
Submitted:
07 April 2025
Posted:
09 April 2025
You are already at the latest version
Abstract
The 2024 outbreak of highly pathogenic avian influenza virus (HPAIV) H5N1 in U.S. dairy cattle presented an unprecedented scenario where the virus infected bovine mammary glands and was detected in milk, raising serious concerns for public health and the dairy industry. Unlike previously described subclinical influenza A virus (IAV) infections in cattle, H5N1 infection induced severe clinical symptoms, including respiratory distress, mastitis, and abnormal milk production. To understand the host immune responses and changes particularly in the mammary gland, we performed scRNA-seq analysis on bovine milk somatic cells (bMSC) in-vitro infected with H5N1 isolate from dairy farm. We identified ten distinct cell clusters and observed a shift toward type-2 immune responses, characterized by T-cells expressing IL13 and GATA3, and three different subtypes of epithelial cells based on expression of genes associated with milk production. Our study revealed temporal dynamics in cytokine expression, with a rapid decline in luminal epithelial cells and an increase in macrophages and dendritic cells, suggesting a role in increased antigen presentation. These findings indicate that bovine H5N1 infection triggers complex immune responses involving both pro-inflammatory and regulatory pathways. This research fills a critical gap in understanding the immune responses of bovine mammary glands to H5N1 infection and highlights the need for further investigation into therapeutic strategies for managing such outbreaks.
Keywords:
highly pathogenic avian influenza virus (HPAIV) H5N1
; Dairy cattle
; Bovine Milk Somatic Cells
; scRNA-seq
Introduction
Highly pathogenic avian influenza viruses (HPAIVs) have predominantly affected avian species, but their capacity to spill over into mammalian hosts poses a significant public health and agricultural concern [1,2,3]. The 2024 outbreak of HPAIV H5N1 in United States dairy cattle represents a novel and alarming example where the virus has infected cattle, specifically targeting cow mammary glands [1,2,3]. Unlike previous influenza A virus (IAV) infections in cattle, which are generally subclinical, the H5N1 outbreak was associated with severe clinical symptoms, including respiratory distress, reduced feed intake, abnormal milk production, and mastitis with elevated somatic cell counts [1,2,3]. More concerning is the detection of high loads infectious viral particles in cow milk, highlighting potential risks for viral transmission through dairy products [1,2,3].
Cattle with bovine H5N1 infection display neutrophilic and lymphoplasmacytic mastitis, which mirrors the immune response seen in bacterial infections of the udder [1,2,3,4,5]. This results in severe inflammation and structural damage to the mammary glands. While bovine mastitis has been extensively studied in the context of bacterial infections, there is limited understanding of how viral infections, particularly by IAV, modulate the mammary gland’s immune responses. To address this gap, we employed single-cell RNA sequencing (scRNA-seq) to profile milk somatic cells infected with bovine H5N1 (bovine-H5N1) to better understand the bovine mammary gland state during bovine H5N1 infection. We observed significant changes in luminal epithelial cells, with distinct cytokine expression patterns reflecting the temporal dynamics of the immune response.
Results
Bovine Milk Somatic Cells (bMSCs) Consist of Diverse Luminal and Immune Cells
Typically milk from healthy cattle consists of approximately 100,000 cells/ml with 30-40% viability, which is low for scRNA-seq experiments [6,7]. To overcome low viability, we enriched our collected cells three times, two times before infection and one time after infection by removing dead cells and pooling the cells from three animals. A total of 10,000 pooled cells/treatment were analyzed using 10X Genomics controller and cellranger pipeline, from which ~6900 cells/treatment were recovered with ~16,500 mean reads/cell [8].
The control and bovine-H5N1 infected cells were initially analyzed individually using the 10X genomics cellranger pipeline [8]. All data were merged together, unsupervised clustering performed using 50 principal components for Principal Components Analysis (PCA) and cells projected on two-dimensional t-distributed stochastic neighbor embedding (t-SNE) plot [9] (Figure 1). We identified 10 cell clusters and utilized the 28 selected gene markers curated from various published databases to identify the cell type of the individual cluster [10,11,12,13] (Figure 1A-B). Seven clusters, constituting ~70% of cells, were identified as immune cells with 48% as monocytes, 10% as T-cells, 7% as macrophages and 3% as dendritic cells (DCs) (Supplementary data). Other three clusters, constituting ~30% cells, were characterized as luminal epithelial cells with 13% as subset-1 expressing low casein genes, 11% as subset-2 expressing medium casein genes, and 7% as subset-3 with high casein gene expression (Figure 1B, Supplementary data).
Differential gene expression from the ScRNA-seq data further resolved the cells subsets and their functional states (Figure 1, Supplementary data). Monocytes subset 1 was comprised of cells expressing genes PKD2L1, CACNA2D3, ENO4 (corresponding to classical monocytes); monocytes subset 2 consisted of cells expressing CCDC42, ZNF713 (corresponding to non-classical monocytes); monocytes subset 3 was comprised of cells expressing genes LAMA3, RNASE12, RTKN2 (corresponding to intermediate monocytes) [10,11,12,13]. The macrophages cluster consisted of cells expressing genes CLDN1, CYP1B1, MMP1, MMP3, RBP4, CSF3, MMP12, HMGA2 which were upregulated with inflammatory stimuli [14,15,16,17,18,19,20]. The DCs cluster comprised of cells expressing genes MGP, GARNL3, RARRES2 (corresponding to plasmacytoid DCs) [21]. T-cell subset 1 was comprised of cells expressing CCL1, IL13, LTA, TNFRSF4, KLRB1, TP73 genes (corresponding to activated T-cells Th2) [22,23,24,25,26,27,28,29], whereas T-cell subset 2 consisted of expressing genes EGFL8, FBXO44, IL-15L corresponding to inhibited Th2 T-cells [30,31,32,33]. Luminal cell subset 1 was comprised of cells expressing genes ISM1, CCL19 (leukocyte chemoattractant), CLEC1A, Ubiquitin-like protein 5 (corresponding to stress response apoptosis inducing protein) [34,35,36,37,38,39]. Luminal cells subset 2 consisted of cells expressing genes ENTPD3, SRRM3, GPM6A (corresponding to breast cancer related genes) [40,41,42]. Luminal subset 3 was comprised of cells expressing DNAH9, PTPRH, DYDC1 genes (corresponding to ciliated luminal epithelial cells) [43,44,45].
Bovine-H5N1 Infection Alters the bMSCs Cellular Diversity and Their Functional State
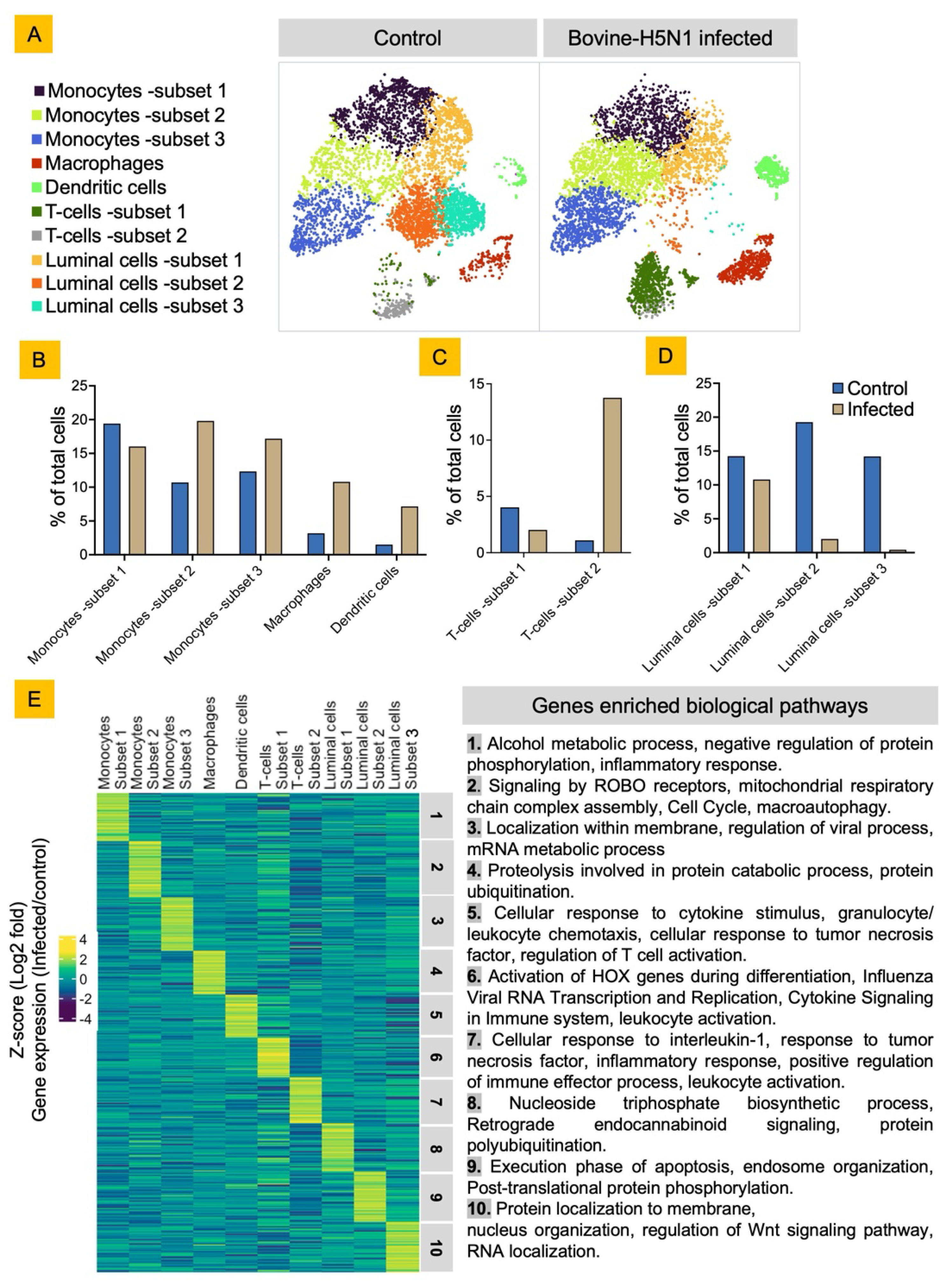
Bovine H5N1 has the distinct tropism for the epithelial cells lining the alveoli of the udder of cows [1,2]. To assess how bovine-H5N1 infection affects the bMSCs, which consists of exfoliated epithelial cells from udder alveoli, we compared the scRNA-seq data from bMSCs either mock-infected or bovine-H5N1 infected after 24 hrs post-infection [46,47] (Figure 2). We combined the reference genomes and annotations for both the bovine genome and H5N1 virus genome to identify viral reads within the cells; however, no viral RNA reads were observed in any of the cells. To validate this, we performed M gene based qRT-PCR on bovine-H5N1 infected bMSCs and MDCK cells (as a positive control) and successfully detected viral RNA in both cell types (Supplementary Figure 1A). Therefore, we concluded that a small number of cells were infected with H5N1. We then quantified the number of cells in each cluster and compared it with or without bovine-H5N1 infection. The dynamics of the cell population changed with the infection with increased immune cells and decreased luminal cells in bovine-infected cells. Only subset 1 of monocytes and luminal cells remained the same in the uninfected and infected cell populations. We assessed the change in functional state of cells by differential gene expression followed by gene ontology (GO) enrichment of the top 50 upregulated genes from each cell cluster after bovine-H5N1 infection. The monocytes subset 1, which corresponds to classical monocytes, revealed upregulation of inflammatory responses with negative regulation of protein phosphorylation process after infection. The luminal cells subset 1 which has high expression of leukocyte chemoattractant showed upregulation of nucleoside triphosphate biosynthetic process and protein polyubiquitination. The GO analysis on subset 2 and 3 of monocytes revealed upregulation of pathways related to cell cycle, macroautophagy, and regulation of viral process. In macrophages, upregulation of proteolysis involved in protein catabolic process after infection was observed. The GO analysis also indicated that bovine-H5N1 activates the dendritic cells and T-cells subsets by upregulating the cellular responses to cytokines and inflammatory stimulations. The subsets 2 and 3 of luminal epithelial cells which have higher expression of casein proteins appeared to be negatively affected by infection with decrease in their number and upregulation of apoptosis pathways (Figure 2, supplementary data).
We further assessed the immunological state of the cells by selecting genes related to Th1, Th2, Th9 and Th17 cytokines (Supplementary Figure 1B). Monocytes and dendritic cells showed strong upregulation of pro-inflammatory cytokine genes such as CSF2, IL1B, and TNF. Subset 3 of luminal cells showed high expression of IL1B and TNF with contrast to subset 2 which showed increased expression of the genes IFNG and TGFB. T-cells (subsets 1 and 2) display differential expression patterns, with subset 2 showing increased expression of IFNG, FOXP3, and TGFBR1/2 (corresponding to regulatory T-cells), while subset 1 showed markers of Th17-related responses such as RORC and IL17A. Macrophages show notable expression of MMP1, MMP3, and MMP12, which are involved in tissue remodeling.
Discussion
The 2024 outbreak of highly pathogenic avian influenza virus (HPAIV) H5N1 in US dairy cattle is unprecedented due to its unique ability to infect cow mammary glands, with high amounts of infectious viral particles found in milk [1,2,3]. This raises concerns for public health and food safety. Unlike previously described subclinical IAV infections in cattle, the H5N1 lead to severe symptoms, including reduced feed intake, respiratory distress, abnormal milk production, and mastitis with elevated somatic cell counts [1,2,3]. In healthy cattle, somatic cells contain exfoliated udder alveoli epithelial cells and immune cells, whereas in cattle with bacterial mastitis, the number of epithelial cells diminished and the number of neutrophils significantly increase [4,5,7]. Bovine-H5N1 infection in the udder exhibits neutrophilic and lymphoplasmacytic mastitis similar to bacterial infection, leading to severe inflammation and structural damage in the mammary glands [1,2,3]. These findings underscore the need for further studies to understand the role of mammary gland immune responses against viral infections.
Here, we employed scRNA-seq to profile immune cells in the bMSC to explore the state of the udder during bovine-H5N1 infection. One significant challenge in scRNA-seq of MSCs is the low viability of cells, which is further exacerbated in in vitro environments [7]. To overcome this, we removed dead cells at multiple stages of the experiment. Another major obstacle was the incomplete annotation of the bovine genome, hindering the accurate characterization of single cells and their activation states. To mitigate this, we leveraged various databases and publications to identify respective proteins, though many genes remain unannotated. Thus, to ensure confidence in cell clustering, we used a higher number of principal components in our PCA analysis, resulting in 10 distinct clusters identified with 28 gene markers [10,11,12,13]. Differential gene expression analysis revealed that the bMSCs skew toward a type-2 immune response with T-cells expressing Th2-related genes, including IL13, CCL1, and GATA3, while macrophages exhibited high expression of matrix metalloproteinases (MMP1, MMP3, MMP12), which are upregulated by Th2 cytokines [12,48,49].
When influenza A virus infects cells, it triggers a complex cascade of immune responses, including the production and release of cytokines and chemokines. This process plays a crucial role in alerting and activating the immune system to combat the viral infection [50,51,52,53]. We observed the number of immune cells after infection increased substantially with upregulation of their activation. Whereas the number of luminal epithelial cells (subset 2 and 3) expressing milk genes decreased. There was a considerable difference between cytokine gene expression among these subsets after infection, with subset 3 expressing high amount of CSF2, IL1B, and TNF genes which corresponds to the first wave of cytokines after infection, and subset 2 cells expressing genes related to the second wave of cytokines, IFNG, TGFB, and IL12B [52,53].
Macrophages and DCs are among the first to respond to infection signals. They recognize common pathogen features through pattern recognition receptors (PRRs). Upon activation, they engulf pathogens and present fragments on their surface and release additional cytokines and chemokines to recruit more immune cells [50]. We observed a similar pattern with the number of both macrophages and DCs increasing during infection and upregulation of genes corresponding to their activation. These cells likely phagocytize epithelial cells, as observed by a decrease in luminal epithelial cell populations occurring within 24 hours, a time frame too short for viral cytolysis. The cytokine release and antigen processing and presentation can further activate other immune cells such as B- and T-cells [50]. We didn’t observe B-cells in our bMSCs; however, we observed two sets of T-cells, both of which correspond to Th2 cells in different functional states: activated and inhibited. The number of inhibited T-cells (subset 1) decreased after infection with upregulation of Th17 related genes RORC, IL17A, and IL17F; however, downregulation of STAT3 (Supplementary Figure 1 and Supplementary data), which is crucial for the development and function of Th17 T-cells, implies a dysregulation of these T-cells [54,55,56]. The number of activated T-cells (subset 2) increased substantially after infection with upregulation of IFNG, STAT3, STAT4, IL9, FOXP3, and TGFBR1, TGFBR2 genes. TGF-β1 induces Foxp3 expression in naive CD4+ T cells, leading to the generation of induced regulatory T cells (iTregs) [57,58,59]. Upregulation of Foxp3 and TGF-β1 receptor genes implies that after infection these T-cells were induced to iTregs via actions of cytokines secreted from infected epithelial and innate immune cells. However, further experiments to explore the re-programming of T-cells is required.
Overall, our in vitro analysis highlights the diverse host immune responses triggered by bovine-H5N1 infection in different cell populations, particularly the upregulation of both pro-inflammatory and regulatory pathways in response to infection. Different epithelial cell subsets in bMSCs exhibit varied responses to H5N1 infection, and not all are permissive to viral replication. The infection in permissive epithelial cells leads to immune cells present in bMSCs to proliferate and secrete cytokines, contributing to inflammation and potential mastitis. These results highlight the need for further research to understand the mechanisms underlying H5N1-induced mastitis and to develop effective prevention and treatment strategies for this emerging disease threat.
Material and Methods
Cells and Viruses
Madin–Darby Canine Kidney (MDCK) cells were cultured as routine in Dulbecco’s Modified Eagle Medium (DMEM) at 37°C, supplemented with 10% fetal bovine serum (FBS) and a 1X antibiotic–antimycotic solution (ThermoFisher Scientific).
The bovine-H5N1 HPAIV (A/dairy cattle/Kansas/5/2024) was isolated from the HPAIV impacted dairy farm in Kansas and propagated in 9-days old SPF free chicken Egg and plaque tittered using MDCK as described previously [60].
Raw Milk Processing and Collection of Bovine Milk Somatic Cells (bMSCs):
Fresh raw milk (50 ml from each quarter) from three multiparous Holstein-Friesian cows was collected via hand milking at the Kansas State University Dairy Teaching and Research Center (Manhattan, KS). Milk was collected directly into the collection vial during forestripping, and milk was collected from each quarter of the udder and composited. The collected raw milk was centrifugated at 400xg for 10 min at 4°C, the supernatant and fat were removed, and the pellet was washed twice with chilled (4oC) RPMI growth media (1x RPMI 1640+ 2% FBS + 1x antibiotic-antimycotic; Thermos scientific). The viable bMSCs were enriched using Dead Cell Removal Kit (Miltenyi Biotec) and bMSCs were resuspended in warm (37oC) RPMI growth media and pooled together for virus infection.
bMSCs Infection with HPAIV Bovine-H5N1
Pooled bMSCs plated at density of 1x106 cells/well in low Ultra-Low attachment 6-well plate (Corning) were infected with 0.01 multiplicity of infection (MOI) with either bovine-H5N1 or infection media. The cells and viruses were then incubated together for 24 hrs at 37oC supplemented with 5% CO2. After 24 hrs incubation, cells were centrifuged at 400xg for 5 mins at 4oC to remove debris and ambient host and viral DNA/RNA. The cells were again subjected to Dead Cell Removal Kit (Miltenyi Biotec) for removing dead cells and debris and to reach the viable cell target of more than 70% live cells for scRNA-seq experiments.
Single-Cell Sequencing Using 10X Genomics Platform
Single-cell suspensions from both control and infected samples processed as above were subjected to single-cell RNAseq (scRNA-seq) using Chromium Next GEM Single Cell 5’ Reagent Kits v2 (10X genomics) following manufacturer’s instructions. Briefly, single-cells suspensions were loaded onto a Chromium Single Cell Chip (10x Genomics) for co-encapsulation with barcoded Gel Beads at a target capture rate of ~10000 individual cells per sample using Chromium Controller (10x Genomics). The co-encapsulation cells with barcoded Gel Beads were subjected to RT-PCR cycles for barcoding capture cell’s mRNA and cDNA synthesis. Synthesized cDNA was brought out of BSL3 using approved protocols. The cDNA was further indexed and processed for Illumina sequencing as per manufacturer’s instructions. The indexed samples were pooled together with equal concertation of DNA molecules and sequenced on an Illumina NextSeq 550 as per 10X Genomics’ instructions.
Bioinformatics Analysis
The sequencing process yielded raw fastq files which were used for input for cellranger package (10X genomics) for scRNA-seq analysis [8]. To quantify and align reads to a host and virus genome, the bovine reference and annotation (ARS-UCD1.2/bosTau9) was combined with A/Dairy Cattle/Kansas/5/ (NCBI Genbank ID PP732373-80) genome and annotation files. The resulting aligned and analyzed files were visualized and further analyzed using Loupe Brower (10X Genomics) as follows. The processed data was cleaned up by removing cells with less than 5000 reads and unique molecular identifier (UMI) barcode, followed by re-clustering using 50 principal components in PCA and 50 neighbors for t-distributed stochastic neighbor embedding (t-SNE) plot [9]. (Figure 1). Identification of cells in the cultures was performed by a literature research and reported gene markers for single-cell types [10,11,12,13]. Each cluster was then re-named based on the cell type.
Differential gene expression between the cluster as well as samples was performed using Loupe Brower (10X genomics) [8]. The resulting genes matrixes were visualized using pheatmap [61]. The top 50 upregulated genes after infection were further analyzed for gene function enrichment using Metascape website [62]. To quantify the cell number for each cluster in each sample, individual clusters were selected and then spitted into samples ID. The resulted count matrix for cells was then visualized using Prism v10.2.2 (GraphPad).
RNA Extraction and qPCR for Viral RNA Detection
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgment and Funding
We thank Dr. Billy Brown and the staff of the KSU Dairy Teaching and Research Center for sample collection and coordination, Dr. Natasha N. Gaudreault for reviewing and providing feedback on the manuscript, Karinne Cortes, Catherine Hickman, Christine Huncovsky, and Michelle Edie for excellent administrative support, Yonghai Li for technical support and all BRI, KSU staff for management and organization of the BSL3 facility. We also like to thank KSU Beocat Director Daniel Andresen and Beocat Systems Administrator Nathan Wells for providing valuable technical expertise. Funding for this study was partially provided through grants from the National Bio and Agro-Defense Facility (NBAF) Transition Fund from the State of Kansas, the MCB Core of the Center of Emerging and Zoonotic Infectious Diseases (CEZID) from National Institute of General Medical Sciences (NIGMS) under award number P20GM130448 (JAR, IM), the NIAID supported Centers of Excellence for Influenza Research and Response (CEIRR, contract number 75N93021C00016 to JAR). The computing for this project was performed on the Beocat Research Cluster at Kansas State University, which is funded in part by NSF grants CNS-1006860, EPS-1006860, EPS-0919443, ACI-1440548, CHE-1726332, and NIH P20GM113109.
Conflicts of Interest
The J.A.R. laboratory received support from Tonix Pharmaceuticals, Xing Technologies, and Zoetis, outside of the reported work. J.A.R. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections, owned by Kansas State University.
References
- Halwe, N. J.; Cool, K.; Breithaupt, A.; Schön, J.; Trujillo, J. D.; Nooruzzaman, M.; Kwon, T.; Ahrens, A. K.; Britzke, T.; McDowell, C. D.; Piesche, R.; Singh, G.; Pinho dos Reis, V.; Kafle, S.; Pohlmann, A.; Gaudreault, N. N.; Corleis, B.; Ferreyra, F. M.; Carossino, M.; Balasuriya, U. B. R.; Hensley, L.; Morozov, I.; Covaleda, L. M.; Diel, D.; Ulrich, L.; Hoffmann, D.; Beer, M.; Richt, J. A. H5N1 Clade 2.3.4.4b Dynamics in Experimentally Infected Calves and Cows. Nature 2024, 1–3. [CrossRef]
- Caserta, L. C.; Frye, E. A.; Butt, S. L.; Laverack, M.; Nooruzzaman, M.; Covaleda, L. M.; Thompson, A. C.; Koscielny, M. P.; Cronk, B.; Johnson, A.; Kleinhenz, K.; Edwards, E. E.; Gomez, G.; Hitchener, G.; Martins, M.; Kapczynski, D. R.; Suarez, D. L.; Alexander Morris, E. R.; Hensley, T.; Beeby, J. S.; Lejeune, M.; Swinford, A. K.; Elvinger, F.; Dimitrov, K. M.; Diel, D. G. Spillover of Highly Pathogenic Avian Influenza H5N1 Virus to Dairy Cattle. Nature 2024, 634 (8034), 669–676. [CrossRef]
- Baker, A. L.; Arruda, B.; Palmer, M. V.; Boggiatto, P.; Davila, K. S.; Buckley, A.; Zanella, G. C.; Snyder, C. A.; Anderson, T. K.; Hutter, C. R.; Nguyen, T.-Q.; Markin, A.; Lantz, K.; Posey, E. A.; Torchetti, M. K.; Robbe-Austerman, S.; Magstadt, D. R.; Gorden, P. J. Dairy Cows Inoculated with Highly Pathogenic Avian Influenza Virus H5N1. Nature 2024, 1–3. [CrossRef]
- Thompson-Crispi, K.; Atalla, H.; Miglior, F.; Mallard, B. A. Bovine Mastitis: Frontiers in Immunogenetics. Front. Immunol. 2014, 5, 493. [CrossRef]
- Sordillo, L. M.; Streicher, K. L. Mammary Gland Immunity and Mastitis Susceptibility. J. Mammary Gland Biol. Neoplasia 2002, 7 (2), 135–146. [CrossRef]
- Li, N.; Richoux, R.; Perruchot, M.-H.; Boutinaud, M.; Mayol, J.-F.; Gagnaire, V. Flow Cytometry Approach to Quantify the Viability of Milk Somatic Cell Counts after Various Physico-Chemical Treatments. PLoS ONE 2015, 10 (12), e0146071. [CrossRef]
- Cinar, M.; Serbester, U.; Ceyhan, A.; Gorgulu, M. Effect of Somatic Cell Count on Milk Yield and Composition of First and Second Lactation Dairy Cows. Ital. J. Anim. Sci. 2015, 14 (1), 3646. [CrossRef]
- Zheng, G. X. Y.; Terry, J. M.; Belgrader, P.; Ryvkin, P.; Bent, Z. W.; Wilson, R.; Ziraldo, S. B.; Wheeler, T. D.; McDermott, G. P.; Zhu, J.; Gregory, M. T.; Shuga, J.; Montesclaros, L.; Underwood, J. G.; Masquelier, D. A.; Nishimura, S. Y.; Schnall-Levin, M.; Wyatt, P. W.; Hindson, C. M.; Bharadwaj, R.; Wong, A.; Ness, K. D.; Beppu, L. W.; Deeg, H. J.; McFarland, C.; Loeb, K. R.; Valente, W. J.; Ericson, N. G.; Stevens, E. A.; Radich, J. P.; Mikkelsen, T. S.; Hindson, B. J.; Bielas, J. H. Massively Parallel Digital Transcriptional Profiling of Single Cells. Nat. Commun. 2017, 8 (1), 14049. [CrossRef]
- Maaten, L. van der; Hinton, G. Visualizing Data Using T-SNE. J. Mach. Learn. Res. 2008, 9 (86), 2579–2605.
- Karlsson, M.; Zhang, C.; Méar, L.; Zhong, W.; Digre, A.; Katona, B.; Sjöstedt, E.; Butler, L.; Odeberg, J.; Dusart, P.; Edfors, F.; Oksvold, P.; von Feilitzen, K.; Zwahlen, M.; Arif, M.; Altay, O.; Li, X.; Ozcan, M.; Mardinoglu, A.; Fagerberg, L.; Mulder, J.; Luo, Y.; Ponten, F.; Uhlén, M.; Lindskog, C. A Single–Cell Type Transcriptomics Map of Human Tissues. Sci. Adv. 2021, 7 (31), eabh2169. [CrossRef]
- Becker, D.; Weikard, R.; Hadlich, F.; Kühn, C. Single-Cell RNA Sequencing of Freshly Isolated Bovine Milk Cells and Cultured Primary Mammary Epithelial Cells. Sci. Data 2021, 8 (1), 177. [CrossRef]
- Szabo, P. A.; Levitin, H. M.; Miron, M.; Snyder, M. E.; Senda, T.; Yuan, J.; Cheng, Y. L.; Bush, E. C.; Dogra, P.; Thapa, P.; Farber, D. L.; Sims, P. A. Single-Cell Transcriptomics of Human T Cells Reveals Tissue and Activation Signatures in Health and Disease. Nat. Commun. 2019, 10 (1), 4706. [CrossRef]
- Zorc, M.; Dolinar, M.; Dovč, P. A Single-Cell Transcriptome of Bovine Milk Somatic Cells. Genes 2024, 15 (3), 349. [CrossRef]
- Van den Bossche, J.; Laoui, D.; Morias, Y.; Movahedi, K.; Raes, G.; De Baetselier, P.; Van Ginderachter, J. A. Claudin-1, Claudin-2 and Claudin-11 Genes Differentially Associate with Distinct Types of Anti-Inflammatory Macrophages In Vitro and with Parasite- and Tumour-Elicited Macrophages In Vivo. Scand. J. Immunol. 2012, 75 (6), 588–598. [CrossRef]
- Šmerdová, L.; Svobodová, J.; Kabátková, M.; Kohoutek, J.; Blažek, D.; Machala, M.; Vondráček, J. Upregulation of CYP1B1 Expression by Inflammatory Cytokines Is Mediated by the P38 MAP Kinase Signal Transduction Pathway. Carcinogenesis 2014, 35 (11), 2534–2543. [CrossRef]
- Gao, D.; Bing, C. Macrophage-Induced Expression and Release of Matrix Metalloproteinase 1 and 3 by Human Preadipocytes Is Mediated by IL-1β via Activation of MAPK Signaling. J. Cell. Physiol. 2011, 226 (11), 2869–2880. [CrossRef]
- Broch, M.; Ramírez, R.; Auguet, M. T.; Alcaide, M. J.; Aguilar, C.; Garcia-España, A.; Richart, C. Macrophages Are Novel Sites of Expression and Regulation of Retinol Binding Protein-4 (RBP4). Physiol. Res. 2010, 59 (2), 299–303. [CrossRef]
- Hollmén, M.; Karaman, S.; Schwager, S.; Lisibach, A.; Christiansen, A. J.; Maksimow, M.; Varga, Z.; Jalkanen, S.; Detmar, M. G-CSF Regulates Macrophage Phenotype and Associates with Poor Overall Survival in Human Triple-Negative Breast Cancer. Oncoimmunology 2015, 5 (3), e1115177. [CrossRef]
- Wang, X.; Wang, J.; Zhao, J.; Wang, H.; Chen, J.; Wu, J. HMGA2 Facilitates Colorectal Cancer Progression via STAT3-Mediated Tumor-Associated Macrophage Recruitment. Theranostics 2022, 12 (2), 963–975. [CrossRef]
- Aristorena, M.; Gallardo-Vara, E.; Vicen, M.; de Las Casas-Engel, M.; Ojeda-Fernandez, L.; Nieto, C.; Blanco, F. J.; Valbuena-Diez, A. C.; Botella, L. M.; Nachtigal, P.; Corbi, A. L.; Colmenares, M.; Bernabeu, C. MMP-12, Secreted by Pro-Inflammatory Macrophages, Targets Endoglin in Human Macrophages and Endothelial Cells. Int. J. Mol. Sci. 2019, 20 (12), 3107. [CrossRef]
- RARRES2 retinoic acid receptor responder 2 [Homo sapiens (human)] - Gene - NCBI. https://www.ncbi.nlm.nih.gov/gene/5919 (accessed 2024-10-15).
- Kokubo, K.; Onodera, A.; Kiuchi, M.; Tsuji, K.; Hirahara, K.; Nakayama, T. Conventional and Pathogenic Th2 Cells in Inflammation, Tissue Repair, and Fibrosis. Front. Immunol. 2022, 13. [CrossRef]
- Upadhyay, V.; Fu, Y.-X. Lymphotoxin Signaling in Immune Homeostasis and the Control of Microorganisms. Nat. Rev. Immunol. 2013, 13 (4), 270–279. [CrossRef]
- Yang, K.; Liang, Y.; Sun, Z.; Liu, L.; Liao, J.; Xu, H.; Zhu, M.; Fu, Y.-X.; Peng, H. T Cell-Derived Lymphotoxin Limits Th1 Response during HSV-1 Infection. Sci. Rep. 2018, 8 (1), 17727. [CrossRef]
- Schreiber, T. H.; Wolf, D.; Bodero, M.; Gonzalez, L.; Podack, E. R. T Cell Costimulation by TNFRSF4 and TNFRSF25 in the Context of Vaccination. J. Immunol. Baltim. Md 1950 2012, 189 (7), 3311–3318. [CrossRef]
- Fergusson, J. R.; Hühn, M. H.; Swadling, L.; Walker, L. J.; Kurioka, A.; Llibre, A.; Bertoletti, A.; Holländer, G.; Newell, E. W.; Davis, M. M.; Sverremark-Ekström, E.; Powrie, F.; Capone, S.; Folgori, A.; Barnes, E.; Willberg, C. B.; Ussher, J. E.; Klenerman, P. CD161int CD8+ T Cells: A Novel Population of Highly Functional, Memory CD8+ T Cells Enriched within the Gut. Mucosal Immunol. 2016, 9 (2), 401–413. [CrossRef]
- Truong, K.-L.; Schlickeiser, S.; Vogt, K.; Boës, D.; Stanko, K.; Appelt, C.; Streitz, M.; Grütz, G.; Stobutzki, N.; Meisel, C.; Iwert, C.; Tomiuk, S.; Polansky, J. K.; Pascher, A.; Babel, N.; Stervbo, U.; Sauer, I.; Gerlach, U.; Sawitzki, B. Killer-like Receptors and GPR56 Progressive Expression Defines Cytokine Production of Human CD4+ Memory T Cells. Nat. Commun. 2019, 10 (1), 2263. [CrossRef]
- Cadilha, B. L.; Benmebarek, M.-R.; Dorman, K.; Oner, A.; Lorenzini, T.; Obeck, H.; Vänttinen, M.; Di Pilato, M.; Pruessmann, J. N.; Stoiber, S.; Huynh, D.; Märkl, F.; Seifert, M.; Manske, K.; Suarez-Gosalvez, J.; Zeng, Y.; Lesch, S.; Karches, C. H.; Heise, C.; Gottschlich, A.; Thomas, M.; Marr, C.; Zhang, J.; Pandey, D.; Feuchtinger, T.; Subklewe, M.; Mempel, T. R.; Endres, S.; Kobold, S. Combined Tumor-Directed Recruitment and Protection from Immune Suppression Enable CAR T Cell Efficacy in Solid Tumors. Sci. Adv. 2021, 7 (24), eabi5781. [CrossRef]
- Miller, M. D.; Krangel, M. S. The Human Cytokine I-309 Is a Monocyte Chemoattractant. Proc. Natl. Acad. Sci. 1992, 89 (7), 2950–2954. [CrossRef]
- Shen, J. Z.; Qiu, Z.; Wu, Q.; Finlay, D.; Garcia, G.; Sun, D.; Rantala, J.; Barshop, W.; Hope, J. L.; Gimple, R. C.; Sangfelt, O.; Bradley, L. M.; Wohlschlegel, J.; Rich, J. N.; Spruck, C. FBXO44 Promotes DNA Replication-Coupled Repetitive Element Silencing in Cancer Cells. Cell 2021, 184 (2), 352-369.e23. [CrossRef]
- Yamaguchi, T.; Chang, C. J.; Karger, A.; Keller, M.; Pfaff, F.; Wangkahart, E.; Wang, T.; Secombes, C. J.; Kimoto, A.; Furihata, M.; Hashimoto, K.; Fischer, U.; Dijkstra, J. M. Ancient Cytokine Interleukin 15-Like (IL-15L) Induces a Type 2 Immune Response. Front. Immunol. 2020, 11. [CrossRef]
- Choi, H.-J.; Yoon, T.-D.; Muhammad, I.; Jeong, M.-H.; Lee, J.; Baek, S.-Y.; Kim, B.-S.; Yoon, S. Regulatory Role of Mouse Epidermal Growth Factor-like Protein 8 in Thymic Epithelial Cells. Biochem. Biophys. Res. Commun. 2012, 425 (2), 250–255. [CrossRef]
- Subhan, F.; Yoon, T.-D.; Choi, H. J.; Muhammad, I.; Lee, J.; Hong, C.; Oh, S.-O.; Baek, S.-Y.; Kim, B.-S.; Yoon, S. Epidermal Growth Factor-like Domain 8 Inhibits the Survival and Proliferation of Mouse Thymocytes. Int. J. Mol. Med. 2013, 32 (4), 952–958. [CrossRef]
- Drouin, M.; Saenz, J.; Gauttier, V.; Evrard, B.; Teppaz, G.; Pengam, S.; Mary, C.; Desselle, A.; Thepenier, V.; Wilhelm, E.; Merieau, E.; Ligeron, C.; Girault, I.; Lopez, M.-D.; Fourgeux, C.; Sinha, D.; Baccelli, I.; Moreau, A.; Louvet, C.; Josien, R.; Poschmann, J.; Poirier, N.; Chiffoleau, E. CLEC-1 Is a Death Sensor That Limits Antigen Cross-Presentation by Dendritic Cells and Represents a Target for Cancer Immunotherapy. Sci. Adv. 2022, 8 (46), eabo7621. [CrossRef]
- Makusheva, Y.; Chung, S.-H.; Akitsu, A.; Maeda, N.; Maruhashi, T.; Ye, X.-Q.; Kaifu, T.; Saijo, S.; Sun, H.; Han, W.; Tang, C.; Iwakura, Y. The C-Type Lectin Receptor Clec1A Plays an Important Role in the Development of Experimental Autoimmune Encephalomyelitis by Enhancing Antigen Presenting Ability of Dendritic Cells and Inducing Inflammatory Cytokine IL-17. Exp. Anim. 2022, 71 (3), 288–304. [CrossRef]
- Yan, Y.; Chen, R.; Wang, X.; Hu, K.; Huang, L.; Lu, M.; Hu, Q. CCL19 and CCR7 Expression, Signaling Pathways, and Adjuvant Functions in Viral Infection and Prevention. Front. Cell Dev. Biol. 2019, 7. [CrossRef]
- Wang, W.; Hawkridge, A. M.; Ma, Y.; Zhang, B.; Mangrum, J. B.; Hassan, Z. H.; He, T.; Blat, S.; Guo, C.; Zhou, H.; Liu, J.; Wang, X.-Y.; Fang, X. Ubiquitin-like Protein 5 Is a Novel Player in the UPR-PERK Arm and ER Stress-Induced Cell Death. J. Biol. Chem. 2023, 299 (7), 104915. [CrossRef]
- Chen, M.; Zhang, Y.; Yu, V. C.; Chong, Y.-S.; Yoshioka, T.; Ge, R. Isthmin Targets Cell-Surface GRP78 and Triggers Apoptosis via Induction of Mitochondrial Dysfunction. Cell Death Differ. 2014, 21 (5), 797–810. [CrossRef]
- Hu, M.; Zhang, X.; Hu, C.; Teng, T.; Tang, Q.-Z. A Brief Overview about the Adipokine: Isthmin-1. Front. Cardiovasc. Med. 2022, 9, 939757. [CrossRef]
- Hennessy, B. T.; Gonzalez-Angulo, A.-M.; Stemke-Hale, K.; Gilcrease, M. Z.; Krishnamurthy, S.; Lee, J.-S.; Fridlyand, J.; Sahin, A.; Agarwal, R.; Joy, C.; Liu, W.; Stivers, D.; Baggerly, K.; Carey, M.; Lluch, A.; Monteagudo, C.; He, X.; Weigman, V.; Fan, C.; Palazzo, J.; Hortobagyi, G. N.; Nolden, L. K.; Wang, N. J.; Valero, V.; Gray, J. W.; Perou, C. M.; Mills, G. B. Characterization of a Naturally Occurring Breast Cancer Subset Enriched in Epithelial-to-Mesenchymal Transition and Stem Cell Characteristics. Cancer Res. 2009, 69 (10), 4116–4124. [CrossRef]
- Lee, N. S.; Evgrafov, O. V.; Souaiaia, T.; Bonyad, A.; Herstein, J.; Lee, J. Y.; Kim, J.; Ning, Y.; Sixto, M.; Weitz, A. C.; Lenz, H.-J.; Wang, K.; Knowles, J. A.; Press, M. F.; Salvaterra, P. M.; Shung, K. K.; Chow, R. H. Non-Coding RNAs Derived from an Alternatively Spliced REST Transcript (REST-003) Regulate Breast Cancer Invasiveness. Sci. Rep. 2015, 5, 11207. [CrossRef]
- Zhou, H.; Li, J.; Zhang, Z.; Ye, R.; Shao, N.; Cheang, T.; Wang, S. RING1 and YY1 Binding Protein Suppresses Breast Cancer Growth and Metastasis. Int. J. Oncol. 2016, 49 (6), 2442–2452. [CrossRef]
- Yamashita, H.; Kotani, T.; Park, J.; Murata, Y.; Okazawa, H.; Ohnishi, H.; Ku, Y.; Matozaki, T. Role of the Protein Tyrosine Phosphatase Shp2 in Homeostasis of the Intestinal Epithelium. PLoS ONE 2014, 9 (3), e92904. [CrossRef]
- Fliegauf, M.; Olbrich, H.; Horvath, J.; Wildhaber, J. H.; Zariwala, M. A.; Kennedy, M.; Knowles, M. R.; Omran, H. Mislocalization of DNAH5 and DNAH9 in Respiratory Cells from Patients with Primary Ciliary Dyskinesia. Am. J. Respir. Crit. Care Med. 2005, 171 (12), 1343–1349. [CrossRef]
- Li, S.; Qiao, Y.; Di, Q.; Le, X.; Zhang, L.; Zhang, X.; Zhang, C.; Cheng, J.; Zong, S.; Koide, S. S.; Miao, S.; Wang, L. Interaction of SH3P13 and DYDC1 Protein: A Germ Cell Component That Regulates Acrosome Biogenesis during Spermiogenesis. Eur. J. Cell Biol. 2009, 88 (9), 509–520. [CrossRef]
- Boutinaud, M.; Herve, L.; Lollivier, V. Mammary Epithelial Cells Isolated from Milk Are a Valuable, Non-Invasive Source of Mammary Transcripts. Front. Genet. 2015, 6, 323. [CrossRef]
- Webster, H. H.; Lengi, A. J.; Corl, B. A. Short Communication: Mammary Epithelial Cell Exfoliation Increases as Milk Yield Declines, Lactation Progresses, and Parity Increases. JDS Commun. 2024. [CrossRef]
- Singh, G.; García-Bernalt Diego, J.; Warang, P.; Park, S.-C.; Chang, L. A.; Noureddine, M.; Laghlali, G.; Bykov, Y.; Prellberg, M.; Yan, V.; Singh, S.; Pache, L.; Cuadrado-Castano, S.; Webb, B.; García-Sastre, A.; Schotsaert, M. Outcome of SARS-CoV-2 Reinfection Depends on Genetic Background in Female Mice. Nat. Commun. 2024, 15 (1), 10178. [CrossRef]
- Singh, G.; Warang, P.; García-Bernalt Diego, J.; Chang, L.; Bykov, Y.; Singh, S.; Pache, L.; Cuadrado-Castano, S.; Webb, B.; Garcia-Sastre, A.; Schotsaert, M. Host Immune Responses Associated with SARS-CoV-2 Omicron Infection Result in Protection or Pathology during Reinfection Depending on Mouse Genetic Background. Res. Sq. 2023, rs.3.rs-3637405. [CrossRef]
- Charles A Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M. J. Principles of Innate and Adaptive Immunity. In Immunobiology: The Immune System in Health and Disease. 5th edition; Garland Science, 2001.
- Ma, W.; Belisle, S. E.; Mosier, D.; Li, X.; Stigger-Rosser, E.; Liu, Q.; Qiao, C.; Elder, J.; Webby, R.; Katze, M. G.; Richt, J. A. 2009 Pandemic H1N1 Influenza Virus Causes Disease and Upregulation of Genes Related to Inflammatory and Immune Responses, Cell Death, and Lipid Metabolism in Pigs. J. Virol. 2011, 85 (22), 11626–11637. [CrossRef]
- Brydon, E. W. A.; Morris, S. J.; Sweet, C. Role of Apoptosis and Cytokines in Influenza Virus Morbidity. FEMS Microbiol. Rev. 2005, 29 (4), 837–850. [CrossRef]
- Gu, Y.; Zuo, X.; Zhang, S.; Ouyang, Z.; Jiang, S.; Wang, F.; Wang, G. The Mechanism behind Influenza Virus Cytokine Storm. Viruses 2021, 13 (7), 1362. [CrossRef]
- Yahia-Cherbal, H.; Rybczynska, M.; Lovecchio, D.; Stephen, T.; Lescale, C.; Placek, K.; Larghero, J.; Rogge, L.; Bianchi, E. NFAT Primes the Human RORC Locus for RORγt Expression in CD4+ T Cells. Nat. Commun. 2019, 10 (1), 4698. [CrossRef]
- Capone, A.; Volpe, E. Transcriptional Regulators of T Helper 17 Cell Differentiation in Health and Autoimmune Diseases. Front. Immunol. 2020, 11. [CrossRef]
- Egwuagu, C. E. STAT3 in CD4+ T Helper Cell Differentiation and Inflammatory Diseases. Cytokine 2009, 47 (3), 149. [CrossRef]
- Wang, J.; Zhao, X.; Wan, Y. Y. Intricacies of TGF-β Signaling in Treg and Th17 Cell Biology. Cell. Mol. Immunol. 2023, 20 (9), 1002–1022. [CrossRef]
- O’Malley, J. T.; Sehra, S.; Thieu, V. T.; Yu, Q.; Chang, H.-C.; Stritesky, G. L.; Nguyen, E. T.; Mathur, A. N.; Levy, D. E.; Kaplan, M. H. Signal Transducer and Activator of Transcription 4 Limits the Development of Adaptive Regulatory T Cells. Immunology 2009, 127 (4), 587. [CrossRef]
- Alvarez, F.; Liu, Z.; Bay, A.; Piccirillo, C. A. Deciphering the Developmental Trajectory of Tissue-Resident Foxp3+ Regulatory T Cells. Front. Immunol. 2024, 15. [CrossRef]
- Singh, G.; Trujillo, J. D.; McDowell, C. D.; Matias-Ferreyra, F.; Kafle, S.; Kwon, T.; Gaudreault, N. N.; Fitz, I.; Noll, L.; Morozov, I.; Retallick, J.; Richt, J. A. Detection and Characterization of H5N1 HPAIV in Environmental Samples from a Dairy Farm. Virus Genes 2024, 60 (5), 517–527. [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps, 2019. https://cran.r-project.org/web/packages/pheatmap/index.html (accessed 2024-09-14).
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A. H.; Tanaseichuk, O.; Benner, C.; Chanda, S. K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10 (1), 1523. [CrossRef]
Figure 1.
Single-cell RNA-seq analysis of bovine milk somatic cells (bMSCs). (A) t-SNE embeddings of merged scRNA-seq profiles from in-vitro uninfected and bovine-H5N1 infected bMSCs collected from three healthy cattle. Distinct cell populations are represented by different colors, and assigned the labels based on expression of selected gene markers. (B) Heatmap showing z-scored average expression of curated cell marker genes that had a fold change >1 and p < 0.05 for at least one cluster. Genes are ordered by the cluster in which they have the highest expression. (C) Heatmap displaying z-scored average expression of top 10 genes in each cluster that had a fold change >1 expressed and p < 0.05 for at least one cluster. Genes are organized by cluster in which they have highest expression.
Figure 1.
Single-cell RNA-seq analysis of bovine milk somatic cells (bMSCs). (A) t-SNE embeddings of merged scRNA-seq profiles from in-vitro uninfected and bovine-H5N1 infected bMSCs collected from three healthy cattle. Distinct cell populations are represented by different colors, and assigned the labels based on expression of selected gene markers. (B) Heatmap showing z-scored average expression of curated cell marker genes that had a fold change >1 and p < 0.05 for at least one cluster. Genes are ordered by the cluster in which they have the highest expression. (C) Heatmap displaying z-scored average expression of top 10 genes in each cluster that had a fold change >1 expressed and p < 0.05 for at least one cluster. Genes are organized by cluster in which they have highest expression.
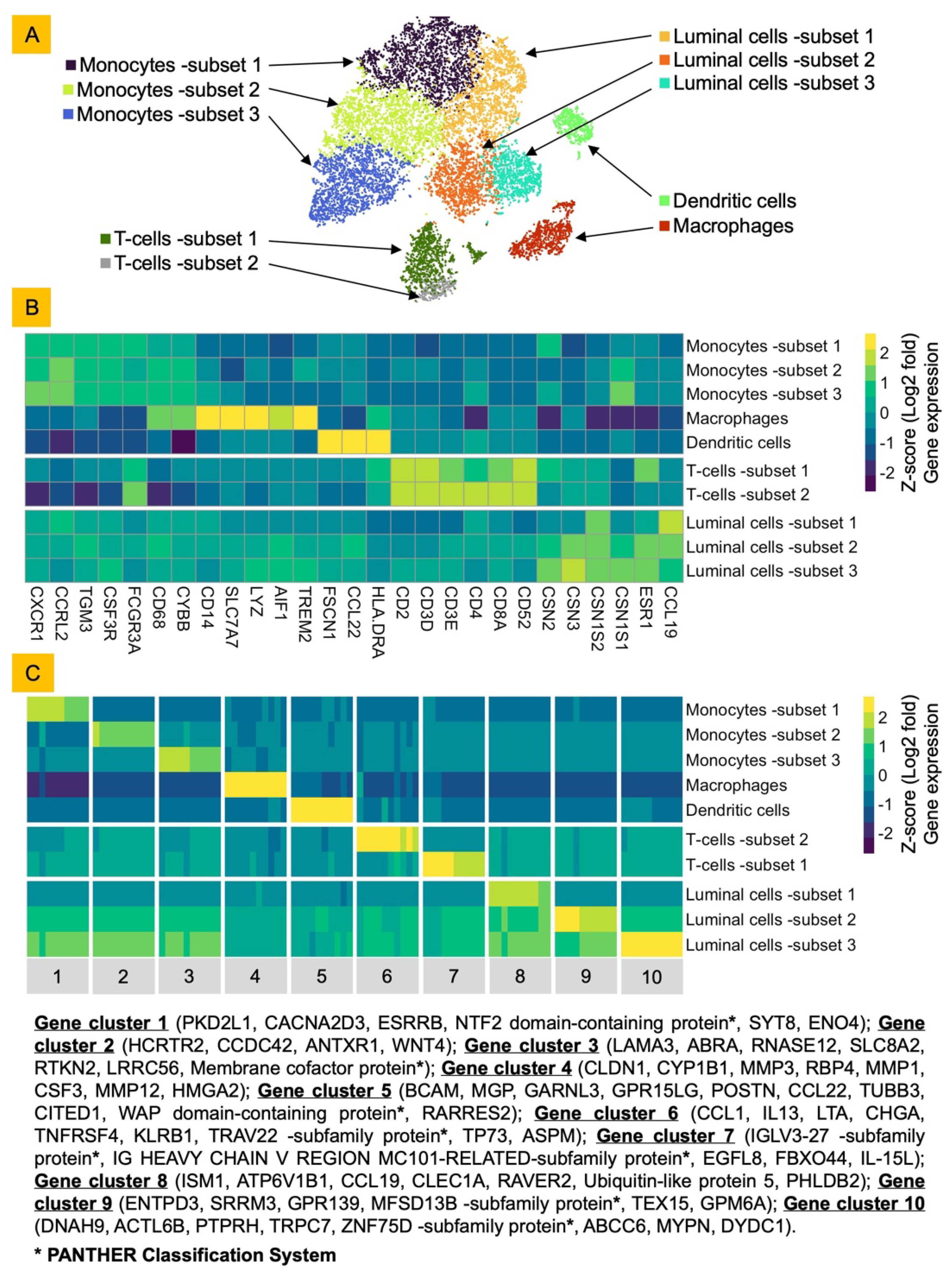
Figure 2.
Comparison of cell populations and gene expression between control and bovine-H5N1 infected bMSCs. (A) t-SNE embeddings of bMSCs either infected bovine-H5N1 or uninfected control cells. Distinct cell populations are represented by different colors, and assigned the labels based on expression of selected gene markers same as Figure 1. (B-D) Bar graphs showing percentage (x-axis) of innate immune (B), T-cells (C), and epithelial (D) cells in total cells, corresponding to different cell types (y-axis) in control (blue) and bovive-H5N1 infected (brown) bMSCs. (E) Heatmap displaying z-scored average expression of top 50 upregulated genes after bovine-H5N1 infection in each cluster that had a fold change >1 expressed and p < 0.05 for at least one cluster. Genes are organized by cluster in which they have highest expression and analyzed for Gene Ontology (GO) enriched processes.
Figure 2.
Comparison of cell populations and gene expression between control and bovine-H5N1 infected bMSCs. (A) t-SNE embeddings of bMSCs either infected bovine-H5N1 or uninfected control cells. Distinct cell populations are represented by different colors, and assigned the labels based on expression of selected gene markers same as Figure 1. (B-D) Bar graphs showing percentage (x-axis) of innate immune (B), T-cells (C), and epithelial (D) cells in total cells, corresponding to different cell types (y-axis) in control (blue) and bovive-H5N1 infected (brown) bMSCs. (E) Heatmap displaying z-scored average expression of top 50 upregulated genes after bovine-H5N1 infection in each cluster that had a fold change >1 expressed and p < 0.05 for at least one cluster. Genes are organized by cluster in which they have highest expression and analyzed for Gene Ontology (GO) enriched processes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



