Submitted:
10 April 2025
Posted:
11 April 2025
You are already at the latest version
Abstract
Broomrapes (Orobanche & Phelipanche spp.) are parasitic weeds that significantly impact the productivity of major crops in Mediterranean region, like tomato (Solanum spp.) and faba bean (Vicia faba) species. This review article discusses extensively management strategies to control broomrapes, ranging from preventive measures to curative approaches. Additionally, it includes meaningful information to intricate molecular mechanisms underlying the broomrape-host interaction, focusing on host recognition of parasitic plant molecular patterns and the hormonal crosstalk that regulates the establishment of parasitism. Moreover, the article highlights the potential of breeding for resistance in cultivated crops such as tomato and faba bean as a sustainable, long-term solution to combat broomrape infestation. This review serves as a valuable resource for both researchers and farmers offering insights to develop, implement and adapt effective and environmentally sustainable management practices for broomrape in Mediterranean agricultural systems.
Keywords:
broomrapes
; broomrape management
; hormonal interactions
; breeding
; tomato
; faba bean
1. Introduction
Broomrapes are obligate root parasitic plants (Orobanche and Phelipanche species) that could cause severe damage to many field crops. In general, the crops affected are members of various families (i.e. Fabaceae, Apiaceae, Cucurbitaceae, Solanaceae, Lamiaceae, Ranunculaceae, and Asteraceae) [1]. In particular, from the long list of these parasitic species, only a limited number of species are considered as important parasitic threat for the Mediterranean cropping systems [1,2,3,4,5]. Those include: Crenate broomrape (O. crenata; hosts are species in Fabaceae, Apiaceae, Cucurbitaceae, Solanaceae, Lamiaceae, Ranunculaceae, and Asteraceae; major problem in faba beans); small broomrape (O. minor; hosts are species in Fabaceae, Solanaceae, Asteraceae, and Apiaceae; major problem in lentils, clover and lucerne); foetid broomrape (O. foetida; hosts are species in Fabaceae; major problem in faba beans, chickpeas, vetch); sunflower broomrape (O. cumana; host is specific and major problem in sunflower); nodding broomrape (O. ceruna; hosts are species in Solanaceae; major problem in tomato); branched broomrape (P. ramosa; hosts are species in Fabaceae, Apiaceae, Brassicaceae, Solanaceae, Cannabiaceae, and Asteraceae; major problem in tomato, tobacco, and oilseed rape); Egyptian broomrape (P. aegyptiaca; hosts are species in Brassicaceae, Cucurbitaceae; major problem in tomato, potato, lentil, and carrot). In this review, given the focus on major Mediterranean crops (i.e. cool-season legumes and industrial tomato), only four species are of great concern such as: O. crenata, O. foetida, P. ramosa, and P. aegyptiaca.
Broomrapes possess several distinctive biological and ecological characteristics that contribute to its persistence and difficulty in control. Over the last 40 years, there have been numerous publications and review papers about all aspects of broomrape life cycle and management methods. Some comprehensive and recent reviews are the followings: [2,6,7,8,9,10]. Based on the published information, the most important characteristics of the broomrapes are the followings: a) Achlorophyllous Nature: having a total lack of chlorophyll, they are entirely dependent on a host plant for its nutrients, water, and energy; b) specialized parasitic growth: being a root parasite, it attaches to the host roots by forming haustoria structures; c) Underground Development: the majority of broomrape's life cycle occurs underground, and manifests itself only at the very last stage of development making difficult to control with conventional methods; d) Prolonged dormancy: The seeds can remain dormant in the soil for extended periods, sometimes up to 20 years, waiting for a suitable host to sprout; e) Host Specificity: Broomrape species tend to have high specificity for certain host plants, often parasitizing only one or a limited number of crops; f) Flowering and Seed Production: Broomrapes can produce thousands of tiny seeds per plant, which are dispersed by wind, water, or human activity, helping it spread across large areas; g) Life Cycle: The seed germination is triggered by specific chemical cues from the host plant roots; h) Low success of control methods: Broomrape's underground parasitic nature makes it resilient to most traditional weed control methods, including herbicides, mechanical weeding, and crop rotation; i) Growth Synchronization with host: There is an impressive synchronization of the parasite to the host for weed growth and development.
In the Mediterranean region, the most important cool-season legumes are: faba beans (Vicia faba L.), field peas (Pisum sativum L.), lentils (Lens culinaris Medik), grass peas (Lathyrus sativus L.), and chickpeas (Cicer arietinum L.); the most important one being the faba beans. All the above legume species are host crops to broomrapes that cause significant yield losses and hence severe economic damage [11]. In addition, tomato (Solanum lycopersicum Mill.) is the vegetable crop with the highest demand and the greatest economic value in the world. In addition, industrial tomato is considered an important and high value crop in the Mediterranean region that suffers significant losses due to the broomrape parasitism [12].
Due to the above-mentioned characteristics, broomrapes are considered one of the most challenging agricultural problems to tackle that could threaten major Mediterranean crops. Despite the huge effort that has been made in the last 50 years, unsuccessful management has exacerbated the problem resulting in continuous spreading into new growing areas [10]. Novel approaches to understanding the mechanism of host-parasite interaction and concentrated efforts to develop resistant cultivars are highly needed more than ever before.
In this review, all the latest research knowledge will be presented in the following’s sections: preventive and curative methods for broomrape management; hormonal interactions related to host signal perception and haustorium development; molecular patterns of host plants to recognize the parasite invaders; and finally breeding efforts for developing resistance cultivars. The focus will be given to Orobanche and Phelipanche species that parasitize major Mediterranean crops (i.e. cool-season legumes with emphasis in faba beans and industrial tomato), although the information presented here could benefit other host crops since either similar control methods exist, or host-plant interactions may be similar.
2. Management Strategies
Significant research efforts to manage broomrape infestations in major Mediterranean crops have been undertaken, with approaches ranging from agronomic practices to biological control methods. Despite these efforts, the efficacy levels remain largely unsatisfactory, and the problem persists in agricultural practice (Table 1). Although significant scientific discoveries have been made in the process, these measures have often had limited success in terms of commercial application. The main reasons for the limited success of these approaches include a) partial protection: Most solutions have only provided partial protection against broomrape; and b) low economic viability: Many of these solutions are not economically feasible particularly for low-input legume cropping systems. Managing broomrape (Orobanche spp.) effectively requires a combination of preventive and curative measures that aim to reduce the risk of infestation and minimize its impact on crops.
2.1. Preventive Measures
Broomrapes are parasitic weeds that rely on host plants for their survival, and hence integrated management strategies focusing on preventing its establishment and spread are essential. Below are presented the key preventive measures for the effective management of broomrapes.
2.1.1. Crops’ Exploitation
Rotation decreases the frequency of host cultivation is one of the main ways that farmers deal with the broomrape-related problem. The key strategy is to rotate susceptible with non-susceptible crops aiming to reduce the weed's impact over time. This key strategy is served by the following three pillars: a) low presence of host crops that could limit broomrapes, reducing its opportunities to parasitize and spread; b) prolong soil seed dormancy since the broomrape cannot parasitize and germinate due to the absence of a suitable host; and c) limit soil seed bank replenishment since no seed rain could come from newly developed broomrape plants. Rotation, however, it is a long-term strategy due to the long viability of seed bank [13], which requires at least a five-course and ideally a nine-course rotation [14,15,16,17]. In such a rotation scheme, should be included non-host crops and fallows to prevent broomrape seed bank increases and manage broomrape outbreaks. In this context, cereals (corn, wheat, barley) or grass could be included to manifest direct and indirect impact on parasitism. Previous research has shown that good rotation crop examples were alfalfa (Medicago sativa), wheat (Triticum spp.), and cultivated oat (Avena sativa) [18,19]. In addition, where severe broomrape problems exist in cool-season legumes (i.e. in Ethiopia), other rotational crops such as common beans (Phaseolus vulgaris L.) and fenugreek (Trigonella foenum-graecum L.) crops are selected [9]. Rotation impact could be enhanced by incorporating trap, catch or allelopathic crops into the scheme, as suggested by studies like Habimana et al. in 2014[20].
Trap crops refer to the cultivation of crop species whose root exudates exhibit high germination inducing activity on broomrape seeds, but these species do not become infected because they are resistant to later stages of the parasitic process indirectly leading to the killing of the young broomrape seedlings due to the lack of proper host. Many other interesting examples of trap crops emerged from a root exudates screening of important crops [13]. Examples of trap crops for P. ramosa include flax, Phaseolus bean, sorghum, maize, sunflower and cucumber [21,22]. Additional trap crop examples include coriander (Coriandrum sativum L.), basil (Ocymum basilicum L.), fenugreek (T. foenum-graecum L.), and clover (Trifolium alexandrinum L.) [4,20,23]. Recent reports have shown that intercropping flax or fenugreek with faba bean cultivars, caused a significant reduction in the infestation levels of O. crenata and increased the faba bean yield compared with sole treatments; flax was more effective in the reducing infestation rate by O. crenata than fenugreek [24]. However, there are only very limited good case-studies of trap-crops and therefore this strategy should be only considered as a component of an IPM approach.
Catch crops could induce high broomrape germination but progression of the parasitism does not materialize due to mechanical, physiological, chemical factors. Catch crops are mostly short-term crops that are harvested or destroyed shortly after sowing (i.e.in 6-8 weeks), mostly at the vegetative stage and prior to parasite emergence or before parasite flowering [25]. In legumes, tori (Brassica campestris var. toria) when managed properly as a catch crop can result in up to a 30% reduction in the size of broomrape seed bank [26]. According to Nadal et al. (2005)[27], using faba beans as a trap crop (i.e. harvested before broomrape can flower and produce seeds) can significantly deplete the broomrape seed bank by more than 30% in a single season. In industrial tomato, flax could be an example of catch crop; the crop was shown 4-6 weeks prior to tomato transplantation and it significantly suppressed P. ramosa infestation [6]. Other good examples are Vigna unguiculata, Hibiscus sabdariffa, Hordeum vulgare and Sorghum bicolor [7], Lotus corniculatus [28]. A large screening survey of several plant species including summer and winter plants, medicinal herbs and important weed species for trapping and catching species for P. ramosa was done [29].
Allelopathic crops could play a role in managing broomrape populations by preventing seed germination, disrupting attachment, and inhibiting growth. These crops could produce allelochemicals that are secondary metabolites that would manifest a variety of effects, both on the parasitic plants themselves and on the surrounding ecosystem. Allelopathic crops have low efficacy on their own and ideally should be part of the rotational scheme or other IPM practices. A good example of such allelopathic crop in legumes is fenugreek (Trigonella foenum-graecum L.) [30]; trigoxazonane was identified from fenugreek root exudates, would be responsible for inhibition of O. crenata seed germination [31]. Another allelopathic crop could be Desmodium uncinatum that was shown to inhibit the development of Striga haustoria [32]. A number of compounds like vanillic, o-coumaric acids or scopoletin that exerted from cereal’s roots [33] are having allelopathic effects [34] that could serve as key allelopathic crops in managing broomrapes. The complexity of the direct and indirect impacts of allelochemicals on parasitic plants indeed makes their use in control strategies quite challenging.
Winter cover crops could play an important role in suppressing the parasitism outbreaks in summer crops such as industrial tomato; in cool-season legumes, they have no practical value. Little research has been done to document the effects of winter cover crops in managing broomrapes. These service crops provide several benefits, including weed control, improving soil health and reducing broomrape seed bank buildup. Winter cover crops are planted in the off-season, typically between harvest and the next planting season, and can suppress broomrape growth through several mechanisms as catch, trap or allelopathic crops, as previously discussed. In this context, Brassica species are releasing glucosinolates [35]; common cereals (i.e. rye) root exudates that could have an allelopathic effect on broomrapes [36]. Haidar et al. (1995a)[37], reported that surface-applied and pre-plant incorporated wheat and barley straw mulch residues had a significant impact on reducing the infestation and growth of P. ramosa in potatoes. Utilizing cover crops by adding dried powdered leaves of radish (Raphanus sativus) to the soil can be an effective method for reducing the incidence of P. ramosa (branched broomrape) and protecting tomato plants (S. lycopersicum) from parasitism [38]. Using cover crops to manage broomrapes could be part of the IPM strategy [39] but it requires a careful selection of cover crops and an understanding of the interactions between these plants and the parasitic broomrapes.
2.1.2. Modifications on Sowing and Transplanting Dates
Modification of sowing in legumes and transplanting in industrial tomato could help to manage broomrape outbreaks. It is known that key environmental factors (i.e. soil temperature and moisture) are critical for the onset of parasitism [40]. In this context, modification of dates could affect all three important stages such as preconditioning, germination and attachment/development of parasitism.
In faba beans, previous reports have shown that one-month shifting (from October to November and from December to January) reduced O. crenata and O. foetida parasitism [14], similar results were also reported by Perez-de-Luque et al., 2004[41]. Also in other field trials, in Ethiopia, delayed sown faba beans had lower levels of parasitism [42], similar results have been demonstrated in chickpeas [43]. Recent reports have shown that 3 weeks delay in faba bean sowing reduced significantly the number of emerged O. crenata shoots for both the resistant (Giza 843) and the susceptible (Nubaria 1) cultivars [24].
In industrial tomato, previous reports have shown that earlier transplanting periods manifested higher yields and lower concentration of broomrape in [44]. However, modification of dates could have a serious negative effect on crop development and final yields; delayed legume sowing might create higher abiotic stresses (i.e. heat stress in flowering, drought situations) whereas early transplanting, in industrial tomato, might subject the crop to suboptimal growing conditions. In addition, modification of dates could benefit scenarios with low or moderate levels of parasite infections [45].
2.1.3. Solarization
Soil solarization has been shown to be an effective non-chemical method in reducing broomrape seed viability in areas with sufficiently hot summer climatic conditions. As such, it is better suited in cool-season legumes but has no practical value in industrial tomato since it will require to take tomato fields out of production for several months during the summer growing season in the Mediterranean region.
In general, solarization can significantly increase top-soil temperatures up to 15 cm; soil should be moist soil and covered with transparent polyethylene sheets for at least 1-2 months. Older reports have shown that soil solarization, based on mulching moist soil with polyethylene sheets for several weeks under solar irradiation, can provide excellent levels of control of Orobanche seeds in the upper soil layers [46]. Several other reports confirmed the significant effects of soil solarization in killing P. ramosa seeds [21,47,48]. Additionally, the great benefits of soil solarization (99% mortality of broomrape viable seeds in the seedbank) were documented when applied for two consecutive summers without any negative impact on tomato yield [47]. The major drawbacks of soil solarization are the following: the high cost for implementing the method, the need to preserve soil moisture during the hot summer period, the lack of irrigation for cool-season legumes. All the above restrictive elements, undermine the feasibility of applying this method despite its high efficacy in controlling broomrape parasitism.
2.1.4. Tillage, Mechanical Cultivation
Tillage and mechanical cultivation could affect the weed soil seed bank, including broomrape seeds. Conventional tillage (i.e. moldboard plow), although may reduce germination stimulation, it ultimately results in seedbank enrichment, as broomrape seeds are viable for more than a decade [49]. In addition, they contribute to a higher seed persistence level by protecting seeds from aging, predation, and induced dormancy due to lower oxygen in deep soil layers [50].
Two distinct non-conventional soil management practices could have an impact on controlling broomrape infestation such as minimum tillage and deep ploughing. Previous reports showed that minimum tillage reduced the amount of viable broomrape seeds incorporated into the soil [51], and hence their potential germination. On the other hand, deep ploughing is a technique often recommended to control broomrape by burying seeds at a depth where they cannot germinate due to insufficient oxygen or unfavorable conditions [52,53,54]. Finaly, both minimum tillage and deep tillage, although could have a moderate efficacy, given their feasibility limitations, seems to be of lower agronomic value practices.
2.1.5. Fertilization
Proper nutrient management can help make the soil less conducive to the establishment of parasitism in crops. Fertilization can influence the soil and plant environment in ways that reduce the likelihood of broomrape seeds attaching to host plants and initiating parasitism in clover and tomato [55]. Nutrient management plays a significant role in promoting tolerance to broomrape parasitism at different stages of crop growth and pre or post attachment stage of the parasite [56].
Direct contact with fertilizer, such as urea and ammonium, may be toxic to broomrape, inhibiting seed germination and seedling growth [11]. Also, the negative effect of ammonium on broomrapes is linked to the plant's limited ability to detoxify ammonium using the enzyme glutamine synthetase [2]. However, other forms (i.e. nitrates) did not exhibit the same inhibitory effects [55,57]. In addition, fertilization could modify the synthesis and exudation of root stimulants (i.e. strigolactones) of the host crop. Phosphorous and nitrogen have been described to down regulate strigolactones exudation in some several crop host species [58]. In industrial tomato, given the fertigation system, it could be important to utilize the fertilization schedule in managing the broomrape invasions whereas in legumes it would be considered of a lesser importance.
2.1.6. Use of Synthetic Stimulants, Suicidal Germination
This method involves the promotion of suicidal germination in broomrape (Orobanche spp.) through the use of synthetic molecules that mimic the natural germination-inducing factors released by host plants, such as strigolactones [59]. This technique is based on the idea of tricking the parasite into germinating in the absence of a suitable host, leading to the death of the parasite seedling since it is unable to acquire the necessary nutrients through parasitism. Previous research on direct application of synthetic strigolactone analogs (i.e. GR7) to deplete soil seed bank [59], proved to be very challenging due to instability issues and the difficulty in achieving consistent and effective application in the field [60]. Newer types of synthetic strigolactone analogs (i.e. Nijmegen-1) that were more stable and better suited for field applications resulted in high suicidal potential [61]. Research has shown the complex interactions between strigolactones, DNA methylation, and the potential for using synthetic compounds to control P. ramosa [62]. In addition, recent studies have demonstrated potential of auxin-derived strigolactone mimics to induce suicidal germination in broomrapes [63]. Other type of stimulants include triazolide strigolactone mimics [64] and Phthalimide-derived strigolactone mimics [65] they have been synthesized as potent selective germinators of broomrape seeds. However, for this method, significant challenges exist such as variable efficacy levels, low stability, high production cost and potential off-target effects in the soil microorganisms.
2.1.7. Polyethylene Plastic Mulching
The use of polyethylene sheet mulching is a cropping system that is used in industrial tomato since it offers significant benefits such as early planting, yield increases, better agrochemical utilization and enhanced weed control. Regarding the broomrape control, the use of polyethylene sheets acts by modifying the temperature and moisture in soil. The plastic sheet can significantly increase soil temperature underneath it, creating an unfavorable environment for broomrape seed germination and root attachment. By enhancing and preserving high soil moisture, it could be more challenging for broomrape seeds to establish a successful parasitic relationship with the tomato plants. Research demonstrated that black polyethylene sheeting applied on the day of transplanting tomato completely controlled broomrape in the field by preventing seeds from germinating and attaching to the plant roots [66]. Similarly, polyethylene mulch application had the highest efficacy in controlling broomrape in aubergines, with greater than 95% control. The polyethylene mulch likely prevented broomrape seed germination and created an unfavorable environment for seed attachment and development [67]. This method has got significant constraints and limitations such as: a smaller scale of potential growing areas, high cost of plastic sheet, equipment and labour, and environmental impact for plastic disposal or recycling.
2.1.8. Use of Crop Tolerance Inducers
Host plants could employ Systemic Acquired Resistance (SAR) that is a plant's defense mechanism that is triggered by an initial localized infection or injury, leading to the activation of defense responses throughout the entire plant to defend on parasitism. As such, salicylic acid and benzothiadiazole application activated broomrape resistance in various legumes such as faba bean [68], pea [41] and red clover [69].
In tomato, both salicylic acid and indole acetic acid can activate the defense mechanisms against P. ramosa parasitism [70]. Priming tomato seeds with different concentrations of the above hormonal inducers significantly reduces the number and biomass of Orobanche spp. tubercles and improves tomato root growth [71]. Based on this principle, experiments have shown that a commercial product (i.e. Bion®), under experimental conditions, could decrease broomrape infection to 80% in hemp and tobacco [72]. Other inducers were also investigated such as L-methionine that have the potential to reduce broomrape invasions on tomatoes [73]. Research has shown the role of biotic inducers of systemic resistance (ISR) in enhancing a plant’s defense against broomrape parasitism. Specifically, Rhizobium leguminosarum, a type of beneficial bacterium, has been shown to induce defense mechanisms that help protect plants from broomrape infection [74]. Additionally, strains of Pseudomonas sp. were documented to trigger induced systemic resistance [75] a commercial product (i.e. Proradix®), in experimental conditions, was shown to have high (80%) control of broomrape parasitism [72]. However, no field applications have been tested the above method under real field conditions.
2.1.9. Use of Resistant/Tolerant cvs
While significant research efforts have been made to develop resistant or tolerant cultivars to broomrape in both legumes and industrial tomatoes, success has been more pronounced in legumes, particularly in faba beans, where a few moderately successful examples of broomrape resistance/tolerance have been achieved. In contrast, there has been no commercially successful broomrape-resistant cultivar developed for industrial tomatoes. ICARDA has developed a multi-site screening program aiming to develop cultivars with resistance to broomrape [76] using as the main source of Orobanche resistance the line F402 identified by Egyptian researchers in early 1970s [77] and some minor sources available in different Mediterranean countries. In this context, over the last 40 years, a number of faba bean cultivars with resistance/tolerance to broomrapes have been developed such as: Baraca, Quijote, Faraon, Navio, Quijote, Najeh, Chourouk, Misr1, Giza843 [78,79,80,81]. Some of these cultivars (i.e. Quijote, Navio, and Najeh) are notable for their unique and effective resistance mechanism against broomrape through a mechanism that involves reduced strigolactone exudation from their roots [82].
Finally, substantial research is needed in this direction to identify a range of broomrape-resistant cultivars with different precocities that gives farmers the flexibility to select the best-performing varieties for their specific environmental and farming conditions. This approach maximizes the potential for controlling broomrape and enhances the sustainability and productivity of agricultural systems by allowing for adaptation to diverse growing conditions and crop management strategies. A full account of the latest developments in breeding for resistance in both faba beans and industrial tomatoes is presented in the following sections of this review.
2.2. Curative Measures
Curative measures to manage broomrapes are generally applied after the parasite has already established itself in the field. Unlike preventive measures, which aim to avoid infestation, curative measures focus on reducing the impact of existing broomrape infestations or attempting to eliminate or control the parasite after it has been established in crops.
2.2.1. Seed Dressing Treatments
Previous research, in faba beans, has shown that seed dressing treatments with imidazolinones proved effective for controlling O. crenata; additionally moderate control using imazethapyr was measured through faba bean and pea seed treatments [83]. In tomato, seed dressing using sulfonylureas (i.e. chlorsulfuron, triasulfuron) in low doses showed some effect on broomrapes with good safety in the crop [84]. However, the seed dressing treatments have low residual effect and need to be supported by other herbicide applications to show their full effectiveness [4].
2.2.2. Foliar Herbicides
In cool-season legumes, being a low-input rain-fed cropping system, only systemic herbicides could be used by foliage spraying, absorption and translocation of herbicide to the roots where broomrape germination and establishment is occurring. ones, incorporated on the leaves and translocated to the roots. In this context, the use of glyphosate with lower doses (at 60-120 g a.i./ha) have shown to effectively control broomrapes in faba beans, lentils , and peas [9,23,85]. In addition, imazethapyr, imazapyr, and chlorsulfuron were effective in controlling broomrapes (i.e. O. crenata, P. aegyptiaca and O. foetida) in legumes [86,87].
In industrial tomato, although lower doses of glyphosate (i.e. 30 to 50 g a.i./ha) were effective against P. ramosa and P. aegyptiaca, they significantly affect crop development and reduced tomato yields [7,88]. In addition, several sulfonylureas (i.e. bensulfuron, chlorsulfuron, nicosulfuron, primsulfuron, trimsulfuron, thifensulfuron, and triasulfuron) have shown to have variable level of effectiveness in broomrapes (i.e. O. aegyptiaca) under real field conditions [89]. Also, imazethapyr applications, although it was very effective in broomrape control, there were severe phytotoxicity problems in tomatoes [70].
2.2.3. Herbigation
Herbigation (i.e. delivery of herbicides through irrigation water) is a method that could be employed in industrial tomato targeting broomrape seeds and young attachments. Previous research has shown that herbigation with some sulfonylurea herbicides (i.e. sulfosulfuron, chlorsulfuron, triasulfuron, rimsulfuron, imazapic, imazamox) could be used to control broomrapes (i.e. P. aegyptiaca and P. ramosa) in tomato through the soil [7,90]. In this context, the most effective herbicide showed to be sulfosulfuron applied at rates of 37.5 – 75g a.i./ha, to control broomrapes (P. aegyptiaca) at a maximum soil depth of 18 cm [50]. However, herbigation effects showed a range of efficacy level (from low to high) due to several complex factors such as the herbicide residual effect, precise identification of the right developmental stage of the parasite, type of irrigation water application, repeated applications, accurate dose application with irrigation [89].
2.2.4. Biological Control
Biological control presents a promising and environmentally sustainable strategy for managing broomrapes in field crops; the high specificity of such agents is one of the key advantages, as this selectivity allows the targeted parasite suppression without harming the crop plants [91]. There has been a plethora of published information showing that several Fusarium spp. and other plant pathogens have the potential to attack various species of broomrapes in field crops [7].
Fusarium oxysporum and Neocosmospora solani (synonyms Nectria haematococca, Haematonectria haematococca) are among the Fusarium species that are most promising because of their rapid growth, intensive sporulation and chlamydospore formation [92]. Other Fusarium species included F. venenatum isolates for tobacco [93]; F. acuminatum, F. fujikuroi, F. proliferatum and F. solani for various crops [94]. Other reports have indicated that Alternaria alternata, Dendrophora sp., Chaetomium sp. and Talaromyces trachyspermus could be potential good agents to control broomrapes [7]. Despite of the huge research effort that has been done in the last 40 years, the development of effective biological control for broomrape remains an ongoing challenge, with the poor field efficacy of known fungal pathogens being the primary barrier to commercialization. However, through approaches such as improving pathogen virulence, optimizing inoculation techniques, and incorporating biocontrol into IPM systems, it would be possible to overcome these challenges [95].
2.2.5. Beneficial Insects
From the long list of 40 species from 22 families of beneficial insects that could be used as biocontrol agents [96], only a limited number showed to be the good case-stidies such as: Phytomyza orobanchia Kalt., Chyliza extenuata, Diaphora mendica and Celypha spp. [97]. The most promising beneficial insect is Phytomyza orobanchia for controlling broomrapes; old reports have shown its ability to control O. cumana (sunflower broomrape) and O. cernua [98]. In the last decades, significant research has shown its potential to control broomrapes in cool-season legumes, mostly in faba beans, by parasitizing seed capsules that causes a great reduction in broomrape seed production ranging between 29-94% depending on Orobanche species [70]. While Phytomyza orobanchia holds promise as a biocontrol agent for Orobanche species, the reality is that its widespread natural distribution and its limited impact on seed production in high-infestation areas make it an imperfect solution for broomrape control. Despite extensive study and attempts at breeding and release, the insect has not demonstrated sufficient efficacy to warrant a commercially viable product.
2.2.6. Hand Weeding
Hand weeding is often the first and most straightforward measure mentioned for controlling broomrape species in field crops, especially when infestations are still light. This manual method involves the physical removal of emerged broomrape plants from the field, followed by their destruction. The impact of hand weeding is minimal on broomrape competition to the host crop but targets to reduce the soil seed bank. In industrial tomato, its feasibility is low given the dense canopy that cover the soil and makes surveillance and removal very laborious. As such, the hand weeding can be effective in small-scale or early-stage infestations, it comes with several limitations and challenges that make it not feasible for large-scale farming.
3. Recognition of Parasitic Plant Molecular Patterns by Hosts
To protect themselves from pathogenic invaders, plants possess passive prerequisites, such as the formation of physical barriers (cuticles, lignified cell walls, etc.) or storage and release of secondary compounds. In addition, plants have evolved an innate immune system to detect pathogens and actively fend them off. The major prerequisite of plant immunity are pattern recognition receptors (PRRs) [99,100,101,102] that recognize herbivore-, microbe- or generally pathogen-associated molecular patterns (HAMPs, MAMPs, or PAMPs) [103,104]. Well-known PAMPs include molecules as chitin of fungal and arthropod origins, flagellin and other proteins of microbial origin, or other compounds like lipopolysaccharides and cell-wall fragments. PAMPs are recognized by plant PRRs in the nano- to picomolar ranges and trigger various plant defense responses measurable as cellular calcium spikes, the emission of the stress-related phytohormone ethylene, the biosynthesis of secondary metabolites (e.g., callose, phytoalexins, lignins, etc.), the assembly of reactive oxygen species (ROS) or the expression of defense-related genes [104]. Taken together, these cellular defense responses restrict the pathogen growth and lead to partial or even full resistance of the host against the pathogen.
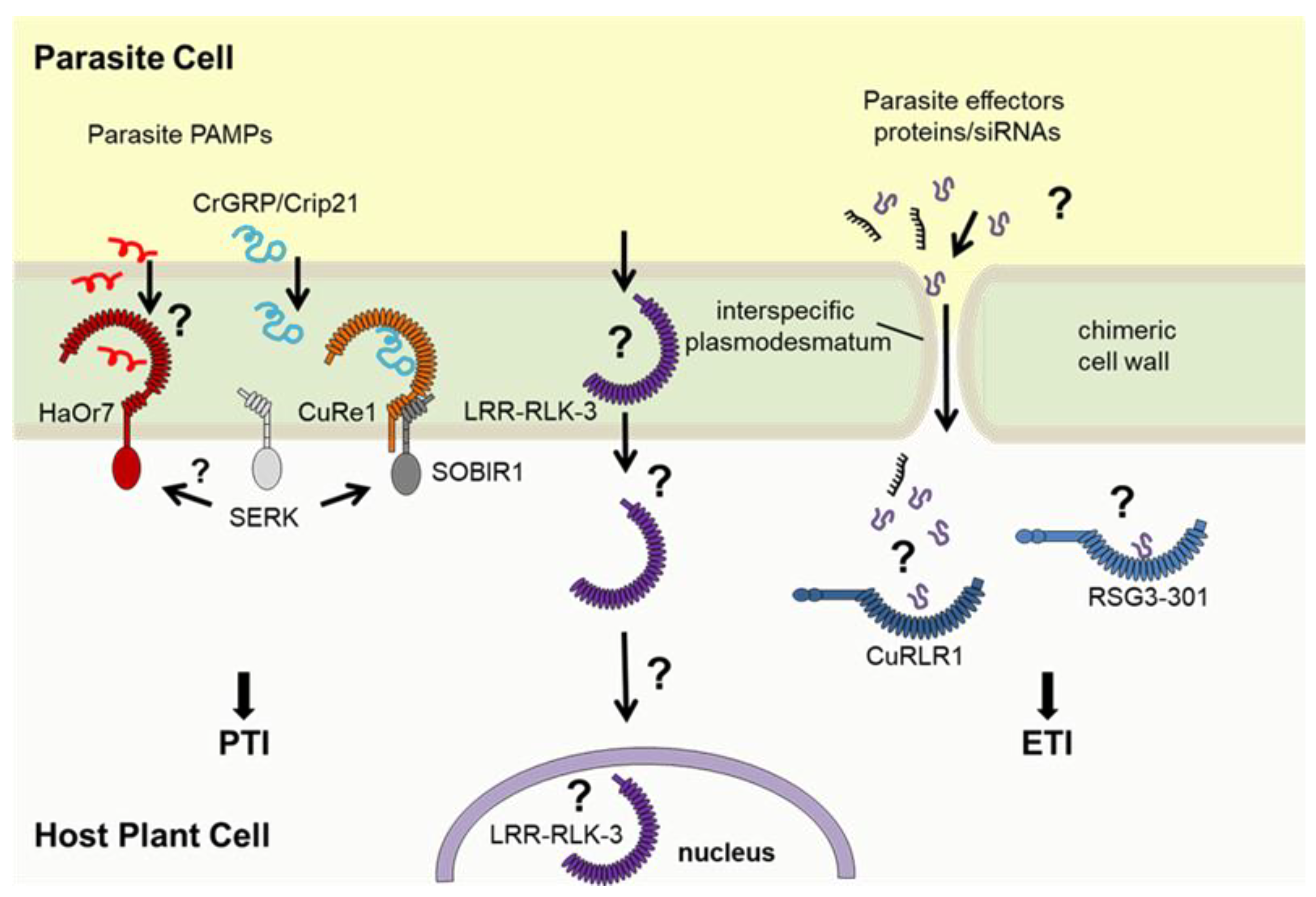
Unlike microbial plant pathogens, parasitic plants are angiosperms and even closely related to their hosts, which makes it more challenging for host plants to recognize them as alien invaders. Molecular patterns that would make parasitic plants distinguishable from "self" may be rare and must be highly specific. To date, only a few plasma membrane-bound PRRs have been identified in host plants that recognize parasitic plants and induce defense mechanisms helping to restrict parasitic plant growth. For example, one gene in sunflower which confers resistance to O. cumana races, was identified through a map-based cloning strategy. This gene, HaOr7 (Helianthus annuus Orobanche resistance 7), encodes a leucine-rich repeat receptor kinase (LRR-RK) that locates to the plasma membrane and the complete HaOr7 protein appears to be present exclusively in O. cumana resistant cultivars of sunflower [105]. Susceptible cultivars contain a variant of the gene that encodes a truncated HaOr7 protein lacking the kinase domain of the LRR-RK HaOR7 and hence seems to be not functional. In the case of successful resistance, an initial penetration of sunflower root tissue by O. cumana is possible, while a connection to the vascular system in the central cylinder of the sunflower roots is blocked due to the HaOr7-triggered resistance reactions [105].
Recently, a leucine-rich repeat receptor-like protein kinase has been identified that enhances tomato resistance to P. aegyptiaca [106]. In this study, field, pot, and rhizotron experiments were conducted to assess 118 tomato varieties against P. aegyptiaca. Further proteomic research, genome-wide analysis, and genetic engineering were performed to determine the LRR-RLK genes that control tomato resistance to P. aegyptiaca. One resistant variety (‘H1015’) was identified which manifested slower growth, partial necrosis, and a lower number of successfully infecting parasites compared to the susceptible variety (‘H2401’). The authors finally identified the LRR-RLK-3 as the essential key-player involved in resistance mechanisms. However, the protein LRR-RLK-3 is not a typical LRR-RLK since it lacks the kinase domain as well as a transmembrane helix domain. Subcellular localization of this truncated protein which appears to be a soluble LRR-domain seems also not to be in the tomato plasma membrane but rather in the ER/nucleus [106]. A binding partner or PAMP of parasitic P. aegyptiaca as well as a mechanistic mode of action for SlLRR-RLK-3 still must be elucidated.
Parasites of the genera Cuscuta comprises about 200 species that all infect the above-ground parts of hosts, preferentially the stems. However, the cellular resistance mechanisms in the host plants show similarities in mechanism and signaling pathways to those triggered by the root-parasitic Orobanchaceae. Interestingly, some species of the Solanaceae, especially of the genus Solanum, show an active defense reaction against selected Cuscuta species [107]. Indeed, the observation made for resistance of different tomato species against diverse Cuscuta species hint to a phenomenon comparable to “race-specific resistance” following a concept described for gene-for-gene interactions. For example, the strong resistance reaction of cultivated tomato (S. lycopersicum) seems very specific to Cuscuta reflexa [108], several other Cuscuta species can successfully infect tomato, including C. pentagona, C. campestris, C. suaveolens and C. europaea [109,110].
The resistance reaction of cultivated tomato against C. reflexa is visible right at the haustorium penetration sites [108,111]. About three to five days after the prehaustorium development, in the late attachment phase, epidermal host cells at the contact sites elongate and burst [112,113]. Sub-epidermal cell layers secrete soluble phenylpropanoids and show an increased activity of peroxidases. Together with synthesized di-fatty acids, omega-di-OH-fatty acids and lignin, these enzyme activities support the crosslinking of cell wall compounds and contribute to form a suberin-like barrier that prevents tomato from C. reflexa haustoria penetration [107].
How these active tomato resistance responses are switched on is only partially understood. One resistance-related gene was mapped on tomato chromosome 8 in an introgression line (IL)-based mapping approach [114]. Since S. lycopersicum shows active defense response against C. reflexa whereas the tomato relative S. pennellii is fully susceptible, the genetic diversity has been explored and a collection of S. lycopersicum x S. pennellii ILs [115,116] has been used for mapping the tomato defense against C. reflexa. The identified tomato gene encodes an LRR receptor protein (LRR-RP) – Cuscuta receptor 1 (CuRe1) – which plays an important role in the recognition of C. reflexa and in the induction of defense-related responses [114]. CuRe1 is a plasma membrane bound receptor that lacks an intracellular kinase domain. To switch-on cellular signaling, it constitutively exists in a heteromeric complex with the adaptor kinase SOBIR1, a requirement that has been described for LRR-RLPs in general [117,118]. CuRe1 recognizes a 116 aa long, glycine-rich protein of the C. reflexa cell wall (CrGRP) as a PAMP. As a minimal epitope, the 21 aa long peptide Crip21 specifically binds to CuRe1 and activates defense responses [119]. CrGRP, or the peptide Crip21, respectively, induce classical PAMP-triggered immunity responses [114,119,120], measurable as an induction of a ROS-burst or the production of the stress-related phytohormone ethylene. Moreover, CuRe1 contributes to host-plant resistance and reduces C. reflexa growth when transformed into otherwise susceptible hosts. However, CuRe1 is not the only relevant resistance gene and tomato harbors additional molecular factors, probably additional receptors, that may act synergistically for a full resistance. For example, loci mapped to chromosomes 1, 2 and 6 in an IL-based screen also appear to be critical for tomato resistance against C. reflexa [121]. Although CuRe1 is a plasma membrane-bound receptor and belongs to the classical set of PTI-related components, the involvement of mechanisms such as effector-triggered immunity (ETI), are probable. Indeed, a CC-NBS-LRR (Coiled-coil-Nucleotide-binding site-LRR) protein has been identified that seems to play an essential role in lignin-based resistance of Solanum habrochaites against C. campestris [122]. This cytosolic receptor, termed CuRLR1 (Cuscuta Receptor for Lignin-based Resistance 1), seems to recognize yet unknown parasitic factors, and acts as a key component in conferring the host resistance against C. campestris together with at least two transcription factors. Phenotypically, the stem cortex in these lines responds with local lignification upon C. campestris attachment, preventing parasite penetration into host.
For Striga–host interactions a race-specific resistance also has been described, following in principle mechanism observed for ETI [123]. S. gesnerioides races have been classified based upon their genetic relatedness and capacity to differentially parasitize cowpea varieties and landraces [124] and several race-specific resistance genes have been mapped in the cowpea genome [125]. Using a molecular marker-assisted positional cloning strategy, [126] subsequently isolated the RSG3-301 gene from cowpea that confers resistance to S. gesnerioides race SG3 and showed that it encodes a typical CC-NBS-LRR protein. The characterization of RSG3-301 led to the suggestion that race-specific Striga resistance in cowpea is an example of effector-triggered immunity (ETI) in which intracellular NLR proteins (such as RSG3-301) are activated either directly or indirectly upon recognition of pathogen/parasite effectors [127].
Figure 1.
PAMPs and recognition by PRRs in plant – parasitic plant interaction. Pathogen-associated molecular patterns (PAMPs) of parasitic plants, like C. reflexa GRP or its peptide epitope Crip21 [119] or other yet unknown patterns are recognized at the host cell surface by membrane-bound pattern-recognition receptors (PRRs), such as HaOr7 [105], CuRe1 [114] or tomato LRR-RLK-3 [106], and induce PAMP-triggered immunity (PTI) in host plant cells. Some host plants possess resistance proteins such as CC-NBS-LRR proteins that locate to the cytosol and may recognize effectors intracellularly. Consequently, those types of receptors induce effector-triggered immunity (ETI), often accompanied by cell wall lignification or a hypersensitive response – leading to resistance against parasitic plants; modified from Albert et al. 2021 [123].
Figure 1.
PAMPs and recognition by PRRs in plant – parasitic plant interaction. Pathogen-associated molecular patterns (PAMPs) of parasitic plants, like C. reflexa GRP or its peptide epitope Crip21 [119] or other yet unknown patterns are recognized at the host cell surface by membrane-bound pattern-recognition receptors (PRRs), such as HaOr7 [105], CuRe1 [114] or tomato LRR-RLK-3 [106], and induce PAMP-triggered immunity (PTI) in host plant cells. Some host plants possess resistance proteins such as CC-NBS-LRR proteins that locate to the cytosol and may recognize effectors intracellularly. Consequently, those types of receptors induce effector-triggered immunity (ETI), often accompanied by cell wall lignification or a hypersensitive response – leading to resistance against parasitic plants; modified from Albert et al. 2021 [123].
4. Hormonal Crosstalk Between Crop-Parasite
Phelipanche and Orobanche spp. are obligate root holoparasitic (devoid of chlorophyll) known as broomrapes, which develop a multicellular infectious organ called haustorium after recognizing hormonal signals released by the host [128]. Various studies demonstrate that holoparasites plants have adopted the ability to detect the same signaling molecules as other beneficial organisms, such as arbuscular mycorrhizal fungi (AM), and trigger the germination of their own seeds in the vicinity of host roots. Direct physical connections between the two ensure the unidirectional transfer of resources (water and nutrients) from host to parasite, but also the exchange of small signaling molecules, such as plant hormones, that concomitantly shape the metabolic activity of both partners [129,130]. Advances in understanding this signaling relationship between host and parasitic plants have greatly improved our comprehension of the evolution of plant parasitism and are facilitating the development of more effective control measures in cases where these parasitic plants have become a plague for species of high agronomic interest (including sunflower, tomato, tobacco, potato, and rapeseed) [131]. However, despite advances in understanding these interactions, a significant challenge remains in determining whether parasitic plants exude hormonally active metabolites in the rhizosphere, although there are indications that this may be the case [129,132,133]. In this review, we provide a bibliographic overview, including the latest findings on hormonal interactions related to host signal perception and haustorium development, where many remains to be deciphered.
Strigolactones (SLs) are the most well-studied class of germination stimulants for holoparasitic seeds. While angiosperms require optimal environmental conditions of light, temperature, and moisture for germination after dormancy release, parasitic plant seeds require the presence of SLs as a chemical signal indicative of host proximity. Additionally, the concentrations of SLs required to induce germination in parasitic plants vary from pM to μM, depending on the strigolactone, likely due to differences in the germination-stimulating receptor or subsequent signaling, possibly reflecting the coevolution of the parasitic plant species with their hosts [134]. The synthesis and exudation of these key chemical signals in parasitism are finely modulated by other biotic factors and changing environmental conditions [129]. The reduction in SL release by infested host roots could be an adaptive strategy to suppress parasitic seed germination [135,136]. However, since SLs are also a plant hormone that controls many aspects of plant development, SL deficiency may also affect post-germination stages of the infection process during the parasite-host interaction. In this regard, SL-deficient tomato mutant plants (SlCCD8 RNAi) infected with pre-germinated seeds of Phelipanche et al., showed a higher level of infection and faster parasite development, suggesting a positive role of SLs in host defense against parasitic plant invasion [137]. To date, the field application of SL analogs such as GR24, methyl phenlactonoate 3 (MP3), and Nijmegen-1 has shown promising results in combating holoparasitism through a suicidal germination approach [61,131]. Another solution lies in the use of monocot trap crops, as they induce the germination of broomrapes through the release of SLs but, unlike dicotyledons, are not parasitized [138].
Other scientific research proposes the use of gibberellin (GAs) agonists as a cheap alternative to natural bioregulators for the control of broomrapes seed banks. Thus, GAs have been shown to elicit in broomrapes seeds a germination activity similar to that of dormant seeds of several autotrophic plant species. The study carried out by Bao et al., (2017)[139] with P. aegyptiaca, reported that as in non-parasitic angiosperms, seed dormancy was mediated simultaneously by an increase in gibberellic acid and a reduction in abscisic acid (ABA). Finally, GAs applied in vitro to the seeds of P. ramosa and Orobanche minor promoted germination in the absence of a host or any other germination stimulator [65].
Genetic approaches have demonstrated the importance of ethylene and auxin signaling pathways for proper haustorium formation during sapling invasion. Cui et al., (2020)[140] proposed that parasitic plants used ethylene as a signal to invade host roots. In their research, mutants exhibiting severe haustorial defects were associated with point mutations in homologs of ETHYLENE RESPONSE 1 (ETR1) and ETHYLENE INSENSITIVE 2 (EIN2), signaling components in response to the gaseous phytohormone ethylene. Application of ethylene signaling inhibitors also caused similar haustorial defects, indicating that ethylene signaling regulated cell proliferation and differentiation in the parasite. Furthermore, genetic disruption of ethylene production in the host also disrupted parasite invasion. Numerous studies indicate that auxins play a crucial role in plant-parasite interactions; however, the responses vary among different species and appear to depend on the type of parasitism involved. Ishida et al., (2016)[133] reported that the localized expression of the auxin biosynthesis gene YUC3 in epidermal cells near the contact site resulted in the accumulation of newly synthesized auxins at the haustorial apex close to the host. This accumulation was necessary and sufficient to prime haustorium formation in the facultative hemiparasitic species Phtheirospermum japonicum. In contrast, auxin showed no haustorium induction in germinated seeds of the obligate holoparasite P. ramosa and had an inhibitory effect when applied together with root exudates [141]. The translocation of nutrients from the host to the broomrape is facilitated by a continuous vascular system at the host-parasite interface [11]. High levels of auxin are deemed essential for establishing the parasite-host xylem connection, as they interfere with the normal development and differentiation of xylem vessels in P. ramosa tubers in infected tomato plants [142]. On the other hand, disruption of normal auxin transport in strigolactone-deficient mutants resulted in increased susceptibility to infection by parasitic plants [137]. In the terminal haustoria of Striga asiatica, the orthologs SOLITARY ROOT (SLR) (INDOL-3-ACETIC ACID INDUCIBLE 14/IAA14) and AUXIN RESPONSE FACTOR 19 (ARF19) were specifically expressed at the early stage of haustorium development [130]. Xiao et al., 2022 reported that the application of an auxin biosynthesis inhibitor increased the rate of haustorium formation and the cessation of cell division in obligate hemiparasite Striga hermonthica[143]. Furthermore, the authors of this study proposed that preventing meristem differentiation by applying auxin or a metabolite with a similar effect could be an alternative approach to suicidal germination by preventing prehaustorium formation and, therefore, infestation by the parasite. Finally, Wakatake et al., (2020) demonstrated that the cooperative action of auxin transporters is responsible for controlling xylem vessel connections between the parasite and the host [144]. In their work, they suggest that these connections could be based on the Sachs model, where auxin “sources” (regions of high concentration) connect with “sinks” (regions of low concentration) in a self-organized manner. In parasitic plants, the apex of the haustorium acts as a source, and the site of xylem plate formation near the root vasculature acts as a sink.
The mechanisms leading to haustorium formation remain largely unknown. However, there is evidence that this event is triggered by a cytokinin (CK) signal, which, unlike auxins, is effective in both obligate and facultative parasitic plants. Bioassays by Goyet et al., (2017) demonstrated through the application of exogenous CKs and the specific CK receptor inhibitor PI-55, that CKs exuded by the roots of the host (Brassica napus) are crucial for haustorium formation and increase the aggressiveness of P. ramosa infection [141]. Roots of broomrape-infested plants were characterized by an enrichment of trans- and cis-Zeatin, CKs with high biological activity; and CK nucleosides, the predominant transport form in plant vascular tissues [129]. These results highlight that CKs are authentic rhizospheric signals for holoparasites. Furthermore, other studies reported that kinetin, a synthetic CK, and 6-benzylaminopurine (BAP) induce prehaustorium-like structures in Striga asiatica and Triphysaria versicolor, obligate and facultative parasites respectively (reviewed in Goyet et al., 2019[145]).
Similar to infections by pathogens and herbivores, parasitic plants also induce defense mechanisms in the parasitized host. The most well-studied responses involve systemic acquired resistance (SAR) induced by jasmonic acid (JA) and salicylic acid (SA), which also involve the expression of pathogenesis-related (PR) genes [146]. Abscisic acid (ABA)-mediated responses have also been established [147]. In addition to its prominent role in plant adaptation to abiotic stress conditions, ABA is intrinsically involved in host-holoparasite interactions. For instance, infection with P. ramosa resulted in increased levels of ABA and the ABA-glucose ester (ABA-GE) in the leaves and roots of wild-type and SL-deficient tomato lines [137]. Another study found that infection of tobacco plants with a mix of Phelipanche spp. led to a significant increase in ABA and phaseic acid (PA), a degradation product with biological activity similar to ABA, in root tissues [129]. As part of a common defense response module, it has been demonstrated that tomato genes related to JA, SA, and ABA are upregulated in the early stages of tomato plant parasitization by P. ramosa and interestingly, this upregulation is accompanied by an increase in the expression of SL biosynthetic genes. Additionally, SL-deficient tomato mutants are characterized by reduced levels of JA, SA, and ABA, making them more susceptible to fungal pathogens [148]. So far, research carried out indicate that plants weakened by abiotic factors become more vulnerable to biotic stress factors such as broomrapes; conversely, under abiotic stress conditions, parasitic plants might encounter potential hosts with already activated SAR. Several reports support the view that abiotic stress-induced hormonal signals could make potential host plants more insensitive or even resistant to parasitization [149,150]. As reported by Foyer et al., (2016) in their research on cross-tolerance, a single stress factor was able to induce different hormonal signaling pathways (SA, JA, ethylene, ABA, and auxins) that led to increased resistance to multiple biotic and abiotic factors [151].
Recently, the results published by Martínez-Melgarejo et al., (2024) align with the information reviewed in the present work. This research analyzed the comprehensive metabolomic profile of rootstocks sensitive and resistant to broomrapes to elucidate the resistance mechanisms conferred by the rootstock and the metabolomic alterations caused by the parasite in the host and vice versa. The results showed that in samples of sensitive rootstocks, SLs, GAs, and CKs were clustered with the photoassimilates of the plant (sugars, amino acids, organic acids), suggesting their role in the induction of sink activity in the host root for the development of broomrapes. Conversely, the ethylene precursors ACC, SA, and JA were negatively correlated with factors that favour parasitism, indicating their participation in the defense against infection and the development of broomrapes [152].
Ultimately, hormonal signals that enhance tolerance to abiotic stress may also bolster resistance to biotic stress through the upregulation of shared defense mechanisms. Therefore, to accurately assess the effects of host defense mechanisms, it is essential to carefully consider the specific species and environmental conditions of both the host and the parasite.
5. Breeding for Resistance
5.1. Industrial Tomato
Cultural practices and herbicide application have been commonly used to control Orobanche and Phelipanche species, though these methods often come with high costs. Breeding for hosts’ resistance offers a more sustainable and cost-effective solution for eliminating this parasitic weed [153]. Cultivated tomatoes are a vital crop and a major nutritious food source for humans, yet many commercial varieties are susceptible to infection by broomrape species. However, knowledge about tomato defense mechanisms against broomrape parasitism remains limited. Despite extensive screening efforts, no fully immune strategy or robust resistance to broomrape has been identified [154]. Breeding for broomrape resistance faces challenges as there is a complex interaction between the host plant and the broomrape species and there is a need to balance resistance with other favorable agronomic traits in tomatoes [90].
However, several research studies discuss various breeding strategies to manage broomrape resistance in tomato. Most of these breeding strategies follow specific steps: The first step is referred to as a pre-breeding method though the selection and exploitation of genetic material tolerant to Orobanche. According to this approach, it is recommended to explore and exploit sources of natural genetic variability regarding broomrape resistance, which is hidden in tomato wild relatives, traditional varieties and accessions [155,156]. Indicatively, in a study in 2016, researchers evaluated a collection of 60 tomato accessions, consisted of cultivated varieties and wild relatives, screening the response of each genotype to P. aegyptiaca infesction, highlighting that screening and selection procedures can be considered as critical steps in pre-breeding to prioritize genetic material for further breeding purposes [157]. The second step is conventional/classical breeding strategies which can be based on the pre-breeding method mentioned above. Traditional breeding techniques, such as hybridization and backcrossing, with traditional or wild tomato genetic materials can effectively introgress favorable traits into S. lycopersicum. While these methods are time-consuming, they enable the transfer of robust genetic resistance from unexplored genetic material into cultivated tomato varieties [157,158]. In 2020 Ghani et.al, developed intraspecific hybrids using S. lycopersicum and two wild relatives, S. pimpinellifolium and Solanum habrochaites, known for their resistance against various abiotic and biotic stress tolerance, and other agronomic traits, claiming their potential use for broomrape tolerance in tomato [159]. Investigation of host resistance mechanisms is also proposed as an important finding to evolve breeding strategies in tomatoes. Regarding resistance against P.aegyptiaca, O. cernua and P.ramosa, a plethora of tomato varieties have been investigated, however only moderate level of resistance has been referred [90,160,161,162]. Rubiales examined the different resistance mechanisms in host tomato plants, including pre-attachment and post-attachment resistance, and how these can vary across broomrape species [163].Towards this end, Marker-Assisted Selection (MAS)would be highly preferable as a potential method for tomato breeding against broomrape parasitism, to efficiently select plants with broomrape resistant characteristics, speeding up the breeding process [153,157,164], however in the existing literature the available data about specific applications are still very few , mainly focusing on functional markers [148].
Several research works also highlight the importance of Quantitative Trait Loci (QTL) mapping to identify genetic regions associated with broomrape resistance [164]. Bai et al. (2020) investigated resistance to P. aegyptiaca in tomatoes by assessing 76 introgression lines (ILs) derived from S. pennellii LA0716. They identified 13 QTLs associated with various resistance traits distributed across several chromosomes. Specifically, IL6-2 and IL6-3, possessed QTLs located on chromosome 6, demonstrated profound tolerance to the holoparasite [165]. These findings are in accordance with recently published results in 2024, on early-stage parasitism by P. ramosa in these two ILs, which, in comparison to a susceptible commercial S. lycopersicum hybrid, revealed remarkable differences in the expression of genes found in several chromosomic regions, including chromosome 6), highly related to biotic stressors [154].
Conventional mutagenesis and TILLING (Targeting Induced Local Lesions IN Genomes) are valuable that alternatives for breeding against broomrape parasitism. Strigolactones (SLs) are plant hormones that play a crucial role in various plant processes, including the stimulation of seed germination in parasitic plants like Phelipanche and Orobanche species. Koltai et.al, (2010) produced through fast-neutron mutagenesis a tomato line resistant to broomrape, named Sl-ORT1 (S. lycopersicum Orobanche ResistantTrait 1). In that specific line they noticed that low transcript levels of SlCCD7 gene were involved in strigolactone’s(SLs) biosynthesis [166]. TILLING is a technique which can be used for the dentification and characterization of specific mutations within genes. Hasegawa et.al, in 2018 used two tomato TILLING genotypes (from Micro-Tom tomato variety) deficient in the CAROTENOID CLEAVAGE DIOXYGENASE 8 (CCD8) gene (which encodes enzymes that play crucial role in the biosynthesis of SLs), and they studied its relevant expression levels, revealing that the mutants down-regulated the expression of the CCD8 gene, highlighting the potential role of CCD8 to broomrape resistant mechanisms [167].
Stacking multiple resistance genes and engineering biochemical pathways, by implementing new breeding approaches and biotechnological tools is an emerging strategy [153,164]. Research utilizing molecular and biotechnological techniques has exploited the key role of strigolactones (SLs), suggesting that manipulating strigolactone pathways could be an effective strategy for developing crop varieties resistant to parasitic weeds [168,169,170]. RNA-interference (RNAi) is a method widely used in tomatoes to explore SLs resistance mechanisms. More specifically, silencing of the CCD7 and CCD8 genes, mentioned above, have been applied, leading to phenotype’s severe alteration [171,172]. Additionally, the expression of a multi-sequence RNAi construct designed to target three critical P. aegyptiaca genes (PaACS, PaM6PR, and PaPrx1) demonstrated resistance to broomrape infestation [173]. Moreover, other molecular studies utilizing -omic approaches, indicate that, in addition to the essential role of SLs-induced resistance mechanisms, multiple cellular processes may be involved in the tomato’s response to broomrapes [154].
Genome editing and transgenic approaches have also been proposed for broomrape management. Research highlights that CRISPR/Cas and other gene-editing technologies are creating new opportunities for targeted modifications in vegetable genomes, potentially strengthening resistance traits that traditional breeding cannot easily achieve. Several studies apply transgenic approaches to introduce resistance genes from other species. More specifically, Radi et.al. in 2006 studied the expression of the sarcotoxin IA gene -which is a gene originally identified in insects, characterized by antimicrobial activity and when expressed in plants provoked resistance against pathogens- driven by a root-specific promoter (tob promoter) in tomato plants. The genetic modification enhanced resistance of tomato plants against broomrape species [174]. Genome editing techniques like knock-out genes relevant to SLs biosynthesis (like CCD8, MAX1, P450 CYP722C) from tomato genotypes, demonstrated resistance against several broomrape species and different levels of pleotropic on phenotype [175,176,177,178]. While these methods are very promising, regulatory challenges and consumer acceptance continue to limit the broad use of transgenic/edited plants for broomrape resistance [153,164].
Overall, various studies recommend a combination of the afore mentioned breeding strategies, emphasizing the importance of integrating both classical and modern breeding techniques to develop durable resistance of tomato varieties and achieve long-term solutions to broomrape infestations in tomato crops[155,158,163] (Figure 2).
5.2. Legumes
Investigations regarding broomrape resistance have included major legumes, like faba bean, chickpeas, peas and lentils, although most research outcomes have arisen from research on faba bean.
Faba bean (Vicia faba L.) is one of the most important legume crops, worldwide, ranking fourth with respect to total production after chickpea (Cicer arietinum L.), field pea (Pisum sativum L.) and lentil (Lens culinaris L.) [179,180]. It is cultivated extensively in all continents with largest producers being China, Ethiopia, United Kingdom and Australia and constitutes a major grain legume in the Mediterranean basin used in human diet and as animal feed for its high protein content (FAOSTAT, 2023). In Tunisia it represents a predominant crop accounting for about 75% of total grain legume are with about 60 k ha and about 400k tons [181,182] and it is of equal importance in Egypt, Algeria and Morocco (FAO 2021) [183]. However, its productivity in the Mediterranean area has been unstable with significant fluctuations in agricultural yield due to a range of factors including abiotic and biotic stresses related to the severe climate change in the region. One of the main challenging abiotic factors is elevated temperature and drought in the area over the last several years as well as the catastrophic effect of infestations by holo-parasitic weeds of the Orobancaceae family. Especially in Tunisia, broomrape infestations are mainly caused by four species: O. crenata, O. foetida, O. cumana / cernua, and P. ramosa / aegyptiaca. Among these, O. foetida poses the most severe threat, with yield losses in faba beans reaching up to 90% [18,184,185] the difficulty to manage this parasite reside in its exceptional fertility that gives rise to about 50,000 to 500,000 seeds/plant [18].
A recent report on a population structure and genetic diversity analysis based on dominant marker ‘RAPD’ revealed a significant genetic disparity within individuals of each of the three studied genus and species respectively O. foetida, O. crenata and P. ramosa [185]. In this context, the three species were clustered into two main metapopulations and then into two genetic groups based on genius and species diversity levels. There were no geographic origin correlation as consequence of low levels of diversity between the populations; the breeding schemes for introgression of resistance to grain legumes against broomrapes can be conducted in one location [185]. In other reports, it was shown that O. foetida Poir. has two different varieties; one in north Africa, var. foetida in Tunisia and var. Broteri in Morocco and Spain [186]. In line with developing legume varieties resistant to broomrapes, previous studies has shown proven that the resistance can be broken by the mutation of the parasite itself which is a significant challenge for breeders [25].
Numerous strategies have been adopted over the years aiming at managing Orobanche parasitism including mechanical, chemical and biological control as well as cultivation practices. However, the capacity of these methods is limited in providing full protection to the host plant against Orobanche attacks. Another effective approach that holds promise and could work in synergy with the above constraining methods is the development of broomrape- resistant varieties through breeding programs.
Legume cultivars showing low susceptibility/partial resistance to Orobanche infestations constitute a valuable genetic resource for studying and understanding defense mechanisms and designing crop improvement programs towards superior genotypes with increased resistance to the parasite. Along these lines, characterization of different fava bean varieties, breeding lines and mutant lines with partial resistance to Orobanche and Phelipanche species highlight research efforts focusing on developing genotypes harboring efficient resistance mechanisms to counteract broomrape attacks [5,164]. Distinct fava bean varieties derived mostly from the Egyptian donor Giza402, showed different degrees of resistance to O.crenata which was most likely linked to prevention of parasite penetration through the vasculature structure of the host [187]. Experiments with two breeding lines, ‘Quijote’ and ‘Navio’, recorded low levels of O.crenate, O. foetida and P. aegyptica seed germination, possibly owing to decreased secretion of germination stimulants in root exudates [187,188]. The study of six faba bean accessions with partial resistance against various broomrape species indicated diversified resistance involving pre- and post- attachment resistance related processes [189]. Similarly, evaluation of a series of genotypes in field and controlled conditions indicated Giza 843, Misr1, and Misr3, as genotypes with good resistance to O. crenata and with significantly higher yields than the susceptible ones. Based on the low effects of parasitism on yield and vegetative growth of host plants, Giza 843 was selected as a promising candidate genotype to be introduced into Moroccan faba bean breeding programs [190]. Recently the Tunisian national faba bean breeding program led by the field crop laboratory of INRAT with collaboration with the CRRGC (the Regional Research Center of Field Crop of Béja) registered new partial resistant varieties to O.foetida and O.crenata in faba beans. These new cultivars belong to the small, seeded type, named “Chourouk” (i.e. with regular tannin), “Zaher” (i.e. with low tannin); and medium-seeded type named “Chams” with good field performance and broomrape resistance as compared to the susceptible varieties examined [18,78,79]. In this context, the first faba bean candidate variety (named Ammar) was deposited for registration in the Tunisian official national catalogue; field trials to be performed in 2024, 2025 in a multi-site field trial (Khamassi 2024, personal communication). All the above mentioned varieties showed a number of distinguished characteristics such as: low Orobanche spp seed germination stimulant production, low numbers of parasite seed germination, a restricted numbers of haustorium attachment and decreased growth of established tubercles; and hence resulting in a low number of final emerged shoots of Orobanche in the host plant [191,192,193,194,195].
Moreover, faba bean mutant lines were generated by gamma irradiation of the highly susceptible to O. crenata and O. foetida variety ‘Badi’. Lower levels of parasite seed germination were detected for these lines suggesting that developing resistant genotypes through mutagenesis may be plausible. However, these genetic material is still under research, facing registration challenges [196].Similar studies have been undertaken to explore the potential of pea germplasm in developing effective resistance to Orobanche infestations. A pea landrace (ROR12) known to guarantee good yields even in heavily infected fields was found to exhibit high levels of resistance to O. crenata, most likely due to interference with the parasite development process leading to delayed emergence of the weed. In addition, ROR12 root exudates displayed decreased strigolactone (SL) accumulation and their ability to stimulate germination of O. crenata seeds in vitro was significantly reduced. Interestingly, induction of the RAMOSUS1 (RMS1) gene encoding the CCD7 enzyme involved in SL biosynthesis was decreased in the infected host plants pointing to altered regulation of the SL biosynthetic pathway upon parasitism that maybe associated with establishment of resistance to O.crenata [197]. Considering the above observations, the ROR12 landrace could serve as a promising candidate for cultivation as well as introduction in breeding programmes aimed to the development of Orobanche resistant pea genotypes. Breeding efforts involving various crosses among P. fulvum, P. sativum ssp. elatius, P. sativum ssp. syriacum and pea landraces led to the development of pea lines with partial resistance to O.crenata. Resistance was associated with reduced tubercle formation both in rhizotron and field conditions and the derived pea lines displayed good performance and relatively high yields in O. crenata infected fields [5].
Delineating host-parasite interactions at the genetic-molecular level has lagged behind in commercial legume crops as compared to tomato where ample -omics and gene editing approaches have become feasible nowadays. However, recent transcriptomic findings in fava bean have begun to illuminate the molecular mechanisms underlying legume responses to parasitic infection and the molecular basis of resistance to Orobanche. An elaborate RNA-seq approach was deployed using a susceptible faba bean variety (Badii) and a resistant variety (Chourouk), at three developmental stages of O. foetida development (before germination, after germination, and tubercle stage) aiming at unraveling the intricate molecular processes underlying susceptibility/resistance in the faba bean/O. foetida interaction [198]. The transcriptomic profile revealed differential gene expression between the resistant and susceptible genotypes and among the different parasitism stages in pathways associated with secondary metabolites such as flavonoids, auxin, thiamine and jasmonic acid. Importantly, differences in gene expression were observed in genes involved in the orobanchol biosynthesis pathway such as the MAX1, MAX1B, and CYP711A3 homologues and in particular for the key gene CYP722C. Notably, knocking out this gene in tomato using CRISPR/Cas9–mediated genome editing resulted in undetectable orobanchol in tomato roots whereas architecture of the edited plant remained unaltered [178] suggesting that CYP722C could serve as good target for developing resistant varieties to broomrapes.
6. Conclusions
6.1. Management Strategies
Management of broomrapes in the Mediterranean cropping systems requires a strategic combination of various preventive and curative measures within the goal is to reduce the infestation and minimize the impact of the parasite on crop yield and quality. Preventive methods are the cornerstone for broomrape management because if the parasite is established, it becomes difficult to control. Preventive measures focus on disrupting the parasite's lifecycle and reducing the chances of infestation. There is a plethora of data for a long list of the preventive methods such as: rotations including trap, catch, allelopathic crops; intercropping; winter cover crops; modify sowing, transplanting dates; solarization; tillage, mechanical cultivation; Fertilization; use of synthetic stimulants for suicidal germination; polyethylene plastic mulching; use of crop tolerance inducers. Curative methods in broomrapes play a smaller role and the focus is on reducing its impact on the current crop. These measures generally aim to manage or limit the spread and damage caused by the parasite by the following tactics: seed dressing treatments, foliar herbicides, herbigation, biological control agents, and handweeding. Integrated management of broomrapes involves combining preventive and curative measures in a way that maximizes their effectiveness. A good example utilizing a combined strategy is the use of a partially resistant faba bean variety (i.e. Hashenge), use of bio-inoculants (R. leguminosarum, Trichoderma harzianum), and perform hand weeding at the flowering crop stage [85]. This integrated management strategy, when implemented together, has been shown to enhance the productivity of the crop while also reducing the impact of the parasitic weed.
6.2. Recognition of Parasitic Plant Molecular Patterns by Hosts
Understanding the recognition of parasitic plant molecular patterns by hosts underscore the complexity and significance of host-parasite interactions in plant ecology and evolution. Understanding these interactions on molecular level is crucial, as parasitic plants can significantly impact agricultural productivity and ecosystem dynamics. Host plants have evolved sophisticated mechanisms to recognize and respond to parasitic threats [123]. Primarily through the detection of pathogen-associated molecular patterns (PAMPs) unique to the parasites, host plants are able to recognize alien invaders.
The recognition of these patterns by host plants triggers a series of defense responses [104] that can halt or mitigate the parasitic invasion. This complex immune response is akin to the pattern recognition mechanisms observed in plant-pathogen interactions and highlights the co-evolutionary arms race between hosts and their parasitic adversaries.
Advancements in molecular biology and genomic technologies have significantly enhanced our capacity to identify these molecular patterns and understand the underlying recognition mechanisms. Unraveling these interactions contributes to our broader understanding of plant immunity and the evolutionary dynamics between species. Additionally, such insights could lead to the development of crops with improved resistance to parasitic plants, thereby enhancing food security and sustainable agricultural practices. Therefore, continued exploration of these interactions will not only advance our knowledge of plant biology but also offer new strategies for managing parasitic plant impacts in natural and agricultural ecosystems.
6.3. Hormonal Crosstalk Between Crop-Parasite
Plant hormones play a fundamental role in interactions between parasitic and crop plants, regulating processes from seed germination in parasitic plants to haustorium formation and host invasion. Strigolactones (SLs) are key chemical signals essential for the germination of parasitic seeds by indicating the proximity of the host, while auxins, gibberellins and ethylene are fundamental for the vascular connection between the parasite and the host, facilitating the flow of nutrients. Additionally, cytokinins (CKs) exuded by the host roots are crucial for haustorium formation, increasing the aggressiveness of the infection. Other defense hormones such as jasmonic acid (JA), salicylic acid (SA) and abscisic acid (ABA) play a key role in the host response to parasitism, activating resistance mechanisms to prevent the infections. Understanding how plant hormones mediate parasitic relationships will facilitate the development of more effective control strategies, such as the use of SL analogues or monocot trap crops. However, many of the hormonal mechanisms involved remain unclear, posing a challenge for optimizing parasitic plant management.
6.4. Breeding for Resistance
Research on broomrape resistance in tomato and legumes emphasizes the development of sustainable and cost-effective solutions by combining breeding strategies and techniques. In tomatoes species, the breeding target is utilizing both conventional methods and advanced molecular techniques to introduce favorable resistant traits. The challenge of these procedure is to keep the balance between resistance to broomrapes with other agronomic traits due to the complex host-parasite interactions. Similarly, in legumes like faba beans, breeding programs have identified the developed resistant varieties through genetic diversity analysis and traditional breeding approaches [163]. Concluding all the above, exploitation of plant breeding strategies and combination of classical and modern techniques is crucial for achieving durable resistance and ensuring long-term crop productivity and sustainability against broomrape infestations.
Author Contributions
“Conceptualization, D.C, E.T., A.K., M.G., F.A., M.A., K.K. ; methodology, D.C., A.K., M.G., P.M., M.A., K.K. ; software, D.C., M.G. ; validation, D.C., M.G. ; formal analysis, D.C., M.G.; investigation, D.C., E.T., M.G., P.M., M.A., K.K.; resources, D.C., A.K., M.G, P.M., M.A., K.K. ; data curation, D.C, M.G.; writing—original draft preparation, D.C., E.T., M.G., P.M., M.A., K.K. ; writing—review and editing, D.C., M.G. ; visualization, M.G. M.A. ; supervision, D.C. ; project administration, D.C.; funding acquisition, D.C., E.T, F.A., M.A., M.K. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Acknowledgments
The authors gratefully acknowledge the support of the project Zeroparasitic: Innovative sustainable solutions for broomrapes: prevention and integrated pest management approaches to overcome parasitism in Mediterranean cropping systems (Project ID: 1485). This work was carried out as part of a collaborative effort among all authors within the framework of the Zeroparasitic project, which provided valuable resources, knowledge exchange, and scientific insight essential for the development of this review.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Parker, C. Parasitic Weeds: A World Challenge. Weed science 2012, 60, 269–276. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Reboud, X.; Gibot-Leclerc, S. Broomrape Weeds. Underground Mechanisms of Parasitism and Associated Strategies for Their Control: A Review. Frontiers in plant science 2016, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, M.; Delavault, P.; Timko, M.P. Management of Infection by Parasitic Weeds: A Review. Plants 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in Parasitic Weeds Management in Legume Crops. A Review. Agronomy for Sustainable Development 2012, 32, 433–449. [Google Scholar] [CrossRef]
- Rubiales, D. Broomrape Threat to Agriculture. Outlooks on Pest Management 2020, 31, 141–145. [Google Scholar] [CrossRef]
- Eizenberg, H.; Goldwasser, Y. Control of Egyptian Broomrape in Processing Tomato: A Summary of 20 Years of Research and Successful Implementation. Plant Disease 2018, 102, 1477–1488. [Google Scholar] [CrossRef]
- Qasem, J.R. Broomrapes (Orobanche Spp.) the Challenge and Management: A Review. Jordan Journal of Agricultural Sciences 2021, 17, 117–150. [Google Scholar] [CrossRef]
- Dor, E.; Goldwasser, Y. “Parasitic Weeds: Biology and Control” Special Issue Editors Summary. Plants 2022, 11, 1891. [Google Scholar] [CrossRef]
- Negewo, T.; Ahmed, S.; Tessema, T.; Tana, T. Biological Characteristics, Impacts, and Management of Crenate Broomrape (Orobanche Crenata) in Faba Bean (Vicia Faba): A Review. Frontiers in Agronomy 2022, 4, 708187. [Google Scholar] [CrossRef]
- Rubiales, D. Managing Root Parasitic Weeds to Facilitate Legume Reintroduction into Mediterranean Rain-Fed Farming Systems. Soil Systems 2023, 7, 99. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. The Effect of Orobanche Crenata Infection Severity in Faba Bean, Field Pea, and Grass Pea Productivity. Frontiers in Plant Science 2016, 7, 1409. [Google Scholar] [CrossRef]
- Lops, F.; Frabboni, L.; Carlucci, A.; Tarantino, A.; Raimondo, M.L.; Disciglio, G. Management of Branched Broomrape in Field Processing Tomato Crop. In Tomato-From Cultivation to Processing Technology; IntechOpen: Rijeka, Croatia, 2022. [Google Scholar]
- Rubiales, D.; Fernández-Aparicio, M.; Wegmann, K.; Joel, D.M. Revisiting Strategies for Reducing the Seedbank of Orobanche and Phelipanche Spp. Weed Research 2009, 49, 23–33. [Google Scholar] [CrossRef]
- Grenz, J.H.; Manschadi, A.M.; Uygur, F.N.; Sauerborn, J. Effects of Environment and Sowing Date on the Competition between Faba Bean (Vicia Faba) and the Parasitic Weed Orobanche Crenata. Field Crops Research 2005, 93, 300–313. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Rao, N.K. Effect of Submergence on Bearing Capacity. Soils and Foundations 1975, 15, 61–66. [Google Scholar] [CrossRef]
- Zhelev, Z.; Zheleva, A.; Halacheva, K. Preparation of a β-Amanitin-Concanavalin A Conjugate of Low Toxicity. Toxicon 1987, 25, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.J.; Rodenburg, J. Parasitic Weed Management. 2023.
- Abbes, Z.; Kharrat, M.; Chaibi, W. Seed Germination and Tubercle Development of Orobanche Foetida and Orobanche Crenata in Presence of Different Plant Species. Tunisian Journal of Plant Protection 2008, 3, 101. [Google Scholar]
- Pérez-de-Luque, A.; Eizenberg, H.; Grenz, J.H.; Sillero, J.C.; Avila, C.; Sauerborn, J.; Rubiales, D. Broomrape Management in Faba Bean. Field Crops Research 2010, 115, 319–328. [Google Scholar] [CrossRef]
- Habimana, S.; Nduwumuremyi, A.; Chinama, R.J.D. Managementof Orobanche in Field Crops: A Review. Journal of soil science and plant nutrition 2014, 14, 43–62. [Google Scholar] [CrossRef]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; 1993.
- Jestin, C.; Molenat, D.; Boulet, C.; Bamme, B.; Leflon, M. Impact of Catch and Non-Host Crops in Cropping Systems to Control Branched Broomrape [Conference Poster]. 2013.
- Restuccia, A.; Marchese, M.; Mauromicale, G.; Restuccia, G. Biological Characteristics and Control of Orobanche Crenata Forsk., a Review. Italian Journal of Agronomy 2009, 4, 53–68. [Google Scholar] [CrossRef]
- Hegazi, E.; Zeid, A.; Attia, M.; Hasaneen, M.; Shall, A.; Eryan, M.; Aly, N.; Showiel, S.; El-Rahman, S.; Taleb, H.; et al. Effect of Intercropping by Flax, Radish and Fenugreek on Faba Bean, Vicia Faba L., Production and Reduction of Orobanche Crenata Forsk Seed Bank. AFF 2024, 13, 52–59. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Pérez-de-Luque, A.; Castillejo, M.A.; Prats, E.; Sillero, J.C.; Rispail, N.; Fondevilla, S. Breeding Approaches for Crenate Broomrape (Orobanche Crenata Forsk.) Management in Pea ( Pisum Sativum L.). Pest Management Science 2009, 65, 553–559. [Google Scholar] [CrossRef]
- Acharya, B.D.; Khattri, G.B.; Chettri, M.K.; Srivastava, S.C. Effect of Brassica Campestris Var. Toria as a Catch Crop on Orobanche Aegyptiaca Seed Bank. Crop protection 2002, 21, 533–537. [Google Scholar] [CrossRef]
- Nadal, S.; Moreno, M.T.; Cubero, J.I.; Rubiales, D. Determinate Faba Bean Young Pod Response to Glyphosate and Crenate Broomrape ( Orobanche Crenata). Journal of Sustainable Agriculture 2005, 25, 19–27. [Google Scholar] [CrossRef]
- Perronne, R.; Makowski, D.; Goffaux, R.; Montalent, P.; Goldringer, I. Temporal Evolution of Varietal, Spatial and Genetic Diversity of Bread Wheat between 1980 and 2006 Strongly Depends upon Agricultural Regions in France. Agriculture, Ecosystems & Environment 2017, 236, 12–20. [Google Scholar]
- Qasem, J.R.S. Parasitic Weeds of Jordan: Species, Hosts, Distribution and Management; Bentham Science Publishers, 2022.
- Fernández-Aparicio, M.; Emeran, A.A.; Rubiales, D. Control of Orobanche Crenata in Legumes Intercropped with Fenugreek (Trigonella Foenum-Graecum). Crop protection 2008, 27, 653–659. [Google Scholar] [CrossRef]
- Evidente, A.; Fernández-Aparicio, M.; Andolfi, A.; Rubiales, D.; Motta, A. Trigoxazonane, a Monosubstituted Trioxazonane from Trigonella Foenum-Graecum Root Exudate, Inhibits Orobanche Crenata Seed Germination. Phytochemistry 2007, 68, 2487–2492. [Google Scholar] [CrossRef]
- Khan, Z.R.; Hassanali, A.; Overholt, W.; Khamis, T.M.; Hooper, A.M.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Control of Witchweed Striga Hermonthica by Intercropping with Desmodium Spp., and the Mechanism Defined as Allelopathic. Journal of chemical ecology 2002, 28, 1871–1885. [Google Scholar] [CrossRef]
- Chon, S.-U.; Kim, Y.-M. Herbicidal Potential and Quantification of Suspected Allelochemicals from Four Grass Crop Extracts. J Agronomy Crop Science 2004, 190, 145–150. [Google Scholar] [CrossRef]
- Baghestani, A.; Lemieux, C.; Leroux, G.D.; Baziramakenga, R.; Simard, R.R. Determination of Allelochemicals in Spring Cereal Cultivars of Different Competitiveness. Weed Science 1999, 47, 498–504. [Google Scholar] [CrossRef]
- Aksoy, E.; Arslan, Z.F.; Tetik, Ö.; Eymirli, S. Using the Possibilities of Some Trap, Catch and Brassicaceaen Crops for Controlling Crenate Broomrape a Problem in Lentil Fields. International Journal of Plant Production 2016, 10, 53–62. [Google Scholar]
- Kitiş, Y.E.; Grenz, J.H.; Sauerborn, J. Effects of Some Cereal Root Exudates on Germination of Broomrapes (Orobanche Spp. and Phelipanche Spp.). Mediterranean Agricultural Sciences 2019, 32, 145–150. [Google Scholar] [CrossRef]
- Haidar, M.A.; Bibi, W.; Abdel-Khalek, N. Effect of Wheat and Barley Residues on Branched Broomrape (Orobanche Ramosa) Growth and Development in Potatoes. 1995.
- Hosh, M.; Tabbache, S.; Haddad, D.; Habak, H. The Effect of Dried Powdered Leaves of Radish (Rhaphanus Sativus L.) in Decreasing the Parasitism of Orobanche Ramosa L. on Tomato (Solanum Lycopersicum L.) Grown in Greenhouses. 2022.
- Calha, I.M.; Santos, J.; Amaral, A.; Cachado, J.; Nunes, A.P. Cover Cropping Effect on P. Ramosa Parasitation in Tomato under Controlled Conditions. In Proceedings of the Actas XVIII Congreso de la Sociedad Española de Malherbología; Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX), 2022; pp. 215–220.
- Kebreab, E.; Murdoch, A.J. Modelling the Effects of Water Stress and Temperature on Germination Rate of Orobanche Aegyptiaca Seeds. Journal of Experimental Botany 1999, 50, 655–664. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Jorrín, J.V.; Rubiales, D. Crenate Broomrape Control in Pea by Foliar Application of Benzothiadiazole (BTH). Phytoparasitica 2004, 32, 21–29. [Google Scholar] [CrossRef]
- Kemal, S.A. Narrowing the Yield Gap of Food Legumes through Integrated Management of Parasitic Weeds in the Highlands of Ethiopia. 2015.
- Rubiales, D.; Alcántara, C.; Gil, J.; Sillero, J.C. Infection of Chickpea (Cicer Arietinum) by Crenate Broomrape (Orobanche Crenata) as Influenced by Sowing Date and Weather Conditions. Agronomie 2003, 23, 359–362. [Google Scholar] [CrossRef]
- Kacan, K.; Tursun, N. Effect of Planting Time and Tomato Varieties on Broomrape (Phelipanche Aegyptiaca) Emergence and Tomato Yield in Western Turkey. Research on Crop 2012, 13, 1070–1077. [Google Scholar]
- Manschadi, A.M.; Sauerborn, J.; Stützel, H. Quantitative Aspects of Orobanche Crenata Infestation in Faba Beans as Affected by Abiotic Factors and Parasite Soil Seedbank. Weed Research 2001, 41, 311–324. [Google Scholar] [CrossRef]
- Jacobsohn, R.; Greenberger, A.; Katan, J.; Levi, M.; Alon, H. Control of Egyptian Broomrape (Orobanche Aegyptiaca) and Other Weeds by Means of Solar Heating of the Soil by Polyethylene Mulching. Weed Science 1980, 28, 312–316. [Google Scholar] [CrossRef]
- Mauromicale, G.; Monaco, A.L.; Longo, A.M.; Restuccia, A. Soil Solarization, a Nonchemical Method to Control Branched Broomrape (Orobanche Ramosa) and Improve the Yield of Greenhouse Tomato. Weed science 2005, 53, 877–883. [Google Scholar] [CrossRef]
- Boz, Ö.; Doğan, M.N.; Öğöt, D. The Effect of Duration of Solarization on Controlling Branched Broomrape (Phelipanche Ramosa L.) and Some Weed Species. 2012.
- Goldwasser, Y.; Rodenburg, J. Integrated Agronomic Management of Parasitic Weed Seed Banks. In Parasitic Orobanchaceae; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2013; pp. 393–413. ISBN 978-3-642-38145-4. [Google Scholar]
- Eizenberg, H.; Lande, T.; Achdari, G.; Roichman, A.; Hershenhorn, J. Effect of Egyptian Broomrape (Orobanche Aegyptiaca) Seed-Burial Depth on Parasitism Dynamics and Chemical Control in Tomato. Weed science 2007, 55, 152–156. [Google Scholar] [CrossRef]
- Ghersa, C.M.; Martınez-Ghersa, M.A. Ecological Correlates of Weed Seed Size and Persistence in the Soil under Different Tilling Systems: Implications for Weed Management. Field Crops Research 2000, 67, 141–148. [Google Scholar] [CrossRef]
- Van Delft, G.J.; Graves, J.D.; Fitter, A.H.; Van Ast, A. Striga Seed Avoidance by Deep Planting and No-Tillage in Sorghum and Maize. International Journal of Pest Management 2000, 46, 251–256. [Google Scholar] [CrossRef]
- Tarantino, E.; Lops, F.; Disciglio, G.; Carlucci, A.; Gatta, G.; Frabboni, L. Contain Phelipanche Ramosa on Processing Tomatoes. 2015.
- Disciglio, G.; Frabboni, L.; Tarantino, A.; Stasi, A. Association between Dynamic Agrivoltaic System and Cultivation: Viability, Yields and Qualitative Assessment of Medical Plants. Sustainability 2023, 15, 16252. [Google Scholar] [CrossRef]
- Jain, R.; Foy, C.L. Nutrient Effects on Parasitism and Germination of Egyptian Broomrape (Orobanche Aegyptiaca). Weed Technology 1992, 6, 269–275. [Google Scholar] [CrossRef]
- Parker, C. Observations on the Current Status of Orobanche and Striga Problems Worldwide. Pest Management Science 2009, 65, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Westwood, J.H.; Foy, C.L. Influence of Nitrogen on Germination and Early Development of Broomrape (Orobanche Spp.). Weed Science 1999, 47, 2–7. [Google Scholar] [CrossRef]
- Yoneyama, K.; Xie, X.; Kim, H.I.; Kisugi, T.; Nomura, T.; Sekimoto, H.; Yokota, T.; Yoneyama, K. How Do Nitrogen and Phosphorus Deficiencies Affect Strigolactone Production and Exudation? Planta 2012, 235, 1197–1207. [Google Scholar] [CrossRef]
- Johnson, A.W.; Rosebery, G.; Parker, C. A Novel Approach to Striga and Orobanche Control Using Synthetic Germination Stimulants. Weed Research 1976, 16, 223–227. [Google Scholar] [CrossRef]
- Babiker, A.G.T.; Hamdoun, A.M.; Rudwan, A.; Mansi, N.G.; Faki, H.H. Influence of Soil Moisture on Activity and Persistence of the Strigol Analogue GR 24. Weed Research 1987, 27, 173–178. [Google Scholar] [CrossRef]
- Zwanenburg, B.; Mwakaboko, A.S.; Kannan, C. Suicidal Germination for Parasitic Weed Control. Pest Management Science 2016, 72, 2016–2025. [Google Scholar] [CrossRef]
- Lechat, M.-M.; Brun, G.; Montiel, G.; Véronési, C.; Simier, P.; Thoiron, S.; Pouvreau, J.-B.; Delavault, P. Seed Response to Strigolactone Is Controlled by Abscisic Acid-Independent DNA Methylation in the Obligate Root Parasitic Plant, Phelipanche Ramosa L. Pomel. Journal of experimental botany 2015, 66, 3129–3140. [Google Scholar] [CrossRef]
- Hỳlová, A.; Pospíšil, T.; Spíchal, L.; Mateman, J.J.; Blanco-Ania, D.; Zwanenburg, B. New Hybrid Type Strigolactone Mimics Derived from Plant Growth Regulator Auxin. New biotechnology 2019, 48, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.; Hylova, A.; Soudek, P.; Petrova, S.; Spichal, L.; Vanek, T. Triazolide Strigolactone Mimics as Potent Selective Germinators of Parasitic Plant Phelipanche Ramosa. Pest Management Science 2019, 75, 2049–2056. [Google Scholar] [CrossRef]
- Cala, A.; Ghooray, K.; Fernández-Aparicio, M.; Molinillo, J.M.; Galindo, J.C.; Rubiales, D.; Macías, F.A. Phthalimide-derived Strigolactone Mimics as Germinating Agents for Seeds of Parasitic Weeds. Pest Management Science 2016, 72, 2069–2081. [Google Scholar] [CrossRef]
- Vouzounis, N.A.; Americanos, P.G. Control of Orobanche (Broomrape) in Tomato and Eggplant. 1998.
- Sokat, Y.; Demirkan, H. Research on the Methods for Controlling Broomrape (Phelipanche Ramosa (L.) Pomel.), Problem in Eggplant Production Areas in Turkey. 2020.
- Sillero, J.C.; Rojas-Molina, M.M.; Ávila, C.M.; Rubiales, D. Induction of Systemic Acquired Resistance against Rust, Ascochyta Blight and Broomrape in Faba Bean by Exogenous Application of Salicylic Acid and Benzothiadiazole. Crop Protection 2012, 34, 65–69. [Google Scholar] [CrossRef]
- Kusumoto, D.; Goldwasser, Y.; Xie, X.; Yoneyama, K.; Takeuchi, Y.; Yoneyama, K. Resistance of Red Clover (Trifolium Pratense) to the Root Parasitic Plant Orobanche Minor Is Activated by Salicylate but Not by Jasmonate. Annals of botany 2007, 100, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Bakewell-Stone, P. Orobanche Ramosa (Branched Broomrape) 2024, 37747.
- Al-Wakeel, S.A.M.; Moubasher, H.; Gabr, M.M.A.; Madany, M.M.Y. Induction of Systemic Resistance in Tomato Plants against Orobanche Ramosa L. Using Hormonal Inducers. 2012.
- Gonsior, G.; Buschmann, H.; Szinicz, G.; Spring, O.; Sauerborn, J. Induced Resistance—An Innovative Approach to Manage Branched Broomrape (Orobanche Ramosa) in Hemp and Tobacco. Weed science 2004, 52, 1050–1053. [Google Scholar] [CrossRef]
- Vurro, M.; Boari, A.; Pilgeram, A.L.; Sands, D.C. Exogenous Amino Acids Inhibit Seed Germination and Tubercle Formation by Orobanche Ramosa (Broomrape): Potential Application for Management of Parasitic Weeds. Biological Control 2006, 36, 258–265. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Simier, P.; Arfaoui, A.; Sifi, B.; Delavault, P.; Zourgui, L.; Belhadj, O. Induction of Phenolic Compounds in Pea ( Pisum Sativum L.) Inoculated by Rhizobium Leguminosarum and Infected with Orobanche Crenata. Journal of Phytopathology 2007, 155, 728–734. [Google Scholar] [CrossRef]
- Gozzo, F. Systemic Acquired Resistance in Crop Protection: From Nature to a Chemical Approach. J. Agric. Food Chem. 2003, 51, 4487–4503. [Google Scholar] [CrossRef]
- Khalil, S.; Malhotra, R.; Saxena, M.C.; Erskine, W.; Kharrat, M. Breeding Faba Bean for Orobanche Resistance. 2004.
- Nassib, A.M.; Ibrahim, A.A.; Saber, H.A. Broomrape (Orobanche Crenata) Resistance in Broad Beans: Breeding Work in Egypt. In Food legume improvement and development: Proceedings...; IDRC: Ottawa, ON, CA, 1978. [Google Scholar]
- Kharrat, M.; Abbes, Z.; Amri, M. A New Faba Bean Small Seeded Variety Najeh Tolerant to Orobanche Registered in the Tunisian Catalogue. Tunisian Journal of Plant Protection 2010, 5. [Google Scholar]
- Amri, M.; Trabelsi, I.; Abbes, Z.; Kharrat, M. Release of a New Faba Bean Variety" Chourouk" Resistant to the Parasitic Plants Orobanche Foetida and O. Crenata in Tunisia. 2019.
- Rubiales, D.; Flores, F.; Emeran, A.A.; Kharrat, M.; Amri, M.; Rojas-Molina, M.M.; Sillero, J.C. Identification and Multi-Environment Validation of Resistance against Broomrapes (Orobanche Crenata and Orobanche Foetida) in Faba Bean (Vicia Faba). Field Crops Research 2014, 166, 58–65. [Google Scholar] [CrossRef]
- El-Mehy, A.A.; El-Gendy, H.M.; Aioub, A.A.; Mahmoud, S.F.; Abdel-Gawad, S.; Elesawy, A.E.; Elnahal, A.S. Response of Faba Bean to Intercropping, Biological and Chemical Control against Broomrape and Root Rot Diseases. Saudi Journal of Biological Sciences 2022, 29, 3482–3493. [Google Scholar] [CrossRef]
- Rubiales, D.; Moral, A.; Flores, F. Agronomic Performance of Broomrape Resistant and Susceptible Faba Bean Accession. Agronomy 2022, 12, 1421. [Google Scholar] [CrossRef]
- Jurado-Expósito, M.; Castejón-Muñoz, M.; García-Torres, L. Broomrape (Orobanche Crenata) Control with Imazethapyr Applied to Pea (Pisum Sativum) Seed. Weed technology 1996, 10, 774–780. [Google Scholar] [CrossRef]
- Punia, S.S. Biology and Control Measures of Orobanche. 2014.
- Misganaw, M. Integrated Weed (Orobanche Crenata) Management on Faba Bean. American Journal of Agricultural Research 2016, 1, 0029–0034. [Google Scholar]
- Garcia-Torres, L.; Lopez-Granados, F. Progress of Herbicide Control of Broomrape (Orobanche Spp.) in Legumes and Sunflower (Helinanthus Annuus L.). 1991.
- Garcia-Torres, L.; Lopez-Granados, F.; Jurado-Exposito, M.; Díaz Sánchez, J. The Present State of Orobanche Spp. Infestations in Andalusia and the Prospects for Its Management. 1998.
- Foy, C.L.; Jacobsohn, R.; Jain, R. Screening of Lycopersicon Spp. for Glyphosate and/or Orobanche Aegyptiaca Pers. Resistance. Weed Research 1988, 28, 383–391. [Google Scholar] [CrossRef]
- Eizenberg, H.; Aly, R.; Cohen, Y. Technologies for Smart Chemical Control of Broomrape (Orobanche Spp. and Phelipanche Spp.). Weed Science 2012, 60, 316–323. [Google Scholar] [CrossRef]
- Hershenhorn, J.; Eizenberg, H.; Dor, E.; Kapulnik, Y.; Goldwasser, Y. Phelipanche Aegyptiaca Management in Tomato. Weed Research 2009, 49, 34–47. [Google Scholar] [CrossRef]
- Gressel, J. Enhancing Microbiocontrol of Weeds. Microbe 2003, 69, 498–502. [Google Scholar]
- Boari, A.; Vurro, M. Evaluation of Fusarium Spp. and Other Fungi as Biological Control Agents of Broomrape (Orobanche Ramosa). Biological Control 2004, 30, 212–219. [Google Scholar] [CrossRef]
- Gibot-Leclerc, S.; Guinchard, L.; Edel-Hermann, V.; Dessaint, F.; Cartry, D.; Reibel, C.; Gautheron, N.; Bernaud, E.; Steinberg, C. Screening for Potential Mycoherbicides within the Endophyte Community of Phelipanche Ramosa Parasitizing Tobacco. FEMS Microbiology Ecology 2022, 98, fiac024. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A. Biological Control of Orobanche Ramosa by Fusarium Solani. International Journal of Advanced Biological and Biomedical Research 2014, 2, 2751–2755. [Google Scholar]
- Vurro, M. Are Root Parasitic Broomrapes Still a Good Target for Bioherbicide Control? Pest Management Science 2024, 80, 10–18. [Google Scholar] [CrossRef]
- Klein, M.; Braverman, Y.; Chizov-Ginzburg, A.; Gol’berg, A.; Blumberg, D.; Khanbegyan, Y.; Hackett, K.J. Infectivity of Beetle Spiroplasmas for New Host Species. BioControl 2002, 47, 427–433. [Google Scholar] [CrossRef]
- Tóth, B.; Mesterházy, Á.; Horváth, Z.; Bartók, T.; Varga, M.; Varga, J. Genetic Variability of Central European Isolates of the Fusarium Graminearum Species Complex. European Journal of Plant Pathology 2005, 113, 35–45. [Google Scholar] [CrossRef]
- Kapralov, S.I. Phytomyza against Broomrape. 1974.
- Ranf, S. Sensing of Molecular Patterns through Cell Surface Immune Receptors. Current opinion in plant biology 2017, 38, 68–77. [Google Scholar] [CrossRef]
- Shiu, S.-H.; Bleecker, A.B. Plant Receptor-Like Kinase Gene Family: Diversity, Function, and Signaling. Sci. STKE 2001, 2001. [Google Scholar] [CrossRef]
- Shiu, S.-H.; Bleecker, A.B. Expansion of the Receptor-like Kinase/Pelle Gene Family and Receptor-like Proteins in Arabidopsis. Plant physiology 2003, 132, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.-H.; Li, W.-H. Origins, Lineage-Specific Expansions, and Multiple Losses of Tyrosine Kinases in Eukaryotes. Molecular biology and evolution 2004, 21, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Albert, M. Peptides as Triggers of Plant Defence. Journal of experimental botany 2013, 64, 5269–5279. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Duriez, P.; Vautrin, S.; Auriac, M.-C.; Bazerque, J.; Boniface, M.-C.; Callot, C.; Carrère, S.; Cauet, S.; Chabaud, M.; Gentou, F. A Receptor-like Kinase Enhances Sunflower Resistance to Orobanche Cumana. Nature Plants 2019, 5, 1211–1215. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, Z.; Cao, X.; Chen, M.; Chen, S.; Zhao, Q.; Zhao, S. A Leucine-Rich Repeat Receptor-like Protein Kinase Enhances Tomato Resistance to Phelipanche Aegyptiaca. Scientia Horticulturae 2024, 337, 113353. [Google Scholar] [CrossRef]
- Kaiser, B.; Vogg, G.; Fürst, U.B.; Albert, M. Parasitic Plants of the Genus Cuscuta and Their Interaction with Susceptible and Resistant Host Plants. Frontiers in plant science 2015, 6, 45. [Google Scholar] [CrossRef]
- Ihl, B.; Tutakhil, N.; Hagen, A.; Jacob, F. Studien an Cuscuta Reflexa Roxb: VII. Zum Abwehrmechanismus von Lycopersicon Esculentum Mill. Flora 1988, 181, 383–393. [Google Scholar] [CrossRef]
- Jiang, L.; Wijeratne, A.J.; Wijeratne, S.; Fraga, M.; Meulia, T.; Doohan, D.; Li, Z.; Qu, F. Profiling mRNAs of Two Cuscuta Species Reveals Possible Candidate Transcripts Shared by Parasitic Plants. PLoS ONE 2013, 8, e81389. [Google Scholar] [CrossRef]
- Ranjan, A.; Ichihashi, Y.; Farhi, M.; Zumstein, K.; Townsley, B.; David-Schwartz, R.; Sinha, N.R. Focus on Weed Control: De Novo Assembly and Characterization of the Transcriptome of the Parasitic Weed Dodder Identifies Genes Associated with Plant Parasitism. Plant Physiology 2014, 166, 1186. [Google Scholar] [CrossRef]
- Runyon, J.B.; Mescher, M.C.; Felton, G.W.; De Moraes, C.M. Parasitism by Cuscuta Pentagona Sequentially Induces JA and SA Defence Pathways in Tomato. Plant Cell & Environment 2010, 33, 290–303. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of Plant Growth by Cytokinin. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Werner, M.; Proksch, P.; Fry, S.C.; Kaldenhoff, R. The Cell Wall-Modifying Xyloglucan Endotransglycosylase/Hydrolase LeXTH1 Is Expressed during the Defence Reaction of Tomato against the Plant Parasite Cuscuta Reflexa. Plant Biology 2004, 6, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Hegenauer, V.; Fürst, U.; Kaiser, B.; Smoker, M.; Zipfel, C.; Felix, G.; Stahl, M.; Albert, M. Detection of the Plant Parasite Cuscuta Reflexa by a Tomato Cell Surface Receptor. Science 2016, 353, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Chitwood, D.H.; Kumar, R.; Headland, L.R.; Ranjan, A.; Covington, M.F.; Ichihashi, Y.; Fulop, D.; Jiménez-Gómez, J.M.; Peng, J.; Maloof, J.N. A Quantitative Genetic Basis for Leaf Morphology in a Set of Precisely Defined Tomato Introgression Lines. The Plant Cell 2013, 25, 2465–2481. [Google Scholar] [CrossRef]
- Eshed, Y.; Zamir, D. An Introgression Line Population of Lycopersicon Pennellii in the Cultivated Tomato Enables the Identification and Fine Mapping of Yield-Associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [CrossRef]
- Liebrand, T.W.H.; Van Den Berg, G.C.M.; Zhang, Z.; Smit, P.; Cordewener, J.H.G.; America, A.H.P.; Sklenar, J.; Jones, A.M.E.; Tameling, W.I.L.; Robatzek, S.; et al. Receptor-like Kinase SOBIR1/EVR Interacts with Receptor-like Proteins in Plant Immunity against Fungal Infection. Proc. Natl. Acad. Sci. U.S.A. 2013, 110, 10010–10015. [Google Scholar] [CrossRef]
- Liebrand, T.W.; van den Burg, H.A.; Joosten, M.H. Two for All: Receptor-Associated Kinases SOBIR1 and BAK1. Trends in plant science 2014, 19, 123–132. [Google Scholar] [CrossRef]
- Hegenauer, V.; Slaby, P.; Körner, M.; Bruckmüller, J.-A.; Burggraf, R.; Albert, I.; Kaiser, B.; Löffelhardt, B.; Droste-Borel, I.; Sklenar, J. The Tomato Receptor CuRe1 Senses a Cell Wall Protein to Identify Cuscuta as a Pathogen. Nature communications 2020, 11, 5299. [Google Scholar] [CrossRef]
- Fürst, U.; Hegenauer, V.; Kaiser, B.; Körner, M.; Welz, M.; Albert, M. Parasitic Cuscuta Factor(s) and the Detection by Tomato Initiates Plant Defense. Communicative & Integrative Biology 2016, 9, e1244590. [Google Scholar] [CrossRef]
- Johnsen, H.R.; Striberny, B.; Olsen, S.; Vidal-Melgosa, S.; Fangel, J.U.; Willats, W.G.T.; Rose, J.K.C.; Krause, K. Cell Wall Composition Profiling of Parasitic Giant Dodder ( C Uscuta Reflexa ) and Its Hosts: A Priori Differences and Induced Changes. New Phytologist 2015, 207, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Jhu, M.-Y.; Farhi, M.; Wang, L.; Philbrook, R.N.; Belcher, M.S.; Nakayama, H.; Zumstein, K.S.; Rowland, S.D.; Ron, M.; Shih, P.M. Heinz-Resistant Tomato Cultivars Exhibit a Lignin-Based Resistance to Field Dodder (Cuscuta Campestris) Parasitism. Plant Physiology 2022, 189, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Axtell, M.J.; Timko, M.P. Mechanisms of Resistance and Virulence in Parasitic Plant–Host Interactions. Plant physiology 2021, 185, 1282–1291. [Google Scholar] [CrossRef]
- Ohlson, E.W.; Timko, M.P. Race Structure of Cowpea Witchweed (Striga Gesnerioides) in West Africa and Its Implications for Striga Resistance Breeding of Cowpea. Weed science 2020, 68, 125–133. [Google Scholar] [CrossRef]
- Li, J.; Lis, K.E.; Timko, M.P. Molecular Genetics of Race-specific Resistance of Cowpea to Striga Gesnerioides (Willd.). Pest Management Science 2009, 65, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Timko, M.P. Gene-for-Gene Resistance in Striga -Cowpea Associations. Science 2009, 325, 1094. [Google Scholar] [CrossRef]
- Jubic, L.M.; Saile, S.; Furzer, O.J.; El Kasmi, F.; Dangl, J.L. Help Wanted: Helper NLRs and Plant Immune Responses. Current opinion in plant biology 2019, 50, 82–94. [Google Scholar] [CrossRef]
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The Haustorium, a Specialized Invasive Organ in Parasitic Plants. Annual Review of Plant Biology 2016, 67, 643–667. [Google Scholar] [CrossRef]
- Mishev, K.; Dobrev, P.I.; Lacek, J.; Filepová, R.; Yuperlieva-Mateeva, B.; Kostadinova, A.; Hristeva, T. Hormonomic Changes Driving the Negative Impact of Broomrape on Plant Host Interactions with Arbuscular Mycorrhizal Fungi. International Journal of Molecular Sciences 2021, 22, 13677. [Google Scholar] [CrossRef]
- Yoshida, S.; Shirasu, K. Plants That Attack Plants: Molecular Elucidation of Plant Parasitism. Current opinion in plant biology 2012, 15, 708–713. [Google Scholar] [CrossRef]
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere Signaling: Insights into Plant–Rhizomicrobiome Interactions for Sustainable Agronomy. Microorganisms 2022, 10, 899. [Google Scholar] [CrossRef] [PubMed]
- Aloni, R. Ecophysiological Implications of Vascular Differentiation and Plant Evolution. Trees 2015, 29, 1–16. [Google Scholar] [CrossRef]
- Ishida, J.K.; Wakatake, T.; Yoshida, S.; Takebayashi, Y.; Kasahara, H.; Wafula, E.; DePamphilis, C.W.; Namba, S.; Shirasu, K. Local Auxin Biosynthesis Mediated by a YUCCA Flavin Monooxygenase Regulates Haustorium Development in the Parasitic Plant Phtheirospermum Japonicum. The Plant Cell 2016, 28, 1795–1814. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.; Sinha, N.; Scholes, J. Parasitic Plants: Physiology, Development, Signaling, and Ecosystem Interactions. Plant Physiology 2021, 185, 1267–1269. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Shirasu, K.; Foo, E. Strigolactones in Plant Interactions with Beneficial and Detrimental Organisms: The Yin and Yang. Trends in plant science 2017, 22, 527–537. [Google Scholar] [CrossRef]
- Zagorchev, L.; Stöggl, W.; Teofanova, D.; Li, J.; Kranner, I. Plant Parasites under Pressure: Effects of Abiotic Stress on the Interactions between Parasitic Plants and Their Hosts. International Journal of Molecular Sciences 2021, 22, 7418. [Google Scholar] [CrossRef]
- Cheng, X.; Floková, K.; Bouwmeester, H.; Ruyter-Spira, C. The Role of Endogenous Strigolactones and Their Interaction with ABA during the Infection Process of the Parasitic Weed Phelipanche Ramosa in Tomato Plants. Frontiers in plant science 2017, 8, 392. [Google Scholar] [CrossRef]
- Inoue, M.; Xie, X.; Yoneyama, K. Barley Is a Potential Trap Crop for Root Parasitic Broomrape Weeds. Journal of Pesticide Science 2024, D24–D34. [Google Scholar] [CrossRef]
- Bao, Y.Z.; Yao, Z.Q.; Cao, X.L.; Peng, J.F.; Xu, Y.; Chen, M.X.; Zhao, S.F. Transcriptome Analysis of Phelipanche Aegyptiaca Seed Germination Mechanisms Stimulated by Fluridone, TIS108, and GR24. PLoS ONE 2017, 12, e0187539. [Google Scholar] [CrossRef]
- Cui, S.; Kubota, T.; Nishiyama, T.; Ishida, J.K.; Shigenobu, S.; Shibata, T.F.; Toyoda, A.; Hasebe, M.; Shirasu, K.; Yoshida, S. Ethylene Signaling Mediates Host Invasion by Parasitic Plants. Sci. Adv. 2020, 6, eabc2385. [Google Scholar] [CrossRef]
- Goyet, V.; Billard, E.; Pouvreau, J.-B.; Lechat, M.-M.; Pelletier, S.; Bahut, M.; Monteau, F.; Spíchal, L.; Delavault, P.; Montiel, G. Haustorium Initiation in the Obligate Parasitic Plant Phelipanche Ramosa Involves a Host-Exudated Cytokinin Signal. Journal of Experimental Botany 2017, 68, 5539–5552. [Google Scholar] [CrossRef] [PubMed]
- Harb, A.M.; Hameed, K.M.; Shibli, R.A. Effect of Triiodobenzoic Acid on Broomrape (Orobanche Ramosa) Infection and Development in Tomato Plants. The Plant Pathology Journal 2004, 20, 81–84. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, J.; Satheesh, V.; Meng, F.; Gao, W.; Dong, J.; Zheng, Z.; An, G.-Y.; Nussaume, L.; Liu, D. SHORT-ROOT Stabilizes PHOSPHATE1 to Regulate Phosphate Allocation in Arabidopsis. Nature Plants 2022, 8, 1074–1081. [Google Scholar] [CrossRef]
- Wakatake, T.; Ogawa, S.; Yoshida, S.; Shirasu, K. An Auxin Transport Network Underlies Xylem Bridge Formation between the Hemi-Parasitic Plant Phtheirospermum Japonicum and Host Arabidopsis. Development 2020, 147, dev187781. [Google Scholar] [CrossRef]
- Goyet, V.; Wada, S.; Cui, S.; Wakatake, T.; Shirasu, K.; Montiel, G.; Simier, P.; Yoshida, S. Haustorium Inducing Factors for Parasitic Orobanchaceae. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Smith, J.L.; De Moraes, C.M.; Mescher, M.C. Jasmonate- and Salicylate-mediated Plant Defense Responses to Insect Herbivores, Pathogens and Parasitic Plants. Pest Management Science 2009, 65, 497–503. [Google Scholar] [CrossRef]
- Rispail, N.; Dita, M.-A.; González-Verdejo, C.; Pérez-de-Luque, A.; Castillejo, M.-A.; Prats, E.; Román, B.; Jorrín, J.; Rubiales, D. Plant Resistance to Parasitic Plants: Molecular Approaches to an Old Foe. New Phytologist 2007, 173, 703–712. [Google Scholar] [CrossRef]
- Torres-Vera, R.; García, J.M.; Pozo, M.J.; López-Ráez, J.A. Expression of Molecular Markers Associated to Defense Signaling Pathways and Strigolactone Biosynthesis during the Early Interaction Tomato-Phelipanche Ramosa. Physiological and Molecular Plant Pathology 2016, 94, 100–107. [Google Scholar] [CrossRef]
- Frost, A.; López-Gutiérrez, J.C.; Purrington, C.B. Fitness of Cuscuta Salina (Convolvulaceae) Parasitizing Beta Vulgaris (Chenopodiaceae) Grown under Different Salinity Regimes. American Journal of Botany 2003, 90, 1032–1037. [Google Scholar] [CrossRef]
- Wiese, A.; Elzinga, N.; Wobbes, B.; Smeekens, S. A Conserved Upstream Open Reading Frame Mediates Sucrose-Induced Repression of Translation. The Plant Cell 2004, 16, 1717–1729. [Google Scholar] [CrossRef]
- Foyer, C.H.; Rasool, B.; Davey, J.W.; Hancock, R.D. Cross-Tolerance to Biotic and Abiotic Stresses in Plants: A Focus on Resistance to Aphid Infestation. Journal of experimental botany 2016, 67, 2025–2037. [Google Scholar] [CrossRef]
- Martínez-Melgarejo, P.A.; Ormazabal, M.; López-García, J.A.; Guerrero-Franco, J.J.; Martín-Rodríguez, J.A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Solanum Rootstock Biodiversity to Deal with Holoparasitism and Fertilizer Use Efficiency. In Proceedings of the IV Asian Horticultural Congress-AHC2023 1404; 2023; pp. 513–520.
- Baghyalakshmi, K.; Sarala, K.; Prabhakararao, K.; Reddy, D.D. Orobanche Menace in Crop Plants: Host Resistance as a Potential Tool to Control. Journal of Pharmacognosy and Phytochemistry 2019, 8, 93–102. [Google Scholar]
- Gerakari, M.; Kotsira, V.; Kapazoglou, A.; Tastsoglou, S.; Katsileros, A.; Chachalis, D.; Hatzigeorgiou, A.G.; Tani, E. Transcriptomic Approach for Investigation of Solanum Spp. Resistance upon Early-Stage Broomrape Parasitism. Current Issues in Molecular Biology 2024, 46, 9047. [Google Scholar] [CrossRef] [PubMed]
- Tokasi, S.; Bannayan Aval, M.; Mashhadi, H.R.; Ghanbari, A. Screening of Resistance to Egyptian Broomrape Infection in Tomato Varieties. Planta Daninha 2014, 32, 109–116. [Google Scholar] [CrossRef]
- Kapazoglou, A.; Gerakari, M.; Lazaridi, E.; Kleftogianni, K.; Sarri, E.; Tani, E.; Bebeli, P.J. Crop Wild Relatives: A Valuable Source of Tolerance to Various Abiotic Stresses. Plants 2023, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Sawafta, M.; Shtaya, M. Search for Resistance to Egyptian Broomrape (Orobanche Aegyptiaca) in Tomato Germplasm. 2016.
- Sawafta, M.S. Resistance of Some Tomato Species to Orobanche Aegyptiaca (Comparative Study). PhD Thesis, 2012.
- Ghani, M.A.; Abbas, M.M.; Amjad, M.; Ziaf, K.; Ali, B.; Shaheen, T.; Awan, F.S.; Khan, A.N. Production and Characterisation of Tomato Derived from Interspecific Hybridisation between Cultivated Tomato and Its Wild Relatives. The Journal of Horticultural Science and Biotechnology 2020, 95, 506–520. [Google Scholar] [CrossRef]
- Dalela, G.G.; Mathur, R.L. Resistance of Varieties of Eggplant, Tomato and Tobacco to Broomrape (Orobanche Cernua Loefl.). PANS Pest Articles & News Summaries 1971, 17, 482–483. [Google Scholar] [CrossRef]
- Avdeyev, Y.I.; Scherbinin, B.M.; Ivanova, L.M.; Avdeyev, A.Y. Studying of Tomato Resistance to Broomrape and Breeding Varieties for Processing. In Proceedings of the VIII International Symposium on the Processing Tomato 613; 2002; pp. 283–290.
- Draie, R. Differential Responses of Commercial Tomato Rootstocks to Branched Broomrape. Science and Education 2017, 5, 15–25. [Google Scholar] [CrossRef]
- Rubiales, D.; Araújo, S.S.; Vaz Patto, M.C.; Rispail, N.; Valdés-López, O. Advances in Legume Research. Frontiers in Plant Science 2018, 9, 501. [Google Scholar] [CrossRef]
- Cuccurullo, A.; Nicolia, A.; Cardi, T. Resistance against Broomrapes (Orobanche and Phelipanche Spp.) in Vegetables: A Comprehensive View on Classical and Innovative Breeding Efforts. Euphytica 2022, 218, 82. [Google Scholar] [CrossRef]
- Exploration of Resistance to Phelipanche Aegyptiaca in Tomato - Bai - 2020 - Pest Management Science - Wiley Online Library. Available online: https://scijournals.onlinelibrary.wiley.com/doi/abs/10.1002/ps.5932?casa_token=6C7CoD5d5SkAAAAA%3ACMmxpiLH4ef35n7x8bfzPRp7GiGX7rm2bfpkqs43fHiWYI6l5c6AVEhpzzaiHUIgR1t_f8POcYswGJLP (accessed on 9 January 2025).
- Koltai, H.; LekKala, S.P.; Bhattacharya, C.; Mayzlish-Gati, E.; Resnick, N.; Wininger, S.; Dor, E.; Yoneyama, K.; Yoneyama, K.; Hershenhorn, J. A Tomato Strigolactone-Impaired Mutant Displays Aberrant Shoot Morphology and Plant Interactions. Journal of Experimental Botany 2010, 61, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Tsutsumi, T.; Fukushima, S.; Okabe, Y.; Saito, J.; Katayama, M.; Shindo, M.; Yamada, Y.; Shimomura, K.; Yoneyama, K. Low Infection of Phelipanche Aegyptiaca in Micro-Tom Mutants Deficient in CAROTENOID CLEAVAGE DIOXYGENASE 8. International journal of molecular sciences 2018, 19, 2645. [Google Scholar] [CrossRef] [PubMed]
- Dor, E.; Yoneyama, K.; Wininger, S.; Kapulnik, Y.; Yoneyama, K.; Koltai, H.; Xie, X.; Hershenhorn, J. Strigolactone Deficiency Confers Resistance in Tomato Line SL-ORT1 to the Parasitic Weeds Phelipanche and Orobanche Spp. Phytopathology® 2011, 101, 213–222. [Google Scholar] [CrossRef]
- Karniel, U.; Koch, A.; Bar Nun, N.; Zamir, D.; Hirschberg, J. Tomato Mutants Reveal Root and Shoot Strigolactone Involvement in Branching and Broomrape Resistance. Plants 2024, 13, 1554. [Google Scholar] [CrossRef] [PubMed]
- En-Nahli, Y.; Abbes, Z.; Trabelsi, I.; Makaza, W.; Kharrat, M.; Ghanem, M.E.; Es-Safi, N.E.; Amri, M. Host–Parasitic Plant Interaction: The Key Role of Strigolactones. In Strigolactones. In Strigolactones, Alkamides and Karrikins in Plants; CRC Press: Boca Raton, FL, USA, 2023; pp. 87–99. [Google Scholar]
- Vogel, J.T.; Walter, M.H.; Giavalisco, P.; Lytovchenko, A.; Kohlen, W.; Charnikhova, T.; Simkin, A.J.; Goulet, C.; Strack, D.; Bouwmeester, H.J.; et al. SlCCD7 Controls Strigolactone Biosynthesis, Shoot Branching and Mycorrhiza-Induced Apocarotenoid Formation in Tomato: SlCCD7 Controls Tomato Branching. The Plant Journal 2009, 61, 300–311. [Google Scholar] [CrossRef]
- Kohlen, W.; Charnikhova, T.; Lammers, M.; Pollina, T.; Tóth, P.; Haider, I.; Pozo, M.J.; De Maagd, R.A.; Ruyter-Spira, C.; Bouwmeester, H.J.; et al. The Tomato CAROTENOID CLEAVAGE DIOXYGENASE 8 ( S l CCD 8 ) Regulates Rhizosphere Signaling, Plant Architecture and Affects Reproductive Development through Strigolactone Biosynthesis. New Phytologist 2012, 196, 535–547. [Google Scholar] [CrossRef]
- Dubey, N.K.; Eizenberg, H.; Leibman, D.; Wolf, D.; Edelstein, M.; Abu-Nassar, J.; Marzouk, S.; Gal-On, A.; Aly, R. Enhanced Host-Parasite Resistance Based on Down-Regulation of Phelipanche Aegyptiaca Target Genes Is Likely by Mobile Small RNA. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Radi, A.; Dina, P.; Guy, A. Expression of Sarcotoxin IA Gene via a Root-Specific Tob Promoter Enhanced Host Resistance against Parasitic Weeds in Tomato Plants. Plant Cell Rep 2006, 25, 297–303. [Google Scholar] [CrossRef]
- Bari, V.K.; Nassar, J.A.; Kheredin, S.M.; Gal-On, A.; Ron, M.; Britt, A.; Steele, D.; Yoder, J.; Aly, R. CRISPR/Cas9-Mediated Mutagenesis of CAROTENOID CLEAVAGE DIOXYGENASE 8 in Tomato Provides Resistance against the Parasitic Weed Phelipanche Aegyptiaca. Scientific reports 2019, 9, 11438. [Google Scholar] [CrossRef]
- Bari, V.K.; Nassar, J.A.; Aly, R. CRISPR/Cas9 Mediated Mutagenesis of MORE AXILLARY GROWTH 1 in Tomato Confers Resistance to Root Parasitic Weed Phelipanche Aegyptiaca. Scientific reports 2021, 11, 3905. [Google Scholar] [CrossRef]
- Bari, V.K.; Nassar, J.A.; Meir, A.; Aly, R. Targeted Mutagenesis of Two Homologous ATP-Binding Cassette Subfamily G (ABCG) Genes in Tomato Confers Resistance to Parasitic Weed Phelipanche Aegyptiaca. J Plant Res 2021, 134, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, T.; Moriyama, D.; Miyamoto, A.; Okamura, H.; Shiotani, N.; Shimizu, N.; Mizutani, M.; Takikawa, H.; Sugimoto, Y. Identification of Novel Canonical Strigolactones Produced by Tomato. Frontiers in Plant Science 2022, 13, 1064378. [Google Scholar] [CrossRef] [PubMed]
- Punia, S.; Dhull, S.B.; Sandhu, K.S.; Kaur, M. Faba Bean ( Vicia Faba ) Starch: Structure, Properties, and in Vitro Digestibility—A Review. Legume Science 2019, 1, e18. [Google Scholar] [CrossRef]
- Alharbi, N.H.; Adhikari, K.N. Factors of Yield Determination in Faba Bean (Vicia Faba). Crop Pasture Sci. 2020, 71, 305–321. [Google Scholar] [CrossRef]
- Khamassi, K.; Babay, E.; Rouissi, M.; Dakhlaoui, A.; Ben Ayed, R.; Hanana, M. Genetic Variability of Tunisian Faba Beans (Vicia Faba L.) Based on Seeds’ Morphophysical Properties as Assessed by Statistical Analysis. Journal of Food Quality 2021, 2021, 1–10. [Google Scholar] [CrossRef]
- Babay, E.; Khamassi, K.; Sabetta, W.; Miazzi, M.M.; Montemurro, C.; Pignone, D.; Danzi, D.; Finetti-Sialer, M.M.; Mangini, G. Serendipitous in Situ Conservation of Faba Bean Landraces in Tunisia: A Case Study. Genes 2020, 11, 236. [Google Scholar] [CrossRef]
- Hadou el hadj, D.; Tellah, S.; Goumeida, K.; Aitouakli, S.; Tifest, C.; Ammi, N.; Ratet, P.; Pulvento, C.; Sellami, M.H. Evaluation of Adaptability of Different Faba Bean Landraces under Mediterranean Field Conditions of Central-Northern Algeria. Agronomy 2022, 12, 1660. [Google Scholar] [CrossRef]
- Bouraoui, M.; Abbes, Z.; Rouissi, M.; Abdi, N.; Hemissi, I.; Kouki, S.; Sifi, B. Effect of Rhizobia Inoculation, N and P Supply on Orobanche Foetida Parasitising Faba Bean (Vicia Faba Minor) under Field Conditions. Biocontrol Science and Technology 2016, 26, 776–791. [Google Scholar] [CrossRef]
- Khamassi, K.; Abbes, Z.; Tani, E.; Katsileros, A.; Guenni, K.; Rouissi, M.; Khoufi, S.; Chaabane, R.; Chachalis, D.; Kharrat, M. Genetic Structure and Diversity Analysis of Tunisian Orobanche Spp. and Phelipanche Spp. Using Molecular Markers. Applied Sciences 2023, 13, 11622. [Google Scholar] [CrossRef]
- Pujadas-Salvà, A.J.; Plaza-Arregui, L. Studies on Thapsia (Apiaceae) from North-Western Africa: A Forgotten and a New Species. Botanical Journal of the Linnean Society 2003, 143, 433–442. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. Escape and True Resistance to Crenate Broomrape (Orobanche Crenata Forsk.) in Grass Pea (Lathyrus Sativus L.) Germplasm. Field Crops Research 2012, 125, 92–97. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Kisugi, T.; Xie, X.; Rubiales, D.; Yoneyama, K. Low Strigolactone Root Exudation: A Novel Mechanism of Broomrape ( Orobanche and Phelipanche Spp.) Resistance Available for Faba Bean Breeding. J. Agric. Food Chem. 2014, 62, 7063–7071. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Rojas-Molina, M.M.; Sillero, J.C. Characterization of Resistance Mechanisms in Faba Bean (Vicia Faba) against Broomrape Species (Orobanche and Phelipanche Spp.). Frontiers in Plant Science 2016, 7, 1747. [Google Scholar] [CrossRef] [PubMed]
- Briache, F.Z.; El Amri, M.; Ennami, M.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Induction of Systemic Resistance to Orobanche Crenata in Lentil by Exogenous Application of Salicylic Acid and Indole Acetic Acid. Journal of Plant Protection Research 2023, 83–96. [Google Scholar]
- Thebti, S.; Bouallegue, A.; Rzigui, T.; En-Nahli, Y.; Horchani, F.; Hosni, T.; Kharrat, M.; Amri, M.; Abbes, Z. Potential Physiological Tolerance Mechanisms in Faba Bean to Orobanche Spp. Parasitism. Frontiers in Plant Science 2024, 15, 1497303. [Google Scholar] [CrossRef]
- Abbes, Z.; Kharrat, M.; Delavault, P.; Chaïbi, W.; Simier, P. Nitrogen and Carbon Relationships between the Parasitic Weed Orobanche Foetida and Susceptible and Tolerant Faba Bean Lines. Plant Physiology and Biochemistry 2009, 47, 153–159. [Google Scholar] [CrossRef]
- Abbes, Z.; Bouallegue, A.; Trabelsi, I.; Trabelsi, N.; Taamalli, A.; Amri, M.; Mhadhbi, H.; Kharrat, M. Investigation of Some Biochemical Mechanisms Involved in the Resistance of Faba Bean (Vicia Faba L.) Varieties to Orobanche Spp. Plant Protection Science 2020, 56. [Google Scholar] [CrossRef]
- Trabelsi, I.; Abbes, Z.; Amri, M.; Kharrat, M. Study of Some Resistance Mechanisms to Orobanche Spp. Infestation in Faba Bean ( Vicia Faba L.) Breeding Lines in Tunisia. Plant Production Science 2016, 19, 562–573. [Google Scholar] [CrossRef]
- Trabelsi, I.; Yoneyama, K.; Abbes, Z.; Amri, M.; Xie, X.; Kisugi, T.; Kim, H.I.; Kharrat, M. Characterization of Strigolactones Produced by Orobanche Foetida and Orobanche Crenata Resistant Faba Bean (Vicia Faba L.) Genotypes and Effects of Phosphorous, Nitrogen, and Potassium Deficiencies on Strigolactone Production. South African Journal of Botany 2017, 108, 15–22. [Google Scholar] [CrossRef]
- Mejri, S.; Mabrouk, Y.; Belhadj, O.; Saidi, M. Orobanche Foetida Resistance in Two New Faba Bean Genotypes Produced by Radiation Mutagenesis. International Journal of Radiation Biology 2018, 94, 671–677. [Google Scholar] [CrossRef]
- Pavan, S.; Schiavulli, A.; Marcotrigiano, A.R.; Bardaro, N.; Bracuto, V.; Ricciardi, F.; Charnikhova, T.; Lotti, C.; Bouwmeester, H.; Ricciardi, L. Characterization of Low-Strigolactone Germplasm in Pea (Pisum Sativum L.) Resistant to Crenate Broomrape (Orobanche Crenata Forsk.). MPMI 2016, 29, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Boukteb, A.; Sato, K.; Gan, P.; Kharrat, M.; Sakouhi, H.; Shibata, A.; Shirasu, K.; Ichihashi, Y.; Bouhadida, M. Global Changes in Gene Expression during Compatible and Incompatible Interactions of Faba Bean (Vicia Faba L.) during Orobanche Foetida Parasitism. PLoS ONE 2024, 19, e0301981. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
A holistic approach of breeding strategies to create tomato resistant genotypes to broomrapes.
Figure 2.
A holistic approach of breeding strategies to create tomato resistant genotypes to broomrapes.
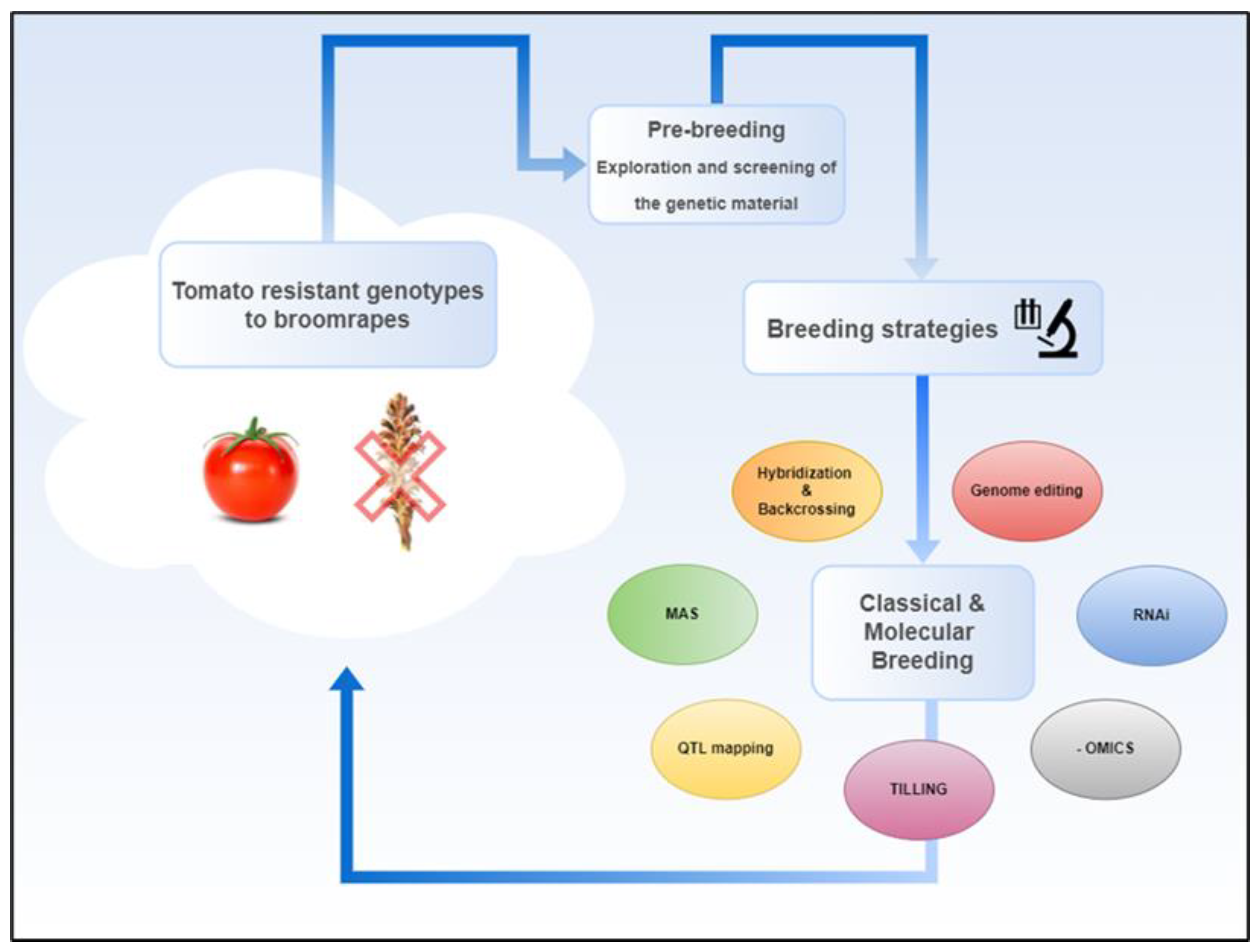

Table 1.
Current status of the list of preventive and curative techniques for controlling broomrapes in major Mediterranean crops such as legumes and industrial tomato. The efficacy levels are based on expert estimation after reviewing the related literature. The mechanism and the primary effects of the techniques refer to parasite life cycle and broomrape stage targeted.
Table 1.
Current status of the list of preventive and curative techniques for controlling broomrapes in major Mediterranean crops such as legumes and industrial tomato. The efficacy levels are based on expert estimation after reviewing the related literature. The mechanism and the primary effects of the techniques refer to parasite life cycle and broomrape stage targeted.
| Technique | Efficacy Levels(a) | Mechanism; primary Effects | |
|---|---|---|---|
| Legumes | Ind. tomato | ||
| I. Preventive | |||
| Rotation | + | + | (M): Disruption of the parasite life cycle by avoiding soil seed bank replenishing (E): soil seed bank; seed germination; pre- and early-attachment of haustorium |
| Winter cover crops | na1 | + | (M): Disruption of the parasite life cycle by creating a hostile soil environment (E): soil seed bank; seed germination; pre-attachment of haustorium |
| Use of trap, catch allelopathic crops; intercropping | + | + | (M): Disruption of the parasite life cycle by reducing final haustorium numbers (multiple tactics) (E): soil seed bank; seed germination; pre-attachment of haustorium |
| Modify sowing, transplanting dates | + | + | (M): Disruption of best conditions for the onset of parasitism (E): soil seed bank; seed germination; pre- and early-attachment of haustorium |
| Solarization | +++ | na2 | (M): Reduction of active soil seed bank by seed decay (E): soil seed bank; seed germination |
| Tillage, mechanical cultivation | ++ | ++ | (M): Reduction of active soil seed bank by physical misplacement of seeds (E): soil seed bank; seed germination |
| Fertilization | + | ++ | (M): Disruption of the parasite life cycle by employing direct (on the parasite seeds) and indirect (on enhancing crop competitive ability) conditions (E): soil seed bank; seed germination; pre-, early-, and late-attachment of haustorium |
| Use of synthetic stimulants | + | + | (M): Lower the onset of parasitism by suicidal germination tactics (E): soil seed bank; seed germination |
| Polyethylene plastic mulching | na3 | +++ | (M): Disruption of the parasite life cycle by physical barriers (E): seed germination; pre-, early-, and late-attachment of haustorium |
| Use of crop tolerance inducers | + | + | (M): Disruption of the parasite life cycle by enhancing crop competitive ability (E): seed germination; pre-, and early-attachment of haustorium |
| Use of resistant/tolerant cvs | ++ | na4 | (M): Low or minimum onset of parasitism by natural crop tolerance/resistance development (E): seed germination; pre-, and early-attachment of haustorium |
| II. Curative | |||
| Seed dressing treatments | ++ | ++ | (M): Lower onset of parasitism by utilizing synthetic substances applied as seed coating, film or pellets (E): seed germination; pre-, and early-attachment of haustorium |
| Foliar herbicides | ++ | + | (M): Lower onset of parasitism by utilizing herbicides to reach target sites via translocation of the haustorium (E): seed germination; pre-, and early-attachment of haustorium |
| Herbigation | na5 | ++ | (M): Lower onset of parasitism by utilizing herbicides to be applied with irrigation (E): seed germination; pre-, early- and late-attachment of haustorium |
| Biological control; beneficial insects | na6 | (M): Disruption of the parasite life cycle by biological agents (multiple tactics) (E): soil seed bank; seed germination; pre-, and early-attachment of haustorium |
|
| Hand weeding | +++ | +++ | (M): Disruption of the parasite life cycle by physical weed removal (E): soil seed bank; late-attachment of haustorium |
(a): Efficacy levels: +, low level; ++ medium level; +++ high level. 1: Non applicable: No timing window for employing winter cover crops. 2: Non applicable: No timing window for employing solarization in summer crops unless fallow. 3: Non applicable: Such a cropping system does not exist in legumes. 4: Non applicable: No current commercial resistant or tolerant cv available in industrial tomato (only research outcomes). 5: Non applicable: Such a strategy (application of herbicides using drip irrigation) does not exist in legumes. 6: Non applicable. No current commercial biological control agent available in industrial tomato (only research outcomes).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



