
Submitted:
09 April 2025
Posted:
09 April 2025
You are already at the latest version
Abstract
As life expectancy continues to increase, it becomes increasingly important to extend healthspan by targeting mechanisms associated with aging. Cellular senescence is recognized as a significant contributor to aging and neurodegenerative disorders. This review examines the emerging role of nutraceuticals and functional foods as potential modulators of cellular senescence, which may, in turn, influence the development of neurodegenerative diseases. An analysis of both epidemiological and experimental studies indicates that bioactive compounds, including polyphenols, vitamins, and spices, possess substantial antioxidants, anti-inflammatory, and epigenetic properties. These nutritional senotherapeutic agents effectively scavenge reactive oxygen species, modulate gene expression, and decrease the secretion of senescence-associated secretory phenotype factors, minimizing cellular damage. Nutraceuticals can enhance mitochondrial function, reduce oxidative stress, and regulate inflammation, key factors in aging and diseases like Alzheimer's and Parkinson's. Furthermore, studies reveal that specific bioactive compounds can reduce senescence markers in cellular models, while others exhibit senostatic and senolytic properties, both directly and indirectly. Diets enriched with these nutraceuticals, such as the Mediterranean diet, have been correlated with improved brain health and the deceleration of aging. Despite these promising outcomes, direct evidence linking these compounds to reducing senescent cell numbers remains limited, highlighting the necessity for further inquiry. This review presents compelling arguments for the potential of nutraceuticals and functional foods to promote longevity and counteract neurodegeneration by exploring their molecular mechanisms. The emerging relationship between dietary bioactives and cellular senescence sets the stage for future research to develop effective preventive and therapeutic strategies for age-related diseases.
Keywords:
Cellular senescence
; Inflammaging
; Healthy aging
; Neurodegenerative diseases
; Nutraceuticals
; Functional foods
; Alzheimer's disease
; Parkinson’s disease
1. Introduction and Background
Life expectancy has dramatically increased in nearly all nations, and the global population has tripled since the mid-twentieth century. By 2030, the global human population is projected to grow to approximately 8.5 billion, with an additional 1.18 billion people expected in the following two decades, reaching 9.7 billion in 2050 [1]. Aging is rapidly accelerating worldwide. By 2050, the number of people over 65 is expected to more than double, reaching 1.5 billion, representing 16% of the global population. While this trend is more intense in developed countries - where 26% of the European and North American population are over 65 - it has also become a significant global phenomenon that affects developing countries [2,3]. Nevertheless, insufficient evidence suggests that an increase in longevity correlates with a more extended period of good health [4]. Indeed, a notable difference exists between lifespan, defined as the total years lived, and healthspan, which refers to the duration without disease [5]. Extending lifespan without postponing the onset of diseases or lessening their severity would worsen the healthspan-lifespan gap. Advanced age is marked by the emergence of various complex health conditions, such as age-related diseases (ARDs) and geriatric syndromes (GSs), also referred to as “chronic or non-communicable” diseases, that are the leading cause of mortality and disability worldwide [6].
Aging is an inescapable, natural and universal feature of most living organisms that results from environmental, genetic, epigenetic and stochastic factors, each contributing to the overall phenotype [7,8]. As humans age, damaging changes accumulate in the molecules, cells, and tissues, leading to a decline in normal physiological functions and a reduced ability to maintain adequate homeostasis. The increased susceptibility to various stressors and reduced ability to adapt to the environment lead to clinical diseases, where genetic, epigenetic and environmental factors play a key role [9]. Geroscience provides a new perspective on gerontology by investigating the link between aging and ARDs. Both epidemiological evidence and experimental research demonstrate that aging is the principal risk factor for ARDs and GSs. Geroscience posits that aging and ARDs/GSs share a fundamental set of biological mechanisms, and twelve biological processes have been identified as the critical pillars of aging and ARDs (Figure 1).
These hallmarks are intricately linked and interconnected and represent the fundamental changes associated with aging (the roots of aging). As aging advances, it broadly supports the onset of ARDs, including chronic obstructive pulmonary disease (COPD), sarcopenia, diabetes, cancer, frailty syndrome, cardiovascular diseases (CVDs), and neurodegenerative disorders like Alzheimer’s and Parkinson’s diseases. Just as a tree derives nourishment from its roots, these health issues represent the fundamental biological alterations of aging.
The hallmarks of aging appear to be closely interconnected, forming a finely controlled network in which cellular senescence and inflammation represent the “umbrella” that encompasses all these mechanisms [10,11].
In common, all hallmarks are time-dependent on the aging process and can be manipulated by laboratory experiments to accelerate or by therapeutic interventions to slow down the aging process [12]. Therefore, medicine’s primary objective should be to tackle the aging process and enhance the mechanisms that can prevent, delay, or counteract ARDs/GSs [13,14]. An integrated hypothesis proposes that ARDs/GSs manifest an accelerated aging process, indicating that the aging phenotype and ARDs/GSs are not distinct entities but the outcomes of the same common set of molecular and cellular processes, likely occurring at varying rates [15]. Which are the determinants that make the aging trajectories more or less steep? Environmental conditions, such as the intensity and types of stressors and lifestyle, are important health factors. However, the body’s ability to respond to and adapt to these stressors is even more crucial. This capacity is influenced at least partly by an individual’s genetic background and epigenetic changes, which play a significant role in various adaptation and remodeling processes.
Hormesis is a potential mechanism that explains the relationship between healthy aging and the development of ARDs/GSs. Lifelong low-intensity stressors activate maintenance and repair mechanisms that positively affect health. However, increasing the intensity of these stressors can surpass the ability of organs and systems to adapt, resulting in detrimental effects. The emerging concept defines aging as malleable. By targeting the hallmarks of biological aging, such as cell senescence and its interdependent features, it is possible to alleviate ARDs and dysfunctions, thereby extending longevity. Additionally, using external molecules to boost the body’s natural cellular defense mechanisms is proposed as a promising anti-aging strategy centered on hormetic-based protection [16].
A recent multi-omics data study has shown that different organs and tissues can age at distinct rates within the same individual [17]. Brain pathologies and changes in brain structure are commonly seen in aging [18], with significant modifications in the brain’s intricate microstructure resulting in cognitive decline [19]. Brain morphology evolves with age and most commonly undergoes significant atrophy [20]. These changes are flanked, if not directly the cause, by cognitive deficits that include memory loss [21,22], reduced motor performance [23], and behavior [24].
Among neurodegenerative diseases, Parkinson’s disease (PD) and Alzheimer’s disease (AD) are the most common. Usually, they have a late debut of the manifestation with a subsequent stage of progression leading to signs of dementia with similar symptoms, such as memory impairment, orientation problems, and difficulties in performing service functions. In central nervous system (CNS) health, the brain aging trajectory is closely linked to cellular damage accumulation and the onset of neurodegenerative processes. An emerging pivotal factor contributing to the decline of brain structure and function is cellular senescence, a state of stable growth arrest, macromolecular damage, and altered metabolism associated with a hypersecretory and pro-inflammatory phenotype known as the senescence-associated secretory phenotype (SASP). Neuroinflammation may be one of the factors responsible for the increased cognitive decline and risk of AD and PD [25].
This article comprehensively reviews recent advancements concerning the impact of various nutraceuticals and foods on cellular senescence. It delves into key factors associated with this process, such as inflammation, macromolecular damage, mitochondrial dysfunction, and oxidative stress. These factors are critical as they represent common pathways linked to aging and neuronal damage. The review highlights how these dietary components may influence the above-mentioned mechanisms, potentially offering therapeutic avenues to mitigate the effects of aging at the cellular level.
2. The Role of Senescence in Aging and Neurodegenerative Diseases
Senescence is considered a highly dynamic, multi-step process over which the properties of senescent cells continuously evolve and diversify context-dependently [26]. Formally described in 1961 by Hayflick and colleagues, cellular senescence was initially observed in normal human fibroblasts that stopped proliferating after a finite number of divisions [27]. Subsequent studies have proven that a variety of stressors, including oxidative stress, DNA damage, oncogene activation, mitochondria deterioration, chemotherapy and exposition of ionizing radiations, can trigger a “stress-induced premature senescence” in vitro [28,29].
Senescence activation leads to several molecular changes and distinct phenotypic alterations, including chromatin remodeling, shortened telomeres, accumulation of DNA damage and reactive oxygen species (ROS), activation of cell cycle inhibitory pathways, lysosome enlargement, macromolecular disruption, metabolic disbalance, apoptosis resistance and SASP [30]. SASP is characterized by synthetizing various biologically active molecules, such as inflammatory mediators, growth factors and extracellular matrix proteins. These factors reinforce the senescent phenotype through autocrine or paracrine signaling and can also affect the microenvironment, influencing neighboring cells and distant locations within the organism [29] (Figure 2).
As the number of senescent cells increases with age, there is increasing evidence suggesting their involvement in the pathogenesis of ARDs [31,32,33], including neurodegenerative diseases such as AD and PD [34]. Moreover, PD and AD are called “protein misfolding diseases” because deposits of improperly folded and modified proteins are detected in specific areas of the patient’s brain, leading to neuronal damage [35]. It has been reported that the final dysfunction and neuronal loss observed in neurodegenerative diseases are often flanked by malfunctions of other types of CNS cells, such as microglia and astrocytes. Various types of cells in the nervous system have been identified as undergoing the senescence process, including neural stem cells, neurons, astrocytes, oligodendrocytes and microglia. In a state of senescence, microglia are neurotoxic and become detrimental in many neurodegenerative diseases by producing inflammatory cytokines, superoxide anions, and nitric oxide, promoting the phenomenon of ‘oxi-inflamm-aging’, which contributes to the neuropathogenesis [36,37,38].
Evidence shows that senescent astrocytes accumulate in AD and PD patients, promoting inflammation through the SASP factors [39,40,41]. Indeed, several SASP factors, including MMP-3, IL-1α, IL-6, and IL-8, are increased in PD and AD brain, indicating that cellular senescence could contribute to neurodegeneration [42,43,44]. In addition, in the brain tissue of PD patients, α-synuclein deposition correlates with increased senescent cell accumulation and higher SA-β-Gal expression, suggesting the role of cellular senescence in the pathogenesis of PD [45]. Conversely, attenuation or elimination of cellular senescence has been shown to alleviate neuroinflammation in AD and PD models [43,46]. Moreover, a recent study revealed that senescent neurons with tau neuropathology are prevalent in patients with AD [47], while the removal of accumulated senescent glial cells attenuated cognitive decline and age-related neurogenerative disorders [48].
Therefore, eliminating senescent cells within the CNS, or at least delaying their senescence, and mitigating the adverse effects of a spreading SASP have been identified as targets for the prophylaxis and adjunctive treatment of neurodegenerative diseases.
Furthermore, the SASP can be viewed as an ‘inflammatory chain reaction’ that promotes damaging effects and contributes to systemic inflammaging; thus, biomolecules with antioxidants and anti-inflammatory properties would be beneficial not only as protectors against senescence induction but also as tools to extinguish the inflammatory ripple effect [38].
3. Nutritional Interventions to Slow Down Aging
The hallmarks of aging constitute an interconnected network of fundamental mechanisms that influence aging and can be modulated by lifestyle factors, including nutrition, to improve human healthspan [49]. Aging is a malleable process characterized by an intra- and inter-individual heterogeneous and dynamic balance between accumulating damage and repair mechanisms. Nutritional interventions that help slow this process can reduce cellular damage and accumulation of senescent cells or enhance the ability of cells, tissues, or the organism to repair or adapt to that damage [50]. In this context, several natural compounds, known as “bioactive compounds,” can interact with biological processes, and when present in food, they are referred to as “nutraceuticals” [51]. As discussed in the next paragraph, many studies focus on identifying bioactive compounds with preventive effects against pathological conditions or with broader anti-aging properties. Moreover, emerging evidence suggests that dietary factors can influence brain health and cognitive function, providing a promising avenue for intervention [52].
In this context, it is also important to highlight that some nutraceuticals may exhibit hormetic behavior, displaying a biphasic dose-response relationship in which low doses provide beneficial effects, whereas high doses may be detrimental. These positive effects at low concentrations arise from stimulating adaptive stress responses, ultimately enhancing the body’s resilience to various stressors. This phenomenon is particularly evident among polyphenols, such as curcumin and resveratrol [53].
Recently, the focus has been on identifying nutraceuticals that mimic metformin and rapamycin without their collateral effects. Among these, researchers have individuated withaferin A, allantoin, ginsenoside, and epigallocatechin gallate as promising candidates for experimental validation [54,55]. They showed strong activation of the cAMP pathway, which was recently found to induce anti-aging effects similar to caloric restriction (CR) via up-regulation of sirtuins (SIRTs) [56]. SIRTs, particularly SIRT1 and SIRT3, are key regulators of cellular metabolism, stress responses, and aging. As NAD+-dependent deacetylases, they are activated under CR, promoting longevity and healthspan by modulating energy metabolism, mitochondrial function, and stress resistance. SIRT1 acts as a nutrient sensor, regulating epigenetic modifications, mitochondrial quality, and anti-inflammatory responses, while SIRT3 enhances mitochondrial protein deacetylation, optimizing oxidative metabolism and aerobic fitness, both contributing to the lifespan-extending effects of CR [57,58,59].
CR consists of a 25–50% calorie reduction compared to a standard diet, with preservation of the vitamins and minerals supply [60]. In addition to SIRTs, CR modulates other key nutrient signaling pathways, including insulin/IGF-1, mTOR, and AMPK, leading to a reduction in oxidative stress, enhancement of mitochondrial function, activation of anti-inflammatory responses, stimulation of neurogenesis, and increased synaptic plasticity, emphasizing the positive impact of CR on brain functions. These effects can delay cellular senescence and may significantly mitigate age-related functional decline [61]. Experimental studies have reported that CR reduces molecular features of cellular senescence in different models [62,63,64]. Interestingly, a recent study demonstrated that moderate CR could decrease circulating biomarkers of cellular senescence in healthy young to middle-aged humans without obesity, highlighting the impact of lifestyle [65]. Moreover, dietary restriction and plant-based dietary patterns have been linked to improved key clinical outcomes associated with aging, especially body composition changes, lipid profile, blood pressure, lipid peroxidation, inflammation, and cardiometabolic risk [66,67,68,69,70,71,72,73]. Despite the mechanisms not being fully elucidated, these benefits suggest that such dietary approaches may be crucial in promoting healthy aging by modulating metabolic and inflammatory pathways central to age-related physiological changes and disease prevention. However, in CR, the timing of initiation is a critical factor; when started at an early age, it is associated with beneficial effects [74]. Conversely, in older adults, CR may exacerbate sarcopenia and osteopenia, contributing to muscle and bone loss [75].
In addition to CR, other dietary patterns have been proposed to promote healthy aging. Among these, the Mediterranean diet (MedDiet) is the most studied. MedDiet is characterized by a high intake of vegetables, fruits, whole grains, and fish, and it has demonstrated significant health benefits, including the prevention of ARDs. Its protective effects are mainly attributed to its rich composition of bioactive compounds that help modulate oxidative stress, inflammation, and metabolic processes, further supporting its role in longevity and overall well-being [76,77,78,79,80] and reducing cognitive impairment [81]. Intriguingly, emerging proofs suggest that adherence to the MedDiet may contribute to delaying cellular senescence [82]. In older adults, adherence to MedDiet has been associated with a lower proportion of endothelial cells with shorter telomeres, an effect likely mediated by decreased ROS production and apoptosis [83]. Furthermore, Mantilla-Escalante suggests that long-term adherence (1 year) to the MedDiet, particularly when enriched with nuts, can modulate the expression of several microRNAs (miRNAs) involved in cellular senescence, including cell cycle regulators and pro-inflammatory markers. MedDiet, through miRNA-mediated gene modulation, may influence fundamental mechanisms of aging and cellular homeostasis [84].
Even if the mechanisms through which food influences aging are not fully understood, several bioactive compounds have been reported to function as epigenetic modulators, influencing gene expression, chromatin organization, DNA methylation patterns, and non-coding RNA expression [85,86]. Interestingly, the human epigenome is influenced by exogenous factors such as nutrition, a field explored in nutritional genomics. Both the quality and quantity of diet have been found to modulate DNA methylation and mental health epigenetically [87].
Additionally, an intriguing hypothesis suggests that bioactive compounds in food may extend healthspan by modulating SASP, indicating new strategies to slow the onset and progression of ARDs [88]. Since the anti-aging effects of natural compounds have only recently begun to be scientifically evaluated, very few notions are available about their properties and ability to exert anti-SASP and/or senolytic activity. However, nutrition is often considered one of the most promising modifiable risk factors for ARDs, including neurodegenerative diseases, a contention fully appreciated in multidomain intervention studies [89,90,91].
While all-natural foods are inherently functional due to their composition, the concept of functional foods emerged from the observation that certain manufactured foods, enhanced with additional ingredients, can further improve human health [92]. This category includes conventional foods enriched with bioactive compounds such as vitamins, minerals, and phytochemicals [94,95], directly impacting nutritional health by enhancing overall well-being or reducing disease risk [93]. Among the various nutraceutical-enriched foods, olive oil, milk, and yoghurt stand out for their potential health benefits. Extra virgin olive oil has been extensively studied for its positive effects on telomere length, diabetes, cognitive function, and various hallmarks of aging, including cellular senescence [94]. Similarly, the impact of fortified milk on inflammaging parameters has been investigated by Martucci et al. in 2020. The fortified milk was enriched with omega-3 fatty acids (EPA, DHA), various vitamins, and trace elements, finding improved levels of micronutrients and the omega-3 index, along with reduced arachidonic acid (AA), homocysteine and omega-6/omega-3 ratio [95]. Yoghurt, rich in anti-inflammatory and B-vitamin content, may help protect against cognitive decline, as shown by Tillisch et al., who reported brain function changes linked to fermented dairy intake [96].
Functional foods play a crucial role in healthy aging by addressing factors like oxidative stress, inflammation, and mitochondrial dysfunction. Their positive effects on aging mechanisms suggest potential benefits for aged people [97]. By modulating many biological mechanisms in mammalian body and cells, it could be proposed the following anti-aging mechanisms of functional foods: i) Stabilizers of mitochondrial membranes and enhancers of mitochondrial function, agents that avoid cell death by apoptosis or necrosis, ii) Metal chelating activities, iii) Antioxidants, iv) Inducers of apoptosis of preneoplastic and neoplastic cells [98,99,100,101].
The distinction between nutraceuticals and functional foods is often blurred due to their intrinsic overlap, as nutraceuticals represent a specific subset of functional foods. Given this complexity, our review will specifically focus on nutraceuticals to provide a more structured and comprehensive analysis of their role in neurodegenerative diseases, specifically AD and PD. By narrowing our scope, we aim to offer a clearer perspective on their mechanisms of action and potential therapeutic applications.
4. Nutraceutical Interventions in Neurodegenerative Disorders: Focus on Parkinson’s and Alzheimer’s Diseases
The aging brain is highly susceptible to neurodegenerative diseases, but the exact mechanisms through which senescence in the CNS contributes to neuropathogenesis remain unclear. The number of senescent cells increases with age, and there is growing evidence suggesting the involvement of cellular senescence in the neuropathogenesis of AD and PD, resulting in a significant increase in chronic neuroinflammation due to the SASP. Therefore, countering and removing senescent cells in the brain, or at least postponing their senescence and alleviating the adverse effects of spreading SASP, could be a strategy for helping to slow the progression of AD and PD or delaying their onset.
This paragraph reviews studies investigating nutraceutical compounds that may mitigate cellular senescence processes in the brain, including neuroinflammation and the reduced expression of anti-apoptotic proteins such as Bcl-2 and Bcl-xl, as well as demonstrating senostatic and senolytic effects. Although direct evidence linking nutraceuticals to cellular senescence in neurodegenerative diseases is currently limited, this field has considerable potential. Various nutraceuticals have shown beneficial effects across numerous models by modulating traits associated with senescence, indicating that further research may provide valuable insights into their advantages.
Given the established role of senescent cells in neurodegenerative diseases, we suggest that nutraceutical compounds affecting senescence-associated features may yield beneficial outcomes in these conditions. However, due to the lack of direct evidence, our discussion will primarily focus on key molecular and cellular mechanisms related to senescence rather than directly indicating their effects on senescent cells in neurodegeneration.
The compounds discussed are categorized based on their natural origins.
4.1. Antioxidant Vitamins
A major contributor to aging and ARDs, such as AD [102] and PD [103] is oxidative stress induced by free radicals. ROS, natural byproducts of cellular metabolism increased in senescent cells, play dual roles in biological systems. At low to moderate concentrations, ROS support essential physiological processes; however, when their levels become excessive, oxidative stress ensues, leading to damage to critical cellular components, including lipids, proteins, and DNA and also can induce cell senescence [104,105,106]. This oxidative stress can directly activate glial cells, mainly by priming astrocytes, resulting in their interaction with neurons and subsequent release of immune mediators such as nitric oxide (NO), additional ROS, pro-inflammatory cytokines, and chemokines. These mediators act as neurotoxins, propagating inflammation within the CNS [107].
Accumulating evidence from mouse models of accelerated senescence indicates that ascorbic acid (AAC) plays a rescuing role in premature aging. Moreover, although the precise role of AAC in the CNS remains only partially understood, studies have demonstrated that its concentration in the cerebrospinal fluid (200–400 mM) far exceeds that found in cerebral parenchyma and plasma (30–60 nM) [108]. Overall, AAC exhibits notable nootropic properties [109] and has been shown to decrease acetylcholinesterase activity in mice [110]. In addition, it facilitates the differentiation of neuronal and astrocyte precursors, thereby promoting synaptic maturation [111]. AAC is also essential for the biosynthesis of catecholamines, peptide amination, myelin formation, and enhancement of synaptic function while providing neuroprotection against glutamate toxicity [112,113].
In PD, dopamine metabolism generates oxidative stress products that contribute to accumulating abnormal proteins that are characteristic of PD [114]. Current therapeutic strategies for PD primarily alleviate symptoms, but they do not halt disease progression, rendering treatment particularly challenging.
Although early studies indicated that AAC supplementation could mitigate oxidative damage in vitro and animal models [115,116], more recent investigations have yielded inconsistent results [117]. Notably, AAC levels are lower in the substantia nigra compared to other brain regions [118,119], heightening its vulnerability to oxidative stress [120]. Furthermore, AAC has been shown to enhance the production of dihydroxyphenylalanine (DOPA); Seitz et al. observed a dose-dependent overproduction of DOPA in the human neuroblastoma cell line SK-N-SH following incubation with AAC (100–500 mM) for 2 hours [121]. Nonetheless, AAC has been demonstrated to improve the absorption of levodopa in elderly PD patients with poor levodopa bioavailability, thereby enhancing its therapeutic efficacy and reducing its side effects [122,123].
Moreover, AAC is critical for brain development; one study reported that AAC treatment induced a tenfold increase in dopaminergic differentiation in CNS precursor cells derived from the E12 rat mesencephalon [124]. In vivo, a cohort study of 1,036 PD patients further supported the neuroprotective role of AAC, demonstrating that higher dietary intake was significantly associated with a reduced risk of PD [125], although some studies have not corroborated these findings [126,127].
In contrast, the neuroprotective effects of vitamin E are thought to arise from its ability to prevent oxidative stress and inhibit apoptosis. Vitamin E has been shown to reverse impaired synaptic plasticity in mouse models [128] and reduce ROS levels in Drosophila models [129]. Additional evidence underscoring the role of oxidative stress in PD includes observations that cellular antioxidants such as glutathione (GSH) are depleted in PD [130] and that treatment with antioxidants, including vitamin E, can protect mice and cultured cells from 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced neurotoxicity [131].
Specific isoforms of vitamin E, such as γ-tocotrienol and δ-tocotrienol, exert neuroprotective effects through the ERβ-PI3K/Akt signaling pathways in SH-SY5Y cells [132]. Moreover, δ-tocotrienol has been found to prevent dopaminergic neuron loss and improve motor function in mouse models of PD; its neuroprotective effect, however, was attenuated by ER inhibitors [133]. In an MPTP-induced PD model in C57/B1 mice, vitamin E-deficient animals were markedly more susceptible to MPTP toxicity, exhibiting increased lethality and greater depletion of dopamine metabolites in the substantia nigra [134]. Perry et al. [135] similarly reported that mice pretreated with daily subcutaneous injections of high-dose α-tocopherol (αT) (2350 mg/kg body weight) for 48 hours before and 72 hours after MPTP administration experienced partial protection against the loss of striatal dopamine and dopaminergic neurons in the substantia nigra. In supporting these experimental findings, a cross-sectional study involving participants over 40 years of age found that higher vitamin E intake was significantly associated with a reduced risk of PD [136].
Conversely, L-AAC has also garnered attention for its beneficial effects on AD [137]. The primary neuroprotective mechanisms attributed to AAC include ROS-scavenging activity, neuroinflammation modulation, Aβ fibrillation inhibition, and chelation of metals such as iron, copper, and zinc [138]. Furthermore, AAC has been shown to protect SH-SY5Y neuroblastoma cells from Aβ-mediated apoptosis [139] and, when administered orally, to reduce oxidative stress and neuroinflammation induced by Aβ fibrils in rats [140].
In contrast, vitamin E is a potent antioxidant that scavenges free radicals primarily through a hydrogen atom transfer mechanism. This reaction yields a non-radical product and a vitamin E radical, which can subsequently react with another radical to form a stable compound, interact with lipids, or be regenerated by reducing agents such as AAC or ubiquinol [141]. Vitamin E plays a crucial role in the brain, is one of the most potent antioxidants, and has shown significant benefits in AD [142]. It counteracts Aβ-induced oxidative stress [143]; for instance, vitamin E has been demonstrated to prevent Aβ₁–₄₂-induced protein oxidation, ROS production, and neurotoxicity in primary rat embryonic hippocampal neuronal cultures [144]. Moreover, although Aβ₁–₄₂ reduces the surface expression of the principal glutamate transporter, GLT-1, in adult mouse astrocytes, this detrimental effect is prevented by a water-soluble analogue of vitamin E [145]. Vitamin E also helps preserve calcium homeostasis, protects against damage from Aβ deposits near cell membranes [146], and prevents oxidative stress-induced Tau hyperphosphorylation [147,148]. Additionally, it can inhibit neuroinflammation by suppressing the production of prostaglandins E₂ and D₂, along with reducing cyclooxygenase and lipoxygenase activity [149,150].
Limited evidence directly links the triad of cellular senescence, neurodegenerative diseases, and antioxidant vitamins. Most studies have focused on other cell types, with only a few providing insight into their potential role in modulating senescence-related pathways. To offer a broader perspective, we highlight some relevant findings. For instance, Jeong et al. reported that AAC significantly reduced SA-β-Gal activity by suppressing cell-cycle inhibitors (p53, p21, p16, and pRb) while stimulating cell-cycle activators (E2F1 and E2F2). This finding demonstrated its ability to prevent cellular senescence in vitro and in vivo using human dermal fibroblasts and hairless mice models [151]. Similarly, another study showed that co-incubation with AAC significantly downregulated SA-β-Gal staining, reduced senescence-associated gene expression, and improved cell migration, mineralized nodule formation, and the expression of dentine sialophosphoprotein (DSPP) and bone sialoprotein (BSP) genes in LPS-treated human apical papilla cells [152]. Limited studies are also available regarding vitamin E supplementation. In particular, vitamin E has been shown to reduce the number of SA-β-Gal positive cells in vitro in human umbilical vein endothelial cells and human fibroblasts [153]. One study revealed that vitamin E ability to decrease SA-β-Gal activity in human fibroblasts depends on the donor’s age and cell passage number.
Specifically, while vitamin E reduces SA-β-Gal levels in cells from both young and aged donors at replicative senescence in fibroblasts from older subjects, this effect occurs even at earlier passages. This may be attributed to the phosphorylated form of vitamin E, α-tocopheryl phosphate (αTP), which accumulates with aging due to a reduced conversion to αT. Notably, αTP is more effective than αT in decreasing CD36 expression and CD36-mediated senescence [154].
To conclude, AAC exerts neuroprotective effects by scavenging ROS, modulating neuroinflammation, and supporting synaptic function, while vitamin E mitigates oxidative stress, preserves membrane integrity, and inhibits apoptosis. Although studies indicate potential positive outcomes on senescence-associated characteristics, findings remain inconsistent in AD and PD, highlighting the need for further research to elucidate their precise mechanisms and therapeutic potential.
4.2. Polyphenols
Dietary polyphenols exhibit robust neuroprotective effects that extend well beyond their well-known antioxidant and anti-inflammatory properties. Circumstantial evidence indicates that these compounds modulate intracellular signaling pathways, alter gene expression, and influence enzyme activities, all contributing to their therapeutic potential in neurodegenerative diseases [155,156].
A growing body of research demonstrates that polyphenols can modulate cellular senescence. For example, chronic treatment of pre-senescent neonatal human dermal fibroblasts with oleuropein aglycone, a prominent polyphenol in extra-virgin olive oil, resulted in a significant reduction in senescent cell numbers, as evidenced by decreased SA-β-Gal activity and lower p16 protein expression [157]. Similarly, compounds such as apigenin, quercetin, kaempferol, and wogonin have been shown to suppress the secretion of SASP markers, including IL-6, IL-8, and IL-1β [158]. Recent studies by Bientinesi et al. revealed that quercetin can prevent doxorubicin-induced senescence [159,160]. Quercetin not only alleviates the deleterious effects of the SASP in both U2OS and normal cells but also protects fibroblasts from ROS-mediated damage, evidenced by reductions in SAHF, Lamin B1 loss, and NF-κB nuclear translocation. Moreover, quercetin exhibits senolytic activity, reducing autophagy while increasing endoplasmic reticulum stress, thereby underscoring its multifaceted role in combating cellular aging. Several benefits have also been demonstrated in human in vivo studies. For instance, Maurya et al. showed that in human red blood cells, these flavonoids reduce malondialdehyde (MDA) levels while increasing GSH and membrane sulfhydryl (-SH) group levels [161].
In the context of PD, dietary polyphenols appear to have beneficial effects. Flavonoids, a major subgroup of polyphenols, protect neurons against oxidative stress, suppress neuroinflammation, and modulate key intracellular signaling pathways critical for neuronal survival. These pathways, including protein kinase and lipid kinase signaling cascades, alter the phosphorylation state of target proteins and influence gene expression [162].
Moreover, histochemical evaluations in 6-OHDA-treated mouse models of PD have shown that green tea (a variant of tea obtained with non-treated leaves of Camellia sinensis) polyphenols markedly reduce ROS levels, lipid peroxidation, and intracellular nitrite/nitrate concentrations [163,164].
Ginkgo biloba extract has been reported to protect against nigrostriatal dopaminergic neurotoxicity in MPTP-induced PD models, with observed reductions in lipid peroxidation and enhancements in the activities of key antioxidant enzymes such as SOD, GPx, and GSH reductase [165]. Notably, Ginkgo biloba extract inhibited monoamine oxidase B (MAO-B) in vitro, reducing dopaminergic neuron degeneration [165,166].
Resveratrol, a nonflavonoid polyphenol found in grapes and berries, has shown promise in mitigating oxidative stress in PD [167,168] while enhancing the number of dopaminergic neurons at the synapses through MAO suppression as well as the prevention of glutamate release [169,170,171].
Additionally, oleuropein and its derivatives have been demonstrated to inhibit ROS accumulation and prevent PD pathology. In vitro, oleuropein aglycone stabilizes α-synuclein monomers, thereby preventing pathological aggregation [172], and it also inhibits α-synuclein fibril elongation, reducing the cytotoxic effects of α-synuclein oligomers [173]. Furthermore, oleuropein activates redox-sensitive transcription factors such as Nrf2, which may enhance the intracellular antioxidant capacity and contribute to neuroprotection [174].
Beyond PD, dietary polyphenols have been shown to have several benefits in AD, mitigating pathological manifestations partly due to their ability to cross the blood-brain barrier [175,176]. Oxidative stress plays a pivotal role in the initiation and progression of Alzheimer’s, and while the body has evolved defence mechanisms to counteract oxidative damage, aging is associated with increased oxidative stress that disrupts cellular architecture [177]. Polyphenols reinforce endogenous antioxidant defences and attenuate protein oxidation [178]. By sequestering reactive oxygen and nitrogen species, these compounds prevent the formation of toxic Aβ oligomers and modulate tau-protein hyperphosphorylation, thereby impeding the development of neurofibrillary tangles (NFTs) [179]. Additionally, polyphenols may help preserve neuronal integrity by interacting with transcription factors such as CREB and NF-κB [180].
Studies on AD transgenic mouse models (APP/PS1 model) and patients’ post-mortem brains have revealed a senescent phenotype in oligodendrocyte progenitor cells (OPCs) within the Aβ plaque environment. Notably, these cells were sensitive to clearance by the senolytic cocktail dasatinib plus quercetin (D+Q). The treatment removed senescent OPCs and ameliorated Aβ plaque-associated inflammation and cognitive deficits in AD mice. [181]. Meanwhile, in PD, direct evidence of the beneficial effects of D+Q has not been observed, even though some advantages have been shown in the aging killifish [182]. Additionally, fisetin, a natural senolytic, has been shown to improve cognitive function in mouse models of AD and dementia [183]. Among the senolytics tested in multiple preclinical studies and increasing clinical trials, fisetin and D+Q appear to be the most effective [184,185].
Animal studies further substantiate the neuroprotective potential of polyphenols. For instance, mice receiving pomegranate juice exhibited significant improvements in both cued and spatial learning tasks, along with reduced hippocampal plaque loads—including both soluble and fibrillar forms of Aβ, as well as lower soluble Aβ1-42 levels [186]. Red wine polyphenols have been shown to interfere with Aβ oligomerization, thereby attenuating Aβ neuropathology and cognitive decline in Tg2576 mice [187]. Mori et al. [188] demonstrated that tannic acid shifts amyloid precursor protein metabolism toward a non-amyloidogenic pathway by lowering β-secretase 1 (BACE1) expression and β-secretase activity, decreasing Aβ peptide levels. Similarly, grape-derived polyphenolics from Vitis vinifera grape seeds significantly inhibited Aβ aggregation in vitro and ameliorated cognitive deterioration in Tg2576 mice when administered orally [189].
Collectively, these findings illustrate the multifaceted neuroprotective potential of dietary polyphenols. By modulating intracellular signaling pathways, gene expression, and enzyme activities, polyphenols offer promising therapeutic avenues for preventing and treating neurodegenerative diseases, highlighting their potential as valuable agents in mitigating age-related cognitive decline and neuronal dysfunction.
4.3. Spices
Over the past decade, numerous studies have underscored various spices’ broad spectrum of anti-aging and anti-senescence properties. For instance, the primary bioactive compounds of black pepper, including piperine, chavicine, and sabinene, exhibit significant pharmacological potential. Notably, black pepper oil has reduced the percentage of doxorubicin-induced senescent cells in CHO-K1 and NIH-3T3 cells [190]. Furthermore, curcumin, the primary component of Curcuma longa, has demonstrated a capacity to mitigate age-related deterioration by counteracting oxidative stress [191], modulating inflammatory pathways [192,193], promoting telomere elongation and telomerase activity [194], and influencing key metabolic regulators such as AMPK [195,196] and SIRTs [197,198]. Similarly, coriander seeds have shown potential in reducing oxidative stress and cellular senescence, as evidenced by decreased expression of senescence markers SA-β-Gal and p21 in cardiac [199] and brain tissues [200] of obese rats.
Beyond their culinary roles, spices have emerged as promising agents for preventing or even counteracting neurodegenerative processes associated with aging. The neuroprotective effects of spices show promising therapeutic potential in PD as well. Curcumin has exhibited multiple protective mechanisms in PD, facilitated by its ability to cross the blood-brain barrier due to its lipophilic nature [201]. Its neuroprotective effects are attributed mainly to its potent antioxidant properties, surpassing conventional antioxidants such as vitamins C and E [202,203]. The ability of curcumin to donate hydrogen ions from its β-diketone moiety is believed to underlie its anti-ROS activity [204]. Notably, pre- or post-treatment with curcumin in 6-OHDA-lesioned rats reduced dopaminergic neuron loss [205], while MES cells treated with curcumin exhibited increased Cu-Zn superoxide dismutase expression and reduced intracellular ROS accumulation [206]. Moreover, curcumin was found to modulate inflammatory processes by decreasing the production of prostaglandins, glutamate, and pro-inflammatory cytokines in the hypothalamus, as well as reducing GFAP levels, a marker of astrocytic proliferation [207].
Similarly, piperine from black pepper has demonstrated neuroprotective effects in PD models. Yang et al. reported that piperine administration ameliorated MPTP-induced motor and cognitive deficits while preventing the loss of tyrosine hydroxylase-positive neurons in the substantia nigra [208]. Additionally, piperine reduced microglial activation, IL-1β expression, and oxidative stress and exhibited anti-apoptotic properties by modulating the Bcl-2/Bax ratio. Interestingly, piperine has been evaluated in combination with quercetin due to its well-documented ability to enhance the bioavailability of other compounds [209]. Combining quercetin and piperine improved MPTP-induced behavioural and neurochemical deficits while mitigating oxidative stress and inflammation in the striatum [210].
Emerging in vitro evidence further supports the beneficial role of cinnamon and its metabolites in PD. Cinnamaldehyde (10 μM) was shown to protect BE(2)-M17 human neuroblastoma cells from MPP+-induced toxicity by inhibiting autophagy [211]. Cinnamon extract (CEppt) has also shown protective effects against 6-OHDA-induced cytotoxicity by enhancing cell viability, reducing apoptosis, and decreasing ROS levels [212]. Furthermore, CEppt has been found to interfere with α-synuclein aggregation by stabilizing its soluble oligomeric form and promoting the disassembly of preformed aggregates [213].
In addition to their anti-inflammatory properties, these bioactive compounds exert antioxidative effects and inhibit acetylcholinesterase activity and Aβ aggregation in AD. Curcumin has demonstrated potent antioxidant activity in both in vitro and in vivo models [214,215,216]. Epidemiological studies have further revealed an inverse correlation between curcumin intake and AD prevalence, particularly in populations with high dietary consumption of this compound, such as India [217,218]. Mechanistically, curcumin enhances macrophage-mediated clearance of Aβ plaques [219], suppresses microglial proliferation [220], attenuates neuroinflammation by downregulating pro-inflammatory cytokines [221,222], and inhibits oxidative stress by preventing free radical formation and propagation [223,224]. Remarkably, in vitro studies suggest that curcumin reduces Aβ levels by modulating APP processing and downregulating BACE1 expression [225]. Additionally, curcumin exhibits a high binding affinity for Aβ aggregates, thereby preventing their formation both in vitro and in vivo [226].
Beyond curcumin, cinnamon has also demonstrated significant neuroprotective properties. Studies have shown that cinnamon exerts potent antioxidant effects by enhancing the activity of endogenous antioxidant enzymes [227,228] while simultaneously exerting anti-inflammatory effects [229,230]. Notably, a CEppt effectively inhibits the formation of toxic Aβ oligomers and protects neuronal PC12 cells from Aβ-induced toxicity, eliminating tetrameric Aβ species in the brain. Moreover, oral administration of this extract in an aggressive AD transgenic mouse model significantly reduced 6 kDa Aβ oligomers, diminished plaque burden, and improved cognitive performance [231].
Piperine, the principal bioactive component of Piper nigrum (black pepper), has been reported to exert neuroprotective effects [232]. In animal models, black pepper administration reduced cholinesterase activity and amyloid plaque formation in the brain [233]. Furthermore, piperine significantly attenuated lipid peroxidation and acetylcholinesterase activity while preserving neuronal density in adult male Wistar rats [232].
Similarly, cardamom oil treatment improved neurobehavioral parameters in male Wistar rats, inhibited acetylcholinesterase activity in the hippocampus and cortex, and enhanced antioxidant enzyme levels while reducing oxidative damage. Additionally, it increased BDNF levels and suppressed Aβ expression in the hippocampus and cortex [234].
Overall, accumulating evidence suggests that spices such as curcumin, cinnamon, black pepper, and cardamom possess significant neuroprotective properties that may be exploited to prevent and treat neurodegenerative diseases, including AD and PD.
4.4. Dietary Fiber
Recent findings suggest that a high-fiber diet may protect against neurodegenerative disorders by supporting metabolism, modulating neuroinflammation, and regulating epigenetics. Unfortunately, although it displays several effects on key mechanisms of cellular senescence, a direct link to senescence itself remains unclear. Dietary fiber, composed of non-digestible and non-absorbable carbohydrates, influences gut microbiota composition and short-chain fatty acids (SCFAs) production, impacting brain function through the microbiota-gut-brain axis [235,236].
Shi et al. studied dietary fiber deficiency (FD) in mice, revealing alterations in hippocampal synaptic ultrastructure, proteome, and microglial-neuroinflammation pathways, affecting cognition and dopamine cholinergic synapses [235].
Gut microbiome alterations have also been linked to PD, with decreased SCFAs, particularly butyrate, observed in PD patients [237]. Similarly, Matt et al. showed that Butyrate administration and high-fiber diets reduced neuroinflammation in aged mice [238].
4.5. Probiotics and Prebiotics
Despite strong evidence supporting the role of probiotics and prebiotics in neuroinflammation and neurodegeneration, their direct impact on cellular senescence remains unclear. However, they have been shown to exert regulatory effects on oxidative stress, inflammation, and metabolism. Notably, they may modulate neuroinflammation, which is at least partially influenced by the presence of senescent cells. Further studies are needed to clarify this relationship and explore their potential in aging and neurodegenerative diseases.
Probiotics and prebiotics influence human health by modulating metabolic regulation, immune response, and neurological function via the gut microbiome [245,246,247,248,249]. Probiotics, particularly Lactobacilli species, have demonstrated benefits in aging by enhancing immunity and maintaining gut microbiota balance, potentially extending the healthspan [250,251,252,253]. Probiotics can regulate neuroinflammation and oxidative stress in neurodegenerative diseases, reducing the risk of disorders like PD and AD [254,255,256,257].
In PD, probiotics improve gut health, mitigate permeability, and reduce neuroinflammation [258]. Cellular studies show that probiotic treatment can shift cytokine production towards an anti-inflammatory profile [259], while in vivo studies suggest protection against dopaminergic neuron loss and neurotrophic factor depletion [260,261,262]. Similarly, specific probiotic strains can restore gut microbiota balance in AD, improve cognitive function, and mitigate pathological features such as Aβ deposition and oxidative stress [263,264]. However, the precise mechanisms remain unclear [265,266], and probiotics’ role as modulators of cellular senescence per se is only beginning to be understood.
Conversely, prebiotics, including fructo- and galacto-oligosaccharides, promote SCFA production and support cognitive function [267]. They modulate oxidative damage, enhance GLP-1 (glucagon-like peptide 1) secretion, and potentially regulate brain glucose metabolism and CNS inflammation via GLP-1 receptors [268,269]. Prebiotics alter gut microbiota composition in PD, reducing pro-inflammatory bacteria and increasing SCFA-producing taxa, with promising neuroprotective effects [270,271]. Intriguingly, their combination with probiotics appears particularly beneficial [272,273].
On the other hand, prebiotic supplementation in AD models has shown improvements in neurotransmitter levels, cognitive function, and Aβ pathology, with chitosan oligosaccharides demonstrating neuroprotective properties in several studies [274].
4.6. Polyunsaturated Fatty Acids (PUFAs)
PUFAs are crucial in neuroprotection, presenting potential therapeutic implications for neurodegenerative diseases. Evidence supports their involvement in modulating inflammation, oxidative stress, and neurotoxicity. However, further research is required to investigate their direct action as senolytic or senostatic agents and their influence on the broader aging process [275]. PUFAs play a fundamental role in neurodevelopment, neurotransmission, and neuromodulation, with potential neuroprotective effects that include reducing neuroinflammation, mitigating neurotoxicity, promoting neural recovery, and preserving blood-brain barrier integrity [276].
Among PUFAs, omega-3 (n-3) and omega-6 (n-6) long-chain polyunsaturated fatty acids (LCPUFAs) are essential for brain function, constituting 30–35% of total brain fatty acids. Docosahexaenoic acid (DHA) and AA are the predominant LCPUFAs in phospholipids, playing key roles in synaptic integrity, plasticity, and cognitive function [277,278]. Neuroinflammation is a major contributor to age-related neurodegeneration, and n-3 LCPUFAs, particularly eicosapentaenoic acid (EPA) and DHA, exhibit anti-inflammatory properties by downregulating IL-6 and TNF-α while enhancing cognitive function [279]. Interestingly, higher brain DHA concentrations correlate with improved cognitive performance, likely due to its effects on membrane fluidity, neurotransmitter release, gene expression, neuroinflammation, and neuronal growth [280,281]. These fatty acids also possess antioxidant and anti-apoptotic effects, mitigating neurotoxicity in preclinical models [282,283].
Research on dietary fats is still emerging in PD, but observational studies suggest that LCPUFA intake may offer neuroprotection [284,285]. For example, a meta-analysis reported that higher LCPUFA consumption is associated with a reduced PD risk, while specific plasma fatty acid levels correlate with motor and non-motor symptoms [286]. Additionally, n-3 PUFAs have demonstrated neuroprotective effects in MPTP-induced PD mice models, preventing neuronal loss and preserving striatal dopamine levels [283,287,288,289,290].
In AD, epidemiological studies and randomized controlled trials (RCTs) indicate that higher n-3 LCPUFA intake correlates with a lower incidence of cognitive impairment and dementia [281,291]. Other RCTs in individuals with mild to moderate AD have reported cognitive improvements following supplementation [292,293]. Animal studies suggest DHA reduces amyloid accumulation, tau pathology, and synaptic dysfunction, with several independent reports confirming reduced Aβ levels in APP transgenic models following DHA-enriched diets [294,295,296,297,298].
5. Conclusions
The review highlights the important role of nutraceuticals and functional foods in reducing aging and neurodegenerative diseases by modulating cellular senescence. It discusses how these natural bioactive compounds possess potent antioxidant, anti-inflammatory, and epigenetic properties that can impact essential cellular pathways associated with aging and the onset of neurodegenerative diseases. We specifically emphasize the importance of polyphenols, vitamins, and spices as nutritional senotherapeutic agents in scavenging ROS, reducing the secretion of inflammatory SASP factors and modulating gene expression alongside other characteristics related to cell senescence (Figure 3).
Collectively, these actions contribute to alleviating cellular damage that is involved in both aging and the onset of neurodegenerative disorders such as AD and PD diseases.
Furthermore, the review highlights that substantial evidence supports the beneficial effects of these nutraceuticals on cellular processes, such as improving mitochondrial function, reducing oxidative stress, and modulating inflammatory responses, direct evidence demonstrating a senolytic effect is still limited.
Most existing studies have primarily focused on elucidating the mechanisms through which these compounds influence senescence-associated characteristics rather than proving a direct reduction of the number of senescent cells. This emerging and relatively new field requires further research to explore these correlations in more detail and to understand the potential benefits of introducing nutraceuticals into preventive strategies. Such interventions may offer a promising approach to extending healthspan by targeting the underlying mechanisms of cellular senescence, although current research is still in its infancy. Ultimately, the review suggests that including nutraceuticals in comprehensive dietary interventions may help lower the risk of neurodegenerative diseases. However, the scarcity of clinical data raises questions about their effectiveness, especially considering the emerging hormetic properties of certain nutraceuticals.
Numerous questions remain unresolved regarding the application of nutraceuticals as senotherapeutics, but there exists a pressing necessity to identify anti-aging strategies that promote active longevity while minimizing disability and functional dependence.
Author Contributions
Conceptualization, S.R., G.B., E.B. and D.M.; writing—original draft preparation, S.R., G.B.; writing—review and editing, E.B. and D.M, supervision and funding acquisition: D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by co-funding from the European Union – Next Generation EU, in the context of The National Recovery and Resilience Plan, Investment Partenariato Esteso PE8 “Conseguenze e sfide dell’invecchiamento”, Project Age-IT, CUP: B83C22004800006, to D.M.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We acknowledge co-funding from Next Generation EU, in the context of the National Recovery and Resilience Plan, Investment PE8 – Project Age-It: “Ageing Well in an Ageing Society”. This resource was co-financed by the Next Generation EU [DM 1557 11.10.2022]. The views and opinions expressed are only those of the authors and do not necessarily reflect those of the European Union or the European Commission. Neither the European Union nor the European Commission can be held responsible for them.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| αT | α-tocopherol |
| αTP | α-tocopheryl phosphate |
| Aβ | Amyloid-β peptide |
| AA | Arachidonic acid |
| AAC | Ascorbic acid |
| AD | Alzheimer’s Disease |
| ARDs | Age-related diseases |
| BACE1 | β-secretase 1 |
| CEppt | Cinnamon extract |
| CNS | Central Nervous System |
| CR | Caloric Restriction |
| D+Q | Dasatinib plus Quercetin |
| DHA | Docosahexaenoic Acid |
| DOPA | Dihydroxyphenylalanine |
| EPA | Eicosapentaenoic Acid |
| FD | Fiber deficiency |
| GLP-1 | Glucagon-like peptide 1 |
| GSH | Glutathione |
| GSs | Geriatric Syndromes |
| IL | Interleukin |
| LCPUFAs | Long-chain polyunsaturated fatty acids |
| MAO-B | Monoamine Oxidase B |
| MDA | Malondialdehyde |
| MedDiet | Mediterranean Diet |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| NFTs | Neurofibrillary tangles |
| PD | Parkinson’s Disease |
| PUFAs | Polyunsaturated Fatty Acids |
| ROS | Reactive Oxygen Species |
| SASP | Senescence-Associated Secretory Phenotype |
| SA-β-gal | Senescence-Associated β-Galactosidase |
| SAHFs | Senescence-associated heterochromatic foci |
| SCFAs | Short-Chain Fatty Acids |
| SIRT | Sirtuin |
References
- WHO World Population Prospects 2022; 2022; ISBN 978-92-1 -148373-4.
- Santana, P.; Grant, M. Global Aging and Health Determinants in a Changing World; INC, 2023; ISBN 9780128237618.
- World Health Organisation World Population Ageing 2019; 2019; ISBN 9789211483260.
- Nemitz, J. Increasing Longevity and Life Satisfaction: Is There a Catch to Living Longer? J Popul Econ 2022, 35, 557–589. [Google Scholar] [CrossRef]
- Olshansky, S.J. From Lifespan to Healthspan. JAMA 2018, 320, 1323. [Google Scholar] [CrossRef] [PubMed]
- Garmany, A.; Yamada, S.; Terzic, A. Longevity Leap: Mind the Healthspan Gap. NPJ Regen Med 2021, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Ostan, R.; Bucci, L.; Capri, M.; Salvioli, S.; Scurti, M.; Pini, E.; Monti, D.; Franceschi, C. Immunosenescence and Immunogenetics of Human Longevity. Neuroimmunomodulation 2008, 15, 224–240. [Google Scholar] [CrossRef]
- Cevenini, E.; Bellavista, E.; Tieri, P.; Castellani, G.; Lescai, F.; Francesconi, M.; Mishto, M.; Santoro, A.; Valensin, S.; Salvioli, S.; Capri, M.; Zaikin, A.; Monti, D.; de Magalhaes, J.; Franceschi, C. Systems Biology and Longevity: An Emerging Approach to Identify Innovative Anti- Aging Targets and Strategies. Curr Pharm Des 2010, 16, 802–813. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and Aging: Signaling Pathways and Intervention Therapies. Signal Transduct Target Ther 2023, 8, 239. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-Aging. An Evolutionary Perspective on Immunosenescence. Ann N Y Acad Sci 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and Anti-Inflammaging: A Systemic Perspective on Aging and Longevity Emerged from Studies in Humans. Mech Ageing Dev 2007, 128, 92–105. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front Med (Lausanne) 2018, 5. [Google Scholar] [CrossRef]
- Sierra, F.; Kohanski, R. Geroscience and the Trans-NIH Geroscience Interest Group, GSIG. 2017, 1–5. [CrossRef]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The Continuum of Aging and Age-Related Diseases: Common Mechanisms but Different Rates. Front Med (Lausanne) 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Mehdi, M.M. Oxidative Stress, Inflammation and Hormesis: The Role of Dietary and Lifestyle Modifications on Aging. Neurochem Int 2023, 164, 105490. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; Frigé, C.; Pellegrini, V.; Scisciola, L.; Santoro, A.; Monti, D.; Rippo, M.R.; Ivanchenko, M.; Olivieri, F.; Franceschi, C. Organ-Specific Biological Clocks: Ageotyping for Personalized Anti-Aging Medicine. Ageing Res Rev 2024, 96, 102253. [Google Scholar] [CrossRef]
- Fjell, A.M.; Walhovd, K.B. Structural Brain Changes in Aging: Courses, Causes and Cognitive Consequences. Rev Neurosci 2010, 21, 187–221. [Google Scholar] [CrossRef] [PubMed]
- Rodrigue, K.M.; Kennedy, K.M. The Cognitive Consequences of Structural Changes to the Aging Brain; Seventh Ed.; Elsevier Inc., 2011; ISBN 9780123808820.
- Nyberg, L.; Wåhlin, A. The Many Facets of Brain Aging. Elife 2020, 9, 18–20. [Google Scholar] [CrossRef]
- Murman, D.L. The Impact of Age on Cognition. Semin Hear 2015, 36, 111–121. [Google Scholar] [CrossRef]
- Fjell, A.M.; Sneve, M.H.; Grydeland, H.; Storsve, A.B.; Amlien, I.K.; Yendiki, A.; Walhovd, K.B. Relationship between Structural and Functional Connectivity Change across the Adult Lifespan: A Longitudinal Investigation. Hum Brain Mapp 2017, 38, 561–573. [Google Scholar] [CrossRef]
- Seidler, R.D.; Bernard, J.A.; Burutolu, T.B.; Fling, B.W.; Gordon, M.T.; Gwin, J.T.; Kwak, Y.; Lipps, D.B. Motor Control and Aging: Links to Age-Related Brain Structural, Functional, and Biochemical Effects. Neurosci Biobehav Rev 2010, 34, 721–733. [Google Scholar] [CrossRef]
- Park, D.C.; Reuter-Lorenz, P. The Adaptive Brain: Aging and Neurocognitive Scaffolding. Annu Rev Psychol 2009, 60, 173–196. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, G.; Luo, Y.; Jiang, L.; Chi, H.; Tian, G. Neuroinflammation in Alzheimer’s Disease: Insights from Peripheral Immune Cells. Immunity & Ageing 2024, 21, 38. [Google Scholar] [CrossRef]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front Cell Dev Biol 2021, 9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L. The Limited in vitro Lifetime of Human Diploid Cell Strains. Exp Cell Res 1965, 37, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; D’Adda Di Fagagna, F. Cellular Senescence: When Bad Things Happen to Good Cells. Nat Rev Mol Cell Biol 2007, 8, 729–740. [Google Scholar] [CrossRef]
- González-Gualda, E.; Baker, A.G.; Fruk, L.; Muñoz-Espín, D. A Guide to Assessing Cellular Senescence in vitro and in vivo. FEBS Journal 2021, 288, 56–80. [Google Scholar] [CrossRef]
- Campisi, J. Aging, Cellular Senescence, and Cancer. Annu Rev Physiol 2013, 75, 685–705. [Google Scholar] [CrossRef]
- M. Laura Idda; Waverly G. McClusky; Valeria Lodde; Rachel Munk; Kotb Abdelmohsen; Martina Rossi; Myriam Gorospe Survey of Senescent Cell Markers with Age in Human Tissues. Aging 2020, 12, 4052–4066. [Google Scholar]
- Krishnamurthy, J.; Torrice, C.; Ramsey, M.R.; Kovalev, G.I.; Al-Regaiey, K.; Su, L.; Sharpless, N.E. Ink4a/Arf Expression Is a Biomarker of Aging. Journal of Clinical Investigation 2004, 114, 1299–1307. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Wilkinson, J.E.; Hughes, B.; Gadela, N.; Ladiges, W.C.; Vo, N.; Niedernhofer, L.J.; Huffman, D.M.; Robbins, P.D. Heterochronic Parabiosis Regulates the Extent of Cellular Senescence in Multiple Tissues. Geroscience 2020, 42, 951–961. [Google Scholar] [CrossRef]
- Si, Z.; Sun, L.; Wang, X. Evidence and Perspectives of Cell Senescence in Neurodegenerative Diseases. Biomedicine and Pharmacotherapy 2021, 137, 111327. [Google Scholar] [CrossRef]
- Boyko, A.A.; Troyanova, N.I.; Kovalenko, E.I.; Sapozhnikov, A.M. Similarity and Differences in Inflammation-Related Haracteristics of the Peripheral Immune System of Patients with Parkinson’s and Alzheimer’s Diseases. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Nakajima, K.; Kohsaka, S. Microglia: Neuroprotective and Neurotrophic Cells in the Central Nervous System. Curr Drug Targets Cardiovasc Haematol Disord 2004, 4, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Diwan, B.; Sharma, R. Nutritional Components as Mitigators of Cellular Senescence in Organismal Aging: A Comprehensive Review. Food Sci Biotechnol 2022, 31, 1089–1109. [Google Scholar] [CrossRef] [PubMed]
- Głowacka, P.; Oszajca, K.; Pudlarz, A.; Szemraj, J.; Witusik-Perkowska, M. Postbiotics as Molecules Targeting Cellular Events of Aging Brain—The Role in Pathogenesis, Prophylaxis and Treatment of Neurodegenerative Diseases. Nutrients 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Crowe, E.P.; Bitto, A.; Moh, M.; Katsetos, C.D.; Garcia, F.U.; Johnson, F.B.; Trojanowski, J.Q.; Sell, C.; Torres, C. Astrocyte Senescence as a Component of Alzheimer’s Disease. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Walker, L.; Jacobs, E.; McAleese, K.E.; Johnson, M.; Attems, J. Do Senescent Cells Play a Role in Alzheimer’s Disease? Alzheimer’s & Dementia 2020, 16, 43820. [Google Scholar] [CrossRef]
- Simmnacher, K.; Krach, F.; Schneider, Y.; Alecu, J.E.; Mautner, L.; Klein, P.; Roybon, L.; Prots, I.; Xiang, W.; Winner, B. Unique Signatures of Stress-Induced Senescent Human Astrocytes. Exp Neurol 2020, 334, 113466. [Google Scholar] [CrossRef]
- Si, Z.; Sun, L.; Wang, X. Evidence and Perspectives of Cell Senescence in Neurodegenerative Diseases. Biomedicine and Pharmacotherapy 2021, 137, 111327. [Google Scholar] [CrossRef]
- Chinta, S.J.; Woods, G.; Demaria, M.; Rane, A.; Zou, Y.; McQuade, A.; Rajagopalan, S.; Limbad, C.; Madden, D.T.; Campisi, J.; et al. Cellular Senescence Is Induced by the Environmental Neurotoxin Paraquat and Contributes to Neuropathology Linked to Parkinson’s Disease. Cell Rep 2018, 22, 930–940. [Google Scholar] [CrossRef]
- Wang, W.Y.; Tan, M.S.; Yu, J.T.; Tan, L. Role of Pro-Inflammatory Cytokines Released from Microglia in Alzheimer’s Disease. Ann Transl Med 2015, 3, 1–15. [Google Scholar] [CrossRef]
- Bae, E.J.; Choi, M.; Kim, J.T.; Kim, D.K.; Jung, M.K.; Kim, C.; Kim, T.K.; Lee, J.S.; Jung, B.C.; Shin, S.J.; Rhee, K. H.; Lee, S. J. TNF-α Promotes α-Synuclein Propagation through Stimulation of Senescence-Associated Lysosomal Exocytosis. Exp Mol Med 2022, 54, 788–800. [Google Scholar] [CrossRef]
- Hou, Y.; Wei, Y.; Lautrup, S.; Yang, B.; Wang, Y.; Cordonnier, S.; Mattson, M.P.; Croteau, D.L.; Bohr, V.A. NAD+ Supplementation Reduces Neuroinflammation and Cell Senescence in a Transgenic Mouse Model of Alzheimer’s Disease via CGAS-STING. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Dehkordi, S.K.; Walker, J.; Sah, E.; Bennett, E.; Atrian, F.; Frost, B.; Woost, B.; Bennett, R.E.; Orr, T.C.; Zhou, Y.; Andhey, P. S.; Colonna, M.; Sudmant, P. H.; Xu, P.; Wang, M.; Zhang, B.; Zare, H.; Orr, M. E. Profiling Senescent Cells in Human Brains Reveals Neurons with CDKN2D/P19 and Tau Neuropathology. Nat Aging 2021, 1, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Bussian, T.; Aziz, A.; Meyer, C.; Swenson, B.; Deursen, J.; Baker, D. Clearance of Senescent Glial Cells Prevents Tau-Dependent Pathology and Cognitive Decline. Nature 2018, 562. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Mathers, J.C. Impact of Nutrition on the Ageing Process. British Journal of Nutrition 2015, 113, S18–S22. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive Compounds: Definition and Assessment of Activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef]
- Morris, M.C. Nutrition and Risk of Dementia: Overview and Methodological Issues. Ann N Y Acad Sci 2016, 1367, 31–37. [Google Scholar] [CrossRef]
- Scuto, M.; Rampulla, F.; Reali, G.M.; Spanò, S.M.; Trovato Salinaro, A.; Calabrese, V. Hormetic Nutrition and Redox Regulation in Gut–Brain Axis Disorders. Antioxidants 2024, 13. [Google Scholar] [CrossRef]
- Aliper, A.; Belikov, A. V.; Garazha, A.; Jellen, L.; Artemov, A.; Suntsova, M.; Ivanova, A.; Venkova, L.; Borisov, N.; Buzdin, A.; Mamoshina, P.; Putin, E.; Swick, A. G.; Moskalev, A.; Zhavoronkov, A. In Search for Geroprotectors: In Silico Screening and in vitro Validation of Signalome-Level Mimetics of Young Healthy State. Aging 2016, 8, 2127–2152. [Google Scholar] [CrossRef]
- Aliper, A.; Jellen, L.; Cortese, F.; Artemov, A.; Semper, D.K.; Moskalev, A.; Swick, A.G.; Zhavoronkov, A. Natural Mimetics of Rapamycin and Minoxidil Obtained via Computational Methods. Aging (Albany NY) 2017, 9, 2245–2268. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Liang, Y.; Zhang, C.; Xu, Z.; Zhang, L.; Fuji, R.; Mu, W.; Li, L.; Jiang, J.; Ju, Y.; Wang, Z. Cyclic AMP Mimics the Anti-Ageing Effects of Calorie Restriction by Up-Regulating Sirtuin. Sci Rep 2015, 5, 12012. [Google Scholar] [CrossRef] [PubMed]
- Naisam, S.; Mohan, A.; Sreekumar, N. Epigenetic Regulation of Human Sirtuin 1 Insights into Aging Mechanisms 2024.
- Effect of Calorie Restriction on the Expression of Sirtuin1 as an Antiaging Biomarker. Makara J Sci 2023, 27. [CrossRef]
- Dhillon, R.S.; Qin, Y.; van Ginkel, P.R.; Fu, V.X.; Vann, J.M.; Lawton, A.J.; Green, C.L.; Manchado-Gobatto, F.B.; Gobatto, C.A.; Lamming, D.W.; Prolla, T.A.; Denu, J.M. SIRT3 Deficiency Decreases Oxidative Metabolism Capacity but Increases Lifespan in Male Mice under Caloric Restriction. Aging Cell 2022, 21. [Google Scholar] [CrossRef]
- Gurău, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafè, M.; Olivieri, F. Anti-Senescence Compounds: A Potential Nutraceutical Approach to Healthy Aging. Ageing Res Rev 2018, 46, 14–31. [Google Scholar] [CrossRef]
- Hadem, I.K.H.; Majaw, T.; Kharbuli, B.; Sharma, R. Beneficial Effects of Dietary Restriction in Aging Brain. J Chem Neuroanat 2019, 95, 123–133. [Google Scholar] [CrossRef]
- Krishnamurthy, J.; Torrice, C.; Ramsey, M.R.; Kovalev, G.I.; Al-Regaiey, K.; Su, L.; Sharpless, N.E. Ink4a/Arf Expression Is a Biomarker of Aging. Journal of Clinical Investigation 2004, 114, 1299–1307. [Google Scholar] [CrossRef]
- Fontana, L.; Mitchell, S.E.; Wang, B.; Tosti, V.; van Vliet, T.; Veronese, N.; Bertozzi, B.; Early, D.S.; Maissan, P.; Speakman, J.R.; Demaria, M. The Effects of Graded Caloric Restriction: XII. Comparison of Mouse to Human Impact on Cellular Senescence in the Colon. Aging Cell 2018, 17, 4–8. [Google Scholar] [CrossRef]
- Wang, C.; Maddick, M.; Miwa, S.; Jurk, D.; Czapiewski, R.; Saretzki, G.; Langie, S.A.S.; Godschalk, R.W.L.; Cameron, K.; von Zglinicki, T. Adult-Onset, Short-Term Dietary Restriction Reduces Cell Senescence in Mice. Aging 2010, 2, 555–566. [Google Scholar] [CrossRef]
- Aversa, Z.; White, T.A.; Heeren, A.A.; Hulshizer, C.A.; Saul, D.; Zhang, X.; Molina, A.J.A.; Redman, L.M.; Martin, C.K.; Racette, S.B.; et al. Calorie Restriction Reduces Biomarkers of Cellular Senescence in Humans. Aging Cell 2024, 23, 1–11. [Google Scholar] [CrossRef]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S.; Tabolacci, C.; Jadeja, R.N. Beneficial Role of Phytochemicals on Oxidative Stress and Age-Related Diseases. Biomed Res Int 2019. [Google Scholar] [CrossRef]
- Ooi, T.C.; Meramat, A.; Rajab, N.F.; Shahar, S.; Ismail, I.S.; Azam, A.A.; Sharif, R. Intermittent Fasting Enhanced the Cognitive Function in Older Adults with Mild Cognitive Impairment by Inducing Biochemical and Metabolic Changes: A 3-Year Progressive Study. Nutrients 2020, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Teng, N.I.M.F.; Shahar, S.; Rajab, N.F.; Manaf, Z.A.; Johari, M.H.; Ngah, W.Z.W. Improvement of Metabolic Parameters in Healthy Older Adult Men Following a Fasting Calorie Restriction Intervention. Aging Male 2013, 16, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Chiavaroli, L.; Nishi, S.K.; Khan, T.A.; Braunstein, C.R.; Glenn, A.J.; Mejia, S.B.; Rahelić, D.; Kahleová, H.; Salas-Salvadó, J.; Jenkins, D.J.A.; Kendall, C.W.C.; Sievenpiper, J.L. Portfolio Dietary Pattern and Cardiovascular Disease: A Systematic Review and Meta-Analysis of Controlled Trials. Prog Cardiovasc Dis 2018, 61, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Ndanuko, R.N.; Tapsell, L.C.; Charlton, K.E.; Neale, E.P.; Batterham, M.J. Dietary Patterns and Blood Pressure in Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Advances in Nutrition 2016, 7, 76–89. [Google Scholar] [CrossRef]
- Fontana, L.; Mitchell, S.E.; Wang, B.; Tosti, V.; van Vliet, T.; Veronese, N.; Bertozzi, B.; Early, D.S.; Maissan, P.; Speakman, J.R.; et al. The Effects of Graded Caloric Restriction: XII. Comparison of Mouse to Human Impact on Cellular Senescence in the Colon. Aging Cell 2018, 17, 4–8. [Google Scholar] [CrossRef]
- Barbaresko, J.; Koch, M.; Schulze, M.B.; Nöthlings, U. Dietary Pattern Analysis and Biomarkers of Low-Grade Inflammation: A Systematic Literature Review. Nutr Rev 2013, 71, 511–527. [Google Scholar] [CrossRef]
- Grande de França, N.A.; Rolland, Y.; Guyonnet, S.; de Souto Barreto, P. The Role of Dietary Strategies in the Modulation of Hallmarks of Aging. Ageing Res Rev 2023, 87, 101908. [Google Scholar] [CrossRef]
- Ravussin, E.; Redman, L.M.; Rochon, J.; Das, S.K.; Fontana, L.; Kraus, W.E.; Romashkan, S.; Williamson, D.A.; Meydani, S.N.; Villareal, D.T.; Smith, Steven R. ; Stein, R.I.; Scott, T.M.; Stewart, T.M.; Saltzman, E.; Klein, S.; Bhapkar, M.; Martin, C.K.; Gilhooly, C.H.; Holloszy, J.O.; Hadley, E.C.; Roberts, S.B. A 2-Year Randomized Controlled Trial of Human Caloric Restriction: Feasibility and Effects on Predictors of Health Span and Longevity. J Gerontol A Biol Sci Med Sci 2015, 70, 1097–1104. [Google Scholar] [CrossRef]
- Panda, S.; Maier, G.; Villareal, D.T. Targeting Energy Intake and Circadian Biology to Engage Mechanisms of Aging in Older Adults With Obesity: Calorie Restriction and Time-Restricted Eating. The Journals of Gerontology: Series A 2023, 78, 79–85. [Google Scholar] [CrossRef]
- Sofi, F.; Cesari, F.; Abbate, R.; Gensini, G.F.; Casini, A. Adherence to Mediterranean Diet and Health Status: Meta-Analysis. BMJ 2008, 337, a1344. [Google Scholar] [CrossRef]
- Xavier Medina, F. Mediterranean Diet, Culture and Heritage: Challenges for a New Conception. Public Health Nutr 2009, 12, 1618–1620. [Google Scholar] [CrossRef] [PubMed]
- Marin, C.; Delgado-Lista, J.; Ramirez, R.; Carracedo, J.; Caballero, J.; Perez-Martinez, P.; Gutierrez-Mariscal, F.M.; Garcia-Rios, A.; Delgado-Casado, N.; Cruz-Teno, C.; Yubero-Serrano, E.M.; Tinahones, F.; Malagon, M.D.M.; Perez-Jimenez, F.; Lopez-Miranda, J. Mediterranean Diet Reduces Senescence-Associated Stress in Endothelial Cells. Age (Omaha) 2012, 34, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Salas-Salvadó, J.; Guasch-Ferré, M.; Lee, C.H.; Estruch, R.; Clish, C.B.; Ros, E. Protective Effects of the Mediterranean Diet on Type 2 Diabetes and Metabolic Syndrome. Journal of Nutrition 2016, 146, 920S–927S. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.T.; Davis, C.R.; Dyer, K.A.; Hodgson, J.M.; Woodman, R.J.; Keage, H.A.D.; Murphy, K.J. A Mediterranean Diet to Improve Cardiovascular and Cognitive Health: Protocol for a Randomised Controlled Intervention Study. Nutrients 2017, 9, 1–29. [Google Scholar] [CrossRef]
- Fekete, M.; Varga, P.; Ungvari, Z.; Tibor, J.; Annamaria, F.; Ágnes, B.; Lehoczki, A.; Mózes, N.; Grosso, G.; Godos, J.; Menyhart, O.; Munkácsy, G.; Tarantini, S.; Yabluchanskiy, A.; Ungvari, A.; Győrffy, B. The Role of the Mediterranean Diet in Reducing the Risk of Cognitive Impairement, Dementia, and Alzheimer ‘s Disease: A Meta - Analysis. Geroscience 2025. [Google Scholar] [CrossRef]
- Andreo-López, M.C.; Contreras-Bolívar, V.; Muñoz-Torres, M.; García-Fontana, B.; García-Fontana, C. Influence of the Mediterranean Diet on Healthy Aging. Int J Mol Sci 2023, 24, 4491. [Google Scholar] [CrossRef]
- Marin, C.; Ramirez, R.; Delgado-Lista, J.; Yubero-Serrano, E.M.; Perez-Martinez, P.; Carracedo, J.; Garcia-Rios, A.; Rodriguez, F.; Gutierrez-Mariscal, F.M.; Gomez, P.; Perez-Jimenez, F.; Lopez-Maria, J. Mediterranean Diet Reduces Endothelial Damage and Improves the Regenerative Capacity of Endothelium. American Journal of Clinical Nutrition 2011, 93, 267–274. [Google Scholar] [CrossRef]
- Mantilla-Escalante, D.C.; López de las Hazas, M.C.; Crespo, M.C.; Martín-Hernández, R.; Tomé-Carneiro, J.; del Pozo-Acebo, L.; Salas-Salvadó, J.; Bulló, M.; Dávalos, A. Mediterranean Diet Enriched in Extra-Virgin Olive Oil or Nuts Modulates Circulating Exosomal Non-Coding RNAs. Eur J Nutr 2021, 60, 4279–4293. [Google Scholar] [CrossRef]
- Bacalini, M.G.; Friso, S.; Olivieri, F.; Pirazzini, C.; Giuliani, C.; Capri, M.; Santoro, A.; Franceschi, C.; Garagnani, P. Present and Future of Anti-Ageing Epigenetic Diets. Mech Ageing Dev 2014, 136–137, 101–115. [Google Scholar] [CrossRef]
- Lee, P.S.; Chiou, Y.S.; Ho, C.T.; Pan, M.H. Chemoprevention by Resveratrol and Pterostilbene: Targeting on Epigenetic Regulation. BioFactors 2018, 44, 26–35. [Google Scholar] [CrossRef]
- Bekdash, R.A. Epigenetics, Nutrition, and the Brain: Improving Mental Health through Diet. Int J Mol Sci 2024, 25, 4036. [Google Scholar] [CrossRef] [PubMed]
- Pazoki-Toroudi, H.; Amani, H.; Ajami, M.; Nabavi, S.F.; Braidy, N.; Kasi, P.D.; Nabavi, S.M. Targeting MTOR Signaling by Polyphenols: A New Therapeutic Target for Ageing. Ageing Res Rev 2016, 31, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Gillette-Guyonnet, S.; Secher, M.; Vellas, B. Nutrition and Neurodegeneration: Epidemiological Evidence and Challenges for Future Research. Br J Clin Pharmacol 2013, 75, 738–755. [Google Scholar] [CrossRef]
- Vellas, B.; Carrie, I.; Gillette-Guyonnet, S.; Touchon, J.; Dantoine, T.; Dartigues, J.F.; Cuffi, M.N.; Bordes, S.; Gasnier, Y.; Robert, P.; et al. Alzheimer’s Disease : Design and Baseline Data. J Prev Alzheimers Dis 2014, 1, 13–22. [Google Scholar]
- Ngandu, T.; Lehtisalo, J.; Solomon, A.; Levälahti, E.; Ahtiluoto, S.; Antikainen, R.; Bäckman, L.; Hänninen, T.; Jula, A.; Laatikainen, T.; Lindström, J.; Mangialasche, F.; Paajanen, T.; Pajala, S.; Peltonen, M.; Rauramaa, R.; Stigsdotter-Neely, A.; Strandberg, T.; Tuomilehto, J.; Soininen, H.; Kivipelto, M. A 2 Year Multidomain Intervention of Diet, Exercise, Cognitive Training, and Vascular Risk Monitoring versus Control to Prevent Cognitive Decline in at-Risk Elderly People (FINGER): A Randomised Controlled Trial. The Lancet 2015, 385, 2255–2263. [Google Scholar] [CrossRef]
- Palou, A.; Serra, F.; Pico, C. General Aspects on the Assessment of Functional Foods in the European Union. Eur J Clin Nutr 2003, 57, S12–S17. [Google Scholar] [CrossRef]
- Siró, I.; Kápolna, E.; Kápolna, B.; Lugasi, A. Functional Food. Product Development, Marketing and Consumer Acceptance—A Review. Appetite 2008, 51, 456–467. [Google Scholar] [CrossRef]
- Jeck, W.R.; Siebold, A.P.; Sharpless, N.E. Review: A Meta-analysis of GWAS and Age-associated Diseases. Aging Cell 2012, 11, 727–731. [Google Scholar] [CrossRef]
- Martucci, M.; Conte, M.; Bucci, L.; Giampieri, E.; Fabbri, C.; Palmas, M.G.; Izzi, M.; Salvioli, S.; Zambrini, A.V.; Orsi, C.; Brigidi, P.; Santoro, A.; Capri, M.; Monti, D.; Franceschi, C. Twelve-Week Daily Consumption of Ad Hoc Fortified Milk with ω-3, D, and Group B Vitamins Has a Positive Impact on Inflammaging Parameters: A Randomized Cross-over Trial. Nutrients 2020, 12, 1–19. [Google Scholar] [CrossRef]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain–Raspaud, S.; Trotin, B.; Naliboff, B.; Mayer, E.A. Consumption of Fermented Milk Product With Probiotic Modulates Brain Activity. Gastroenterology 2013, 144, 1394–1401.e4. [Google Scholar] [CrossRef]
- Navarro-Hortal, M.D.; Romero-Márquez, J.M.; Jiménez-Trigo, V.; Xiao, J.; Giampieri, F.; Forbes-Hernández, T.Y.; Grosso, G.; Battino, M.; Sánchez-González, C.; Quiles, J.L. Molecular Bases for the Use of Functional Foods in the Management of Healthy Aging: Berries, Curcumin, Virgin Olive Oil and Honey; Three Realities and a Promise. Crit Rev Food Sci Nutr 2023, 63, 11967–11986. [Google Scholar] [CrossRef] [PubMed]
- Driver, C. Mitochondrial Interventions in Aging and Longevity. In Modulating Aging and Longevity; Springer Netherlands: Dordrecht, 2003; pp. 205–217. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; El-Sawi, M.R. Melatonin Reduces Oxidant Damage and Promotes Mitochondrial Respiration: Implications for Aging. Ann N Y Acad Sci 2002, 959, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, D.J.; Parise, G.; Tarnopolsky, M.A. Nutritional and Exercise-Based Therapies in the Treatment of Mitochondrial Disease. Curr Opin Clin Nutr Metab Care 2002, 5, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc Natl Acad Sci U S A 1993, 90, 7915–7922. [Google Scholar] [CrossRef]
- Lyras, L.; Cairns, N.J.; Jenner, A.; Jenner, P.; Halliwell, B. An Assessment of Oxidative Damage to Proteins, Lipids, and DNA in Brain from Patients with Alzheimer’s Disease. J Neurochem 1997, 68, 2061–2069. [Google Scholar] [CrossRef]
- Bej, E.; Cesare, P.; Volpe, A.R.; d’Angelo, M.; Castelli, V. Oxidative Stress and Neurodegeneration: Insights and Therapeutic Strategies for Parkinson’s Disease. Neurol Int 2024, 16, 502–517. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free Radicals, Metals and Antioxidants in Oxidative Stress-Induced Cancer. Chem Biol Interact 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Stadtman, E.R. Role of Oxidant Species in Aging. Curr Med Chem 2004, 11, 1105–1112. [Google Scholar] [CrossRef]
- von Zglinicki, T. Oxidative Stress and Cell Senescence as Drivers of Ageing: Chicken and Egg. Ageing Res Rev 2024, 102, 102558. [Google Scholar] [CrossRef]
- May, J.M. Vitamin C Transport and Its Role in the Central Nervous System. In Water Soluble Vitamins. Subcellular Biochemistry; O. Stanger; 2012; Volume 56, pp. 85–103.
- Harrison, F.E.; May, J.M. Vitamin C Function in the Brain: Vital Role of the Ascorbate Transporter SVCT2. Free Radic Biol Med 2009, 46, 719–730. [Google Scholar] [CrossRef]
- Harrison, F.E.; Hosseini, A.H.; McDonald, M.P.; May, J.M. Vitamin C Reduces Spatial Learning Deficits in Middle-Aged and Very Old APP/PSEN1 Transgenic and Wild-Type Mice. Pharmacol Biochem Behav 2009, 93, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, D.; Parle, M.; Kulkarni, S.K. Comparative Brain Cholinesterase-Inhibiting Activity of Glycyrrhiza Glabra, Myristica Fragrans, Ascorbic Acid, and Metrifonate in Mice. J Med Food 2006, 9, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Chang, M.; Park, C.; Kim, H.; Kim, J.; Son, H.; Lee, Y.; Lee, S. Ascorbate-induced Differentiation of Embryonic Cortical Precursors into Neurons and Astrocytes. J Neurosci Res 2003, 73, 156–165. [Google Scholar] [CrossRef]
- Harrison, F.; Bowman, G.; Polidori, M. Ascorbic Acid and the Brain: Rationale for the Use against Cognitive Decline. Nutrients 2014, 6, 1752–1781. [Google Scholar] [CrossRef]
- Nualart, F. Vitamin C Transporters, Recycling and the Bystander Effect in the Nervous System: SVCT2 versus Gluts. J Stem Cell Res Ther 2014, 04. [Google Scholar] [CrossRef]
- Belluzzi, E.; Bisaglia, M.; Lazzarini, E.; Tabares, L.C.; Beltramini, M.; Bubacco, L. Human SOD2 Modification by Dopamine Quinones Affects Enzymatic Activity by Promoting Its Aggregation: Possible Implications for Parkinson’s Disease. PLoS One 2012, 7, e38026. [Google Scholar] [CrossRef]
- Man Anh, H.; Linh, D.M.; My Dung, V.; Thi Phuong Thao, D. Evaluating Dose- and Time-Dependent Effects of Vitamin C Treatment on a Parkinson’s Disease Fly Model. Parkinsons Dis 2019, 2019, 1–14. [Google Scholar] [CrossRef]
- Casani, S.; Gómez-Pastor, R.; Matallana, E.; Paricio, N. Antioxidant Compound Supplementation Prevents Oxidative Damage in a Drosophila Model of Parkinson’s Disease. Free Radic Biol Med 2013, 61, 151–160. [Google Scholar] [CrossRef]
- Etminan, M.; Gill, S.S.; Samii, A. Intake of Vitamin E, Vitamin C, and Carotenoids and the Risk of Parkinson’s Disease: A Meta-Analysis. Lancet Neurol 2005, 4, 362–365. [Google Scholar] [CrossRef]
- Grünewald, R.A. Ascorbic Acid in the Brain. Brain Res Rev 1993, 18, 123–133. [Google Scholar] [CrossRef]
- Mefford, I.N.; Oke, A.F.; Adams, R.N. Regional Distribution of Ascorbate in Human Brain. Brain Res 1981, 212, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Méndez, R.; Rivas-Arancibia, S. Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain. Front Physiol 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Seitz, G.; Gebhardt, S.; Beck, J.F.; Böhm, W.; Lode, H.N.; Niethammer, D.; Bruchelt, G. Ascorbic Acid Stimulates DOPA Synthesis and Tyrosine Hydroxylase Gene Expression in the Human Neuroblastoma Cell Line SK-N-SH. Neurosci Lett 1998, 244, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, H.; Hamamoto, M.; Ueda, M.; Nito, C.; Yamaguchi, H.; Katayama, Y. The Effect of Ascorbic Acid on the Pharmacokinetics of Levodopa in Elderly Patients with Parkinson Disease. Clin Neuropharmacol 2004, 27, 270–273. [Google Scholar] [CrossRef]
- Nikolova, G.; Karamalakova, Y.; Gadjeva, V. Reducing Oxidative Toxicity of L-Dopa in Combination with Two Different Antioxidants: An Essential Oil Isolated from Rosa Damascena Mill., and Vitamin C. Toxicol Rep 2019, 6, 267–271. [Google Scholar] [CrossRef]
- Yan, J.; Studer, L.; McKay, R.D.G. Ascorbic Acid Increases the Yield of Dopaminergic Neurons Derived from Basic Fibroblast Growth Factor Expanded Mesencephalic Precursors. J Neurochem 2001, 76, 307–311. [Google Scholar] [CrossRef]
- Hughes, K.C.; Gao, X.; Kim, I.Y.; Rimm, E.B.; Wang, M.; Weisskopf, M.G.; Schwarzschild, M.A.; Ascherio, A. Intake of Antioxidant Vitamins and Risk of Parkinson’s Disease. Movement Disorders 2016, 31, 1909–1914. [Google Scholar] [CrossRef]
- Yang, F.; Wolk, A.; Håkansson, N.; Pedersen, N.L.; Wirdefeldt, K. Dietary Antioxidants and Risk of Parkinson’s Disease in Two Population-based Cohorts. Movement Disorders 2017, 32, 1631–1636. [Google Scholar] [CrossRef]
- Miyake, Y.; Fukushima, W.; Tanaka, K.; Sasaki, S.; Kiyohara, C.; Tsuboi, Y.; Yamada, T.; Oeda, T.; Miki, T.; Kawamura, N.; Fukuyama, H.; Hirota, Y.; Nagai, M. ; Dietary Intake of Antioxidant Vitamins and Risk of Parkinson’s Disease: A Case–Control Study in Japan. Eur J Neurol 2011, 18, 106–113. [Google Scholar] [CrossRef]
- Schirinzi, T.; Martella, G.; Imbriani, P.; Di Lazzaro, G.; Franco, D.; Colona, V.L.; Alwardat, M.; Sinibaldi Salimei, P.; Mercuri, N.B.; Pierantozzi, M.; Pisani, A. ; Dietary Vitamin E as a Protective Factor for Parkinson’s Disease: Clinical and Experimental Evidence. Front Neurol 2019, 10. [Google Scholar] [CrossRef]
- Casani, S.; Gómez-Pastor, R.; Matallana, E.; Paricio, N. Antioxidant Compound Supplementation Prevents Oxidative Damage in a Drosophila Model of Parkinson’s Disease. Free Radic Biol Med 2013, 61, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.; Kesler, N. Monoamine Oxidase and Mitochondrial Respiration. J Neurochem 1999, 73, 2310–2315. [Google Scholar] [CrossRef] [PubMed]
- Ithayarasi, A.P.; Shyamala Devi, C. Effect of Alpha-Tocopherol on Mitochondrial Electron Transport in Experimental Myocardial Infarction in Rats. Indian J Biochem Biophys 1998, 35, 115–119. [Google Scholar] [PubMed]
- Nakaso, K.; Tajima, N.; Horikoshi, Y.; Nakasone, M.; Hanaki, T.; Kamizaki, K.; Matsura, T. The Estrogen Receptor β-PI3K/Akt Pathway Mediates the Cytoprotective Effects of Tocotrienol in a Cellular Parkinson’s Disease Model. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2014, 1842, 1303–1312. [Google Scholar] [CrossRef]
- Nakaso, K.; Horikoshi, Y.; Takahashi, T.; Hanaki, T.; Nakasone, M.; Kitagawa, Y.; Koike, T.; Matsura, T. Estrogen Receptor-Mediated Effect of δ-Tocotrienol Prevents Neurotoxicity and Motor Deficit in the MPTP Mouse Model of Parkinson’s Disease. Neurosci Lett 2016, 610, 117–122. [Google Scholar] [CrossRef]
- Odunze, I.N.; Klaidman, L.K.; Adams, J.D. MPTP Toxicity in the Mouse Brain and Vitamin E. Neurosci Lett 1990, 108, 346–349. [Google Scholar] [CrossRef]
- Perry, T.L.; Yong, V.W.; Clavier, R.M.; Jones, K.; Wright, J.M.; Foulks, J.G.; Wall, R.A. Partial Protection from the Dopaminergic Neurotoxin N-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine by Four Different Antioxidants in the Mouse. Neurosci Lett 1985, 60, 109–114. [Google Scholar] [CrossRef]
- Hao, X.; Li, H.; Li, Q.; Gao, D.; Wang, X.; Wu, C.; Wang, Q.; Zhu, M. Dietary Vitamin E Intake and Risk of Parkinson’s Disease: A Cross-Sectional Study. Front Nutr 2024, 10. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X. Antioxidant Therapies for Alzheimer’s Disease. Oxid Med Cell Longev 2012, 2012, 1–17. [Google Scholar] [CrossRef]
- Choudhry, F.; Howlett, D.R.; Richardson, J.C.; Francis, P.T.; Williams, R.J. Pro-Oxidant Diet Enhances β/γ Secretase-Mediated APP Processing in APP/PS1 Transgenic Mice. Neurobiol Aging 2012, 33, 960–968. [Google Scholar] [CrossRef]
- Huang, J.; May, J.M. Ascorbic Acid Protects SH-SY5Y Neuroblastoma Cells from Apoptosis and Death Induced by β-Amyloid. Brain Res 2006, 1097, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Corral, S.; Tan, D.; Reiter, R.J.; Valdivia-Velázquez, M.; Martínez-Barboza, G.; Pablo Acosta-Martínez, J.; Ortiz, G.G. Orally Administered Melatonin Reduces Oxidative Stress and Proinflammatory Cytokines Induced by Amyloid- β Peptide in Rat Brain: A Comparative, in vivo Study versus Vitamin C and E. J Pineal Res 2003, 35, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Role of Vitamin E as a Lipid-Soluble Peroxyl Radical Scavenger: In vitro and in vivo Evidence. Free Radic Biol Med 2014, 66, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ulatowski, L.M.; Manor, D. Vitamin E and Neurodegeneration. Neurobiol Dis 2015, 84, 78–83. [Google Scholar] [CrossRef]
- Yatin, S.M.; Varadarajan, S.; Butterfield, D.A. Vitamin E Prevents Alzheimer’s Amyloid ß-Peptide (1-42)-Induced Neuronal Protein Oxidation and Reactive Oxygen Species Production. Journal of Alzheimer’s Disease 2000, 2, 123–131. [Google Scholar] [CrossRef]
- Yatin, S.M.; Varadarajan, S.; Butterfield, D.A. Vitamin E Prevents Alzheimer’s Amyloid ß-Peptide (1-42)-Induced Neuronal Protein Oxidation and Reactive Oxygen Species Production. Journal of Alzheimer’s Disease 2000, 2, 123–131. [Google Scholar] [CrossRef]
- Scimemi, A.; Meabon, J.S.; Woltjer, R.L.; Sullivan, J.M.; Diamond, J.S.; Cook, D.G. Amyloid-β 1–42 Slows Clearance of Synaptically Released Glutamate by Mislocalizing Astrocytic GLT-1. The Journal of Neuroscience 2013, 33, 5312–5318. [Google Scholar] [CrossRef]
- Mattson, M.P. Pathways towards and Away from Alzheimer’s Disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef]
- Pérez, M.; Cuadros, R.; Smith, M.A.; Perry, G.; Avila, J. Phosphorylated, but Not Native, Tau Protein Assembles Following Reaction with the Lipid Peroxidation Product, 4-hydroxy-2-nonenal. FEBS Lett 2000, 486, 270–274. [Google Scholar] [CrossRef]
- Gamblin, T.C.; King, M.E.; Kuret, J.; Berry, R.W.; Binder, L.I. Oxidative Regulation of Fatty Acid-Induced Tau Polymerization. Biochemistry 2000, 39, 14203–14210. [Google Scholar] [CrossRef]
- Jiang, Q.; Yin, X.; Lill, M.A.; Danielson, M.L.; Freiser, H.; Huang, J. Long-Chain Carboxychromanols, Metabolites of Vitamin E, Are Potent Inhibitors of Cyclooxygenases. Proceedings of the National Academy of Sciences 2008, 105, 20464–20469. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Elson-Schwab, I.; Courtemanche, C.; Ames, B.N. γ-Tocopherol and Its Major Metabolite, in Contrast to α-Tocopherol, Inhibit Cyclooxygenase Activity in Macrophages and Epithelial Cells. Proceedings of the National Academy of Sciences 2000, 97, 11494–11499. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-H.; Kim, M.-B.; Kim, C.; Hwang, J.-K. Inhibitory Effect of Vitamin C on Intrinsic Aging in Human Dermal Fibroblasts and Hairless Mice. Food Sci Biotechnol 2017. [Google Scholar] [CrossRef] [PubMed]
- Teawcharoensopa, C.; Srisuwan, T. The Potential Use of Ascorbic Acid to Recover the Cellular Senescence of Lipopolysaccharide-Induced Human Apical Papilla Cells: An in vitro Study. Clin Oral Investig 2023, 28, 49. [Google Scholar] [CrossRef]
- La Fata, G.; Seifert, N.; Weber, P.; Mohajeri, M.H. Vitamin E Supplementation Delays Cellular Senescence In vitro. Biomed Res Int 2015, 2015, 1–11. [Google Scholar] [CrossRef]
- Ricciarelli, R.; Azzi, A.; Zingg, J. Reduction of Senescence-associated Beta-galactosidase Activity by Vitamin E in Human Fibroblasts Depends on Subjects’ Age and Cell Passage Number. BioFactors 2020, 46, 665–674. [Google Scholar] [CrossRef]
- Mandel, S.A.; Amit, T.; Weinreb, O.; Youdim, M.B.H. Understanding the Broad-Spectrum Neuroprotective Action Profile of Green Tea Polyphenols in Aging and Neurodegenerative Diseases. Journal of Alzheimer’s Disease 2011, 25, 187–208. [Google Scholar] [CrossRef]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or Signalling Molecules? Free Radic Biol Med 2004, 36, 838–849. [Google Scholar] [CrossRef]
- Menicacci, B.; Cipriani, C.; Margheri, F.; Mocali, A.; Giovannelli, L. Modulation of the Senescence-Associated Inflammatory Phenotype in Human Fibroblasts by Olive Phenols. Int J Mol Sci 2017, 18, 2275. [Google Scholar] [CrossRef]
- Lim, H.; Park, H.; Kim, H.P. Effects of Flavonoids on Senescence-Associated Secretory Phenotype Formation from Bleomycin-Induced Senescence in BJ Fibroblasts. Biochem Pharmacol 2015, 96, 337–348. [Google Scholar] [CrossRef]
- Bientinesi, E.; Lulli, M.; Becatti, M.; Ristori, S.; Margheri, F.; Monti, D. Doxorubicin-Induced Senescence in Normal Fibroblasts Promotes in vitro Tumour Cell Growth and Invasiveness: The Role of Quercetin in Modulating These Processes. Mech Ageing Dev 2022, 206. [Google Scholar] [CrossRef] [PubMed]
- Bientinesi, E.; Ristori, S.; Lulli, M.; Monti, D. Quercetin Induces Senolysis of Doxorubicin-Induced Senescent Fibroblasts by Reducing Autophagy, Preventing Their pro-Tumour Effect on Osteosarcoma Cells. Mech Ageing Dev 2024, 220. [Google Scholar] [CrossRef] [PubMed]
- Maurya, P.K.; Kumar, P.; Nagotu, S.; Chand, S.; Chandra, P. Multi-Target Detection of Oxidative Stress Biomarkers in Quercetin and Myricetin Treated Human Red Blood Cells. RSC Adv 2016, 6, 53195–53202. [Google Scholar] [CrossRef]
- Jung, U.J.; Kim, S.R. Beneficial Effects of Flavonoids Against Parkinson’s Disease. J Med Food 2018, 21, 421–432. [Google Scholar] [CrossRef]
- Guo, S.; Yan, J.; Yang, T.; Yang, X.; Bezard, E.; Zhao, B. Protective Effects of Green Tea Polyphenols in the 6-OHDA Rat Model of Parkinson’s Disease Through Inhibition of ROS-NO Pathway. Biol Psychiatry 2007, 62, 1353–1362. [Google Scholar] [CrossRef]
- Levites, Y.; Youdim, M.B.H.; Maor, G.; Mandel, S. Attenuation of 6-Hydroxydopamine (6-OHDA)-Induced Nuclear Factor-KappaB (NF-ΚB) Activation and Cell Death by Tea Extracts in Neuronal Cultures. Biochem Pharmacol 2002, 63, 21–29. [Google Scholar] [CrossRef]
- Rojas, P.; Serrano-García, N.; Mares-Sámano, J.J.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Ögren, S.O. EGb761 Protects against Nigrostriatal Dopaminergic Neurotoxicity in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced Parkinsonism in Mice: Role of Oxidative Stress. European Journal of Neuroscience 2008, 28, 41–50. [Google Scholar] [CrossRef]
- Wu, W.-R.; Zhu, X.-Z. Involvement of Monoamine Oxidase Inhibition in Neuroprotective and Neurorestorative Effects of Ginkgo Biloba Extract against MPTP-Induced Nigrostriatal Dopaminergic Toxicity in C57 Mice. Life Sci 1999, 65, 157–164. [Google Scholar] [CrossRef]
- Khan, Mohd. M.; Ahmad, A.; Ishrat, T.; Khan, M.B.; Hoda, Md.N.; Khuwaja, G.; Raza, S.S.; Khan, A.; Javed, H.; Vaibhav, K.; et al. Resveratrol Attenuates 6-Hydroxydopamine-Induced Oxidative Damage and Dopamine Depletion in Rat Model of Parkinson’s Disease. Brain Res 2010, 1328, 139–151. [Google Scholar] [CrossRef]
- Wang, H.; Dong, X.; Liu, Z.; Zhu, S.; Liu, H.; Fan, W.; Hu, Y.; Hu, T.; Yu, Y.; Li, Y.; et al. Resveratrol Suppresses Rotenone-induced Neurotoxicity Through Activation of SIRT1/Akt1 Signaling Pathway. Anat Rec 2018, 301, 1115–1125. [Google Scholar] [CrossRef]
- Gao, Z.-B.; Chen, X.-Q.; Hu, G.-Y. Inhibition of Excitatory Synaptic Transmission by Trans-Resveratrol in Rat Hippocampus. Brain Res 2006, 1111, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, S.-J. Inhibitory Effect of Glutamate Release from Rat Cerebrocortical Nerve Terminals by Resveratrol. Neurochem Int 2009, 54, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Yáñez, M.; Fraiz, N.; Cano, E.; Orallo, F. Inhibitory Effects of Cis- and Trans-Resveratrol on Noradrenaline and 5-Hydroxytryptamine Uptake and on Monoamine Oxidase Activity. Biochem Biophys Res Commun 2006, 344, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Palazzi, L.; Bruzzone, E.; Bisello, G.; Leri, M.; Stefani, M.; Bucciantini, M.; Polverino de Laureto, P. Oleuropein Aglycone Stabilizes the Monomeric α-Synuclein and Favours the Growth of Non-Toxic Aggregates. Sci Rep 2018, 8, 8337. [Google Scholar] [CrossRef]
- Mohammad-Beigi, H.; Aliakbari, F.; Sahin, C.; Lomax, C.; Tawfike, A.; Schafer, N.P.; Amiri-Nowdijeh, A.; Eskandari, H.; Møller, I.M.; Hosseini-Mazinani, M.; Christiansen, G.; Ward, J.L.; Morshedi, D.; Otzen, D.E. ; Oleuropein Derivatives from Olive Fruit Extracts Reduce α-Synuclein Fibrillation and Oligomer Toxicity. Journal of Biological Chemistry 2019, 294, 4215–4232. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; Hou, C.; Yang, L.; Li, H.; Guo, J.; Huo, C.; Wang, M.; Miao, Y.; Liu, J.; Kang, Y. ; Oleuropein Improves Mitochondrial Function to Attenuate Oxidative Stress by Activating the Nrf2 Pathway in the Hypothalamic Paraventricular Nucleus of Spontaneously Hypertensive Rats. Neuropharmacology 2017, 113, 556–566. [Google Scholar] [CrossRef]
- Lange, K.W.; Li, S. Resveratrol, Pterostilbene, and Dementia. BioFactors 2018, 44, 83–90. [Google Scholar] [CrossRef]
- Ader, P. Bioavailability and Metabolism of the Flavonol Quercetin in the Pig. Free Radic Biol Med 2000, 28, 1056–1067. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; Abete, P. ; Oxidative Stress, Aging, and Diseases. Clin Interv Aging 2018, Volume 13, 757–772. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef]
- Zheng, Q.; Kebede, M.T.; Kemeh, M.M.; Islam, S.; Lee, B.; Bleck, S.D.; Wurfl, L.A.; Lazo, N.D. Inhibition of the Self-Assembly of Aβ and of Tau by Polyphenols: Mechanistic Studies. Molecules 2019, 24, 2316. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, B.N.; Parylak, S.L.; Gage, F.H. Mechanisms of Dietary Flavonoid Action in Neuronal Function and Neuroinflammation. Mol Aspects Med 2018, 61, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kishimoto, Y.; Grammatikakis, I.; Gottimukkala, K.; Cutler, R.G.; Zhang, S.; Abdelmohsen, K.; Bohr, V.A.; Misra Sen, J.; Gorospe, M.; et al. Senolytic Therapy Alleviates Aβ-Associated Oligodendrocyte Progenitor Cell Senescence and Cognitive Deficits in an Alzheimer’s Disease Model. Nat Neurosci 2019, 22, 719–728. [Google Scholar] [CrossRef]
- Van houcke, J.; Mariën, V.; Zandecki, C.; Ayana, R.; Pepermans, E.; Boonen, K.; Seuntjens, E.; Baggerman, G.; Arckens, L. A Short Dasatinib and Quercetin Treatment Is Sufficient to Reinstate Potent Adult Neuroregenesis in the Aged Killifish. NPJ Regen Med 2023, 8. [Google Scholar] [CrossRef]
- Currais, A.; Farrokhi, C.; Dargusch, R.; Armando, A.; Quehenberger, O.; Schubert, D.; Maher, P. Fisetin Reduces the Impact of Aging on Behavior and Physiology in the Rapidly Aging SAMP8 Mouse. Journals of Gerontology - Series A Biological Sciences and Medical Sciences 2018, 73, 299–307. [Google Scholar] [CrossRef]
- Wissler Gerdes, E.O.; Misra, A.; Netto, J.M.E.; Tchkonia, T.; Kirkland, J.L. Strategies for Late Phase Preclinical and Early Clinical Trials of Senolytics. Mech Ageing Dev 2021, 200. [Google Scholar] [CrossRef]
- Song, S.; Lam, E.W.F.; Tchkonia, T.; Kirkland, J.L.; Sun, Y. Senescent Cells: Emerging Targets for Human Aging and Age-Related Diseases. Trends Biochem Sci 2020, 45, 578–592. [Google Scholar] [CrossRef]
- Hartman, R.E.; Shah, A.; Fagan, A.M.; Schwetye, K.E.; Parsadanian, M.; Schulman, R.N.; Finn, M.B.; Holtzman, D.M. Pomegranate Juice Decreases Amyloid Load and Improves Behavior in a Mouse Model of Alzheimer’s Disease. Neurobiol Dis 2006, 24, 506–515. [Google Scholar] [CrossRef]
- Ho, L.; Chen, L.H.; Wang, J.; Zhao, W.; Talcott, S.T.; Ono, K.; Teplow, D.; Humala, N.; Cheng, A.; Percival, S.S.; Ferruzzi, M.; Janle, E.; Dickstein, D.L.; Pasinetti, G.M. ; Heterogeneity in Red Wine Polyphenolic Contents Differentially Influences Alzheimer’s Disease-Type Neuropathology and Cognitive Deterioration. Journal of Alzheimer’s Disease 2009, 16, 59–72. [Google Scholar] [CrossRef]
- Mori, T.; Rezai-Zadeh, K.; Koyama, N.; Arendash, G.W.; Yamaguchi, H.; Kakuda, N.; Horikoshi-Sakuraba, Y.; Tan, J.; Town, T. Tannic Acid Is a Natural β-Secretase Inhibitor That Prevents Cognitive Impairment and Mitigates Alzheimer-like Pathology in Transgenic Mice. Journal of Biological Chemistry 2012, 287, 6912–6927. [Google Scholar] [CrossRef]
- Wang, J.; Ho, L.; Zhao, W.; Ono, K.; Rosensweig, C.; Chen, L.; Humala, N.; Teplow, D.B.; Pasinetti, G.M. Grape-Derived Polyphenolics Prevent A Oligomerization and Attenuate Cognitive Deterioration in a Mouse Model of Alzheimer’s Disease. Journal of Neuroscience 2008, 28, 6388–6392. [Google Scholar] [CrossRef] [PubMed]
- Nugraheni, N.; Ahlina, F.N.; Salsabila, I.A.; Haryanti, S.; Meiyanto, E. Anti-Senescence Activity of Indonesian Black Pepper Essential Oil (Piper Nigrum L.) on Ovarian CHO-K1 and Fibroblast NIH-3T3 Cells; Vol. 45.
- Chilelli, N.; Ragazzi, E.; Valentini, R.; Cosma, C.; Ferraresso, S.; Lapolla, A.; Sartore, G. Curcumin and Boswellia Serrata Modulate the Glyco-Oxidative Status and Lipo-Oxidation in Master Athletes. Nutrients 2016, 8, 745. [Google Scholar] [CrossRef] [PubMed]
- Karimian, M.S.; Pirro, M.; Majeed, M.; Sahebkar, A. Curcumin as a Natural Regulator of Monocyte Chemoattractant Protein-1. Cytokine Growth Factor Rev 2017, 33, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, W.; Suszek, M.; Wnuk, M.; Lewinska, A.; Wasiak, E.; Sikora, E.; Bielak-Zmijewska, A. Curcumin Elevates Sirtuin Level but Does Not Postpone in vitro Senescence of Human Cells Building the Vasculature. Oncotarget 2016, 7, 19201–19213. [Google Scholar] [CrossRef]
- Taka, T.; Changtam, C.; Thaichana, P.; Kaewtunjai, N.; Suksamrarn, A.; Lee, T.R.; Tuntiwechapikul, W. Curcuminoid Derivatives Enhance Telomerase Activity in an in vitro TRAP Assay. Bioorg Med Chem Lett 2014, 24, 5242–5246. [Google Scholar] [CrossRef]
- Kim, T.; Davis, J.; Zhang, A.J.; He, X.; Mathews, S.T. Curcumin Activates AMPK and Suppresses Gluconeogenic Gene Expression in Hepatoma Cells. Biochem Biophys Res Commun 2009, 388, 377–382. [Google Scholar] [CrossRef]
- Liu, Z.; Cui, C.; Xu, P.; Dang, R.; Cai, H.; Liao, D.; Yang, M.; Feng, Q.; Yan, X.; Jiang, P. Curcumin Activates AMPK Pathway and Regulates Lipid Metabolism in Rats Following Prolonged Clozapine Exposure. Front Neurosci 2017, 11. [Google Scholar] [CrossRef]
- Ray Hamidie, R.D.; Yamada, T.; Ishizawa, R.; Saito, Y.; Masuda, K. Curcumin Treatment Enhances the Effect of Exercise on Mitochondrial Biogenesis in Skeletal Muscle by Increasing CAMP Levels. Metabolism 2015, 64, 1334–1347. [Google Scholar] [CrossRef]
- Juturu, V.; Sahin, K.; Pala, R.; Tuzcu, M.; Ozdemir, O.; Orhan, C.; Sahin, N. Curcumin Prevents Muscle Damage by Regulating NF-KB and Nrf2 Pathways and Improves Performance: An in vivo Model. J Inflamm Res 2016, Volume 9, 147–154. [Google Scholar] [CrossRef]
- Namirah, I.; Wimbanu, K.S.; Rompies, A.M.E.; Prayogo, Y.S.; Arozal, W.; Fadilah, F.; Hanafi, M.; Hardiany, N.S. The Effect of Ethanol-Based Coriander (Coriandrum Sativum L.) Seed Extract on Oxidative Stress, Antioxidant Level and Cellular Senescence in the Heart of Obese Rat. J Pharm Pharmacogn Res 2024, 12, 1111–1120. [Google Scholar] [CrossRef]
- Hardiany, N.S.; Dewi, P.K.K.; Dewi, S.; Tejo, B.A. Exploration of Neuroprotective Effect from Coriandrum Sativum L. Ethanolic Seeds Extracts on Brain of Obese Rats. Sci Rep 2024, 14, 603. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-Y.; Cheng, Y.; Cui, J.; Zhang, Y.; Yu, L.-C. Potential Protection of Curcumin against Amyloid β-Induced Toxicity on Cultured Rat Prefrontal Cortical Neurons. Neurosci Lett 2009, 463, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Muthian, G.; Mackey, V.; Prasad, K.; Charlton, C. Curcumin and an Antioxidant Formulation Protect C57BL/6J Mice from MPTP-Induced Parkinson’s Disease like Changes: Potential Neuroprotection for Neurodegeneration. Journal of Parkinsonism and Restless Legs Syndrome 2018, Volume 8, 49–59. [Google Scholar] [CrossRef]
- Canistro, D.; Chiavaroli, A.; Cicia, D.; Cimino, F.; Curro, D.; Dell Agli, M.; Ferrante, C.; Giovannelli, L.; Leone, S.; Martinelli, G.; Milella, L.; Pagano, E.; Piazza, S.; Ponticelli, M.; Recinella, L.; Ristori, S.; Sangiovanni, E.; Smeriglio, A.; Speciale, A.; Trombetta, D.; Vivarelli, F. ; The Pharmacological Basis of the Curcumin Nutraceutical Uses: An Update. Pharmadvances 2021, 03, 421. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Karlsen, J.; Henegouwen, G.B. Studies on Curcumin and Curcuminoids VIII. Photochemical Stability of Curcumin. Zeitschrift für Lebensmittel-Untersuchung und -Forschung 1986, 183, 116–122. [Google Scholar] [CrossRef]
- Zbarsky, V.; Datla, K.P.; Parkar, S.; Rai, D.K.; Aruoma, O.I.; Dexter, D.T. Neuroprotective Properties of the Natural Phenolic Antioxidants Curcumin and Naringenin but Not Quercetin and Fisetin in a 6-OHDA Model of Parkinson’s Disease. Free Radic Res 2005, 39, 1119–1125. [Google Scholar] [CrossRef]
- Wang, J.; Du, X.-X.; Jiang, H.; Xie, J.-X. Curcumin Attenuates 6-Hydroxydopamine-Induced Cytotoxicity by Anti-Oxidation and Nuclear Factor-KappaB Modulation in MES23.5 Cells. Biochem Pharmacol 2009, 78, 178–183. [Google Scholar] [CrossRef]
- Huang, W.-T.; Niu, K.-C.; Chang, C.-K.; Lin, M.-T.; Chang, C.-P. Curcumin Inhibits the Increase of Glutamate, Hydroxyl Radicals and PGE2 in the Hypothalamus and Reduces Fever during LPS-Induced Systemic Inflammation in Rabbits. Eur J Pharmacol 2008, 593, 105–111. [Google Scholar] [CrossRef]
- Yang, W.; Chen, Y.-H.; Liu, H.; Qu, H.-D. Neuroprotective Effects of Piperine on the 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Parkinson’s Disease Mouse Model. Int J Mol Med 2015, 36, 1369–1376. [Google Scholar] [CrossRef]
- Chaudhri, S.K.; Jain, S. A Systematic Review of Piperine as a Bioavailability Enhancer. Journal of Drug Delivery and Therapeutics 2023, 13, 133–136. [Google Scholar] [CrossRef]
- Singh, S.; Jamwal, S.; Kumar, P. Neuroprotective Potential of Quercetin in Combination with Piperine against 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Neurotoxicity. Neural Regen Res 2017, 12, 1137. [Google Scholar] [CrossRef] [PubMed]
- Bae, W.-Y.; Choi, J.-S.; Jeong, J.-W. The Neuroprotective Effects of Cinnamic Aldehyde in an MPTP Mouse Model of Parkinson’s Disease. Int J Mol Sci 2018, 19, 551. [Google Scholar] [CrossRef] [PubMed]
- Ramazani, E.; YazdFazeli, M.; Emami, S.A.; Mohtashami, L.; Javadi, B.; Asili, J.; Tayarani-Najaran, Z. Protective Effects of Cinnamomum Verum, Cinnamomum Cassia and Cinnamaldehyde against 6-OHDA-Induced Apoptosis in PC12 Cells. Mol Biol Rep 2020, 47, 2437–2445. [Google Scholar] [CrossRef]
- Shaltiel-Karyo, R.; Davidi, D.; Frenkel-Pinter, M.; Ovadia, M.; Segal, D.; Gazit, E. Differential Inhibition of α-Synuclein Oligomeric and Fibrillar Assembly in Parkinson’s Disease Model by Cinnamon Extract. Biochimica et Biophysica Acta (BBA) - General Subjects 2012, 1820, 1628–1635. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; Yousef, M.I.; Radwan, F.M.E. Ameliorating Effect of Curcumin on Sodium Arsenite-Induced Oxidative Damage and Lipid Peroxidation in Different Rat Organs. Food and Chemical Toxicology 2009, 47, 249–254. [Google Scholar] [CrossRef]
- Venkatesan, N.; Punithavathi, D.; Arumugam, V. Curcumin Prevents Adriamycin Nephrotoxicity in Rats. Br J Pharmacol 2000, 129, 231–234. [Google Scholar] [CrossRef]
- Strimpakos, A.S.; Sharma, R.A. Curcumin: Preventive and Therapeutic Properties in Laboratory Studies and Clinical Trials. Antioxid Redox Signal 2008, 10, 511–546. [Google Scholar] [CrossRef]
- Ganguli, M.; Chandra, V.; Kamboh, M.I.; Johnston, J.M.; Dodge, H.H.; Thelma, B.K.; Juyal, R.C.; Pandav, R.; Belle, S.H.; DeKosky, S.T. Apolipoprotein E Polymorphism and Alzheimer Disease. Arch Neurol 2000, 57, 824. [Google Scholar] [CrossRef]
- Ng, T.-P.; Chiam, P.-C.; Lee, T.; Chua, H.-C.; Lim, L.; Kua, E.-H. Curry Consumption and Cognitive Function in the Elderly. Am J Epidemiol 2006, 164, 898–906. [Google Scholar] [CrossRef]
- Zhang, L.; Fiala, M.; Cashman, J.; Sayre, J.; Espinosa, A.; Mahanian, M.; Zaghi, J.; Badmaev, V.; Graves, M.C.; Bernard, G.; et al. Curcuminoids Enhance Amyloid-β Uptake by Macrophages of Alzheimer’s Disease Patients. Journal of Alzheimer’s Disease 2006, 10, 1–7. [Google Scholar] [CrossRef]
- Ambegaokar, S.S.; Wu, L.; Alamshahi, K.; Lau, J.; Jazayeri, L.; Chan, S.; Khanna, P.; Hsieh, E.; Timiras, P.S. Curcumin Inhibits Dose-Dependently and Time-Dependently Neuroglial Cell Proliferation and Growth. Neuro Endocrinol Lett 2003, 24, 469–473. [Google Scholar] [PubMed]
- Kim, G.-Y.; Kim, K.-H.; Lee, S.-H.; Yoon, M.-S.; Lee, H.-J.; Moon, D.-O.; Lee, C.-M.; Ahn, S.-C.; Park, Y.C.; Park, Y.-M. Curcumin Inhibits Immunostimulatory Function of Dendritic Cells: MAPKs and Translocation of NF-ΚB as Potential Targets. The Journal of Immunology 2005, 174, 8116–8124. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Kim, D.S.H.L. Discovery of Natural Products from Curcuma l Onga That Protect Cells from Beta-Amyloid Insult: A Drug Discovery Effort against Alzheimer’s Disease. J Nat Prod 2002, 65, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.H.L.; Park, S.-Y.; Kim, J.-Y. Curcuminoids from Curcuma Longa L. (Zingiberaceae) That Protect PC12 Rat Pheochromocytoma and Normal Human Umbilical Vein Endothelial Cells from ΒA(1–42) Insult. Neurosci Lett 2001, 303, 57–61. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, W.; Sun, Y.J.; Hu, M.; Li, F.; Zhu, D.Y. Neuroprotective Effect of Curcumin on Focal Cerebral Ischemic Rats by Preventing Blood–Brain Barrier Damage. Eur J Pharmacol 2007, 561, 54–62. [Google Scholar] [CrossRef]
- Liu, H.; Li, Z.; Qiu, D.; Gu, Q.; Lei, Q.; Mao, L. The Inhibitory Effects of Different Curcuminoids on β-Amyloid Protein, β-Amyloid Precursor Protein and β-Site Amyloid Precursor Protein Cleaving Enzyme 1 in SwAPP HEK293 Cells. Neurosci Lett 2010, 485, 83–88. [Google Scholar] [CrossRef]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin Has Potent Anti-amyloidogenic Effects for Alzheimer’s Β-amyloid Fibrils in vitro. J Neurosci Res 2004, 75, 742–750. [Google Scholar] [CrossRef]
- Pagliari, S.; Forcella, M.; Lonati, E.; Sacco, G.; Romaniello, F.; Rovellini, P.; Fusi, P.; Palestini, P.; Campone, L.; Labra, M.; Bulbarelli, A.; Bruni, I.; Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum Verum J. Presl) Bark Extract after In vitro Digestion Simulation. Foods 2023, 12, 452. [CrossRef]
- Moselhy, S.S.; Ali, H.K.H. Hepatoprotective Effect of Cinnamon Extracts against Carbon Tetrachloride Induced Oxidative Stress and Liver Injury in Rats. Biol Res 2009, 42, 93–98. [Google Scholar] [CrossRef]
- Gunawardena, D.; Karunaweera, N.; Lee, S.; van Der Kooy, F.; Harman, D.G.; Raju, R.; Bennett, L.; Gyengesi, E.; Sucher, N.J.; Münch, G. Anti-Inflammatory Activity of Cinnamon (C. Zeylanicum and C. Cassia) Extracts – Identification of E-Cinnamaldehyde and o-Methoxy Cinnamaldehyde as the Most Potent Bioactive Compounds. Food Funct 2015, 6, 910–919. [Google Scholar] [CrossRef]
- Schink, A.; Naumoska, K.; Kitanovski, Z.; Kampf, C.J.; Fröhlich-Nowoisky, J.; Thines, E.; Pöschl, U.; Schuppan, D.; Lucas, K. Anti-Inflammatory Effects of Cinnamon Extract and Identification of Active Compounds Influencing the TLR2 and TLR4 Signaling Pathways. Food Funct 2018, 9, 5950–5964. [Google Scholar] [CrossRef] [PubMed]
- Frydman-Marom, A.; Levin, A.; Farfara, D.; Benromano, T.; Scherzer-Attali, R.; Peled, S.; Vassar, R.; Segal, D.; Gazit, E.; Frenkel, D.; Ovadia, M. ; Orally Administrated Cinnamon Extract Reduces β-Amyloid Oligomerization and Corrects Cognitive Impairment in Alzheimer’s Disease Animal Models. PLoS One 2011, 6, e16564. [Google Scholar] [CrossRef] [PubMed]
- Chonpathompikunlert, P.; Wattanathorn, J.; Muchimapura, S. Piperine, the Main Alkaloid of Thai Black Pepper, Protects against Neurodegeneration and Cognitive Impairment in Animal Model of Cognitive Deficit like Condition of Alzheimer’s Disease. Food and Chemical Toxicology 2010, 48, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Subedee, L. Preventive Role of Indian Black Pepper in Animal Models of Alzheimer’s Disease. Journal of Clinical And Diagnostic Research 2015. [Google Scholar] [CrossRef]
- Auti, S.T.; Kulkarni, Y.A. Neuroprotective Effect of Cardamom Oil Against Aluminum Induced Neurotoxicity in Rats. Front Neurol 2019, 10. [Google Scholar] [CrossRef]
- Shi, H.; Ge, X.; Ma, X.; Zheng, M.; Cui, X.; Pan, W.; Zheng, P.; Yang, X.; Zhang, P.; Hu, M.; Tang, R.; Zheng, K.; Huang, X.; Yu, Y. ; A Fiber-Deprived Diet Causes Cognitive Impairment and Hippocampal Microglia-Mediated Synaptic Loss through the Gut Microbiota and Metabolites. Microbiome 2021, 9, 1–20. [Google Scholar] [CrossRef]
- Gareau, M.G.; Gareau, M.G.; Lyte, M.; Cryan, J.F. Microbiota-Gut-Brain Axis and Cognitive Function Abbreviations 5-HT Serotonin ANS Autonomic Nervous System BDNF Brain Derived Neurotropic Factor CD Crohn’s Disease CREB CAMP Response Element Binding Protein CRF Corticotrophin-Releasing Factor DA Dopamine. Adv Exp Med Biol 2014, 817, 357–371. [Google Scholar] [CrossRef]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.H. Short Chain Fatty Acids and Gut Microbiota Differ between Patients with Parkinson’s Disease and Age-Matched Controls. Parkinsonism Relat Disord 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Matt, S.M.; Allen, J.M.; Lawson, M.A.; Mailing, L.J.; Woods, J.A.; Johnson, R.W. Butyrate and Dietary Soluble Fiber Improve Neuroinflammation Associated with Aging in Mice. Front Immunol 2018, 9. [Google Scholar] [CrossRef]
- Martins, I.J.; Fernando, W.M.A.D.B. High Fibre Diets and Alzheimer’s Disease. Food Nutr Sci 2014, 05, 410–424. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J Lipid Res 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Zilberter, Y.; Zilberter, M. The Vicious Circle of Hypometabolism in Neurodegenerative Diseases: Ways and Mechanisms of Metabolic Correction. J Neurosci Res 2017, 95, 2217–2235. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; Schwierzeck, V.; Utermöhlen, O.; Chun, E.; Garrett, W.S.; McCoy, K.D.; Diefenbach, A.; Staeheli, P.; Stecher, B.; Amit, I.; Prinz, M. ; Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat Neurosci 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Ho, L.; Ono, K.; Tsuji, M.; Mazzola, P.; Singh, R.; Pasinetti, G.M. Protective Roles of Intestinal Microbiota Derived Short Chain Fatty Acids in Alzheimer’s Disease-Type Beta-Amyloid Neuropathological Mechanisms. Expert Rev Neurother 2018, 18, 83–90. [Google Scholar] [CrossRef]
- Zhou, Y.; Xie, L.; Schröder, J.; Schuster, I.S.; Nakai, M.; Sun, G.; Sun, Y.B.Y.; Mariño, E.; Degli-Esposti, M.A.; Marques, F.Z.; Grubman, A.; Polo, J.M.; Mackay, C.R. ; Dietary Fiber and Microbiota Metabolite Receptors Enhance Cognition and Alleviate Disease in the 5xFAD Mouse Model of Alzheimer’s Disease. Journal of Neuroscience 2023, 43, 6460–6475. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus Plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front Microbiol 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Yi, H.; Wang, L.; Xiong, Y.; Wang, Z.; Qiu, Y.; Wen, X.; Jiang, Z.; Yang, X.; Ma, X. Lactobacillus Reuteri LR1 Improved Expression of Genes of Tight Junction Proteins via the MLCK Pathway in IPEC-1 Cells during Infection with Enterotoxigenic Escherichia Coli K88. Mediators Inflamm 2018, 2018. [Google Scholar] [CrossRef]
- Perdigón, G.; Maldonado Galdeano, C.; Valdez, J.C.; Medici, M. Interaction of Lactic Acid Bacteria with the Gut Immune System. Eur J Clin Nutr 2002, 56, S21–S26. [Google Scholar] [CrossRef]
- Sichetti, M.; De Marco, S.; Pagiotti, R.; Traina, G.; Pietrella, D.; Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L.; et al. Effect of Lactobacillus Rhamnosus HN001 and Bifidobacterium Longum BB536 on the Healthy Gut Microbiota Composition at Phyla and Species Level: A Preliminary Study. Front Microbiol 2019, 10, S21–S26. [Google Scholar] [CrossRef]
- Marotta, A.; Sarno, E.; Casale, A. Del; Pane, M.; Mogna, L.; Amoruso, A.; Felis, G.E.; Fiorio, M. Effects of Probiotics on Cognitive Reactivity, Mood, and Sleep Quality. Front Psychiatry 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Sharma, R.; Padwad, Y. Probiotic Bacteria as Modulators of Cellular Senescence: Emerging Concepts and Opportunities. Gut Microbes 2020, 11, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus Rhamnosus HN001 and Bifidobacterium Longum BB536 on the Healthy Gut Microbiota Composition at Phyla and Species Level: A Preliminary Study. World J Gastroenterol 2017, 23, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health Benefits of Probiotics: A Review. ISRN Nutr 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Azad, F.J.; Talaei, A.; Rafatpanah, H.; Yousefzadeh, H.; Jafari, R.; Jafari, M.; Talaei, A.; Hosseini, R.F.; Jabari, F. Copyright© Autumn; 2014; Vol. 13.
- Woo, J.Y.; Gu, W.; Kim, K.A.; Jang, S.E.; Han, M.J.; Kim, D.H. Lactobacillus Pentosus Var. Plantarum C29 Ameliorates Memory Impairment and Inflammaging in a d-Galactose-Induced Accelerated Aging Mouse Model. Anaerobe 2014, 27, 22–26. [Google Scholar] [CrossRef]
- Grompone, G.; Martorell, P.; Llopis, S.; González, N.; Genovés, S.; Mulet, A.P.; Fernández-Calero, T.; Tiscornia, I.; Bollati-Fogolín, M.; Chambaud, I.; Foligné, B.; Montserrat, A.; Ramón, D. ; Anti-Inflammatory Lactobacillus Rhamnosus CNCM I-3690 Strain Protects against Oxidative Stress and Increases Lifespan in Caenorhabditis Elegans. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Lin, X.; Xia, Y.; Wang, G.; Xiong, Z.; Zhang, H.; Lai, F.; Ai, L. Lactobacillus Plantarum AR501 Alleviates the Oxidative Stress of D-Galactose-Induced Aging Mice Liver by Upregulation of Nrf2-Mediated Antioxidant Enzyme Expression. J Food Sci 2018, 83, 1990–1998. [Google Scholar] [CrossRef]
- Eifler, N.; Vetsch, M.; Gregorini, M.; Ringler, P.; Chami, M.; Philippsen, A.; Fritz, A.; Müller, S.A.; Glockshuber, R.; Engel, A.; et al. Cytotoxin ClyA from Escherichia Coli Assembles to a 13-Meric Pore Independent of Its Redox-State. EMBO Journal 2006, 25, 2652–2661. [Google Scholar] [CrossRef]
- Gazerani, P. Probiotics for Parkinson’s Disease. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Magistrelli, L.; Amoruso, A.; Mogna, L.; Graziano, T.; Cantello, R.; Pane, M.; Comi, C. Probiotics May Have Beneficial Effects in Parkinson’s Disease: In vitro Evidence. Front Immunol 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Castelli, V.; D’Angelo, M.; Lombardi, F.; Alfonsetti, M.; Antonosante, A.; Catanesi, M.; Benedetti, E.; Palumbo, P.; Cifone, M.G.; Giordano, A.; Desideri, G.; Cimini, A. ; Effects of the Probiotic Formulation SLAB51 in in vitro and in vivo Parkinson’s Disease Models. Aging 2020, 12, 4641–4659. [Google Scholar] [CrossRef]
- Liao, J.F.; Cheng, Y.F.; You, S.T.; Kuo, W.C.; Huang, C.W.; Chiou, J.J.; Hsu, C.C.; Hsieh-Li, H.M.; Wang, S.; Tsai, Y.C. Lactobacillus Plantarum PS128 Alleviates Neurodegenerative Progression in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Mouse Models of Parkinson’s Disease. Brain Behav Immun 2020, 90, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Srivastav, S.; Neupane, S.; Bhurtel, S.; Katila, N.; Maharjan, S.; Choi, H.; Hong, J.T.; Choi, D.Y. Probiotics Mixture Increases Butyrate, and Subsequently Rescues the Nigral Dopaminergic Neurons from MPTP and Rotenone-Induced Neurotoxicity. Journal of Nutritional Biochemistry 2019, 69, 73–86. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, C.; Li, J. V.; Marchesi, J.R.; Plummer, S.; Garaiova, I.; Good, M.A. Long-Term Multi-Species Lactobacillus and Bifidobacterium Dietary Supplement Enhances Memory and Changes Regional Brain Metabolites in Middle-Aged Rats. Neurobiol Learn Mem 2017, 144, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Distrutti, E.; O’Reilly, J.A.; McDonald, C.; Cipriani, S.; Renga, B.; Lynch, M.A.; Fiorucci, S. Modulation of Intestinal Microbiota by the Probiotic VSL#3 Resets Brain Gene Expression and Ameliorates the Age-Related Deficit in LTP. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Jeong, J.J.; Woo, J.Y.; Kim, K.A.; Han, M.J.; Kim, D.H. Lactobacillus Pentosus Var. Plantarum C29 Ameliorates Age-Dependent Memory Impairment in Fischer 344 Rats. Lett Appl Microbiol 2015, 60, 307–314. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Sugahara, H.; Shimada, K.; Mitsuyama, E.; Kuhara, T.; Yasuoka, A.; Kondo, T.; Abe, K.; Xiao, J.Z. Therapeutic Potential of Bifidobacterium Breve Strain A1 for Preventing Cognitive Impairment in Alzheimer’s Disease. Sci Rep 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Chandra, S.; Sisodia, S.S.; Vassar, R.J. The Gut Microbiome in Alzheimer’s Disease: What We Know and What Remains to Be Explored. Mol Neurodegener 2023, 18, 1–21. [Google Scholar] [CrossRef]
- Mossello, E.; Ballini, E.; Boncinelli, M.; Monami, M.; Lonetto, G.; Mello, A.M.; Tarantini, F.; Baldasseroni, S.; Mannucci, E.; Marchionni, N. Glucagon-like Peptide-1, Diabetes, and Cognitive Decline: Possible Pathophysiological Links and Therapeutic Opportunities. Exp Diabetes Res 2011, 2011. [Google Scholar] [CrossRef]
- Franco-Robles, E.; López, M.G. Implication of Fructans in Health: Immunomodulatory and Antioxidant Mechanisms. Scientific World Journal 2015, 2015. [Google Scholar] [CrossRef]
- Hall, D.A.; Voigt, R.M.; Cantu-Jungles, T.M.; Hamaker, B.; Engen, P.A.; Shaikh, M.; Raeisi, S.; Green, S.J.; Naqib, A.; Forsyth, C.B.; Chen, T.; Manfready, R.; Ouyang, B.; Rasmussen, H.E.; Sedghi, S.; Goetz, C.G.; Keshavarzian, A. ; An Open Label, Non-Randomized Study Assessing a Prebiotic Fiber Intervention in a Small Cohort of Parkinson’s Disease Participants. Nat Commun 2023, 14. [Google Scholar] [CrossRef]
- Becker, A.; Schmartz, G.P.; Gröger, L.; Grammes, N.; Galata, V.; Philippeit, H.; Weiland, J.; Ludwig, N.; Meese, E.; Tierling, S.; Walter, J.; Schwiertz, A.; Spiegel, J.; Wagenpfeil, G.; Faßbender, K.; Keller, A.; Unger, M.M. ; Effects of Resistant Starch on Symptoms, Fecal Markers, and Gut Microbiota in Parkinson’s Disease — The RESISTA-PD Trial. Genomics Proteomics Bioinformatics 2022, 20, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Barichella, M.; Pacchetti, C.; Bolliri, C.; Cassani, E.; Iorio, L.; Pusani, C.; Pinelli, G.; Privitera, G.; Cesari, I.; Faierman, S.A.; Caccialanza, R.; Pezzoli, G.; Cereda, E. ; Probiotics and Prebiotic Fiber for Constipation Associated with Parkinson Disease. Neurology 2016, 87, 1274–1280. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Raja Ali, R.A.; Abdul Manaf, M.R.; Ahmad, N.; Tajurruddin, F.W.; Qin, W.Z.; Md Desa, S.H.; Ibrahim, N.M. Multi-Strain Probiotics (Hexbio) Containing MCP BCMC Strains Improved Constipation and Gut Motility in Parkinson’s Disease: A Randomised Controlled Trial. PLoS One 2020, 15, 1–17. [Google Scholar] [CrossRef]
- Chen, D.; Yang, X.; Yang, J.; Lai, G.; Yong, T.; Tang, X.; Shuai, O.; Zhou, G.; Xie, Y.; Wu, Q. Prebiotic Effect of Fructooligosaccharides from Morinda Officinalis on Alzheimer’s Disease in Rodent Models by Targeting the Microbiota-Gut-Brain Axis. Front Aging Neurosci 2017, 9, 1–28. [Google Scholar] [CrossRef]
- Sarparast, M.; Dattmore, D.; Alan, J.; Lee, K.S.S. Cytochrome P450 Metabolism of Polyunsaturated Fatty Acids and Neurodegeneration. Nutrients 2020, 12, 1–40. [Google Scholar] [CrossRef]
- Şimşek, H.; Uçar, A. Polyunsaturated Fatty Acids as a Nutraceutical for Age-Related Neurodegenerative Diseases: Current Knowledge and Future Directions. Clinical Nutrition Open Science 2024, 56, 65–73. [Google Scholar] [CrossRef]
- Luchtman, D.W.; Song, C. Cognitive Enhancement by Omega-3 Fatty Acids from Child-Hood to Old Age: Findings from Animal and Clinical Studies. Neuropharmacology 2013, 64, 550–565. [Google Scholar] [CrossRef]
- Youdim, K.A.; Martin, A.; Joseph, J.A. Essential Fatty Acids and the Brain: Possible Health Implications. International Journal of Developmental Neuroscience 2000, 18, 383–399. [Google Scholar] [CrossRef]
- Joffre, C.; Dinel, A.L.; Chataigner, M.; Pallet, V.; Layé, S. N-3 Polyunsaturated Fatty Acids and Their Derivates Reduce Neuroinflammation during Aging. Nutrients 2020, 12, 1–25. [Google Scholar] [CrossRef]
- Uauy, R.; Dangour, A.D. Nutrition in Brain Development and Aging: Role of Essential Fatty Acids. Nutr Rev 2006, 64. [Google Scholar] [CrossRef]
- Kerdiles, O.; Layé, S.; Calon, F. Omega-3 Polyunsaturated Fatty Acids and Brain Health: Preclinical Evidence for the Prevention of Neurodegenerative Diseases. Trends Food Sci Technol 2017, 69, 203–213. [Google Scholar] [CrossRef]
- Singh, P.K.; Gupta, M.K.; Nath, R. Omega-3 Fatty Acid as a Protectant in Lead-Induced Neurotoxicity; Elsevier Inc., 2023; ISBN 9780323900522.
- Bousquet, M.; Saint-Pierre, M.; Julien, C.; Salem, N.; Cicchetti, F.; Calon, F. Beneficial Effects of Dietary Omega-3 Polyunsaturated Fatty Acid on Toxin-induced Neuronal Degeneration in an Animal Model of Parkinson’s Disease. The FASEB Journal 2008, 22, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, H.; Fung, T.T.; Logroscino, G.; Schwarzschild, M.A.; Hu, F.B.; Ascherio, A. Prospective Study of Dietary Pattern and Risk of Parkinson Disease. American Journal of Clinical Nutrition 2007, 86, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- De Lau, L.M.L.; Bornebroek, M.; Witteman, J.C.M.; Hofman, A.; Koudstaal, P.J.; Breteler, M.M.B. Dietary Fatty Acids and the Risk of Parkinson Disease: The Rotterdam Study. Neurology 2005, 64, 2040–2045. [Google Scholar] [CrossRef]
- Kamel, F.; Goldman, S.M.; Umbach, D.M.; Chen, H.; Richardson, G.; Barber, M.R.; Meng, C.; Marras, C.; Korell, M.; Kasten, M.; et al. Dietary Fat Intake, Pesticide Use, and Parkinson’s Disease. Parkinsonism Relat Disord 2014, 20, 82–87. [Google Scholar] [CrossRef]
- Bousquet, M.; St-Amour, I.; Vandal, M.; Julien, P.; Cicchetti, F.; Calon, F. High-Fat Diet Exacerbates MPTP-Induced Dopaminergic Degeneration in Mice. Neurobiol Dis 2012, 45, 529–538. [Google Scholar] [CrossRef]
- Bousquet, M.; Gibrat, C.; Saint-Pierre, M.; Julien, C.; Calon, F.; Cicchetti, F. Modulation of Brain-Derived Neurotrophic Factor as a Potential Neuroprotective Mechanism of Action of Omega-3 Fatty Acids in a Parkinsonian Animal Model. Prog Neuropsychopharmacol Biol Psychiatry 2009, 33, 1401–1408. [Google Scholar] [CrossRef]
- Bousquet, M.; Calon, F.; Cicchetti, F. Impact of Omega-3 Fatty Acids in Parkinson’s Disease. Ageing Res Rev 2011, 10, 453–463. [Google Scholar] [CrossRef]
- Calon, F.; Cicchetti, F. Can We Prevent Parkinson’s Disease with n-3 Polyunsaturated Fatty Acids? Future Lipidol 2008, 3, 133–137. [Google Scholar] [CrossRef]
- Morgese, M.G.; Schiavone, S.; Bove, M.; Colia, A.L.; Dimonte, S.; Tucci, P.; Trabace, L. N-3 PUFA Prevent Oxidative Stress in a Rat Model of Beta-Amyloid-Induced Toxicity. Pharmaceuticals 2021, 14. [Google Scholar] [CrossRef]
- Olde Rikkert, M.G.M.; Verhey, F.R.; Blesa, R.; Von Arnim, C.A.F.; Bongers, A.; Harrison, J.; Sijben, J.; Scarpini, E.; Vandewoude, M.F.J.; Vellas, B.; et al. Tolerability and Safety of Souvenaid in Patients with Mild Alzheimer’s Disease: Results of Multi-Center, 24-Week, Open-Label Extension Study. Journal of Alzheimer’s Disease 2015, 44, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Faxén-Irving, G.; Freund-Levi, Y.; Eriksdotter-Jönhagen, M.; Basun, H.; Hjorth, E.; Palmblad, J.; Vedin, I.; Cederholm, T.; Wahlund, L.O. Effects on Transthyretin in Plasma and Cerebrospinal Fluid by Dha-Rich n-3 Fatty Acid Supplementation in Patients with Alzheimer’s Disease: The Omegad Study. Journal of Alzheimer’s Disease 2013, 36, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Lim, G.P.; Yang, F.; Morihara, T.; Teter, B.; Ubeda, O.; Rostaing, P.; Triller, A.; Salem, N.; Ashe, K.H.; et al. Docosahexaenoic Acid Protects from Dendritic Pathology in an Alzheimer’s Disease Mouse Model. Neuron 2004, 43, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Lim, G.P.; Morihara, T.; Yang, F.; Ubeda, O.; Salem, N.; Frautschy, S.A.; Cole, G.M. Dietary N-3 Polyunsaturated Fatty Acid Depletion Activates Caspases and Decreases NMDA Receptors in the Brain of a Transgenic Mouse Model of Alzheimer’s Disease. European Journal of Neuroscience 2005, 22, 617–626. [Google Scholar] [CrossRef]
- Arsenault, D.; Julien, C.; Tremblay, C.; Calon, F. DHA Improves Cognition and Prevents Dysfunction of Entorhinal Cortex Neurons in 3xTg-AD Mice. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Casali, B.T.; Corona, A.W.; Mariani, M.M.; Karlo, J.C.; Ghosal, K.; Landreth, G.E. Omega-3 Fatty Acids Augment the Actions of Nuclear Receptor Agonists in a Mouse Model of Alzheimer’s Disease. Journal of Neuroscience 2015, 35, 9173–9181. [Google Scholar] [CrossRef]
- Perez, S.E.; Berg, B.M.; Moore, K.A.; He, B.; Counts, S.E.; Fritz, J.J.; Hu, Y.S.; Lazarov, O.; Lah, J.J.; Mufson, E.J. DHA Diet Reduces AD Pathology in Young APPswe/PS1ΔE9 Transgenic Mice: Possible Gender Effects. J Neurosci Res 2010, 88, 1026–1040. [Google Scholar] [CrossRef]
Figure 1.
The Hallmarks of aging. Aging is a multifactorial process at various levels, from molecules to cells, organs to systems, ultimately affecting the entire organism. In 2023, López-Otín and colleagues refined the framework of aging hallmarks, identifying 12 key features: dysbiosis, stem cell exhaustion, chronic inflammation (inflammaging), altered intercellular communication, mitochondrial dysfunction, epigenetic alterations, impaired macroautophagy, loss of proteostasis, deregulated nutrient sensing, telomere attrition, genomic instability, and cellular senescence.
Figure 1.
The Hallmarks of aging. Aging is a multifactorial process at various levels, from molecules to cells, organs to systems, ultimately affecting the entire organism. In 2023, López-Otín and colleagues refined the framework of aging hallmarks, identifying 12 key features: dysbiosis, stem cell exhaustion, chronic inflammation (inflammaging), altered intercellular communication, mitochondrial dysfunction, epigenetic alterations, impaired macroautophagy, loss of proteostasis, deregulated nutrient sensing, telomere attrition, genomic instability, and cellular senescence.
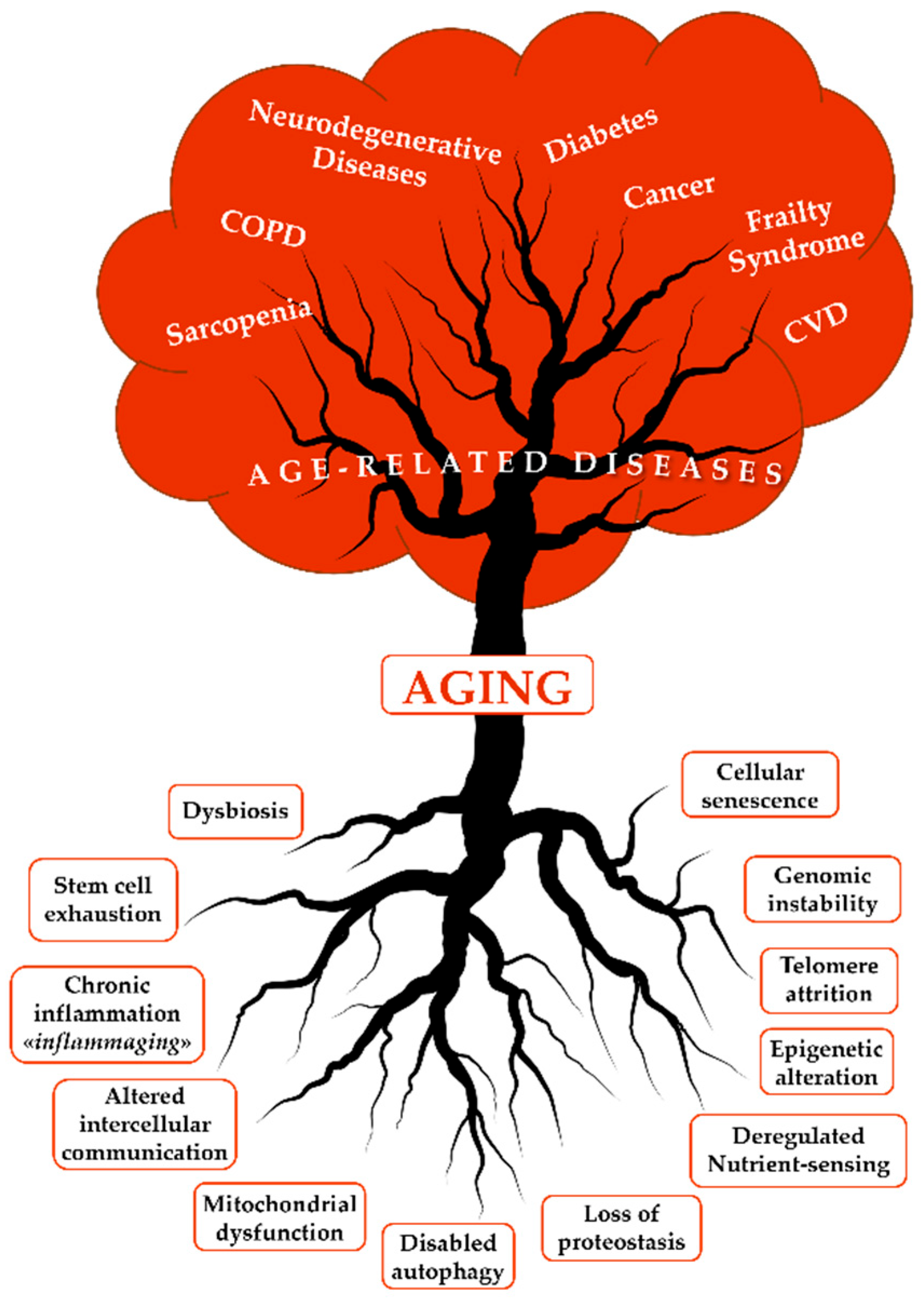
Figure 2.
The Hallmarks of cellular senescence. Senescent cells undergo distinct morphological and molecular alterations defining cellular senescence’s hallmarks. At the genomic level, senescence is characterized by stable cell cycle arrest, primarily driven by the p16INK4a/Rb and p21CIP1/p53 pathways. Chromatin reorganization is a key feature, marked by senescence-associated heterochromatic foci (SAHFs) and γ-H2AX foci, indicating DNA damage. Lysosomal alterations are also evident, with increased lysosomal mass and activity, elevated expression of senescence-associated β-galactosidase (SA-β-Gal), and lipofuscin accumulation. Senescent cells exhibit notable morphological changes, including enlargement, flattening, and modifications in membrane structure. A feature of nuclear dysfunction is the downregulation of Lamin B1, leading to nuclear envelope instability. The senescent surfaceome is enriched with specific channels and receptors, such as NOTCH1, DPP4, and B2M. Mitochondrial dysfunction is another critical feature, leading to increased ROS production and deregulated nutrient sensing. SIRT1 and SIRT3 play pivotal roles in mitochondrial homeostasis and oxidative stress regulation. SIRT1, located in the nucleus, modulates mitochondrial function by deacetylating FOXO3a, reducing inflammatory protein expression, and activating PGC-1α to promote mitochondrial biogenesis. Oxidative stress inactivates SIRT3, leading to SOD2 hyperacetylation and increased mitochondrial ROS (mtROS), creating a vicious cycle of oxidative stress and mitochondrial dysfunction. SIRT3-mediated deacetylation of FOXO3a and SOD2 counteracts ROS accumulation by upregulating antioxidant defenses, including catalase. Finally, a defining feature of senescence is the SASP, characterized by the secretion of pro-inflammatory cytokines (e.g., IL-1β, IL-6, TNF-α, etc.) and matrix metalloproteinases (MMPs), often encapsulated in extracellular vesicles (EVs). ↑ increase; ↓decrease.
Figure 2.
The Hallmarks of cellular senescence. Senescent cells undergo distinct morphological and molecular alterations defining cellular senescence’s hallmarks. At the genomic level, senescence is characterized by stable cell cycle arrest, primarily driven by the p16INK4a/Rb and p21CIP1/p53 pathways. Chromatin reorganization is a key feature, marked by senescence-associated heterochromatic foci (SAHFs) and γ-H2AX foci, indicating DNA damage. Lysosomal alterations are also evident, with increased lysosomal mass and activity, elevated expression of senescence-associated β-galactosidase (SA-β-Gal), and lipofuscin accumulation. Senescent cells exhibit notable morphological changes, including enlargement, flattening, and modifications in membrane structure. A feature of nuclear dysfunction is the downregulation of Lamin B1, leading to nuclear envelope instability. The senescent surfaceome is enriched with specific channels and receptors, such as NOTCH1, DPP4, and B2M. Mitochondrial dysfunction is another critical feature, leading to increased ROS production and deregulated nutrient sensing. SIRT1 and SIRT3 play pivotal roles in mitochondrial homeostasis and oxidative stress regulation. SIRT1, located in the nucleus, modulates mitochondrial function by deacetylating FOXO3a, reducing inflammatory protein expression, and activating PGC-1α to promote mitochondrial biogenesis. Oxidative stress inactivates SIRT3, leading to SOD2 hyperacetylation and increased mitochondrial ROS (mtROS), creating a vicious cycle of oxidative stress and mitochondrial dysfunction. SIRT3-mediated deacetylation of FOXO3a and SOD2 counteracts ROS accumulation by upregulating antioxidant defenses, including catalase. Finally, a defining feature of senescence is the SASP, characterized by the secretion of pro-inflammatory cytokines (e.g., IL-1β, IL-6, TNF-α, etc.) and matrix metalloproteinases (MMPs), often encapsulated in extracellular vesicles (EVs). ↑ increase; ↓decrease.
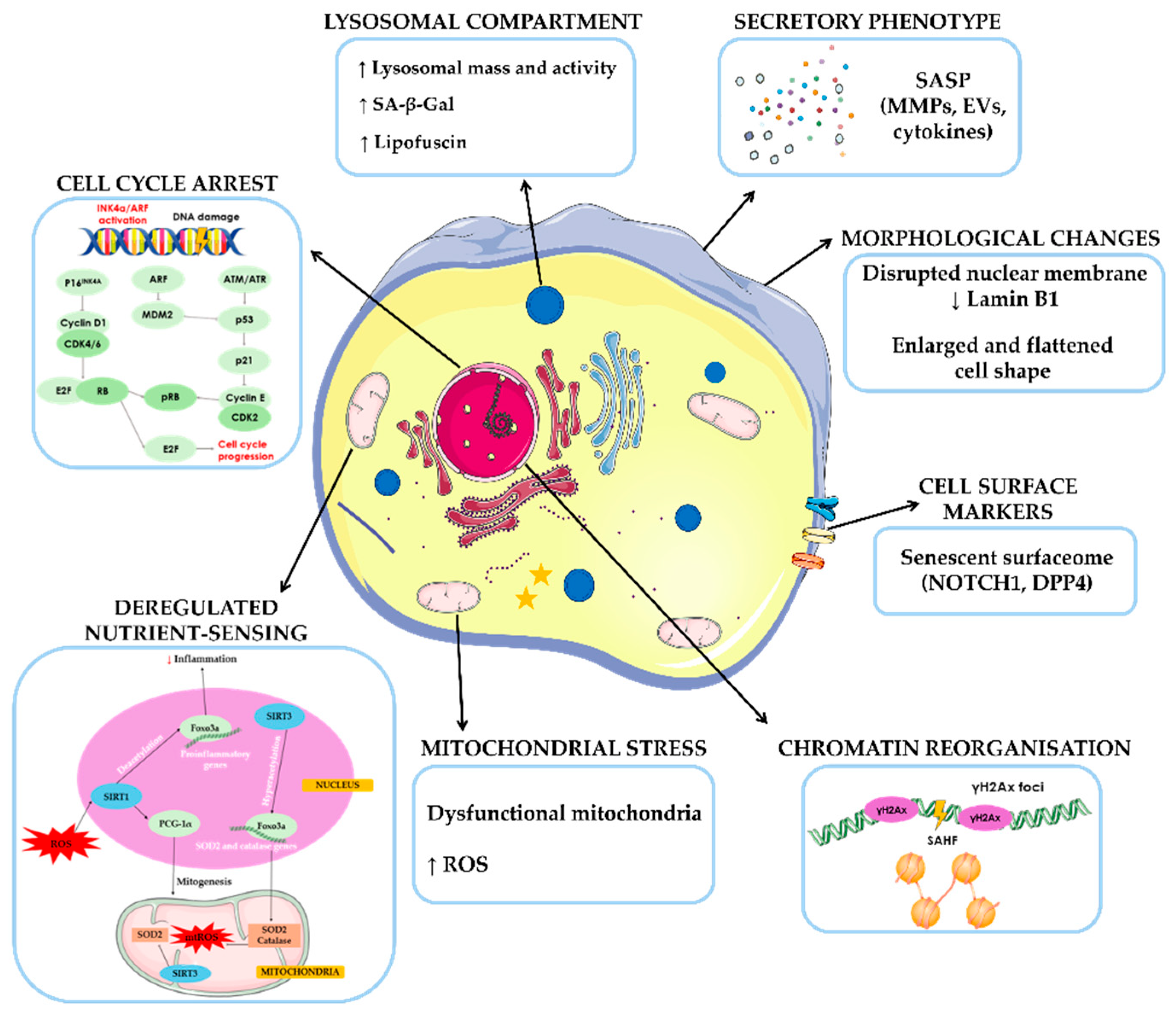
Figure 3.
The Benefits of Nutraceuticals and Functional Foods in Aging and Neurodegenerative Diseases. Nutraceuticals and functional foods are crucial in combatting aging, providing a natural and accessible approach to promoting longevity and healthspan. Among the most researched bioactive compounds are antioxidant vitamins, polyphenols, spices, dietary fiber, probiotics, prebiotics, and PUFAs. These substances offer protective and rejuvenating effects at the cellular level, helping to alleviate the hallmarks of aging and the harmful traits of senescent cells. Nutraceuticals can support cellular homeostasis and slow down age-related functional decline by reducing oxidative stress, moderating chronic inflammation, preserving DNA integrity, preventing telomere shortening, and restoring metabolic balance. Additionally, they aid in maintaining lysosomal function and guarding against the structural changes associated with senescence, including morphological alterations and cellular enlargement. Through their diverse mechanisms of action, these bioactive compounds assist in delaying aging and reducing the risk of ARDs, such as neurodegeneration.
Figure 3.
The Benefits of Nutraceuticals and Functional Foods in Aging and Neurodegenerative Diseases. Nutraceuticals and functional foods are crucial in combatting aging, providing a natural and accessible approach to promoting longevity and healthspan. Among the most researched bioactive compounds are antioxidant vitamins, polyphenols, spices, dietary fiber, probiotics, prebiotics, and PUFAs. These substances offer protective and rejuvenating effects at the cellular level, helping to alleviate the hallmarks of aging and the harmful traits of senescent cells. Nutraceuticals can support cellular homeostasis and slow down age-related functional decline by reducing oxidative stress, moderating chronic inflammation, preserving DNA integrity, preventing telomere shortening, and restoring metabolic balance. Additionally, they aid in maintaining lysosomal function and guarding against the structural changes associated with senescence, including morphological alterations and cellular enlargement. Through their diverse mechanisms of action, these bioactive compounds assist in delaying aging and reducing the risk of ARDs, such as neurodegeneration.
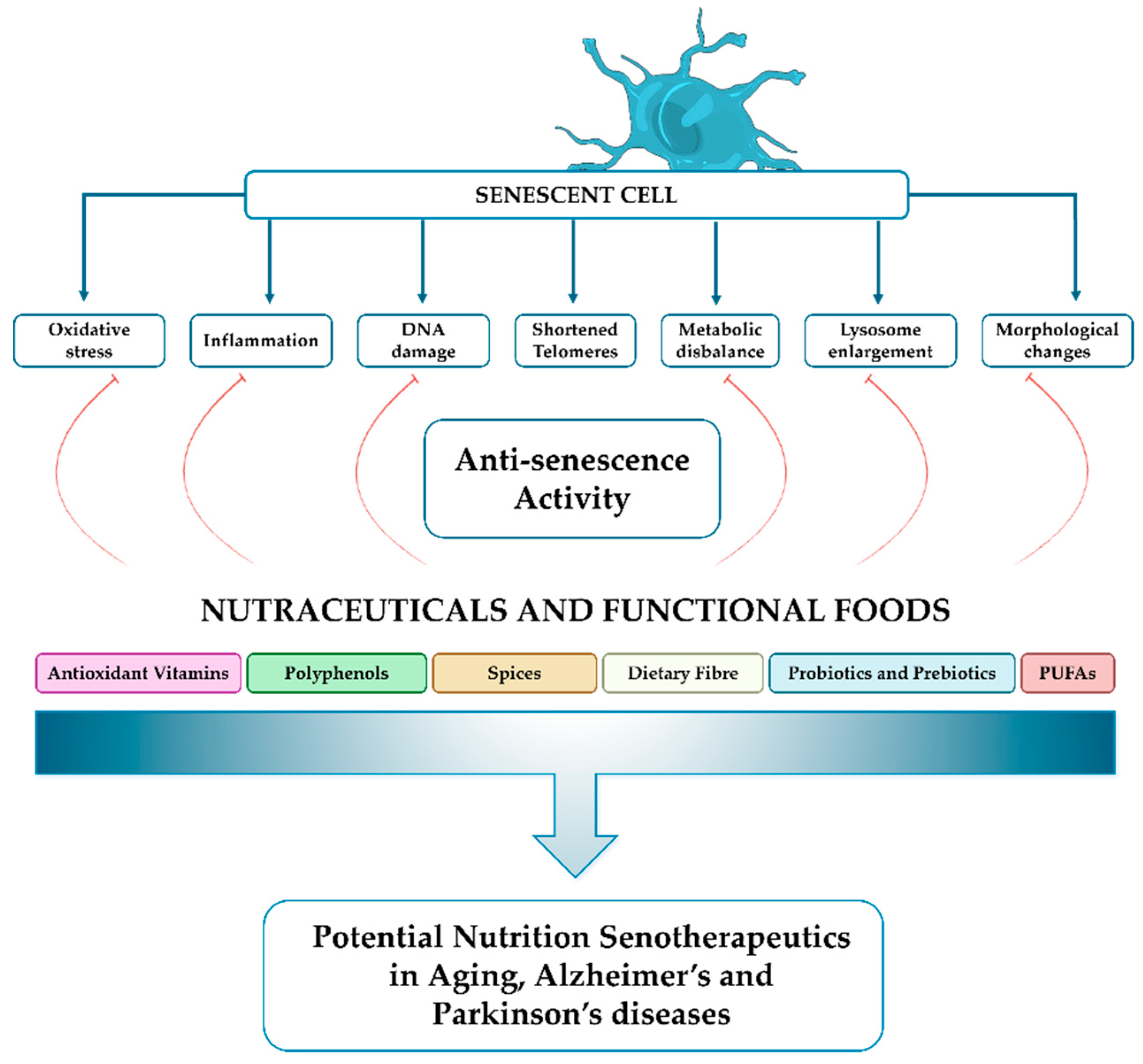
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



