
Submitted:
08 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Fungal infections caused by Candida spp. cause potentially fatal infections with high morbidity and mortality rates, particularly in immunocompromised individuals. Their resistance to conventional antifungal drugs is closely associated with biofilm formation, which increasingly limits available therapeutic options, making medical practice a major challenge worldwide. In this context, plant derived compounds are an excellent alternative to be investigated. The objective of this study was to evaluate the antifungal potential of the flavonoid diosmin (DIO) and its synergistic capacity with commercial antifungals against clinical isolates of Candida spp. Susceptibility testing by broth microdilution method showed antifungal activity of DIO against all isolates tested, with minimum inhibitory concentration (MIC90) values ranging from 1150 to 2251 µg/mL. Furthermore, we report the ability of DIO to inhibit biofilm and mature biofilm formation, with a greater inhibitory effect on fungal biofilm formation; this effect with DIO was significantly greater in most cases than that shown by amphotericin B (AFB). Intracellular leakage experiments (260/280 nm) showed damage to the cell membrane. This indicates that the antifungal action of DIO could be associated with damage to the cell membrane integrity and consequent death of these pathogens. Synergism experiments with DIO and fluconazole (FLZ) and DIO and AFB revealed a significant synergistic effect against Candida spp. These results highlight the antifungal and synergistic potential of the flavonoid DIO against antifungal resistant Candida spp. Furthermore, these findings serve as a basis for future studies aimed at elucidating DIO's antimicrobial mechanisms of action and contribute to the search for novel compounds from natural sources with antimicrobial potential.
Keywords:
flavonoid
; diosmin
; antifungal
; Candida spp
; antibiofilm
; synergism
1. Introduction
Candida spp. are a common cause of healthcare associated infections (HAIs) globally; approximately 1565000 people contract invasive candidiasis each year, particularly immunosuppressed individuals, including: critically ill patients in intensive care units (ICUs), people on long term broad spectrum antibiotic therapy and, recently, people with complications of post-COVID-19 disease. Invasive candidiasis is often misdiagnosed and is responsible for 995000 deaths (63.6% of all cases) annually. Africa is estimated to have a higher proportion of the invasive candidiasis burden than the rest of the world [1-3]. Bloodstream infections due to these pathogens are associated with mortality of up to approximately 40% according to the Excellence Center of Medical Mycology (ECMM), Cologne, Germany [4,5]. Furthermore, Candida spp., are the most common cause of fungal endocarditis, implicated in more than 50% of cases, are associated with high morbidity and mortality (0.70%) and present numerous challenges during clinical care [6]. These pathogens can persist within the host through the development of pathogenicity and multidrug resistance, which often leads to the failure of therapeutic strategies. A particular feature of the pathogenicity of Candida spp. it is their ability to form biofilms, which protects them from external factors such as the host's immune system defenses and antifungal drugs, multiplying by more than 1000 their resistance to first-line antifungals, compared to the planktonic growth of the same strains [7,8]. The rapid spread of multi resistant Candida spp. strains [1,9-12], associated with therapeutic failure and high mortality is alarming, representing this situation as a substantial threat to the prognosis of patients. In this context, the search for and development of new compounds with activity against Candida spp. that are safe, tolerable, and effective is urgent today. Since ancient times, natural products, particularly plant-based ones, have made a valuable contribution to pharmacotherapy, especially in infectious diseases and cancer [13,14]. Plants play a primary role as a source of specialized metabolites with recognized medicinal properties, due to their wide chemical diversity, these metabolites can be used directly as bioactive compounds, as drug prototypes or used as pharmacological tools for different targets [15], which is why they are an excellent alternative to be investigated.
Flavonoids are phytochemicals widely known for their health promoting pharmacological properties, including antimicrobial activity [16]. Diosmin (DIO) (diosmetin 7-O-rutinoside), Figure 1, is a flavone glycoside derived from hesperidin, a flavanone found abundantly in citrus fruits; it has anti-inflammatory, antioxidant, antidiabetic, antihyperlipidemic, antifibrotic, anticancer and hepatoprotective effects, among others [17-21]. Regarding the antimicrobial activity of this flavonoid, its potential against Pseudomonas aeruginosa has been reported, reducing its virulence mechanisms [22]; Escherichia coli, Pseudomonas putida, Staphylococcus aureus [23], varicella-zoster virus (VZV) [24], and its synergistic effect with amoxicillin-clavulanic acid (AMC) against Mycobacterium tuberculosis [25], among other biological activities, however, its potential against pathogenic yeasts of the Candida genus has not been documented. We hypothesize that the flavonoid DIO has activity against Candida spp. and the fungal biofilms of these pathogens. The objective of this investigation was to evaluate the antifungal potential of DIO against clinical isolates of Candida spp., estimate their ability to inhibit biofilms and explore their possible effect on the yeast membrane, contributing to the search for new compounds of natural origin that can serve as adjuvants in the treatment of pathogenic yeasts resistant to antifungal drugs.
2. Results
2.1. Susceptibility Testing
DIO showed antifungal activity against all clinical isolates of Candida spp. studied. We observed a reduction in the growth percentage of yeasts treated with DIO, compared to untreated isolates used as controls. Figure 2 shows the similar trend among isolates to increase the percentage of growth reduction as the concentration of DIO increases. Table 1 shows the MIC values; MIC90 values of DIO were obtained between 1150 and 2251 µg/mL and MIC50 values between 660.4 and 1199 µg/mL. FLZ MIC90 values ranged from 2.3 to 241.8 µg/mL, with the majority of strains tested resistant to FLZ. This effect on Candida spp. was shown to be dependent on DIO concentration.
It is observed that the efficacy of DIO was different between strains of the same species.
2.2. Biofilm Reduction
All Candida spp. isolates evaluated in this study produced biofilms on polystyrene microplates, as shown in Figure 3a; showing strong biofilm biomass production, the isolates C. tropicalis, C. haemulonii and C. parapsilosis (OD590 > 3), while the isolates C. auris, C. albicans and C. krusei (OD590: 1.1-3.0) were moderate biofilm producers. The C. glabrata isolate (OD590: 0.1-1.0) was a weak biofilm producer. Adding the MIC90 of DIO to yeast inhibited biofilm formation by 35.35–87.85% after 1 h of exposure, while the percentage inhibition of biofilm formation in cells treated with AFB ranged from 6.49–91.26%, as shown in Table 2; in most isolates, the inhibitory effect of DIO on biofilm formation was greater than that of AFB, as shown in Figure 3b. In C. albicans and C. glabrata isolates, no inhibitory effect on biofilm formation was evident from DIO or AFB. The effect of DIO and AFB was less evident in the inhibition of mature biofilms; as seen in Figure 3c, an inhibitory effect was only evident against mature biofilms of the isolates C. tropicalis, C. krusei and C. auris. The behavior of DIO and AFB was similar, with significant differences observed only with C. auris. The percentages of inhibition of mature biofilms by DIO and AFB on Candida spp., are shown in Table 3.
Table 2 shows the percentages of inhibition of DIO biofilm formation in Candida spp. during 24 hours of exposure.
Table 3 shows the percentages of DIO inhibition against mature Candida spp. biofilms after 24 hours of incubation.
2.3. Leakage of Nucleic Acids and Proteins through the Fungal Membrane
The action of DIO on the membrane integrity of Candida spp. was evaluated by assays for the release of intracellular constituents that absorb at 260/280 nm, such as nucleic acids and proteins. These assays were performed at 0, 30, 60, and 120 min after treatment with the MIC90 of DIO for each isolate. As seen in Figure 4, OD260/OD280 values in DIO-treated groups were significantly higher from baseline compared to FLZ treated groups, where minimal or even no (in some cases) release of intracellular material was observed in all Candida spp. isolates. These results indicate damage to fungal cell membrane permeability caused by DIO.
2.4. Diosmin Action in Combination with Commercial Antifungals
Figure 5 shows the synergistic effect of DIO and the commercial antifungals FLZ and AFB against most of the Candida spp. isolates evaluated. The DIO-FLZ combination exhibited synergistic effects against four of the seven Candida spp. isolates tested (Ca02, Ct74, Cg01, and Cp02), while the DIO-AFB combination exhibited synergism against five of the seven strains tested (Ca02, Ct74, Cg01, Ch01, and Cau75). No interaction was observed against strain Ck01 in any of the tested combinations (DIO-FLZ and DIO-AFB). The DIO-FLZ combination did not interact against isolate Ch01; similarly, the DIO-AFB combination did not interact against isolate Cp02.
Table 4 shows that the minimum inhibitory concentrations (MIC90) at which 90% of the yeasts were inhibited in synergistic activity were substantially lower than the individual MIC90 values of DIO and the antifungals used.
3. Discussion
Candida spp. are pathogens of great global concern. These yeasts' remarkable ability to adapt to the hospital environment is primarily due to their multi drug resistance to conventional drugs used for their control and the expression of various virulence factors, including the production of powerful biofilms. This limits therapeutic options and increases morbidity and mortality rates, worsening the patient situation and increasing global financial costs. This situation sparks interest and drives the search for new molecules with antifungal properties. In this context, chemical compounds of natural or synthetic origin become an excellent alternative.
In this research we demonstrate that DIO, a flavonoid present in citrus fruits, has antifungal activity against clinical isolates of Candida spp., and this effect is concentration dependent. Results consistent with our study document the potential of DIO against methicillin resistant and methicillin sensitive Staphylococcus aureus (MRSA) strains; it has been shown that diosmetin (aglycone form of DIO) significantly suppresses the pharmacokinetic activity of MRSA in a dose-dependent manner. This could cause ATP deficiency and affect the bacterial efflux pump, which would contribute to the antibacterial action of diosmetin against MRSA [25,26]. Similar studies [22] reported the antibacterial effect of DIO against P. aeruginosa suggesting that this compound could be a potential candidate for the development of a new agent targeting P. aeruginosa infections by reducing its virulence mechanisms, showing a negative regulation of the associated genes (lasI and pvdS). DIO has been implicated in partial inhibition of ATP synthesis in E. coli, and DIO and diosmetin have been virtually coupled to the enzymes LdtMt1 and LdtMt2, which are involved in M. tuberculosis (Mtb) cell wall biosynthesis [21]. MIC and BMC antimicrobial assays have shown that pompia and lemon juices high in DIO and other flavonoids have inhibitory and antibiofilm effects against pathogenic bacteria including P. aeruginosa, S. aureus and Enterococcus faecalis [28]. The antiviral effect of DIO has also been documented [24], indicating that DIO significantly inhibited the varicella zoster virus (VZV). The antimicrobial effect of DIO has been poorly reported against bacteria, and the role of DIO in the treatment of infectious diseases such as malaria, dengue, chikungunya, SARS-CoV-2, and leishmaniasis may likely be investigated in the future [23]. This study reports for the first time the effect of DIO against clinical isolates of pathogenic yeasts Candida spp.
The damage to the permeability of the fungal cell membrane of Candida spp. caused by DIO was evidenced in the intracellular material leakage experiments (260/280 nm), which showed a significant and early release of intracellular material when the fungal cells were treated with DIO; these results are consistent with the results reported by [23], who showed antibacterial activity of DIO against strains of E. coli, P. putida and S. aureus, reporting that among the mechanisms of action of this compound is the alteration of the permeability of the cell membrane, as well as the formation of holes in the bacterial cell wall, the inhibition of transduction, the inhibition of respiratory enzymes due to the formation of free radicals and the inactivation of several enzymes. Our results suggest that damage to the cell membrane of Candida spp. is at least one of the mechanisms of antifungal action of this flavonoid.
Candida spp. have been widely documented for their ability to form potent biofilms [29], which increase resistance to both antifungal drugs and the host immune response, causing persistent infections; these biofilms vary depending on the source of the infection [30]. The Candida spp. isolates in this study were all biofilm producers (Figure 3a). We report the potential of DIO against fungal biofilms; this is in line with studies documented by [22], who recorded a high potential of DIO to disrupt both bacterial biofilm formation and eradication, including a significant reduction in biofilm biomass, exopolysaccharide and extracellular DNA production in P. aeruginosa, where treatment of the biofilm with DIO resulted in the lowest percentage of live microbial cells. When comparing the efficacy of DIO with AFB, we highlighted the role of DIO in inhibiting biofilm formation after 1 hour of treatment and in most cases it was significantly higher than with AFB (Figure 3b); however, the effect of DIO and AFB against mature biofilms was lower (Figure 3c), with similar effects; this is consistent with studies reported by [31], which indicate the ability of liposomal AFB to inhibit further biofilm growth, but its ineffectiveness in eradicating mature biofilms, even at high doses.
On the other hand, effective therapeutic regimens for the treatment of Candida spp. are limited. In this study, we highlight the synergistic effect of DIO in combination with the antifungals FLZ and AFB against antifungal resistant Candida spp. clinical isolates. We emphasize the synergistic effect of DIO-AFB against the multidrug resistant strain of C. auris Cau75, achieving substantial reductions in the MIC90 values of both treatments (Table 4). These results are consistent with studies demonstrating the synergistic effect of DIO and its aglycone form (diosmetin) with other drugs. The combination of DIO and amoxicillin-clavulanic acid has shown synergistic inhibition of mycobacterial growth, demonstrating greater mycobactericidal activity against Mycobacterium marinum [21,25]. Likewise, the synergistic effect of diosmetin with erythromycin against the ABC transporter overexpressed in MRSA has been documented [26]. Besides, the synergistic effects of DIO have also been reported against different types of cancer [32], revealing the synergistic chemotherapeutic effects of DIO in combination with BEZ-235 in the colorectal cancer cell line HCT-116. Likewise, the synergistic effect of DIO and interferon-α has been shown in metastatic pulmonary melanoma [33]. DIO in combination with other flavonoids has also shown a synergistic effect, in fact, hesperidin and DIO increased the cytotoxic activity of cisplatin on hepatocellular carcinoma [34]; likewise, DIO in combination with naringenin enhanced apoptosis in colon cancer cells [35]. Our results suggest that DIO could act as an important adjuvant in the treatment of Candida spp. multidrug resistant to antifungal drugs.
Our results provide important new information on the antifungal potential of the flavonoid DIO against Candida spp., demonstrating its action against the cell membrane and its inhibitory effect against fungal biofilms; in addition, its synergistic potential with the commercial antifungal FLZ. We also provide information that serves as a basis for future research to elucidate the antifungal mechanisms of action of this compound, which could serve as an adjuvant for the treatment of infections caused by these pathogenic yeasts. Importantly, plant derived compounds can potentially be used to combat multidrug resistant and biofilm forming strains of Candida spp., thus becoming a promising alternative to antifungal drugs.
4. Materials and Methods
4.1. Reagents
The RPMI 1640, phosphate buffered saline (PBS), and yeast peptone dextrose broth (YPD) were obtained from Thermo Fisher Scientific, Waltham, MA, USA; 3-N-morpholinopropanesulfonic acid (MOPS) was obtained from Merck; potato dextrose broth (PDB), sabouraud dextrose agar (SDA), sabouraud dextrose broth (SDB), amphotericin B (AFB), crystal violet and Fluconazole (FLZ) used in this study were obtained from Sigma-Aldrich, USA; glacial acetic acid was obtained from Carlo Erba Reagents, Italy.
4.2. Diosmin (DIO)
Diosmin, 3′,5,7-trihydroxy-4′-methoxyflavone-7-rutinoside, (C28H32O15) molecular weight: 608.54, was purchased from Sigma-Aldrich Inc, CAS number: 520-27-4.
4.3. Strains
Nine clinical isolates of Candida spp. including: C. albicans (Ca02 and Ca77), C. glabrata (Cg01), C. tropicalis (Ct03 and Ct74), C. krusei (Ck01), C. haemulonii (Ch01), C. parapsilosis (Cp02), and C. auris (Cau75) were used in this study. The isolates were cultured from the blood and urine culture samples of patients hospitalized at the Social Health Service S.A.S. in the city of Sincelejo, Colombia. All microorganisms were identified by standard methods: Vitek 2 Compact, Biomerieux SA, YST Vitek 2 Card and AST-YS08 Vitek 2 Card (Ref 420739). SDA medium and BBL CHROMagar Candida medium were used to maintain the cultures until the tests were carried out.
4.4. Antifungal Susceptibility Testing
The minimum inhibitory concentration (MIC) of DIO against clinical isolates of Candida spp., was defined as the lowest concentration at which 90% of fungal growth was inhibited (MIC90) compared to the control. The MIC90 was determined by broth microdilution assays using 96 well microtiter plates (Nunclon Delta, Thermo Fisher Scientific, Waltham, MA, USA) as described in the Clinical Laboratory Standards Institute (CLSI) method (M27-A3) [36] and the European Committee for Antimicrobial Susceptibility Testing (EUCAST) method [37], with minor modifications. Serial dilutions were made in RPMI 1640 broth (pH 7.0) buffered with 0.165 M MOPS to obtain final concentrations of 3.90 to 1000 µg/mL of DIO in each reaction well. Stock solutions of DIO were prepared at 20000 µg/mL in DMSO and FLZ at 1500 µg/mL in 10% DMSO in distilled water. Assays were performed with a final volume of 200 µL per well as follows: 100 µL of fungal inoculum at a concentration of 1×106 CFU/mL and 100 µL of DIO adjusted to achieve the concentrations described above in a final reaction system. Candida spp. isolates without DIO and FLZ were used as growth controls and positive controls, respectively; wells with culture medium without inoculum and without DIO were used as negative controls. For each test, controls were run with different concentrations of DIO in culture medium without inoculum. The plates were incubated at 37 °C for 24 h. Inhibition of fungal growth by DIO was determined by the change in optical density using a SYNERGY LX microplate reader (Biotek), at 530 nm, from the start of incubation to the final time (24 h) [38], and the percentage reduction in growth was calculated using the following equation:
where, ODt24: optical density of the test well at 24 h post-inoculation; ODt0: optical density of the test well at 0 h post-inoculation; ODgc24: optical density of the growth control well at 24 h post-inoculation; ODgc0: optical density of the growth control well at 0 h post inoculation.
%Reduction = (1 − (ODt24 − ODt0/ODgc24 − ODgc0)) × 100
4.5. Quantitative Evaluation of Biofilm Inhibition
Clinical isolates of Candida spp. were evaluated to quantify biofilm reduction in the presence of DIO following the methodology reported by [38], with some modifications. For biofilm formation, yeast colonies on SDA with 24 h of incubation were used to standardize the inoculum to a concentration of 1×106 cells/mL. Subsequently, 200 µL of the fungal inoculum was cultured in 96-well plates in YPD broth and incubated at 37 °C for 48 h. The broth was then removed from the microplates and 200 µL of the DIO MIC90 was added to each isolate in YPD broth, incubating at 37 °C for 1 h. Subsequently, floating cells were removed and the biofilm at the bottom of the wells was washed with deionized water three times. Six replicates of each sample were prepared. Cultures without DIO were used as a control and AFB as a positive control. Biofilm reduction was quantified by staining the wells with 0.1% crystal violet for 20 min. The samples were washed with deionized water until excess dye was removed. Finally, they were immersed in 250 µL of 30% glacial acetic acid. Absorbance values were measured at 590 nm (OD590) using a SYNERGY LX microplate reader (Biotek). Biofilm production was grouped into the following categories: OD590 < 0.1: non-producers (NP), OD590 0.1–1.0: weak producers (WP), OD590 1.1–3.0: moderate producers (MP), and OD590 > 3.0: strong producers (SP). For biofilm inhibition assays, the standardized bacterial inoculum was incubated simultaneously with the MIC90 of DIO. Biofilm reduction was calculated using the following equation:
where, AbsCO: absorbance of the control and AbsDIO: absorbance of the sample treated with DIO
% Biofilm reduction: AbsCO − AbsDIO/AbsCO × 100
4.6. Leakage of Nucleic Acids and Proteins through the Fungal Membrane
The release of intracellular material was measured according to the methodology proposed by [38], with some modifications. Yeasts grown in SDB were centrifuged at 3000× g for 20 min, washed three times and resuspended in 20 mL of PBS (pH 7.0). The cell suspension was then treated with DIO (MIC90 for each isolate) and incubated at 37 °C for 0, 30, 60 and 120 min. Subsequently, 2 mL of the samples were collected and centrifuged at 3000× g for 20 min. To determine the concentration of the released constituents, 2 mL of supernatant was used to measure the absorbance at 260/280 nm in a Spectroquant® Prove 300 UV/Vis spectrophotometer. Samples without DIO and samples with FLZ were used as controls. All assays were performed in triplicate.
4.7. Diosmin Action in Combination with Commercial Antifungals
To obtain the fractional inhibitory concentration indices (FICIs) of DIO in combination with FLZ and AFB against Candida spp., we followed the methodology proposed by [39]. Serial dilutions were made in RPMI 1640 broth containing 0.1% 2,3,5-triphenyltetrazolium chloride (CTT), reaching final concentrations in the range of 1000—3.9 µg/mL, 256—0.25 µg/mL and 32—0.062 µg/mL for DIO, FLZ and AFB, respectively. The assay was performed with a total volume of 200 µL per well, distributed as follows: 50 µL of DIO + 50 µL of FLZ and AFB were added to reach the concentrations described previously, as well as 100 µL of fungal inoculum at a concentration of 1×106 CFU/mL. Absorbance readings were measured with a Chromate 4300 ELISA reader at a wavelength of 630 nm, and subsequently, after 24 h of incubation at 37 °C, measurements were performed again. The FICI were calculated using the following equation:
FICIs = MIC90 DIO in combination/ MIC90 DIO single + MIC90 antifungal in combination/ MIC90 antifungal single
The results were interpreted following the approach used in [39]. The FICIs are considered to have a synergistic effect (FIC index ≤ 1.0), a commutative effect (FIC index = 1), no interaction (1.0 < FIC index ≤ 2.0) or an antagonistic effect (FIC index > 2.0).
4.8. Statistical Analysis
The results were analyzed using GraphPad Prism software version 8.0 and Microsoft Excel version 2024. Initially, the Shapiro-Wilk test was used to determine the distribution of the data. Pearson's correlation coefficients were then used to measure the degree of linearity, the correlation between DIO concentration and the percentage reduction in fungal growth. To compare the effects of DIO and the antifungal agent AFB on biofilm reduction, Games-Howell's post hoc tests were used; this test was also used to compare the effects of DIO and the antifungal agent on intracellular membrane leakage (260/280 nm).
5. Conclusions
In this study, we investigated the antifungal potential of the flavonoid diosmin against clinical isolates of Candida spp., as well as its role in biofilm inhibition. We also explored its action against the cell membrane of these pathogens. We demonstrated the antifungal action of DIO against Candida spp. this effect being associated with damage to cell membrane integrity, in addition to its action against fungal biofilms. It is necessary to continue these studies, with the aim of elucidating the mechanisms of antifungal action of DIO, and its possible synergistic action with drugs, since this flavonoid shows promise as an alternative tool for the treatment and control of multi-resistant nosocomial pathogens such as Candida spp.
Author Contributions
Conceptualization, O.I.C.-M., N.R.C. and A.A.-O.; methodology, O.I.C.-M., N.R.C. and A.A.-O.; validation, O.I.C.-M., N.R.C. and A.A.-O.; formal analysis, O.I.C.-M. and A.A.-O.; investigation, O.I.C.-M., N.R.C. and A.A.-O; writing—original draft preparation, O.I.C.-M. and A.A.-O.; writing—review and editing, O.I.C.-M., N.R.C. and A.A.-O.; supervision, O.I.C.-M. and A.A.-O.; project administration, A.A.-O.; funding acquisition, A.A.-O. and O.I.C.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by resources from the FCB-02-22 project of the University of Córdoba, Montería, Colombia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article and the
supporting information.
Acknowledgments
We would like to thank the University of Córdoba and the Microbiological Interactions Research Group (GIIM).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ibe, C.; Pohl, C. H. Epidemiology and drug resistance among Candida pathogens in Africa: Candida auris could now be leading the pack. The Lancet Microbe. Elsevier Ltd, 2024. [CrossRef]
- Naranjo-Bravo, J.; Romero-Romero, D.; Contreras-Rodríguez, A.; Aguilera-Arreola, M. G.; Parra-Ortega, B. Candida isolation during COVID-19: microbiological findings of a prospective study in a regional hospital. Arch Med Res 2024, 55, 6. [CrossRef]
- Koulenti, D.; Karvouniaris, M.; Paramythiotou, E.; Koliakos, N.; Markou, N.; Paranos, P.; Meletiadis, J.; Blot, S. Severe candida infections in critically ill patients with COVID-19. Journal of Intensive Medicine. Chinese Medical Association, 2023, pp 291–297. [CrossRef]
- Salmanton-García, J.; Cornely, O. A.; Stemler, J.; Barać, A.; Steinmann, J.; Siváková, A.; Akalin, E. H.; Arikan-Akdagli, S.; Loughlin, L.; Toscano, C.; Narayanan, M.; Rogers, B.; Willinger, B.; Akyol, D.; Roilides, E.; Lagrou, K.; Mikulska, M.; Denis, B.; Ponscarme, D.; Scharmann, U.; Azap, A.; Lockhart, D.; Bicanic, T.; Kron, F.; Erben, N.; Rautemaa-Richardson, R.; Goodman, A. L.; Garcia-Vidal, C.; Lass-Flörl, C.; Gangneux, J. P.; Taramasso, L.; Ruiz, M.; Schick, Y.; Van Wijngaerden, E.; Milacek, C.; Giacobbe, D. R.; Logan, C.; Rooney, E.; Gori, A.; Akova, M.; Bassetti, M.; Hoenigl, M.; Koehler, P. Attributable mortality of candidemia – results from the ECMM candida III multinational European observational cohort study. Journal of Infection 2024, 89, 3. [CrossRef]
- Aslanov, P. B.; Ermakova, D. L.; Nabieva, D. A. Surveillance and prevention of fungal infections in the neonatal intensive care unit. International Journal of Infectious Diseases 2025, 152, 107729. [CrossRef]
- Thompson, G. R.; Jenks, J. D.; Baddley, J. W.; Lewis, J. S.; Egger, M.; Schwartz, I. S.; Boyer, J.; Patterson, T. F.; Chen, S. C. A.; Pappas, P. G.; Hoenigl, M. Fungal endocarditis: pathophysiology, epidemiology, clinical presentation, diagnosis, and management. Clinical Microbiology Reviews 2023, 36, 3. [CrossRef]
- Žiemytė, M.; Rodríguez-Díaz, J. C.; Ventero-Martín, M. P.; Mira, A.; Ferrer, M. D. Real-time monitoring of biofilm growth identifies andrographolide as a potent antifungal compound eradicating candida biofilms. Biofilm 2023, 5, 100134. [CrossRef]
- Cavalheiro, M.; Teixeira, M. C. Candida biofilms: threats, challenges, and promising strategies. Frontiers in Medicine 2018, 5, 28. [CrossRef]
- Brassington, P. J. T.; Klefisch, F.-R.; Graf, B.; Pfüller, R.; Kurzai, O.; Walther, G.; Barber, A. E. Genomic reconstruction of an azole-resistant Candida parapsilosis outbreak and the creation of a multi-locus sequence typing scheme: a retrospective observational and genomic epidemiology study. Lancet Microbe 2025, 6, 1, 100949. [CrossRef]
- Ramos, L. S.; Barbosa, P. F.; Lorentino, C. M. A.; Lima, J. C.; Braga, A. L.; Lima, R. V.; Giovanini, L.; Casemiro, A. L.; Siqueira, N. L. M.; Costa, S. C.; Rodrigues, C. F.; Roudbary, M.; Branquinha, M. H.; Santos, A. L. S. The multidrug-resistant Candida auris, Candida haemulonii complex and phylogenetic related species: insights into antifungal resistance mechanisms. Current Research in Microbial Sciences 2025, 8, 100354. [CrossRef]
- Arendrup, M. C.; Arikan-Akdagli, S.; Jørgensen, K. M.; Barac, A.; Steinmann, J.; Toscano, C.; Arsenijevic, V. A.; Sartor, A.; Lass-Flörl, C.; Hamprecht, A.; Matos, T.; Rogers, B. R. S.; Quiles, I.; Buil, J.; Özenci, V.; Krause, R.; Bassetti, M.; Loughlin, L.; Denis, B.; Grancini, A.; White, P. L.; Lagrou, K.; Willinger, B.; Rautemaa-Richardson, R.; Hamal, P.; Ener, B.; Unalan-Altintop, T.; Evren, E.; Hilmioglu-Polat, S.; Oz, Y.; Ozyurt, O. K.; Aydin, F.; Růžička, F.; Meijer, E. F. J.; Gangneux, J. P.; Lockhart, D. E. A.; Khanna, N.; Logan, C.; Scharmann, U.; Desoubeaux, G.; Roilides, E.; Talento, A. F.; van Dijk, K.; Koehler, P.; Salmanton-García, J.; Cornely, O. A.; Hoenigl, M. European candidaemia is characterised by notable differential epidemiology and susceptibility pattern: results from the ECMM Candida III study. Journal of Infection 2023, 87, 5, 428–437. [CrossRef]
- Díaz-García, J.; Machado, M.; Alcalá, L.; Reigadas, E.; Sánchez-Carrillo, C.; Pérez-Ayala, A.; Gómez-García de la Pedrosa, E.; González-Romo, F.; Merino, P.; Cuétara, M. S.; García-Esteban, C.; Quiles-Melero, I.; Zurita, N. D.; Muñoz-Algarra, M.; Durán-Valle, M. T.; Martínez-Quintero, G. A.; Sánchez-García, A.; Muñoz, P.; Escribano, P.; Guinea, J.; Mesquida, A.; Gómez, A.; Muñoz, R. P.; González, M. del C. V.; Romo, F. G.; Merino-Amador, P.; Muñoz Clemente, O. M.; Berenguer, V. A.; Lobato, O. C.; Bernal, G.; Zurita, N.; Cobos, A. G.; Romero, I. S.; San Juan Delgado, F.; Romero, Y. G.; Fraile Torres, A. M. Antifungal resistance in Candida spp. within the intra-abdominal cavity: study of resistance acquisition in patients with serial isolates. Clinical Microbiology and Infection 2023, 29, 12, 1604.e1-1604.e6. [CrossRef]
- Atanasov, A.; Zotchev, S.; Dirsch, V.; Taskforce, T. I. N. P. S.; Supuran, C. Natural products in drug discovery: advances and opportunities. Nature Reviews 2021, 20, 200–216. [CrossRef]
- Naman, C. B.; Benatrehina, P. A.; Kinghorn, A. D. Pharmaceuticals, Plant Drugs. In Breeding Genetics and Biotechnology, Second Edi.; Elsevier, 2016, 2. (pp. 93-99).
- Avato, P. Editorial to the Special Issue – “Natural Products and Drug Discovery". Molecules 2020, 25, 1128. [CrossRef]
- Ciriminna, R.; Petri, G. L.; Angellotti, G.; Luque, R.; Fabiano Tixier, A.; Meneguzzo, F.; Pagliaro, M. Citrus flavonoids as antimicrobials. Chem Biodivers 2025, e202403210. [CrossRef]
- Gerges, S. H.; Wahdan, S. A.; Elsherbiny, D. A.; El-Demerdash, E. Pharmacology of diosmin, a citrus flavone glycoside: an updated review. Eur J Drug Metab Pharmacokinet. 2022. 47, 1, 1-18 . [CrossRef]
- Cazaubon, M.; Benigni, J. P.; Steinbruch, M.; Jabbour, V.; Gouhier-Kodas, C. Is there a difference in the clinical efficacy of diosmin and micronized purified flavonoid fraction for the treatment of chronic venous disorders? review of available evidence. Vasc Health Risk Manag 2021, 17, 591-600. [CrossRef]
- Hassanein, E. H. M.; Althagafy, H. S.; Baraka, M. A.; Amin, H. Hepatoprotective effects of diosmin: a narrative review. Naunyn-Schmiedeberg’s Archives of Pharmacology 2025, 398, 1, 279-295 . [CrossRef]
- Klimek-szczykutowicz, M.; Szopa, A.; Ekiert, H. Citrus limon (Lemon) phenomenon—a review of the chemistry, pharmacological properties, applications in the modern pharmaceutical, food, and cosmetics industries, and biotechnological studies. Plants, 2020, 9, 1, 119. [CrossRef]
- Zheng, Y.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Chen, Z.; Wu, L. Metabolism and pharmacological activities of the natural health-benefiting compound diosmin. Food Funct. 2020, 11, 10,8472-8492. doi: 10.1039/d0fo01598a.
- Carević, T.; Kolarević, S.; Kolarević, M. K.; Nestorović, N.; Novović, K.; Nikolić, B.; Ivanov, M. Citrus flavonoids diosmin, myricetin and neohesperidin as inhibitors of Pseudomonas aeruginosa: evidence from antibiofilm, gene expression and in vivo analysis. Biomed Pharmacother 2024, 181, 117642. [CrossRef]
- Mustafa, S.; Akbar, M.; Khan, M. A.; Sunita, K.; Parveen, S.; Pawar, J. S.; Massey, S.; Agarwal, N. R.; Husain, S. A. Plant metabolite diosmin as the therapeutic agent in human diseases. Curr Res Pharmacol Drug Discov 2022. 3, 100122. [CrossRef]
- Yarmolinsky, L.; Nakonechny, F.; Budovsky, A.; Zeigerman, H.; Khalfin, B.; Sharon, E.; Yarmolinsky, L.; Ben-Shabat, S.; Nisnevitch, M. Antimicrobial and antiviral compounds of Phlomis viscosa poiret. Biomedicines 2023, 11, 2. [CrossRef]
- Pushkaran, A. C.; Vinod, V.; Vanuopadath, M.; Nair, S. S.; Nair, S. V.; Vasudevan, A. K.; Biswas, R.; Mohan, C. G. Combination of repurposed drug diosmin with amoxicillin-clavulanic acid causes synergistic inhibition of mycobacterial growth. Sci Rep 2019, 9, 1. [CrossRef]
- Chan, B. C. L.; Ip, M.; Gong, H.; Lui, S. L.; See, R. H.; Jolivalt, C.; Fung, K. P.; Leung, P. C.; Reiner, N. E.; Lau, C. B. S. synergistic effects of diosmetin with erythromycin against ABC transporter over-expressed methicillin-resistant Staphylococcus aureus (MRSA) RN4220/PUL5054 and inhibition of MRSA pyruvate kinase. Phytomedicine 2013, 20, 7, 611–614. [CrossRef]
- Aires, A.; Marrinhas, E.; Carvalho, R.; Dias, C.; Saavedra, M. J. Phytochemical composition and antibacterial activity of hydroalcoholic extracts of Pterospartum tridentatum and Mentha pulegium against Staphylococcus aureus isolates. Biomed Res Int 2016, 2016. [CrossRef]
- Barberis, A.; Deiana, M.; Spissu, Y.; Azara, E.; Fadda, A.; Serra, P. A.; D’Hallewin, G.; Pisano, M.; Serreli, G.; Orrù, G.; Scano, A.; Steri, D.; Sanjust, E. Antioxidant, antimicrobial, and other biological properties of pompia juice. Molecules 2020, 25, 14. [CrossRef]
- Contreras, O.; Angulo, A.; Santafé, G. Antifungal potential of isoespintanol extracted from Oxandra xylopioides diels (Annonaceae) against intrahospital isolations of Candida spp. Heliyon 2022, 8, e11110. [CrossRef]
- Guembe, M.; Cruces, R.; Peláez, T.; Mu, P.; Bouza, E. Assessment of biofilm production in candida isolates according to species and origin of infection. Enferm Infecc Microbiol Clin 2017, 35, 1, 37–40. [CrossRef]
- Kawai, A.; Yamagishi; Mikamo, H. Time-lapse tracking of Candida tropicalis biofilm formation and the antifungal efficacy of liposomal amphotericin B. Jpn. J. Infect. Dis. 2017, 70, 559–564. [CrossRef]
- Helmy, M. W.; Ghoneim, A. I.; Katary, M. A.; Elmahdy, R. K. The synergistic anti-proliferative effect of the combination of diosmin and BEZ-235 (Dactolisib) on the HCT-116 colorectal cancer cell line occurs through inhibition of the PI3K/Akt/MTOR/NF-ΚB Axis. Mol Biol Rep 2020, 47, 3, 2217–2230. [CrossRef]
- Alvarez, N. A. ´; Vicente, V.; Martínez, C. Synergistic Effect of Diosmin and Interferon-a on Metastatic Pulmonary Melanoma. Cancer Biother Radiopharm 2009, 24, 3, 347-52. [CrossRef]
- Artanti, A. N.; Jenie, R. I.; Rumiyati, R.; Meiyanto, E. Hesperidin and diosmin increased cytotoxic activity cisplatin on hepatocellular carcinoma and protect kidney cells senescence. Asian Pac J Cancer Prev 2024, 25, 12, 4247–4255. [CrossRef]
- Zeya, B.; Nafees, S.; Imtiyaz, K.; Uroog, L.; Fakhri, K. U.; Rizvi, M. A. M. Diosmin in combination with naringenin enhances apoptosis in colon cancer cells. Oncol Rep 2022, 47, 1. [CrossRef]
- Cantón, E.; Martín, E.; Espinel-Ingroff, A. Métodos estandarizados por el CLSI para el estudio de la sensibilidad a los antifúngicos (Documentos M27-A3, M38-A y M44-A). Rev Iberoam Micol 2007, 15, 1–17.
- Rodriguez-Tudela, J. L. Method for determination of minimal inhibitory concentration (MIC) by broth dilution of fermentative yeasts. Clinical Microbiology and Infection 2003, 9, 467–474. [CrossRef]
- Contreras, O.; Angulo, A.; Santafé, G. Mechanism of antifungal action of monoterpene isoespintanol against clinical isolates of Candida tropicalis. Molecules 2022, 27, 5808. [CrossRef]
- Contreras-Martínez, O. I.; Angulo-Ortíz, A.; Santafé Patiño, G.; Sierra Martinez, J.; Berrio Soto, R.; de Almeida Rodolpho, J. M.; de Godoy, K. F.; de Freitas Aníbal, F.; de Lima Fragelli, B. D. Synergistic antifungal effect and in vivo toxicity of a monoterpene isoespintanol obtained from Oxandra xylopioides Diels. Molecules 2024, 29, 18. [CrossRef]
Figure 1.
Structure of the flavonoid diosmin (DIO), diosmetin 7-O-rutinoside.
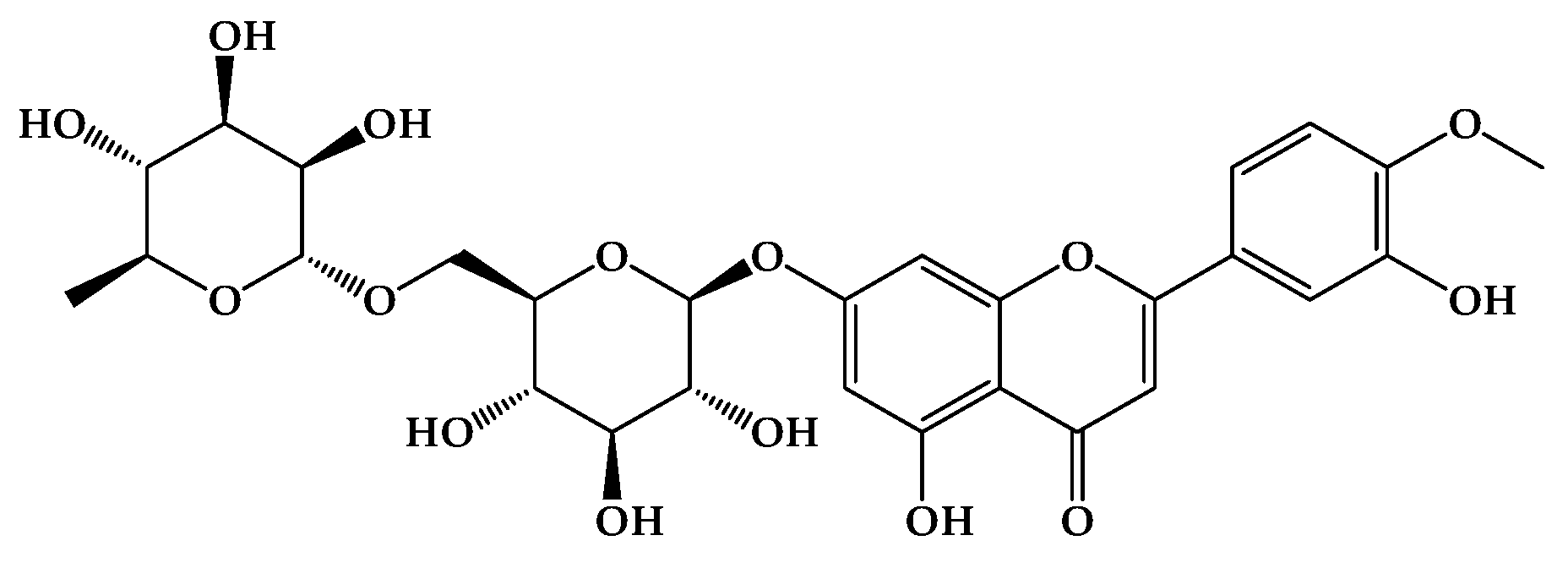
Figure 2.
Growth reduction of Candida spp. isolates exposed to DIO (MIC90 of each isolate). A strong, positive linear relationship is observed between DIO concentration and the percentage reduction in Candida spp. growth; that is, as the DIO concentration increases, so does the percentage reduction in Candida spp. growth. This is consistent with the Pearson correlation coefficient (0.82 < r < 0.99) across all isolates. Furthermore, hypothesis testing for the correlation coefficient yielded a p-value < 0.05, indicating a significant linear relationship with 95% confidence.
Figure 2.
Growth reduction of Candida spp. isolates exposed to DIO (MIC90 of each isolate). A strong, positive linear relationship is observed between DIO concentration and the percentage reduction in Candida spp. growth; that is, as the DIO concentration increases, so does the percentage reduction in Candida spp. growth. This is consistent with the Pearson correlation coefficient (0.82 < r < 0.99) across all isolates. Furthermore, hypothesis testing for the correlation coefficient yielded a p-value < 0.05, indicating a significant linear relationship with 95% confidence.
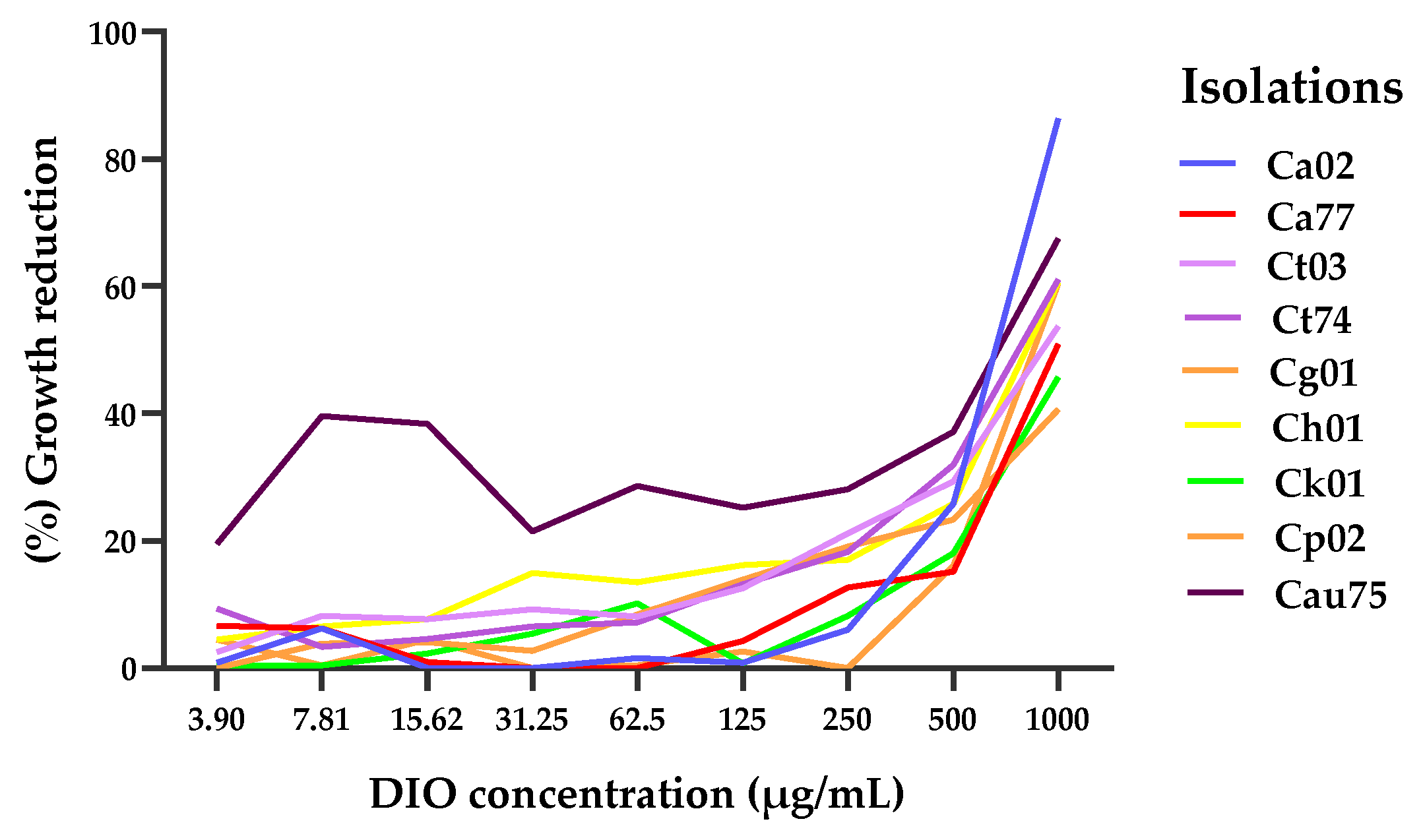
Figure 3.
Action of DIO and AFB on Candida spp. biofilms; (a) biofilm formation at 37 °C for 28 h, where an OD590 > 3 indicates strong biomass production in the biofilms. (b) percentage of biofilm reduction after 1 h of treatment with MIC90 of DIO for each isolate and AFB (2 µg/mL). The results of the ANOVA (Brown-Forsythe test) with a value of *** p < 0.001 (C. parapsilosis) and ** p < 0.01 (C. tropicalis, C. haemulonii, C. auris) and the Games-Howell's post hoc test, with a confidence level of 95%, indicate that there are significant differences between the effect of DIO and the effect of AFB on biofilm inhibition in these isolates.
Figure 3.
Action of DIO and AFB on Candida spp. biofilms; (a) biofilm formation at 37 °C for 28 h, where an OD590 > 3 indicates strong biomass production in the biofilms. (b) percentage of biofilm reduction after 1 h of treatment with MIC90 of DIO for each isolate and AFB (2 µg/mL). The results of the ANOVA (Brown-Forsythe test) with a value of *** p < 0.001 (C. parapsilosis) and ** p < 0.01 (C. tropicalis, C. haemulonii, C. auris) and the Games-Howell's post hoc test, with a confidence level of 95%, indicate that there are significant differences between the effect of DIO and the effect of AFB on biofilm inhibition in these isolates.
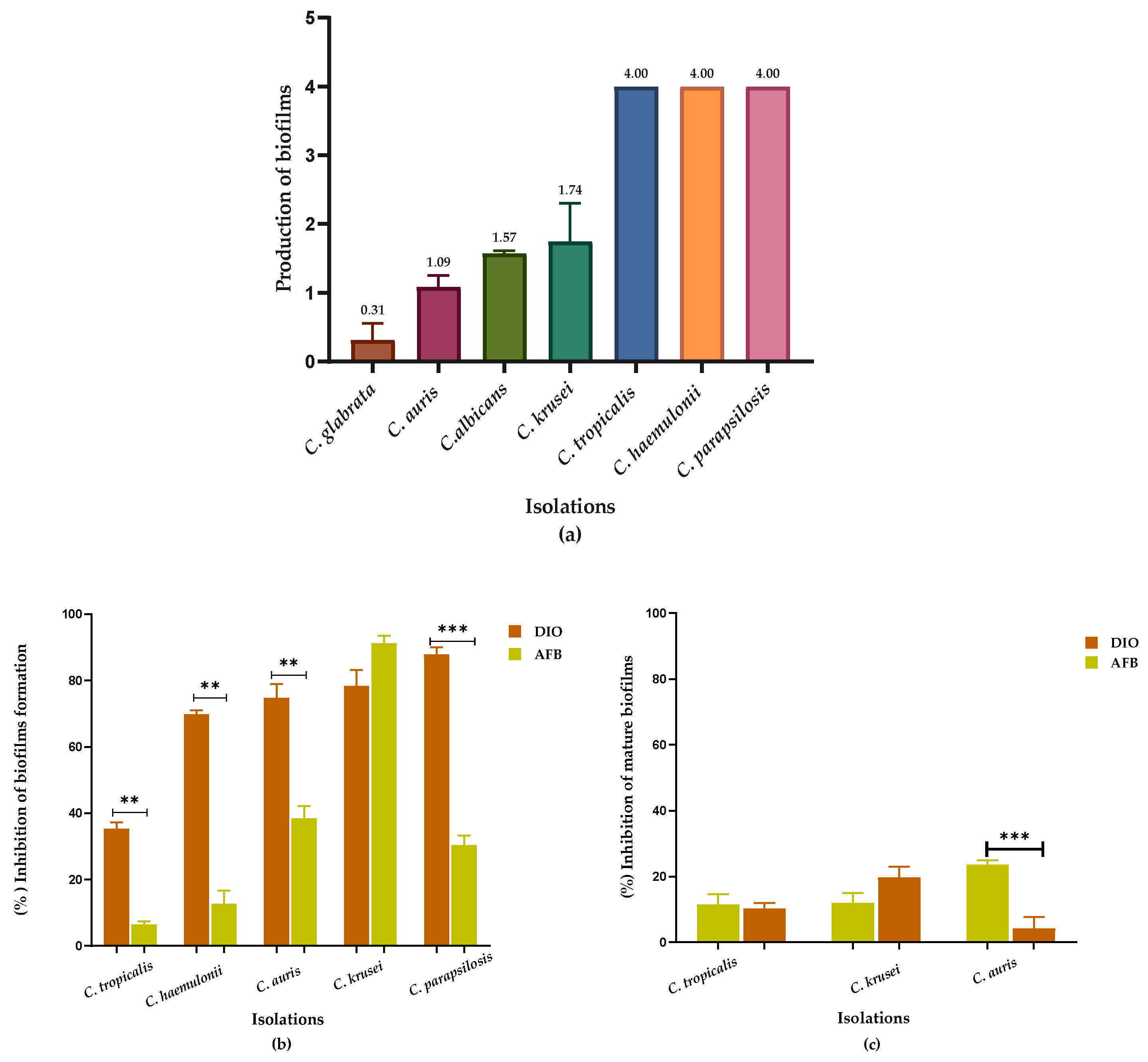
Figure 4.
Intracellular content was released at 260/280 nm as a function of time, from Candida spp. treated with DIO (MIC90 µg/mL) and FLZ (MIC90 µg/mL). Results are expressed as the absorbance of the sample (treated with DIO) minus the absorbance of the control (samples without DIO). Analyses of variance (Brown Forsythe ANOVA test) revealed p values < 0.05 (260/280 nm), the Games-Howell´s post hoc test, with a 95% confidence level, showed significant differences between the effect of DIO and FLZ on the release of intracellular material from all Candida spp. isolates evaluated, demonstrating damage to the fungal membrane permeability caused by DIO.
Figure 4.
Intracellular content was released at 260/280 nm as a function of time, from Candida spp. treated with DIO (MIC90 µg/mL) and FLZ (MIC90 µg/mL). Results are expressed as the absorbance of the sample (treated with DIO) minus the absorbance of the control (samples without DIO). Analyses of variance (Brown Forsythe ANOVA test) revealed p values < 0.05 (260/280 nm), the Games-Howell´s post hoc test, with a 95% confidence level, showed significant differences between the effect of DIO and FLZ on the release of intracellular material from all Candida spp. isolates evaluated, demonstrating damage to the fungal membrane permeability caused by DIO.
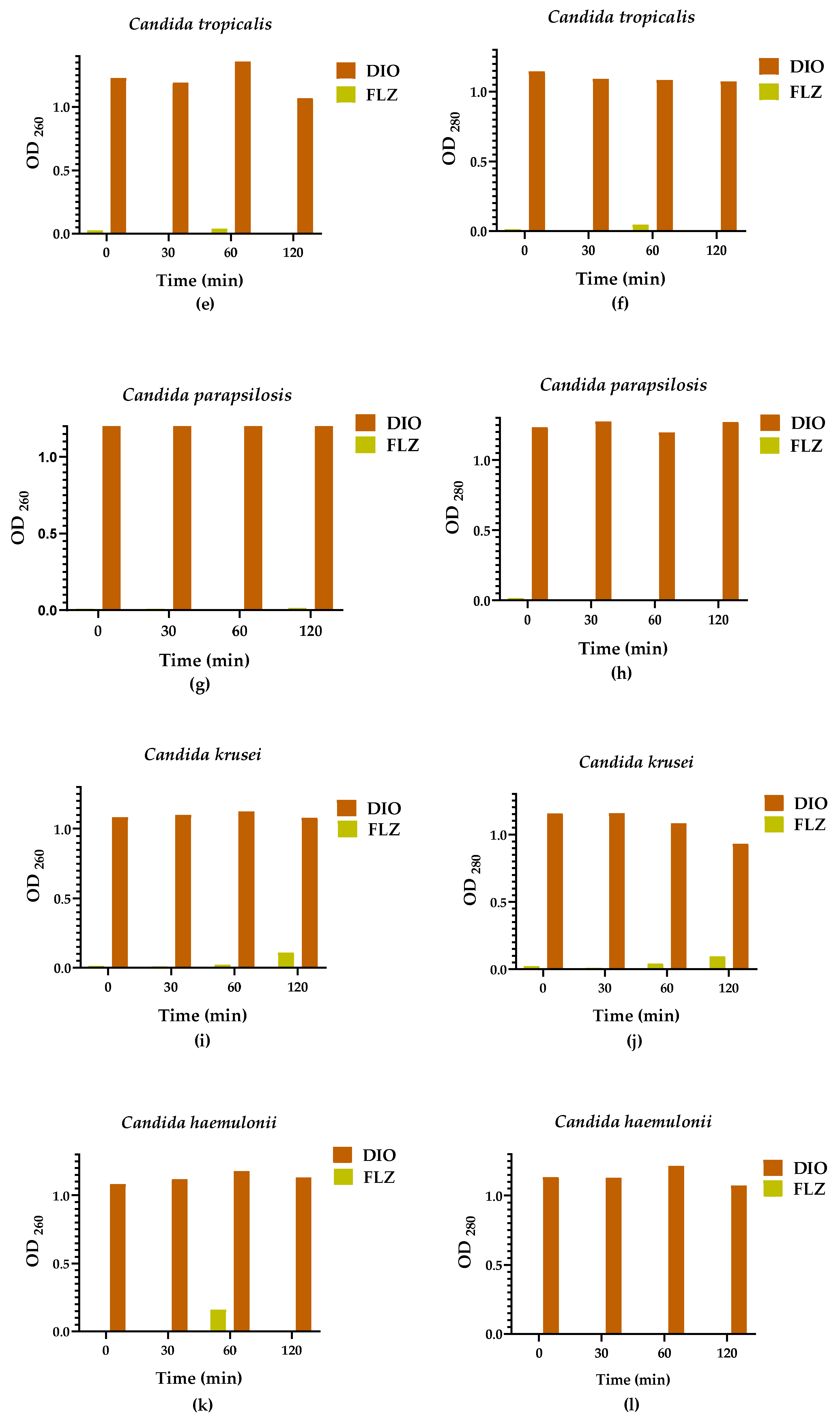
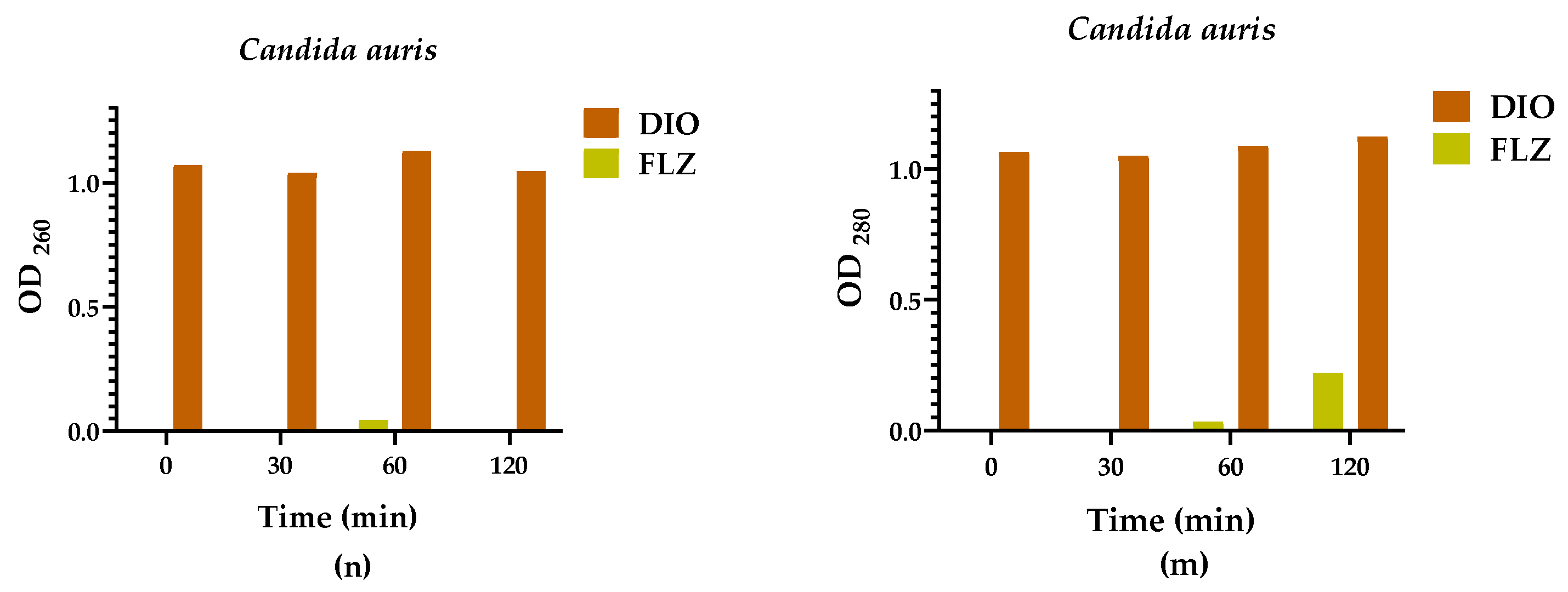
Figure 5.
Effect of DIO in combination with FLZ and AFB against Candida spp. The fractional inhibitory concentration (FCI) indices of the different clinical isolates of Candida spp., are shown.
Figure 5.
Effect of DIO in combination with FLZ and AFB against Candida spp. The fractional inhibitory concentration (FCI) indices of the different clinical isolates of Candida spp., are shown.
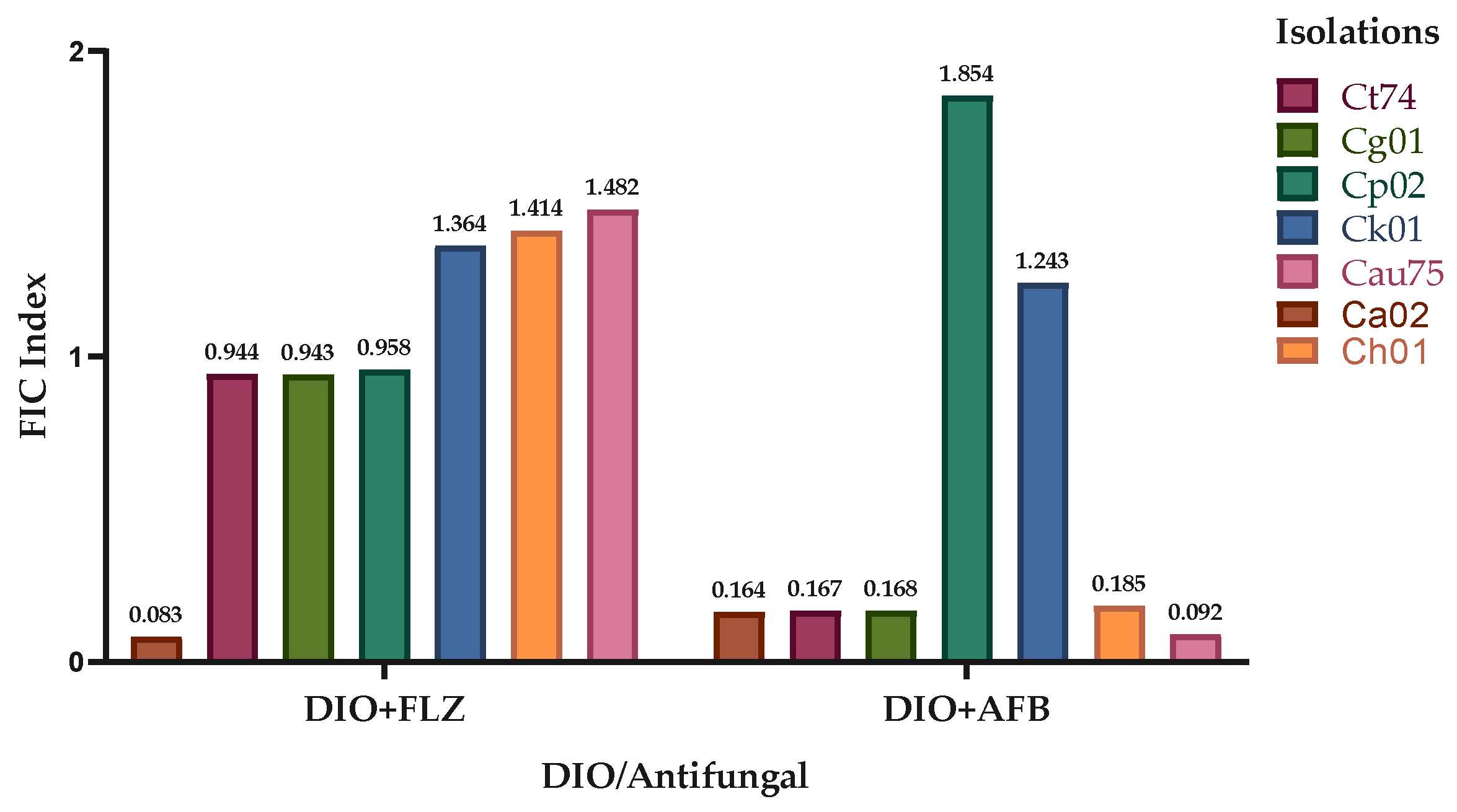

Table 1.
Minimum inhibitory concentration (MIC90) values (µg/mL) of DIO vs FLZ against Candida spp.
Table 1.
Minimum inhibitory concentration (MIC90) values (µg/mL) of DIO vs FLZ against Candida spp.
| Candida spp. | DIO MIC50 | DIO MIC90 | FLZ MIC90 |
| Ca02 | 660.4 | 1150 | 55.34 |
| Ca77 | 1079 | 1952 | 2.3 |
| Ct03 | 916 | 1756 | 154.5 |
| Ct74 | 808.8 | 1526 | 241.8 |
| Cg01 | 1199 | 2251 | 214.3 |
| Ch01 | 853.2 | 1655 | 133.9 |
| Ck01 | 1168 | 2118 | 69.99 |
| Cp02 | 947 | 1670 | 42.09 |
| Cau75 | 669.7 | 1786 | 75.65 |
Table 2.
Percentages of inhibition of biofilm formation of DIO vs AFB in Candida spp.
| Candida spp. isolates | DIO | AFB |
| C. albicans | 00.00 ± 4.39 | 00.00 ± 4.48 |
| C. glabrata | 00.00 ± 6.63 | 00.00 ± 4.77 |
| C. tropicalis | 35.35 ± 1.88 | 6.49 ± 0.92 |
| C. haemulonii | 69.81 ± 1.16 | 12.75 ± 3.92 |
| C. auris | 74.75 ± 4.22 | 38.49 ± 3.71 |
| C. krusei | 78.41 ± 4.79 | 91.26 ± 2.17 |
| C. parapsilosis | 87.85 ± 2.18 | 30.43 ±2 .78 |
Table 3.
Percentages of DIO inhibition against mature biofilms, DIO vs AFB in Candida spp.
| Candida spp. isolates | DIO | AFB |
| C. albicans | 00.00 ± 3.12 | 00.00 ± 3.14 |
| C. glabrata | 00.00 ± 5.68 | 00.00 ± 5.41 |
| C. tropicalis | 11.53 ± 3.09 | 10.2 ± 1.67 |
| C. haemulonii | 00.00 ± 0.00 | 00.00 ± 0.00 |
| C. auris | 23.65 ± 1.24 | 4.24 ± 3.48 |
| C. krusei | 11.92 ± 2.99 | 19.67 ± 3.32 |
| C. parapsilosis | 00.00 ± 0.00 | 00.00 ± 0.00 |
Table 4.
MIC90 values, individually and in combination, of DIO, FLZ and AFB against Candida spp.
| Candida spp. | MIC90 Single | MIC90 in Combination | FIC Indices | Effect | |||||
|---|---|---|---|---|---|---|---|---|---|
| FLZ | AFB | DIO | FLZ-DIO | AFB-DIO | FLZ-DIO | AFB-DIO | FLZ-DIO | AFB-DIO | |
| Ca02 | 55.34 | 9.91 | 1150 | 2.2 - 49.9 | 0.82 - 93.99 | 0.08 | 0.16 | Sng | Sng |
| Ct74 | 241.8 | 9.87 | 1526 | 112.6 – 730.6 | 1.18 - 73.32 | 0.94 | 0.17 | Sng | Sng |
| Cg01 | 214.3 | 4.11 | 2251 | 103.5 - 1036 | 0.35 - 186.9 | 0.94 | 0.17 | Sng | Sng |
| Ch01 | 133.9 | 8.89 | 1655 | 94.7 - 1170 | 0.82 - 153.5 | 1.41 | 0.18 | S.I. | Sng |
| Ck01 | 69.99 | 3.30 | 2118 | 46.9 - 1469 | 2.08 - 1298 | 1.36 | 1.24 | S.I. | S.I. |
| Cp02 | 42.09 | 4.05 | 1670 | 20.3 - 792.7 | 3.791- 1534 | 0.96 | 1.85 | Sng | S.I. |
| Cau75 | 75.65 | 18.41 | 1786 | 55.4 - 1338 | 0.853-82.89 | 1.48 | 0.09 | S.I. | Sng |
Sng: synergism; S.I.: no interaction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



