
Submitted:
15 April 2025
Posted:
15 April 2025
You are already at the latest version
Abstract
The temperate terrestrial Australasian orchid genus Caladenia has 181 taxa in Western Australia, including 23 endangered species. They have relatively specific pollinators and fungi, so are difficult to propagate and conserve. This project focuses on three threatened Caladenia species from isolated habitats in the Western Australian wheatbelt. Despite many attempts to optimise orchid propagation methods, inexact and inconsistent success criteria make comparisons of them difficult. Our aims were to measure the efficiency, complexity and effectiveness of propagation methods for rare and common Caladenia species, document their development in relatively natural conditions and produce robust plants for translocation. We utilised three orchid seed germination methods (1) in vitro asymbiotic synthetic agar media without fungi, (2) ex situ seed baiting over concentrated soil organic matter (OM), and (3) a novel method using permeable pouches in non-sterile substrates containing OM from orchid habitats (the FORGE system). Methods were compared by measuring growth rates and survival of protocorms and seedlings. There were major differences between methods and species. In vitro asymbiotic germination on Petri plates produced many protocorms, but only a few survived explanting. Protocorms from non-sterile methods were substantially more resilient in the incubator or greenhouse. The FORGE system was most efficient, as seedlings grew 20 times faster and could be moved to new pouches for further growth. Seed baiting protocorms also survived in pouches. Several stages of growth in pouches produced large seedlings with tuber initiation, suitable for outplanting that year. Our observations lead to refined definitions of growth stages and success criteria for orchid propagation. We recommend further development of FORGE systems for efficiently producing orchid seedlings pre-adjusted to growth in nature, especially for conservation. This system also has great potential for fungal isolation and morphological or physiological studies, based on orchid development in near-natural conditions.

Keywords:
orchid conservation
; Western Australia
; Caladenia
; orchid propagation
; seedling development
; soil seed baiting
; non-sterile germination
1. Introduction
The Orchidaceae is the largest family of angiosperms with over 30,000 species, and also has the most threatened species [1,2]. They occur in many habitats in most areas of the world, but the majority are tropical epiphytic species [3]. Among their characteristics are relatively specific symbiotic relationships with mycorrhizal fungi [4,5,6], which are required for germinating the dust-like seeds that lack a substantial endosperm [7,8]. In Australia, terrestrial orchid species are very susceptible to ecological change from environmental threats such as dryland salinity, hydrology, land clearing, invasive weeds and grazing, as well as accidental destruction, small population sizes and climatic extremes [9,10,11,12]. There are 44 endangered and 85 possibly threatened orchid taxa in Western Australia (florabase.dbca.wa.gov.au accessed 30-12-2024). Threats to rare orchids are most extreme in the wheatbelt of Western Australia due to extensive land clearing (80%), resulting in highly fragmented and degraded areas remaining.
There is a substantial body of research into optimising and developing propagation methods for rare plants, as part of efficient and effective conservation programs [13,14]. Conservation of Caladenia, like other terrestrial orchids, usually requires collecting seed for in vitro propagation, either symbiotically (with the mycorrhizal fungi) or asymbiotically [5,15,16,17] (Table 1). This is especially important for Caladenia species which are at high risk of extinction in the wild. Orchids are most often propagated in vitro (under sterile conditions) using complex culture media without a mycorrhizal fungus. These asymbiotic methods were first used by Knudson [18], and there now are a wide diversity of orchid seed germination media used for commercial orchid propagation and media often require optimisation for different species [17,19]. This can lead to consistent large-scale production of protocorms without the need to isolate and test fungal strains, so may be preferrable for rare and endangered species when seed is limited [5]. However, this can lead to high mortality rates of terrestrial orchids in the nursery, especially due to the lack of compatible symbiotic fungi [20,21].
The other standard in vitro method is to germinate orchid seeds symbiotically by including compatible fungi in culture media [22,23,24,25,26,27]. This method often produces similar germination rates than asymbiotic methods, but is usually faster (Table 1). Major problems can still arise when transferring the developing seedlings into the nursery [16,20,21,28,29,30]. A related method of orchid propagation utilises orchid seed and mycorrhizal fungi (which may be combined into alginate beads) placed in a growth substrate [31,32]. Seed sowing into natural habitats has also been successful, but is rarely utilised [33,34]. Finally, ex situ soil baiting, which was developed to test if compatible mycorrhizal fungi for orchid species are present in soil samples, was also suggested as a potential tool for terrestrial orchid propagation [35]. In situ baiting, where packets containing seed are placed in or on substrates, is also routinely used to detect or isolate compatible fungi, but living plants are rarely recovered [36,37,38]. Related methods utilising orchid seed in packets have been successful used to propagate epiphytic orchids [30,39]. Despite the time required, complexity and cost of orchid propagation methods, they usually are much more efficient than orchid reproduction in nature where only a few seeds out of the thousands or millions produced by a plant germinate successfully [7,40]. Orchid propagation studies summarised in Table 1 are primarily for conservation purposes, so focus on production of large seedlings with tubers for translocation. These data also primary concern Australian orchids, especially Caladenia species, but outcomes for terrestrial orchids from other regions were similar. Case studies of successful translocation of Caladenia are described by Reiter et al. [26], Janissen et al. [41] and Reiter and Menz [42].
The orchid genus of Caladenia (spider or fairy orchids) is endemic to the Australasian region and includes 181 species only found in southwestern Western Australia, including 23 threatened species (florabase.dbca.wa.gov.au accessed 30-12-2024). These are geophytic terrestrial orchids with a single hairy leaf and no true roots. This limits their nutrient uptake ability, so they are heavily reliant on relatively specific mycorrhizal fungi throughout their entire lifetime. Caladenia species also have extremely specialised relationships with pollinators, via sexual or visual deception [53,54]. Caladenia williamsiae (Williams Spider Orchid), C. melanema (Ballerina Orchid) and C. graniticola (Granite Spider Orchid) were selected for this study in consultation with a conservation agency staff, and common co-occurring orchid species were included for comparison. The overall objective of this study was to compare the relative effectiveness, efficiency and costs of seed-based propagation methods to produce new plants for conservation actions for three endangered Caladenia species. This required comparative propagation trials using the same orchid species and comparisons with published data. A secondary objective of propagation trails was to produce seedlings of orchids suitable for translocation. The third objective was to develop microcosms where symbiotic orchid seeding development could be studied under realistic conditions, avoiding artefacts of in vitro methods. This work is part the Wheatbelt Orchid Rescue Project which was funded from 2007 to 2011 and based on collaborations between scientists, community groups and a government department [55]. Volunteers played a very important role in this project, as explained elsewhere [56].
2. Materials and Methods
2.1 Study species
This project focused on three of the rarest threatened orchids found in the wheat-belt of WA, all of which are restricted to extremely small areas in the West Australian wheatbelt [56,57]. Caladenia melanema (Figure 1A) grows under eucalyptus and melaleuca tall shrubland on sandy soils, on the outskirts of salt lakes about 270 km SE of Perth. It grows 20 cm tall, with cream to pale yellow 5 cm wide flowers in August to September. Caladenia graniticola (Figure 1B) grows beneath tall shrubs and sheoaks (Allocasuarina huegeliana) on the margins of granite outcrops in a few locations in the southeast wheatbelt 270-300 km southeast of Perth. Their maroon and green flowers are 5 cm across, flowering in late September to October. Caladenia williamsiae (Figure 1C) grows under dense shrubs, in kwongan vegetation red-brown loamy lateritic gravel soil on ridges about 130 km southeast of Perth. It has greenish-yellow and red flowers 3 cm wide in August to early September.
2.2 Sources of seed and soils
Seed of rare and co-occurring common orchids required for seed baiting and propagation was obtained from wild populations [55,56]. Seed bags were placed on manually cross-pollinated flowers to ensure good quality seed was collected (Figure 2A). These were made of readily available, thin, tough, and highly flexible interfacing fabric that is, allows light transmission and survives exposure to the sun (non-woven polyester interfacing fabric). Squares of this fabric were wrapped several times around pollinated flowers or developing pods and gently fastened above and below them with twist ties (Figure 2BC). This allows seeds to be collected after reaching maturity and prevents grazing (Figure 2D). Seed was cleaned by sieving, died and stored at 4°C in vials. The same seed lots for each species of orchid were used for all propagation types, which also included several co-occurring common Caladenia species and Microtis media, which is a very widespread orchid (Table 2). Exact locations where soil and seed are collected cannot be provided, but the approximate locations used here are consistent with recovery plans for each species [55].
2.3 Seed germination on sterile media
Asymbiotic seed germination utilised pre-packaged medium ‘W3’ from Western Orchid Laboratories, a culture media routinely used for Australian orchids grown in the absence of mycorrhizal fungi (http://www.westernorchids.com.au/). The supplied dry ingredients and 60g of banana pulp were suspended in 1.1 L of deionised water, which was autoclaved before pouring into Petri dishes in a laminar flow hood.
Orchid seeds were surface sterilised in packets made from folding filter paper, closed using a staple, dipped into 70% methanol, and then left in 3% hydrogen peroxide for 10 minutes [28,58]. Seeds were transferred to the asymbiotic medium in the Petri dishes in sterile conditions, approximately 50-100 seeds per plate, and Petri dishes were sealed by wrapping a 5 cm wide strip of Gladwrap around the sides 2-3 times. Plates were stored in a tightly closing lidded plastic storage container that had been sterilised by spraying with methanol. Storage boxes were placed in a constant temperature room at 22˚C. Protocorms were measured from photographs as described below after 4, 6, 8 and 10 weeks (Figures 3, 4).
2.4 Orchid seed baiting
Ex situ seed baiting detects compatible mycorrhizal fungi in soils by germinating orchid seeds over soil collected from natural habitats and incubated under controlled conditions [35]. Living seeds of West Australian orchids expanded rapidly on exposure to water because they germinate during the first winter after seed is dispersed and lack a persistent soil seed bank [37]. Topsoil was collected from habitats of the 3 rare orchids, while avoiding impacts to them (Table 2). Soil sampled had a pH range of 5-6 and consisted primarily of fine sand with up to 10% silt and clay. Relatively coarse soil organic matter (OM) was separated from other components by dry sieving between a 6 mm and 2 mm mesh sieves and manually removing rocks and sticks.
Seed baiting plates were prepared by placing sieved soil OM in 100 x 100 x 10 mm square Petri plates, spread to about 7 mm depth, then covered with a sufficient sterile fine white sand to level the surface [35]. Plates were then wetted with deionised water and covered with a 95 x 95 mm square of 100 µm pore size nylon mesh (silk screen fabric). Plates were left for 48 hours before seed was added. Seed was uniformly sprinkled on 10 mm squares of 0.45 µm pore size nitro-cellulose filter as shown in Figure 3C (25-50 seeds per square). Each baiting plate received nine seed types on separate squares placed on top of the mesh (Figure 3D). Plates were kept in an air-conditioned room at 22˚C. Protocorms were measured 4, 6, 8 and 10 weeks after sowing from digital photographs, as described below. In total 170-200 protocorms were measured for orchids each time. Soils that germinated seeds successfully were used in subsequent trials or in new baiting plates. In total there were 43 soil samples, 8 orchid species and 10 seed lots. This method also measured seed viability by counting imbibed swollen seeds with ruptured seed coats - a pre-germination process that does not require fungi (Figure 3AB).
2.5 Seed germination in non-sterile conditions
A novel non-sterile orchid propagation method based on ex situ seed baiting was developed for this study [59]. This method (Fungal Orchid Realistic Germination Environments, FORGE) utilised seed packets placed in a standard potting mix used for Australian terrestrial orchids that incorporates soil OM known to contain compatible fungi (Figure 7). The 100 x 30 mm seed packets were made by sandwiching 100 µm nylon mesh (or thin interfacing fabric) and clear plastic (from a thick plastic bag), joined by heat sealing the bottom. Seed packets were then placed in plastic 1000 ml (650 x120 x 70 mm) rectangular heat-resistant Polypropylene food containers (Figure 4E). These containers contained approx. 500 ml of autoclaved growth substrate mixed with 20% soil OM with active compatible fungus from baiting plates as inoculum. This substrate consisted of 50% autoclaved finely chopped sheoak branchlets and 50% coarse sand, fine perlite or finely chopped coir (coconut peat) to provide aeration and stability (Figure 4). Orchid growth substrates based on fallen sheoak branchlets (Allocasuarina or Casuarina spp.), which resemble conifer needles, are commonly used to grow orchids in Australia. These can be substituted by other types of litter that is relatively resistant to decomposition. Orchid seed was sprinkled inside dampened seed pouches (folded open) or onto a single layer of nylon mesh which were then placed along the edges of containers (Figure 4E). Containers were placed in an incubator (20˚C day, 15˚C night, 12 hours light per day). Protocorms were counted, measured and photographed after 4, 6, 8 and 10 weeks.
2.6 Continued growth and translocation of seedlings
Seedlings from the soil baiting and asymbiotic experiments were transplanted after developing a substantial leaf (Figure 5C, Table 3). Seedlings from seed baiting plates were also transplanted into 100 x 30 mm pouches, lined up and spaced 10 mm apart (Figure 5A). Seedlings with a small leaf (stage 3 or 4) could be picked up with fine forceps and arranged in a row on the dampened fabric of growing pouches (Figure 5A). Each permeable pouch had 10 seedlings arranged 10 mm apart (Figure 5). These pouches were placed in rows in containers filled with the standard growth substrate mixed with 10-20% inoculum (OM from a plate or container were seeds geminated), as explained above. Pouches were angled at 45° with mesh side down and buried so only leaves were exposed to light and placed on the side of containers (6 pouches with up to 60 seedlings per container). Once seedlings had leaves 5-10 mm long (Figure 5), they were transplanted into new pouches and fresh growth media for further growth in a growth cabinet until they reach sufficient size for establishment in soil or potting mix for tuber formation.
Seedlings were transplanted into the greenhouse (23˚C day, 8˚C night) once reaching stage 5 (Figure 5). Eight 200 mm diameter plastic plant-pots were filled with soil collected from sites were orchids originated, with 5-10% OM inoculum from containers mixed in as inoculum. Window pots with a 100 x 100 mm section cut out and a 120 x 120 mm clear plastic insert glued in place were used (Figure 6). This allowed dropper growth and tuber formation to be easily observed in the glasshouse. Window pots were placed in an intact pot the same size to exclude light. Orchid seedlings were placed in a 100 x 100 mm translocation pouches made of interfacing material and clear plastic sealed on sides and bottom as described above (Figure 6C). There was one pouch per pot (facing the window) and 5 seedlings per pouch, placed so leaves extended above the soil surface. Interfacing fabric covered pots to increase humidity levels and seedlings watered using deionised water. Leaf and dropper lengths were measured weekly.
As a small research translocation trial, seedlings of 6 Caladenia species were transplanted into three field locations close to where these rare orchids occurred along with a common Caladenia species. These orchids were C. williamsiae along with C. falcata, C. melanema with C. flava and C. graniticola with C. roei. They were transplanted into 100 x 100 mm pouches, as in the greenhouse. Five seedlings were in each pouch, and between 1 and 2 pouches were planted at each location, depending on the number of available seedlings (Figure S3). Pouches were planted with approximately 30 ml of inoculated growth media from incubator containers (containing soil organisms from the same site where translocation occurred). One m tall chicken wire cages were secured to the ground over seedlings (Figure S3) for grazing protection (kangaroos and rabbits).
2.7 Measurements and data analysis
In this study, the relative efficiency of orchid propagation methods was compared by determining the number of sufficiently large orchid seedlings produced per container (Petri dish or germination pouch) and the time required for them to grow. Orchid seed germination rates per seed were not determined because they were strongly influenced by resource competition due to seedling density [60] and it was not possible to precisely regulate sowing densities. Protocorm width was measured once trichomes were present, using calibrated images taken with a digital microscope camera and the measurement tools in Adobe Photoshop CS3 ExtendedTM. Measurements switched from protocorm size to leaf length as seedlings grew, because protocorm growth slowed as leaves developed.
Graphs of protocorm, or leaf sizes over time were created in Microsoft Excel and a trend line used to calculate average growth rates, and 95% confidence intervals were used to compare means. Differences in protocorm sizes between species and time were analysed using Permanova in Primer, using square root transformation and Bray-Curtis similarity [61].
3. Results
3.1 Asymbiotic sterile germination
This method was very laborious, complex and required access to an autoclave, laminar flow hoods, and incubators (see Section 6 below). Orchid seed germination was also slow and highly variable, requiring only 4 weeks to reach trichome stage for C. williamsiae, but over 6 months for C. falcata and C. roei, where some plates failed to germinate, and others had very few protocorms (Figure 7). Protocorm growth rates (Figures 8A, S2A) varied within a species (e.g. C. williamsiae 0.034 to 0.077 mm/week) and between species (e.g. C. melanema 0.028 vs. C. williamsiae, 0.077 mm/week). The number of protocorms produced also varied between species, from C. falcata with only 9 protocorms, to C. williamsiae with 129 protocorms (Table 3). Protocorms were very small after 10 weeks (averaging 0.46 mm diameter), with an average of 37 per surviving plate. Overall, approximately 50% of all plates were lost to contamination. Figure 7 illustrates protocorm development C. williamsiae, but other species were substantially smaller at the same age.
3.2 Ex situ baiting
Living seeds swelled substantially when first exposed to water, rupturing the seed coat (Figure 3B). This allows seed viability to be efficiently measured from the proportion of swollen seed on baiting plates (all over 90%). All germination on baiting plates occurred within 4-8 weeks, but there was substantial variability in germination between soil samples (Figures 8B, 9). This experiment (30 soils, 10 orchids) resulted in 945 protocorms, averaging 22 per square, with an average of three squares with protocorms per plate. Protocorm growth rates varied substantially both within and between species (Figures 9B, S2B). For example, growth of C. graniticola varied from 0.037 to 0.094 mm/week and C. melanema grew slower (0.022 mm/week) than C. graniticola (0.094 mm/week). By week 9 growth had slowed or stopped and protocorm health started to decline. The average they were 0.38 mm in diameter, with 66 per plate. (Table 3). Evidence for strong competition between protocorms in the same container was also observed (Figure 9C). The number of substantial protocorms produced varied between species, although not as much as with the asymbiotic method described below (Table 3). For example, C. falcata only produced 28 protocorms, while C. latifolia produced 94. One species did not germinate at all in this experiment (C. dimidia). Overall, C. latifolia produced the most protocorms, and C. graniticola had the fastest growth on ex situ baiting plates (Figure 8B).
Baiting trials with C. melanema, C. williamsiae and C. graniticola demonstrated that mycorrhizal fungi capable of germinating seed were present in some soil samples from suitable looking habitats, but not others (Figure 9A). The results of three seed baiting experiments were similar, so only one is presented in Figure 9A. Rare orchids germinated in fewer soil samples than the common orchid C. flava or the weedy orchid Microtis media (Figure 9A). Thus, it seems that fungi compatible with rare orchids were often less common than those from common orchids, even in soils collected where rare orchids occur. Ex situ baiting identified OM samples containing inoculum of compatible fungi for all orchids except C. williamsiae. These samples were used to provide compatible fungi for orchid propagation, as explained below. Squares of seed that failed to germinate could be moved to other plates or containers, as they remined alive in a pregerminated state for several months.
3.3 Symbiotic germination in non-sterile conditions
We developed a new germination pouch system (FORGE) which allowed orchid seed to be incubated in a non-sterile environment using natural inoculum sources (Figure 4). Pouches enclosed germinating orchid seeds between transparent plastic and permeable mesh layers placed in contact with organic matter containing highly active mycorrhizal fungi, identified as explained above. Seed germination commenced within 4 weeks in the FORGE system, which was relatively easy to set up but required careful management (see below). This system allowed accurate measurement of orchid growth without disturbance (Figure 9B), clearly revealing developmental stages (Figure 4). Growth data was based primarily on leaf lengths, once protocorm growth slowed (Figure 10). Seedlings growth was exponentially rather than linear, and mor than an order of magnitude faster than other germination methods (compare y axis scales in Figures 8 to Figure 10). On average seedlings were 7.5 mm long after 10 weeks, and 54 were produced in a container with two pouches (Table 3). Some of these seedlings were large enough for transfer to pots, but most were moved to growing pouches (see below). Most Caladenia seeding grew about 1.4 mm per week in length, but one orchid (C. roei) germinated earlier and grew much faster than the others, reaching 5 mm long in 6 weeks (Figure 10B).
Caladenia roei also produced the most seedlings, with up to 72 in a pouch (Figure 4). We also found that packets could be returned to containers after larger seedlings were transplanted, so smaller seedlings continued to grow and harvested later. Overall, most large seedlings transplanted in to growing containers in the incubator (226) were produced by the FORGE method (Table 3). It was noted that seedling growth and health tended to decline if they were left in pouches for too long, presumably due to competition for declining resources and structural collapse of growing media. This required careful observation of seedlings so they could be moved to new containers to maintain rapid growth, as explained below (e.g. after 6-10 weeks). It was also important to avoid excess moisture in containers, keep them relatively cool, and inspect for pest, especially fungus gnats, regularly.
3.4 Seedlings in the incubator
A modification of the germination pouch system was used to protect and track growth of orchid seedlings, as shown in Figure 5. After reaching a suitable size (leaves 2-10 mm), protocorms were transplanted into new pouches placed in potting mix inoculated with mycorrhizal fungi along with a portion of their previous culture substrate (Figure 5). Growth in the incubator was relatively uniform across all species, but sizes differed considerably at the start (Figures 10C, S3). The fastest leaf growth was seen in C. graniticola and C. roei, and slowest in C. williamsiae. Droppers began to form on C. graniticola (14), C. roei (13), C. williamsiae (3) and C. falcata (4), but most were transplanted into the greenhouse or field before this happened (Caladenia species do not form roots).
Transplanting asymbiotic protocorms into growth pouches in the incubator with a substrate lacking compatible fungal inoculum resulted in mortality for all species except C. williamsiae, where very slow growth resulted and only 28 seedlings survived for use in the greenhouse and field (Table 3). Seedlings transplanted from soil baiting plates with some substrate inoculum survived transfer to pouches better, but with variable results. Of these, C. graniticola, C. latifolia and C. flava survived the best (Table 3). Seedlings originating from seed pouches transplanted into growing pouches with some original substrate had the fastest growth (Figure 9C), were largest overall and most successful when transplanted into the greenhouse or field (Table 3).
3.5 Seedlings in the glasshouse or field
Permeable translocation pouches in window pots allowed seedling growth and tuber formation to be observed and measured non-destructively (Figure 6). These pouches also helped to protect seedlings in the field and glasshouse, presumably by reducing moisture loss and interactions with soil animals. However, further work is required to optimize methods for their use in translocation trails. Greenhouse leaf growth largely reflected incubator growth and survival, with C. melanema having consistently slow leaf growth and droppers in only one pot. Caladenia graniticola had fast leaf growth in some pots, but poor dropper growth (Figure S2D). C. williamsiae (from asymbiotic culture) had the smallest seedlings at the start and leaves did not grow in the greenhouse, but one dropper extended (Figure S2F). Caladenia roei had the most consistent growth of leaves and droppers overall (Figure S2H). Dropper growth varied from 0.07 to 0.7 mm per day (Figure S2). It is likely that the mixed results we observed in the glasshouse were due to variations in the quality of soils used to grow plants, since they absorbed water poorly and became very hard when dry. Mixing soil with orchid growing substrate may have resolved these issues.
As a small translocation trial, seedlings of six Caladenia species were transplanted into three field location close to habitats where the rare orchids grew, in combination with one common species (C. williamsiae with C. falcata, C. melanema with C. flava and C. graniticola with C. roei). One month after transplanting, the C. graniticola and C. roei were the only species with visible leaves at two sites, due to a lack of rain and infrequent watering [56]. These also were the largest seedlings transplanted, with leaves 10 - 25 mm long. A few leaves of C. melanema emerged in the same cage the following year (Figure S3D). Ongoing monitoring of these trials was not possible, and larger trials are required.
3.6 Comparison of methods
We observed a high degree of variation between species for each seed germination method, but the asymbiotic in vitro method was much slower and more erratic than other methods overall (Table 4). There were no visible impacts of sterilisation on orchids seeds or batches with low viability observed in any method. We also found the use of Petri plates impractical due to contamination risks, but use of flasks would be even more difficult and expensive (Table S2). Non-sterile symbiotic germination in seed packets was the most rapid and efficient method in our trials, resulting in hundreds of large seedlings, but results varied between species. Another key advantage was that seedlings could be harvested from the same pouches once the first lot of protocorms had been removed. Orchid seed germination rates in seed baiting plates or pouches using the same inoculum source were similar, presumably due to the identity and activity of symbiotic fungi (Table 4).
Statistical analysis of the protocorm size between asymbiotic germination and seed baiting found significant differences between orchid species and times but not methods (Table S1). A significant time x size interaction suggest growth rates were different. The FORGE method was not included in this analysis because sizes of protocorms and seedlings were measured differently (protocorm vs. leaf length). However, seedlings from the FORGE method were 20x larger after 10 weeks (9.25 ± 2.4 vs. 0.33 ± 0.047, average mm ± 95% CI).
Most orchids grew faster in symbiotic, non-sterile systems (up to 1.4 mm/week for C. roei), compared to axenic asymbiotic media (up to 0.03 mm/week for C. williamsiae). For all propagation methods, rare Caladenia species were not harder to propagate than common species, except for C. williamsiae where compatible fungi were not detected. Very widespread (C. flava) or weedy (M. media) orchids germinated more often than rare orchids (Figure 4A), as expected from fungal specificity studies [62,63]. An unexpected finding was that the same orchid which failed to germinate in non-sterile systems (C. williamsiae) was most productive in axenic asymbiotic culture. This may be related to higher fungal specificity, scarcity of fungal inoculum, or genetic issues such as inbreeding (this species has a very restricted range under 2 ha [55]). Additional work is required to resolve propagation of this species which is one of the most threatened orchids globally.
We found that comparisons between published orchid germination data were difficult because germination stages were not consistently defined, and data on protocorm size relative to age was not provided. As shown in Table 1, germination is most often defined by stage 3 or 4 (leaf primordia or leaf), but some studies used stage 2 (trichomes), which can be misleading since some fungi only germinate orchid seed to Stage 2 [62,64,65]. Initial germination can be followed by protocorm death due to hypersensitivity (fungal suppression) or transition to parasitism by the fungus [64]. It has been recommended that “compatible fungi be designated as those that support the orchid host to an advanced seedling stage where the leaf exceeds the protocorm in length”, because smaller seedlings were unlikely to survive transplantation [62].
Comparisons of studies including 7 asymbiotic and 12 symbiotic trails that included 69 terrestrial orchid species revealed that symbiotic germination was twice as fast as asymbiotic sterile culture on average (Figure 11A). Thus, protocorm growth rates were more important than germination rates, because all methods had adequate germination. Inclusion of our data and other studies of early orchid growth in non-sterile systems, revealed that seedling growth is substantially faster in semi-natural than in all axenic culture methods (Figure 11B). The latter also tend to be more expensive and time consuming (Table S2, Figure 11).
4. Discussion
A wide diversity of methods media types which are routinely used for non-symbiotic seed germination and tissue culture propagation of orchids [19]. Here we were only tested one axenic method due to time and capacity limitations, but the relative efficiency of other methods was compared using published data, especially from Australia for Caladenia species (Figure 11). Financial cost for these methods were also estimated (Table S2). Costs for materials are relatively low, but expenses from specialised equipment (such as an autoclave and laminar flow unit) and specialised labour are much higher for in vitro methods (Tables 4, S2). The main cost for each method is attributed to labour, due to time taken for set-up, maintenance and waiting for adequately sized seedlings to grow.
It has been suggested that Ex vitro methods use more seed than in vitro symbiotic methods, so are less appropriate for endangered taxa with scarce seed resources [5,66]. However, most earlier studies did not include non-sterile methods which we found to be the most efficient way of propagating endangered orchids, due to more rapid seedling growth and higher rates of survival. Approximately 50% of seed germinated in the FORGE packets, which was similar to the asymbiotic method, but most seedlings that survived were from the former method. In contrast, C. williamsiae, only produced protocorms using the asymbiotic method, some of which survived explanting but were extremely slow to grow. Caladenia species can be difficult to grow or translocate successfully [5,28,67] so it is extremely important to use the most successful propagation methods available [68], as explained for each method below.
4.1 In vitro seed germination
We found that the asymbiotic sterile method produced many protocorms, but had the most erratic rates of germination. It also had the lowest transplant survival rate, as is often the case for these methods [29,45,69]. Other studies also reported germination in asymbiotic methods to be extremely slow (Table 1), but growth can also be rapid [70]. These differences may be due to use of complex media formulations, which may need to be tailored for individual orchids [17]. Contamination due to high levels of nutrients and sugars is also a major problem for asymbiotic media, especially in Petri plates [28,29] and survival of seedlings after culture can be very low [21]. Similar issues also occur with in vitro symbiotic culture, but germination was generally faster and seedlings larger than to asymbiotic sterile seed germination in earlier studies with Caladenia species (e.g. Clements et al. [24]; Chou & Chang [49]; Wright et al. [69] and Bustam et al. [16]). Batty et al. [28] developed a sand over agar container system which allows seedlings to acclimate more successfully after axenic culture.
4.2 Non-sterile seed germination
A single Ex situ seed baiting experiment using 29 soil samples efficiently detected compatible fungi for all but one orchid species in soils sourced from their habitats. This experiment used concentrated organic matter, which functions as a reservoir of fungi inoculum [35]. We also used ex situ baiting to measure protocorm growth rates in a relatively natural situation, and successfully transplanted protocorms into containers for further growth. Other key outcomes were identification of soil samples with active compatible fungi for propagation trials and identification of suitable sites for translocation of orchids. Ex situ seed baiting has also been used to test seed viability and isolate fungi from protocorms [38,63,71,72,73,74]. However, adequate replication is required due to natural variations in inoculum levels of mycorrhizal fungi in soils (e.g. Batty et al. [37,75]; Feuerherdt et al. [76]; Diez [77]; Jacquemyn et al. [78]).
The new FORGE method introduced here (seed pouches incubated in non-sterile conditions) is worthy of further development for propagation of rare and common orchids. We used this system directly for seedling production and indirectly to acclimate seedlings produced by other methods. The FORGE method resulted in seedlings that were much larger and survived better after transplanting. These included all the seedlings transplanted into the greenhouse or field, except for C. williamsiae where highly compatible fungi were not recovered. The FORGE method is an extension of ex situ seed baiting, so had similar germination results. Germination occurred after 4-8 weeks in both systems, but seedling growth was more rapid and consistent in larger FORGE containers, probably due to reduced crowding. Declining growth of seedlings also eventually occurred in FORGE germination pouches, due to resource competition and declining growing conditions, but was remedied by transferring seedlings into new containers with fresh media. Orchid seedling growth in growing pouches continued at a steady rate for 1-2 months until droppers were initiated. At this stage seedlings required transfer to larger or less-crowded containers (or pots) for tuber formation. Thus, the FORGE system requires a series of growing stages where seedlings are moved to new containers to maintain rapid growth, as shown in Figure 12.
It was noted that seedling growth rates varied considerably between species and inoculum sources in both seed baiting plates and germination pouches (e.g. exceptionally rapid growth of C. roei). This probably results from variations in fungal inoculum levels and orchid-fungus compatibility. This suggests that additional trials to select effective fungal strains would result in more rapid growth of a wider diversity of orchids. A key advantage of the FORGE system is that it avoids the need to isolate fungi, identify effective isolates for long-term seedling growth, or periodically re-isolate fungi due to declining compatibility in culture [79,80]. Inoculation orchids in the FORGE system also has the potential to revitalise fungal isolates which have lost effectiveness in axenic culture media. Mycorrhizal fungus diversity and identity in our germination trials is unknown, but consist of organisms from the same habitats as orchids, thus avoiding transferring non-indigenous organisms with translocated plants. Fungi used in FORGE systems could also be easily stored as dried growth substrates.
The FORGE system is similar to other methods used to successfully propagate orchids, such as non-sterile symbiotic seed germination of tropical epiphytic orchids using packets placed in situ [81,82]. Other similar methods utilise seed on sterile potting mix inoculated with fungi from culture, or sowing seeds around the base of an established plant [45,46,69,75,83,84], or alginate beads containing both orchid seed and mycorrhizal fungi [31,50]. In general, non-sterile symbiotic methods can produce variable results, but in our study were the most efficient and successful overall (Table 3).
Growth of orchid species in the FORGE system could be extended by transferring them to new growing containers or pots in the glasshouse along with small amounts of growing media as fungal inoculum. These seedlings had to be transferred several times before they were large enough for translocation, due to declining growing conditions in containers and increased competition as seedings grow. We also noted that the rate of leaf growth declined when droppers were initiated due to resource competition within seedlings, as also observed by Batty et al. [75]. As an extension of the FORGE system, we also trialled the use of larger pouches of the same materials to aid survival of seedlings in the glasshouse or field. This provided some protection form grazing and increase desiccation tolerance, but most seedlings were still lost to extreme drought conditions in the field.
The FORGE system and window pots allowed us to observe orchid seedling development in relatively natural conditions from germination to tuber formation. Protocorm or seedling development is also less affected by competition for space or depleted resources than in closed culture systems, because they can be transferred to new pouches when required (Figure 12). Thus, this approach provides more realistic data on orchid seed germination and growth dynamics and allows developmental stages to be observed easily without the abnormalities often present in sterile systems. FORGE is a highly flexible system which can be adapted for other methods of seed germination, fungal isolation, providing material for scientific experiments, etc. Experiments can occur in semi-natural conditions where nutrients and soil biota can be manipulated or managed (e.g. Schweiger et al. [85]; Mehra et al. [86]). The FORGE system is also ideal for studying beneficial or hostile interactions with soil organisms (helper bacteria, antagonists, biocontrol of soil animals feeding on fungi or protocorms, etc.), and for genomic studies of expressed or supressed genes (expressome, epigenetics) by providing material of exact growth stages growing in realistic conditions.
A common theme in orchid production trials is gradual attrition in seedling numbers across all stages of production (Table S2). Addressing this issue is more important than increasing rates of seed germination, but most research focuses on the latter. The orchid propagation system presented here allows more efficient observations and diagnosis of causes of seedling loss during intermediate and later growth stages. However, further research is required to optimise methods for transplanting seedlings to the glasshouse or field, where survival and growth was lower.
4.3 Germination and development of orchids in semi-natural conditions
According to Yeung & Lee [87], experts disagree about the nature of the protocorm, designating it either as a seedling-like or embryo-like structure. It should be noted that protocorms are not equivalent to seedlings, because the latter includes a protocorm as well as a leaf, root, etc. It could also be argued that all stages are seedlings, even though the first stages are more like an embryo. We recommend that substantial leaf formation be used to define the protocorm-seedling transition, because the protocorm gradually declines in importance after that. However, some orchids do not have leaves, so presence of the first substantial organ characteristic of adult orchid should be used (e.g. protocorm to rhizome transition in myco-heterotrophic orchids).
Protocorms are radially symmetrical before cell division and enlargement leads to more complex shapes that vary considerably between orchid species [87,88]. Morphologists generally recognise three main stages in early orchid development from protocorm, to pre-seedling (shoot initiation), to seedling (root initiation). Here the pre-seedling stage is referred to as a seedling, since root initiation did not occur. This stage also represents a key metabolic transition when leaves become major sources of energy and sinks for nutrients. We also recognise two other stages that are major structural and functional transitions - advanced protocorms and advanced seedlings (Table 5). This results in five discrete transition stages from seed to adult plants, which are similar to those used in germination studies (Table 1). These stages are rigorously defined in Table 5, and their functional consequences listed.
We observed that protocorm growth slows after leaf initiation, followed by an exponential leaf growth phase, then leaf growth slows or stops when droppers form (Figure S2). Allocation of resources to shoots normally slows or stops when tuber formation begins, and would normally also slow due to root formation [89]. Thus, seedling size had to be measured in different ways for different stages (e.g. protocorm length, leaf length, dropper length, then tuber size). Overall length (protocorm plus leaf, plus dropper) would also be effective for long-term studies. Caladenia protocorms illustrated here (Figures 3, 7. 8) seem to be simpler in structure than other terrestrial orchids studied, due to the lack of roots, scales or a cotyledon-like structures observed in temperate terrestrial orchids [87,90]. Protocorms are quickly replaced by roots and shoots in most orchids, but not in Caladenia seedlings, where they are essential during the first year, functioning like the mycorrhizal stem collar in adult plants. Another feature of Caladenia and other similar Diuridae orchids is that tuber formation starts with a dropper produced laterally from the protocorm and remains connected to the leaf that way (Figure 5E).
5. Conclusions
Orchid propagation using symbiotic fungi from soil organic matter as inoculum in a non-sterile environment has now shown to be an effective means of orchid propagation, but is not in common use [39]. We developed the FORGE system, an efficient and effective container-based, non-sterile propagation system and tested it on rare and common species of Caladenia (the largest Australian orchid genus). We also compared results with a sterile germination method commonly used in Australia, using the same orchid species and seed batches. We found that most orchids germinated and grew faster in the FORGE system than in sterile culture, and produced more robust seedlings. This shows there can be substantial advantages compared to sterile germination systems, especially for later stages of seedling growth, as listed in Box 1. However, additional research is required to optimise these new methods (Box 1). Orchid propagation and other conservation actions requires a better understanding of the role of symbiotic fungi, especially relationships with other soil organisms and key resources that sustain them. The FORGE system produced seedlings with normal growth rates, morphology and physiology, providing samples for scientific research lacking artifacts from sterile systems. It also allowed detailed non-destructive observations of all stages of protocorm and seedling development, leading to revised definitions of germination stages and recommendations for designating germination success in orchids (Table 5).
The FORGE system is a diverse system where different components will be relevant, based on the nature of orchids and their circumstances (Figure 12). The main use scenario is likely to be orchid conservation, via rapid seedling production for translocation in combination with identification of target sites with compatible fungi via ex situ seed baiting. This is especially relevant in situations where lab-based methods are too expensive or impractical for use. We found seedlings growth in a “realistic environment” was 20 times faster than in axenic systems, and they reached a suitable size for explanting in a nursery after 6 - 10 weeks. Other advantages include pre-adjustment to growth in non-sterile soils, production of mycorrhizal inoculum along with orchids, and use of locally indigenous fungi. The FORGE system could also considerably reduce seedling costs for horticulture, but requires further research to test applicability for diverse orchid types.
Box 1. Comparing orchid propagation methods
A. Advantages of non-sterile germination methods
- 1.
- Ex situ orchid seed baiting is normally used to determine if inoculum of compatible fungi is present in soils, but also measures seed viability (by counting imbibed seeds with coats ruptured by embryo enlargement). This method efficiently detected soil samples and fractions that contained fungus inoculum compatible with specific orchids, usually on the first attempt.
- 2.
- Non-sterile orchid seed germination utilising fungi present in soil organic material was a comparatively efficient and rapid method for orchid propagation (FORGE). The equipment and supplies required are readily available, inexpensive and containers can be reused many times. This method avoids the need for complex and expensive laboratory equipment and associated training.
- 3.
- The pouch system allows transplantation of seedlings along with substrate colonised by fungi into new containers at an optimum stage for further growth so rapid growth continues (Figures 5, 6). It is also possible to leave smaller protocorms for further growth. We were also transferred protocorms from in vitro culture or soil baiting into pouches, but these were less robust (Figure 5).
- 4.
- Regular observation of seeding growth allows intervention when growth slows, or pests appear (e.g. fungus gnats, nematodes, or slime moulds). Action can then be taken to address these issues (e.g. changing growing conditions, relocating seedlings, or application of control agents).
- 5.
- Orchid germination and growth in the FORGE system follows a normal sequence of development, in contrast to in vitro systems where seedlings tend to be abnormal. Developmental stages that are normally invisible in the soil can be studied under relatively natural conditions and easily photographed in plastic pouches without disturbance using inexpensive portable microscopes, phones or cameras. Transplanting seedings into window pots (Figure 6) allows observations to continue. Continuous observation of seedling development is also ideal for research on their development or physiology.
- 6.
- The use of natural inoculum sources containing indigenous fungi from orchid habitats should result in more robust seedlings for translocation and avoids introduction of non-local fungi.
- 7.
- 8.
- Large seedlings were available for translocation much sooner than in other propagation systems (the same year). Seedlings also survive better and grow more rapidly than those from in vitro methods, presumably because they were pre-adjusted to soil conditions.
B. Challenges for development of large-scale non-sterile germination
This approach requires ex situ seed baiting to identify soils which contain inoculum of compatible fungi (we found suitable soils for most orchids on the first attempt), or another effective source of fungal inoculum.
- 9.
- Management of seedlings in FORGE microcosms requires inspections several times a week and occasional additions of small amounts of water.
- 10.
- Maintaining suitable temperature and substrate moisture is important to avoid over-abundance of harmful soil animals, slime moulds, etc.
- 11.
- Predators of orchid seeds and fungi may be present, so must be monitored and controlled (e.g. fungus gnats, nematodes, slime moulds, etc.). Mites are present, but have limited impacts, unlike sterile culture systems where they are a major source of contamination. Larger soil animals such as snails and millipedes can be manually removed.
- 12.
- Organic substrates supporting orchid growth eventually become depleted, collapsed or soggy, but can be augmented, or seedlings transplanted into a mixture of new and old substrate.
- 13.
- Living soil systems should be isolated from sterile culture facilities to avoid spread of harmful soil organisms.
- 14.
- The FORGE system is relatively new and requires further optimisation to increase consistency and efficiency for a wider diversity of orchid genera. Additional research is required to test substrates, growing conditions, plant density, possible nutrient supplements, management of soil animals, fungal diversity and the role of plant genetics in germination responses. However, this optimisation is unlikely to be more arduous than what is required for successful outcomes from sterile culture methods, which produce seedlings that must also survive in non-sterile environments after explanting.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgements
This paper includes results of a 4th year since project in Biological Sciences at the University of Western Australia by EA supervised by MB in 2009. Funding was provided by Lotterywest to the West Australian Native Orchid Study and Conservation Group, coordinated by Gerald Stack. The Friends of Kings Park Orchid Carers Group and other volunteers had a major role in propagating orchids, especially Lyn Rowland, Lyn Roberts, Val Preston, Phylis Robertson, Nur Koshkuson and Mary-Ann Andrews). Staff of the West Australian Department of Conservation, especially Andrew Brown, Erica Shedly and Marie Edgley, contributed substantially to fieldwork. The University of Western Australia provided facilities for orchid propagation. Valuable assistance was provided by Gary Cass, Hai Ngo, Rob Creasy, Dr Pauline Grierson, Tim Morald, Jeremy Bougoure, Margaret Collins and Tammy Edmonds.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vitt, P.; Taylor, A.; Rakosy, D.; Kreft, H.; Meyer, A.; Weigelt, P.; Knight, T.M. Global conservation prioritization for the Orchidaceae. Scientific reports 2023, 13, 6718.
- Freudenstein, J.V. Orchid phylogenetics and evolution: history, current status and prospects. Annals of Botany 2024, mcae202.
- Givnish, T.J.; Zuluaga, A.; Marques, I.; Lam, V.K.; Gomez, M.S.; Iles, W.J.; Ames, M.; Spalink, D.; Moeller, J.R.; Briggs, B.G. Phylogenomics and historical biogeography of the monocot order Liliales: out of Australia and through Antarctica. Cladistics 2016, 32, 581–605.
- Rasmussen, H.N.; Rasmussen, F.N. Orchid mycorrhiza: Implications of a mycophagous life style. Oikos 2009, 118, 334–345. [CrossRef]
- Wright, M.; Cross, R.; Cousens, R.; May, T.; McLean, C. Taxonomic and functional characterisation of fungi from the Sebacina vermifera complex from common and rare orchids in the genus Caladenia. Mycorrhiza 2010, 20, 375,390.
- Oktalira, F.T.; Whitehead, M.R.; Linde, C.C. Mycorrhizal specificity in widespread and narrow-range distributed Caladenia orchid species. Fungal Ecology 2019, 42, 100869.
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. The New Phytologist 2000, 145, 367–421.
- Brundrett, M.C. Scientific approaches to Australian temperate terrestrial orchid conservation. Australian Journal of Botany 2007, 55, 293–307. [CrossRef]
- Phillips, R.D.; Brown, A.P.; Dixon, K.W.; Hopper, S.D. Orchid biogeography and factors associated with rarity in a biodiversity hotspot, the southwest Australian floristic region. Journal of Biogeography 2011, 38, 487–501.
- Gale, S.W.; Fischer, G.A.; Cribb, P.J.; Fay, M.F. Orchid conservation: bridging the gap between science and practice. Botanical Journal of the Linnean Society 2018, 186, 425–434.
- Wraith, J.; Pickering, C. A continental scale analysis of threats to orchids. Biological conservation 2019, 234, 7–17.
- Wraith, J.; Pickering, C. Tourism and recreation a global threat to orchids. Biodiversity and Conservation 2017, 26, 3407–3420.
- Dillon, R.; Monks, L.; Coates, D. Establishment success and persistence of threatened plant translocations in south west Western Australia: An experimental approach. Australian Journal of Botany 2018, 66, 338–346.
- Zimmer, H.C.; Auld, T.D.; Cuneo, P.; Offord, C.A.; Commander, L.E. Conservation translocation–an increasingly viable option for managing threatened plant species. Australian Journal of Botany 2020, 67, 501–509.
- Dowling, N.; Jusaitis, M. Asymbiotic in vitro germination and seed quality assessment of australian terrestrial orchids. Australian Journal of Botany 2012, 60, 592–601.
- Bustam, B.M.; Dixon, K.W.; Bunn, E. In vitro propagation of temperate australian terrestrial orchids: Revisiting Asymbiotic Compared with Symbiotic Germination. Botanical Journal of the Linnean Society 2014, 176, 556–566.
- Jolman, D.; Batalla, M.I.; Hungerford, A.; Norwood, P.; Tait, N.; Wallace, L.E. The challenges of growing orchids from seeds for conservation: An assessment of asymbiotic techniques. Appl Plant Sci 2022, 10, e11496. [CrossRef]
- Knudson, L. Nonsymbiotic germination of orchid seeds. Botanical Gazette 1922, 73, 1–25. [CrossRef]
- Arditti, J. Micropropagation of Orchids; John Wiley & Sons: Malden, MA, USA, 2009;
- Huynh, T.T.; Thomson, R.; Mclean, C.B.; Lawrie, A.C. Functional and genetic diversity of mycorrhizal fungi from single plants of Caladenia formosa (Orchidaceae). Annals of Botany 2009, 104, 757–765.
- Freestone, M.; Linde, C.; Swarts, N.; Reiter, N. Asymbiotic germination of Prasophyllum (Orchidaceae) requires low mineral concentration. Australian Journal of Botany 2023, 71, 67–78.
- Hadley, G. Cellulose as a carbon source for orchid mycorrhiza. New Phytologist 1969, 68, 933–939. [CrossRef]
- Warcup, J.H. Symbiotic Germination of Some Australian Terrestrial Orchids. New Phytologist 1973, 72, 387–392.
- Clements, M.A.; Muir, H.; Cribb, P.J. A preliminary report on the symbiotic germination of European terrestrial orchids. Kew Bulletin 1986, 437–445.
- Zettler, L.W.; Piskin, K.A. Mycorrhizal fungi from protocorms, seedlings and mature plants of the eastern prairie fringed orchid, Platanthera leucophaea (Nutt.) Lindley: a comprehensive list to augment conservation. The American Midland Naturalist 2011, 166, 29–39.
- Reiter, N.; Whitfield, J.; Pollard, G.; Bedggood, W.; Argall, M.; Dixon, K.; Davis, B.; Swarts, N. Orchid re-introductions: an evaluation of success and ecological considerations using key comparative studies from Australia. Plant ecology 2016, 217, 81–95.
- Reiter, N.; Lawrie, A.C.; Linde, C.C. Matching symbiotic associations of an endangered orchid to habitat to improve conservation outcomes. Annals of Botany 2018, 122, 947–959.
- Batty, A.L.; Brundrett, M.C.; Dixon, K.W.; Sivasithamparam, K. New methods to improve symbiotic propagation of temperate terrestrial orchid seedlings from axenic culture to soil. Australian Journal of Botany 2006, 54, 367–374.
- Johnson, T.R.; Stewart, S.L.; Dutra, D.; Kane, M.E.; Richardson, L. Asymbiotic and symbiotic seed germination of Eulophia alta (Orchidaceae)—preliminary evidence for the symbiotic culture advantage. Plant cell, Tissue and organ culture 2007, 90, 313–323.
- Jamja, T.; Bora, S.; Tabing, R.; Tagi, N.; Chaurasiya, A.K.; Devi, N.; Yangfo, M. Symbiotic germination in orchids: an overview of ex situ and in situ symbiotic seed germination. Ecology Environment and Conservation 2023, 29, 1251–1265.
- Sommerville, K.D.; Siemon, J.P.; Wood, C.B.; Offord, C.A. Simultaneous encapsulation of seed and mycorrhizal fungi for long-term storage and propagation of terrestrial orchids. Australian Journal of Botany 2008, 56, 609–615.
- Yang, H.; Li, N.-Q.; Gao, J.-Y. A novel method to produce massive seedlings via symbiotic seed germination in orchids. Frontiers in Plant Science 2023, 14, 1114105.
- Wright, M.; French, G.; Cross, R.; Cousens, R.; Andrusiak, S.; McLean, C.B. Site amelioration for direct seeding of Caladenia tentaculata improves seedling recruitment and survival in natural habitat. Lankesteriana International Journal on Orchidology 2007, 7, 430–432.
- De Hert, K.; Jacquemyn, H.; Provoost, S.; Honnay, O. Absence of recruitment limitation in restored dune slacks suggests that manual seed introduction can be a successful practice for restoring orchid populations. Restoration ecology 2013, 21, 159–162.
- Brundrett, M.; Scade, A.; Batty, A.; Dixon, K.; Sivasithamparam, K. Development of in situ and ex situ seed baiting techniques to detect mycorrhizal fungi from terrestrial orchid habitats. Mycological Research 2003, 107, 1210–1220. [CrossRef]
- Whigham, D.F.; O’Neill, J.P.; Rasmussen, H.N.; Caldwell, B.A.; McCormick, M.K. Seed longevity in terrestrial orchids–potential for persistent in situ seed banks. Biological conservation 2006, 129, 24–30.
- Batty, A.L.; Dixon, K.W.; Sivasithamparam, K. Soil seed-bank dynamics of terrestrial orchids. Lindleyana 2000, 15, 227–236.
- Zhao, D.-K.; Selosse, M.-A.; Wu, L.; Luo, Y.; Shao, S.-C.; Ruan, Y.-L. Orchid reintroduction based on seed germination-promoting mycorrhizal fungi derived from protocorms or seedlings. Frontiers in plant science 2021, 12, 701152.
- Brundrett, M.C. A Proposed framework for efficient and cost-effective terrestrial orchid conservation. www.preprints.org 2020, doi:doi:10.20944/preprints202004.0465.v1.
- Benzing, D.H. Vascular epiphytism: Taxonomic participation and adaptive diversity. Annals of the Missouri Botanical Garden 1987, 74, 183–204.
- Janissen, B.; French, G.; Selby-Pham, J.; Lawrie, A.C.; Huynh, T. differences in emergence and flowering in wild, re-introduced and translocated populations of an endangered terrestrial orchid and the influences of climate and orchid mycorrhizal abundance. Australian Journal of Botany 2021, 69, 9–20.
- Reiter, N.; Menz, M.H. Optimising conservation translocations of threatened Caladenia (orchidaceae) by identifying adult microsite and germination niche. Australian Journal of Botany 2022, 70, 231–247.
- Collins, M.T.; Dixon, K.W. Micropropagation of an Australian terrestrial orchid Diuris longifolia R. Br. Australian Journal of Experimental Agriculture 1992, 32, 131–135.
- Zettler, L.W.; McInnis, J. symbiotic seed germination and development of Spiranthes cernua and Goodyera pubescens (Orchidaceae: Spiranthoideae). Lindleyana, 1993, 8, 155-162.
- Oddie, R.L.A.; Dixon, K.W.; McComb, J.A. influence of substrate on asymbiotic and symbiotic in vitro germination and seedling growth of two Australian terrestrial orchids. Lindleyana 1994, 9, 183–189.
- Quay, L.; McComb, J.A.; Dixon, K.W. Methods for ex vitro germination of Australian terrestrial orchids. HortScience 1995, 30, 1445–1446.
- Batty, A.L.; Dixon, K.W.; Brundrett, M.; Sivasithamparam, K. Constraints to symbiotic germination of terrestrial orchid seed in a mediterranean bushland. New Phytologist 2001, 152, 511–520.
- Yamato, M.; Iwase, K. Introduction of Asymbiotically Propagated Seedlings of Cephalanthera falcata (Orchidaceae) into natural habitat and investigation of colonized mycorrhizal fungi. Ecological Research 2008, 23, 329–337. [CrossRef]
- Chou, L.-C.; Chang, D.C.-N. Asymbiotic and symbiotic seed germination of Anoectochilus formosanus and Haemaria discolor and their F1 Hybrids. Botanical Bulletin of Academia Sinica 2004, 45, 143–147.
- Batty, A.L.; Dixon, K.W.; Brundrett, M.; Sivasithamparam, K. Long-term storage of mycorrhizal fungi and seed as a tool for the conservation of endangered Western Australian terrestrial orchids. Australian Journal of Botany 2001, 49, 619–628.
- Swarts, N.D.; Sinclair, E.A.; Francis, A.; Dixon, K.W. Ecological specialization in mycorrhizal symbiosis leads to rarity in an endangered orchid. Molecular Ecology 2010, 19, 3226–3242.
- Freestone, M.; Linde, C.; Swarts, N.; Reiter, N. Ceratobasidium orchid mycorrhizal fungi reveal intraspecific variation and interaction with different nutrient media in symbiotic germination of Prasophyllum (Orchidaceae). Symbiosis 2022, 87, 255–268. [CrossRef]
- Phillips, R.D.; Faast, R.; Bower, C.C.; Brown, G.R.; Peakall, R. Implications of pollination by food and sexual deception for pollinator specificity, fruit set, population genetics and conservation of Caladenia (Orchidaceae). Australian Journal of Botany 2009, 57, 287–306;
- Brundrett, M.C.; Ladd, P.G.; Keighery, G.J. Pollination strategies are exceptionally complex in southwestern Australia—A globally significant ancient biodiversity hotspot. Australian Journal of Botany 2024, 72, 1–70. [CrossRef]
- Brundrett, M.C. Using vital statistics and core habitat maps to manage critically endangered orchids in the West Australian wheatbelt. Australian Journal of Botany 2016, 64, 51–64.
- Brundrett, M. Wheatbelt Orchid Rescue Project: Case Studies of Collaborative Orchid Conservation in Western Australia; University of Western Australia: Nedlands, Western Australia, 2011;
- Brundrett, M. Identification and Ecology of Southwest Australian Orchids; Western Australian Naturalists’ Club Inc.: Perth, Western Australia, 2014;
- Brundrett, M.; Sivasithamparam, K.; Ramsay, M.; Krauss, S.; Taylor, R.; Bunn, E.; Hicks, A.; Karim, N.; Debeljak, N.; Mursidawati, S.; et al. Orchid Conservation Techniques Manual; First International Orchid Conservation Congress: Kings Park, Western Australia, 2001;
- Brundrett, M.; Ager, E. Wheatbelt Orchid Rescue Project Final Report: Seed Collecting, Soil Baiting and Propagation of Orchids; The University of Western Australia: Nedlands WA, 2011;
- Rasmussen, H.; Johansen, B.; Andersen, T.F. Density-dependent interactions between seedlings of Dactylorhiza majalis (Orchidaceae) in symbiotic in vitro culture. Physiologia Plantarum 1989, 77, 473–478.
- Anderson, M.J. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. PRIMER-E, Plymouth Marine Laboratory 2008, 214.
- Bonnardeaux, Y.; Brundrett, M.; Batty, A.; Dixon, K.; Koch, J.; Sivasithamparam, K. Diversity of mycorrhizal fungi of terrestrial orchids: compatibility webs, brief encounters, lasting relationships and alien invasions. Mycological research 2007, 111, 51–61.
- De Long, J.R.; Swarts, N.D.; Dixon, K.W.; Egerton-Warburton, L.M. Mycorrhizal preference promotes habitat invasion by a native Australian orchid: Microtis media. Annals of botany 2012, 111, 409–418.
- Hadley, G. Non-specificity of symbotic infection in orchid mycorrhiza. New Phytologist 1970, 69, 1015–1023. [CrossRef]
- Masuhara, G.; Katsuya, K. In situ and in vitro specificity between Rhizoctonia spp. and Spiranthes sinensis (Persoon) Ames, var. amoena (M. Bieberstein) Hara (Orchidaceae). New Phytologist 1994, 127, 711–718.
- Raleigh, R.E. Propagation and Biology of Arachnorchis (orchidaceae) and their mycorrhizal fungi. PhD Thesis, Royal Melbourne Institute of Technology (Australia), 2005.
- Scade, A.; Brundrett, M.; Batty, A.; Dixon, K.; Sivasithamparam, K. Survival of transplanted terrestrial orchid seedlings in urban bushland habitats with high or low weed cover. Australian Journal of Botany 2006, 54, 383–389. [CrossRef]
- Silcock, J.L.; Simmons, C.L.; Monks, L.; Dillon, R.; Reiter, N.; Jusaitis, M.; Vesk, P.A.; Byrne, M.; Coates, D.J. Threatened plant translocation in Australia: a review. Biological Conservation 2019, 236, 211–222.
- Wright, M.; Cross, R.; Dixon, K.; Huynh, T.; Lawrie, A.; Nesbitt, L.; Pritchard, A.; Swarts, N.; Thomson, R. Propagation and reintroduction of Caladenia. Australian Journal of Botany 2009, 57, 373–387.
- Dutra, D.; Johnson, T.R.; Kauth, P.J.; Stewart, S.L.; Kane, M.E.; Richardson, L. Asymbiotic seed germination, in vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell, Tissue and Organ Culture 2008, 94, 11–21.
- Zi, X.-M.; Sheng, C.-L.; Goodale, U.M.; Shao, S.-C.; Gao, J.-Y. In situ seed baiting to isolate germination-enhancing fungi for an epiphytic orchid, Dendrobium aphyllum (Orchidaceae). Mycorrhiza 2014, 24, 487–499.
- Cruz-Higareda, J.B.; Luna-Rosales, B.S.; Barba-Alvarez, A. A novel seed baiting technique for the epiphytic orchid Rhynchostele cervantesii, a means to acquire mycorrhizal fungi from protocorms. Lankesteriana 2015, 15, 67–76.
- Higaki, K.; Rammitsu, K.; Yamashita, Y.; Yukawa, T.; Ogura-Tsujita, Y. A method for facilitating the seed germination of a mycoheterotrophic orchid, Gastrodia pubilabiata, using decomposed leaf litter harboring a basidiomycete fungus, Mycena sp. Botanical studies 2017, 58, 1-7 (59).
- Yang, W.-K.; Li, T.-Q.; Wu, S.-M.; Finnegan, P.M.; Gao, J.-Y. Ex situ seed baiting to isolate germination-enhancing fungi for assisted colonization in Paphiopedilum spicerianum, a critically endangered orchid in China. Global Ecology and Conservation 2020, 23, e01147. [CrossRef]
- Batty, A.L.; Brundrett, M.C.; Dixon, K.W.; Sivasithamparam, K. In situ symbiotic seed germination and propagation of terrestrial orchid seedlings for establishment at field sites. Australian Journal of Botany 2006, 54, 375–381.
- Feuerherdt, L.; Petit, S.; Jusaitis, M. Distribution of mycorrhizal fungus associated with the endangered pink-lipped spider orchid (Arachnorchis (Syn. Caladenia) behrii) at Warren Conservation Park in South Australia. New Zealand Journal of Botany 2005, 43, 367–371.
- Diez, J.M. Hierarchical patterns of symbiotic orchid germination linked to adult proximity and environmental gradients. Journal of Ecology 2007, 95, 159–170.
- Jacquemyn, H.; Brys, R.; Lievens, B.; Wiegand, T. Spatial variation in below-ground seed germination and divergent mycorrhizal associations correlate with spatial segregation of three co-occurring orchid species. Journal of Ecology 2012, 100, 1328–1337.
- Reiter, N.; Dimon, R.; Arifin, A.; Linde, C. Culture age of Tulasnella Affects Symbiotic Germination of the Critically Endangered Wyong sun orchid Thelymitra adorata (Orchidaceae). Mycorrhiza 2023, 33, 409–424.
- Harzli, I.; Özdener Kömpe, Y. Impact of fungal symbionts of co-occurring orchids on the seed germination of Serapias orientalis and Spiranthes spiralis. Current Microbiology 2025, 82, 79.
- Shao, S.-C.; Burgess, K.S.; Cruse-Sanders, J.M.; Liu, Q.; Fan, X.-L.; Huang, H.; Gao, J.-Y. Using in situ symbiotic seed germination to restore over-collected medicinal orchids in southwest China. Frontiers in Plant Science 2017, 8, 888.
- Wang, X.-J.; Wu, Y.-H.; Ming, X.-J.; Wang, G.; Gao, J.-Y. Isolating ecological-specific fungi and creating fungus-seed bags for epiphytic orchid conservation. Global Ecology and Conservation 2021, 28, e01714.
- Aewsakul, N.; Maneesorn, D.; Serivichyaswat, P.; Taluengjit, A.; Nontachaiyapoom, S. Ex vitro symbiotic seed germination of Spathoglottis plicata blume on common orchid cultivation substrates. Scientia Horticulturae 2013, 160, 238–242.
- Yao, N.; Wang, T.; Cao, X. Epidendrum radicans fungal community during ex situ germination and isolation of germination-enhancing fungi. Microorganisms 2022, 10, 1841.
- Schweiger, J.M.-I.; Bidartondo, M.I.; Gebauer, G. Stable isotope signatures of underground seedlings reveal the organic matter gained by adult orchids from mycorrhizal fungi. Functional ecology 2018, 32, 870–881.
- Mehra, S.; Morrison, P.D.; Coates, F.; Lawrie, A.C. Differences in carbon source utilisation by orchid mycorrhizal fungi from common and endangered species of Caladenia (Orchidaceae). Mycorrhiza 2017, 27, 95–108.
- Yeung, E.C.-T.; Lee, Y.-I. The orchid protocorm. In Orchid Propagation; Yeung, E.C.-T., Lee, Y.-I., Eds.; Springer Protocols Handbooks; Springer US: New York, NY, 2024; pp. 3–15 ISBN 978-1-0716-4030-2.
- Veyret, Y. Development of the embryo and the young seedling stages of orchids. In The orchids: scientific studies; Withner, C., L., Ed.; John Wiley & Sons: New York, 1974; pp. 223–265.
- Kauth, P.J.; Kane, M.E. In vitro ecology of Calopogon tuberosus var. tuberosus (Orchidaceae) seedlings from distant populations: implications for assessing ecotypic differentiation. The Journal of the Torrey Botanical Society 2009, 136, 433–444.
- Leroux, G.; Barabé, D.; Vieth, J. Morphogenesis of the protocorm of Cypripedium acaule (Orchidaceae). Plant Systematics and Evolution 1997, 205, 53–72.
- González-Orellana, N.; Salazar Mendoza, A.; Tremblay, R.L.; Ackerman, J.D. Host suitability for germination differs from that of later stages of development in a rare epiphytic orchid. Lankesteriana 2024, 24, 93–114.
Figure 1.
This project focussed on three threatened West Australian orchid species - Caladenia graniticola (A), Caladenia melanema (B), and Caladenia williamsiae (C).
Figure 1.
This project focussed on three threatened West Australian orchid species - Caladenia graniticola (A), Caladenia melanema (B), and Caladenia williamsiae (C).

Figure 2.
A. Artificial pollination of Caladenia graniticola to allow seed collection. B. Seed bags used to protect developing seedpods. CD. Opening seed bags to retrieve seeds.
Figure 2.
A. Artificial pollination of Caladenia graniticola to allow seed collection. B. Seed bags used to protect developing seedpods. CD. Opening seed bags to retrieve seeds.

Figure 3.
Orchid seed and ex situ seed baiting examples. A. Dried orchid seed on 1 mm graph paper. B. Healthy imbibed seed with swollen embryos (Stage 1). C. Seed squares (10x10 mm) prepared for seed baiting. D. Seed baiting plate with 20 seed squares. E. Caladenia graniticola protocorms on baiting plate. F. Focus stacked image of two protocorms from E showing trichomes extending outwards, brown seed coat remnants, and leaf initiation (arrows).
Figure 3.
Orchid seed and ex situ seed baiting examples. A. Dried orchid seed on 1 mm graph paper. B. Healthy imbibed seed with swollen embryos (Stage 1). C. Seed squares (10x10 mm) prepared for seed baiting. D. Seed baiting plate with 20 seed squares. E. Caladenia graniticola protocorms on baiting plate. F. Focus stacked image of two protocorms from E showing trichomes extending outwards, brown seed coat remnants, and leaf initiation (arrows).

Figure 4.
Non-sterile symbiotic seedling germination in semi-permeable pouches. A-D. The same seedlings of Caladenia graniticola observed over time in a germination pouch (scale bar = 5 mm). E. Germination pouch in 0.6 L container with potting mix.
Figure 4.
Non-sterile symbiotic seedling germination in semi-permeable pouches. A-D. The same seedlings of Caladenia graniticola observed over time in a germination pouch (scale bar = 5 mm). E. Germination pouch in 0.6 L container with potting mix.

Figure 5.
A. Transplanting seedlings from non-sterile symbiotic germination into a pouch. B-D. Development of seedlings of Caladenia graniticola after transplanting into growth pouches at 5-weeks old. E. magnified view of seedlings showing dropper initiation (arrow). F. Transplanted seedlings of Caladenia williamsiae germinated asymbiotically in vitro. Scale bars are 100 mm.
Figure 5.
A. Transplanting seedlings from non-sterile symbiotic germination into a pouch. B-D. Development of seedlings of Caladenia graniticola after transplanting into growth pouches at 5-weeks old. E. magnified view of seedlings showing dropper initiation (arrow). F. Transplanted seedlings of Caladenia williamsiae germinated asymbiotically in vitro. Scale bars are 100 mm.

Figure 6.
Later growth stages of Caladenia roei seedlings. A. Advanced seedlings in an incubator in a 1 l container. B. Tuber formation in a container. C-D. Older seedlings with droppers (arrows), growing in translocation pouches within window pots in the glasshouse. E. Tubers formed in a translocation pouch.
Figure 6.
Later growth stages of Caladenia roei seedlings. A. Advanced seedlings in an incubator in a 1 l container. B. Tuber formation in a container. C-D. Older seedlings with droppers (arrows), growing in translocation pouches within window pots in the glasshouse. E. Tubers formed in a translocation pouch.

Figure 7.
A-D. Seedlings of Caladenia williamsiae growing asymbiotically in vitro media after 4, 6, 8, or 10 weeks of growth (scale 1 mm).
Figure 7.
A-D. Seedlings of Caladenia williamsiae growing asymbiotically in vitro media after 4, 6, 8, or 10 weeks of growth (scale 1 mm).

Figure 8.
A. Protocorm growth rates for 5 Caladenia species in asymbiotic culture. B. Protocorm growth rates for 5 Caladenia species in soil baiting plates, a semi-natural symbiotic environment.
Figure 8.
A. Protocorm growth rates for 5 Caladenia species in asymbiotic culture. B. Protocorm growth rates for 5 Caladenia species in soil baiting plates, a semi-natural symbiotic environment.
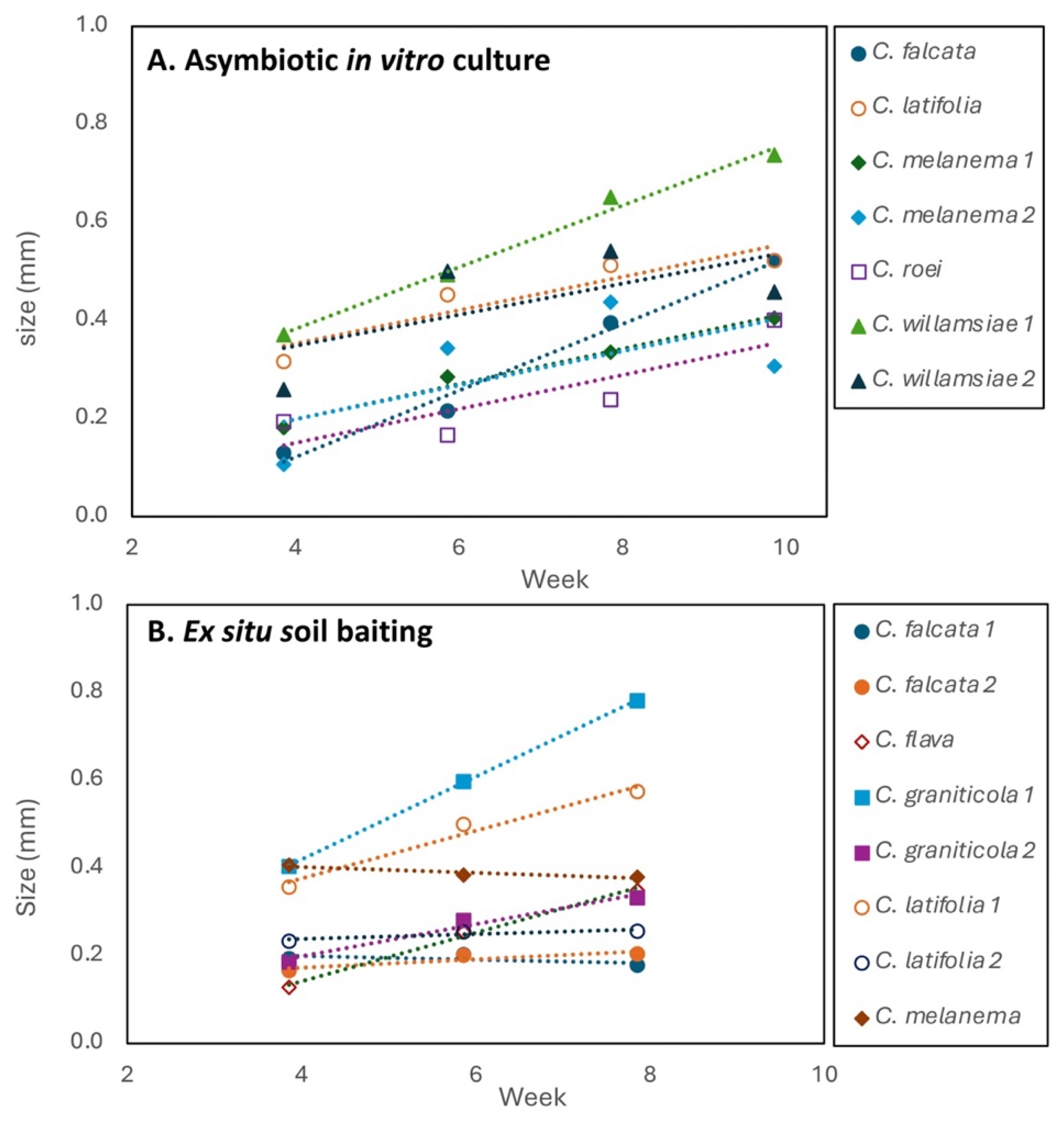
Figure 9.
A. Ex situ seed baiting results for 3 Caladenia graniticola seed sources in comparison with 2 common orchids in 7 soil samples from natural habitats (C = Caladenia, M = Microtis). B. Protocorm sizes from seed baiting, showing separate plates or seed sources (C. melonema, C. graniticola). C. Relationship between protocorm density and size on these plates.
Figure 9.
A. Ex situ seed baiting results for 3 Caladenia graniticola seed sources in comparison with 2 common orchids in 7 soil samples from natural habitats (C = Caladenia, M = Microtis). B. Protocorm sizes from seed baiting, showing separate plates or seed sources (C. melonema, C. graniticola). C. Relationship between protocorm density and size on these plates.
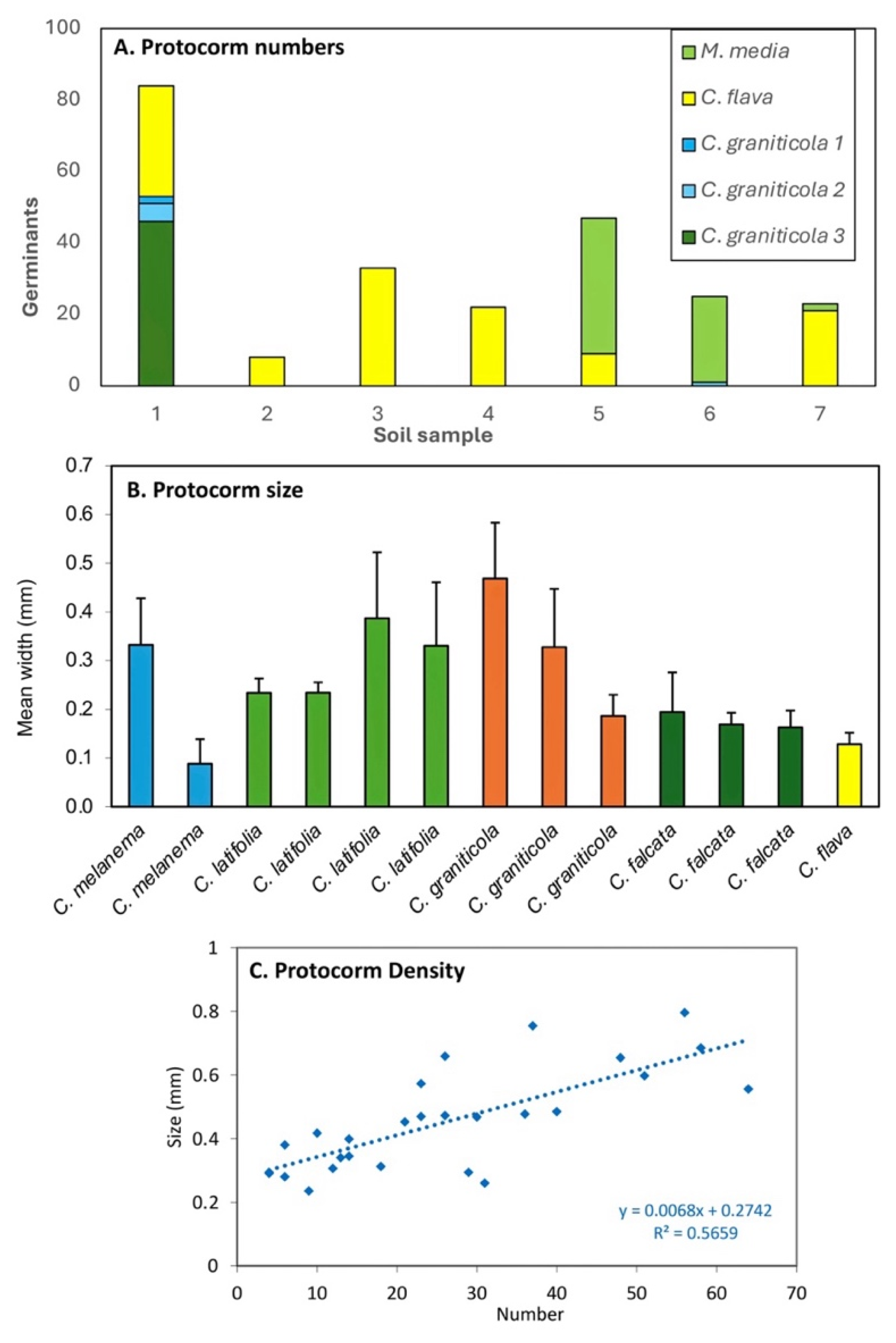
Figure 10.
A. Protocorm growth rates for Caladenia species in non-sterile symbiotic culture in germination pouches. B. Separate growth data for leaves and protocorms. C. Ongoing leaf growth of seedlings transplanted into growing pouches over a month. All values are averages for 10-20 protocorms in a pouch.
Figure 10.
A. Protocorm growth rates for Caladenia species in non-sterile symbiotic culture in germination pouches. B. Separate growth data for leaves and protocorms. C. Ongoing leaf growth of seedlings transplanted into growing pouches over a month. All values are averages for 10-20 protocorms in a pouch.
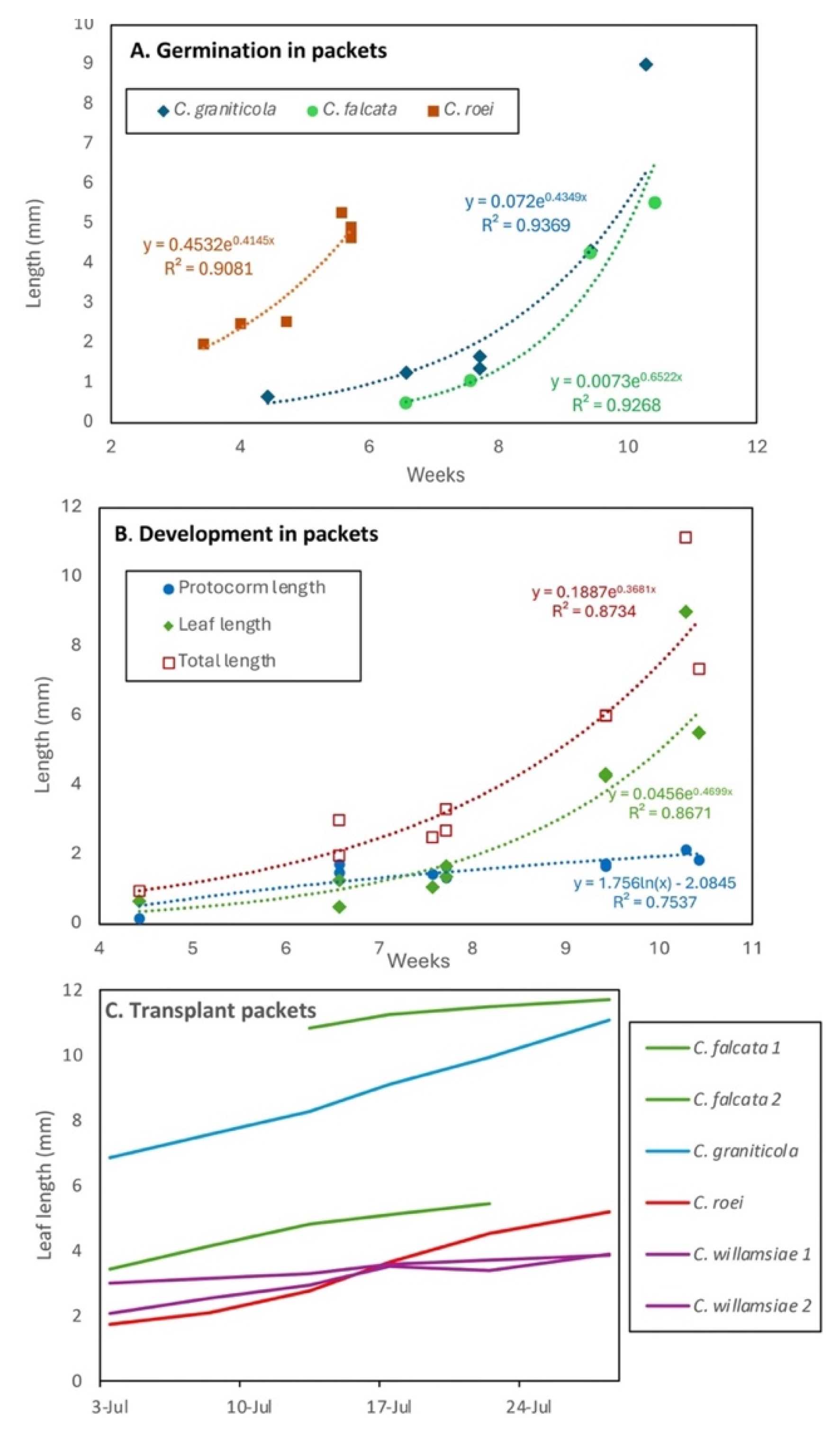
Figure 11.
Growth rate comparisons for propagation methods using data from Table 1. A. Asymbiotic (7 studies) and symbiotic (12) axenic germination showing averages for all species and growth stage at harvest (best results only if multiple methods used). B. Average time required to transition between growth stages for methods in Table 1 (stage 2 = trichome initiation, 3 = leaf primordia, 4 = small leaf, 5 = large leaf).
Figure 11.
Growth rate comparisons for propagation methods using data from Table 1. A. Asymbiotic (7 studies) and symbiotic (12) axenic germination showing averages for all species and growth stage at harvest (best results only if multiple methods used). B. Average time required to transition between growth stages for methods in Table 1 (stage 2 = trichome initiation, 3 = leaf primordia, 4 = small leaf, 5 = large leaf).
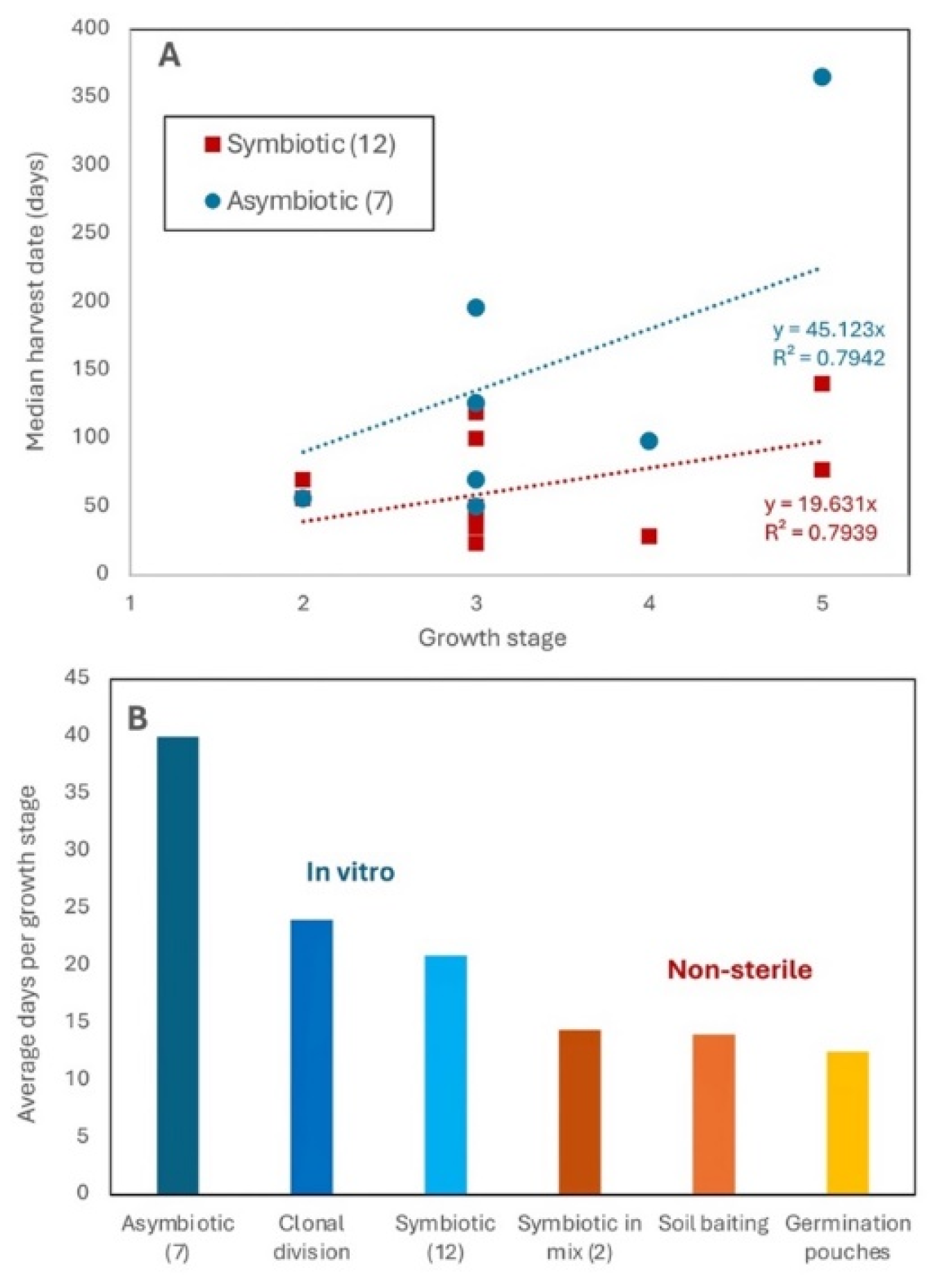
Figure 12.
Orchid propagation flowchart using adaptive approaches based on non-sterile FORGE methods.
Figure 12.
Orchid propagation flowchart using adaptive approaches based on non-sterile FORGE methods.
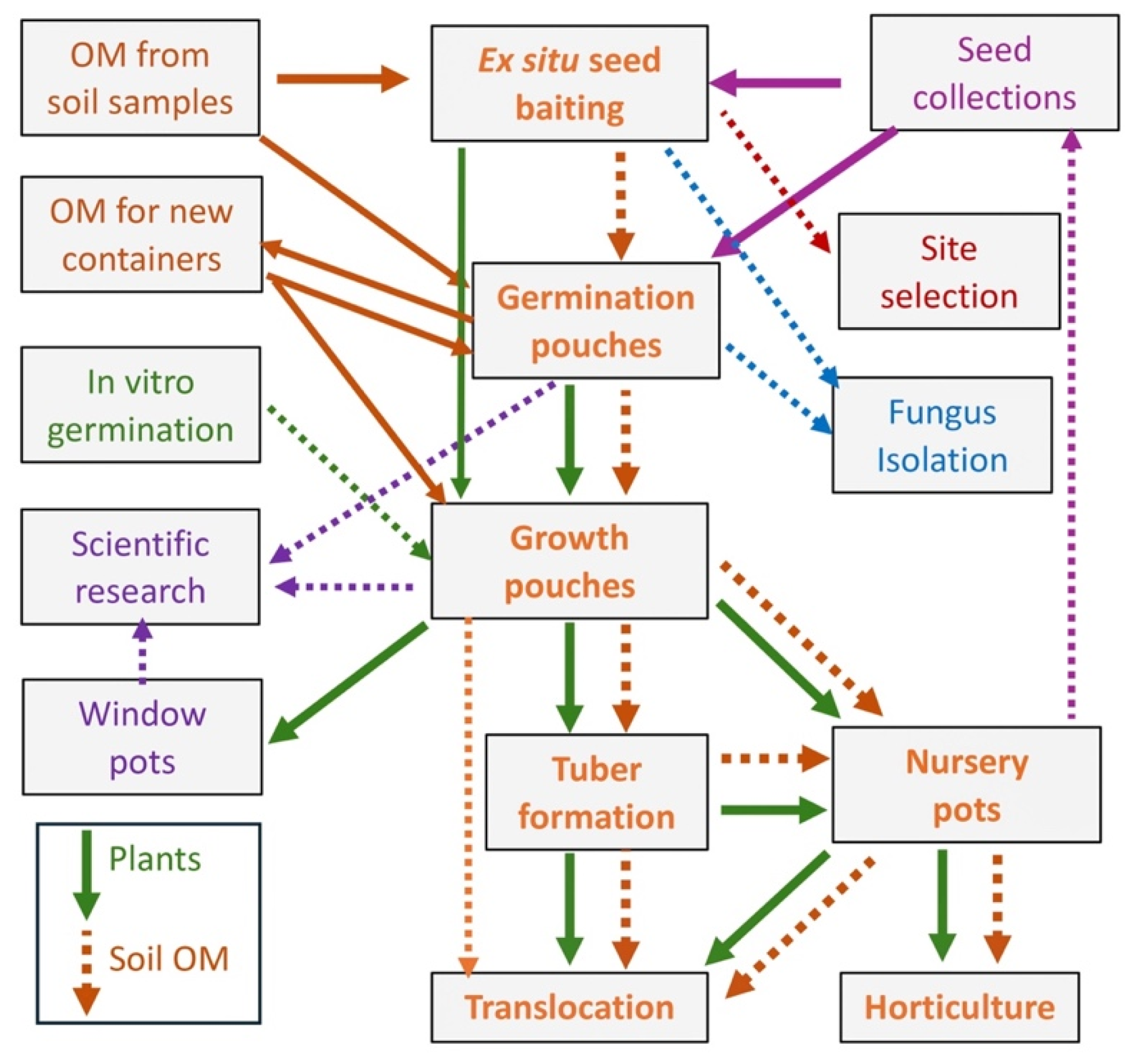

Table 1.
Relative germination times and yields for orchid propagation methods from selected references (Australian orchids unless otherwise stated).
Table 1.
Relative germination times and yields for orchid propagation methods from selected references (Australian orchids unless otherwise stated).
| Method | Orchids | Time taken | Success criteria | Yield | Reference |
| Symbiotic | Dactylorhiza elata, Orchis spp. (European) | 39-161 days | Stage 3-4 | ND | [24] |
| Symbiotic | 7 Thelymitra, 2 Diuris, 2 Pterostylis | 24-65 days | Stage 3 | ND | [23] |
| Clonal division | Diuris magnifica (was D. longifolia) | 120 days | Stage 5 (roots) | 5-55% | [43] |
| Symbiotic | Spiranthes cenua, Goodyera pubescens (European) | 23 days | Stage 3 | 8-95% | [44] |
| Symbiotic | Elythranthera emarginata, Diuris magnifica | 17 weeks | Stage 3 (trichomes) | 0.3-42% | [45] |
| Asymbiotic | As above | 28 weeks | As above | 11-52% | [45] |
| Symbiotic | Caladenia latifolia, D. magnifica (inoculated sterile potting mix) | 10 weeks | Stage 4 (leaf) | 80-230 per 270 ml | [46] |
| Symbiotic | Caladenia arenicola, Diuris magnifica, Pterostylis sanguinea, Thelymitra crinita | 4-6 weeks | Stage 3 (trichomes) | 31-59% | [47] |
| Asymbiotic | Eulophia alta (African) | 18 weeks | Stage 3 (trichomes) | 20-88% | [29] |
| Symbiotic | As above | 6 weeks | Stage 3 | 44.3% | [29] |
| Asymbiotic | Cephalanthera falcata, Anoectochilus formosanus, Haemaria discolor (Asian) | 2-5 months | Stage 4-6 | 50-70% | [48] |
| Asymbiotic | A. formosanus, H. discolor (Asian) | 50 days | Stage 3 (trichomes) | 72-75% | [49] |
| Symbiotic | As above | 50 days | Stage 3 | 80-81% | [49] |
| Soil baiting | Disa bracteata, Microtis media, Pterostylis sp., Caladenia arenicola, Diuris sp., Caladenia latifolia | 4-8 weeks | Stage 2-4 | 2 - 27% | [35] |
| Symbiotic | Caladenia arenicola, Diuris magnifica, Thelymitra crinita | 10-12 weeks | Stage 3-4 (leaf size) | 35 -53 % | [28] |
| Symbiotic | Caladenia formosa | 4 weeks | Stage 3-4 (green leaf) | 4-21% | [20] |
| Symbiotic | Caladenia arenicola, Pterostylis sanguinea (seed and fungi in alginate beads) | 8 weeks | Stage 3-5 (leaf size) | 560-2300 / m2 | [50] |
| Symbiotic | Caladenia huegelii, C. discoidea, C. arenicola, C. flava, C. longicauda | 10 weeks | Stage 2 (trichomes) | 35-85% | [51] |
| Asymbiotic | Pterostylis nutans, Microtis arenaria,我Thelymitra pauciflora, Prasophyllum pruinosum | 10 weeks | Stage 3 -4 | 10-95% | [15] |
| Symbiotic | Caladenia latifolia | 8 weeks | Stage 2 | 95% | [16] |
| Asymbiotic | D. magnifica, Thelymitra benthamiana, Spiculaea ciliata, Cyanicula gemmata, Elythranthera brunonis, Ericksonella saccharata, Pheladenia deformis, Eriochilus dilatatus, Microtis media, C. huegelii | 8 weeks | As above | 60-95% | [16] |
| Symbiotic | Prasophyllum frenchii | 3 months | Stage 3 (leaf primordia) | 0-5% | [52] |
| Asymbiotic | Prasophyllum 18 spp. | 12 months | Stage 5 (leaf) | 0-93% | [21] |
Table 2.
Seed and soil collections and replicates per orchid in propagation trails.
| Caladenia species | Seed batches | Seed sites | Soils (sites) | Baiting plates | Axenic plates | Germ pouch | Grow pouch | Pots |
|---|---|---|---|---|---|---|---|---|
| C. graniticola | 22 | 4 | 20 (4) | 20 | 4 | 2 | 10 | 4 |
| C. melanema | 7 | 3 | 5 (2) | 6 | 4 | 1 | 5 | 4 |
| C. williamsiae | 4 | 1 | 4 (1) | 5 | 4 | 6 | 6 | |
| C. roei | 2 | 1 | 5 | 2 | 2 | 7 | 3 | |
| C. dimidia | 1 | 1 | 2 | 2 | 4 | |||
| C. latifolia | 3 | 1 | 2 | 2 | ||||
| C. falcata | 1 | 1 | 6 | 3 | 2 | 5 | 3 | |
| C. flava | 4 | 1 | 3 | 1 | ||||
| C. radialis | 2 | 1 | 5 | 3 |
Table 3.
Total seedling numbers relative to production method or stage (numbers per container).
| Asymbiotic Sterile | Soil Baiting | Seed Packets | Incubator | Greenhouse | Field | |
|---|---|---|---|---|---|---|
| C. williamsiae | 129 | 1 | 64 | 28 | 28 | |
| C. melanema | 94 | 42 | 13 | 42 | 20 | 20 |
| C. graniticola | 256 | 28 | 51 | 20 | 20 | |
| C. falcata | 9 | 155 | 157 | 25 | 50 | 10 |
| C. roei | 29 | 193 | 108 | 48 | 80 | 20 |
| C. latifolia | 96 | 93 | 5 | 10 | ||
| C. flava | 159 | 10 | 10 | |||
| Total | 357 | 899 | 311 | 333 | 218 | 108 |
Table 4.
Cost and effort comparison for orchid propagation methods based on data in Table S2.
Table 4.
Cost and effort comparison for orchid propagation methods based on data in Table S2.
| Factor | Asymbiotic | Soil Baiting | Seed Packets |
|---|---|---|---|
| Preparation time (days) | 2 | 3 | 3 |
| Set up time (days) | 2 | 1 | 1 |
| Maintenance time (hrs./week) | 1 | 1 | 2 |
| Multiplies fungi associates in soil | no | no | yes |
| Complexity of methods | high | medium | low |
| Loss due to contamination or similar | 50% | 15% | 10% |
| Typical growth rate (mm/week) | 0.04 | 0.04 | 1.4 |
| Yield (average size mm) | 0.46 | 0.38 | 7.5 |
| Yield (seedlings per container) | 19 | 66 | 54 |
| Time to produce seedlings (stage 3) | 2-6 months | 1-2 months | 1-2 months |
| Further growth required for outplanting | months | weeks | weeks (if needed) |
| Survival in greenhouse | low | medium | high |
| Cost per 100 seedlings in 2009 (Table S1) | $ 3.79 | $ 1.09 | $ 1.30 |
| Overall Ranking | * | ** | *** |
Table 5.
Orchid developmental stages from seeds to reproductive adult plants (TF = transplantable in the FORGE system, SN = suitable for transfer to a nursery, SO = suitable for outplanting or translocation).
Table 5.
Orchid developmental stages from seeds to reproductive adult plants (TF = transplantable in the FORGE system, SN = suitable for transfer to a nursery, SO = suitable for outplanting or translocation).
| Stage | Description | Morphology | Roles | Status |
|---|---|---|---|---|
| 0 | Seed | Small embryo in dry seed coat | Dispersal and quiescence | Waiting |
| 1 | Imbibed seed | Spherical swollen embryo, split seed coat (if viable and non-dormant) | Irreversible transition to growth or death due to water uptake | Pre-germination |
| 2 | Protocorm | Growth from cell division starts, leading to cell differentiation and trichome initiation (ovoid shape) | Attraction of fungi and potential initiation of symbiosis by fungal colonisation | Germination starts |
| 3 | Advanced protocorm | Substantial growth (change in shape - longer, broader, etc.), many long trichomes, fungal coils, leaf primordia (if relevant) | Establishment of functional mycorrhizas, resource acquisition, initiation of photosynthesis (if relevant) | Germination success – TF |
| 4 | Seedling | Substantial leaf growth (or stem, root, or rhizome), protocorm growth slows | Metabolic balance shifts towards autotrophy (if relevant), fuelling rapid growth | Growth – TF |
| 5 | Advanced seedling | Organ differentiation to form root, or dropper, or rhizome (varies with orchid) | Switch towards adult growth phase and resource storage | Consolidation我– TF, SN |
| 6 | Adult plant | Vegetative organs complete (tuber, corm, rhizome, stem - varies with orchid), plant dormancy or quiescence starts (if relevant) | Completion of structures required for successful perennation | Completion and survival – SN, SO |
| 7 | Reproductive plant | Flowering and seed set or clonal division (usually on a subsequent year) | Reproduction potentially leading to local persistence and spread | Establishment and expansion – SO |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



