Submitted:
03 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
The field of biotechnology highly values natural compounds isolated from medicinal plants, but their discovery, purification, biofunctional characterization, and biochemical validation have historically been time-consuming and laborious processes. To more efficiently discover new bioactive substances, two innovative approaches have emerged: the predicted data mining approach (PDMA) and biotransformation-guided purification (BGP). PDMA is a computational method that predicts biotransformation potential, identifying potential substrates for specific enzymes from numerous candidate compounds to generate new compounds. BGP combines enzymatic biotransformation with traditional purification techniques to directly identify and isolate biotransformed products from crude extract fractions. This review examines recent research employing BGP or PDMA for novel compound discovery. These studies demonstrate that both approaches effectively discover novel bioactive molecules from natural sources, enhance bioactivity and solubility of existing compounds, and provide alternatives to traditional methods. The findings highlight the potential of integrating traditional medicinal knowledge with modern enzymatic and computational tools to advance drug discovery and development.
Keywords:
biotransformation
; enzymatic synthesis
; glycosylation
; glycosyltransferase
; glycoside hydrolase
; flavonoids
; predicted data mining
; phenolic compounds
; triterpenoids
1. Introduction
Natural products have long been fundamental resources for the development of health foods and drugs. The majority of natural products are secondary metabolites found in plants [1]. These secondary metabolites include alkaloids, terpenoids, and phenolic compounds [2,3]. The increasing need for new drugs to treat various human diseases has sparked a strong and sustained interest within the scientific community in discovering new bioactive compounds [4].
Isolating new compounds is the first step in the development of new drugs from herb plants. Traditionally, there are two strategies to isolate new compounds: isolating each natural compound from crude extract or chemical synthesis in a laboratory [4]. The discovery and isolation of new compounds from natural resources, such as medicinal plants, have been laborious and time-consuming. Moreover, numerous natural products have been isolated and identified in past decades; as a result, it has become increasingly difficult to isolate new compounds nowadays. On the other hand, organic synthesis is a promising method for candidate compounds; however, the synthetic procedures are usually harmful to the environment, such as toxic chemical reagents under high pressure or high temperature. Sometimes, it is difficult to achieve high production yields for complex chemical structures. Thus, an environmentally friendly approach to obtaining new compounds becomes more important. Instead of empirical screening, enzymatic biotransformation and particularly separation for candidate targets have provided more efficient approaches to generate novel compounds [2,3,5,6].
The structural modification of natural products using biotransformation is one effective way to produce regioselective and/or stereoselective compounds with specific bioactivities [2,3]. Hydroxylation, O-methylation and O-glycosylation are three major modifications in nature [1,7]. Various enzymes have been employed to modify functional groups onto compound backbones, such as hydroxyl, glycosyl, or methyl groups [2,3,5]. These enzymatic modifications often yield derivatives with significantly enhanced bioactivities or physicochemical properties compared to the original precursors.
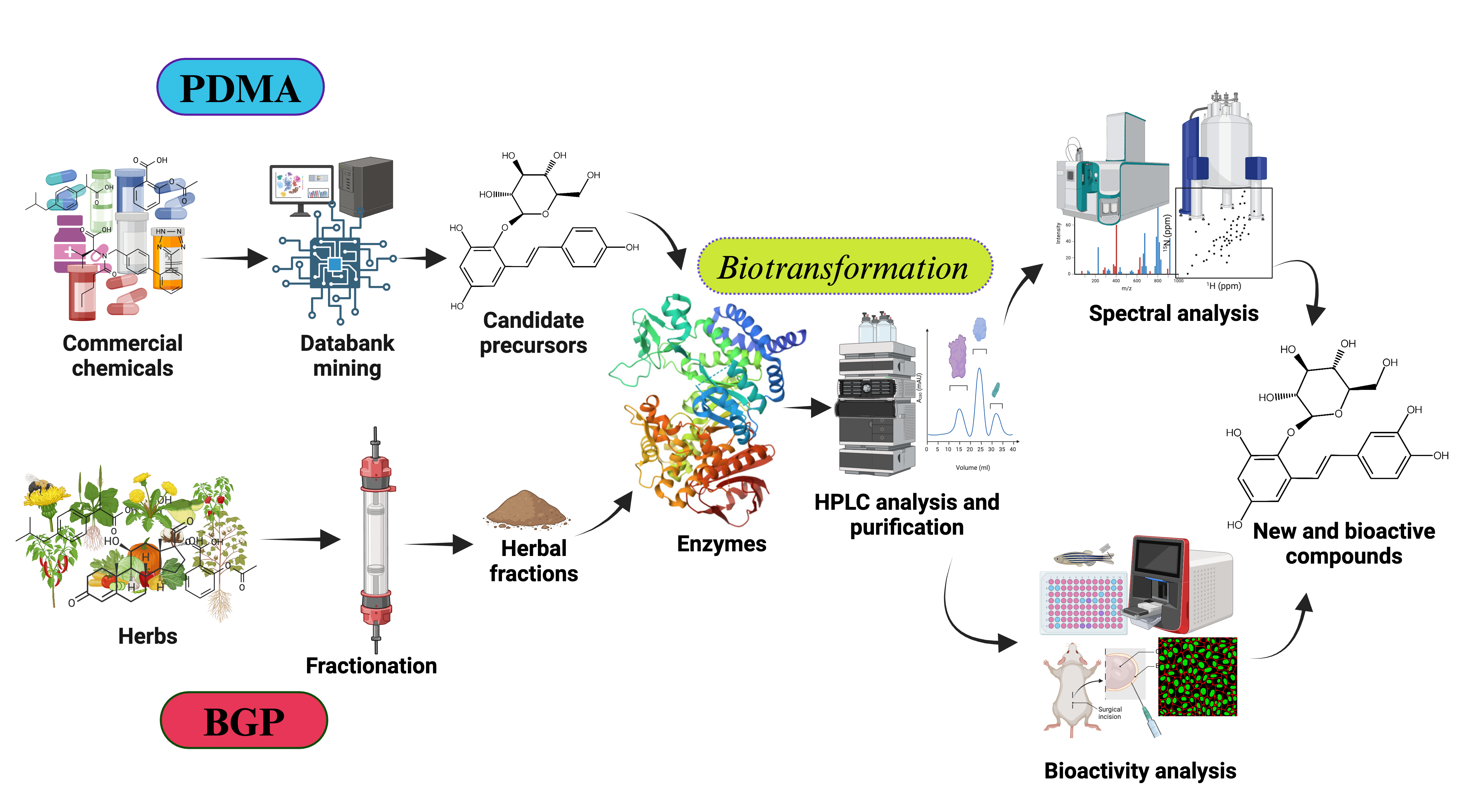
Although enzymatic synthesis offers a good choice to initiate finding new compounds in the new drug development, there are still some drawbacks and hence limit its applications. For example, the uncertainty of biotransformation product may not be a new compound after identification of chemical structures. one needs a systematic method to predict the novelty of the biotransformation products before designing experiments. Another limitation is the high cost of some commercial pure precursor, which means less amount in the raw materials and/or difficulty to isolate. For such high-priced precursors, it is inconvenient to study their biotransformation products. In reality, many identified compounds are not commercially available precursors; thus, it is much more difficult for further applications. To overcome these limitations, predicted data mining approaches (PDMA) was recently developed as in silico analysis to predict putative enzymatic biotransformation substrates for novel bioactive compounds (Figure 1) [8,9,10,11]. Indeed, PDMA may quickly screen out candidate therapeutic agents derived from available precursors in natural sources. On the other hand, a particularly effective strategy that integrates enzymatic biotransformation with an upstream process is biotransformation-guided purification (BGP, Figure 1) [13,14,15,16]. BGP uses herbal extracts as starting materials to find high-cost or non-commercially available compounds, which there are higher chances to find biotransformable (new) compounds. By monitoring the formation of new-peak compounds, BGP could guide the direct identification and isolation of biotransformed products from complex mixtures, following enzymatic validation with desired properties or bioactivities. This minireview collects studied cases using PDMA or BGP approaches to quickly produce 10 new compounds from either commercial precursors in PDMA cases or herbal extracts in BGP cases (Table 1).
2. Finding New Compounds by Predicted Data Mining Approach (PDMA)
The scientific community is continuously pursuing novel compounds with biological activities. Enzyme-catalyzed biotransformation is a method that uses the catalytic power of enzymes to modify compound structures. Biotransformation has garnered significant attention in drug development due to its high specificity, efficiency, and ability to carry out reactions that are challenging for chemical synthesis [2,3,5,6]. However, conventional enzymatic biotransformation primarily relies on a trial-and-error approach, which is time-consuming and involves randomly combining different enzymes and precursors in experiments, leading to low efficiency and high costs.
To overcome these limitations, PDMA (Figure 2) has been developed. The main idea of PDMA is to utilize the known catalytic properties of enzymes and precursor/chemical structures to predict putative products through in silico computational screening. Based on known chemical databases, this strategy aims to efficiently filter out novel compounds, which reduces the time and cost of experimental validations and prevents resource wastage.
This section will delve into the core principles, general workflow, and advantages of PDMA compared to traditional methods. Five recent studies have demonstrated how PDMA has been applied to various enzymatic biotransformation reactions (hydroxylation, glycosylation, and methylation). PDMA demonstrates its effectiveness in discovering novel compounds with potential applications in pharmacology and biotechnology.
PDMA generally follows six steps (Figure 2) although specific implementation details may vary depending on the research objectives and the enzymes used:
- Setting Screening Criteria (selecting suitable enzymatic reaction): Define clear screening criteria based on the target enzyme's known catalytic mechanism, substrate preference, and desired product characteristics. These criteria may include specific functional groups, structural features, physicochemical properties of precursor compounds, available at an industrial scale. For example, hydroxylation by tyrosinase (BmTYR) requires precursors contained a phenyl group mimicking the structure of tyrosine. glycosylation by glycosyltransferases (GTs) needs precursors with at least one hydroxyl group that can be glycosylated. O-Methylation by O-methyltransferases (OMTs) needs precursors with a catechol structure.
- Screening Candidate Precursors: Based on the defined screening criteria, potential candidate precursors could be screened from commercial chemical or natural product databases. These databases usually contain a vast amount of compound structures and related information. In some cases, customized catalogs of commercially available compounds are used.
- Predicting Biotransformation Product Structure: For the selected candidate precursors, the structures of potential biotransformation products under the action of the target enzyme are determined using chemical drawing software (such as Reaxys® or SciFindern®). This step requires researchers to have a certain knowledge on the enzyme's catalytic mechanism; for instance, BmTYR primarily catalyzes ortho-hydroxylation, GTs catalyze the transfer of sugar moieties, and OMTs catalyze the transfer of methyl groups.
- Verifying Product Novelty: The predicted biotransformation product structures are uploaded to chemical databases (e.g., Reaxys®, PubChem®, or SciFindern®) to verify their novelty, confirming whether each product is a known compound. Only precursors that yield novel derivatives are further selected for subsequent experimental validation.
- In Vitro Biotransformation and Product Identification: The selected precursors are reacted with the target enzyme in vitro. The biotransformed products are analyzed using isolation methods, such as high-performance liquid chromatography (HPLC). Once the putative new compounds are purified, their chemical structures can be identified using techniques such as mass spectrometry (MS) and nuclear magnetic resonance (NMR).
- Bioactivity Evaluation: Alternatively, the identified compounds may undergo bioactivity testing to evaluate their potential application value. The tested activities may include antioxidant, anti-inflammatory, anticancer, and anti-diabetic properties, etc.
The first reported case of PDMA involved the hydroxylation of Dragon's Blood components by BmTYR to enhance antioxidant and anti-α-glucosidase activities (Figure 3) [8]. Kim et al. have developed an efficient hydroxylation system for flavonoids using BmTYR in the presence of ascorbic acid and boric acid [17]. In our previous study, we also applied tyrosinase to hydroxylate soybean isoflavone, daidzin, and genistin to produce hydroxylated isoflavones [18]. These studies have revealed that BmTYR is a promiscuous biocatalyst for flavonoid hydroxylation. Thus, BmTYR was selected as the first trial to validate the main idea of PDMA. The catalytic criteria of BmTYR included containing a phenyl group mimicking tyrosine. PDMA further screened 475 compounds and identified loureirin A and loureirin B as two candidate precursors. The data mining further confirmed the predicted products, 3'-hydroxyloureirin A and 3'-hydroxyloureirin B, are novel flavonoids. Thus, the biotransformation was conducted, and the experimental validation confirmed that BmTYR indeed convert loureirin A and loureirin B into the two flavonoids. In addition, bioactivity assays revealed that these new compounds exhibited significantly higher antioxidant activity than ascorbic acid and possessed remarkable anti-α-glucosidase activity. PDMA quickly screened hundreds of compounds and identified two potential novel flavonoids via enzymatic biotransformation.
Therefore, Therefore, PDMA was further applied to facilitate the ortho-hydroxylation by BmTYR to generate compounds with enhanced antioxidant and anti-inflammatory activities (Figure 3) [9]. This study screened 764 commercially available chemical compounds that are suitable as substrates of BmTYR to produce new catecholic compounds. Only isoxsuprine was selected as a potential substrate due to its phenolic structure. PDMA predicted that BmTYR would catalyze the hydroxylation of isoxsuprine, yielding a novel catechol product. Experimental validation confirmed that isoxsuprine could be biotransformed into the novel compound 3''-hydroxyisoxsuprine. Bioactivity assays further demonstrated that 3''-hydroxyisoxsuprine exhibited 40-fold higher antioxidant activity and 10-fold higher anti-inflammatory activity than isoxsuprine.
PDMA was further applied to screen 412 commercial natural compounds for potential glycosylation by a bacterial glycosyltransferase (GT), BsUGT489 from Bacillus subtilis American type culture collection (ATCC) 6633 strain. In nature, many compounds could be glycosylated by GT to form glycosides. Glycosylation of molecules is one of the promising strategies to improve water solubility. After glycosylation, the water solubility of the glycosylated derivatives would be greatly improved [19,20,21,22]. Thus, glycosylation of medicines can also improve the oral bioavailability of the original molecules [23]. Based on the advantages described above, glycosylation coupled with PDMA to find new and bioactive glycosides was also developed. By coupling GT with PDMA, the screening criteria included industrial-scale availability, possessing one hydroxyl group, and the predicted glucoside product being novel in the SciFindern® database. Corylin was predicted to be glycosylated by BsUGT489 and yield a novel glucoside. Molecular docking analysis also predicted corylin as a compatible substrate. Experimental validation and structural analysis further validated the new glucoside, corylin-7-O-β-glucoside (Figure 4) [10]. The new glucoside demonstrated significantly enhanced aqueous solubility, retained anti-inflammatory activity, and exhibited more potent anti-melanoma activity than the precursor, corylin. Through a traditional approach, another clinical team also identified corylin from 22 studied compounds, which is the one potential compound promoted longevity in eukaryotic cells [24]. PDMA is indeed a cost-effective strategy to quickly survey potential drug development and could be extended to clinical trials or disease therapy in the future.
Another report screening 1143 natural compounds showed that skullcapflavone II glycosylated by BsUGT489 may produce a highly soluble and anti-melanoma flavone glucoside (Figure 4) [11]. This study utilized PDMA combined with molecular docking analysis to predict that glucoside derivatives of skullcapflavone II would be novel flavonoid compounds with higher binding potential to oncogenic proteins. Experimentally, BsUGT489 was proven effective in glycosylating skullcapflavone II into a novel compound skullcapflavone II-6'-O-β-glucoside. Moreover, the novel glucoside exhibited 272-fold higher aqueous solubility than its precursor and demonstrated noteworthy anti-melanoma activity.
Since PDMA is a functionally validated method for hydroxylation and glycosylation, PDMA coupled with a bacterial O-methyltransferases (OMTs) was investigated to find new methyl compounds. Methylation is also a common modification on natural products [25]. In nature, methylation is mainly catalyzed by OMTs, which transfer the methyl group of a methyl donor molecule, S-adenosyl methionine (SAM), to an acceptor molecule [26]. Many O-methyl natural products have been identified with multiple bioactivities, especially in plant metabolites [1,7,27,28]. The bioactivities of some natural products can be greatly improved after O-methylation using bacterial OMTs [29,30]. Accordingly, a recent study utilized PDMA coupled with a Streptomyces peucetius O-methyltransferase (SpOMt2884) to screen 4364 commercially available natural compounds with catechol structures as potential substrates. Screening criteria were based on the catalytic characteristics of SpOMt2884 and the novelty of the predicted methylated products in the SciFinder® database. Eight precursors were predicted to form novel methyl derivatives, and seven of them (including plantagoside and protosappanin B) were successfully biotransformed by SpOMT2884. Two novel compounds, 4'-O-methyl plantagoside and 5'-O-methyl plantagoside, were identified in advance by NMR as new methylated plantagoside derivatives (Figure 5). This study demonstrated that PDMA does indeed help to find many new derivatives. However, the bioactivity of these methylated compounds was restricted to the quick functional screening. Nevertheless, other bioactivities of these compounds could be investigated via high-throughput drug screening assay in the future.
Compared with traditional trial-and-error methods, the application of PDMA across different enzyme types (hydrolases and transferases) and chemical modifications (hydroxylation, glycosylation, and methylation) highlights its broad applicability. All these applications exhibit great potential in the fields of pharmacology and biotechnology. In short, PDMA offers the following significant advantages:
- High Efficiency: PDMA enables rapid in silico screening of a large number of compounds, targeting potential precursors and thereby significantly reducing the time required to find suitable biotransformation substrates.
- Reduced Cost: By minimizing the number of trial-and-error experiments, PDMA helps to lower the consumption of experimental reagents, enzymes, and human resources, as well as reducing costs associated with clinical trials.
- Predicting Novelty: PDMA predicts whether the product is a novel compound before experimentation, avoiding the risk of redundant research on known compounds and increasing the likelihood of discovering new entities.
- Knowledge-Based Guidance: PDMA can predict outcomes based on the enzyme's characteristics and the precursor's structure, experimental design and helping researchers better understand the potential results of biotransformation reactions.
- Applicable to Various Enzymes and Reactions: The PDMA concept is not limited to specific enzymes or reaction types. It can be adapted based on the catalytic properties of different enzymes and applied to various biotransformation processes, including hydroxylation, glycosylation, and methylation.
3. Finding New Compounds by Biotransformation-Guided Purification (BGP)
In most natural product biotransformations, pure compounds (precursors) are necessary substrates. There are two major ways to obtain these pure compounds: either purifying them from natural sources or purchasing them from chemical companies. However, the traditional way to isolate pure compounds from plants, for example, is often laborious, time-consuming, and yields limited amounts of the desired molecules for drug discovery [1]. Moreover, few new compounds are identified from such purified compounds, since most have already been characterized in past decades. For commercially prepared pure compounds, the high cost of some pure precursors limits such investigations. Even when precursors are available, these compounds may not be novel after biotransformation. All these disadvantages create a high risk of failure in isolating new drugs.
In contrast, herbal crude extracts serve as another low-cost precursor resource, but their complex mixture of various compounds can be difficult to use in biotransformation. To overcome this challenge, BGP has emerged as a complementary strategy for generating novel bioactive compounds [Figure 6a]. BGP represents an innovative approach that integrates enzymatic biotransformation with traditional purification techniques (chromatography). The workflow of BGP involves: (1) crude extraction of the natural source, (2) partial purification or fractionation of the crude extract, (3) enzymatic biotransformation of the fractionated extract, and (4) purification and identification of the biotransformed products [Figure 6a]. In contrast, the workflow of chemical purification involves multiple-step chromatography to isolate each natural compound step by step [Figure 6b].
BGP can quickly isolate biotransformable compounds from crude extracts. For example, the Chinese medicinal fungus Ganoderma lucidum possesses over 300 fungal triterpenoids; however, only three Ganoderma triterpenoid glycosides have been found so far. This phenomenon may be due to a lack of specific glycosyltransferases (GTs) required to form fungal terpenoid glycosides. Indeed, more than 20,000 GT1 enzymes have been identified [31], but only a few GT1 genes belong to fungi, which may explain why fewer terpenoid glycosides are found in fungi. Thus, Ganoderma lucidum extract is a suitable crude extract for use in BGP, coupled with recombinant GTs to produce new glycosides.
Ding et al. (2023) successfully utilized BGP coupling with the Bacillus GT, BsUGT489, to generate novel Ganoderma triterpenoid saponins from a commercial Ganoderma extract (Figure 7) [13]. The commercial Ganoderma extract was partially separated into three fractions by preparative HPLC equipped with a C18 column, and these fractions were directly biotransformed by recombinant BsUGT489. One major biotransformation product could be purified from fraction F1 and identified as a novel saponin, ganoderic acid C2 (GAC2)-3-O-β-glucoside, confirmed by NMR and mass spectral analyses. Further validation revealed that commercial GAC2 indeed could be biotransformed by BsUGT489 into four novel saponins, including GAC2-26-O-β-glucoside, GAC2-15-O-β-glucoside, GAC2-3-O-β-glucoside and a novel diglucoside, GAC2-3,15-O-β-diglucoside. Importantly, GAC2-3-O-β-glucoside and GAC2-3,15-O-β-diglucoside exhibited significantly higher aqueous solubility (17-fold and 200-fold, respectively) than GAC2. Additionally, GAC2-3-O-β-glucoside retained significant anti-α-glucosidase activity, comparable to the anti-diabetes drug acarbose. This case study reveals that the efficient BGP not only easily discovers novel compounds but also isolates biotransformable compounds for the desired physicochemical properties, such as improved solubility or enhanced bioactivities.
In addition to GT1s, GHs are another type of enzyme capable of glycosylation of flavonoids [32,33]. GT1s glycosylate flavonoids via a β-glycosidic linkage. In contrast to GT1s, GHs can glycosylate flavonoids via an α-glycosidic linkage. Both α-glycosidic and β-glycosidic flavonoids are more soluble than flavonoids [10,19,23,34]. However, GT1s use expensive uridine diphosphate-glucose (UDP-G) as the sugar donor [35] , whereas GHs use cheaper sugars, such as starch, maltodextrin, maltose, and sucrose, as donors during glycosylation [32]. Therefore, GHs are generally used for the bio-industrial production of glycosylated molecules. Moreover, all plant glycoside derivatives of natural products could be produced by its own GT1s, which means plant glycosides are almost β-form. Thus, GHs are promiscuously suitable enzymes for the production of new α-glycosides from natural compounds. The carbohydrate-activating enzyme (CAZY) databank already records 1.9 million GHs, classified into 190 distinct families [36]. Since many putative enzymes were still unverified, new glycosylated derivatives of natural products that bear suitable structures as acceptors can be designed and synthesized with new enzymes in the future. There are several successful cases as follows:
Amylosucrase (AS, E.C. 2.4.1.4) is a versatile sucrose-hydrolyzing enzyme that belongs to GH family 13 (GH13) [37,38,39,40,41]. AS can catalyze the synthesis of α-1,4-glucans using sucrose as the sole substrate. Furthermore, it can mediate the glycosylation of various small molecules with sucrose. Recombinant Deinococcus geothermalis amylosucrase (DgAS) is a very famous AS in glycosylation of natural products [37,41]. At least 10 natural products have been demonstrated to be suitable substrates of DgAS, such as simple phenolics, hydroquinone [42]; flavonoids, catechin [43], rutin [44], daidzin [45,46,47], puerarin [48], 8-hydroxydaidzein [21,49], isoquercitrin [40]; Ganoderma triterpenoids, ganoderic acid A (GAG), ganoderic acid G (GAG), and ganoderic acid F (GAF) [50].
Based on the powerful glycosylation activity of DgAS toward natural products, Chang et al. (2024) employed the BGP approach with DgAS to produce a novel glycoside from the extracts of the Chinese herb Baizhi (Angelica dahurica) (Figure 8) [14]. Baizhi is known for its anti-inflammatory and analgesic properties, and the researchers aimed to enhance the properties of its bioactive constituents through glycosylation. Initial biotransformation of the crude Baizhi extract with DgAS yielded limited products due to unknown inhibition by chemicals or compounds in the extract. Thus, the extract was further partitioned into four fractions by HPLC, and fraction 2 yielded a significant amount of a putative novel compound upon reaction with DgAS. The biotransformed compound was confirmed as byakangelicin-7’’-O-α-glucopyranoside (BG-G3), a new alpha glucoside derivative of byakangelicin (Figure 8). Further experiments validated that DgAS indeed glycosylate pure byakangelicin to produce BG-G3. Remarkably, the aqueous solubility of byakangelicin-7’’-O-α-glucoside was found to be over 29,000-fold greater than that of byakangelicin. This significant improvement in solubility extends a potential clinical trial for enhanced bioavailability and therapeutic efficacy. This study exemplifies BGP concept may overcome the unknown inhibition or low efficiency of enzyme within crude extract and quickly target novel derivatives or biotransformable compounds with improved pharmaceutical properties.
From the above Baizhi case, biotransformation efficiency can be improved to use fraction mixtures as the precursors. This might be because some competitive substrates or inhibitors were removed through chromatographic fractionation, and the fraction mixtures become more suitable for biotransformation in BGP. BGP indeed saves a lot of effort and cost for target compounds (precursors), instead of using commercial pure compounds or isolated precursors one by one.
In addition to glycosylation, Wu et al. used BGP coupling with BmTYR (ortho-hydroxylation) to produce butin from licorice (Glycyrrhiza) (Figure 9) [16]. Licorice is a widely used traditional Chinese medicine and contains various flavonoids with valuable bioactivities [51]. Thus, the researchers employed BmTYR to catalyze the biotransformation of a commercial licorice extract. However, BmTYR failed to catalyze significant product from either the methanol crude extract or the ethyl acetate extract. To overcome this, the ethyl acetate extract was further fractionated using Sephadex LH-20 chromatography. One fraction (F2) yielded a significant biotransformation product upon incubation with BmTYR. This product was subsequently purified using preparative C-18 via high-performance liquid chromatography (HPLC) and identified as butin (3′-hydroxyliquiritigenin) through nuclear magnetic resonance (NMR) and mass spectral analyses. The study further confirmed that butin was produced from liquiritigenin, a major flavonoid in licorice, through BmTYR-catalyzed hydroxylation (Figure 9). Notably, butin exhibited over 100-fold stronger antioxidant activity compared to its precursor, liquiritigenin. Moreover, the BGP process yielded a significantly higher amount of butin (0.15% from licorice medicine) compared to traditional isolation methods from other natural sources (0.0001% to 0.0055%). This case study highlights that BGP is not only used to identify novel bioactive molecules, but also to enhance their bioactivity and achieve a higher yield of desired compounds at low cost.
Wu et al. (2022) also applied BGP to the Chinese herb Ha-Soo-Oh (Polygonum multiflorum) using BmTYR as a biocatalyst, leading to the discovery of a new stilbene glucoside, 2,3,5,3′,4′-pentahydroxystilbene-2-O-β-glucoside (PSG) (Figure 9) [15]. Direct biotransformation of the Ha-Soo-Oh extract with BmTYR resulted in the formation of a major product, which was purified by preparative HPLC and identified as PSG using mass spectrometry and NMR. Further investigation confirmed that PSG was a hydroxylation product of 3,5,4′-trihydroxystilbene-2-O-β-glucoside (TSG), a main stilbene glucoside in Ha-Soo-Oh and a quality index for this herb (Figure 9). The new compound PSG demonstrated a 4.7-fold higher 1,1-diphenyl-2-picrylhydrazine (DPPH) free radical scavenging activity than its precursor TSG. This study further underscores the utility of BGP in discovering novel bioactive compounds from traditional Chinese medicines and in quickly identifying valuable precursors
Previous studies highlight the significant advantages of the BGP strategy in natural product research:
- Discovery of Novel Bioactive Compounds: BGP facilitates the identification of new molecules that are structurally related to known bioactive precursors but possess altered or enhanced properties.
- Enhanced Bioactivity: Enzymatic biotransformation can modify the functional groups of precursor molecules, leading to derivatives with significantly improved bioactivity, as demonstrated by the enhanced antioxidant activity of butin and PSG.
- Improved Physicochemical Properties: BGP can be used to generate derivatives with enhanced pharmaceutical properties, such as significantly increased aqueous solubility, as seen with the Ganoderma glucosides GAC2-3-O-β-glucoside and GAC2-3,15-O-β-diglucoside and byakangelicin-7’’-O-α-glucoside from Baizhi.
- Increased Yield of Active Ingredients: By selectively biotransforming a specific precursor within a complex mixture and then purifying the valuable product, BGP can sometimes lead to higher yields of the target compound compared to direct isolation from the natural source, as observed in the production of butin.
- Cost-Effectiveness: BGP can utilize crude or partially purify extracts as starting materials, potentially reduce the need for extensive initial purification of precursors, and lead the process more economical.
- Efficiency in Screening Biotransformable Compounds: BGP offers an efficient method to screen complex natural extracts for compounds that can be biotransformed by specific enzymes, rather than testing expensive pure compounds individually.
However, the application of BGP also involves certain considerations. First, the success of BGP heavily relies on the choice of an appropriate enzyme with the desired catalytic activity and substrate specificity. In the case of Baizhi, the used enzyme DgAS could catalyze unique biotransformation, α-glycosylation of flavonoids, which does not occur in natural plants. Thus, applying a unique functional enzyme with BGP is a good choice to get new and bioactive compounds. Secondary, crude extracts contain various chemicals/compounds that may inhibit the activity of the biotransformation enzyme, as observed in the initial attempts with licorice and Baizhi extracts. Thus, partial purification or suitable fractionation of the extract need to be established before biotransformation. Additional chromatography fractionations might also isolate more biotransformable fractions as trade-off. Third, BGP might need to optimize biotransformation conditions for different herbal extracts. Even for pure compounds, the biotransformation enzyme might have different activity under optimal pH and temperature. For example, DgAS showed optimal conversion yields at pH 6 toward triterpenoids GAA, GAG, and GAF [50] and toward isoflavone puerarin [48]; at pH 7 toward 8-hydroxydaidzein [21]; at pH 8 toward isoquercitrin [40]. Therefore, one needs to carefully optimize reaction parameters, such as enzyme concentration, substrate concentration, pH, temperature, and incubation time, which is crucial for achieving efficient biotransformation. Finally, BGP may isolate complex mixtures of products from crude extracts, but it requires efficient purification and analytical techniques for the isolation and identification of each desired compound.
4. Conclusions
This review highlights the significant potential of PDMA and BGP as powerful and efficient strategies for the modification and discovery of bioactive natural products. PDMA serves as an effective in silico method to predict potential biotransformation reactions and guide the isolation of new derivatives with desired characteristics. The successful application of PDMA in developing promising compounds like 3′-hydroxyloureirin A and B, skullcapflavone II-6′-O-β-glucoside, 3''-hydroxyisoxsuprine, and various methylated products demonstrates its effectiveness in generating derivatives with improved anti-melanoma, anti-inflammatory, and antioxidant activities. In the future, the development of PDMA can be further enhanced by integrating more advanced computational simulation techniques, such as molecular docking and molecular dynamics simulations, to more accurately model precursor-enzyme interactions. Furthermore, integrating more comprehensive chemical and biological databases and developing more intelligent screening algorithms will help improve the prediction accuracy and efficiency of PDMA. With the continuous advancement of biotechnology and computer science, PDMA stands out as a knowledge-based method for the next generation of bioactive molecule discovery, bringing more innovation and breakthroughs to drug development, functional foods, and cosmetics.
Complementarily, BGP may help to explore unknown functional compounds or rare active ingredients retained in crude extracts. BGP allows for the direct isolation of novel compounds resulting from enzymatic transformations of fractionated herbs, which often exhibit enhanced properties such as antioxidant activity, solubility, and bioactivity. BGP has emerged as a cost-efficient and versatile process for the discovery and isolation of novel and/or bioactive molecules from natural raw materials. By seamlessly integrating enzymatic biotransformation with available purification methodologies, BGP helps researchers overcome the disadvantages and limitations of traditional natural product research. For instance, applying BGP to licorice, Ha-Soo-Oh, Ganoderma, and Baizhi has unequivocally demonstrated a cost-effective strategy for yielding novel compounds with enhanced bioactivity and improved physicochemical properties. As the repertoire of characterized biocatalytic enzymes continues to expand, BGP holds immense promise for accelerating the discovery of new drugs and functional ingredients from the vast and largely untapped resources of the natural world, particularly those found in traditional medicines.
Funding
This research was funded by the National Science and Technology Council of Taiwan under grant number MOST 111-2221-E-024 -001-MY3.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The author thank Dr. Tzi-Yuan Wang, Biodiversity Research Center, Academia Sinica, for his kind help in the discussion and correction of the writing in the manuscript draft.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Ekiert, H.M.; Szopa, A. Biological activities of natural products. Molecules 2020, 25, 5769. [Google Scholar] [CrossRef] [PubMed]
- Muffler, K.; Leipold, D.; Scheller, M.-C.; Haas, C.; Steingroewer, J.; Bley, T.; Neuhaus, H.E.; Mirata, M.A.; Schrader, J.; Ulber, R. Biotransformation of triterpenes. Process Biochemistry 2011, 46, 1–15. [Google Scholar] [CrossRef]
- Cao, H.; Chen, X.; Jassbi, A.R.; Xiao, J. Microbial biotransformation of bioactive flavonoids. Biotechnology Advances 2015, 33, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Wozniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Critical Review of Food Scienes and Nutritions 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Sultana, N.; Saify, Z.S. Enzymatic biotransformation of terpenes as bioactive agents. Journal of Enzyme Inhibition and Medicinal Chemistry 2013, 28, 1113–1128. [Google Scholar] [CrossRef]
- Andres, P.; Francisco, R.; Andres, G.-G.; Antonio, M. Microbial transformation of triterpenoids. Mini-Reviews in Organic Chemistry 2009, 6, 307–320. [Google Scholar] [CrossRef]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomedicine and Pharmacotherapy 2021, 140, 111596. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Ding, H.-Y.; Wang, T.-Y.; Cai, C.-Z.; Chang, T.-S. Antioxidant and anti-α-glucosidase activities of biotransformable dragon’s blood via predicted data mining approach. Process Biochemistry 2023, 130, 166–173. [Google Scholar] [CrossRef]
- Wu, C.Y.; Ding, H.Y.; Wang, T.Y.; Liu, C.W.; Wu, J.Y.; Chang, T.S. Development of a new isoxsuprine hydrochloride-based hydroxylated compound with potent antioxidant and anti-inflammatory activities. Journal of Microbiology and Biotechnology 2024, 34, 1–9. [Google Scholar] [CrossRef]
- Chang, T.S.; Wu, J.Y.; Ding, H.Y.; Tayo, L.L.; Suratos, K.S.; Tsai, P.W.; Wang, T.Y.; Fong, Y.N.; Ting, H.J. Predictive production of a new highly soluble glucoside, corylin-7-O-beta-glucoside with potent anti-inflammatory and anti-melanoma activities. Applied Biochemistry and Biotechnology 2024. [Google Scholar] [CrossRef]
- Chang, T.S.; Ding, H.Y.; Wang, T.Y.; Wu, J.Y.; Tsai, P.W.; Suratos, K.S.; Tayo, L.L.; Liu, G.C.; Ting, H.J. In silico-guided synthesis of a new, highly soluble, and anti-melanoma flavone glucoside: Skullcapflavone II-6'-O-beta-glucoside. Biotechnology and Applied Biochemistry 2024. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-S.; Ding, H.-Y.; Wu, J.-Y.; Lee, C.-C.; Yang, Z.; Liu, Y.-C.; Wang, T.-Y. New methyl compounds using the predicted data mining approach (PDMA), coupled with the biotransformation of Streptomyces peucetius O-methyltransferase. Biocatalysis and Biotransformation 2025, 43, 84–96. [Google Scholar] [CrossRef]
- Ding, H.Y.; Wang, T.Y.; Wu, J.Y.; Zhang, Y.R.; Chang, T.S. Novel Ganoderma triterpenoid saponins from the biotransformation-guided purification of a commercial Ganoderma extract. Journal Bioscience Bioengineering 2023, 135, 402–410. [Google Scholar] [CrossRef]
- Chang, T.S.; Ding, H.Y.; Wu, J.Y.; Wang, M.L.; Ting, H.J. Biotransformation-guided purification of a novel glycoside derived from the extracts of Chinese berb Baizhi. Journal Bioscience Bioengineering 2024, 137, 47–53. [Google Scholar] [CrossRef]
- Wu, J.Y.; Ding, H.Y.; Wang, T.Y.; Hsu, M.H.; Chang, T.S. A new stilbene glucoside from biotransformation-guided purification of Chinese herb Ha-Soo-Oh. Plants (Basel) 2022, 11, 2286. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-Y.; Ding, H.-Y.; Wang, T.-Y.; Cai, C.-Z.; Chang, T.-S. Application of biotransformation-guided purification in Chinese medicine: an example to produce butin from licorice. Catalysts 2022, 12, 718. [Google Scholar] [CrossRef]
- Lee, S.H.; Baek, K.; Lee, J.E.; Kim, B.G. Using tyrosinase as a monophenol monooxygenase: A combined strategy for effective inhibition of melanin formation. Biotechnology and Bioengineering 2016, 113, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.M.; Wang, D.S.; Chang, T.S. Improving free radical scavenging activity of soy isoflavone glycosides daidzin and genistin by 3’-hydroxylation using recombinant Escherichia coli. Molecules 2016, 21, 1723. [Google Scholar] [CrossRef]
- Chang, T.S.; Wang, T.Y.; Chiang, C.M.; Lin, Y.J.; Chen, H.L.; Wu, Y.W.; Ting, H.J.; Wu, J.Y. Biotransformation of celastrol to a novel, well-soluble, low-toxic and anti-oxidative celastrol-29-O-beta-glucoside by Bacillus glycosyltransferases. Journal of Bioscience and Bioengineering 2021, 131, 176–182. [Google Scholar] [CrossRef]
- Wu, J.Y.; Wang, T.Y.; Ding, H.Y.; Zhang, Y.R.; Lin, S.Y.; Chang, T.S. Enzymatic synthesis of novel vitexin glucosides. Molecules 2021, 26, 6274. [Google Scholar] [CrossRef]
- Chang, T.S.; Wang, T.Y.; Yang, S.Y.; Kao, Y.H.; Wu, J.Y.; Chiang, C.M. Potential industrial production of a well-soluble, alkaline-stable, and anti-inflammatory isoflavone glucoside from 8-hydroxydaidzein glucosylated by recombinant amylosucrase of Deinococcus geothermalis. Molecules 2019, 24, 2236. [Google Scholar] [CrossRef]
- Chang, T.-S.; Chiang, C.-M.; Wu, J.-Y.; Tsai, Y.-L.; Ting, H.-J. Production of a new triterpenoid disaccharide saponin from sequential glycosylation of ganoderic acid A by 2 Bacillus glycosyltransferases. Bioscience, Biotechnology, and Biochemistry 2021, 85, 687–690. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. International Journal of Pharmacy 2019, 570, 118642. [Google Scholar] [CrossRef]
- Wang, T.H.; Tseng, W.C.; Leu, Y.L.; Chen, C.Y.; Lee, W.C.; Chi, Y.C.; Cheng, S.F.; Lai, C.Y.; Kuo, C.H.; Yang, S.L.; et al. The flavonoid corylin exhibits lifespan extension properties in mouse. Nature Communications 2022, 13, 1238. [Google Scholar] [CrossRef] [PubMed]
- Kopycki, J.G.; Rauh, D.; Chumanevich, A.A.; Neumann, P.; Vogt, T.; Stubbs, M.T. Biochemical and structural analysis of substrate promiscuity in plant Mg2+-dependent O-methyltransferases. Journal of Molecular Biology 2008, 378, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kim, H.; Hur, H.G.; Lim, Y.; Ahn, J.H. Regioselectivity of 7-O-methyltransferase of poplar to flavones. Journal of Biotechnology 2006, 126, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: an overview. Journal of Nutrition Science 2016, 5, e47. [Google Scholar] [CrossRef]
- Solnier, J.; Martin, L.; Bhakta, S.; Bucar, F. Flavonoids as novel efflux pump inhibitors and antimicrobials against both environmental and pathogenic intracellular mycobacterial species. Molecules 2020, 25, 734. [Google Scholar] [CrossRef]
- Chiang, C.M.; Chang, Y.J.; Wu, J.Y.; Chang, T.S. Production and anti-melanoma activity of methoxyisoflavones from the biotransformation of genistein by two recombinant Escherichia coli strains. Molecules 2017, 22, 87. [Google Scholar] [CrossRef]
- Chiang, C.M.; Ding, H.Y.; Tsai, Y.T.; Chang, T.S. Production of two novel methoxy-isoflavones from biotransformation of 8-hydroxydaidzein by recombinant Escherichia coli expressing O-methyltransferase SpOMT2884 from Streptomyces peucetius. Intertnational Journal of Molecular Science 2015, 16, 27816–27823. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, Z.; Zhang, L.; Wang, J.; Wu, C. Glycosyltransferase GT1 family: Phylogenetic distribution, substrates coverage, and representative structural features. Computional and Structrucral Biotechnology Journal 2020, 18, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, W.; Guang, C.; Zhang, W.; Mu, W. Glycosylation of flavonoids by sucrose- and starch-utilizing glycoside hydrolases: A practical approach to enhance glycodiversification. Critical Review on Food Scienes and Nutrition 2023, 63, 1–18. [Google Scholar] [CrossRef]
- Slamova, K.; Kapesova, J.; Valentova, K. "Sweet flavonoids": glycosidase-catalyzed modifications. International Journal of Molecular Sciences 2018, 19, 2126. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Ding, H.-Y.; Wang, T.-Y.; Tsai, Y.-L.; Ting, H.-J.; Chang, T.-S. Improving aqueous solubility of natural antioxidant mangiferin through glycosylation by maltogenic amylase from Parageobacillus galactosidasius DSM 18751. Antioxidants 2021, 10, 1817. [Google Scholar] [CrossRef]
- Mestrom, L.; Przypis, M.; Kowalczykiewicz, D.; Pollender, A.; Kumpf, A.; Marsden, S.R.; Bento, I.; Jarzebski, A.B.; Szymanska, K.; Chrusciel, A.; et al. Leloir glycosyltransferases in applied biocatalysis: a multidisciplinary approach. International Journal of Molecular Science 2019, 20, 5263. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Research 2014, 42, D490–495. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, W.; Zhang, W.; Zhang, T.; Guang, C.; Mu, W. Amylosucrase as a transglucosylation tool: From molecular features to bioengineering applications. Biotechnology Advances 2018, 36, 1540–1552. [Google Scholar] [CrossRef]
- Park, H.-S.; Choi, K.-H.; Park, Y.-D.; Park, C.-S.; Cha, J.-H. Enzymatic Synthesis of Polyphenol Glycosides by Amylosucrase. Journal of Life Science 2011, 21, 1631–1635. [Google Scholar] [CrossRef]
- Moulis, C.; Andre, I.; Remaud-Simeon, M. GH13 amylosucrases and GH70 branching sucrases, atypical enzymes in their respective families. Cellular and Molecular Life Sciences 2016, 73, 2661–2679. [Google Scholar] [CrossRef]
- Rha, C.S.; Kim, H.G.; Baek, N.I.; Kim, D.O.; Park, C.S. Using Amylosucrase for the Controlled Synthesis of Novel Isoquercitrin Glycosides with Different Glycosidic Linkages. Journal of Agricultural and Food Chemistry 2020, 68, 13798–13805. [Google Scholar] [CrossRef]
- Seo, D.H.; Yoo, S.H.; Choi, S.J.; Kim, Y.R.; Park, C.S. Versatile biotechnological applications of amylosucrase, a novel glucosyltransferase. Food Science and Biotechnology 2020, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.H.; Jung, J.H.; Ha, S.J.; Cho, H.K.; Jung, D.H.; Kim, T.J.; Baek, N.I.; Yoo, S.H.; Park, C.S. High-yield enzymatic bioconversion of hydroquinone to alpha-arbutin, a powerful skin lightening agent, by amylosucrase. Applied Microbiology and Biotechnology 2012, 94, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.K.; Kim, H.H.; Seo, D.H.; Jung, J.H.; Park, J.H.; Baek, N.I.; Kim, M.J.; Yoo, S.H.; Cha, J.; Kim, Y.R.; et al. Biosynthesis of (+)-catechin glycosides using recombinant amylosucrase from Deinococcus geothermalis DSM 11300. Enzyme Microbiology and Technology 2011, 49, 246–253. [Google Scholar] [CrossRef]
- Kim, M.D.; Jung, D.H.; Seo, D.H.; Jung, J.H.; Seo, E.J.; Baek, N.I.; Yoo, S.H.; Park, C.S. Acceptor specificity of amylosucrase from Deinococcus radiopugnans and its application for synthesis of rutin derivatives. Journal of Microbiology and Biotechnology 2016, 26, 1845–1854. [Google Scholar] [CrossRef]
- Kim, E.R.; Rha, C.S.; Jung, Y.S.; Choi, J.M.; Kim, G.T.; Jung, D.H.; Kim, T.J.; Seo, D.H.; Kim, D.O.; Park, C.S. Enzymatic modification of daidzin using heterologously expressed amylosucrase in Bacillus subtilis. Food Science and Biotechnology 2019, 28, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Rha, C.S.; Kim, E.R.; Kim, Y.J.; Jung, Y.S.; Kim, D.O.; Park, C.S. Simple and efficient production of highly soluble daidzin glycosides by amylosucrase from Deinococcus geothermalis. Journal of Agricultural and Food Chemistry 2019, 67, 12824–12832. [Google Scholar] [CrossRef]
- Jung, Y.S.; Kim, Y.J.; Kim, A.T.; Jang, D.; Kim, M.S.; Seo, D.H.; Nam, T.G.; Rha, C.S.; Park, C.S.; Kim, D.O. Enrichment of polyglucosylated isoflavones from soybean isoflavone aglycones using optimized amylosucrase transglycosylation. Molecules 2020, 25, 181. [Google Scholar] [CrossRef]
- Ding, H.-Y.; Wang, T.-Y.; Wu, J.-Y.; Tsai, Y.-L.; Chang, T.-S. Enzymatic synthesis of novel and highly soluble puerarin glucoside by Deinococcus geothermalis amylosucrase. Molecules 2022, 27, 4074. [Google Scholar] [CrossRef]
- Chiang, C.-M.; Wang, T.-Y.; Wu, J.-Y.; Zhang, Y.-R.; Lin, S.-Y.; Chang, T.-S. Production of new isoflavone diglucosides from glycosylation of 8-hydroxydaidzein by Deinococcus geothermalis amylosucrase. Fermentation 2021, 7, 232. [Google Scholar] [CrossRef]
- Wu, J.Y.; Ding, H.Y.; Luo, S.Y.; Wang, T.Y.; Tsai, Y.L.; Chang, T.S. Novel glycosylation by amylosucrase to produce glycoside anomers. Biology (Basel) 2022, 11, 822. [Google Scholar] [CrossRef]
- Wahab, S.; Annadurai, S.; Abullais, S.S.; Das, G.; Ahmad, W.; Ahmad, M.F.; Kandasamy, G.; Vasudevan, R.; Ali, M.S.; Amir, M. Glycyrrhiza glabra (Licorice): A comprehensive review on its phytochemistry, biological activities, clinical evidence and toxicology. Plants (Basel) 2021, 10, 2751. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Two novel functional approaches, PDMA and BGP, to find new and/or bioactive compounds from either commercial chemicals (PDMA) or herbal extracts (BGP).
Figure 1.
Two novel functional approaches, PDMA and BGP, to find new and/or bioactive compounds from either commercial chemicals (PDMA) or herbal extracts (BGP).
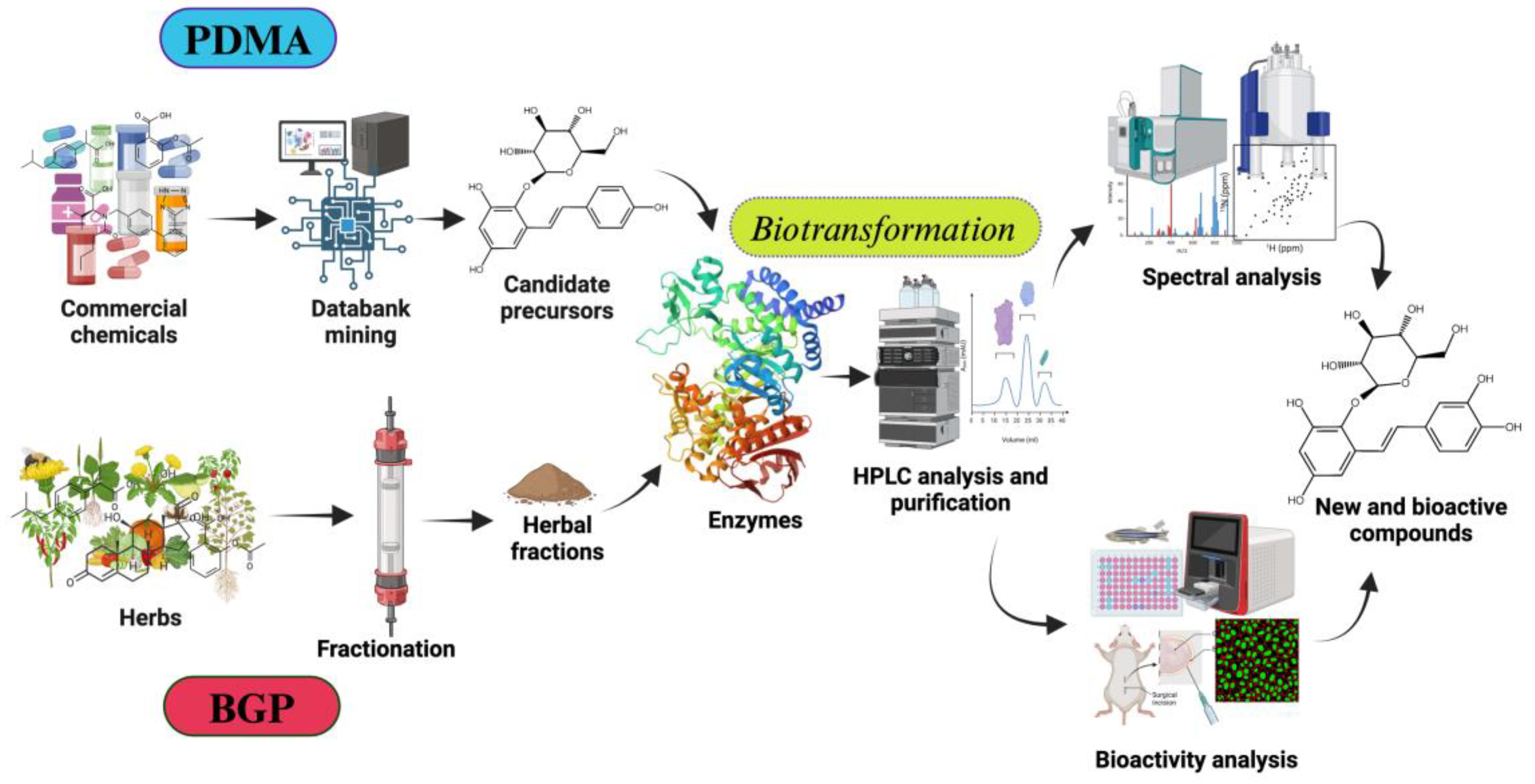
Figure 2.
Flowchart of the predicted data mining approach (PDMA).
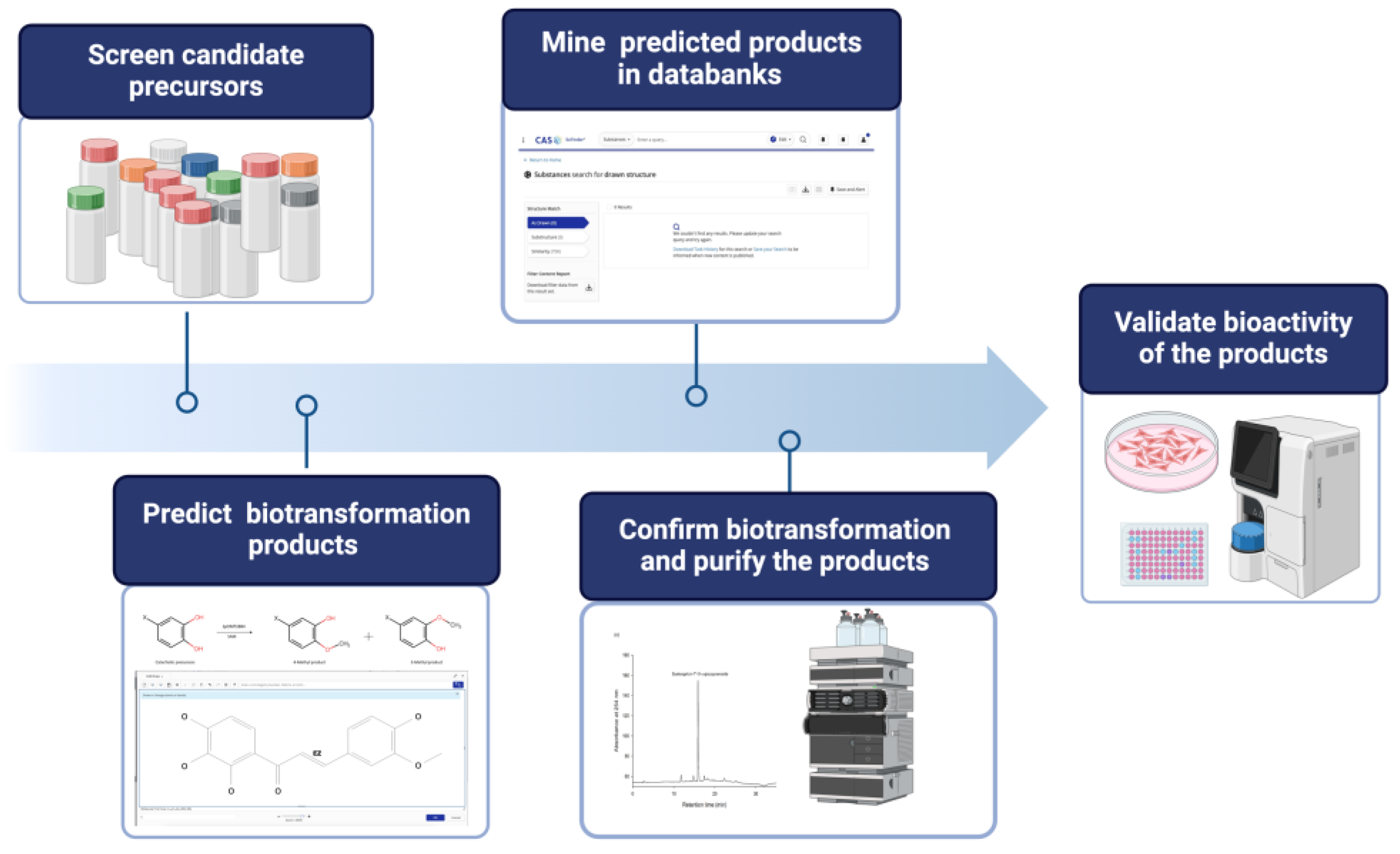
Figure 3.
Enzymatic synthesis of new catecholic compounds from loureirin A/B or isoxsuprine by BmTYR through PDMA [8,9].
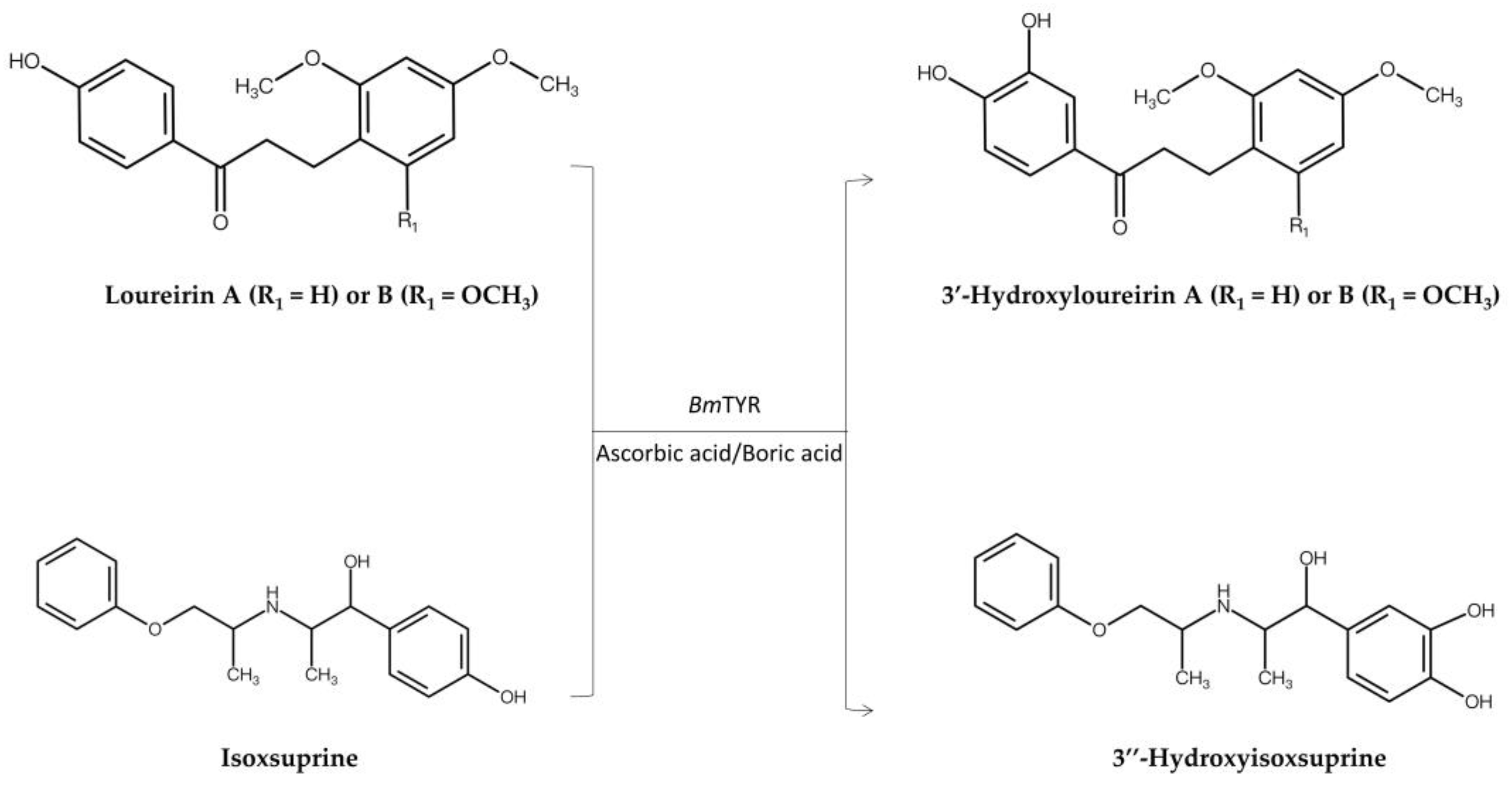
Figure 4.
Enzymatic synthesis of new glucosides from corylin or skullcapflavone II by BsUGT489 through PDMA. UDP-G: Uridine diphosphate glucose. UDP: Uridine diphosphate.
Figure 4.
Enzymatic synthesis of new glucosides from corylin or skullcapflavone II by BsUGT489 through PDMA. UDP-G: Uridine diphosphate glucose. UDP: Uridine diphosphate.
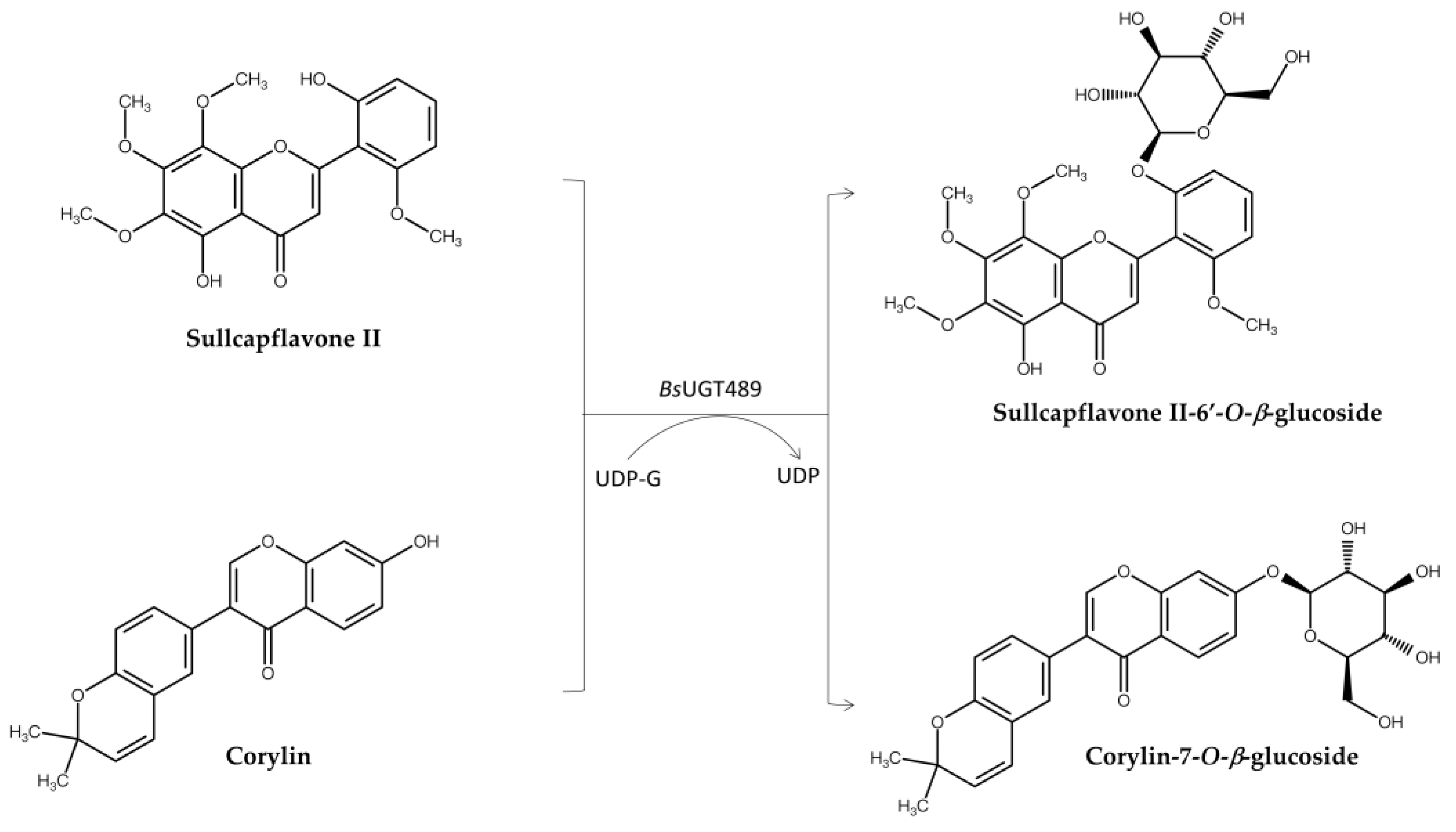
Figure 5.
Enzymatic synthesis of new methyl compounds from plantagoside by SpOMt2884 through PDMA. SAM: S-Adenosylmethionine. SAH: S-Adenosylhomocysteine.
Figure 5.
Enzymatic synthesis of new methyl compounds from plantagoside by SpOMt2884 through PDMA. SAM: S-Adenosylmethionine. SAH: S-Adenosylhomocysteine.
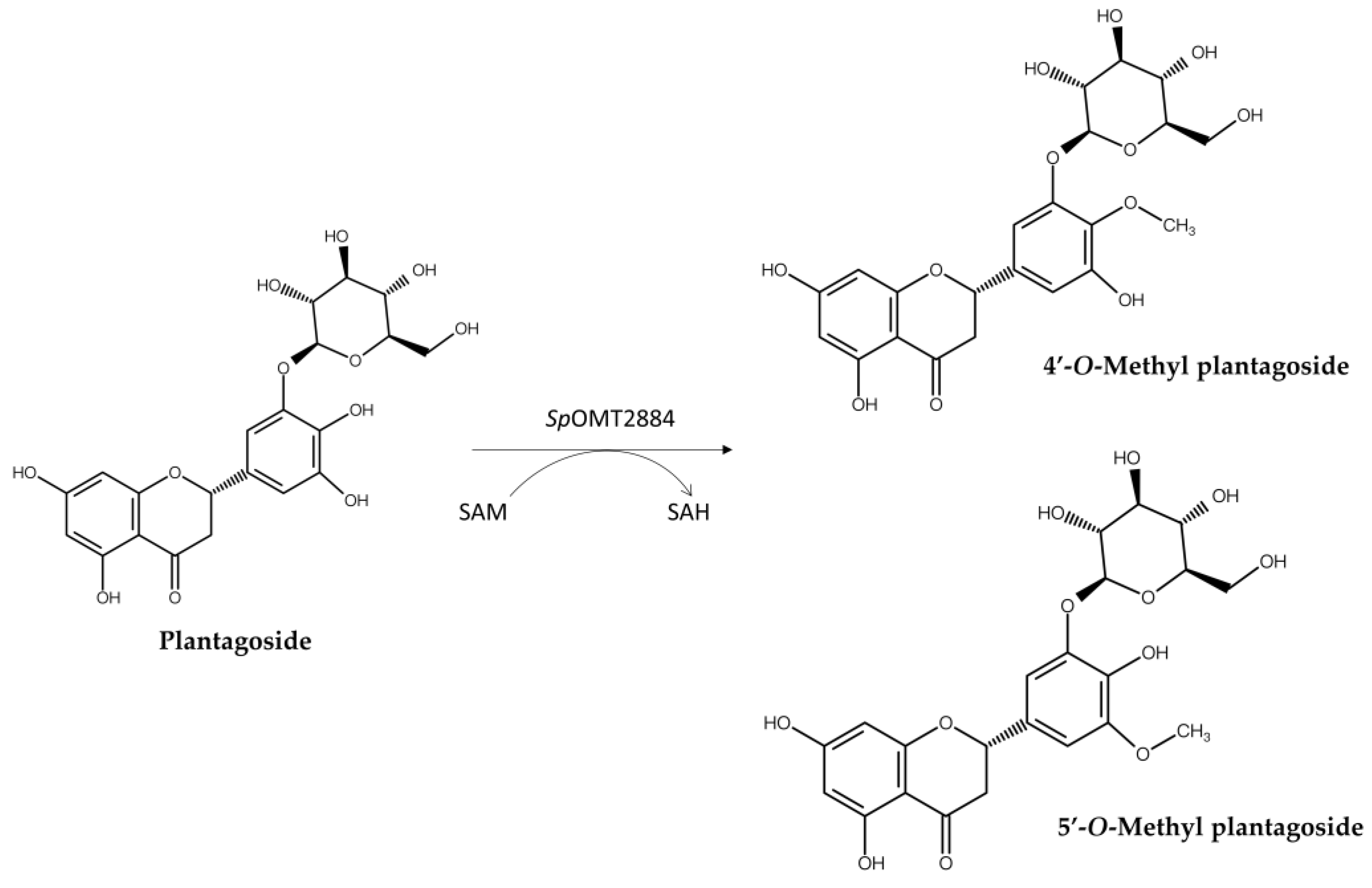
Figure 6.
Concepts between biotransformation-guided purification (BGP) (a) and chemical purification process (b).
Figure 6.
Concepts between biotransformation-guided purification (BGP) (a) and chemical purification process (b).
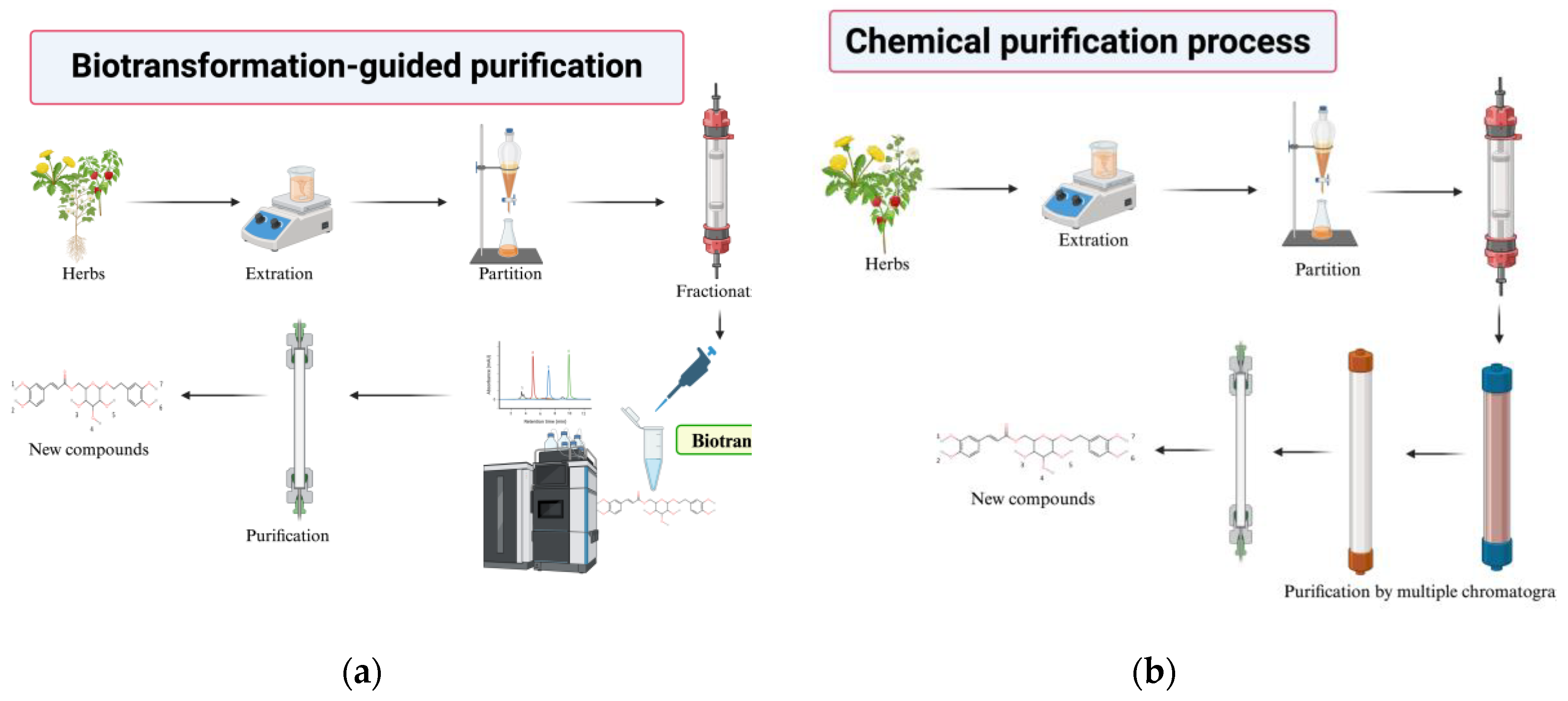
Figure 7.
Enzymatic synthesis of a new glucoside from Ganoderma lucidum by BsUGT489 through BGP [13].
Figure 7.
Enzymatic synthesis of a new glucoside from Ganoderma lucidum by BsUGT489 through BGP [13].
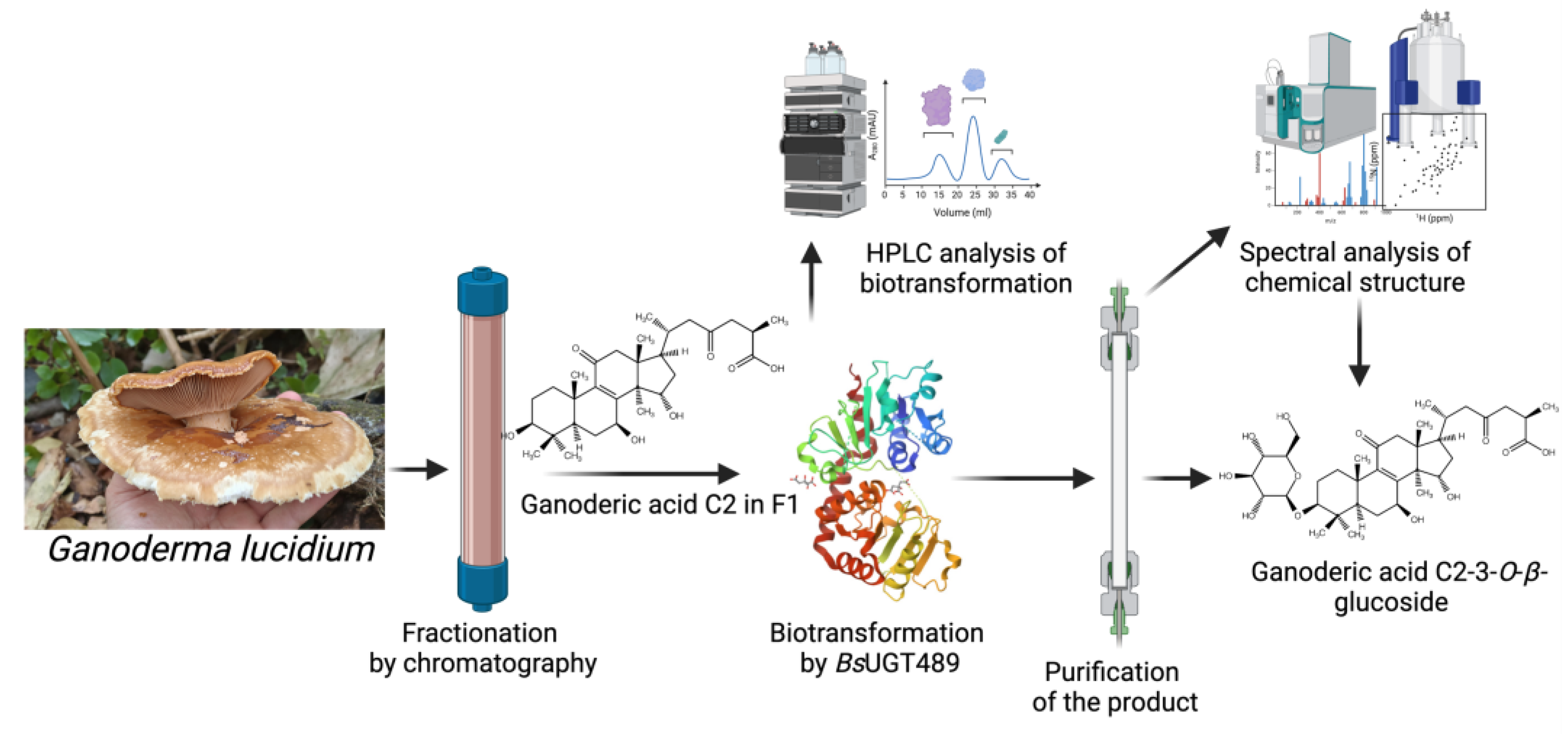
Figure 8.
Enzymatic synthesis of a new glucoside from Baizhi herb by DgAS through BGP.
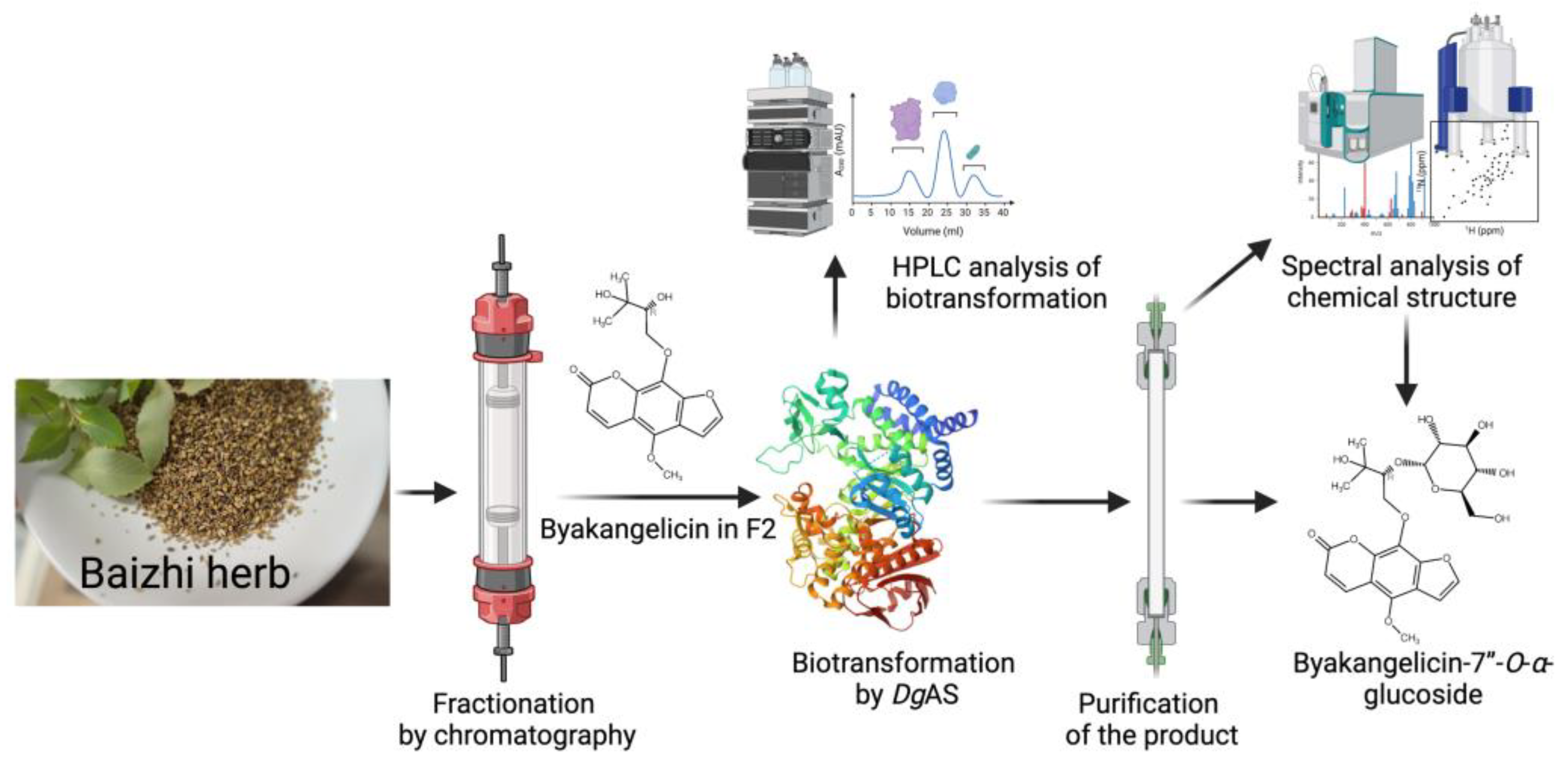
Figure 9.
Enzymatic synthesis of new catecholic compounds from licorice or Ha-Soo-Oh by BmTYR through BGP [15,16].
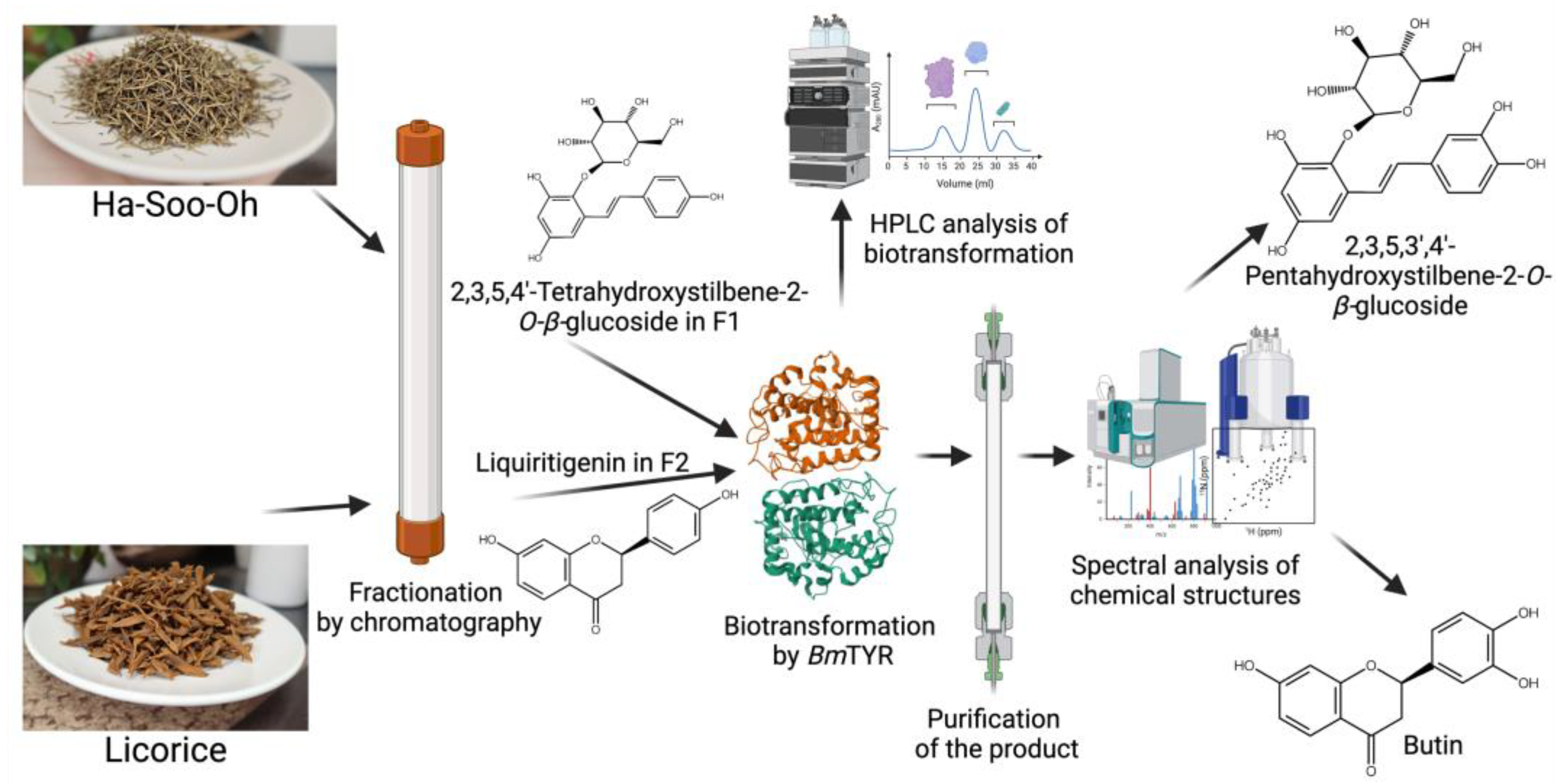

Table 1.
Enzymatic synthesis of new compounds by either PDMA or BGP approach.
| Strategy | Enzyme | Precursor | Novel Product | Property of the New Compounds | Illustration | Reference |
|---|---|---|---|---|---|---|
| Predicted data mining approach (PDMA) | BmTYR1,2 | Loureirin A Loureirin B |
3’-Hydroxyloureirin A 3’-Hydroxyloureirin B |
Improve both antioxidant and anti-α-glucosidase activity | Figure 3 | [8] |
| BmTYR | Isoxsuprine | 3’’-Hydroxyisoxsuprine | Improve both antioxidant and anti-inflammatory activity | Figure 3 | [9] | |
| BsUGT4891,3 | Corylin | Corylin-7-O-β-glucoside | Improve both anti-inflammatory and anti-melanoma activity | Figure 4 | [10] | |
| BsUGT489 | Skullcapflavone II | Sullcapflavone II-6’-O-β-glucoside | Improve both solubility and anti-melanoma activity | Figure 4 | [11] | |
| SpOMT28841,4 | Plantagoside | 4′-O-Methyl plantagoside 5′-O-Methyl plantagoside |
No mention | Figure 5 | [12] | |
| Biotransformation-guided purification (BGP) | BsUGT489 | Ganoderma extract | Ganoderic acid C2-3-O-β-glucoside | Improve solubility and maintain anti-α-glucosidase activity | Figure 7 | [13] |
| DgAS1,5 | Baizhi herb | Byakangelicin-7’’-O-α-glucoside | Improve solubility | Figure 8 | [14] | |
| BmTYR | Ha-Soo-Oh herb | 2,3,5,3′,4′-Pentahydroxystilbene-2-O-β-glucoside | Improve antioxidant activity | Figure 9 | [15] | |
| BmTYR | Licorice herb | Butin | Improve antioxidant activity | Figure 9 | [16] |
1 Recombinant enzyme isolated from Escherichia coli. 2 Genetic source: Bacillus megaterium. 3 Genetic source: Bacillus subtills. 4 Genetic Source: Streptomyces peucetius. 5 Genetic source: Deinococcus geothermalis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



