Submitted:
10 April 2025
Posted:
11 April 2025
You are already at the latest version
Abstract
Originally identified as a potential receptor for opioids, sigma-1 receptor is a class of intracellular receptors expressed in various tissues including neurons. As a resident protein in the mitochondria-associated endoplasmic reticulum (ER) membrane, sigma-1 receptor has been found involved in various biological and disease processes including stress responses, neurotransmission, and neurodegenerative diseases. Herein we discuss the history, structure, functions, and pharmacology of sigma-1 receptor and its pathological implications in neurodegenerative diseases.
Keywords:
sigma-1 receptor
; sigma receptor
; TMEM97
; neurodegenerative disorders
; ER stress
; Alzheimer’s disease
; amyotrophic lateral sclerosis
1. Introduction
Sigma receptors were discovered in the 1970s when the scientific community was actively exploring the mechanisms of action of opioids, focusing on their interactions with various receptors in the brain. W.R. Martin investigated the effects of various drugs, including opioids and opioid-like compounds, on the central nervous system. It was observed that SKF-10047, a congener of morphine, caused mydriasis, tachypnea, tachycardia, and mania in contrast with morphine, a mu receptor agonist, and ketocyclazocine, a kappa receptor agonist. Its unique binding profile did not align with the traditional opioid receptors known at the time—mu, kappa, and delta, suggesting the existence of a previously unidentified receptor type. The "sigma" classification, derived from the Greek letter σ, is used to denote its distinct binding properties [1]. This discovery has sparked considerable interests in finding additional pathways through which the opioid drugs could exert their effects.
The 1980s and 1990s were pivotal for sigma receptor research. Studies by Tam and Cook (1984) pointed out that the effects of sigma ligands could not be reversed by naloxone, an opioid antagonist, which is a defining feature of opioid receptors, suggesting a pharmacological distinction from opioid receptors [2]. It was eventually accepted that sigma receptors are not a typical opioid receptor due to their unique binding sites, affinities, and pharmacological profile, as well as their naloxone insensitivity [3].
Later, based on pharmacological profiles, binding characteristics, and tissue distribution patterns, sigma receptors were categorized into sigma-1 receptor (S1R) and sigma-2 receptors (S2R) [4]. For instance, the sigma-1 receptor has a high affinity for (+)-pentazocine, while the sigma-2 receptor does not. Conversely, certain ligands distinguish sigma-2 receptors by their higher affinity for these receptors compared to sigma-1 receptors. This differential ligand binding was one of the initial methods used to distinguish between the two receptor subtypes. In addition, the difference in molecular weight as determined by photoaffinity labeling studies and other biochemical techniques were noticed.
The cloning of the sigma-1 receptor in 1996 by Hanner et al. was a significant milestone, revealing its molecular structure and function [5]. The gene for the sigma-1 receptor encodes a protein that does not resemble any traditional G protein-coupled receptors but instead, actually shares some characteristics with molecular chaperones. In contrast, the molecular identity of the sigma-2 receptor remained elusive until much later in 2017, with its gene revealing it to be distinct from sigma-1 receptor and involved in different cellular processes [6], setting a significant advancement in sigma receptor research.
Initially misconstrued as a variant of opioid receptors, sigma receptors have emerged as a distinct class of proteins, playing distinct roles in cellular signaling, neurophysiology, and pharmacology. Their discoveries have opened new avenues to understand the molecular underpinnings of various diseases and develop novel therapeutic strategies.
2. Sigma-1 Receptor: Structure, Functions and Pharmacology
The sigma-1 receptor is a novel protein localized to the endoplasmic reticulum (ER) that interacts with lipid rafts [1,7,8,9]. The first cloning took place in 1996 when it was found to be on gene 9p13 [5,10]. The sigma-1 receptor crystal structure was identified shortly thereafter [5,11,12,13], which is a homo-trimer with a unique transmembrane domain for each region [14,15]. Within this homo-trimer, oligomerization is notably increased with sigma agonists and decreased with antagonists [16]. Certain multi-level oligomers induce the ability to form heteromers with other receptors, such as the dopamine receptor [17,18]. With a molecular weight of 25kDa, it possesses no similarities to other known proteins in the genome [9,19].
Following the discovery of the sigma-1 receptor was the sigma-2 receptor, which is less researched, however it resembles similar properties of S1R due to binding patterns to SKF-10047 and its location within lipid rafts [7,20,21,22]. Most notably, sigma-2 receptor is potentially a putative binding site for progesterone receptor membrane component 1 (Pgrmc1) [23]. Eventually, the sigma-2 receptor was later identified as the known protein TMEM97 [6]. Currently, S1R and S2R are primarily differentiated based on ligand binding assays. H3(+) pentazocine has relatively selective affinity for sigma-1, whereas H3-DTG or H3-(+)-3-PPP combined with a sigma-1 masking agent, such as pentazocine, is selective for sigma-2 [24,25].
The sigma-1 receptor plays many roles within the cell. It can translocate between the ER, mitochondria, and cell membrane [26,27,28,29]. In addition, the sigma-1 receptor couples with G-proteins, ion channels, the IP3 receptor, and the glutamate receptor [29,30,31]. Specifically, it is expressed in multiple vital organs such as the heart, liver, and kidney as well as immune cells [32]. The sigma-1 receptor can bind a wide variety of ligands, such as antipsychotics, antidepressants, and neurosteroids [3,33]. Currently, it is suggested to play a role in cell survival, as tumors and other cancers show highly expressed levels of sigma-1 receptor. Furthermore, it is suggested to play a role in neuroprotection through acting as a molecular chaperone protein [8,34,35,36,37].
In the resting state, the sigma-1 receptor is under the classification of a mitochondria-associated-ER membrane domain (MAM), as it resides near the mitochondria in ceramide and cholesterol rich lipid microdomains with BiP, an ER chaperone protein [27,38]. Under times of cell stress, the ER becomes injured, causing the sigma-1 receptor to dissociate from BiP. Upon dissociation, it can bind IP3 receptors, leading to an increase in cell survival through calcium signaling between the ER and mitochondria. Previous testing has shown that sigma-1 receptor agonists increase this stress-like response, while antagonists cause the opposite effect [27].
Overall, the sigma-1 receptor is shown to play a non-regulatory role in normal conditions, but act as a chaperone protein in times of stress to benefit cell survival [30]. In addition to modulating the IP3 pathway, sigma-1 receptor is shown to interfere with dopaminergic and cholinergic transmission, manipulating the ion channels in these various families [39,40,41,42]. Here we review the pharmacologic and pathologic implications of the sigma receptors in the nervous system.
3. Sigma-1 Receptor in Nerve Injury
Injuries to the nervous system can take place with many mechanisms, such as mechanical trauma or an ischemic insult in both the central and peripheral nervous system [43,44,45]. When a nerve undergoes injury, it has been studied that there is an increase in S1R expression [46]. S1R is theorized to play a protective role in ischemic stroke by preventing neural apoptosis and inflammation while increasing neurotrophic factors [47,48]. One of the key mechanisms in apoptosis following ischemic stroke is endoplasmic reticulum stress, which can be lessened with S1R activation [48,49].
Strokes are one of the most common causes of disability in the world, with the majority being ischemic [50]. Generally, the prognosis of a stroke can be derived from the extent of ischemia, which represents the amount of dead neural tissue [51]. There is hope that S1R agonism can reduce the size of an infarct and promote better outcomes [52]. Not only could S1R agonism result in a smaller infarct, but there are potential mechanisms to restore damage already done in white matter injury after a stroke [53]. Following an ischemic stroke, macrophages are critical for a process known as efferocytosis, which clears dead neurons from the infarct and induces neural repair and inflammation resolution [54]. The role of S1R in this topic has recently been unraveled, and studies show that S1R knockout (KO) models led to impaired macrophage function and worsened brain damage after ischemic stroke [55]. The functional outcomes from S1R agonism after stroke are not limited to functional outcomes but also cognition [56].
Strokes also carry the risk of damaging the blood-brain barrier (BBB) integrity and increasing permeability to substances otherwise unable to enter the brain [57]. S1R activation may attenuate this damage by inducing the BBB astrocytes to increase levels of glia-derived neurotrophic factor (GDNF) [58,59]. BBB integrity may be compromised due to pericyte detachment, which can also be ameliorated with S1R activation [60,61]. Another pathology seen in stroke is spreading depolarizations (SDs), which are depolarizations of neural cells due to a failure of ion homeostasis after an insult [62,63]. S1R agonists were able to resolve the SDs and promote neural survival and reduce apoptosis [64]. A rather unfavorable outcome of ischemic stroke is reperfusion injury, characterized by a worsening of damaged cells after blood flow is returned, causing inflammation and apoptosis [65]. Upregulation of S1R may promote favorable outcomes after reperfusion injury [66]
Interestingly, in models of traumatic brain injury (TBI), mice deficient in sigma receptors had better outcomes, such as less coordination impairments and neurological deficits after 1 year [67]. Other studies of TBI find that S1R agonism led to better neurological function, including restoration of blood flow and less brain edema, suggesting a biphasic role in TBI [68]. This paradoxical effect is perhaps due to long-term S1R activation resulting in unfavorable long term outcome but are beneficial in the acute phase. The acute phase of TBI is marked by inflammation, which is dampened with S1R activation [69]. Implications of S2R on TBI is much less studied, however modulation may result in more favorable outcomes [70]. As far as spinal cord injury, S1R activation resulted in effective recovery after a mechanical insult to the spine. This was done through reducing neuroinflammation and decreasing the amount of neural apoptosis and ferroptosis [71,72]. Ferroptosis is a rather novel mechanism for cell death related to the buildup of lipid oxidation products and has been described as an unfavorable mechanism resulting in neuronal cell death after injury [73,74]. Additional results concluded that ferroptosis was significantly upregulated after spinal cord injury [71].
4. Sigma-1 Receptor in Neurodegenerative Disorders
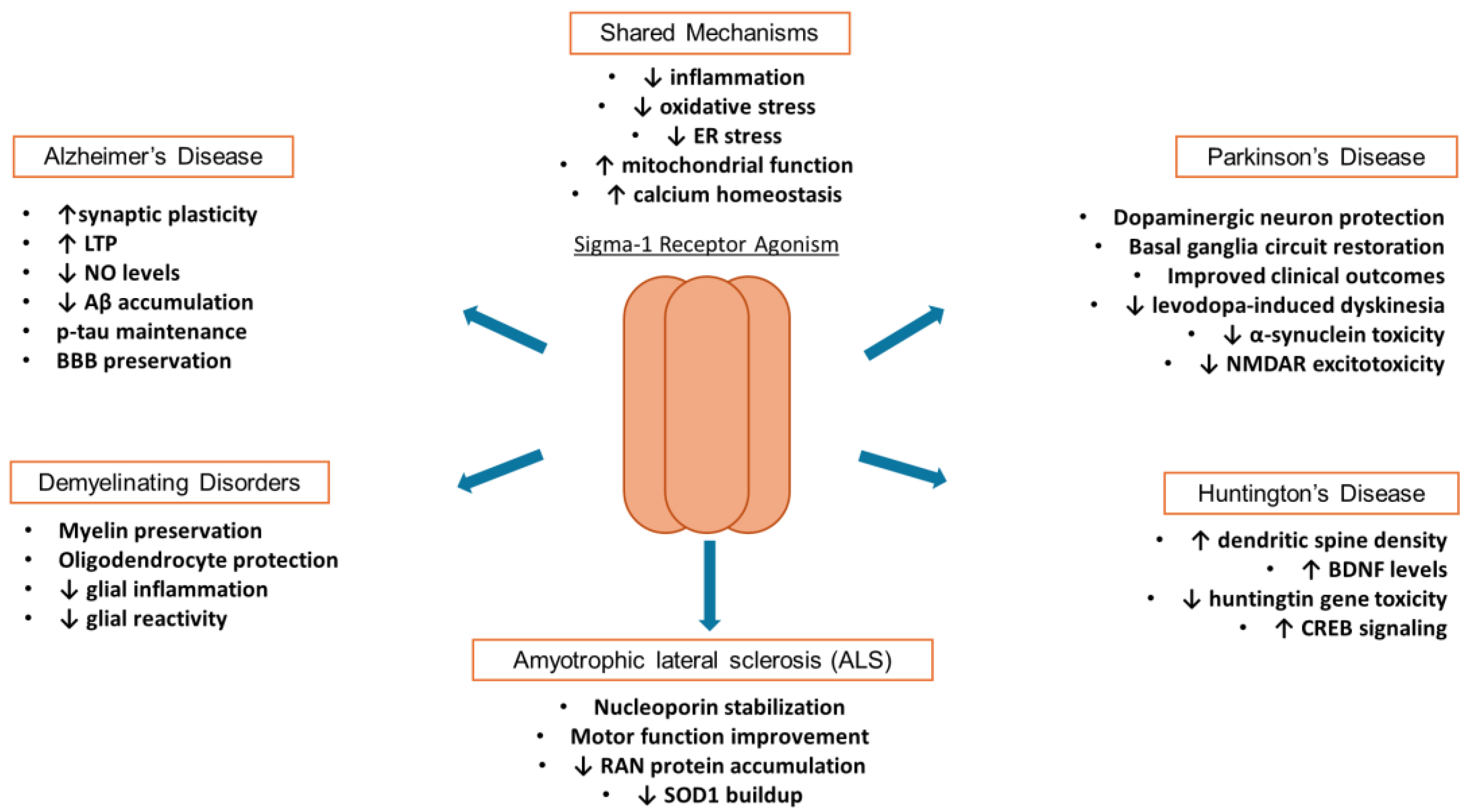
Sigma receptors are implicated in a variety of neurodegenerative disorders due to their functioning in calcium homeostasis, mitochondrial function, and oxidative stress regulation [29,75,76,77]. In fact, genetic polymorphisms related to S1R have been shown to influence development of Alzheimer’s disease (AD) [78]. Other mutations of S1R, such as loss-of-function, have been linked to the development of amyotrophic lateral sclerosis (ALS) as well as frontotemporal dementia (FTD) [79,80]. This is perhaps due to the loss of long-term potentiation seen in S1R KO mice [81]. Other neurological disorders have had favorable outcomes with sigma-receptor modulation, and these include Huntington’s Disease (HD), multiple sclerosis (MS), and Parkinson’s Disease (PD) [82,83,84]. In recent years, the sigma-1 receptor has gained attention as potential therapeutic targets for mitigating these diseases, among others [85]. The overlapping mechanisms of S1R mediated neuroprotection are summarized in Figure 1. Additionally, the utility of S2R ligands is rising in interest for their potential in providing neuroprotection [86].
S1R agonism has been linked to neuroprotection through mechanisms involving synaptic plasticity through brain-derived neurotrophic factor (BDNF) dependent mechanisms [87,88,89]. Other mechanisms for neuroprotection studied with S1R agonism include reduction of intracellular nitric oxide (NO) by inhibiting NO synthase [90,91]. Prevention of oxidative stress is another critical aspect to slow the development of neurodegeneration, and sigma-receptor ligands often act through this mechanism [92,93,94]. All these mechanisms play a central role in homeostatic plasticity, which is the stabilization of neural pathways, and loss of this plasticity is central to neurodegeneration [95,96]. Gross manifestations of neuroprotection have been studied in mouse models using modalities such as novel object recognition, showing favorable outcomes with sigma-1 agonism [97,98].
Neuroinflammation is another key factor in the progression of neurodegenerative disorders [99]. Downregulation of S1R has been shown to increase inflammation markers and induce dysregulation of the surrounding microglia [100]. S1R agonists and allosteric modulators have been shown to decrease neuroinflammation levels through reduction of microglial recruitment and inhibition of pro-inflammatory cytokines, and the resulting attenuation of gliosis has potential to slow cognitive impairment from various degenerative conditions, such a chronic epilepsy [101,102,103,104].
4.1. Sigma-1 Receptor in Alzheimer’s Disease
Mitochondrial dysfunction is a hallmark of neurodegeneration in many of neurodegenerative disorders, including Alzheimer’s Disease (AD) [105]. Restoration of mitochondrial stability is a sought-after avenue for S1R modulation [106]. S1R agonists, such as N, N-Dimethyltryptamine (DMT), have been shown to restore levels of S1R and preserve mitochondrial function. Chronic treatment with this ligand led general neuroprotection as well as a slowing of beta-amyloid accumulation, the hallmark of Alzheimer’s Disease [107,108,109,110,111,112]. A similar effect is seen with pridopidine and PRE-084, other S1R agonists, which restored mitochondrial dysfunction through lowering levels of reactive oxygen species [82,113]. Agonists of S1R also had synergistic neuroprotective roles when combined with acetylcholinesterase inhibitors, the current treatment for AD [114]. Paradoxically, other studies have shown that MAM induction leads to increased amyloid-beta accumulation, which can be reduced with S1R downregulation [115].
Another critical hallmark in the development of AD is deposition of neurofibrillary tangles, characterized by hyperphosphorylated tau protein [116]. In healthy individuals, it has been discovered that S1R assists with maintaining normal levels of phosphorylation on tau proteins [117]. Additional studies show that the presence of functional S1R is imperative to ensure that the development of AD does not take place [118]. Less studied mechanisms to slow the progression of AD include their role in alleviating disruption of the BBB, which may be achieved through increasing levels of vascular endothelial growth factor (VEGF) and low-density lipoprotein receptor-related protein 1 (LRP-1) [119]. Newly studied S2R ligands have also showed promising results in beta-amyloid induced neurologic dysfunction [120]. This impact may be related to restoration of calcium homeostasis offered by sigma-2 ligands [121]. Additionally, reductions in neuroinflammation from S1R have offered promising results in models of Alzheimer’s Disease [111,112]. Oxidative damage plays a critical role in the development of neurologic impairment [122]. PRE-084 can provide antioxidant properties during times of cell stress, offering safety from toxicity and prolonging neuroprotection through preservation of synaptic connections [123,124].
4.2. Sigma-1 Receptor in Demyelinating Disorders
Demyelinating disorders, both inherited and acquired, have been studied in respect to their response to S1R agonism as well. Krabbe Disease is an autosomal recessive disorder marked by neurodegeneration and resultant demyelination [125]. Treatment with donepezil in models of Krabbe Disease had both preservation of myelin as well as a reduction in reactivity of glial cells, contributing to neuroprotection [126]. This remains true for other, rare genetic neurodegenerative disorders such as Wolfram Syndrome and Vanishing White Matter Disease (VWM) [127,128,129,130]. MS is an acquired and autoimmune disorder that leads to inflammation as well as demyelination and loss of neuronal structures [131]. Studies investigatnig models of MS concluded that S1R agonists were able to attenuate worsening clinical course, offering a promising avenue for future clinical indications [103,132]. The mechanism of action for protection in MS is thought to be due to protection of oligodendroglia from apoptosis and reactive oxygen species [83].
4.3. Sigma-1 Receptor in ALS
ALS is a devastating neurodegenerative disease, with the most common inherited form stemming from a mutation in the C9orf72 gene [133]. S1R is critical to maintaining MAMs, and instability of these domains can predispose to the development of ALS [134,135]. Unregulated autophagy can lead to this critical condition, among other neurodegenerative disorders [136]. A common selective serotonin reuptake inhibitor (SSRI), fluvoxamine, is a S1R agonist that has been shown to restore regulation of autophagy in inherited ALS through stabilization of nucleoporins [137]. Other treatments with agonists such as pridopidine, PRE-084, and SA4503 led to improvement of motor behavior and neuroprotection in mouse models of ALS [138,139]. Interestingly, in the same study, BD1063, an antagonist of S1R, had a similar neuroprotective effect [139]. Another mechanism leading to the development of ALS is the accumulation of RAN proteins within the nervous system [140]. Overexpression of S1R led to less accumulation of RAN, offering a new potential mechanism for treatment [141]. Additionally, a common hallmark in ALS are mutations in the Cu/Zn superoxide dismutase (SOD1) gene, leading to accumulation of neurofilaments. Treatment with pridopidine led to a reduction in this buildup, opening new avenues for treatment of this critical disease [142,143].
4.4. Sigma-1 Receptor in Huntington’s Disease
Huntington’s Disease is an inherited neurodegenerative disorder marked by a progressive loss of neurons through one’s lifespan, with resultant debilitating, uncontrolled movements [144]. Like other neurodegenerative disorders, mitochondrial dysfunction is critical to the pathogenesis [145]. In models of HD, pridopidine restored the antioxidant response and decreased levels of reactive oxygen species within the mitochondria [82,146]. Other studies have found that S1R agonism can reduce endoplasmic reticulum stress or restore calcium homeostasis, contributing to attenuation of disease progression [147,148,149,150]. Development of new treatment modalities are critical, as there are no disease-modifying medications for HD available currently. Ongoing clinical trials offer promising results and show less decline in patients [151].
4.5. Sigma-1 Receptor in Parkinson’s Disease
Parkinson’s Disease is one of the most common neurodegenerative disorders characterized by a general slowing of movement as well as tremor and rigidity [152]. Toxic accumulation of alpha-synuclein in the nervous system can lead to mitochondrial dysfunction and the degeneration resulting in this pathology [153]. Accumulation of alpha-synuclein can be worsened with S1R deficiencies, showing its importance in preventing this neurodegenerative disorder [154]. Antagonists of S2R potentially attenuate the alpha-synuclein induced neurodegeneration and offer a novel treatment modality [155]. S1R agonists, alone or when combined with nicotinic agonists, have also offered promising protection of the dopaminergic neurons commonly impacted in PD [156,157]. In addition, there are therapeutic strategies emerging using S1R agonism to restore already damaged neurons in PD [158]. Levodopa is a common treatment in PD, however common side effects include dyskinesia [159]. S1R agonists not only offered neuroprotection but were able to decrease levodopa induced dyskinesia [160,161]. S1R antagonists may also play a role in altering the progression of PD. One study found that S1R inhibits the transient receptor potential canonical (TRPC) channel, which is important for calcium regulation and maintaining cell viability. S1R antagonism let to reversal of this inhibition which resulted in dopaminergic neuroprotection [162]. Other studies show that lower levels of S1R reduce neurotoxicity in dopaminergic cells through suppression of the NMDA receptor (NMDAR) and resulting excitotoxicity [163].
5. Conclusions
In conclusion, as shown in Figure 1, sigma receptors have emerged as critical modulators of cellular homeostasis and neuroprotection. Their widespread expression has paved the road for extensive research into their pathologic and pharmacologic roles. The sigma receptors represent an intricate pharmacologic target with implications in multiple disease states of the nervous system. Their versatile roles across a variety of pathologies emphasizes their potential as novel therapeutic targets in future research. Contradictory findings in certain pathologies warrant the need for temporal and tissue specific modulation to define the conditions in which sigma receptor activation or inhibition is most beneficial. Combination therapies may soon offer use to provide mitigation for presently uncurable diseases.
Supplementary Materials
None.
Author Contributions
Conceptualization, N.D. and D.N.; writing—original draft preparation, N.D. and X.F.; writing—review and editing, D.N.; visualization, X.F. and N.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Dr. Michael Crider, School of Pharmacy at Southern Illinois University Edwardsville, for critical reading and suggestions of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martin, W.R.; Eades, C.G.; Thompson, J.A.; Huppler, R.E.; Gilbert, P.E. The effects of morphine- and nalorphine- like drugs in the nondependent and morphine-dependent chronic spinal dog. J Pharmacol Exp Ther 1976, 197, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.W.; Cook, L. Sigma opiates and certain antipsychotic drugs mutually inhibit (+)-[3H] SKF 10,047 and [3H]haloperidol binding in guinea pig brain membranes. Proc Natl Acad Sci U S A 1984, 81, 5618–5621. [Google Scholar] [CrossRef]
- Su, T.P. Evidence for sigma opioid receptor: binding of [3H]SKF-10047 to etorphine-inaccessible sites in guinea-pig brain. J Pharmacol Exp Ther 1982, 223, 284–290. [Google Scholar] [CrossRef]
- Quirion, R.; Bowen, W.D.; Itzhak, Y.; Junien, J.L.; Musacchio, J.M.; Rothman, R.B.; Su, T.P.; Tam, S.W.; Taylor, D.P. A proposal for the classification of sigma binding sites. Trends Pharmacol Sci 1992, 13, 85–86. [Google Scholar] [CrossRef]
- Hanner, M.; Moebius, F.F.; Flandorfer, A.; Knaus, H.G.; Striessnig, J.; Kempner, E.; Glossmann, H. Purification, molecular cloning, and expression of the mammalian sigma1-binding site. Proc Natl Acad Sci U S A 1996, 93, 8072–8077. [Google Scholar] [CrossRef] [PubMed]
- Alon, A.; Schmidt, H.R.; Wood, M.D.; Sahn, J.J.; Martin, S.F.; Kruse, A.C. Identification of the gene that codes for the sigma(2) receptor. Proc Natl Acad Sci U S A 2017, 114, 7160–7165. [Google Scholar] [CrossRef] [PubMed]
- Su, T.P.; Hayashi, T. Understanding the molecular mechanism of sigma-1 receptors: towards a hypothesis that sigma-1 receptors are intracellular amplifiers for signal transduction. Curr Med Chem 2003, 10, 2073–2080. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T. The sigma receptor: evolution of the concept in neuropsychopharmacology. Curr Neuropharmacol 2005, 3, 267–280. [Google Scholar] [CrossRef]
- Palmer, C.P.; Mahen, R.; Schnell, E.; Djamgoz, M.B.; Aydar, E. Sigma-1 receptors bind cholesterol and remodel lipid rafts in breast cancer cell lines. Cancer Res 2007, 67, 11166–11175. [Google Scholar] [CrossRef]
- Prasad, P.D.; Li, H.W.; Fei, Y.J.; Ganapathy, M.E.; Fujita, T.; Plumley, L.H.; Yang-Feng, T.L.; Leibach, F.H.; Ganapathy, V. Exon-intron structure, analysis of promoter region, and chromosomal localization of the human type 1 sigma receptor gene. J Neurochem 1998, 70, 443–451. [Google Scholar] [CrossRef]
- Kekuda, R.; Prasad, P.D.; Fei, Y.J.; Leibach, F.H.; Ganapathy, V. Cloning and functional expression of the human type 1 sigma receptor (hSigmaR1). Biochem Biophys Res Commun 1996, 229, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Seth, P.; Leibach, F.H.; Ganapathy, V. Cloning and structural analysis of the cDNA and the gene encoding the murine type 1 sigma receptor. Biochem Biophys Res Commun 1997, 241, 535–540. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Zheng, S.; Gurpinar, E.; Koehl, A.; Manglik, A.; Kruse, A.C. Crystal structure of the human σ1 receptor. Nature 2016, 532, 527–530. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Bertoni, M.; Kiefer, F.; Biasini, M.; Bordoli, L.; Schwede, T. Modeling protein quaternary structure of homo- and hetero-oligomers beyond binary interactions by homology. Sci Rep 2017, 7, 10480. [Google Scholar] [CrossRef]
- Hong, W.C. Distinct Regulation of sigma (1) Receptor Multimerization by Its Agonists and Antagonists in Transfected Cells and Rat Liver Membranes. J Pharmacol Exp Ther 2020, 373, 290–301. [Google Scholar] [CrossRef]
- Navarro, G.; Moreno, E.; Bonaventura, J.; Brugarolas, M.; Farré, D.; Aguinaga, D.; Mallol, J.; Cortés, A.; Casadó, V.; Lluís, C.; et al. Cocaine inhibits dopamine D2 receptor signaling via sigma-1-D2 receptor heteromers. PLoS One 2013, 8, e61245. [Google Scholar] [CrossRef] [PubMed]
- Aguinaga, D.; Medrano, M.; Vega-Quiroga, I.; Gysling, K.; Canela, E.I.; Navarro, G.; Franco, R. Cocaine Effects on Dopaminergic Transmission Depend on a Balance between Sigma-1 and Sigma-2 Receptor Expression. Front Mol Neurosci 2018, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Fontanilla, D.; Hajipour, A.R.; Pal, A.; Chu, U.B.; Arbabian, M.; Ruoho, A.E. Probing the steroid binding domain-like I (SBDLI) of the sigma-1 receptor binding site using N-substituted photoaffinity labels. Biochemistry 2008, 47, 7205–7217. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptors (sigma(1) binding sites) form raft-like microdomains and target lipid droplets on the endoplasmic reticulum: roles in endoplasmic reticulum lipid compartmentalization and export. J Pharmacol Exp Ther 2003, 306, 718–725. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptors at galactosylceramide-enriched lipid microdomains regulate oligodendrocyte differentiation. Proc Natl Acad Sci U S A 2004, 101, 14949–14954. [Google Scholar] [CrossRef]
- Gebreselassie, D.; Bowen, W.D. Sigma-2 receptors are specifically localized to lipid rafts in rat liver membranes. Eur J Pharmacol 2004, 493, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zeng, C.; Chu, W.; Pan, F.; Rothfuss, J.M.; Zhang, F.; Tu, Z.; Zhou, D.; Zeng, D.; Vangveravong, S.; et al. Identification of the PGRMC1 protein complex as the putative sigma-2 receptor binding site. Nat Commun 2011, 2, 380. [Google Scholar] [CrossRef]
- Vaupel, D.B. Naltrexone fails to antagonize the sigma effects of PCP and SKF 10,047 in the dog. Eur J Pharmacol 1983, 92, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Chu, U.B.; Ruoho, A.E. Sigma Receptor Binding Assays. Curr Protoc Pharmacol 2015, 71, 1.34–31. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.P. Intracellular dynamics of sigma-1 receptors (sigma(1) binding sites) in NG108-15 cells. J Pharmacol Exp Ther 2003, 306, 726–733. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca(2+) signaling and cell survival. Cell 2007, 131, 596–610. [Google Scholar] [CrossRef]
- Aydar, E.; Palmer, C.P.; Klyachko, V.A.; Jackson, M.B. The sigma receptor as a ligand-regulated auxiliary potassium channel subunit. Neuron 2002, 34, 399–410. [Google Scholar] [CrossRef]
- Su, T.P.; Hayashi, T.; Maurice, T.; Buch, S.; Ruoho, A.E. The sigma-1 receptor chaperone as an inter-organelle signaling modulator. Trends Pharmacol Sci 2010, 31, 557–566. [Google Scholar] [CrossRef]
- Brimson, J.M.; Brown, C.A.; Safrany, S.T. Antagonists show GTP-sensitive high-affinity binding to the sigma-1 receptor. Br J Pharmacol 2011, 164, 772–780. [Google Scholar] [CrossRef]
- Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Herrero-Labrador, R.; Burgueño, J.; Zamanillo, D.; Garzón, J. The calcium-sensitive Sigma-1 receptor prevents cannabinoids from provoking glutamate NMDA receptor hypofunction: implications in antinociception and psychotic diseases. Int J Neuropsychopharmacol 2014, 17, 1943–1955. [Google Scholar] [CrossRef]
- Vilner, B.J.; John, C.S.; Bowen, W.D. Sigma-1 and sigma-2 receptors are expressed in a wide variety of human and rodent tumor cell lines. Cancer Res 1995, 55, 408–413. [Google Scholar]
- Bermack, J.E.; Debonnel, G. Distinct modulatory roles of sigma receptor subtypes on glutamatergic responses in the dorsal hippocampus. Synapse 2005, 55, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Cobos, E.J.; Entrena, J.M.; Nieto, F.R.; Cendán, C.M.; Del Pozo, E. Pharmacology and therapeutic potential of sigma(1) receptor ligands. Curr Neuropharmacol 2008, 6, 344–366. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Maurice, T.; Su, T.P. Ca(2+) signaling via sigma(1)-receptors: novel regulatory mechanism affecting intracellular Ca(2+) concentration. J Pharmacol Exp Ther 2000, 293, 788–798. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Regulating ankyrin dynamics: Roles of sigma-1 receptors. Proc Natl Acad Sci U S A 2001, 98, 491–496. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptor ligands: potential in the treatment of neuropsychiatric disorders. CNS Drugs 2004, 18, 269–284. [Google Scholar] [CrossRef]
- Hayashi, T.; Fujimoto, M. Detergent-resistant microdomains determine the localization of sigma-1 receptors to the endoplasmic reticulum-mitochondria junction. Mol Pharmacol 2010, 77, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Carnally, S.M.; Johannessen, M.; Henderson, R.M.; Jackson, M.B.; Edwardson, J.M. Demonstration of a direct interaction between sigma-1 receptors and acid-sensing ion channels. Biophys J 2010, 98, 1182–1191. [Google Scholar] [CrossRef]
- Crottès, D.; Martial, S.; Rapetti-Mauss, R.; Pisani, D.F.; Loriol, C.; Pellissier, B.; Martin, P.; Chevet, E.; Borgese, F.; Soriani, O. Sig1R protein regulates hERG channel expression through a post-translational mechanism in leukemic cells. J Biol Chem 2011, 286, 27947–27958. [Google Scholar] [CrossRef]
- Balasuriya, D.; Stewart, A.P.; Crottès, D.; Borgese, F.; Soriani, O.; Edwardson, J.M. The sigma-1 receptor binds to the Nav1.5 voltage-gated Na+ channel with 4-fold symmetry. J Biol Chem 2012, 287, 37021–37029. [Google Scholar] [CrossRef] [PubMed]
- Kourrich, S.; Hayashi, T.; Chuang, J.Y.; Tsai, S.Y.; Su, T.P.; Bonci, A. Dynamic interaction between sigma-1 receptor and Kv1.2 shapes neuronal and behavioral responses to cocaine. Cell 2013, 152, 236–247. [Google Scholar] [CrossRef]
- Burnett, M.G.; Zager, E.L. Pathophysiology of peripheral nerve injury: a brief review. Neurosurg Focus 2004, 16, E1. [Google Scholar] [CrossRef]
- Lee, R.H.C.; Lee, M.H.H.; Wu, C.Y.C.; Couto, E.S.A.; Possoit, H.E.; Hsieh, T.H.; Minagar, A.; Lin, H.W. Cerebral ischemia and neuroregeneration. Neural Regen Res 2018, 13, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Lee, A.Y.W. Traumatic Brain Injuries: Pathophysiology and Potential Therapeutic Targets. Front Cell Neurosci 2019, 13, 528. [Google Scholar] [CrossRef]
- Schonfeld, E.; Johnstone, T.M.; Haider, G.; Shah, A.; Marianayagam, N.J.; Biswal, S.; Veeravagu, A. Sigma-1 receptor expression in a subpopulation of lumbar spinal cord microglia in response to peripheral nerve injury. Sci Rep 2023, 13, 14762. [Google Scholar] [CrossRef]
- Nardai, S.; László, M.; Szabó, A.; Alpár, A.; Hanics, J.; Zahola, P.; Merkely, B.; Frecska, E.; Nagy, Z. N,N-dimethyltryptamine reduces infarct size and improves functional recovery following transient focal brain ischemia in rats. Exp Neurol 2020, 327, 113245. [Google Scholar] [CrossRef]
- Zhai, M.; Liu, C.; Li, Y.; Zhang, P.; Yu, Z.; Zhu, H.; Zhang, L.; Zhang, Q.; Wang, J.; Wang, J. Dexmedetomidine inhibits neuronal apoptosis by inducing Sigma-1 receptor signaling in cerebral ischemia-reperfusion injury. Aging (Albany NY) 2019, 11, 9556–9568. [Google Scholar] [CrossRef] [PubMed]
- Morihara, R.; Yamashita, T.; Liu, X.; Nakano, Y.; Fukui, Y.; Sato, K.; Ohta, Y.; Hishikawa, N.; Shang, J.; Abe, K. Protective effect of a novel sigma-1 receptor agonist is associated with reduced endoplasmic reticulum stress in stroke male mice. J Neurosci Res 2018, 96, 1707–1716. [Google Scholar] [CrossRef]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Saunders, D.E.; Clifton, A.G.; Brown, M.M. Measurement of infarct size using MRI predicts prognosis in middle cerebral artery infarction. Stroke 1995, 26, 2272–2276. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, D.A.; Dalwadi, D.; Kim, S.; Metzger, D.; Oppong-Gyebi, A.; Das-Earl, P.; Schetz, J.A. Treatment of Stroke at a Delayed Timepoint with a Repurposed Drug Targeting Sigma 1 Receptors. Transl Stroke Res 2024, 15, 1035–1049. [Google Scholar] [CrossRef]
- Song, W.; Yao, Y.; Zhang, H.; Hao, X.; Zhou, L.; Song, Z.; Wei, T.; Chi, T.; Liu, P.; Ji, X.; et al. Sigma-1 Receptor Activation Improves Oligodendrogenesis and Promotes White-Matter Integrity after Stroke in Mice with Diabetic Mellitus. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Wang, R.; Jiang, M.; Ye, Q.; Smith, A.D.; Chen, J.; Shi, Y. Macrophages reprogram after ischemic stroke and promote efferocytosis and inflammation resolution in the mouse brain. CNS Neurosci Ther 2019, 25, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, Q.; Tao, W.; Qin, P.; Chen, J.; Yang, H.; Chen, J.; Liu, H.; Dai, Q.; Zhen, X. Sigma-1 receptor-regulated efferocytosis by infiltrating circulating macrophages/microglial cells protects against neuronal impairments and promotes functional recovery in cerebral ischemic stroke. Theranostics 2023, 13, 543–559. [Google Scholar] [CrossRef]
- Yabuki, Y.; Shinoda, Y.; Izumi, H.; Ikuno, T.; Shioda, N.; Fukunaga, K. Dehydroepiandrosterone administration improves memory deficits following transient brain ischemia through sigma-1 receptor stimulation. Brain Res 2015, 1622, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Suzuki, H.; Travis, Z.D.; Zhang, J.H. The Stroke-Induced Blood-Brain Barrier Disruption: Current Progress of Inspection Technique, Mechanism, and Therapeutic Target. Curr Neuropharmacol 2020, 18, 1187–1212. [Google Scholar] [CrossRef]
- Liu, D.; Yang, L.; Liu, P.; Ji, X.; Qi, X.; Wang, Z.; Chi, T.; Zou, L. Sigma-1 receptor activation alleviates blood-brain barrier disruption post cerebral ischemia stroke by stimulating the GDNF-GFRα1-RET pathway. Exp Neurol 2022, 347, 113867. [Google Scholar] [CrossRef]
- Zhao, Q.; Yu, S.; Ling, Y.; Hao, S.; Liu, J. The Protective Effects of Dexmedetomidine against Hypoxia/Reoxygenation-Induced Inflammatory Injury and Permeability in Brain Endothelial Cells Mediated by Sigma-1 Receptor. ACS Chem Neurosci 2021, 12, 1940–1947. [Google Scholar] [CrossRef]
- Yang, S.; Jin, H.; Zhu, Y.; Wan, Y.; Opoku, E.N.; Zhu, L.; Hu, B. Diverse Functions and Mechanisms of Pericytes in Ischemic Stroke. Curr Neuropharmacol 2017, 15, 892–905. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Wei, Q.; Leng, S.; Li, C.; Han, B.; Bai, Y.; Zhang, H.; Yao, H. Activation of Sigma-1 Receptor Enhanced Pericyte Survival via the Interplay Between Apoptosis and Autophagy: Implications for Blood-Brain Barrier Integrity in Stroke. Transl Stroke Res 2020, 11, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Dohmen, C.; Sakowitz, O.W.; Fabricius, M.; Bosche, B.; Reithmeier, T.; Ernestus, R.I.; Brinker, G.; Dreier, J.P.; Woitzik, J.; Strong, A.J.; et al. Spreading depolarizations occur in human ischemic stroke with high incidence. Ann Neurol 2008, 63, 720–728. [Google Scholar] [CrossRef]
- Leao, A.A. The slow voltage variation of cortical spreading depression of activity. Electroencephalogr Clin Neurophysiol 1951, 3, 315–321. [Google Scholar] [CrossRef]
- Szabó, Í.; Varga, V.; Dvorácskó, S.; Farkas, A.E.; Körmöczi, T.; Berkecz, R.; Kecskés, S.; Menyhárt, Á.; Frank, R.; Hantosi, D.; et al. N,N-Dimethyltryptamine attenuates spreading depolarization and restrains neurodegeneration by sigma-1 receptor activation in the ischemic rat brain. Neuropharmacology 2021, 192, 108612. [Google Scholar] [CrossRef]
- Soares, R.O.S.; Losada, D.M.; Jordani, M.C.; Evora, P.; Castro, E.S.O. Ischemia/Reperfusion Injury Revisited: An Overview of the Latest Pharmacological Strategies. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Dai, L.A.; Chen, X.Y.; Li, W.J.; Yang, J.H.; Lin, M.J.; Li, X.S.; Zeng, Y.F.; Chen, S.W.; Xie, Z.L.; Zhu, Z.L.; et al. Sigma-1 Receptor and Binding Immunoglobulin Protein Interact with Ulinastatin Contributing to a Protective Effect in Rat Cerebral Ischemia/Reperfusion. World Neurosurg 2022, 158, e488–e494. [Google Scholar] [CrossRef]
- Stelfa, G.; Vavers, E.; Svalbe, B.; Serzants, R.; Miteniece, A.; Lauberte, L.; Grinberga, S.; Gukalova, B.; Dambrova, M.; Zvejniece, L. Reduced GFAP Expression in Bergmann Glial Cells in the Cerebellum of Sigma-1 Receptor Knockout Mice Determines the Neurobehavioral Outcomes after Traumatic Brain Injury. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Liu, L.; Min, X.; Mi, L.; Chai, Y.; Chen, F.; Wang, J.; Yue, S.; Zhang, J.; Deng, Q.; et al. Activation of Sigma-1 Receptor Alleviates ER-Associated Cell Death and Microglia Activation in Traumatically Injured Mice. J Clin Med 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Ma, Y.; Ren, Z.; Xu, B.; Zhang, Y.; Chen, J.; Yang, B. Sigma-1 Receptor Modulates Neuroinflammation After Traumatic Brain Injury. Cell Mol Neurobiol 2016, 36, 639–645. [Google Scholar] [CrossRef]
- Vázquez-Rosa, E.; Watson, M.R.; Sahn, J.J.; Hodges, T.R.; Schroeder, R.E.; Cintrón-Pérez, C.J.; Shin, M.K.; Yin, T.C.; Emery, J.L.; Martin, S.F.; et al. Neuroprotective Efficacy of a Sigma 2 Receptor/TMEM97 Modulator (DKR-1677) after Traumatic Brain Injury. ACS Chem Neurosci 2019, 10, 1595–1602. [Google Scholar] [CrossRef]
- Tan, R.; Sui, C.; Diao, Y.; Shi, G.; Hu, X.; Hao, Z.; Li, C.; Hao, M.; Xie, M.; Zhu, T. Activation of the sigma-1 receptor ameliorates neuronal ferroptosis via IRE1α after spinal cord injury. Brain Res 2024, 1838, 149011. [Google Scholar] [CrossRef] [PubMed]
- Gaja-Capdevila, N.; Hernández, N.; Yeste, S.; Reinoso, R.F.; Burgueño, J.; Montero, A.; Merlos, M.; Vela, J.M.; Herrando-Grabulosa, M.; Navarro, X. EST79232 and EST79376, Two Novel Sigma-1 Receptor Ligands, Exert Neuroprotection on Models of Motoneuron Degeneration. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Kenny, E.M.; Fidan, E.; Yang, Q.; Anthonymuthu, T.S.; New, L.A.; Meyer, E.A.; Wang, H.; Kochanek, P.M.; Dixon, C.E.; Kagan, V.E.; et al. Ferroptosis Contributes to Neuronal Death and Functional Outcome After Traumatic Brain Injury. Crit Care Med 2019, 47, 410–418. [Google Scholar] [CrossRef]
- Maurice, T.; Su, T.P. The pharmacology of sigma-1 receptors. Pharmacol Ther 2009, 124, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Urani, A.; Romieu, P.; Roman, F.J.; Maurice, T. Enhanced antidepressant effect of sigma(1) (sigma(1)) receptor agonists in beta(25-35)-amyloid peptide-treated mice. Behav Brain Res 2002, 134, 239–247. [Google Scholar] [CrossRef]
- Genovese, I.; Giamogante, F.; Barazzuol, L.; Battista, T.; Fiorillo, A.; Vicario, M.; D'Alessandro, G.; Cipriani, R.; Limatola, C.; Rossi, D.; et al. Sorcin is an early marker of neurodegeneration, Ca(2+) dysregulation and endoplasmic reticulum stress associated to neurodegenerative diseases. Cell Death Dis 2020, 11, 861. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, L.; Halliday, G.; Dobson-Stone, C.; Wang, Y.; Tang, H.D.; Cao, L.; Deng, Y.L.; Wang, G.; Zhang, Y.M.; et al. Genetic polymorphisms in sigma-1 receptor and apolipoprotein E interact to influence the severity of Alzheimer's disease. Curr Alzheimer Res 2011, 8, 765–770. [Google Scholar] [CrossRef]
- Luty, A.A.; Kwok, J.B.; Dobson-Stone, C.; Loy, C.T.; Coupland, K.G.; Karlström, H.; Sobow, T.; Tchorzewska, J.; Maruszak, A.; Barcikowska, M.; et al. Sigma nonopioid intracellular receptor 1 mutations cause frontotemporal lobar degeneration-motor neuron disease. Ann Neurol 2010, 68, 639–649. [Google Scholar] [CrossRef]
- Al-Saif, A.; Al-Mohanna, F.; Bohlega, S. A mutation in sigma-1 receptor causes juvenile amyotrophic lateral sclerosis. Ann Neurol 2011, 70, 913–919. [Google Scholar] [CrossRef]
- Snyder, M.A.; McCann, K.; Lalande, M.J.; Thivierge, J.P.; Bergeron, R. Sigma receptor type 1 knockout mice show a mild deficit in plasticity but no significant change in synaptic transmission in the CA1 region of the hippocampus. J Neurochem 2016, 138, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Naia, L.; Ly, P.; Mota, S.I.; Lopes, C.; Maranga, C.; Coelho, P.; Gershoni-Emek, N.; Ankarcrona, M.; Geva, M.; Hayden, M.R.; et al. The Sigma-1 Receptor Mediates Pridopidine Rescue of Mitochondrial Function in Huntington Disease Models. Neurotherapeutics 2021, 18, 1017–1038. [Google Scholar] [CrossRef]
- Lisak, R.P.; Nedelkoska, L.; Benjamins, J.A. Sigma-1 receptor agonists as potential protective therapies in multiple sclerosis. J Neuroimmunol 2020, 342, 577188. [Google Scholar] [CrossRef] [PubMed]
- Francardo, V. Sigma-1 receptor: a potential new target for Parkinson's disease? Neural Regen Res 2014, 9, 1882–1883. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Kaushal, N.; Robson, M.J.; Matsumoto, R.R. Sigma receptors as potential therapeutic targets for neuroprotection. Eur J Pharmacol 2014, 743, 42–47. [Google Scholar] [CrossRef]
- Mondal, S.; Hegarty, E.; Sahn, J.J.; Scott, L.L.; Gökçe, S.K.; Martin, C.; Ghorashian, N.; Satarasinghe, P.N.; Iyer, S.; Sae-Lee, W.; et al. High-Content Microfluidic Screening Platform Used To Identify σ2R/Tmem97 Binding Ligands that Reduce Age-Dependent Neurodegeneration in C. elegans SC_APP Model. ACS Chem Neurosci 2018, 9, 1014–1026. [Google Scholar] [CrossRef]
- Lenoir, S.; Lahaye, R.A.; Vitet, H.; Scaramuzzino, C.; Virlogeux, A.; Capellano, L.; Genoux, A.; Gershoni-Emek, N.; Geva, M.; Hayden, M.R.; et al. Pridopidine rescues BDNF/TrkB trafficking dynamics and synapse homeostasis in a Huntington disease brain-on-a-chip model. Neurobiol Dis 2022, 173, 105857. [Google Scholar] [CrossRef]
- Geva, M.; Kusko, R.; Soares, H.; Fowler, K.D.; Birnberg, T.; Barash, S.; Wagner, A.M.; Fine, T.; Lysaght, A.; Weiner, B.; et al. Pridopidine activates neuroprotective pathways impaired in Huntington Disease. Hum Mol Genet 2016, 25, 3975–3987. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat Rev Neurosci 2013, 14, 401–416. [Google Scholar] [CrossRef]
- Goyagi, T.; Goto, S.; Bhardwaj, A.; Dawson, V.L.; Hurn, P.D.; Kirsch, J.R. Neuroprotective effect of sigma(1)-receptor ligand 4-phenyl-1-(4-phenylbutyl) piperidine (PPBP) is linked to reduced neuronal nitric oxide production. Stroke 2001, 32, 1613–1620. [Google Scholar] [CrossRef]
- Vagnerova, K.; Hurn, P.D.; Bhardwaj, A.; Kirsch, J.R. Sigma 1 receptor agonists act as neuroprotective drugs through inhibition of inducible nitric oxide synthase. Anesth Analg 2006, 103, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Zampieri, D.; Fortuna, S.; Calabretti, A.; Romano, M.; Menegazzi, R.; Schepmann, D.; Wünsch, B.; Collina, S.; Zanon, D.; Mamolo, M.G. Discovery of new potent dual sigma receptor/GluN2b ligands with antioxidant property as neuroprotective agents. Eur J Med Chem 2019, 180, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Rui, M.; Rossino, G.; Coniglio, S.; Monteleone, S.; Scuteri, A.; Malacrida, A.; Rossi, D.; Catenacci, L.; Sorrenti, M.; Paolillo, M.; et al. Identification of dual Sigma1 receptor modulators/acetylcholinesterase inhibitors with antioxidant and neurotrophic properties, as neuroprotective agents. Eur J Med Chem 2018, 158, 353–370. [Google Scholar] [CrossRef]
- Su, T.C.; Lin, S.H.; Lee, P.T.; Yeh, S.H.; Hsieh, T.H.; Chou, S.Y.; Su, T.P.; Hung, J.J.; Chang, W.C.; Lee, Y.C.; et al. The sigma-1 receptor-zinc finger protein 179 pathway protects against hydrogen peroxide-induced cell injury. Neuropharmacology 2016, 105, 1–9. [Google Scholar] [CrossRef]
- Turrigiano, G. Homeostatic synaptic plasticity: local and global mechanisms for stabilizing neuronal function. Cold Spring Harb Perspect Biol 2012, 4, a005736. [Google Scholar] [CrossRef]
- Pozo, K.; Goda, Y. Unraveling mechanisms of homeostatic synaptic plasticity. Neuron 2010, 66, 337–351. [Google Scholar] [CrossRef]
- Szczepańska, K.; Bojarski, A.J.; Popik, P.; Malikowska-Racia, N. Novel object recognition test as an alternative approach to assessing the pharmacological profile of sigma-1 receptor ligands. Pharmacol Rep 2023, 75, 1291–1298. [Google Scholar] [CrossRef]
- Sahlholm, K.; Valle-León, M.; Fernández-Dueñas, V.; Ciruela, F. Pridopidine Reverses Phencyclidine-Induced Memory Impairment. Front Pharmacol 2018, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl Neurodegener 2020, 9, 42. [Google Scholar] [CrossRef]
- Iwamoto, M.; Nakamura, Y.; Takemura, M.; Hisaoka-Nakashima, K.; Morioka, N. TLR4-TAK1-p38 MAPK pathway and HDAC6 regulate the expression of sigma-1 receptors in rat primary cultured microglia. J Pharmacol Sci 2020, 144, 23–29. [Google Scholar] [CrossRef]
- Orciani, C.; Do Carmo, S.; Foret, M.K.; Hall, H.; Bonomo, Q.; Lavagna, A.; Huang, C.; Cuello, A.C. Early treatment with an M1 and sigma-1 receptor agonist prevents cognitive decline in a transgenic rat model displaying Alzheimer-like amyloid pathology. Neurobiol Aging 2023, 132, 220–232. [Google Scholar] [CrossRef]
- Ji, J.; Gao, C.; Wang, Q.; Jia, X.; Tian, H.; Wei, Y.; Liu, Z.; Wang, Y.; Guo, L. The sigma-1 receptor-TAMM41 axis modulates neuroinflammation and attenuates memory impairment during the latent period of epileptogenesis. Animal Model Exp Med 2025, 8, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Gao, T.; Jia, X.; Gao, C.; Tian, H.; Wei, Y.; Lu, W.; Liu, Z.; Wang, Y. SKF83959 Attenuates Memory Impairment and Depressive-like Behavior during the Latent Period of Epilepsy via Allosteric Activation of the Sigma-1 Receptor. ACS Chem Neurosci 2022, 13, 3198–3209. [Google Scholar] [CrossRef]
- Heiss, K.; Raffaele, M.; Vanella, L.; Murabito, P.; Prezzavento, O.; Marrazzo, A.; Aricò, G.; Castracani, C.C.; Barbagallo, I.; Zappalà, A.; et al. (+)-Pentazocine attenuates SH-SY5Y cell death, oxidative stress and microglial migration induced by conditioned medium from activated microglia. Neurosci Lett 2017, 642, 86–90. [Google Scholar] [CrossRef]
- Johri, A.; Beal, M.F. Mitochondrial dysfunction in neurodegenerative diseases. J Pharmacol Exp Ther 2012, 342, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Cavendish, J.Z.; Sarkar, S.N.; Colantonio, M.A.; Quintana, D.D.; Ahmed, N.; White, B.A.; Engler-Chiurazzi, E.B.; Simpkins, J.W. Mitochondrial Movement and Number Deficits in Embryonic Cortical Neurons from 3xTg-AD Mice. J Alzheimers Dis 2019, 70, 139–151. [Google Scholar] [CrossRef]
- Cheng, D.; Lei, Z.G.; Chu, K.; Lam, O.J.H.; Chiang, C.Y.; Zhang, Z.J. N, N-Dimethyltryptamine, a natural hallucinogen, ameliorates Alzheimer's disease by restoring neuronal Sigma-1 receptor-mediated endoplasmic reticulum-mitochondria crosstalk. Alzheimers Res Ther 2024, 16, 95. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; LeVine, H. , 3rd. Alzheimer's disease and the amyloid-beta peptide. J Alzheimers Dis 2010, 19, 311–323. [Google Scholar] [CrossRef]
- Borbély, E.; Varga, V.; Szögi, T.; Schuster, I.; Bozsó, Z.; Penke, B.; Fülöp, L. Impact of Two Neuronal Sigma-1 Receptor Modulators, PRE084 and DMT, on Neurogenesis and Neuroinflammation in an Aβ(1-42)-Injected, Wild-Type Mouse Model of AD. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Maurice, T.; Volle, J.N.; Strehaiano, M.; Crouzier, L.; Pereira, C.; Kaloyanov, N.; Virieux, D.; Pirat, J.L. Neuroprotection in non-transgenic and transgenic mouse models of Alzheimer's disease by positive modulation of σ(1) receptors. Pharmacol Res 2019, 144, 315–330. [Google Scholar] [CrossRef]
- Hall, H.; Iulita, M.F.; Gubert, P.; Flores Aguilar, L.; Ducatenzeiler, A.; Fisher, A.; Cuello, A.C. AF710B, an M1/sigma-1 receptor agonist with long-lasting disease-modifying properties in a transgenic rat model of Alzheimer's disease. Alzheimers Dement 2018, 14, 811–823. [Google Scholar] [CrossRef]
- Fisher, A.; Bezprozvanny, I.; Wu, L.; Ryskamp, D.A.; Bar-Ner, N.; Natan, N.; Brandeis, R.; Elkon, H.; Nahum, V.; Gershonov, E.; et al. AF710B, a Novel M1/σ1 Agonist with Therapeutic Efficacy in Animal Models of Alzheimer’s Disease. Neurodegener Dis 2016, 16, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Lahmy, V.; Long, R.; Morin, D.; Villard, V.; Maurice, T. Mitochondrial protection by the mixed muscarinic/σ1 ligand ANAVEX2-73, a tetrahydrofuran derivative, in Aβ25-35 peptide-injected mice, a nontransgenic Alzheimer's disease model. Front Cell Neurosci 2014, 8, 463. [Google Scholar] [CrossRef] [PubMed]
- Freyssin, A.; Carles, A.; Guehairia, S.; Rubinstenn, G.; Maurice, T. Fluoroethylnormemantine (FENM) shows synergistic protection in combination with a sigma-1 receptor agonist in a mouse model of Alzheimer's disease. Neuropharmacology 2024, 242, 109733. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, R.; Black, S.E.; Lotlikar, M.S.; Fenn, R.H.; Jorfi, M.; Kovacs, D.M.; Tanzi, R.E. Axonal generation of amyloid-β from palmitoylated APP in mitochondria-associated endoplasmic reticulum membranes. Cell Rep 2021, 35, 109134. [Google Scholar] [CrossRef]
- Rawat, P.; Sehar, U.; Bisht, J.; Selman, A.; Culberson, J.; Reddy, P.H. Phosphorylated Tau in Alzheimer's Disease and Other Tauopathies. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Pokrass, M.J.; Klauer, N.R.; Nohara, H.; Su, T.P. Sigma-1 receptor regulates Tau phosphorylation and axon extension by shaping p35 turnover via myristic acid. Proc Natl Acad Sci U S A 2015, 112, 6742–6747. [Google Scholar] [CrossRef]
- Maurice, T.; Strehaiano, M.; Duhr, F.; Chevallier, N. Amyloid toxicity is enhanced after pharmacological or genetic invalidation of the σ(1) receptor. Behav Brain Res 2018, 339, 1–10. [Google Scholar] [CrossRef]
- An, Y.; Qi, Y.; Li, Y.; Li, Z.; Yang, C.; Jia, D. Activation of the sigma-1 receptor attenuates blood-brain barrier disruption by inhibiting amyloid deposition in Alzheimer's disease mice. Neurosci Lett 2022, 774, 136528. [Google Scholar] [CrossRef]
- Christmann, U.; Díaz, J.L.; Pascual, R.; Bordas, M.; Álvarez, I.; Monroy, X.; Porras, M.; Yeste, S.; Reinoso, R.F.; Merlos, M.; et al. Discovery of WLB-89462, a New Drug-like and Highly Selective σ(2) Receptor Ligand with Neuroprotective Properties. J Med Chem 2023, 66, 12499–12519. [Google Scholar] [CrossRef]
- Yi, B.; Sahn, J.J.; Ardestani, P.M.; Evans, A.K.; Scott, L.L.; Chan, J.Z.; Iyer, S.; Crisp, A.; Zuniga, G.; Pierce, J.T.; et al. Small molecule modulator of sigma 2 receptor is neuroprotective and reduces cognitive deficits and neuroinflammation in experimental models of Alzheimer's disease. J Neurochem 2017, 140, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: a review of upstream and downstream antioxidant therapeutic options. Curr Neuropharmacol 2009, 7, 65–74. [Google Scholar] [CrossRef]
- Lasbleiz, C.; Peyrel, A.; Tarot, P.; Sarniguet, J.; Crouzier, L.; Cubedo, N.; Delprat, B.; Rossel, M.; Maurice, T.; Liévens, J.C. Sigma-1 receptor agonist PRE-084 confers protection against TAR DNA-binding protein-43 toxicity through NRF2 signalling. Redox Biol 2022, 58, 102542. [Google Scholar] [CrossRef] [PubMed]
- Estévez-Silva, H.M.; Cuesto, G.; Romero, N.; Brito-Armas, J.M.; Acevedo-Arozena, A.; Acebes, Á.; Marcellino, D.J. Pridopidine Promotes Synaptogenesis and Reduces Spatial Memory Deficits in the Alzheimer's Disease APP/PS1 Mouse Model. Neurotherapeutics 2022, 19, 1566–1587. [Google Scholar] [CrossRef]
- Suzuki, K.; Suzuki, Y. Globoid cell leucodystrophy (Krabbe's disease): deficiency of galactocerebroside beta-galactosidase. Proc Natl Acad Sci U S A 1970, 66, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Papakyriakopoulou, P.; Valsami, G.; Dev, K.K. The Effect of Donepezil Hydrochloride in the Twitcher Mouse Model of Krabbe Disease. Mol Neurobiol 2024, 61, 8688–8701. [Google Scholar] [CrossRef]
- Barrett, T.G.; Bundey, S.E. Wolfram (DIDMOAD) syndrome. J Med Genet 1997, 34, 838–841. [Google Scholar] [CrossRef]
- Fischer, T.T.; Nguyen, L.D.; Ehrlich, B.E. Boosting ER-mitochondria calcium transfer to treat Wolfram syndrome. Cell Calcium 2022, 104, 102572. [Google Scholar] [CrossRef]
- Crouzier, L.; Danese, A.; Yasui, Y.; Richard, E.M.; Liévens, J.C.; Patergnani, S.; Couly, S.; Diez, C.; Denus, M.; Cubedo, N.; et al. Activation of the sigma-1 receptor chaperone alleviates symptoms of Wolfram syndrome in preclinical models. Sci Transl Med 2022, 14, eabh3763. [Google Scholar] [CrossRef]
- Atzmon, A.; Herrero, M.; Sharet-Eshed, R.; Gilad, Y.; Senderowitz, H.; Elroy-Stein, O. Drug Screening Identifies Sigma-1-Receptor as a Target for the Therapy of VWM Leukodystrophy. Front Mol Neurosci 2018, 11, 336. [Google Scholar] [CrossRef]
- Noyes, K.; Weinstock-Guttman, B. Impact of diagnosis and early treatment on the course of multiple sclerosis. Am J Manag Care 2013, 19, s321–331. [Google Scholar] [PubMed]
- Collina, S.; Rui, M.; Stotani, S.; Bignardi, E.; Rossi, D.; Curti, D.; Giordanetto, F.; Malacrida, A.; Scuteri, A.; Cavaletti, G. Are sigma receptor modulators a weapon against multiple sclerosis disease? Future Med Chem 2017, 9, 2029–2051. [Google Scholar] [CrossRef] [PubMed]
- Renton, A.E.; Majounie, E.; Waite, A.; Simon-Sanchez, J.; Rollinson, S.; Gibbs, J.R.; Schymick, J.C.; Laaksovirta, H.; van Swieten, J.C.; Myllykangas, L.; et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron 2011, 72, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Ilieva, H.; Tamada, H.; Nomura, H.; Komine, O.; Endo, F.; Jin, S.; Mancias, P.; Kiyama, H.; Yamanaka, K. Mitochondria-associated membrane collapse is a common pathomechanism in SIGMAR1- and SOD1-linked ALS. EMBO Mol Med 2016, 8, 1421–1437. [Google Scholar] [CrossRef]
- Watanabe, S.; Horiuchi, M.; Murata, Y.; Komine, O.; Kawade, N.; Sobue, A.; Yamanaka, K. Sigma-1 receptor maintains ATAD3A as a monomer to inhibit mitochondrial fragmentation at the mitochondria-associated membrane in amyotrophic lateral sclerosis. Neurobiol Dis 2023, 179, 106031. [Google Scholar] [CrossRef]
- Chua, J.P.; De Calbiac, H.; Kabashi, E.; Barmada, S.J. Autophagy and ALS: mechanistic insights and therapeutic implications. Autophagy 2022, 18, 254–282. [Google Scholar] [CrossRef]
- Lin, C.Y.; Wu, H.E.; Weng, E.F.; Wu, H.C.; Su, T.P.; Wang, S.M. Fluvoxamine Exerts Sigma-1R to Rescue Autophagy via Pom121-Mediated Nucleocytoplasmic Transport of TFEB. Mol Neurobiol 2024, 61, 5282–5294. [Google Scholar] [CrossRef]
- Estévez-Silva, H.M.; Mediavilla, T.; Giacobbo, B.L.; Liu, X.; Sultan, F.R.; Marcellino, D.J. Pridopidine modifies disease phenotype in a SOD1 mouse model of amyotrophic lateral sclerosis. Eur J Neurosci 2022, 55, 1356–1372. [Google Scholar] [CrossRef]
- Gaja-Capdevila, N.; Hernández, N.; Navarro, X.; Herrando-Grabulosa, M. Sigma-1 Receptor is a Pharmacological Target to Promote Neuroprotection in the SOD1(G93A) ALS Mice. Front Pharmacol 2021, 12, 780588. [Google Scholar] [CrossRef]
- Zu, T.; Liu, Y.; Banez-Coronel, M.; Reid, T.; Pletnikova, O.; Lewis, J.; Miller, T.M.; Harms, M.B.; Falchook, A.E.; Subramony, S.H.; et al. RAN proteins and RNA foci from antisense transcripts in C9ORF72 ALS and frontotemporal dementia. Proc Natl Acad Sci U S A 2013, 110, E4968–4977. [Google Scholar] [CrossRef]
- Lee, P.T.; Liévens, J.C.; Wang, S.M.; Chuang, J.Y.; Khalil, B.; Wu, H.E.; Chang, W.C.; Maurice, T.; Su, T.P. Sigma-1 receptor chaperones rescue nucleocytoplasmic transport deficit seen in cellular and Drosophila ALS/FTD models. Nat Commun 2020, 11, 5580. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, G.A.; Clark, A.W.; Rooke, K.; Pramatarova, A.; Krizus, A.; Suchowersky, O.; Julien, J.P.; Figlewicz, D. SOD1 mutation is associated with accumulation of neurofilaments in amyotrophic lateral sclerosis. Ann Neurol 1996, 39, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, A.; Gradus, T.; Altman, T.; Maimon, R.; Saraf Avraham, N.; Geva, M.; Hayden, M.; Perlson, E. Targeting the Sigma-1 Receptor via Pridopidine Ameliorates Central Features of ALS Pathology in a SOD1(G93A) Model. Cell Death Dis 2019, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.B.; Gusella, J.F. Huntington's disease. Pathogenesis and management. N Engl J Med 1986, 315, 1267–1276. [Google Scholar] [CrossRef]
- Carmo, C.; Naia, L.; Lopes, C.; Rego, A.C. Mitochondrial Dysfunction in Huntington's Disease. Adv Exp Med Biol 2018, 1049, 59–83. [Google Scholar] [CrossRef]
- Eddings, C.R.; Arbez, N.; Akimov, S.; Geva, M.; Hayden, M.R.; Ross, C.A. Pridopidine protects neurons from mutant-huntingtin toxicity via the sigma-1 receptor. Neurobiol Dis 2019, 129, 118–129. [Google Scholar] [CrossRef]
- Shenkman, M.; Geva, M.; Gershoni-Emek, N.; Hayden, M.R.; Lederkremer, G.Z. Pridopidine reduces mutant huntingtin-induced endoplasmic reticulum stress by modulation of the Sigma-1 receptor. J Neurochem 2021, 158, 467–481. [Google Scholar] [CrossRef]
- Kusko, R.; Dreymann, J.; Ross, J.; Cha, Y.; Escalante-Chong, R.; Garcia-Miralles, M.; Tan, L.J.; Burczynski, M.E.; Zeskind, B.; Laifenfeld, D.; et al. Large-scale transcriptomic analysis reveals that pridopidine reverses aberrant gene expression and activates neuroprotective pathways in the YAC128 HD mouse. Mol Neurodegener 2018, 13, 25. [Google Scholar] [CrossRef]
- Bol'shakova, A.V.; Kraskovskaya, N.A.; Gainullina, A.N.; Kukanova, E.O.; Vlasova, O.L.; Bezprozvanny, I.B. Neuroprotective Effect of σ1-Receptors on the Cell Model of Huntington's Disease. Bull Exp Biol Med 2017, 164, 252–258. [Google Scholar] [CrossRef]
- Ryskamp, D.; Wu, J.; Geva, M.; Kusko, R.; Grossman, I.; Hayden, M.; Bezprozvanny, I. The sigma-1 receptor mediates the beneficial effects of pridopidine in a mouse model of Huntington disease. Neurobiol Dis 2017, 97, 46–59. [Google Scholar] [CrossRef]
- McGarry, A.; Auinger, P.; Kieburtz, K.; Geva, M.; Mehra, M.; Abler, V.; Grachev, I.D.; Gordon, M.F.; Savola, J.M.; Gandhi, S.; et al. Additional Safety and Exploratory Efficacy Data at 48 and 60 Months from Open-HART, an Open-Label Extension Study of Pridopidine in Huntington Disease. J Huntingtons Dis 2020, 9, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Alexoudi, A.; Alexoudi, I.; Gatzonis, S. Parkinson's disease pathogenesis, evolution and alternative pathways: A review. Rev Neurol (Paris) 2018, 174, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Calabresi, P.; Mechelli, A.; Natale, G.; Volpicelli-Daley, L.; Di Lazzaro, G.; Ghiglieri, V. Alpha-synuclein in Parkinson's disease and other synucleinopathies: from overt neurodegeneration back to early synaptic dysfunction. Cell Death Dis 2023, 14, 176. [Google Scholar] [CrossRef]
- Hong, J.; Wang, L.; Zhang, T.; Zhang, B.; Chen, L. Sigma-1 receptor knockout increases α-synuclein aggregation and phosphorylation with loss of dopaminergic neurons in substantia nigra. Neurobiol Aging 2017, 59, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Limegrover, C.S.; Yurko, R.; Izzo, N.J.; LaBarbera, K.M.; Rehak, C.; Look, G.; Rishton, G.; Safferstein, H.; Catalano, S.M. Sigma-2 receptor antagonists rescue neuronal dysfunction induced by Parkinson's patient brain-derived α-synuclein. J Neurosci Res 2021, 99, 1161–1176. [Google Scholar] [CrossRef]
- Vetel, S.; Foucault-Fruchard, L.; Tronel, C.; Buron, F.; Vergote, J.; Bodard, S.; Routier, S.; Sérrière, S.; Chalon, S. Neuroprotective and anti-inflammatory effects of a therapy combining agonists of nicotinic α7 and σ1 receptors in a rat model of Parkinson's disease. Neural Regen Res 2021, 16, 1099–1104. [Google Scholar] [CrossRef]
- Wang, M.; Wan, C.; He, T.; Han, C.; Zhu, K.; Waddington, J.L.; Zhen, X. Sigma-1 receptor regulates mitophagy in dopaminergic neurons and contributes to dopaminergic protection. Neuropharmacology 2021, 196, 108360. [Google Scholar] [CrossRef]
- Francardo, V.; Geva, M.; Bez, F.; Denis, Q.; Steiner, L.; Hayden, M.R.; Cenci, M.A. Pridopidine Induces Functional Neurorestoration Via the Sigma-1 Receptor in a Mouse Model of Parkinson's Disease. Neurotherapeutics 2019, 16, 465–479. [Google Scholar] [CrossRef]
- Pandey, S.; Srivanitchapoom, P. Levodopa-induced Dyskinesia: Clinical Features, Pathophysiology, and Medical Management. Ann Indian Acad Neurol 2017, 20, 190–198. [Google Scholar] [CrossRef]
- Johnston, T.H.; Geva, M.; Steiner, L.; Orbach, A.; Papapetropoulos, S.; Savola, J.M.; Reynolds, I.J.; Ravenscroft, P.; Hill, M.; Fox, S.H.; et al. Pridopidine, a clinic-ready compound, reduces 3,4-dihydroxyphenylalanine-induced dyskinesia in Parkinsonian macaques. Mov Disord 2019, 34, 708–716. [Google Scholar] [CrossRef]
- Fox, S.H.; Metman, L.V.; Nutt, J.G.; Brodsky, M.; Factor, S.A.; Lang, A.E.; Pope, L.E.; Knowles, N.; Siffert, J. Trial of dextromethorphan/quinidine to treat levodopa-induced dyskinesia in Parkinson's disease. Mov Disord 2017, 32, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sukumaran, P.; Singh, B.B. Sigma1 Receptor Inhibits TRPC1-Mediated Ca(2+) Entry That Promotes Dopaminergic Cell Death. Cell Mol Neurobiol 2021, 41, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Sha, S.; Zhou, L.; Wang, C.; Yin, J.; Chen, L. Sigma-1 receptor deficiency reduces MPTP-induced parkinsonism and death of dopaminergic neurons. Cell Death Dis 2015, 6, e1832. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of sigma-1 receptor agonism on neurodegenerative disorders. Agonism of sigma-1 receptor can dampen inflammation and reduce ER and oxidative stresses while promotes mitochondrial functions and calcium homeostasis. .
Figure 1.
Effects of sigma-1 receptor agonism on neurodegenerative disorders. Agonism of sigma-1 receptor can dampen inflammation and reduce ER and oxidative stresses while promotes mitochondrial functions and calcium homeostasis. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



