
Submitted:
04 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
The endocannabinoid system (ECS) regulates and controls many of our most critical bodily functions including healthy muscles however, its potential role(s) in physiological skeletal muscle function and its disorders are unclear. Earlier using the Cre/LoxP system we generated and characterized a tamoxifen-inducible skel-etal muscle-specific CB1R knockdown (skmCB1-KD) mouse model. Here, we sought to elucidate the mechanisms underlying reduced muscle force generation. To address this, we measured calcium movements following electrical stimula-tion-induced muscle fatigue, evaluated store-operated calcium entry (SOCE), and conducted functional analysis of respiring mitochondria. Based on our measure-ments, it is likely that the reduced muscle performance recorded in vivo may oc-cur due to complementary changes in ATP production by mitochondria. Further-more, in skmCB1-KD mice, we observed a significant decrease in one component of the respiratory chain and a delayed mitochondrial membrane potential dissipa-tion upon the application of an un-coupler.

Keywords:
skeletal endocannabinoid system
; murine skeletal muscle
; in vivo muscle force
; mitochondrial respiration
; calcium homeostasis
; store-operated calcium entry
1. Introduction
Skeletal muscle, the largest organ in our body, comprises about 40 to 50% of total body mass and serves crucial roles for overall health, not only for maintaining posture and enabling mobility but also for regulating whole-body homeostasis and metabolism [1,2].
Muscle contraction underlying force generation is a result of a strict sequence of processes called excitation-contraction coupling (ECC) [3,4]. Any alteration of this well-orchestrated sequence of events could lead to altered force generation. During muscle contraction and force generation, chemical energy (ATP) is continuously synthesized and broken down into ADP and AMP. ATP, which acts as fuel for the cell through oxidative phosphorylation (OXPHOS) is produced by the mitochondria, a key organelle that occupies about 10–15% of the muscle fibre volume [5]. Mitochondria, these highly dynamic organelles that constantly undergo fission and fusion, are essential in decoding intracellular calcium (Ca2+) signals, thus contributing to the spatio–temporal distribution of intracellular Ca2+ concentrations [6,7,8,9]. Mitochondrial morphology is essential for ATP synthesis and this dynamism allows for the maintenance of the organelle membrane potential (ψm), which serves as the driving force for ATP synthesis within the electron transport chain (ETC).
Store-operated calcium entry (SOCE) is a Ca2+ influx pathway activated upon sarcoplasmic reticulum (SR) Ca2+ store depletion [10]. This process is based on the assembly and interaction of two proteins: stromal interaction molecule 1 (STIM1), a calcium sensor located in the SR membrane, and Orai1, a calcium channel located in the plasmalemma [11,12,13]. There are two distinct appearances of SOCE. The phasic (p)SOCE reflects the transient nature of the Ca2+ flux and is activated with every action potential when small Ca2+ depletions occur [14]. On the other hand, chronic (c)SOCE is slower and longer-lasting and is activated following a partial (or virtually almost complete) SR depletion. This is either induced ex vivo through pharmacological activation of RyR1 or SERCA, or it occurs in vivo due to leaky RyR1s in pathological states like myopathies [15,16].
The endocannabinoid system (ECS) is a broadly distributed signaling network that is involved in a wide array of physiological processes such as mood and emotion, pain modulation, immune response, appetite, and metabolism [17,18,19,20].
The components of the system are the cannabinoid receptors (CB1R and CB2R), endocannabinoids (the two main ones are anandamide and 2-arachidonoyl-glycerol), and various enzymes and transport systems involved in their synthesis and degradation [21,22,23]. CB1Rs (encoded by the Cnr1 gene) are the most abundant in muscle being involved in cellular functions like autophagy [24], energy metabolism [25,26], and exerting several biochemical effects, including ATP production and modulation of reactive oxygen species (ROS) [27]. In addition to CB1Rs located on the cell membrane, referred to as peripheral CB1 (pCB1R) [28,29], a population is present in muscle mitochondria (mtCB1R), which regulates mitochondrial oxidative activity [30,31]. A recent study showed that the global deletion of CB1Rs induced a fast- to slow-twitch fibre type conversion in mouse m. gastrocnemius, which increased its oxidative capacity and affected its antioxidant defense systems [32].
Using a combination of genetic and pharmacological approaches, we aimed to unravel the intricate interplay between ECS signaling and skeletal muscle physiology. Our work sheds light on the role of ECS, specifically CB1Rs in skeletal muscle, providing new insight into the molecular mechanisms of ECS-mediated regulation of skeletal muscle function, force generation, mitochondria energetics, and overall muscle contraction. This creates new opportunities for therapeutic approaches focused on enhancing muscle health and function, potentially reducing the risk of muscle toxicity caused by various drugs.
2. Results
2.1. In Vivo Muscle Force and Motor Coordination are Depressed in skmCB1-KD Mice
Our workgroup previously generated and characterized a tamoxifen-inducible skeletal muscle-specific CB1R knockdown (KD) mouse model (skmCB1-KD; [33]). However, a key question was left unaddressed: what was the mechanism behind the altered force production observed in these animals? Here, we decided to further investigate these aspects and tried to elucidate the causes and reveal the molecular mechanisms underlying altered muscle performance in skmCB1-KD mice.
To induce skmCB1R-KD, we treated mice for 2 months with tamoxifen starting at one month of animal age (Figure 1A). The chosen tamoxifen treatment substantially reduces CB1R at mRNA and protein levels, as demonstrated in our earlier work [33] . The CB1 downregulation (on mRNA level) was verified here in m. tibialis anterior samples (inset Figure 1A). Genotyping beforehand at 3 to 4 weeks of age allowed to assign animals into a Cre-recombinase negative (Cre-/-) and positive (Cre+/-) group, which served as control or allowed for skeletal muscle-specific tamoxifen-induced KD of CB1, respectively. Regular measurements of body weight and assessment of in vivo force using Rota-Rod (Figure 1B) were performed immediately before, 1, and 2 months after the initiation of tamoxifen treatment. The average weight of the mice was comparable across all tested time points before and after tamoxifen administration and did not differ between control (Cre-/-) and CB1R-KD (Cre+/-) mice (Figure 1C). To assess in vivo force production, we measured the latency to fall for each mouse while the accelerating speed of the Rota-Rod was increased gradually from 4 to 300 rpm in 5 min (see the protocol in Figure 1B). The averaged data of these experiments clearly showed that already 1 month into the tamoxifen treatment the CB1R-KD (Cre+/-) mice performed worse, and their motor coordination was hindered (Figure 1D). While the latency to fall is a widely used parameter to evaluate motor performance, it is also influenced by motor coordination and learning. As a second assessment of in vivo force, we therefore also performed measurements of the grip strength. When normalized to body weight the in vivo grip force was markedly declined at the end of the tamoxifen diet period in CB1R-KD mice when compared to the control (Figure 1E, 4.65±0.23 mN/g for Cre-/- vs 3.77±0.37* mN/g for Cre+/-, *p<0.05). This finding is in agreement with our previous findings (Figure 1H, [33]).
2.2. SOCE Activity is Preserved in skmCB1-KD Mice
We and others have shown earlier that store-operated calcium entry (SOCE) plays an important role in regulating fibre calcium content [14,34,35] and may thus contribute to the reduction in muscle function.
Here, we sought to investigate SOCE using the skinned fibre technique [36], which allows high-resolution measurement of t-system membrane Ca2+-fluxes including SOCE [36,37]. Briefly, the t-tubular system of m. extensor digitorum longus (EDL) fibres from Cre-/- and Cre+/- mice was loaded with the low-affinity Ca2+ sensitive dye Rhod-5N before fibre skinning (Figure 2A). Calibrated measurement of Rhod-5N fluorescence from within the t-tubular system revealed a slight but significant difference in steady-state t-system Ca2+ ([Ca2+]tsys) under resting conditions when comparing Cre-/- to Cre+/- (compare columns 1 and 3 in Figure 2A). We then induced SOCE by activating the RyR1s via exposure of the fibre to a solution containing nominally 0 mM free Mg2+ and Ca2+ , and 30 mM caffeine. Afterwards, we assessed the reuptake of Ca2+ into the t-system after re-exposing the fibre to a physiological free [Ca2+]cyto of 67 nM. Steady-state Ca2+ levels after re-uptake were smaller compared to respective baseline values for both groups, however, the measured values turned out to be significantly smaller for Cre+/- specimens (Figure 2B, columns 2 and 4). The pharmacological activation of SOCE and concomitant depletion of the t-system is fully reversible such that the t-system reloads with Ca2+ upon restoration of physiological [Ca2+]cyto and [Mg2+]cyto values [38]. Such sequence of depletion and reuptake was repeated at different [Ca2+]cyto concentrations of 28, 67, 200 and 1342 nM for Cre-/- and Cre+/- fibres; original representative traces can be seen in Figure 2C and D, respectively. Evaluating [Ca2+]tsys after re-uptake revealed consistently smaller (albeit non-significant) values in Cre+/- compared to Cre-/- for all Ca2+ concentrations tested (Figure 2E). However, peak t-system uptake flux was not altered under the same conditions (Figure 2F), except for the highest concentration tested. When we evaluated the SOCE flux determined as the peak t-system Ca2+ release flux upon 30 mM caffeine application in zero Ca2+/Mg2+conditions, we noticed a reduction (albeit non-significant) in the absolute values by trend (Figure 2G). The peak t-system fluxes derived in this manner are, however dependent on the driving force between the t-system and the cytosol. Because [Ca2+]t-sys was reduced in Cre+/- fibres (Figure 2E), we therefore plotted respective flux values over the ambient steady state [Ca2+]t-sys values immediately before depletion. A linear fit to these data is proportional to the t-system Ca2+ permeability and as such a relative measure of SOCE. Here, the linear relationship follows from the Goldman-Hodgkin-Katz current equation under the assumption of a sufficiently negative resting membrane potential close to -90 mV and a much larger Ca2+ concentration in the t-system compared to the cytosol [10] (methods), two conditions that are readily met in the chosen preparation. When fitting respective data, both Cre-/- and Cre+/- data sets showed a good linear correlation but, were not different from each other (F-test; p=0.77).
In agreement with SOCE being unaltered in CB1R-KD fibres, the protein expression of Orai1 and STIM1, the two key proteins thought to underlie SOCE in skeletal muscle [11,12,13], were likewise unaltered in Cre+/- specimens when compared to Cre-/- (Supplementary Figure S1A–C).
Collectively, these data suggest that the knockdown of CB1R did not alter SOCE but affected the ability of the fibre's t-system to readily take up Ca2+ after depletion.
2.3. Calcium homeostasis Is Preserved in skmCB1-KD Mice
Investigating a global CB1 knockout (KO) murine mouse model our workgroup proposed earlier that CB1Rs are involved in the regulation of Ca2+ homeostasis of the skeletal muscle via a Gi protein and through a PKA-mediated mechanism [40]. Furthermore, our laboratory demonstrated that an immediate refilling of the SR following muscle activation would also require the proper functioning of SOCE [34].
Thus, to specifically investigate the putative involvement of ECS in these processes within skeletal muscle, we monitored the decay of the Ca2+ release following electrical stimulation in control (Cre-/-) and skmCB1-KD (Cre+/-) fibres. This was done in a series of Ca2+ transients triggered by voltage depolarizations with the pattern illustrated in Figure 3A (top). In Figure 3A the fluorescence transient depicted as F/F0 is shown as a white trace in a single m. flexor digitorum brevis (FDB) fibre isolated from a Cre+/- mouse.
In Figure 3B the SR Ca2+ content decline induced by voltage clamp stimulation was fitted with an exponential function for one representative Cre-/- and one Cre+/- FDB fibre. For the given examples the SR Ca2+ decline at the end of the protocol was nearly identical in the two samples (190.7 µM for Cre-/- vs 158.7 µM for Cre+/-). One must note that the light green trace for Cre+/- remains below the dark green Cre-/- values, possibly suggesting a higher propensity for depletion and thus fatigue in the Cre+/- animals. Overall, this data aligns with the SOCE experiments performed on skinned EDL fibres (Figure 2G). Nevertheless, the calculated [Ca2+]SR was only reduced by trend in the Cre+/- fibres and thus, unaffected by Cnr1 genetic ablation (Figure 3C, 555.2±114.9 µM for Cre-/- vs 464.7±54.1 µM for Cre+/-).
In summary, our data suggests moderate fatigability, compromised t-system Ca2+ handling, and negligible changes in SOCE activity in skmCB1-KD Cre+/- muscles. Our findings are in line with unaltered Orai1 flux values (K1 values, Figure 3D), as calculated with our previously introduced model [34] and similar Orai1 protein expression levels in Cre-/- and Cre+/- fibres (Supplementary Figure S1A,B).
The process of Ca2+ clearance from the cytosol in skeletal muscles is majorly accomplished by the SERCA pumps, which mediate the uptake of Ca2+ into the SR stores. In this work, the calculated SERCA pump activity was found to be preserved (Figure 3E) following CB1 downregulation (4.26±0.32 mMs-1 for Cre-/- vs 3.99±0.29 mMs-1 for Cre+/-).
Interestingly, when we measured the resting intracellular calcium concentration ([Ca2+]i) in FDB fibres loaded with the ratiometric dye Fura-2 AM we found a small but significant reduction in Cre+/- fibres (Figure 3F, 47.1±0.6 nM for Cre-/- vs 44.9±0.8* nM for Cre+/-, with *p<0.03).
Taken together, our data suggest minor alterations of Ca2+ homeostasis in CB1R-KD fibres However, we believe these alterations might not be the (sole) reason behind the smaller in vivo grip force observed in this study here, and the decreased in vitro tetanic force demonstrated in our earlier work [33].
2.4. Mitochondrial Function Undergoes Modest Alterations in skmCB1-KD Mice
Mitochondria play a key role in maintaining skeletal muscle function, regulating oxidative stress, calcium homeostasis, and cellular respiration. Earlier we proposed the role of CB1Rs and thus cannabinoid signalling in general, in maintaining proper muscle function and mitochondrial morphology [33]. Here, we aimed to investigate whether the previously observed altered mitochondrial morphology following skeletal CB1R downregulation is also accompanied by changes in mitochondrial function. To test our hypothesis, we used high-resolution respirometry and two different protocols (short and long) to analyze potential changes in the mitochondrial respiration of m. gastrocnemius fibres isolated from the hind limbs of Cre-/- and Cre+/- mice.
First, we applied the short respiration protocol (Figure 4A). This allowed us to determine OXPHOS capacity and leak respiration before and after oligomycin (Omy) application, which inhibits ATP synthesis resulting in a resting or un-phosphorylated state (Figure 4C). Furthermore, routine or baseline respiration, residual oxygen consumption (ROX), and the respiratory control ratio (RCR) were also measured (Figure 4E). We did not observe any differences in these parameters upon CB1R knockdown. An exception was the increase of ROX (4.32±0.54 pmol/s mL for Cre-/- vs 7.07±0.92* pmol/s mL for Cre+/-, *p<0.05), which is obtained after inhibition of the electron transport chain pathway. Various cellular enzymes that consume O2 and promote autoxidation reactions give rise to ROX, including peroxidase and oxidase activities that partially contribute to reactive oxygen species (ROS) production. The oxidative stress, thus ROS state at rest was verified with the non-ratiometric fluorescent dye CellRox Green but we did not find significant differences between the two specimens (Supplementary Figure 2).
Baseline and ROX were determined using the longer respiration protocol as well, along with OXPHOS linked to complex I and II, and complex IV activity (Figure 4B). Among these functional values, only complex IV showed a change, specifically a significant decrease in Cre+/- (Figure 4F, 169.7±11.3 pmol/s mL vs 126.8±8.8* pmol/s mL, *p<0.01). The respiratory control ratio (RCR) was unchanged (Figure 4D).
2.5. Mitochondrial Dynamics Marker Protein Levels are Preserved in skmCB1-KD Mice
Since our previous [33] and current findings pointed towards morphological and functional mitochondrial alterations in skeletal muscles upon CB1R downregulation we next decided to examine the expression level of proteins involved in the maintenance of mitochondrial calcium homeostasis and dynamics. Mitochondrial Ca2+ uptake is driven by the calcium-sensitive regulator MICU1, which modulates mitochondrial function by enhancing ATP production through the calcium-dependent activation of specific metabolic enzymes. Here, using molecular biology tools, we reconfirmed what we described in our prior work [33]: MICU1 was significantly downregulated on protein level in Cre+/- specimens. In the present work, we checked for the mitochondrial content of the voltage-dependent anion channel (VDAC) which was found unaffected by CB1 knockdown (Supplementary Figure S3A,C). These findings were paired with no detectable fibre type switch (at least on mRNA level) (Supplementary Figure S4). Since no changes were seen on mRNA level, this aspect was not investigated further.
We also examined mitochondrial fusion-related proteins: Mfn2 and Opa1 and found no changes, albeit a decline by trend in the latter could be identified (Supplementary Figure S5A,C,D). Drp1, a protein involved in fission processes and known to affect mitochondrial degradation, respiration and ATP synthesis, showed similarly a decline by trend following CB1 knockdown (Supplementary Figure S5A,B), but the change did not reach statistical significance.
2.6. FCCP-Dependent Dissipation of Mitochondrial Membrane Potential is Delayed in skmCB1-KD Mice
The mitochondrial membrane potential (ΔΨm) generated by the proton pumps of complexes I, III and IV is an essential component in the process of energy storage during oxidative phosphorylation, and it provides the driving force for ATP synthesis. Together with the proton gradient ΔΨm, it forms the transmembrane potential of hydrogen ions, which is harnessed to make ATP. Since our respirometry data shed light on a decline of the Complex IV activity in Cre+/- mice (Figure 4D), we were interested in exploring this aspect further.
To this end, we loaded single isolated FDB fibres from both strains with 20 nM TMRE - a dye that stains exclusively live mitochondria in the cell. While continuously scanning the fibre of interest with a confocal microscope, we followed the process of ΔΨm dissipation upon the application of 1 µM FCCP, a well-known un-coupler of the electron transport chain (Figure 5A for Cre-/- and Figure 5C for Cre+/-). Exponential fitting for individual cells (similar to those presented in panels B and D) was used to assess the decay time (τ) for n=14 Cre-/- and n=15 Cre+/- cells obtained from 4-4 animals, respectively. The obtained τ values were averaged and plotted for both groups (Figure 5E). Based on the individual exponential fits the average decay times were: τCre-/- = 125.1±17.9 sec and τCre+/-= 211.3±32.1* sec, with *p<0.02; Figure 5F), meaning that the normalized TMRE fluorescence dissipated slower in the Cre+/- fibres. To exclude possible artifacts due to dye bleaching or leakage we repeated the experiments in the absence of FCCP in 5 control cells from a wild-type C57Bl6 mouse, but essentially no signal loss was detected (Figure 5E, cyan symbols).
3. Discussion
In this study, we characterized the in vivo muscle grip force generation and locomotor coordination in skmCB1-KD mice (referred to as Cre+/- throughout the manuscript) compared to littermate control Cre-/- animals. Furthermore, we analyzed various aspects of cellular calcium homeostasis and mitochondrial respiration.
When compared to control Cre-/- mice, we found that Cre+/- mice had reduced in vivo grip strength force and altered motor coordination (Figure 1). Gonzalez-Mariscal et al. ([41]) proposed that muscle-specific genetic ablation of CB1R influences muscle metabolism by decreasing fat accumulation, promoting oxidative phosphorylation, increasing muscle mass, and inducing a shift in muscle composition characterized by an enrichment of oxidative fibres. Ultimately, these changes were deemed to contribute to enhanced physical performance and an overall improvement in whole-body metabolism in CB1R knockout (KO) mice. Our current results about altered grip force development (including our earlier results on declined in vitro force production as described in Singlár et al. 2022 [33]) may seem somewhat contradictory to these findings. The main difference between the two studies might result from: a) the extent of CB1R knockdown; our inducible CB1R-KD model exhibits ~23% reduction in CB1 on protein level. On the other hand, the study by González-Mariscal et al. did not directly characterize CB1 protein expression, but only the expression of Cre recombinase. Considering it is a constitutive KO model, the degree of KD is presumably greater than 23%; b) the study by González-Mariscal et al. used a constitutive CB1-KO model, where over time compensation mechanisms are likely to occur and influence overall muscle function [41].
CB1 is a G protein-coupled receptor, characterized by its association with heterotrimeric G proteins (Gα, Gβ and Gγ subunits). Upon receptor activation, the Gα subunit and the Gβγ dimeric complex dissociate from the receptor. Subsequently, the Gαi/o and Gαs subfamilies inhibit or activate adenylyl cyclase which prevents (Gαi/o) or stimulates (Gαs) the conversion of adenosine triphosphate to adenosine 3′,5′-cyclic monophosphate (cAMP). Formerly, our workgroup studied a systemic CB1-KO mouse model and found that cannabinoid signaling inhibits sarcoplasmic Ca2+ release and regulates excitation-contraction coupling in mammalian skeletal muscles via Gi-PKA-mediated mechanism [40]. On the other hand, a few years ago, Iannotti et al. demonstrated that in C2C12 myoblasts the effects of 2-AG or synthetic CB1 agonists were unaffected by pertussis toxin, indicating that CB1 coupling in these cells does not involve Gi proteins [42]. Instead, CB1 stimulation induced the hydrolysis of PIP2 (Phosphatidylinositol 4,5-bisphosphate), which relies on Gq-PLC-IP3 axis activation, thus intracellular calcium concentration elevation. Keeping in mind that CB1 mRNA and protein expression profile during murine C2C12 myotube formation is highly time-sensitive, one possible explanation to the above-described discrepancy could be the experimental model studied: Oláh et al. evaluated terminally differentiated C2C12 myotubes and adult FDB fibres from CB1-KO mice, whereas Iannotti et al. examined differentiating C2C12 myoblasts.
Ultimately, we propose drawing an analogy with the CB1-KO model characterized by Oláh et al. [40]: in our skmCB1-KD mouse model, signaling mediated by the Gi/o protein through the inhibition of adenylyl cyclase-cAMP-PKA axis may serve as a significant downstream effect of CB1 downregulation potentially accounting for the altered aspects of calcium homeostasis presented here (Figure 2 and Figure 3). However, we must acknowledge that we did not conduct experiments in this study to confirm this hypothesis, and further investigations are needed to test this.
Most of the muscles present a heterogeneous composition of slow- and fast-type fibres [43]. The spectrum of MyHC isoforms is not fixed and can shift according to the following scheme: I↔IIa↔IIx↔IIb. Iannotti et al. reported that global CB1-KO mice have larger and more numerous muscle fibres compared to control animals [42]. Likewise, Senese et al. [32] working on the same systemic CB1-KO model described elevated oxidative stress and a marked enrichment of type I slow oxidative fibres upon systemic lack of CB1 in gastrocnemius muscles. In our skmCB1-KD mice however, we did not detect signs of fibre type switch when examining MHC isoforms on mRNA level in TA muscles (a mixed composition muscle, Supplementary Figure S4).
It is now well established that long-lasting stimulation of muscle fibres may result in the increase of cytoplasmic Ca2+ concentration accompanied by the reduction of SR Ca2+ release, SR store content, and enhanced ROS production. The metabolic changes in the working muscle (e.g. reduced ATP, increased ADP) may lead to changes in cellular Ca2+ homeostasis that contribute to physiological fatigue, as reduced Ca2+ release may eventually cause reduced force output. Alterations in [Ca2+] across organelles have been related to a variety of pathologies, where the persistent intracellular Ca2+-overload impairs ATP production, promotes excessive ROS production, and provokes the apoptotic cascade and mitochondrial damage. Interestingly, in our skmCB1-KD mice, we found significantly declined [Ca2+]cyto values (Figure 3F) which would normally directly influence [Ca2+]mito uptake as a compensatory mechanism to normalize the [Ca2+]cyto [44]. We did not measure mitochondrial calcium here, however, in our earlier work we did not identify signs of mitochondrial calcium overload (see Figure 6 from Singlár et al. [33] ). In this study, changes in oxidative stress were observed through CellRox Green staining in FDB fibres from both specimens but there was no evidence of excessive ROS production upon skmCB1 downregulation (Supplementary Figure S2).
Significantly lower levels of resting intracellular calcium were registered upon Cnr1 genetic ablation in FDB fibres (Figure 3B). This may serve as an explanation for the lower calculated [Ca2+]SR levels which, however, did not result in enhanced SOCE activity as inferred from measurements in intact FDB (Figure 3D) and skinned EDL fibres (Figure 2). One feasible explanation for the minimal changes in SOCE could be the modest efficiency of CB1 knockdown induced by 2-month-long tamoxifen diet. In the present work we assessed the CB1 downregulation in TA muscles on mRNA level (Figure 1A, inset). In our earlier work [33] we have quantified in slow (SOL), fast (EDL) and mixed muscles (TA) the level of CB1 deficiency and it was quite dramatic on mRNA level (66.44%, 81.34% and 66.04% for EDL, SOL and TA, respectively but more modest on protein level (~23% in TA%). It remains elusive however, if an extended tamoxifen diet period would be more suitable to achieve a larger CB1 protein knockdown. During prolonged muscle stimulation (voltage or pharmacologically induced) it is speculated that pSOCE, in cooperation with the t-system Ca2+- uptake proteins Na+/Ca2+ exchanger (NCX) and/or plasma membrane Ca2+ ATPase (PMCA) - equilibrates Ca2+ gradients across the t-system membrane in a bidirectional manner. These events lead to an influx of Ca2+ if [Ca2+]cyto transients are lower and an efflux of Ca2+ if [Ca2+]cyto transients are higher than “normal”. Moreover, [Ca2+]cyto sets SR load and, according to Koenig et al. [45], also affects pSOCE. Here, we have analyzed the protein levels of the PMCA and no alterations were detected when comparing the TA muscle samples from Cre+/- and Cre-/- specimens (Supplementary Figure S1D,E).
The decline in SR Ca2+ concentration due to voltage-induced electrical stimulation in FDB fibres was moderate in the skmCB1-KD Cre+/- muscles (Figure 3B). This could indicate a slight impairment in SOCE dynamics within the Cre+/- muscle fibres, potentially leading to a more significant depletion of Ca2+ from the SR (for details see Eq. A10 from Sztretye et al. (2017) [34]). It was established that phasic SOCE depends on [Ca2+]i, thus the intracellular calcium levels albeit significantly decreased in Cre+/- (the change is physiologically irrelevant) are only mildly reflected in the pSOCE measurements. Notably, the reduction in resting Ca2+ observed upon Cnr1 genetic depletion was modest which may translate to insignificant changes in free Ca2+ concentration of the SR.
In a working muscle mitochondria will release ROS which in turn could affect SOCE. Nevertheless, SOCE is completely controlled by the SR near membrane Ca2+ content. The uptake of Ca2+ by the mitochondria is controlled by the [Ca2+]cyto, which is set by the Ca2+ permeability of the t-system (a balance of SOCE and PMCA activity [35,46]). On the other hand, more and more studies elaborate on the role of regulatory proteins that contribute to the connection between the sarco(endo)plasmic reticulum (SR) Ca2+ store and mitochondria contact sites referred to as mitochondria-associated membranes (MAMs). In mammalian skeletal muscle fibres, most mitochondria are in contact with the SR, providing a direct link between Ca2+ cycling and energy demand [47]. It is now generally accepted that mitochondria accumulate high [Ca2+] following physiological stimulation when [Ca2+]cyto rises from 0.1 μM to about 2–3 μM. The SR holds ~90 % of the fibre Ca2+ content at steady state. The mitochondria hold < 5 % of the fibre Ca2+ content in healthy muscle [35] and until recently it was not considered a significant candidate to sequester compelling amounts of Ca2+ during SR Ca2+ release. Nevertheless, the proximity between the SR and the mitochondria ensures the formation of high [Ca2+] microdomains, allowing mitochondria to rapidly take up Ca2+. For Ca2+ to enter the mitochondrial matrix, it requires the crossing of the outer and inner mitochondrial membranes (OMM and IMM). The initial passage across the permeable OMM occurs through the voltage-dependent anion channel (VDAC), which is highly expressed in OMM. Therefore, the VDAC expression levels constrain the mitochondrial Ca2+ influx, overexpression of VDAC increases the mitochondrial Ca2+ uptake, whereas silencing VDAC reduces it [48]. Here, we have analyzed by Western Blot the levels of VDAC which showed an almost 2-fold increase in Cre+/- muscles (Supplementary Figure S4A,C). This increase however did not prove to be significant.
Furthermore, the accumulation of Ca2+ in the mitochondrial matrix occurs down to its electrochemical gradient via the mitochondrial calcium uniporter (MCU). MCU uses the electrochemical potential (ΔΨm ~ -180 mV) established across the IMM to drive the Ca2+ entry into the matrix [49]. MICU1 protein as part of the MCU holocomplex acts as a gatekeeper, keeping the channel closed under resting conditions and establishing a Ca²⁺ threshold for mitochondrial uptake [50]. In a very recent study by Hasan et al. (2024) MICU1/2 loss increased mitochondrial calcium influx and sensitized mitochondria to calcium overload injury [51]. In our skmCB1-KD muscles MICU1 levels were significantly depressed upon Cnr1 genetic manipulation (Supplementary Figure S4B,D).
Treatment with protonophores like FCCP, disrupts the ΔΨm, entirely abolishing the mitochondrial Ca2+ uptake process and depleting buffered mitochondrial Ca2+ [48,52]. In our experiments with FCCP we observed significantly slower dissipation of ΔΨm upon CB1 ablation in Cre+/- muscles (Figure 5E,F). We propose that this phenomenon, along with decreased Complex IV activity (Figure 4D) may contribute to a diminished ATP turnover and, consequently a lower rate of energy production. This may explain the reduced in vivo grip force generation (as well as the reduced in vitro force findings presented by Singlár et al. 2022 [33]) and potentially lead to increased fatigability in Cre+/- specimens.
The mitochondrial energy production is carried by the five complexes inserted in the IMM. In summary, pyruvate, a by-product of glucose metabolism, enters the mitochondria through VDAC and is enzymatically converted into acetyl-CoA within the matrix. Further, the acetyl-CoA enters the Krebs cycle, where it undergoes oxidation, resulting in the production of NADH and FADH2 [53]. NADH is subsequently oxidized by complex I (NADH dehydrogenase), which transfers electrons to ubiquinone, reducing it to ubiquinol. Complex II (succinate dehydrogenase) is the only membrane-bound enzyme in the Krebs cycle, catalyzing the conversion of succinate to fumarate [54]. Complex III (ubiquinol-cytochrome C oxidoreductase) catalyzes the transport of H+ from the matrix to IS, coupled with the movement of electrons from ubiquinol to cytochrome C. After receiving the electrons, cytochrome C transports them to complex IV (cytochrome C oxidase). Lastly, complex IV then transfers the electrons to molecular oxygen, forming water. This process also drives the translocation of two protons (H⁺) from the matrix to the IS for each water molecule produced [55]. In our mitochondrial respiration experiments performed on freshly isolated m. gastrocnemius fibers we found a marked decline in Complex IV activity (Figure 4D) which could be associated with changes in organelle energy status, thus ATP provision. In the current study, an examination of TA muscle fibers (Supplementary Figure S4), which have a mixed fibre composition similar to the gastrocnemius muscle used for the respirometry experiments, did not reveal any evidence of a fibre type shift towards an oxidative phenotype (at least at the mRNA level). Nevertheless, we cannot fully exclude this possibility.
CB1Rs have been proposed to regulate nitric oxide (NO) production through nitric oxide synthase (nNOS and eNOS), thus their role has been associated vascular health, cannabinoids-mediated neuroprotection, and mitochondrial biogenesis in adipose tissue and placenta [56,57]. Considering that NO is a direct inhibitor of complex IV, one cannot exclude the possibility that the regulation of NOS/NO production might be involved in the impaired complex IV activity seen in CB1R-KD muscles. This hypothesis however, was not verified in the present study.
Mitochondrial dynamics is a delicate balance between division (fission) and fusion processes that maintain the shape and structure of mitochondria. For healthy mitochondrial function fission and fusion events must be equally balanced. The proteins regulating fission are Drp1 and Fis1, whereas fusion is controlled by Mfn1, Mfn2, and Opa1. Mfn2 is located at both ER and mitochondrial membranes where it has been reported to strengthen ER–mitochondria contacts and facilitate mitochondrial Ca2+ uptake [58,59]. Recently, it was proposed that Mnf2 acts as a negative regulator of ER–mitochondrial tethers, which, by reducing the number of contacts, avoids toxic Ca2+ accumulation [60]. Earlier, we described significantly altered mitochondrial morphology upon muscle-specific CB1 downregulation [33]. In the present study, the mitochondrial dynamics-related proteins examined (Mfn2, Opa1, Drp1) displayed no changes in Cre+/- mice (Supplementary Figure S5).
Figure 6 summarizes our findings with emphasis on the putative role of the skeletal endocannabinoid system in proper muscle force generation, cellular calcium homeostasis, and mitochondria function. Based on our past [33] and current data on the skmCB1-KD mice we propose, that CB1Rs are involved in physiological muscle force generation and maintenance of adequate cellular and mitochondrial function. Adjusting the cannabinoid signaling in muscle may be a valuable approach in certain diseases (e.g. myopathies) and conditions related to reduced force generation, and drug-induced muscle toxicity and wasting cases.
4. Materials and Methods
4.1. Animal Care
Animal experiments adhered to the guidelines of the European Community (86/609/EEC) and were designed to minimize animal suffering and distress. The experimental protocol was approved by the Institutional Animal Care Committee of the University of Debrecen (3-1/2019/DEMAB). The mice were housed in plastic cages with mesh covers and fed ad libitum with pelleted mouse chow and water. Room illumination was an automated cycle of 12 h light and 12 h dark, and the room temperature was maintained within the range 22–25°C.
4.2. Generation of the Muscle-Specific CB1 Knockdown Mouse Strain
Tamoxifen-inducible muscle-specific CB1 knockdown mice were obtained as described in Singlár et al., (2022). Mixed-gender, tamoxifen-inducible muscle-specific CB1 knockdown mice (hereinafter referred to as Cre+/-) and littermates controls (Cre-/-) were used here.
4.3. Tamoxifen Diet
To induce CB1 ablation, tamoxifen diet (per os) was started immediately after the separation of the Cre+/- and Cre-/- pups from the mother at the age of 4 weeks. Littermates were fed for 2 months without interruption (while monitoring normal body weight gain and performing grip strength and Rota-Rod tests see Figure 1A) and then were used for the subsequent experiments. The tamoxifen-supplemented chow (Envigo, TD 130857) contained 500 mg tamoxifen/kg, providing 80 mg tamoxifen/kg body weight per day assuming 20-25 g body weight and 3-4 g daily food intake.
4.4. Molecular Biology
4.4.1. Genotyping
Genotyping was performed at 3 to 4 weeks of age and the procedure helped to identify the animals and allowed their classification into experimental groups.
Total genomic DNA was isolated from finger biopsies and screened for the presence of the HSA-Cre recombinase cassette by PCR (Biometra Advanced Twin 48 G, 230 V, Analytik Jena GmbH, Jena, Germany) as described in detail earlier in Singlár et al. (2022) [33].
4.4.2. Western Blot Analysis
M. tibialis anterior (TA) skeletal muscle tissues were homogenized in lysis buffer (20 mM Tris–HCl, 5 mM EGTA, Protease Inhibitor Cocktail (Sigma, Saint Louis, USA) with an HT Mini homogenizer (OPS Diagnostics USA). Six-fold concentrated electrophoresis sample buffer (20 mM Tris–HCl, pH 7.4, 0.01% bromophenol blue dissolved in 10% SDS, 100 mM β-mercaptoethanol) was added to total lysates to adjust equal protein concentration of samples and then the mixture was boiled for 5 min at 90°C. 20 μg of total protein was loaded to each lane and separated on a 10% SDS–polyacrylamide gel. Proteins were transferred to nitrocellulose membranes, blocked with 5% non-fat milk dissolved in phosphate saline buffer (PBS), and then membranes were incubated with the appropriate primary antibodies overnight at 4°C. The primary antibodies used for specific labeling are summarized in Table 3. After washing for 30 minutes in TBS supplemented with 1% Tween-20 (TBST), membranes were incubated with HRP-conjugated secondary antibodies (Blotting Grade Goat Anti-Rabbit IgG (H+L) (Human IgG Absorbed) Horseradish Peroxidase Conjugate (cat.no. 170-6515, Bio-Rad, Hercules, CA, USA) and Blotting Grade Affinity Purified Goat Anti-Mouse IgG (H+L) Horseradish Peroxidase Conjugate (cat.no. 170-6516). Membranes were developed and signals were detected using enhanced chemiluminescence (Thermo Fisher Scientific, Waltham, MA, USA). Optical density of signals was measured by ImageJ software (NIH, Bethesda, MD, USA) and results were normalized to the optical density of α-actinin (1:1000, cat.no. sc-166524, Santa Cruz Biotechnology, Dallas, TX, USA) and tubulin (1:4000, cat. no. T5168, Sigma, Saint Louis, USA), respectively (see Table 3).
4.4.3. qPCR
TRI reagent (cat. no. TR118MRC, Cincinnati, OH, USA) was used to isolate total ribonucleic acid (RNA) fractions from homogenized skeletal muscle specimens of m. flexor digitorum brevis (FDB), m. extensor digitorum longus (EDL), m. soleus (SOL), and m. tibialis anterior (TA) from tamoxifen-fed Cre+/− and Cre−/− mice. Before use, the separated RNA samples were kept at -80 °C dissolved in nuclease-free water (NFW). A spectrophotometer set to 260 nm (NanoDrop ND1000; Promega Biosciences, Madison, WI, USA) was used to measure the RNA yield and purity. After treating the separated RNA samples with DNase and RNase inhibitor (Ambion, Austin, TX, USA), a High-Capacity cDNA Reverse Transcription Kit (cat. no. 00735667, Thermo Fisher, Waltham, MA, USA) was used to perform reverse transcription.
500 ng of the extracted total RNAs were reverse transcribed into complementary DNA (cDNA) following the manufacturer's instructions. Random hexamers were used in a 25 µL reaction volume to carry out cDNA synthesis. SYBRGreen mix (cat.no 4367659, Thermo Fisher Scientific) was utilized for quantitative RT-PCR. A Light Cycler 480 Master instrument (Roche, Basel, Switzerland) was used for the amplification (cat. no. for plates, Roche: 04729692001; cat. no. for sealing foils, Roche: 04729757001). The investigated genes are summarized in Table 4; all primers were synthesized by Invitrogen and were as follows: myosin heavy chain 1 (MHCI), myosin heavy chain 2A (MHCIIA), and myosin heavy chain 2X (MHCIIX). 18sRNA housekeeping gene was used as the internal control (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA).
45 cycles of 3 min at 95 °C were performed. The comparative Ct technique was used to determine the relative expression values for each transcript of interest. Each sample was normalized to its own internal control gene expression after being conducted in triplicate. Subsequent analysis was conducted using the mean values that were obtained.
4.5. In Vivo Experiments
4.5.1. Body Weight Measurement
The body weight was measured at the beginning of the tamoxifen diet (at 1-month-old age) and weekly afterward until the end of the 2-month feeding period (3-month-old age) for each individual mouse in both Cre-/- and Cre+/- groups. In Figure 1C we present the average monthly body weight monitoring.
4.5.2. Grip Strength Test
Forepaw force was measured as previously described [33]. Briefly, the animals were gently dragged away from the grip test meter by their tails after they had successfully grabbed its bar for a brief period. A computer linked to the test meter recorded the maximum force before the animal released the bar, digitizing it at a frequency of 2 kHz. Each animal underwent the test ten to fifteen times to get an averaged single data point which was then normalized to body weight. Grip strength was assessed before, during, and after the tamoxifen diet at 1, 2, and 3 months of age, respectively.
4.5.3. Rota-Rod Test
The mice were placed on a rotating cylinder (cat.no YPFB00, Dev Scientific and Engineering, India) and the duration (in seconds) spent on the rotating rod before falling was recorded. The device was set to an accelerating protocol (rpm consistently increased over time – accelerating from 4 to 300 rpm in 300 sec). The mice were acclimated to the Rota-Rod before testing. The acclimatization consisted of three trials with 10-minute intervals between trials at a constant speed (4 rpm) for 1 minute. The performance of the mouse was tested after 30 min of rest and the time the mouse remained on the Rota-Rod was recorded. On the day of the examination, the accelerating protocol was performed three times and the average of the three trials was calculated for each animal. The test was carried out at various time points of the tamoxifen diet: before (1-month-old), during (2-month-old), and after (3-month-old).
4.6. In Vitro Experiments
Mice were anesthetized and sacrificed in compliance with the guidelines of the European Community (86/609/EEC). After CO2 overdose and cervical dislocation, m. flexor digitorum brevis (FDB), m. extensor digitorum longus (EDL), m. tibialis anterior (TA) and m. gastrocnemius muscles from the hind limb were dissected manually under a transmitted light microscope using thin forceps and fine precision surgical scissors.
4.6.1. Isolation of Single FDB Fibres
All calcium measurements were carried out on skeletal muscle fibres from the FDB or EDL muscle of the mouse. Calcium free Ringer’s solution (containing in mM: 136 NaCl, 5 KCl, 1 MgCl2, 10 HEPES; 10 glucose; pH 7.2) was used during the dissection of the muscle. Single muscle fibres from FDB were enzymatically dissociated in minimal essential media containing 0.2% Type I collagenase (Sigma) at 37 °C for 45-50 minutes. To release single fibres, the FDB muscles were mechanically dissociated and then triturated gently in a normal Ringer solution (same as above but supplemented with 2.6 mM CaCl2). The isolated fibres were then placed in culture dishes and stored at 4 °C in the refrigerator until use. Only fibres with clearly visible striations, no swelling or surface membrane damage were selected for subsequent experiments.
4.6.2. Mitochondrial Membrane Potential Measurement, Oxidative Stress Measurement, Confocal Microscopy, and Image Processing
Isolated FDB fibres from 3-month-old Cre-/- and Cre+/- mice were loaded with 20 nM tetramethylrhodamine methyl ester (TMRE) prepared in normal Ringer’s solution at room temperature for 15 min. After 20 min the excess dye was washed. To monitor TMRE fluorescence, a time series of images was acquired using a Zeiss 5Live confocal microscope (Zeiss, Oberkochen, Germany) equipped with a 20x air objective and 543-nm laser. Sequential images were taken at 10-second intervals. 20 seconds after the start of the time series 1 μM FCCP was perfused onto the fibre while the decay of TMRE fluorescence indicating the mitochondrial membrane potential (ψm) dissipation was continually monitored. Using the Zeiss Live analyzer software (Zeiss, Oberkochen, Germany) an arbitrarily chosen region of interest (ROI) was marked on all subsequent images, and the obtained fluorescence values were fitted with an exponential function using Eq.3 to determine the decay time (τ). The averages of the individual decay times were plotted.
In the CellRox Green experiments (cat.no. C10444, Thermo Fisher Scientific, Waltham, MA, USA) FDB fibres were loaded in normal Ringer’s solution with the dye in 1:500 dilution for 30 min at 37°C. Excitation wavelength was at 488 nm and detection at λ>520 nm. An arbitrary ROI (marked with a rectangle) was selected in parallel with the longitudinal axis of the fibre and the fluorescence was plotted. The ROI selection was always done in a way to avoid areas of dye accumulation (e.g. nuclei). Fluorescence was calculated similarly as described previously [61]. In short, at the peaks (I-band fluorescence, representing mitochondria (FI-band)) and at troughs (A-band fluorescence, representing baseline, (FA-band)) fluorescence was measured, then the normalized mitochondrial fluorescence expressed as Fmito was calculated with the equation:
Fmito=(FI-band-FA-band)/FA-band
4.6.3. Assessment of Mitochondrial Oxygen Consumption Using High-Resolution Respirometry
Mitochondrial oxygen consumption (O2 flux) was assessed in m. gastrocnemius (7 mg/chamber) using High-Resolution FluoRespirometry (Oxygraph-2k, Oroboros Instruments, Innsbruck, Austria). The isolated muscles were incubated in BIOPS solution containing saponin to ensure membrane permeabilization. After stabilization of baseline respiration, we used a short and a long respirometric protocol (see below).
In the short respirometric protocol (Figure 4A), the integrity of the outer mitochondrial membrane was tested with exogenous cytochrome c (10 µM) following OXPHOS II stimulation (0.5 µM Rotenone (Rot), 10 mM succinate (S), and 2.5 mM ADP). Respiratory control ratio (RCR), an index of respiration coupled to ADP-ATP conversion was expressed as a ratio of OXPHOS II to the ATP synthase-inhibited (oligomycin, Omy; 2.5 μM) LEAK state. The electron transport system-independent respiration (or residual oxygen consumption; ROX) was determined after complex III inhibition with antimycin A (2.5 μM).
In the long respirometric protocol (Figure 4B), complex I-linked oxidative phosphorylation (OXPHOS I) was measured in the presence of complex I-linked substrates (10 mM glutamate and 2 mM malate) and ADP (2.5 mM). Rotenone (Rot; 0.5 µM) was used to (a) inhibit complex I and (b) assess complex II-linked oxidative phosphorylation (OXPHOS II) in the presence of succinate (S; 10 mM) and adenylate. After inhibition of complex III (antimycin A; 2.5 µM), complex IV respiratory activity was measured with ascorbate (2 mM) and artificial substrate N,N,N',N'-Tetramethyl-p-phenylenediamine dihydrochloride (TMPD; 0.5 mM). Ascorbate was added before TMPD to avoid uncontrollable autoxidation of the electron donor. Sodium azide (NaN3; 100 mM) was finally administered to block complex IV-linked mitochondrial respiration.
Both the short and long protocol measurements were performed in a Mir05 respiration medium under continuous magnetic stirring (750 rpm) at 37 °C. The DatLab software (Oroboros Instruments, Innsbruck, Austria) was used for online display, respirometry data acquisition, and analysis.
4.6.4. T-System Ca2+-Uptake Measurement
Mixed-gender Cre-/- (n=5) and Cre+/- (n=7) mice were investigated and t-system uptake measurements were performed as previously described in Cully et al. (2016) [36]. Briefly, euthanasia was performed via cervical dislocation, followed by rapid dissection of the whole m. extensor digitorum longus (EDL) muscle, which was then placed in a glass Petri dish covered with Sylgard and submerged in paraffin oil. A bundle of fibres was exposed to a standard Ringer solution containing 2.5 mM of the low-affinity Ca2+-sensitive dye Rhod-5N for 10 minutes. Individual fibres were isolated from the Rhod-5N loaded bundle and skinned by mechanically removing the sarcolemma. Skinned fibres were subsequently transferred to a custom-made chamber constructed on top of a 1.5 mm coverslip filled with an intracellular solution containing (in mM) 90 HEPES, 10 EGTA, 9.86 MgO2, 9.7 CaCO3, 8 Na2ATP, 10 NaCP, with pH adjusted to 7.1 with KOH (yielding a free Ca2+ concentration of 67 nM). Adjustment of the total [Ca2+] and [Mg2+] resulted in free [Ca2+] concentrations of 28, 67, 200, and 1342 nM, and free [Mg2+] concentrations of 1 mM, respectively. The chosen calcium concentrations are a consequence of the mixing of different EGTA-buffered solutions (one low, one high in free Ca2+) following Cully et al. 2016 [62]. These solutions are based on earlier work by George Stephenson [63]. Depletion of the t-system was induced in a nominally Ca2+ and Mg2+ free solution with the addition of 30 mM caffeine (cat.no. C0750, Sigma). The chamber was transferred onto the stage of a confocal microscope for continuous Rhod-5N fluorescence recording. Minimal (Fmin) and maximal (Fmax) dye fluorescence were determined in the presence of maximal (5 mM) and nominally Ca2+-free solutions containing 25 µM of ionomycin (cat.no. I9657, Sigma) and A23187 (cat.no. C7522, Sigma). The spatially averaged real-time fluorescent signal (F) was converted to calcium concentration using the formula:
with a KD of 0.872 mM (Cully et al., 2016). Uptake fluxes were calculated as the signal’s first-time derivative.
[Ca2+] = β KD*(F-Fmin)/(Fmax-F)
4.6.5. Resting Myoplasmic [Ca2+]i Measurement
The resting intracellular Ca2+ concentration was measured using Fura-2 AM fluorescent calcium indicator. Briefly, single FDB fibres were mounted on a glass coverslip with 5 µM Fura-2 AM dye for 1 h. Fibres were then washed with fresh normal Tyrode’s solution (137 mM NaCl, 5.4 mM KCl, 0.5 mM MgCl2, 1.8 mM CaCl2, 11.8 mM Hepes-NaOH, and 1 g/L glucose, pH 7.4) and then imaged with the CoolLED pE-340fura set up (CoolLED Ltd. Hampshire, UK). The acquisition rate was set to 10 Hz. Fura-2 ratios from myoplasmic areas of interest were calculated as R = F340/F380 using Zeiss Zen software (Zeiss, Oberkochen, Germany) and then converted to resting free Ca2+ concentrations using an in situ calibration curve for Fura-2 AM according to the method of Grynkiewicz et al. (1985) [64] based on Eq.1 (see above) where β Kd = 0.3.
4.6.6. Whole-Cell Voltage Clamp
The experimental design was as described in our earlier report [34]. Briefly, isolated FDB fibres were voltage-clamped (Axoclamp 2B, Axon Instruments, Union City, CA, USA) and imaged using a confocal microscope (Zeiss 5 Live, Oberkochen, Germany). Fibres were dialyzed with 50 µM Rhod-2-containing internal solution (see below). The experimental temperature was 20–22°C and the holding potential was −80 mV. Pipette resistance varied between 1 and 2 MΩ. The experiments were performed in the presence of 10 mM EGTA, so that the endogenous buffers in the removal process were almost negligible. Correction for linear capacitive currents was performed by analog compensation.
The intracellular calcium ([Ca2+]i) values were determined from the fluorescence (F) and the background fluorescence (F0) in line scan images analyzed by an in-house custom-made program using the following parameters taken from Royer et al. (2015) [65]: Kd Rhod-2 = 1.58 μM and kON = 0.07 μM−1 ms−1 and kOFF = 130 s−1. Ca2+-release flux was derived from cytosolic Ca2+ transients and subjected to a removal-model fit analysis, which calculates release flux as that necessary to account for the evolution of [Ca2+]i(t) in a single-compartment model that includes quantitatively specified processes of removal as originally described by Melzer et al. [66] and further optimized by Royer et al. [65]. In brief, the model considers that [EGTA] is sufficiently high, and thus only four forms of Ca2+ must be considered: free, bound to the monitoring dye, bound to EGTA, and sequestered in the SR. From the release flux (the flux exiting through the release channels), the net flux leaving the SR can be derived by subtraction of the pump-removal flux.
The evolution of SR Ca2+ content during repetitive stimulation in both mouse groups was calculated from the amount of calcium released minus the removal flux, which contains the uptake of Ca2+ into the SR by the SERCA pump.
To estimate the average amount of calcium in the SR from the depolarizing train, single-exponential functions were fitted to the eight points of the series. The exponential decay was assessed by the equation below:
where x is the number of tetanic pulses applied, b is the time constant of SR depletion and a is the remaining SR calcium content.
y=y0+ae−bx
4.6.7. External Bath Solution (in mM):
140 TEA-CH3SO3, 1 CaCl2, 3.5 MgCl2, 10 Hepes, 1 4-AP, 0.5 CdCl2, 0.3 LaCl3, 0.001 TTX (citrate), and 0.05 BTS (N-benzyl-p-toluene sulphonamide; Sigma-Aldrich). pH was adjusted to 7.2 with TEA-OH and osmolality was adjusted to 320 mOsm with TEA methanesulfonate.
4.6.8. Internal (pipette) Solution (in mM):
110 N-methylglucamine, 110 L-glutamic acid, 10 EGTA, 10 Tris, 10 glucose, 5 Na ATP, 5 phosphocreatine Tris, 0.1 Rhod-2, 3.56 CaCl2, and 7.4 mM MgCl2 were added for a nominal 1 mM [Mg2+] and 100 nM [Ca2+]. pH was set to 7.2 with NaOH and osmolality to 320 mOsm with N-methylglucamine.
4.7. Quantification and Statistical Analysis
Pooled data were expressed as mean±standard error of the mean (SEM). The differences between Cre-/- and Cre+/- mice were assessed using one-way analysis of variance (ANOVA) and all pairwise Bonferroni's multiple comparison methods using the statistical program Prism 9 (GraphPad Software, San Diego, CA, USA). Student’s t-test was used to test the significance and a p-value of less than 0.05 was considered statistically significant. G-Power (http://www.gpower.hhu.de) software was used to estimate the number of specimens included in the in vivo experiments (α=0.05; assumed effect size: 1; calculated power: 0.9548089).
5. Conclusions
Understanding the role of the skeletal endocannabinoid system in health (and disease) is of utmost importance to be able to address proper therapies in conditions where a defective ECS activity occurs. Here, we give a detailed functional and molecular characterization of a transgenic mouse model upon skeletal muscle-specific Cnr1 genetic ablation. Our in vivo analysis revealed altered motor coordination and grip strength following CB1R knockdown. On the cellular level, we uncovered alterations in certain aspects of calcium homeostasis and hindered ATP production by the mitochondria. Thus, we propose a putative link between ECS (more precisely CB1R) and physiological muscle force generation along with the role of ECS in maintaining physiological cellular and mitochondrial homeostasis and function.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Document S1. Supplementary Figure S1. Western Blot analysis of SOCE partner proteins and PMCA. Supplementary Figure S2. Oxidative stress measurement with CellRox Green in FDB fibers. Supplementary Figure S3. Examination of mitochondrial proteins VDAC and MICU1 in TA muscles. Supplementary Figure S4. No fiber type change was detected on mRNA level upon CB1R downregulation. Supplementary Figure S5. Mitochondrial dynamics-related proteins show no change in Cre+/- mice.
Author Contributions
M.S., A.K.P., X.K., F.A.I., and L.C. designed the experiments. Research on genotyping: body weight, grip force, and Rota-Rod was performed by Z.S. and N.G. Molecular biology experiments were performed by N.G. and Z.S. Functional studies, and research on calcium imaging were performed by Z.S., N.G., M.S., P.S., E.L. Z.M.K., L.J. M.S.: P.S., Z.S., A.K.P., and E.L. analyzed the results. M.S. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Hungarian National Research, Development and Innovation Office: NKFIH-FK-142481 (M.S.), and NKFIH 2019-2.1.11-TÉT-2019-00102 (M.S.). Project no. TKP2020-NKA-04 has been implemented with the support provided by the National Research, Development, and Innovation Fund of Hungary, financed under the TKP2021-EGA funding scheme.
Institutional Review Board Statement
Animal experiments were in compliance with the guidelines of the European Community (86/609/EEC). The animal study protocol was approved by the Institutional Animal Care Committee of the University of Debrecen (3-1/2019/DEMAB).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on reasonable request.
Acknowledgments
The authors would like to thank Moran Kanaan (UniTalent student, University of Debrecen) for her help with molecular biology experiments and Judit Reményi-Puskár (University of Debrecen) for excellent technical assistance. The authors wish to thank Bradley Launikonis (University of Queensland, Brisbane, Australia) for fruitful discussion on the topic and critical review of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wolfe, R.R. The underappreciated role of muscle in health and disease. Am. J. Clin. Nutr. 2006, 84, 475–482. [Google Scholar] [CrossRef]
- Merz, K.E.; Thurmond, D.C. Role of skeletal muscle in insulin resistance and glucose uptake. Compr. Physiol. 2020, 10, 785–809. [Google Scholar] [CrossRef] [PubMed]
- Franzini-Armstrong, C.; Jorgensen, A.O. Structure and development of E-C coupling units in skeletal muscle. Annu. Rev. Physiol. 1994, 56, 509–534. [Google Scholar] [CrossRef]
- Meissner, G.; Lu, X. Dihydropyridine receptor-ryanodine receptor interactions in skeletal muscle excitation-contraction coupling. Biosci. Rep. 1995, 15, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, B.R. Quantitative Ultrastructure of Mammalian Skeletal Muscle. Compr. Physiol. 1983, 73–112. [Google Scholar] [CrossRef]
- Marcucci, L.; Canato, M.; Protasi, F.; Stienen, G.J.M.; Reggiani, C. A 3D diffusional-compartmental model of the calcium dynamics in cytosol, sarcoplasmic reticulum and mitochondria of murine skeletal muscle fibers. PLoS One 2018, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, R.; De Stefani, D.; Raffaello, A.; Mammucari, C. Mitochondria as sensors and regulators of calcium signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 566–578. [Google Scholar] [CrossRef]
- Marcucci, L.; Nogara, L.; Canato, M.; Germinario, E.; Raffaello, A.; Carraro, M.; Bernardi, P.; Pietrangelo, L.; Boncompagni, S.; Protasi, F.; et al. Mitochondria can substitute for parvalbumin to lower cytosolic calcium levels in the murine fast skeletal muscle. Acta Physiol. 2024, 240, 1–14. [Google Scholar] [CrossRef]
- Butera, G.; Vecellio Reane, D.; Canato, M.; Pietrangelo, L.; Boncompagni, S.; Protasi, F.; Rizzuto, R.; Reggiani, C.; Raffaello, A. Parvalbumin affects skeletal muscle trophism through modulation of mitochondrial calcium uptake. Cell Rep. 2021, 35, 109087. [Google Scholar] [CrossRef]
- Launikonis, B.S.; Ríos, E. Store-operated Ca2+ entry during intracellular Ca2+ release in mammalian skeletal muscle. J. Physiol. 2007, 583, 81–97. [Google Scholar] [CrossRef]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.-H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.; Kim, M.L.; Heo, W. Do; Jones, J.T.; Myers, J.W.; Ferrell, J.E.J.; Meyer, T. STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Hayashi, K.; Fujii, Y.; Mizushima, A.; Watarai, H.; Wakamori, M.; Numaga, T.; Mori, Y.; Iino, M.; Hikida, M.; et al. Coupling of STIM1 to store-operated Ca2+ entry through its constitutive and inducible movement in the endoplasmic reticulum. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 16704–16709. [Google Scholar] [CrossRef] [PubMed]
- Koenig, X.; Choi, R.H.; Launikonis, B.S. Store-operated Ca2+ entry is activated by every action potential in skeletal muscle. Commun. Biol. 2018, 1. [Google Scholar] [CrossRef]
- Zhang, S.L.; Yu, Y.; Roos, J.; Kozak, J.A.; Deerinck, T.J.; Ellisman, M.H.; Stauderman, K.A.; Cahalan, M.D. STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. Nature 2005, 437, 902–905. [Google Scholar] [CrossRef]
- Onopiuk, M.; Brutkowski, W.; Young, C.; Krasowska, E.; Róg, J.; Ritso, M.; Wojciechowska, S.; Arkle, S.; Zabłocki, K.; Górecki, D.C. Store-operated calcium entry contributes to abnormal Ca2+ signalling in dystrophic mdx mouse myoblasts. Arch. Biochem. Biophys. 2015, 569, 1–9. [Google Scholar] [CrossRef]
- Cavuoto, P.; McAinch, A.J.; Hatzinikolas, G.; Janovská, A.; Game, P.; Wittert, G.A. The expression of receptors for endocannabinoids in human and rodent skeletal muscle. Biochem. Biophys. Res. Commun. 2007, 364, 105–110. [Google Scholar] [CrossRef]
- Ruiz de Azua, I.; Lutz, B. Multiple endocannabinoid-mediated mechanisms in the regulation of energy homeostasis in brain and peripheral tissues. Cell. Mol. Life Sci. 2019, 76, 1341–1363. [Google Scholar] [CrossRef]
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid signalling and the deteriorating brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The emerging role of the endocannabinoid system in endocrine regulation and energy balance. Endocr. Rev. 2006, 27, 73–100. [Google Scholar] [CrossRef]
- Hudson, B.D.; Hébert, T.E.; Kelly, M.E. Physical and functional interaction between CB1 cannabinoid receptors and β2-adrenoceptors. Br. J. Pharmacol. 2010, 160, 627–642. [Google Scholar] [CrossRef]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, M.; Ricci, G.; Suglia, A.; Manfrevola, F.; Mackie, K.; Fasano, S.; Pierantoni, R.; Chioccarelli, T.; Cobellis, G. Analysis of endocannabinoid system in rat testis during the first spermatogenetic wave. Front. Endocrinol. (Lausanne). 2018, 9. [Google Scholar] [CrossRef]
- Sepúlveda, C.; Rodríguez, J.M.; Monsalves-Álvarez, M.; Donoso-Barraza, C.; Pino-de la Fuente, F.; Matías, I.; Leste-Lasserre, T.; Zizzari, P.; Morselli, E.; Cota, D.; et al. The CB1 cannabinoid receptor regulates autophagy in the tibialis anterior skeletal muscle in mice. Biol. Res. 2023, 56, 1–12. [Google Scholar] [CrossRef]
- Eckardt, K.; Sell, H.; Taube, A.; Koenen, M.; Platzbecker, B.; Cramer, A.; Horrighs, A.; Lehtonen, M.; Tennagels, N.; Eckel, J. Cannabinoid type 1 receptors in human skeletal muscle cells participate in the negative crosstalk between fat and muscle. Diabetologia 2009, 52, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Dalle, S.; Schouten, M.; Deboutte, J.; de Lange, E.; Ramaekers, M.; Koppo, K. The molecular signature of the peripheral cannabinoid receptor 1 antagonist AM6545 in adipose, liver and muscle tissue. Toxicol. Appl. Pharmacol. 2024, 491, 117081. [Google Scholar] [CrossRef] [PubMed]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef]
- Esposito, I.; Proto, M.C.; Gazzerro, P.; Laezza, C.; Miele, C.; Alberobello, A.T.; D’Esposito, V.; Beguinot, F.; Formisano, P.; Bifulco, M. The cannabinoid CB1 receptor antagonist rimonabant stimulates 2-deoxyglucose uptake in skeletal muscle cells by regulating the expression of phosphatidylinositol-3-kinase. Mol. Pharmacol. 2008, 74, 1678–1686. [Google Scholar] [CrossRef]
- Crespillo, A.; Suárez, J.; Bermúdez-Silva, F.J.; Rivera, P.; Vida, M.; Alonso, M.; Palomino, A.; Lucena, M.A.; Serrano, A.; Pérez-Martín, M.; et al. Expression of the cannabinoid system in muscle: Effects of a high-fat diet and CB1 receptor blockade. Biochem. J. 2011, 433, 175–185. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Reguero, L.; Puente, N.; Lutz, B.; Chaouloff, F.; Rossignol, R.; Piazza, P.V.; Benard, G.; Grandes, P.; Marsicano, G. Studying mitochondrial CB1 receptors: Yes we can. Mol. Metab. 2014, 3, 339. [Google Scholar] [CrossRef]
- Mendizabal-Zubiaga, J.; Melser, S.; Bénard, G.; Ramos, A.; Reguero, L.; Arrabal, S.; Elezgarai, I.; Gerrikagoitia, I.; Suarez, J.; De Fonseca, F.R.; et al. Cannabinoid CB1 receptors are localized in striated muscle mitochondria and regulate mitochondrial respiration. Front. Physiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Senese, R.; Petito, G.; Silvestri, E.; Ventriglia, M.; Mosca, N.; Potenza, N.; Russo, A.; Manfrevola, F.; Cobellis, G.; Chioccarelli, T.; et al. Effect of CB1 Receptor Deficiency on Mitochondrial Quality Control Pathways in Gastrocnemius Muscle. Biology (Basel). 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Singlár, Z.; Ganbat, N.; Szentesi, P.; Osgonsandag, N.; Szabó, L.; Telek, A.; Fodor, J.; Dienes, B.; Gönczi, M.; Csernoch, L.; et al. Genetic Manipulation of CB1 Cannabinoid Receptors Reveals a Role in Maintaining Proper Skeletal Muscle Morphology and Function in Mice. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Sztretye, M.; Geyer, N.; Vincze, J.; Al-Gaadi, D.; Oláh, T.; Szentesi, P.; Kis, G.; Antal, M.; Balatoni, I.; Csernoch, L.; et al. SOCE Is Important for Maintaining Sarcoplasmic Calcium Content and Release in Skeletal Muscle Fibers. Biophys. J. 2017, 113, 2496–2507. [Google Scholar] [CrossRef]
- Lamboley, C.R.; Pearce, L.; Seng, C.; Meizoso-Huesca, A.; Singh, D.P.; Frankish, B.P.; Kaura, V.; Lo, H.P.; Ferguson, C.; Allen, P.D.; et al. Ryanodine receptor leak triggers fiber Ca(2+) redistribution to preserve force and elevate basal metabolism in skeletal muscle. Sci. Adv. 2021, 7, eabi7166. [Google Scholar] [CrossRef]
- Cully, T.R.; Edwards, J.N.; Murphy, R.M.; Launikonis, B.S. A quantitative description of tubular system Ca2+ handling in fast- and slow-twitch muscle fibres. J. Physiol. 2016, 594, 2795–2810. [Google Scholar] [CrossRef] [PubMed]
- Lamb, G.D.; Stephenson, D.G. Measurement of force and calcium release using mechanically skinned fibers from mammalian skeletal muscle. J. Appl. Physiol. 2018, 125, 1105–1127. [Google Scholar] [CrossRef]
- Cully, T.R.; Choi, R.H.; Bjorksten, A.R.; Stephenson, D.G.; Murphy, R.M.; Launikonis, B.S. Junctional membrane Ca(2+) dynamics in human muscle fibers are altered by malignant hyperthermia causative RyR mutation. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 8215–8220. [Google Scholar] [CrossRef]
- Brini, M.; Carafoli, E. The plasma membrane Ca2+ ATPase and the plasma membrane sodium calcium exchanger cooperate in the regulation of cell calcium. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Oláh, T.; Bodnár, D.; Tóth, A.; Vincze, J.; Fodor, J.; Reischl, B.; Kovács, A.; Ruzsnavszky, O.; Dienes, B.; Szentesi, P.; et al. Cannabinoid signalling inhibits sarcoplasmic Ca2+ release and regulates excitation–contraction coupling in mammalian skeletal muscle. J. Physiol. 2016, 594, 7381–7398. [Google Scholar] [CrossRef]
- González-Mariscal, I.; Montoro, R.A.; O’Connel, J.F.; Kim, Y.; Gonzalez-Freire, M.; Liu, Q.R.; Alfaras, I.; Carlson, O.D.; Lehrmann, E.; Zhang, Y.; et al. Muscle cannabinoid 1 receptor regulates Il-6 and myostatin expression, governing physical performance and whole-body metabolism. FASEB J. 2019, 33, 5850–5863. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Silvestri, C.; Mazzarella, E.; Martella, A.; Calvigioni, D.; Piscitelli, F.; Ambrosino, P.; Petrosino, S.; Czifra, G.; Bíró, T.; et al. The endocannabinoid 2-AG controls skeletal muscle cell differentiation via CB1 receptor-dependent inhibition of Kv7 channels. Proc. Natl. Acad. Sci. 2014, 111, E2472–E2481. [Google Scholar] [CrossRef] [PubMed]
- Gaglianone, R.B.; Santos, A.T.; Bloise, F.F.; Ortiga-Carvalho, T.M.; Costa, M.L.; Quirico-Santos, T.; da Silva, W.S.; Mermelstein, C. Reduced mitochondrial respiration and increased calcium deposits in the EDL muscle, but not in soleus, from 12-week-old dystrophic mdx mice. Sci. Rep. 2019, 9, 1986. [Google Scholar] [CrossRef]
- Mareedu, S.; Million, E.D.; Duan, D.; Babu, G.J. Abnormal Calcium Handling in Duchenne Muscular Dystrophy: Mechanisms and Potential Therapies. Front. Physiol. 2021, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Koenig, X.; Choi, R.H.; Schicker, K.; Singh, D.P.; Hilber, K.; Launikonis, B.S. Mechanistic insights into store-operated Ca 2+ entry during excitation-contraction coupling in skeletal muscle. Biochim. Biophys. Acta - Mol. Cell Res. 2019, 1866, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.; Meizoso-Huesca, A.; Seng, C.; Lamboley, C.R.; Singh, D.P.; Launikonis, B.S. Ryanodine receptor activity and store-operated Ca2+ entry: Critical regulators of Ca2+ content and function in skeletal muscle. J. Physiol. 2023, 601, 4183–4202. [Google Scholar] [CrossRef]
- Bleck, C.K.E.; Kim, Y.; Willingham, T.B.; Glancy, B. Subcellular connectomic analyses of energy networks in striated muscle. Nat. Commun. 2018, 9, 5111. [Google Scholar] [CrossRef]
- Garbincius, J.F.; Elrod, J.W. Mitochondrial calcium exchange in physiology and disease. Physiol. Rev. 2022, 102, 893–992. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Zhao, F. MCU-Dependent mROS Generation Regulates Cell Metabolism and Cell Death Modulated by the AMPK/PGC-1α/SIRT3 Signaling Pathway. Front. Med. 2021, 8, 674986. [Google Scholar] [CrossRef]
- Pallafacchina, G.; Zanin, S.; Rizzuto, R. Recent advances in the molecular mechanism of mitochondrial calcium uptake. F1000Research 2018, 7. [Google Scholar] [CrossRef]
- Hasan, G. IP3Rs and nSOCE—Tied Together at Two Ends. Contact 2024, 7, 25152564241231092. [Google Scholar] [CrossRef]
- Mammucari, C.; Raffaello, A.; Vecellio Reane, D.; Gherardi, G.; De Mario, A.; Rizzuto, R. Mitochondrial calcium uptake in organ physiology: from molecular mechanism to animal models. Pflugers Arch. 2018, 470, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- McCommis, K.S.; Finck, B.N. Mitochondrial pyruvate transport: a historical perspective and future research directions. Biochem. J. 2015, 466, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Casanova, A.; Wevers, A.; Navarro-Ledesma, S.; Pruimboom, L. Mitochondria: It is all about energy. Front. Physiol. 2023, 14, 1114231. [Google Scholar] [CrossRef]
- Mailloux, R.J. Teaching the fundamentals of electron transfer reactions in mitochondria and the production and detection of reactive oxygen species. Redox Biol. 2015, 4, 381–398. [Google Scholar] [CrossRef]
- Tedesco, L.; Valerio, A.; Cervino, C.; Cardile, A.; Pagano, C.; Vettor, R.; Pasquali, R.; Carruba, M.O.; Marsicano, G.; Lutz, B.; et al. Cannabinoid Type 1 Receptor Blockade Promotes Oxide Synthase Expression in White Adipocytes. Diabetes 2008, 57, 2028–2036. [Google Scholar] [CrossRef]
- Abán, C.E.; Accialini, P.L.; Etcheverry, T.; Leguizamón, G.F.; Martinez, N.A.; Farina, M.G. Crosstalk Between Nitric Oxide and Endocannabinoid Signaling Pathways in Normal and Pathological Placentation. Front. Physiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Filadi, R.; Greotti, E.; Turacchio, G.; Luini, A.; Pozzan, T.; Pizzo, P. Mitofusin 2 ablation increases endoplasmic reticulum-mitochondria coupling. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, E2174–E2181. [Google Scholar] [CrossRef]
- Casellas-Díaz, S.; Larramona-Arcas, R.; Riqué-Pujol, G.; Tena-Morraja, P.; Müller-Sánchez, C.; Segarra-Mondejar, M.; Gavaldà-Navarro, A.; Villarroya, F.; Reina, M.; Martínez-Estrada, O.M.; et al. Mfn2 localization in the ER is necessary for its bioenergetic function and neuritic development. EMBO Rep. 2021, 22, e51954. [Google Scholar] [CrossRef]
- Murphy, E.; Steenbergen, C. Regulation of Mitochondrial Ca(2+) Uptake. Annu. Rev. Physiol. 2021, 83, 107–126. [Google Scholar] [CrossRef]
- Sztretye, M.; Singlár, Z.; Szabó, L.; Angyal, Á.; Balogh, N.; Vakilzadeh, F.; Szentesi, P.; Dienes, B.; Csernoch, L. Improved tetanic force and mitochondrial calcium homeostasis by astaxanthin treatment in mouse skeletal muscle. Antioxidants 2020, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cully, T.R.; Edwards, J.N.; Murphy, R.M.; Launikonis, B.S. A quantitative description of tubular system Ca2+ handling in fast- and slow-twitch muscle fibres. J. Physiol. 2016, 594, 2795–2810. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, D.G.; Williams, D.A. Calcium-activated force responses in fast- and slow-twitch skinned muscle fibres of the rat at different temperatures. J. Physiol. 1981, 317, 281–302. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef] [PubMed]
- Royer, L.; Pouvreau, S.; Ríos, E. Evolution and modulation of intracellular calcium release during long-lasting, depleting depolarization in mouse muscle. J. Physiol. 2008, 586, 4609–4629. [Google Scholar] [CrossRef]
- Melzer, W.; Rios, E.; Schneider, M.F. A general procedure for determining the rate of calcium release from the sarcoplasmic reticulum in skeletal muscle fibers. Biophys. J. 1987, 51, 849–863. [Google Scholar] [CrossRef]
Figure 1.
In vivo grip force and coordination experiments. (A) Experimental scheme for the evaluation of control (Cre-/-) and skmCB1-KD (Cre+/-) mice. Tamoxifen feeding was started at one month of age and lasted two months (created with BioRender). Inset shows the normalized CB1 mRNA expression measured via qPCR in m. tibialis anterior muscles. 18sRNA was used as housekeeping gene. n=2 technical replicates from 4 and 3 biological replicates for Cre-/- and Cre+/-, respectively. Student’s t-test was used to assess significance with ** at p<0.009. (B) Rota-Rod was used to assess coordination in mice and the average latency to fall was recorded using the protocol as illustrated. The acceleration speed was gradually increased from 4 to 300 rpm in 5 min. On the day of the evaluation, 3 trials were recorded with 15 min intervals and the average of these trials was noted for each animal. (C) Body weight monitoring during tamoxifen feeding. For each mouse, the body weight was assessed weekly. The graph presents the monthly average body weight starting from the beginning of the tamoxifen feeding at one month of age. N is the number of specimens investigated in each group (here, and all subsequent panels). (D) The average latency to fall was recorded at different age time points in Cre-/- an Cre+/- mice during the Rota-Rod test. Note that after one month of tamoxifen feeding the Cre+/- animals performed significantly worse and this tendency was maintained until the end of feeding. Significance was assessed with Student’s t-test with *p ≤ 0.05. (E) Grip force normalized to body weight in Cre−/− and Cre+/− mice. *p ≤ 0.05.
Figure 1.
In vivo grip force and coordination experiments. (A) Experimental scheme for the evaluation of control (Cre-/-) and skmCB1-KD (Cre+/-) mice. Tamoxifen feeding was started at one month of age and lasted two months (created with BioRender). Inset shows the normalized CB1 mRNA expression measured via qPCR in m. tibialis anterior muscles. 18sRNA was used as housekeeping gene. n=2 technical replicates from 4 and 3 biological replicates for Cre-/- and Cre+/-, respectively. Student’s t-test was used to assess significance with ** at p<0.009. (B) Rota-Rod was used to assess coordination in mice and the average latency to fall was recorded using the protocol as illustrated. The acceleration speed was gradually increased from 4 to 300 rpm in 5 min. On the day of the evaluation, 3 trials were recorded with 15 min intervals and the average of these trials was noted for each animal. (C) Body weight monitoring during tamoxifen feeding. For each mouse, the body weight was assessed weekly. The graph presents the monthly average body weight starting from the beginning of the tamoxifen feeding at one month of age. N is the number of specimens investigated in each group (here, and all subsequent panels). (D) The average latency to fall was recorded at different age time points in Cre-/- an Cre+/- mice during the Rota-Rod test. Note that after one month of tamoxifen feeding the Cre+/- animals performed significantly worse and this tendency was maintained until the end of feeding. Significance was assessed with Student’s t-test with *p ≤ 0.05. (E) Grip force normalized to body weight in Cre−/− and Cre+/− mice. *p ≤ 0.05.
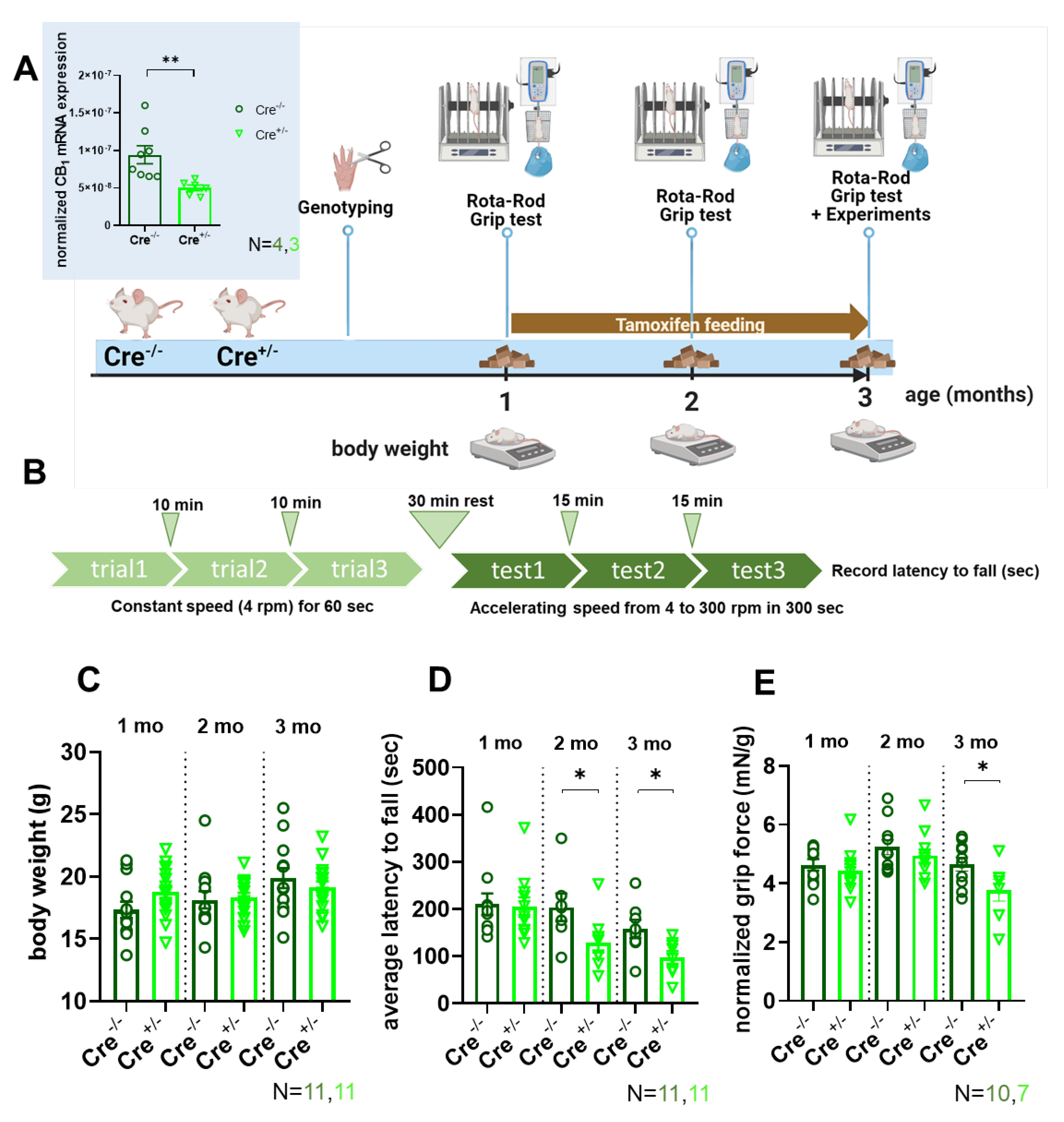
Figure 2.
Measurement of SOCE in skinned EDL fibres (A) Schematic representation of the protocol steps to perform measurements of SOCE in skinned muscle fibres of mouse EDL muscles: (1) Isolated EDL muscle fibre under paraffin oil with intact sarcolemma. (2) Incubation of the muscle fibre with a low-affinity Ca2+-sensitive dye (Rhod-5N) (3) Skinning of the fibres under paraffin oil traps Rhod-5N in the sealed t-system which allows measuring [Ca2+]t-sys and enables full access to the cytoplasm. (B) The steady state [Ca2+]t-sys at baseline and after reuptake of Ca2+ following full depletion in Ca2+ and Mg2+ free and 30 mM caffeine solution (see panels C and D) in Cre-/- and Cre+/- mice. Note that in the Cre+/- animals the reuptake was significantly smaller than in the Cre-/- specimens. The numbers of animals/fibres are given in brackets. ANOVA with Tukey’s post hoc test was performed. * and *** indicate statistical significance at p<0.05 and p<0.001, respectively. (C) and (D) Typical recording of [Ca2+]t-sys derived from the calibrated fluorescence of Rhod-5N in a control Cre-/- (C) and a skmCB1-KD Cre+/- (D) skinned fibre following steps 1–3 presented in panel A. The fibre is bathed in an internal solution containing 1 mM free Mg2+ and 67 nM free Ca2+. SOCE is induced through direct activation of the RyR1s by exposing the fibre to a “release” solution containing 0 mM free Mg2+ and 30 mM caffeine. Activation of pSOCE is seen as a steep depletion of [Ca2+]t-sys. The depletion is fully reversible as the t-system reloads with Ca2+ upon restoration of physiological [Ca2+]cyto and [Mg2+]cyto. Red traces show the t-system release flux calculated as the first-time derivative of the dark or bright green traces. Arrows indicate the start of the solution exchange to known calcium concentration content solutions. (E), (F) and (G) show [Ca2+]t-sys, the peak t-system release flux and the peak t-system uptake flux plotted as a function of [Ca2+]cyto. Note that the peak t-system uptake flux was altered at the highest concentration tested. *p ≤ 0.05. (H) Strong linear correlation between the peak t-system release flux (mMs-1) and [Ca2+]t-sys (mM), but no difference between Cre-/- and Cre+/- fibres. The t-system Ca2+-uptake proteins Na+/Ca2+ exchanger (NCX) and/or plasma membrane Ca2+ ATPase (PMCA) are known to equilibrate Ca2+ gradients across the t-system membrane in a bidirectional manner [38,39]. Ultimately, we found no changes in the expression of the latter on the protein level (Supplementary Figure S1D,E) suggesting that the observed reduction in t-system Ca2+-reuptake capacity may not relate to changes in these proteins but rather reflect changes at the level of the SR, such as altered SERCA or perhaps RyR1 function.
Figure 2.
Measurement of SOCE in skinned EDL fibres (A) Schematic representation of the protocol steps to perform measurements of SOCE in skinned muscle fibres of mouse EDL muscles: (1) Isolated EDL muscle fibre under paraffin oil with intact sarcolemma. (2) Incubation of the muscle fibre with a low-affinity Ca2+-sensitive dye (Rhod-5N) (3) Skinning of the fibres under paraffin oil traps Rhod-5N in the sealed t-system which allows measuring [Ca2+]t-sys and enables full access to the cytoplasm. (B) The steady state [Ca2+]t-sys at baseline and after reuptake of Ca2+ following full depletion in Ca2+ and Mg2+ free and 30 mM caffeine solution (see panels C and D) in Cre-/- and Cre+/- mice. Note that in the Cre+/- animals the reuptake was significantly smaller than in the Cre-/- specimens. The numbers of animals/fibres are given in brackets. ANOVA with Tukey’s post hoc test was performed. * and *** indicate statistical significance at p<0.05 and p<0.001, respectively. (C) and (D) Typical recording of [Ca2+]t-sys derived from the calibrated fluorescence of Rhod-5N in a control Cre-/- (C) and a skmCB1-KD Cre+/- (D) skinned fibre following steps 1–3 presented in panel A. The fibre is bathed in an internal solution containing 1 mM free Mg2+ and 67 nM free Ca2+. SOCE is induced through direct activation of the RyR1s by exposing the fibre to a “release” solution containing 0 mM free Mg2+ and 30 mM caffeine. Activation of pSOCE is seen as a steep depletion of [Ca2+]t-sys. The depletion is fully reversible as the t-system reloads with Ca2+ upon restoration of physiological [Ca2+]cyto and [Mg2+]cyto. Red traces show the t-system release flux calculated as the first-time derivative of the dark or bright green traces. Arrows indicate the start of the solution exchange to known calcium concentration content solutions. (E), (F) and (G) show [Ca2+]t-sys, the peak t-system release flux and the peak t-system uptake flux plotted as a function of [Ca2+]cyto. Note that the peak t-system uptake flux was altered at the highest concentration tested. *p ≤ 0.05. (H) Strong linear correlation between the peak t-system release flux (mMs-1) and [Ca2+]t-sys (mM), but no difference between Cre-/- and Cre+/- fibres. The t-system Ca2+-uptake proteins Na+/Ca2+ exchanger (NCX) and/or plasma membrane Ca2+ ATPase (PMCA) are known to equilibrate Ca2+ gradients across the t-system membrane in a bidirectional manner [38,39]. Ultimately, we found no changes in the expression of the latter on the protein level (Supplementary Figure S1D,E) suggesting that the observed reduction in t-system Ca2+-reuptake capacity may not relate to changes in these proteins but rather reflect changes at the level of the SR, such as altered SERCA or perhaps RyR1 function.
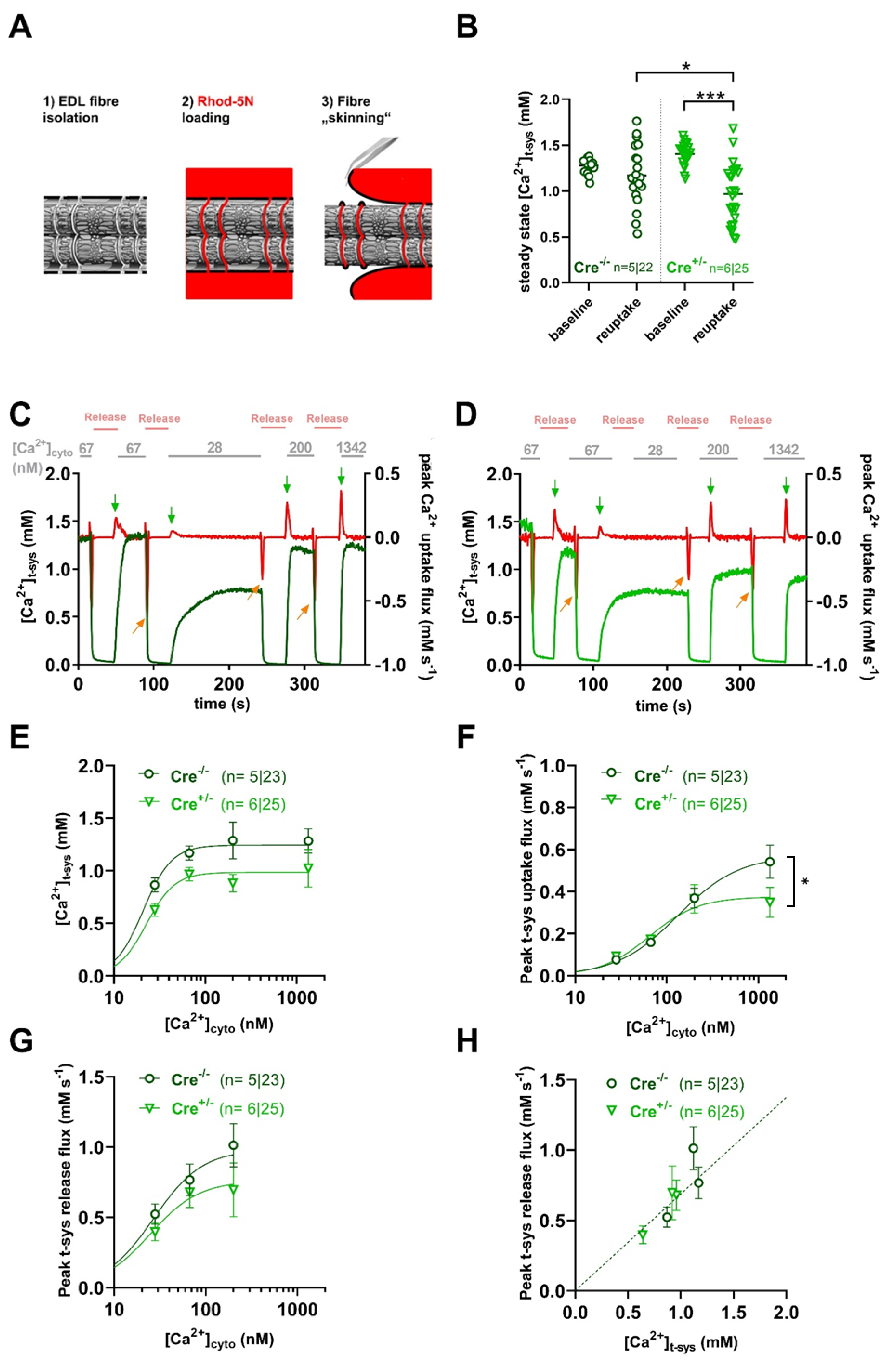
Figure 3.
Intracellular calcium homeostasis assessment in FDB fibres. (A) Representative line-scan image of Rhod-2 fluorescence normalized to the baseline value F0(x) in a Cre+/- FDB fibre subjected to successive rectangular depolarizing voltage steps (top black trace) under whole-cell voltage clamp. The Ca2+ transients were elicited by 200 ms long membrane depolarizations to +30 mV applied once per second. The white trace is the temporal profile of the normalized fluorescence obtained by averaging 50 lines in the spatial domain normalized to average resting F0(x) values. (B) For one representative Cre-/- and one Cre+/- fibre the SR Ca2+ content was estimated from the amount released during each depolarizing pulse minus the removal flux. A single-exponential function was fitted to the points obtained following 8 consecutive stimuli (see Eq. A10 in Appendix A in Sztretye et al. (2017) [34]) with the following parameters: y0=190.7 vs 158.7; a=375.4 vs 276.3; and b=0.65 vs 1.11, respectively where b is the time constant of SR depletion, and a is the remaining SR calcium content after the last applied stimulus. (C) [Ca2+]SR values were calculated for 7 and 11 FDB fibres from 3 Cre-/- and 6 Cre+/- animals, respectively. The calculation of [Ca2+]SR was based on similar exponential fits as seen in (B). On average, in the Cre+/- fibres (bright green) the SR calcium content was slightly lower than in the control Cre-/- specimens. This difference was not statistically different. (D) Pooled data of the calculated fluxes through Orai1 channels (K1) as predicted by our previously introduced model (see Appendix A in Sztretye et al. (2017) [34]) show non-significant calcium flux changes upon CB1 knockdown in the Cre+/- fibres. (E) The calculated SERCA pump activity (pVmax) is preserved in Cre+/- fibres. (F) The resting intracellular calcium concentration is decreased following CB1R knockdown. Results presented as violin plots are from 109 and 72 fibres, from 3 Cre-/- and 6 Cre+/- animals, respectively. Dashed lines depict the median (central horizontal line in the box) while the dotted lines indicate the upper and lower quartiles. * indicates statistical significance at p<0.03.
Figure 3.
Intracellular calcium homeostasis assessment in FDB fibres. (A) Representative line-scan image of Rhod-2 fluorescence normalized to the baseline value F0(x) in a Cre+/- FDB fibre subjected to successive rectangular depolarizing voltage steps (top black trace) under whole-cell voltage clamp. The Ca2+ transients were elicited by 200 ms long membrane depolarizations to +30 mV applied once per second. The white trace is the temporal profile of the normalized fluorescence obtained by averaging 50 lines in the spatial domain normalized to average resting F0(x) values. (B) For one representative Cre-/- and one Cre+/- fibre the SR Ca2+ content was estimated from the amount released during each depolarizing pulse minus the removal flux. A single-exponential function was fitted to the points obtained following 8 consecutive stimuli (see Eq. A10 in Appendix A in Sztretye et al. (2017) [34]) with the following parameters: y0=190.7 vs 158.7; a=375.4 vs 276.3; and b=0.65 vs 1.11, respectively where b is the time constant of SR depletion, and a is the remaining SR calcium content after the last applied stimulus. (C) [Ca2+]SR values were calculated for 7 and 11 FDB fibres from 3 Cre-/- and 6 Cre+/- animals, respectively. The calculation of [Ca2+]SR was based on similar exponential fits as seen in (B). On average, in the Cre+/- fibres (bright green) the SR calcium content was slightly lower than in the control Cre-/- specimens. This difference was not statistically different. (D) Pooled data of the calculated fluxes through Orai1 channels (K1) as predicted by our previously introduced model (see Appendix A in Sztretye et al. (2017) [34]) show non-significant calcium flux changes upon CB1 knockdown in the Cre+/- fibres. (E) The calculated SERCA pump activity (pVmax) is preserved in Cre+/- fibres. (F) The resting intracellular calcium concentration is decreased following CB1R knockdown. Results presented as violin plots are from 109 and 72 fibres, from 3 Cre-/- and 6 Cre+/- animals, respectively. Dashed lines depict the median (central horizontal line in the box) while the dotted lines indicate the upper and lower quartiles. * indicates statistical significance at p<0.03.
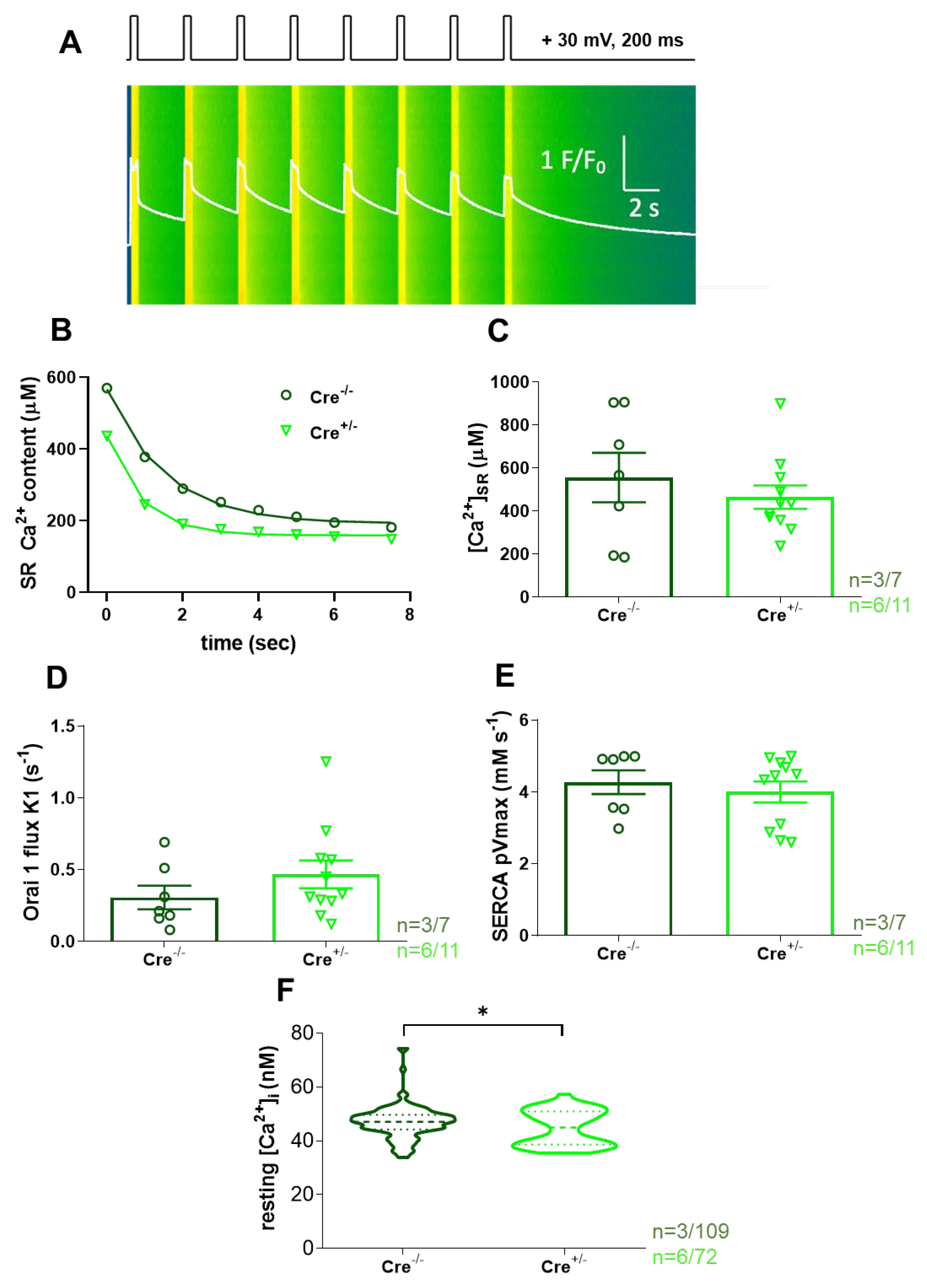
Figure 4.
Mitochondrial respiration measurement performed on gastrocnemius muscle fibers. (A) Representative traces of the short treatment protocol for mitochondrial respiration measurement; the lines indicate the chamber O2 concentrations (blue and cyan) and O2 consumption (dark and bright green) in the Cre-/- and Cre+/- muscles samples, respectively. (B) Representative plots of the long treatment protocol; the lines show chamber O2 concentrations (blue and cyan) and O2 consumption (dark and bright green) in Cre-/- and Cre+/- mice muscles. (C) Measured LEAKS state, oxidative phosphorylation (OXPHOS), LEAKOmy state are presented as O2 flux/V (pmol/[s × mL]). (D) Measured complex I-linked oxidative phosphorylation (complex I), complex II-linked oxidative phosphorylation (complex II), and complex IV respiratory activity (complex IV) are presented as O2 flux/V (pmol/[s × mL]). The box plots demonstrate the median (horizontal line in the box) and the 25th (lower whisker) and 75th (upper whisker) percentiles (n=6). * indicates statistical significance at p<0.01. (E) Baseline, residual oxygen consumption (ROX), and respiratory control ratio (RCR) are plotted as the ratio of OXPHOS and LEAKOmy status. * indicates statistical significance at *p<0.05. Abbreviations: Asc-TMPD, ascorbate and N,N,N',N'-Tetramethyl-p-phenylenediamine dihydrochloride; AmA, antimycin A; au, arbitrary unit; CytC, cytochrome c; Glu, glutamate and malate; Omy, oligomycin; Rot, rotenone; S, succinate; NaN3, Sodium azide.
Figure 4.
Mitochondrial respiration measurement performed on gastrocnemius muscle fibers. (A) Representative traces of the short treatment protocol for mitochondrial respiration measurement; the lines indicate the chamber O2 concentrations (blue and cyan) and O2 consumption (dark and bright green) in the Cre-/- and Cre+/- muscles samples, respectively. (B) Representative plots of the long treatment protocol; the lines show chamber O2 concentrations (blue and cyan) and O2 consumption (dark and bright green) in Cre-/- and Cre+/- mice muscles. (C) Measured LEAKS state, oxidative phosphorylation (OXPHOS), LEAKOmy state are presented as O2 flux/V (pmol/[s × mL]). (D) Measured complex I-linked oxidative phosphorylation (complex I), complex II-linked oxidative phosphorylation (complex II), and complex IV respiratory activity (complex IV) are presented as O2 flux/V (pmol/[s × mL]). The box plots demonstrate the median (horizontal line in the box) and the 25th (lower whisker) and 75th (upper whisker) percentiles (n=6). * indicates statistical significance at p<0.01. (E) Baseline, residual oxygen consumption (ROX), and respiratory control ratio (RCR) are plotted as the ratio of OXPHOS and LEAKOmy status. * indicates statistical significance at *p<0.05. Abbreviations: Asc-TMPD, ascorbate and N,N,N',N'-Tetramethyl-p-phenylenediamine dihydrochloride; AmA, antimycin A; au, arbitrary unit; CytC, cytochrome c; Glu, glutamate and malate; Omy, oligomycin; Rot, rotenone; S, succinate; NaN3, Sodium azide.
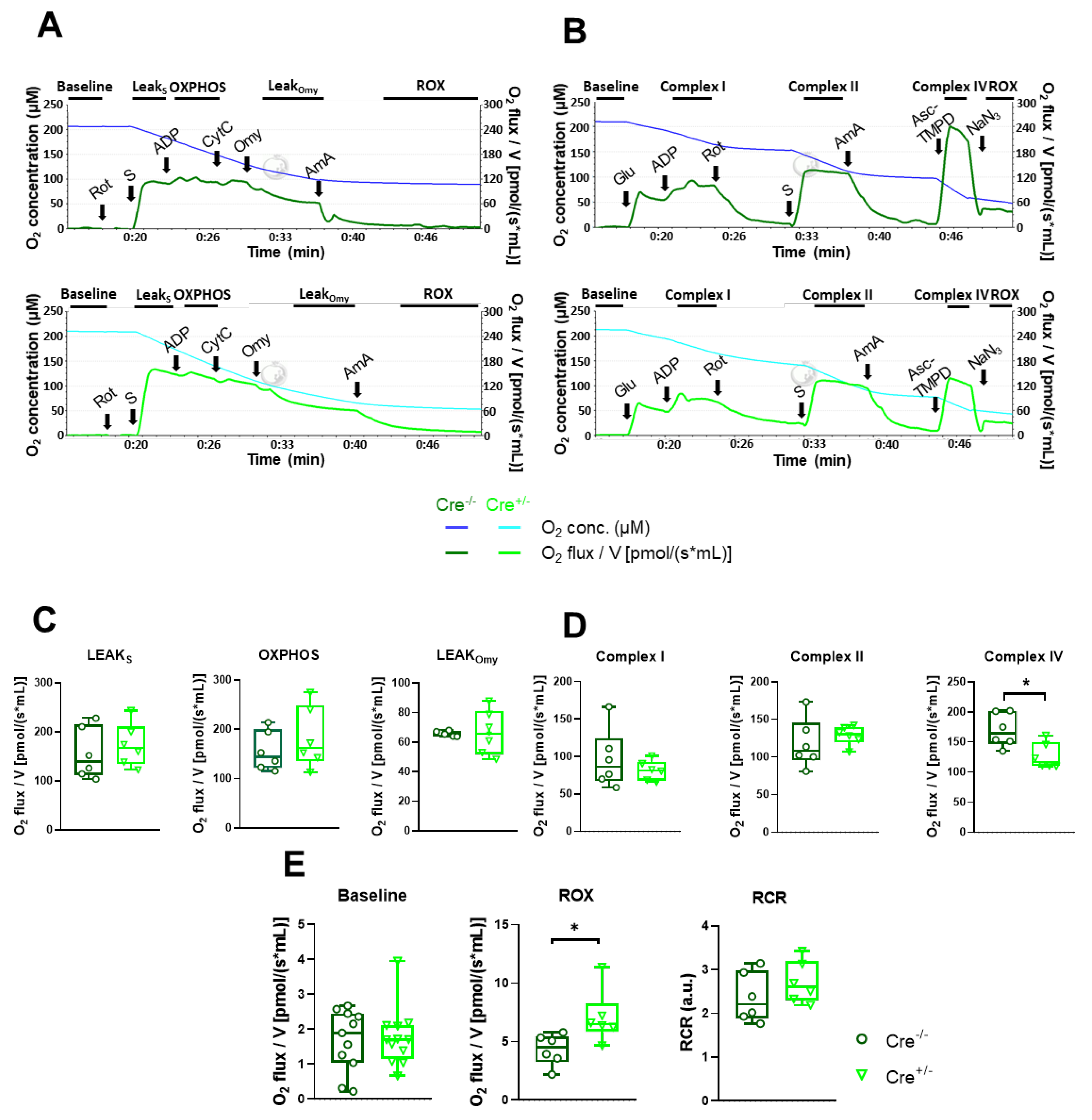
Figure 5.
Measurement of FCCP-dependent dissipation of mitochondrial membrane potential (Δψm) in FDB fibres. (A) (C) Representative confocal image series of TMRE fluorescence recorded on a FDB fibre from a Cre-/- (A) and a Cre+/-(C) mouse, respectively before (10 sec) and during the application of 1 µM FCCP (indicated in panel (B) and (D) by the blue horizontal line). Note the fading over time of the dye fluorescence as the un-coupler poisoned the organelle leading to Δψm loss. (B) (D) Normalized TMRE fluorescence for the two representative cells presented in panels (A) and (C). The red symbols and arrows indicate the time points when the image series shown in (A) and (C) were taken. The exponential fit (solid line) allowed for the calculation of Δψm decay times (τ) for the two representative examples as follows: τ =111.7 sec for Cre-/- and τ =121.13 sec for Cre+/-. (E) The averaged normalized TMRE fluorescence decay was slower in Cre+/- mice. Data are plotted as means ±SEM. In cyan is the average of 5 control (C57/Bl6) FDB cells where no FCCP was administered. (F) Averaged decay time values were significantly higher following FCCP administration in the skmCB1-KD specimens. *indicates statistical significance at *p<0.02. Abbreviations: TMRE, tetramethylrhodamine, ethyl ester; FCCP, carbonyl cyanide-p-trifluoromethoxyphenylhydrazone.
Figure 5.
Measurement of FCCP-dependent dissipation of mitochondrial membrane potential (Δψm) in FDB fibres. (A) (C) Representative confocal image series of TMRE fluorescence recorded on a FDB fibre from a Cre-/- (A) and a Cre+/-(C) mouse, respectively before (10 sec) and during the application of 1 µM FCCP (indicated in panel (B) and (D) by the blue horizontal line). Note the fading over time of the dye fluorescence as the un-coupler poisoned the organelle leading to Δψm loss. (B) (D) Normalized TMRE fluorescence for the two representative cells presented in panels (A) and (C). The red symbols and arrows indicate the time points when the image series shown in (A) and (C) were taken. The exponential fit (solid line) allowed for the calculation of Δψm decay times (τ) for the two representative examples as follows: τ =111.7 sec for Cre-/- and τ =121.13 sec for Cre+/-. (E) The averaged normalized TMRE fluorescence decay was slower in Cre+/- mice. Data are plotted as means ±SEM. In cyan is the average of 5 control (C57/Bl6) FDB cells where no FCCP was administered. (F) Averaged decay time values were significantly higher following FCCP administration in the skmCB1-KD specimens. *indicates statistical significance at *p<0.02. Abbreviations: TMRE, tetramethylrhodamine, ethyl ester; FCCP, carbonyl cyanide-p-trifluoromethoxyphenylhydrazone.
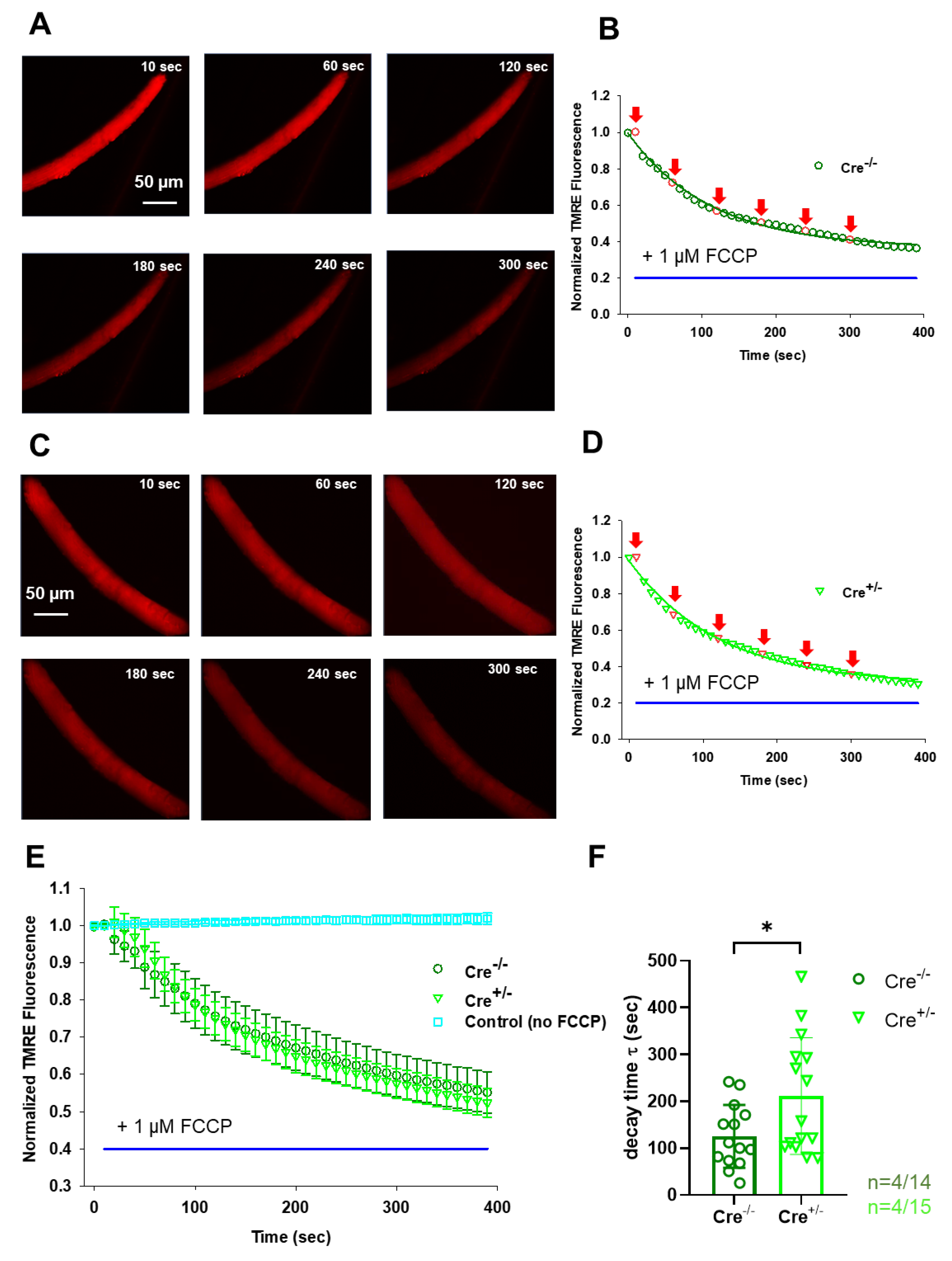
Figure 6.
Molecular insights into the downregulation of skmCB1R and its impact on muscle plasticity in murine skeletal muscles. Green arrows show the path followed by calcium ions; black arrows indicate the path followed by ATP to induce muscle contraction and underlie SERCA function; red arrows indicate the changes obtained in our current experiments following tamoxifen-induced muscle-specific CB1 downregulation. Abbreviations: pCB1R, plasma membrane cannabinoid receptor type 1; mtCB1R, mitochondrial cannabinoid receptor type 1; DHPR, dihydropyridine receptor; PMCA, plasma membrane calcium ATPase; Orai1, Calcium release-activated calcium channel protein 1; STIM1, Stromal interaction molecule 1; RyR1, ryanodine receptor type 1; SERCA, sarco(endo)plasmic reticulum calcium ATPase; NCX, sodium calcium exchanger; ATP, adenosine trisphosphate; ADP, adenosine diphosphate; VDAC, voltage activated anion channel; MCU, mitochondrial calcium uniporter; ROS, reactive oxygen species; TCA, Tricarboxylic acid; MAM, mitochondria-associated membrane.
Figure 6.
Molecular insights into the downregulation of skmCB1R and its impact on muscle plasticity in murine skeletal muscles. Green arrows show the path followed by calcium ions; black arrows indicate the path followed by ATP to induce muscle contraction and underlie SERCA function; red arrows indicate the changes obtained in our current experiments following tamoxifen-induced muscle-specific CB1 downregulation. Abbreviations: pCB1R, plasma membrane cannabinoid receptor type 1; mtCB1R, mitochondrial cannabinoid receptor type 1; DHPR, dihydropyridine receptor; PMCA, plasma membrane calcium ATPase; Orai1, Calcium release-activated calcium channel protein 1; STIM1, Stromal interaction molecule 1; RyR1, ryanodine receptor type 1; SERCA, sarco(endo)plasmic reticulum calcium ATPase; NCX, sodium calcium exchanger; ATP, adenosine trisphosphate; ADP, adenosine diphosphate; VDAC, voltage activated anion channel; MCU, mitochondrial calcium uniporter; ROS, reactive oxygen species; TCA, Tricarboxylic acid; MAM, mitochondria-associated membrane.
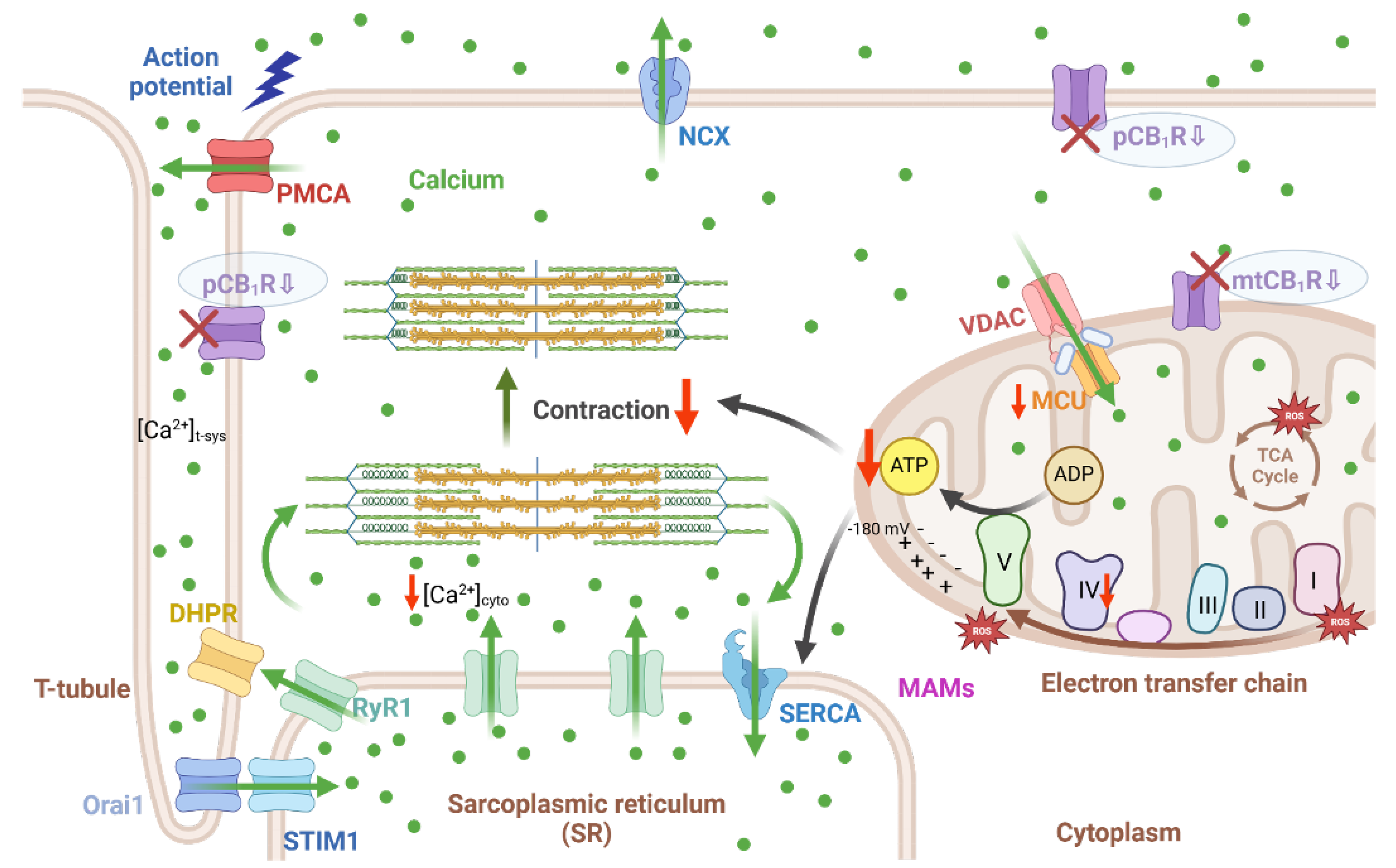

Table 3.
List if antibodies used in Western Blot analysis.
| Name | Cat. no. | Company | Dilution |
|---|---|---|---|
| anti-Orai1 | MA5-15777 | Invitrogen | 1:500 |
| anti-STIM1 | AB-9870 | Sigma | 1:1000 |
| anti-PMCA | 5F10 | Abcam | 1:1000 |
| anti-VDAC | MA5-35349 | Invitrogen | 1:1000 |
| anti-MICU1 | PA5-77364 | Invitrogen | 1:200 |
| anti-Drp1 | C6C7 | Cell Signaling Technology | 1:1000 |
| anti-Mfn2 | D2D10 | Cell Signaling Technology | 1:1000 |
| anti-Opa1 | MA-16149 | Invitrogen | 1:1000 |
| anti-α-actinin | sc-166524 | Santa Cruz Biotechnology | 1:1000 |
| anti-tubulin | T5168 | Sigma | 1:4000 |
Table 4.
Primer sequences used for identifying myosin heavy chain isoforms in muscle samples.
| Name | Forward primer | Reverse primer |
|---|---|---|
| MHCI | 5’-GAGTAGCTCTTGTGCTACCCAGC-3’ | 5’-AATTGCTTTATTCTGCTTCCACC-3’ |
| MHCIIA | 5’-GCAAGAAGCAGATCCAGAAAC-3’ | 5’-GGTCTTCTTCTGTCTGGTAAGTAAGC-3’ |
| MHCIIX | 5’-GCAACAGGAGATTTCTGACCTCAC-3’ | 5’-CCAGAGATGCCTCTGCTTC-3’ |
| 18s RNA | 5’-GGGAGCCTGAGAAACGGC-3’ | 5’-GGGTCGGGAGTGGGTAATTTT-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



