetes medications, metformin accelerated wound healing in diabetic rats while improving both dermis and epidermis regeneration. These composite systems also demonstrated reduced inflammatory cell infiltration and edema, suggesting better control of the wound healing environment [103].
Sustained release of metformin from scaffold systems appears to be crucial for optimal wound healing outcomes. Collagen/PLGA nanofibrous scaffold membranes designed for controlled metformin release have shown particular success in diabetic wound models, leading to increased collagen content and more effective wound closure [103]. The drug’s multiple beneficial properties, including anti-inflammatory, anti-fibrotic, and antioxidant effects, work together to enhance the overall wound healing process [95].
Metformin can also be encapsulated in nanoparticles that interact with the ECM, enhancing drug penetration and therapeutic effects in cancer models. For instance, lysozyme-functionalized metformin-loaded nanoparticles have shown improved anticancer efficacy by modifying the ECM of tumor cells [104].
Metformin-loaded extracellular matrix represents a specialized drug delivery system that combines metformin with the natural scaffold properties of extracellular matrix (ECM). The ECM serves as a biocompatible carrier that can release metformin in a controlled manner while maintaining its natural biological functions. This approach addresses two key challenges in drug delivery: achieving sustained release of therapeutic compounds and providing a supportive environment for tissue repair and regeneration. The development of metformin-loaded ECM emerged from the need to improve drug delivery efficiency while leveraging the inherent benefits of natural tissue matrices (Table 2).
9. Challenges and Limitations
The scientific literature reveals significant contradictions in how metformin affects ECM-related processes, particularly in angiogenesis and tissue remodeling. In vitro and in vivo studies have shown paradoxical effects where metformin demonstrates both anti-angiogenic activity while simultaneously enhancing pro-angiogenic mediators [105]. This duality is particularly evident in diabetic conditions, where metformin has shown pro-angiogenic effects in wound healing, cardiovascular disease, and tumor models [106].
The molecular pathways through which metformin affects ECM remodeling also show conflicting patterns. While AMPK activation is widely recognized as metformin’s primary mechanism for reducing oxidative stress and inflammation in cardiac tissue [68,107]. research has revealed that metformin’s effects on collagen synthesis may operate independently of AMPK activation [73]. This suggests the existence of multiple, potentially parallel mechanisms through which metformin influences ECM dynamics, complicating our understanding of its therapeutic effects [68].
These contradictory findings have important implications for clinical applications, particularly in conditions where ECM remodeling plays a crucial role, such as wound healing and cardiovascular disease. In diabetic contexts, metformin has shown beneficial effects on endothelial function and angiogenesis. yet the precise mechanisms and conditions under which these effects manifest remain unclear [106].
10. Future Directions
While current evidence strongly supports the multifaceted role of metformin in 3D cell culture models and ECM regulation, several promising directions remain for future exploration. First, integrating ECM-based 3D models into preclinical studies, including organotypic cultures and patient-derived cells, could enhance translational relevance and better predict clinical outcomes. Second, the development of smart drug delivery systems—particularly those utilizing bioengineered or decellularized ECM scaffolds—may optimize metformin’s therapeutic efficacy and targeting capacity in specific tissues. Additionally, further investigation into the lesser-known molecular pathways influenced by metformin within fibrotic and tumor microenvironments is warranted. Emerging technologies like organ-on-a-chip and multi-tissue 3D constructs also offer a powerful platform to study metformin’s systemic effects, especially in complex diseases like diabetes with cardiovascular or renal comorbidities. Finally, incorporating patient-specific cells into 3D ECM models may facilitate personalized medicine approaches and help stratify metformin responsiveness across diverse populations.
11. Conclusions
Metformin, a common diabetes medication, has shown promise in regulating the extracellular matrix (ECM) within 3D cell cultures. Beyond its metabolic effects, metformin influences ECM composition and remodeling pathways, suggesting potential therapeutic benefits in cancer, fibrosis, and metabolic diseases. Studies have demonstrated its ability to alter ECM stiffness, modulate cell-ECM interactions, and inhibit fibrotic progression through both AMPK-dependent and independent mechanisms. Additionally, its incorporation into tissue engineering and drug delivery systems enhances its regenerative potential in wound healing and bone repair. However, conflicting findings regarding its effects on angiogenesis and ECM remodeling highlight the complexity of its action and the need for further research to optimize its clinical applications. Advancing 3D culture models will be crucial in uncovering the full therapeutic potential of metformin in ECM-related diseases and tissue engineering.
Funding
This work was supported by Isfahan University of Medical Sciences.
Conflicts of interest
The authors declare no conflict of interest.
References
- Dutta, S.; Shah, R.B.; Singhal, S.; Dutta, S.B.; Bansal, S.; Sinha, S.; Haque, M. Metformin: A Review of Potential Mechanism and Therapeutic Utility Beyond Diabetes. Drug Des. Dev. Ther. 2023, 17, 1907–1932. [Google Scholar] [CrossRef]
- Naseri, A.; Sanaie, S.; Hamzehzadeh, S.; Seyedi-Sahebari, S.; Hosseini, M.-S.; Gholipour-Khalili, E.; Majidazar, R.; Seraji, P.; Daneshvar, S.; Rezazadeh-Gavgani, E. Metformin: new applications for an old drug. J. Basic Clin. Physiol. Pharmacol. 2022, 34, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Incio, J.; Suboj, P.; Chin, S.M.; Vardam-Kaur, T.; Liu, H.; Hato, T.; Babykutty, S.; Chen, I.; Deshpande, V.; Jain, R.K.; et al. Metformin Reduces Desmoplasia in Pancreatic Cancer by Reprogramming Stellate Cells and Tumor-Associated Macrophages. PLOS ONE 2015, 10, e0141392. [Google Scholar] [CrossRef] [PubMed]
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.; et al. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Advanced Drug Delivery Reviews. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: implications for fibrotic diseases and cancer. Dis. Model. Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Vaicik, M.K.; Kortesmaa, J.T.; Movérare-Skrtic, S.; Kortesmaa, J.; Soininen, R.; Bergström, G.; Ohlsson, C.; Chong, L.Y.; Rozell, B.; Emont, M.; et al. Laminin α4 Deficient Mice Exhibit Decreased Capacity for Adipose Tissue Expansion and Weight Gain. PLOS ONE 2014, 9, e109854. [Google Scholar] [CrossRef]
- Yang-Jensen, K.C.; Jørgensen, S.M.; Chuang, C.Y.; Davies, M.J. Modification of extracellular matrix proteins by oxidants and electrophiles. Biochem. Soc. Trans. 2024, 52, 1199–1217. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Y.; Long, H.; Liang, Z.; He, Y.; Zhou, Y.; Li, S.; Bao, J. Tissue-specific micropattern array chips fabricated via decellularized ECM for 3D cell culture. MethodsX 2023, 11, 102463. [Google Scholar] [CrossRef]
- Urciuolo, F.; Imparato, G.; Netti, P.A. In vitro strategies for mimicking dynamic cell–ECM reciprocity in 3D culture models. Front. Bioeng. Biotechnol. 2023, 11, 1197075. [Google Scholar] [CrossRef]
- Sainio, A.; Järveläinen, H. Extracellular matrix-cell interactions: Focus on therapeutic applications. Cell. Signal. 2020, 66, 109487. [Google Scholar] [CrossRef] [PubMed]
- He, L. Metformin and Systemic Metabolism. Trends Pharmacol Sci. 2020, 41, 868–81. [Google Scholar] [CrossRef] [PubMed]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia. 2017, 60, 1577–85. [Google Scholar] [CrossRef]
- Wang, Y.; An, H.; Liu, T.; Qin, C.; Sesaki, H.; Guo, S.; Radovick, S.; Hussain, M.; Maheshwari, A.; Wondisford, F.E.; et al. Metformin Improves Mitochondrial Respiratory Activity through Activation of AMPK. Cell Rep. 2019, 29, 1511–1523.e5. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, R.; Parsons, H.L.; Wambolt, R.B.; Paulson, K.; Sharma, V.; Dyck, J.R.B.; Brownsey, R.W.; Allard, M.F. Metabolic actions of metformin in the heart can occur by AMPK-independent mechanisms. Am. J. Physiol. Circ. Physiol. 2008, 294, H2497–H2506. [Google Scholar] [CrossRef]
- Dziubak, A.; Wójcicka, G.; Wojtak, A.; Bełtowski, J. Metabolic Effects of Metformin in the Failing Heart. Int. J. Mol. Sci. 2018, 19, 2869. [Google Scholar] [CrossRef]
- Hsu, S.-K.; Cheng, K.-C.; Mgbeahuruike, M.O.; Lin, Y.-H.; Wu, C.-Y.; Wang, H.-M.D.; Yen, C.-H.; Chiu, C.-C.; Sheu, S.-J. New Insight into the Effects of Metformin on Diabetic Retinopathy, Aging and Cancer: Nonapoptotic Cell Death, Immunosuppression, and Effects beyond the AMPK Pathway. Int. J. Mol. Sci. 2021, 22, 9453. [Google Scholar] [CrossRef]
- Andrzejewski, S.; Siegel, P.M.; St-Pierre, J. Metabolic Profiles Associated With Metformin Efficacy in Cancer. Front. Endocrinol. 2018, 9, 372. [Google Scholar] [CrossRef]
- Haghshenas, M.; Firouzabadi, N.; Akbarizadeh, A.R.; Rashedinia, M. Combination of metformin and gallic acid induces autophagy and apoptosis in human breast cancer cells. Res. Pharm. Sci. 2023, 18, 663–675. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Mokgalaboni, K.; Ziqubu, K.; Cirilli, I.; Marcheggiani, F.; Louw, J.; et al. Adipokines as a therapeutic target by metformin to improve metabolic function: A systematic review of randomized controlled trials. Pharmacol. Res. 2021, 163, 105219. [Google Scholar] [CrossRef]
- Hristova, M.G. Metabolic Syndrome and Neurotrophins: Effects of Metformin and Non-Steroidal Antiinflammatory Drug Treatment. Eurasian J. Med. 2011, 42, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front Mol Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Li, S.; Yang, K.; Chen, X.; Zhu, X.; Zhou, H.; Li, P.; Chen, Y.; Jiang, Y.; Li, T.; Qin, X.; et al. Simultaneous 2D and 3D cell culture array for multicellular geometry, drug discovery and tumor microenvironment reconstruction. Biofabrication 2021, 13, 045013. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Vera, Y.M.; Valdés, J.; Pérez-Navarro, Y.; Mandujano-Lazaro, G.; Marchat, L.A.; Ramos-Payán, R.; Nuñez-Olvera, S.I.; Pérez-Plascencia, C.; López-Camarillo, C. Three-Dimensional 3D Culture Models in Gynecological and Breast Cancer Research. Front. Oncol. 2022, 12, 826113. [Google Scholar] [CrossRef]
- Vella, N.; Fenech, A.G.; Magri, V.P. 3D cell culture models in research: applications to lung cancer pharmacology. Front. Pharmacol. 2024, 15, 1438067. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, S.H.; D'Avanzo, C.; Hebisch, M.; Sliwinski, C.; Bylykbashi, E.; Washicosky, K.J.; Klee, J.B.; Brüstle, O.; ETanzi, R.; et al. A 3D human neural cell culture system for modeling Alzheimer's disease. Nat. Protoc. 2015, 10, 985–1006. [Google Scholar] [CrossRef]
- Bak, A.; Koch, H.; van Loo, K.M.; Schmied, K.; Gittel, B.; Weber, Y.; Ort, J.; Schwarz, N.; Tauber, S.C.; Wuttke, T.V.; et al. Human organotypic brain slice cultures: a detailed and improved protocol for preparation and long-term maintenance. J. Neurosci. Methods 2024, 404, 110055. [Google Scholar] [CrossRef]
- Bloise, N.; Giannaccari, M.; Guagliano, G.; Peluso, E.; Restivo, E.; Strada, S.; Volpini, C.; Petrini, P.; Visai, L. Growing Role of 3D In Vitro Cell Cultures in the Study of Cellular and Molecular Mechanisms: Short Focus on Breast Cancer, Endometriosis, Liver and Infectious Diseases. Cells 2024, 13, 1054. [Google Scholar] [CrossRef]
- Ryoo, H.; Giovanni, R.; Kimmel, H.; Jain, I.; Underhill, G.H. Combinatorial Microgels for 3D ECM Screening and Heterogeneous Microenvironmental Culture of Primary Human Hepatic Stellate Cells. Adv. Sci. 2024, 11, e2303128. [Google Scholar] [CrossRef]
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. Br. J. Pharmacol. 2018, 176, 82–92. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular Matrix Degradation and Remodeling in Development and Disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Zhang, H. Extracellular matrix: an important regulator of cell functions and skeletal muscle development. Cell Biosci. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed]
- Eble, J.A.; Niland, S. The extracellular matrix in tumor progression and metastasis. Clin. Exp. Metastasis 2019, 36, 171–198. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular Matrix Remodeling of Adipose Tissue in Obesity and Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Ding, F.; Gong, L.; Gu, X. Extracellular Matrix Scaffolds for Tissue Engineering and Regenerative Medicine. Curr. Stem Cell Res. Ther. 2017, 12, 233–246. [Google Scholar] [CrossRef]
- Browe, D.C.; Díaz-Payno, P.J.; Freeman, F.E.; Schipani, R.; Burdis, R.; Ahern, D.P.; Nulty, J.M.; Guler, S.; Randall, L.D.; Buckley, C.T.; et al. Bilayered extracellular matrix derived scaffolds with anisotropic pore architecture guide tissue organization during osteochondral defect repair. Acta Biomater. 2022, 143, 266–281. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Akther, F.; Little, P.; Li, Z.; Nguyen, N.-T.; Ta, H.T. Hydrogels as artificial matrices for cell seeding in microfluidic devices. RSC Adv. 2020, 10, 43682–43703. [Google Scholar] [CrossRef]
- Nicolas, J.; Magli, S.; Rabbachin, L.; Sampaolesi, S.; Nicotra, F.; Russo, L. 3D Extracellular Matrix Mimics: Fundamental Concepts and Role of Materials Chemistry to Influence Stem Cell Fate. Biomacromolecules 2020, 21, 1968–1994. [Google Scholar] [CrossRef]
- Ferreira, L.P.; Gaspar, V.M.; Mano, J.F. Decellularized Extracellular Matrix for Bioengineering Physiomimetic 3D in Vitro Tumor Models. Trends Biotechnol. 2020, 38, 1397–1414. [Google Scholar] [CrossRef] [PubMed]
- Valdoz, J.C.; Johnson, B.C.; Jacobs, D.J.; Franks, N.A.; Dodson, E.L.; Sanders, C.; et al. The ECM: To Scaffold, or Not to Scaffold, That Is the Question. Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, F.; Serpelloni, S.; Francis, L.W.; Taraballi, F. Mechanical Studies of the Third Dimension in Cancer: From 2D to 3D Model. Int. J. Mol. Sci. 2021, 22, 10098. [Google Scholar] [CrossRef]
- Luo, T.; Nocon, A.; Fry, J.; Sherban, A.; Rui, X.; Jiang, B.; Xu, X.J.; Han, J.; Yan, Y.; Yang, Q.; et al. AMPK Activation by Metformin Suppresses Abnormal Extracellular Matrix Remodeling in Adipose Tissue and Ameliorates Insulin Resistance in Obesity. Diabetes 2016, 65, 2295–2310. [Google Scholar] [CrossRef]
- Li, Z.; Ding, Q.; Ling, L.P.; Wu, Y.; Meng, D.X.; Li, X.; et al. Metformin attenuates motility, contraction, and fibrogenic response of hepatic stellate cells in vivo and in vitro by activating AMP-activated protein kinase. World J Gastroenterol. 2018, 24, 819–32. [Google Scholar] [CrossRef]
- Gao, X.; Qiao, X.; Xing, X.; Huang, J.; Qian, J.; Wang, Y.; Zhang, Y.; Zhang, X.; Li, M.; Cui, J.; et al. Matrix Stiffness-Upregulated MicroRNA-17-5p Attenuates the Intervention Effects of Metformin on HCC Invasion and Metastasis by Targeting the PTEN/PI3K/Akt Pathway. Front. Oncol. 2020, 10, 1563. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, W.; Liu, Z.; Sheng, S.; Xiong, W.; He, R.; Zhang, X.; Ma, L.; Ju, Z. Effect of Metformin on Cardiac Metabolism and Longevity in Aged Female Mice. Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.E.; Karaarslan, N.; Yilmaz, I.; Yasar, D.S.; Akalan, H.; Ozbek, H. A study of the effects of metformin, a biguanide derivative, on annulus fibrosus and nucleus pulposus cells. Turk. Neurosurg. 2019, 30, 434–441. [Google Scholar] [CrossRef]
- Liao, Z.; Li, S.; Lu, S.; Liu, H.; Li, G.; Ma, L.; Luo, R.; Ke, W.; Wang, B.; Xiang, Q.; et al. Metformin facilitates mesenchymal stem cell-derived extracellular nanovesicles release and optimizes therapeutic efficacy in intervertebral disc degeneration. Biomaterials 2021, 274, 120850. [Google Scholar] [CrossRef]
- Ma, W.-Q.; Sun, X.-J.; Zhu, Y.; Liu, N.-F. Metformin attenuates hyperlipidaemia-associated vascular calcification through anti-ferroptotic effects. Free. Radic. Biol. Med. 2021, 165, 229–242. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Jeong, H.G. Metformin blocks migration and invasion of tumour cells by inhibition of matrix metalloproteinase-9 activation through a calcium and protein kinase Calpha-dependent pathway: phorbol-12-myristate-13-acetate-induced/extracellular signal-regulated kinase/activator protein-1. Br J Pharmacol. 2010, 160, 1195–211. [Google Scholar] [PubMed]
- Blandino, G.; Valerio, M.; Cioce, M.; Mori, F.; Casadei, L.; Pulito, C.; Sacconi, A.; Biagioni, F.; Cortese, G.; Galanti, S.; et al. Metformin elicits anticancer effects through the sequential modulation of DICER and c-MYC. Nat. Commun. 2012, 3, 865. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-J.; Xu, J.; Ye, H.-Y.; Wang, X.-B. Metformin prevents PFKFB3-related aerobic glycolysis from enhancing collagen synthesis in lung fibroblasts by regulating AMPK/mTOR pathway. Exp. Ther. Med. 2021, 21, 581. [Google Scholar] [CrossRef]
- Xu, J.; Xiang, P.; Liu, L.; Sun, J.; Ye, S. Metformin inhibits extracellular matrix accumulation, inflammation and proliferation of mesangial cells in diabetic nephropathy by regulating H19/miR-143-3p/TGF-β1 axis. J Pharm Pharmacol. 2020, 72, 1101–9. [Google Scholar] [CrossRef]
- Ma, T.; Tian, X.; Zhang, B.; Li, M.; Wang, Y.; Yang, C.; Wu, J.; Wei, X.; Qu, Q.; Yu, Y.; et al. Low-dose metformin targets the lysosomal AMPK pathway through PEN2. Nature 2022, 603, 159–165. [Google Scholar] [CrossRef]
- Park, I.H.; Um, J.Y.; Hong, S.M.; Cho, J.S.; Lee, S.H.; Lee, S.H.; et al. Metformin reduces TGF-β1-induced extracellular matrix production in nasal polyp-derived fibroblasts. Otolaryngol Head Neck Surg. 2014, 150, 148–53. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Wang, L.; Liu, X.; Deng, Y.; Yang, S.; Pan, W.; Zhang, X.; Xu, G.M.; Xiao, S.M.; Deng, C.M. Metformin Eliminates Lymphedema in Mice by Alleviating Inflammation and Fibrosis: Implications for Human Therapy. Plast. Reconstr. Surg. 2024, 154, 1128e–1137e. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, C.; Han, R.; Lu, C.; Li, L.; Hu, C.; Feng, M.; Chen, H.; He, Y. Metformin attenuates TGF-β1-induced pulmonary fibrosis through inhibition of transglutaminase 2 and subsequent TGF-β pathways. 3 Biotech 2020, 10, 287. [Google Scholar] [CrossRef]
- Vella, S.; Conaldi, P.G.; Florio, T.; Pagano, A. PPAR Gamma in Neuroblastoma: The Translational Perspectives of Hypoglycemic Drugs. PPAR Res. 2016, 2016, 3038164. [Google Scholar] [CrossRef]
- Boutaud, M.; Auger, C.; Verdier, M.; Christou, N. Metformin Treatment Reduces CRC Aggressiveness in a Glucose-Independent Manner: An In Vitro and Ex Vivo Study. Cancers 2023, 15, 3724. [Google Scholar] [CrossRef]
- Al Hassan, M.; Fakhoury, I.; El Masri, Z.; Ghazale, N.; Dennaoui, R.; El Atat, O.; Kanaan, A.; El-Sibai, M. Metformin Treatment Inhibits Motility and Invasion of Glioblastoma Cancer Cells. Anal. Cell. Pathol. 2018, 2018, 5917470. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Zhang, Z.; Li, Y.; Yi, J.; Zhang, W.; Chen, D.; Ao, J.; Xiao, Z.-X.; Yi, Y. Integrin β1-Mediated Cell–Cell Adhesion Augments Metformin-Induced Anoikis. Int. J. Mol. Sci. 2019, 20, 1161. [Google Scholar] [CrossRef]
- Schexnayder, C.; Broussard, K.; Onuaguluchi, D.; Poché, A.; Ismail, M.; McAtee, L.; Llopis, S.; Keizerweerd, A.; McFerrin, H.; Williams, C. Metformin Inhibits Migration and Invasion by Suppressing ROS Production and COX2 Expression in MDA-MB-231 Breast Cancer Cells. Int. J. Mol. Sci. 2018, 19, 3692. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Al-Sammarraie, N.; DiPette, D.J.; Singh, U.S. Correction: Metformin impairs Rho GTPase signaling to induce apoptosis in neuroblastoma cells and inhibits growth of tumors in the xenograft mouse model of neuroblastoma. Oncotarget 2016, 7, 42843. [Google Scholar] [CrossRef]
- Nocquet, L.; Juin, P.P.; Souazé, F. Mitochondria at Center of Exchanges between Cancer Cells and Cancer-Associated Fibroblasts during Tumor Progression. Cancers 2020, 12, 3017. [Google Scholar] [CrossRef]
- Choi, S.M.; Jang, A.-H.; Kim, H.; Lee, K.H.; Kim, Y.W. Metformin Reduces Bleomycin-induced Pulmonary Fibrosis in Mice. J. Korean Med Sci. 2016, 31, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Jiang, Z.; Li, J.; Lin, H.; Xu, B.; Liao, X.; Fu, Z.; Ao, H.; Guo, G.; Liu, M. Metformin Improves Burn Wound Healing by Modulating Microenvironmental Fibroblasts and Macrophages. Cells 2022, 11, 4094. [Google Scholar] [CrossRef]
- Loi, H.; Kramar, S.; Laborde, C.; Marsal, D.; Pizzinat, N.; Cussac, D.; Roncalli, J.; Boal, F.; Tronchere, H.; Oleshchuk, O.; et al. Metformin Attenuates Postinfarction Myocardial Fibrosis and Inflammation in Mice. Int. J. Mol. Sci. 2021, 22, 9393. [Google Scholar] [CrossRef]
- Pchejetski, D.; Foussal, C.; Alfarano, C.; Lairez, O.; Calise, D.; Guilbeau-Frugier, C.; Schaak, S.; Seguelas, M.-H.; Wanecq, E.; Valet, P.; et al. Apelin prevents cardiac fibroblast activation and collagen production through inhibition of sphingosine kinase 1. Eur. Hear. J. 2011, 33, 2360–2369. [Google Scholar] [CrossRef]
- Kluivers, K.B.; Lince, S.L.; Ruiz-Zapata, A.M.; Post, W.M.; Cartwright, R.; Kerkhof, M.H.; Widomska, J.; De Witte, W.; Pecanka, J.; Kiemeney, L.A.; et al. Molecular Landscape of Pelvic Organ Prolapse Provides Insights into Disease Etiology. Int. J. Mol. Sci. 2023, 24, 6087. [Google Scholar] [CrossRef]
- Wang, L.; Zhong, N.-N.; Wang, X.; Peng, B.; Chen, Z.; Wei, L.; Li, B.; Li, Y.; Cheng, Y. Metformin Attenuates TGF-β1-Induced Fibrosis in Salivary Gland: A Preliminary Study. Int. J. Mol. Sci. 2023, 24, 16260. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Song, J.; Zhang, Y.; Chen, S.; Ruan, H.; Fan, C. Metformin prevents peritendinous fibrosis by inhibiting transforming growth factor-β signaling. Oncotarget 2017, 8, 101784–101794. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Ma, X.; Feng, W.; Fu, Y.; Lu, Z.; Xu, M.; Shen, Q.; Zhu, Y.; Zhang, Y. Metformin attenuates cardiac fibrosis by inhibiting the TGFβ1–Smad3 signalling pathway. Cardiovasc. Res. 2010, 87, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Armendariz, A.I.; Barroso, M.M.; El Agha, E.; Herold, S. 3D In Vitro Models: Novel Insights into Idiopathic Pulmonary Fibrosis Pathophysiology and Drug Screening. Cells 2022, 11, 1526. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Boina, C.; Douanier, A.; Gasque, P. Deciphering the Antifibrotic Property of Metformin. Cells 2022, 11, 4090. [Google Scholar] [CrossRef] [PubMed]
- Zhang J, Yang L, Zhang Q, Shi X, Hua F, Ma J, et al., editors. Nod-like Receptor Protein 3 (Nlrp3) Inflammasomes Inhibition by Metformin Limits Myocardial Ischemia/reperfusion Injury2020.
- Xu, Z.; Ye, H.; Xiao, W.; Sun, A.; Yang, S.; Zhang, T.; Sha, X.; Yang, H. Metformin Attenuates Inflammation and Fibrosis in Thyroid-Associated Ophthalmopathy. Int. J. Mol. Sci. 2022, 23, 15508. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Miao, N.; Xu, J.; Gan, X.; Xu, D.; Zhou, L.; Xue, H.; Zhang, W.; Lu, L. Metformin Prevents Renal Fibrosis in Mice with Unilateral Ureteral Obstruction and Inhibits Ang II-Induced ECM Production in Renal Fibroblasts. Int. J. Mol. Sci. 2016, 17, 146. [Google Scholar] [CrossRef] [PubMed]
- Śmieszek, A.; Tomaszewski, K.A.; Kornicka, K.; Marycz, K. Metformin Promotes Osteogenic Differentiation of Adipose-Derived Stromal Cells and Exerts Pro-Osteogenic Effect Stimulating Bone Regeneration. J. Clin. Med. 2018, 7, 482. [Google Scholar] [CrossRef]
- Mohandas, S.; Gayatri, V.; Kumaran, K.; Gopinath, V.; Paulmurugan, R.; Ramkumar, K.M. New Frontiers in Three-Dimensional Culture Platforms to Improve Diabetes Research. Pharmaceutics 2023, 15, 725. [Google Scholar] [CrossRef]
- Paris, F.; Pizzuti, V.; Marrazzo, P.; Pession, A.; Alviano, F.; Bonsi, L. Perinatal Stem Cell Therapy to Treat Type 1 Diabetes Mellitus: A Never-Say-Die Story of Differentiation and Immunomodulation. Int. J. Mol. Sci. 2022, 23, 14597. [Google Scholar] [CrossRef]
- Okere, B.; Alviano, F.; Costa, R.; Quaglino, D.; Ricci, F.; Dominici, M.; Paolucci, P.; Bonsi, L.; Iughetti, L. In vitro differentiation of human amniotic epithelial cells into insulin-producing 3D spheroids. Int. J. Immunopathol. Pharmacol. 2015, 28, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Ghandour, F.; Kassem, S.; Simanovich, E.; Rahat, M.A. Glucose Promotes EMMPRIN/CD147 and the Secretion of Pro-Angiogenic Factors in a Co-Culture System of Endothelial Cells and Monocytes. Biomedicines 2024, 12, 706. [Google Scholar] [CrossRef]
- Okkelman, I.A.; Dmitriev, R.I.; Foley, T.; Papkovsky, D.B. Use of Fluorescence Lifetime Imaging Microscopy (FLIM) as a Timer of Cell Cycle S Phase. PLOS ONE 2016, 11, e0167385. [Google Scholar] [CrossRef]
- Ben Sahra, I.; Regazzetti, C.; Robert, G.; Laurent, K.; Le Marchand-Brustel, Y.; Auberger, P.; et al. Metformin, independent of AMPK, induces mTOR inhibition and cell-cycle arrest through REDD1. Cancer Res. 2011, 71, 4366–4372. [Google Scholar] [CrossRef]
- Lundquist, I.; Al-Amily, I.M.; Abaraviciene, S.M.; Salehi, A. Metformin Ameliorates Dysfunctional Traits of Glibenclamide- and Glucose-Induced Insulin Secretion by Suppression of Imposed Overactivity of the Islet Nitric Oxide Synthase-NO System. PLOS ONE 2016, 11, e0165668. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jang, J. Construction of 3D hierarchical tissue platforms for modeling diabetes. APL Bioeng. 2021, 5, 041506. [Google Scholar] [CrossRef] [PubMed]
- Chinnapaka, S.; Yang, K.S.; Flowers, Q.; Faisal, M.; Nerone, W.V.; Rubin, J.P.; Ejaz, A. Metformin Improves Stemness of Human Adipose-Derived Stem Cells by Downmodulation of Mechanistic Target of Rapamycin (mTOR) and Extracellular Signal-Regulated Kinase (ERK) Signaling. Biomedicines 2021, 9, 1782. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Y.; Wu, K.; Wang, X. Dual Effects of Metformin on Adipogenic Differentiation of 3T3-L1 Preadipocyte in AMPK-Dependent and Independent Manners. Int. J. Mol. Sci. 2018, 19, 1547. [Google Scholar] [CrossRef]
- Yang, L.; Jia, X.; Fang, D.; Cheng, Y.; Zhai, Z.; Deng, W.; Du, B.; Lu, T.; Wang, L.; Yang, C.; et al. Metformin Inhibits Lipid Droplets Fusion and Growth via Reduction in Cidec and Its Regulatory Factors in Rat Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2022, 23, 5986. [Google Scholar] [CrossRef]
- Irrechukwu, O.; Yeager, R.; David, R.; Ekert, J.; Saravanakumar, A.; Choi, C.K. Applications of microphysiological systems to disease models in the biopharmaceutical industry: Opportunities and challenges. Altex 2023, 40, 485–518. [Google Scholar] [CrossRef]
- Kostrzewski, T.; Cornforth, T.; ASnow, S.; Ouro-Gnao, L.; Rowe, C.; Large, E.M.; Hughes, D.J. Three-dimensional perfused human in vitro model of non-alcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Ka, M.; Hawkins, E.; Pouponnot, C.; Duvillié, B. Modelling human diabetes ex vivo: a glance at maturity onset diabetes of the young. Front. Endocrinol. 2024, 15, 1427413. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Yu, H.; Jin, Y.; Mo, J.; Sui, J.; Qian, X.; Chen, T. Metformin Facilitates Osteoblastic Differentiation and M2 Macrophage Polarization by PI3K/AKT/mTOR Pathway in Human Umbilical Cord Mesenchymal Stem Cells. Stem Cells Int. 2022, 2022, 1–12. [Google Scholar] [CrossRef]
- Harmanci, S.; Dutta, A.; Cesur, S.; Sahin, A.; Gunduz, O.; Kalaskar, D.M.; Ustundag, C.B. Production of 3D Printed Bi-Layer and Tri-Layer Sandwich Scaffolds with Polycaprolactone and Poly (vinyl alcohol)-Metformin towards Diabetic Wound Healing. Polymers 2022, 14, 5306. [Google Scholar] [CrossRef]
- Jiang, L.-L.; Liu, L. Effect of metformin on stem cells: Molecular mechanism and clinical prospect. World J. Stem Cells 2020, 12, 1455–1473. [Google Scholar] [CrossRef]
- Qin, W.; Chen, J.-Y.; Guo, J.; Ma, T.; Weir, M.D.; Guo, D.; Shu, Y.; Lin, Z.-M.; Schneider, A.; Xu, H.H.K. Novel Calcium Phosphate Cement with Metformin-Loaded Chitosan for Odontogenic Differentiation of Human Dental Pulp Cells. Stem Cells Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Zhu, M.; Zhao, Z.; Xu, H.H.K.; Dai, Z.; Yu, K.; Xiao, L.; Schneider, A.; Weir, M.D.; Oates, T.W.; Bai, Y.; et al. Effects of Metformin Delivery via Biomaterials on Bone and Dental Tissue Engineering. Int. J. Mol. Sci. 2022, 23, 15905. [Google Scholar] [CrossRef]
- Ebrahimi, L.; Farzin, A.; Ghasemi, Y.; Alizadeh, A.; Goodarzi, A.; Basiri, A.; Zahiri, M.; Monabati, A.; Ai, J. Metformin-Loaded PCL/PVA Fibrous Scaffold Preseeded with Human Endometrial Stem Cells for Effective Guided Bone Regeneration Membranes. ACS Biomater. Sci. Eng. 2020, 7, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Smieszek, A.; Kornicka, K.; Szłapka-Kosarzewska, J.; Androvic, P.; Valihrach, L.; Langerova, L.; Rohlova, E.; Kubista, M.; Marycz, K. Metformin Increases Proliferative Activity and Viability of Multipotent Stromal Stem Cells Isolated from Adipose Tissue Derived from Horses with Equine Metabolic Syndrome. Cells 2019, 8, 80. [Google Scholar] [CrossRef]
- Śmieszek, A.; Szydlarska, J.; Mucha, A.; Chrapiec, M.; Marycz, K. Enhanced cytocompatibility and osteoinductive properties of sol–gel-derived silica/zirconium dioxide coatings by metformin functionalization. J. Biomater. Appl. 2017, 32, 570–586. [Google Scholar] [CrossRef]
- Fang, C.-H.; Sun, C.-K.; Lin, Y.-W.; Hung, M.-C.; Lin, H.-Y.; Li, C.-H.; Lin, I.-P.; Chang, H.-C.; Sun, J.-S.; Chang, J.Z.-C. Metformin-Incorporated Gelatin/Nano-Hydroxyapatite Scaffolds Promotes Bone Regeneration in Critical Size Rat Alveolar Bone Defect Model. Int. J. Mol. Sci. 2022, 23, 558. [Google Scholar] [CrossRef] [PubMed]
- Yusuf Aliyu, A.; Adeleke, O.A. Nanofibrous Scaffolds for Diabetic Wound Healing. Pharmaceutics. 2023, 15. [Google Scholar] [CrossRef]
- Wang, A.; Madden, L.A.; Paunov, V.N. Enhanced anticancer effect of lysozyme-functionalized metformin-loaded shellac nanoparticles on a 3D cell model: role of the nanoparticle and payload concentrations. Biomater. Sci. 2024, 12, 4735–4746. [Google Scholar] [CrossRef] [PubMed]
- Dallaglio, K.; Bruno, A.; Cantelmo, A.R.; Esposito, A.I.; Ruggiero, L.; Orecchioni, S.; Calleri, A.; Bertolini, F.; Pfeffer, U.; Noonan, D.M.; et al. Paradoxic effects of metformin on endothelial cells and angiogenesis. Carcinog. 2014, 35, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Rodrigues, I.; Andrade, S.; Costa, R.; Soares, R. Metformin Reduces Vascular Assembly in High Glucose-Treated Human Microvascular Endothelial Cells in An AMPK-Independent Manner. Cell J. 2021, 23, 174–183. [Google Scholar] [CrossRef]
- Gundewar, S.; Calvert, J.W.; Jha, S.; Toedt-Pingel, I.; Ji, S.Y.; Nunez, D.; Ramachandran, A.; Anaya-Cisneros, M.; Tian, R.; Lefer, D.J. Activation of AMP-Activated Protein Kinase by Metformin Improves Left Ventricular Function and Survival in Heart Failure. Circ. Res. 2009, 104, 403–411. [Google Scholar] [CrossRef]
Figure 1.
Application of Metformin.
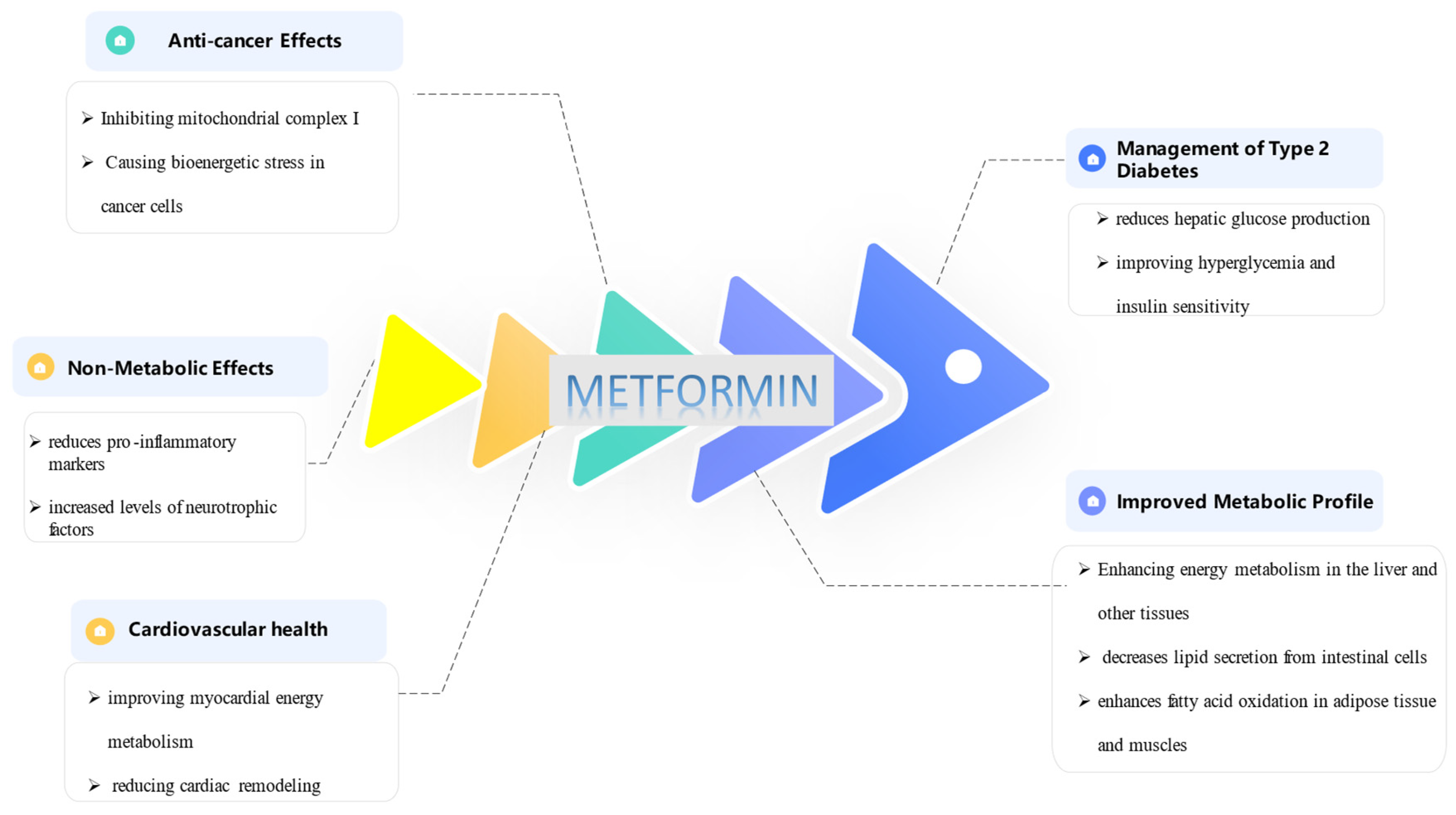
Figure 2.
Key Mechanisms underlying Metformin’s Effects on the ECM:1. Modulation of miRNAs: Research has demonstrated that miR-143-3p regulates cell proliferation and apoptosis by targeting ERK8(extracellular signal-regulated kinase),2. Inhibition of Aerobic Glycolysis,3. AMPK Activation: Metformin activates AMPK (Mitogen-activated protein kinase) inhibits the TGF-β1/Smad3 signaling pathway that leads to a decrease protein synthesis in collagen deposition and fibrotic gene expression,4. TGF-β Inhibition: Metformin suppresses TGF-β signaling, a key driver of fibrosis, LOXL1 (Lysyl oxidase like 1)- an enzyme involved in the crosslinking of collagen and elastin- TIMP1 (Tissue inhibitor of metalloproteinases 1)- TIMP1 is a protein that inhibits matrix metalloproteinases (MMPs) which is degrade the ECM-,α-SMA(alpha-smooth muscle actin)- expressed in myofibroblasts which are a major source of ECM production-,EMT(Epithelial-Mesenchymal Transition)- biological process where epithelial cells (cells that line the surfaces of your body) lose their characteristics and gain those of mesenchymal cells-,HSCs(Hepatic Stellate Cells),MFBs Myofibroblasts.
Figure 2.
Key Mechanisms underlying Metformin’s Effects on the ECM:1. Modulation of miRNAs: Research has demonstrated that miR-143-3p regulates cell proliferation and apoptosis by targeting ERK8(extracellular signal-regulated kinase),2. Inhibition of Aerobic Glycolysis,3. AMPK Activation: Metformin activates AMPK (Mitogen-activated protein kinase) inhibits the TGF-β1/Smad3 signaling pathway that leads to a decrease protein synthesis in collagen deposition and fibrotic gene expression,4. TGF-β Inhibition: Metformin suppresses TGF-β signaling, a key driver of fibrosis, LOXL1 (Lysyl oxidase like 1)- an enzyme involved in the crosslinking of collagen and elastin- TIMP1 (Tissue inhibitor of metalloproteinases 1)- TIMP1 is a protein that inhibits matrix metalloproteinases (MMPs) which is degrade the ECM-,α-SMA(alpha-smooth muscle actin)- expressed in myofibroblasts which are a major source of ECM production-,EMT(Epithelial-Mesenchymal Transition)- biological process where epithelial cells (cells that line the surfaces of your body) lose their characteristics and gain those of mesenchymal cells-,HSCs(Hepatic Stellate Cells),MFBs Myofibroblasts.
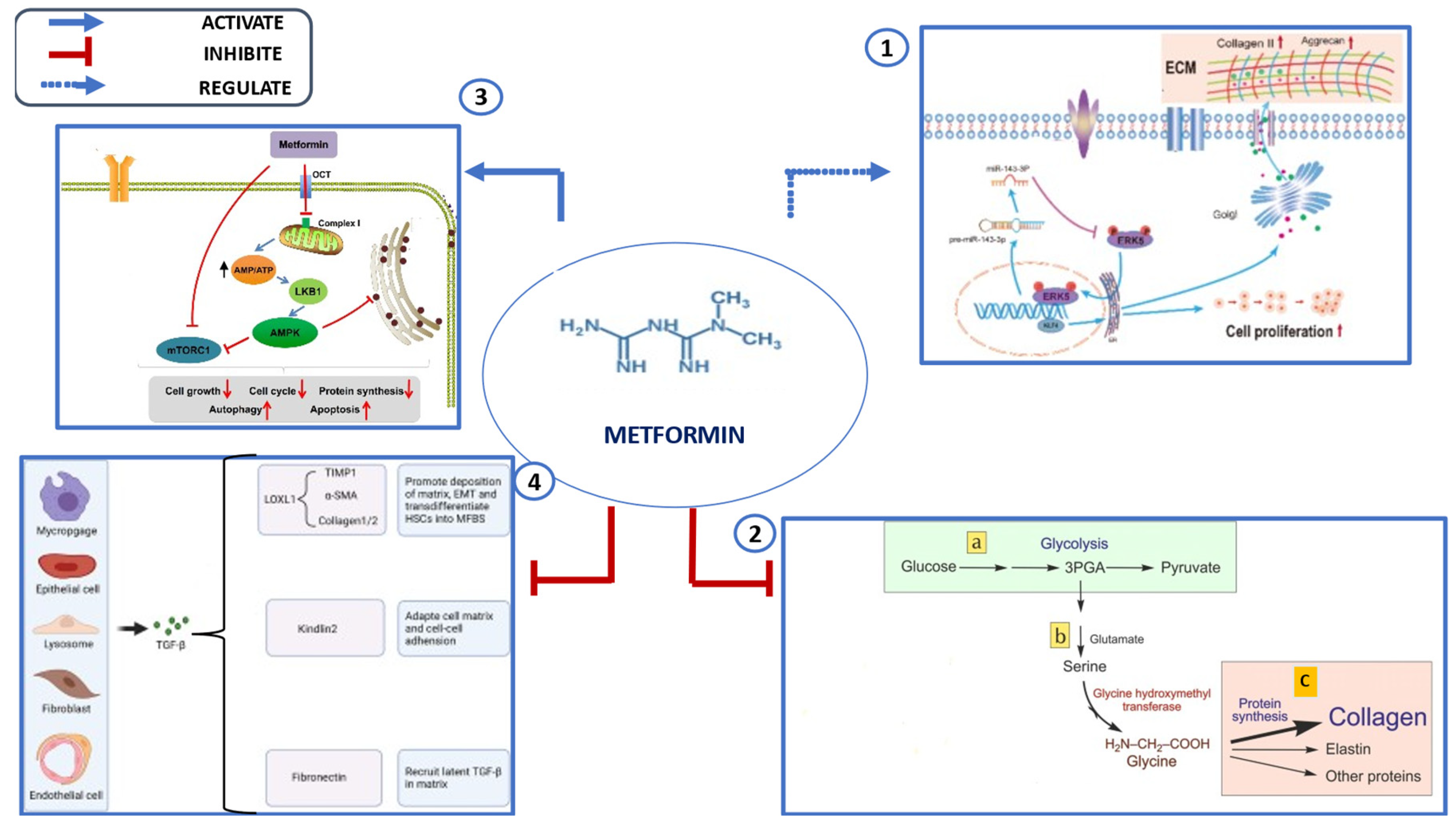

Table 1.
Application of Metformin in 3D disease models.
| Disease Models | Effects of Metformin | Key Mechanisms | Ref. | |
|---|---|---|---|---|
| CANCER | Reduces sphere-forming ability | Upregulates integrin β1 expression | [59] | |
| Targets cancer stem/progenitor cells | Inhibits stemness pathways | [60,61] | ||
| Decreases invasive capacity | Disrupts large single-cluster formations in multicellular spheroids | [62,63] | ||
| Inhibits cell migration and invasion | Enhances ECM adhesion | [64,65] | ||
| FIBROSIS | Reduces ECM production [collagen types I and III, elastin, hyaluronic acid] | Decreases COL1A1, COL3A1, elastin, and hyaluronic acid expression | [3,70] | |
| Suppresses fibroblast to myofibroblast transformation | Decreases α-SMA expression and inhibits TGF-β, PDGF-β, and SMAD-2 signaling | [3,45,71] | ||
| Promotes fibrosis reversion | Induces AMPK activation and phenotypic switch to lipo-fibroblasts via BMP-2 and PPAR-γ phosphorylation | [74,75] | ||
| Diabetes & Metabolic Diseases | Inhibits cell proliferation in intestinal organoids | AMPK activation and p53-dependent activation of REDD1 lead to mTOR inhibition and cell cycle arrest | [84,85] | |
| Protects pancreatic beta cells | Maintains cell viability under high glucose conditions and prevents fatty acid-induced apoptosis via AMPK-mediated autophagy | [86,87] | ||
| Impairs adipogenesis | Enhances stemness in adipose-derived stem cells via autophagy activation and mTOR inhibition | [88] | ||
| Reduces liver lipid content | Exhibits anti-steatotic properties by lowering fatty acid consumption | [91,92] | ||
Table 2.
Other application of metformin with ECM.
| Application | Effects of Metformin | Key Mechanisms | Ref. |
|---|---|---|---|
| Tissue Regeneration | Stimulates osteogenesis in mesenchymal stem cells (MSCs) | Enhances differentiation into osteoblasts, regulates immune responses (M1 to M2 macrophage shift) | [94] |
| Wound Healing | Enhances healing, increases angiogenesis, improves epithelialization, promotes hair follicle formation & collagen deposition | Immunomodulatory & anti-inflammatory effects | [95] |
| Bone Tissue Engineering | Improves osteogenic differentiation & bone formation | Calcium phosphate cement scaffolds & polylactic acid/polycaprolactone composites | [96] |
| Dental Tissue Engineering | Supports dental pulp cell viability & enhances odontogenic differentiation | Calcium phosphate cement-chitosan composites | [97] |
| Periodontal Regeneration | Enhances bone regeneration | Polycaprolactone/polyvinyl alcohol membranes with 10 wt% metformin | [98,99] |
| Cranio-Maxillo-Facial Defects | Improves clinical outcomes in bone defect treatments | Metformin-loaded scaffolds | [98] |
| Implant Integration | Enhances cell proliferation & metabolic activity of adipose-derived stem cells | Sol-gel coatings with metformin for metallic implants | [100,101] |
| Diabetic Wound Healing | Accelerates healing, reduces inflammation, improves dermis & epidermis regeneration | Composite scaffolds (chitosan/gelatin/polycaprolactone, polyvinyl pyrrolidone nanofibers) | [103] |
| Controlled Drug Delivery | Provides sustained metformin release, improves wound healing rates | Collagen/PLGA nanofibrous scaffold membranes | [103] |
| Cancer Therapy | Enhances drug penetration & anticancer effects | Lysozyme-functionalized metformin-loaded nanoparticles modifying ECM | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



