
Submitted:
03 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
Despite the known impacts of weaning on animal health, the underlying molecular mechanisms remain unclear, particularly how psychological and nutritional stress differentially affect gut health and immune function over time. This study hypothesized that early weaning exerts distinct short- and long-term effects on lamb stress physiology, immunity, and gut health, mediated by specific molecular pathways. Methods: Twelve pairs of full-sibling male Hu sheep lambs were assigned to control (CON) or early-weaned (EW) groups. Plasma stress/immune markers were dynamically monitored, and intestinal morphology, antioxidant capacity, apoptosis, and transcriptomic profiles were analyzed at 5- and 28-days post-weaning. Results: Early weaning triggered transient psychological stress, elevating hypothalamic-pituitary-adrenal (HPA) axis hormones (cortisol, catecholamines) and inflammatory cytokines (TNF-α) within 1 day (P < 0.05); however, stress responses were transient and recovered by day 7. Sustained intestinal remodeling was observed in EW lambs, featuring reduced ileal villus height, increased crypt depth (P < 0.05), and oxidative damage (MDA levels doubled vs. CON; P < 0.01). Compensatory epithelial adaptation included increased crypt depth but paradoxically reduced villus tip apoptosis (decreased TUNEL+ cells), suggesting stress-driven suppression of programmed cell death to preserve epithelial integrity. Transcriptome analysis revealed significant changes in gene expression related to immune function, fat digestion, and metabolism. Key DEGs included APOA4, linked to lipid transport adaptation; NOS2, associated with nitric oxide-mediated immune-metabolic crosstalk; and mitochondrial gene COX1, reflecting energy metabolism dysregulation. Protein-protein interaction analysis revealed NOS2 as a hub gene interacting with IDO1 and CXCL11, connecting oxidative stress to immune cell recruitment. Conclusion: These findings underscore the intricate interplay between metabolic stress, immune function, and gut health during early weaning. Early weaning exerts minimal lasting psychological stress but drives persistent gut dysfunction through nutritional-metabolic reprogramming. This study elucidates the molecular mechanisms driving these changes. Future studies should address strategies to mitigate oxidative stress and optimize lipid utilization during the weaning transition.
Keywords:
early weaning
; gut health
; transcriptomics
; oxidative stress
; immune
Introduction
Weaning is a critical period in the growth of lambs, marking the transition from milk to solid feed. This transition involves substantial nutritional and psychological changes that can impose significant stress on the animal, potentially affecting gut health, metabolism, and immune function [1,2]. While the physiological, emotional, and behavioral impacts of weaning on young ruminants have been widely studied[3,4,5], the exact mechanisms—especially how psychological and nutritional stress influence gut health—remain poorly understood. The relative contributions of psychological versus nutritional stress in affecting gut health, as well as their short- and long-term impacts, have not been fully characterized. Additionally, the rapid turnover of gut tissues complicates our understanding of both immediate and lasting effects on gut morphology and function.
Previous studies demonstrated that early weaning induces both psychological and nutritional stress in lambs[6,7]. Psychological stress primarily arises from separation from the mother[8], activating the hypothalamic-pituitary-adrenal (HPA) axis and increasing levels of stress hormones such as cortisol (CORT) and catecholamines[9]. This activation triggers an emergency response in the body, altering heart rate, blood pressure, body temperature, muscle tension, and metabolic rate[10]. While these responses are adaptive in the short term, they may have negative consequences, particularly for young animals with developing immune systems[2]. By contrast, nutritional stress results from the abrupt shift in diet, with structural carbohydrates replacing lactose and milk fats as the primary energy source. Our previous research showed that early weaning at 21 days significantly affects nutrient intake and digestion efficiency in lambs, particularly reducing fat digestion[7]. This dietary change demands significant adaptation by the lamb’s digestive system, and alterations in nutrient intake can disrupt metabolism and other physiological processes.
Gut health is particularly sensitive to stress. During stress, the gut mucosa undergoes significant remodeling, characterized by dysregulated epithelial cell proliferation, differentiation, and apoptosis[11]. These alterations disrupt the delicate balance of intestinal homeostasis, often manifesting as villus atrophy, crypt hyperplasia, and impaired barrier function. Specifically, stress-induced hyperproliferation of crypt base cells, coupled with accelerated differentiation and reduced villus cell survival, leads to structural and functional compromise of the intestinal epithelium[12]. Disruption of gut immunity, a critical component of the overall immune system, can increase susceptibility to infections. Post-weaning diarrhea (PWD), a common issue in young animals[13], typically occurs within 3 to 10 days after weaning, and is a leading cause of morbidity and mortality[14]. PWD is also associated with reduced weight gain and long-term production performance[15]. Furthermore, weaning-induced oxidative stress, lipid peroxidation, and inflammation also exacerbate gut damage[16,17].
Despite the known impacts of weaning on animal health, the underlying molecular mechanisms remain unclear. Specifically, how psychological and nutritional stress differentially affect gut health and immune function in the short and long term is not well understood. The present study hypothesized that early weaning has distinct short-term and long-term effects on lamb stress, immunity, and gut health, mediated by specific molecular mechanisms. Early weaning is expected to affect gut nutrient digestion and absorption, particularly the absorption of fats, by modulating genes involved in these processes. These molecular changes are anticipated to influence key metabolic and immune regulatory pathways, leading to both immediate and prolonged consequences for lamb gut health, metabolism, and immune function.
By investigating the effects of early weaning on stress hormones, immune indicators, and gut health, this study aimed to identify the key genes and molecular pathways involved. Transcriptome analysis was performed to probe the molecular mechanisms linking metabolic stress and immune dysregulation to gut dysfunction. The findings enhance our understanding of the short- and long-term impacts of early weaning on lamb health, and highlight potential strategies for mitigating the adverse effects of weaning on gut function and immunity.
Materials and Methods
Animal Care
Animal procedures employed in this study were reviewed and approved by the Gansu Agricultural University’s Academic Committee and the National Natural Science Foundation of China (Approval No. 31660670). Informed consent was obtained from all animal owners prior to the commencement of the study.
Experimental Design and Animal Handling
To minimize the impact of genetic background differences on the experimental results, 12 pairs of full-sibling neonatal male Hu lambs were selected from a commercial sheep farm. Within each pair, one lamb was assigned to the control group (CON, birth weight = 3.35 ± 0.76 kg, n = 12) and the other to the early weaning group (EW, birth weight = 3.26 ± 0.62 kg, n = 12). From birth to 6 days of age, lambs were housed indoors with their ewes to ensure sufficient colostrum intake. At 7 days of age, all lambs were separated from their dams and fed exclusively with a milk replacer (2% of average body weight per day) containing 23.22% crude protein (CP) and 13.20% lipid. At 21 days of age, all lambs in the EW group were abruptly weaned, while those in the CON group continued to receive artificial feed until 49 days of age. Throughout the study, all lambs had ad libitum access to water and were provided a pelletized starter diet from 7 days of age (Supplementary Material Table S1). Diets were formulated to meet the requirements of the “Feeding standard of meat-producing sheep and goats (NY/T 816-2004)” published in China.
Sample Collection
Blood samples were collected from all lambs via jugular venipuncture in the morning before feeding at 0 days (21 days of age), 1 day (22 days of age), 2 days (23 days of age), 3 days (24 days of age), 7 days (28 days of age), 14 days (35 days of age), 21 days (42 days of age), and 28 days (49 days of age) post-weaning. Each blood sample was collected into a 5 mL lithium heparin (LH) anticoagulant tube and a 2 mL K2EDTA anticoagulant tube. After collection, blood in the LH tube was centrifuged at 3000g for 15 min to harvest plasma, which was stored at -20°C for measurement of stress-related hormones, HPT, and TNF-α. Blood in the K2EDTA tube was analyzed immediately for hematological parameters.
At 26 days of age (5 days post-weaning) and 49 days of age (28 days post-weaning), six lambs from each group were randomly selected and slaughtered via jugular vein exsanguination. The lambs were restrained to minimize stress prior to slaughter, and exsanguination was performed swiftly to ensure minimal suffering. The lambs were not anaesthetized during the procedure, as the method of exsanguination was deemed humane and in accordance with the guidelines for the ethical treatment of experimental animals approved by the Gansu Agricultural University’s Academic Committee. Immediately post-slaughter, samples were collected from the middle section of the duodenum, the anterior section of the jejunum (specifically from a segment located 0.5 to 1.0 m posterior to the end of the duodenum), the middle section of the ileum, and the middle section of the colon (at the center of the colonic loop). Before collecting samples, they were thoroughly rinsed with 1× phosphate-buffered saline (PBS). Samples were fixed in 4% paraformaldehyde for histological examination. Additionally, samples from the middle section of the ileum were snap-frozen in liquid nitrogen and stored at -80°C for total RNA extraction.
Measurement of Hematological Parameters
A PROKAN PE6800 Blood Analyzer (PROKAN Electronics Inc., Shenzhen, China) was used to estimate the number of total WBCs, NEUs, LYMs, RBCs, and the concentration of Hb in K2EDTA anticoagulant blood samples.
Measurement of Plasma Stress-Related Hormones, Haptoglobin, and TNF-α
The concentrations of plasma CORT, NE, HPT, and TNF-α were measured using appropriate enzyme-linked immunosorbent assay (ELISA) kits (Abcam, Cambridge, UK). Determinations were performed using a microplate reader (Thermo Fisher Scientific, Vantaa, Finland).
Measurement of Intestinal Morphology
Duodenum, jejunum, ileum, and colon samples were fixed with 4% paraformaldehyde, embedded in paraffin, sectioned (5μm), and stained with hematoxylin and eosin (HE). The slices were observed using an optical microscope (BA210 Digital, Motic China Group Co. Ltd., Xiamen, China), and 12 complete intestinal villi were randomly selected from each slice. The height, width, and depth of the fossa and muscularis of intestinal villi were measured using an image analysis system (Motic Image Plus 2.0, Motic China Group Co. Ltd., Xiamen, China).
Measurement of Apoptosis in Ileal Cells
TUNEL staining was performed using a TUNEL kit (Roche, Basel, Switzerland) according to the manufacturer's instructions to assess apoptosis in intestinal tissue samples. Briefly, ileum tissue samples fixed with 4% paraformaldehyde were embedded in paraffin and sliced (5μm). The TUNEL reaction mixture was then prepared, applied to tissue sections, and incubated at 37°C in a humidified chamber. The stained sections were observed under a microscope (BA210 Digital, Motic China Group Co. Ltd., Xiamen, China) to detect apoptotic cells.
Measurement of Antioxidant and Immune Indices in Ileum
The collected intestinal tissues were ground in liquid nitrogen and suspended in pre-cooled 1× PBS. After full homogenization, intestinal tissues were centrifuged at 3000g for 10 min at 4℃ to obtain supernatants and prepare 10% tissue extracts. The activities of total superoxide dismutase (SOD) and GSH-PX, and the concentrations of MDA and IgA were measured using appropriate kits (Nanjing Jiancheng Institute of Biological Engineering, Nanning, China). Determinations were performed using a Thermo Scientific™ Varioskan™ LUX multimode microplate reader (Thermo Fisher Scientific, Vantaa, Finland).
mRNA Library Construction and Sequencing
Total RNA was extracted from ileum samples using an RNA extraction kit (Takara, Kusatsu, Japan). RNA integrity was analyzed by 1% agarose gel electrophoresis, and RNA purity and concentration were determined using a Nanodrop2000 instrument (Thermo Fisher Scientific, Wilmington, United States). The quality and purity of total RNA were further assessed using a Bioanalyzer 2100 instrument and a LabChip kit (Agilent, Santa Clara, USA), ensuring that all samples had RNA integrity number (RIN) values >7.0. Approximately 10 μg of total RNA was used to isolate poly(A) mRNA with poly-T oligo attached magnetic beads (Invitrogen, Carlsbad, USA). Following purification, mRNAs were fragmented into small pieces using divalent cations at an elevated temperature. These fragments were reverse-transcribed, and the final cDNA library was obtained using an mRNA Seq sample preparation kit (Illumina, San Diego, CA, USA) following the manufacturer's instructions. The average insert size for paired-end libraries was 300 ± 50 bp. Subsequently, paired-end sequencing was performed on an Illumina HiSeq 4000 platform (Illumina) according to manufacturer's instructions.
Raw data generated by sequencing were filtered using Cutadapt to exclude unqualified sequences. Adapter reads, reads with undetermined base information within the total number of raw reads exceeding 5%, and low-quality reads (base number of mass value Q ≤10 accounting for >20% of the whole read) were removed. Clean reads were then obtained by verifying sequence quality using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/), including Q20, Q30, and GC content. All downstream analyses were performed using high-quality clean data. Raw sequence data have been submitted to the NCBI Short Read Archive under accession code SRP567385.
Reads were aligned against the UCSC sheep reference genome (http://genome.ucsc.edu/) using the HISAT package, which initially removes a portion of reads based on their quality, then maps them to the reference genome. HISAT builds a database of potential splice junctions and confirms them by comparing previously unmapped reads against the database of putative junctions. Mapped reads for each sample were assembled using StringTie[18], and all sample transcriptomes were then merged to reconstruct a comprehensive transcriptome using Perl scripts. StringTie was employed to assess gene expression levels by calculating fragments per kilobase per million mapped fragments (FPKM) [18]. DEGs were identified using an R package based on |log2 FC| >1 and P <0.05. Functional categories of DEGs were established using Kyoto Encyclopedia of Genes and Genomes (KEGG) database. KEGG enrichment analyses were performed using KOBAS (http://bioinfo.org/kobas/). The PPI network of DEGs at 49 days of age was constructed using the STRING database (version 12.0) with a confidence score threshold of >0.15.
Statistical Analysis
Data were analyzed using SPSS version 22.0 (IBM Corp., Armonk, NY, USA). Two-way analysis of variance (ANOVA) within the general linear model (GLM) framework was conducted to evaluate the main effects of weaning strategy and age, as well as their interaction, on hematological parameters, plasma stress-related hormones, and intestinal morphology. Additionally, t-tests were performed to determine the significance of differences between EW and CON groups for each variable at the same age. Each dependent variable was tested for normality and homogeneity of variance prior to analysis. Statistical significance was set at P <0.05.
Results
Hematological Responses
Interactions between weaning and age had no significant effect on total white blood cell (WBC) count, neutrophil (NEU) count, lymphocyte (LYM) count, red blood cell (RBC) count, and hemoglobin (Hb) concentration (P >0.05; Table 1). Weaning had a significant effect on the number of WBC and LYM in lamb blood (P <0.05), and age had a significant effect on the number of LYM, RBC, and Hb concentration (P <0.05). The LYM and RBC count, and Hb concentration of the control group increased with age, and were highest on 28 days after weaning (P <0.05). However, NEU/LYM decreased with age, and were significantly lower at 2 days after weaning than after 28 days (P <0.05). WBC count in the weaning group was highest on day 1 and day 3 after weaning, significantly higher than on day 0 (P <0.05). LYM count was highest on 28 days after weaning (P <0.05). NEU were highest on day 1 after weaning, significantly higher than on day 0 (P <0.05). NEU/LYM were highest on day 1 after weaning, significantly higher than on day 0 and day 28 (P <0.05). The number of WBC, LYM, and NEU in the weaning group was significantly higher than in the control group at 1 day and 3 days after weaning, 3 days after weaning, and 1 day after weaning, respectively (P <0.05), while there were no significant differences in hematological indicators at other days of age between weaning and control groups (P >0.05).
Plasma Stress-Related Hormones, Haptoglobin, and TNF-α
The effects of weaning and interactions between weaning and age on plasma cortisol (CORT), haptoglobin (HPT), norepinephrine (NE) and tumor necrosis factor-alpha (TNF-α) concentrations in lambs were not significant (P >0.05; Table 2). Age had a significant effect on plasma NE concentration (P <0.05). There were no significant differences between days of age for concentrations of CORT, HPT, NE, and TNF-α in the control group (P >0.05), but the concentration of NE in the weaning group was higher at 2 days after weaning than at 0 and 7 days after weaning (P <0.05). TNF-α was significantly higher on day 1 after weaning than on day 0 in the weaning group (P <0.05). CORT, HPT, and TNF-α levels in the weaning group were significantly higher than in the control group on day 1 after weaning (P <0.05), and NE levels were significantly higher than in the control group on day 2 after weaning (P <0.05). There were no significant differences in other days of age between the two groups (P >0.05).
Intestinal Morphology
Interactions between weaning and age had no significant effect on the morphology of the duodenum, jejunum, ileum, and colon of lambs (P >0.05; Table 3). Weaning significantly reduced the height of ileum villi (P <0.05), and increased the depth of jejunum and ileum crypts (P <0.05). The height of villi in the weaning group on days 26 and 49 was significantly lower than in the control group (P <0.05). The depth of jejunum and ileum crypts in the weaning group was significantly higher than in the control group at 26 days (P <0.05), but there was no significant differences between the two groups at 49 days (P >0.05). Weaning had no significant effect on the morphology of duodenum and colon (P >0.05).
Apoptosis of Ileal Cells
As shown in Figure 1, terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining revealed that apoptotic cells in ileum tissues were predominantly localized to the epithelial cells at the villus tips, characterized by distinct brown nuclear staining. Weaning reduced the proportion of TUNEL staining positive intestinal villi and the proportion of apoptotic cells in TUNEL staining positive intestinal villi epithelial cells.
Antioxidant and Immune Indices in Ileum
Interactions between weaning and age had no significant effect on the activities of superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px), and levels of malondialdehyde (MDA) and immunoglobulin A (IgA), in ileum of lambs (P >0.05; Table 4). Age had no significant effect on SOD and GSH-Px activities, and IgA concentration (P >0.05), while MDA concentration increased significantly with increasing days of age (P <0.05). Weaning had no significant effect on the activities of SOD and GSH-Px (P >0.05), but significantly reduced the concentration of IgA (P <0.05) and increased the concentration of MDA (P <0.1). IgA concentration in the weaning group was significantly lower than in the control group at 26 and 49 days (P <0.05), while MDA concentration was significantly higher at 49 days (P <0.05), and there was no significant difference between the two groups at 26 days (P >0.05).
RNA Sequencing (RNA-seq) Data Mapping and Annotation
A total of 24 cDNA libraries were sequenced from the ileum tissues of all experimental lambs (n = 6 per group). After removing adaptors and filtering, we obtained 1187.7 M valid reads. After mapping clean reads to the ovine genome, 87.70–89.47% of reads were successfully aligned, and 55.30–66.92% of reads had unique genomic locations. Moreover, 80.31–83.44% of reads were paired end-mapped reads (Supplementary Table S2).
Differentially Expressed Genes (DEGs)
In the RNA-seq analysis, 18,716 genes were detected in the jejunum of all 24 individuals. Comparing the gene expression profiles of lambs unweaned (CON49) and weaned at 21 days (EW49) detected 233 differentially expressed genes (DEGs; Figure 2). Among these, 83 were downregulated and 150 upregulated. Comparing the EW26 and CON26 groups detected only 29 DEGs, of which 17 were upregulated and 12 were downregulated. Comparing EW49 and EW26 groups detected 690 DEGs, among which 567 were upregulated and 123 were downregulated. Comparing CON49 and CON26 groups detected 753 DEGs, among which 618 were upregulated and 135 were downregulated.
KEGG Pathway Analysis of DEGs
To explore the biological functions of the identified DEGs, KEGG enrichment analysis was performed (Figure 3). At 49 days of age there were 21 significantly enriched pathways (P <0.05) in the EW group compared with the CON group. The most notable pathways were primarily associated with immune-related processes, including viral protein interactions with cytokine and cytokine receptors, chemokine signaling pathways, and cytokine-cytokine receptor interactions. Additionally, pathways related to lipid absorption and metabolism, such as cholesterol metabolism and fat digestion and absorption, were significantly enriched. By contrast, at 26 days of age, only a single pathway, biosynthesis of amino acids, was significantly enriched (P <0.05) in the EW group compared to the CON group.
Protein-Protein Interaction (PPI) Network of DEGs
The PPI network of DEGs at 49 days of age was constructed using the STRING database (version 12.0) with a confidence score threshold of >0.15. The resulting network includes 160 nodes and 109 edges, with an average node degree of 1.36 (Figure 4).
Further integration of PPI network analysis with KEGG pathway enrichment analysis revealed significant interactions among the most significantly enriched pathways, particularly those related to immune processes, metabolic pathways, and lipid absorption and metabolism. Notably, the top 20 most significant DEGs were highly concentrated within the PPI network, with prominent associations with KEGG pathways related to metabolic processes and lipid metabolism, including cholesterol and lipid absorption and metabolism. Key genes including cytochrome c oxidase subunit 1 (COX1), nitric oxide synthase 2 (NOS2), indoleamine 2,3-dioxygenase 1 (IDO1), and C-X-C motif chemokine ligand 11 (CXCL11) were identified as pivotal nodes, linking these enriched pathways, and highlighting their central roles in immune-related processes and metabolic regulation (Figure 4).
Discussion
Our previous research showed that early-weaned lambs (weaned at 21 days) had lower growth performance by 49 days, with body weights of 9.74 kg versus 12.88 kg and average daily gains of 146.79 g/d versus 258.57 g/d in unweaned lambs[7]. Despite increased solid feed intake, early weaning reduced nutrient utilization, evidenced by lower crude protein and fat digestibility. Notably, daily fat digestion per kg BW was significantly lower in early-weaned lambs (0.40 g/kg BW) compared to unweaned lambs (2.65 g/kg BW), reflecting a 6.6-fold reduction in fat digestion capacity, highlighting the profound metabolic challenges imposed by the abrupt dietary transition[7]. Weaning stress impacts lambs in two major ways: psychologically and nutritionally[8]. Regarding psychological impacts, separation from ewes and the weaning process exert substantial influences on lambs[6]. The HPA axis is the primary regulatory system responding to stress[19]. When an individual experiences stress or threat, the HPA axis is activated, triggering a series of endocrine responses. Activation of the HPA axis puts the body into a state of heightened alert, causing significant changes in heart rate, blood pressure, body temperature, muscle tension, and metabolic levels[20]. Studies have shown that weaning stress significantly increases serum CORT levels in calves and lambs[2,9,21]. Additionally, the immature adaptive immune system of young animals relies heavily on the HPA axis as the main regulator of the immune system, providing homeostatic feedback through glucocorticoids, and influencing immune responses[22]. In weaned calves treated with lipopolysaccharide, there are significant changes in blood leukocyte distribution, cytokines, and acute phase proteins[23,24], with neutrophils serving as biomarkers for weaning stress[25,26]. Since lymphocytes are closely related to adaptive immune function[27], our results suggest that immune function, especially adaptive immunity, is still developing up to 49 days of age. Although nutrient intake is known to be crucial in regulating immune system function, with both deficiency and excess negatively affecting immunity and pathogen susceptibility[3,28], some studies indicate that nutrition has limited effects on immune responses in weaned lambs[4,29]. Our results showed that acute stress responses in lambs following weaning lasted only 1 to 3 days, with stress biomarkers such as CORT, catecholamines, and TNF-α returning to normal levels after 3 days, suggesting that short-term psychological stress does not have long-lasting effects on blood immune indicators or hormone levels.
Regarding nutritional impacts, our prior research demonstrated that early weaning, while rapidly increases solid feed intake, the abrupt cessation of liquid milk consumption leads to reduced digestibility and total digestion of crude protein and fat, leading to a shift in energy metabolism in lambs[7]. We hypothesize that these changes, combined with the psychological stress of weaning, disrupt the gut’s metabolic functions, ultimately impairing gut health. This is reflected in our intestinal morphology results showing that early weaning led to reduced villus height and increased crypt depth, indicative of gut mucosal damage and impaired epithelial cell renewal. Numerous studies demonstrated that weaning stress leads to intestinal mucosal damage in mammals, manifested as reduced villus height and increased crypt depth[30,31]. The epithelial layer covering the small intestine forms the physical barrier of the gut and is among the most rapidly renewing tissue structures in mammals, with a cell life cycle of only 4−5 days[32]. Rapid and continuous renewal of intestinal epithelial cells depends on Lgr5+-labeled crypt base columnar cells within crypts, which proliferate and differentiate into transit-amplifying cells and migrate upwards[33]. Intestinal stem cells located at the base of crypts constantly proliferate and differentiate, moving out of crypts and towards villi to replace damaged epithelial cells. Deepened crypts indicate frequent proliferation and differentiation of intestinal stem cells, which is also a marker of epithelial cell damage[34]. Research has also shown that weaning stress impairs tight junctions between epithelial cells and increases mucosal permeability, allowing bacteria, toxins, and allergens to more easily cross the gut barrier, leading to inflammation or immune reactions[30,35]. These changes are accompanied by increased epithelial cell apoptosis, observable by TUNEL staining, and the process is potentially driven by metabolic and oxidative stress.
These findings highlight the impact of early weaning on gut health, particularly regarding mucosal integrity and cell renewal. Although psychological stress associated with weaning may not have long-lasting effects on hormones and blood immune markers, direct structural changes in the gut suggest that the early life stage is crucial, during which lambs are more susceptible to gastrointestinal challenges[36], and nutritional changes brought on by early weaning can have significant effects. Further research is needed to explore the mechanisms underlying the impact of weaning on gut health.
In this study, the MDA concentration in ileal tissue of the weaned group at 49 days of age was twice that of the non-weaned group, indicating that weaning caused oxidative damage to ileal cells, which may mediate the impact of weaning on gut health. MDA is a product of lipid peroxidation and is an important marker of oxidative damage in cells. Reactions involving oxygen radicals and lipid peroxidation play a significant role in metabolism[37]. It is generally believed that generation of reactive oxygen species (ROS) and the body’s ability to clear oxygen radicals are coordinated and in dynamic balance. When this balance is disrupted lipid peroxidation occurs, leading to MDA production, which alters the fluidity and permeability of cell membranes, ultimately causing changes in cell structure and function[38,39]. The degree of lipid peroxidation in villus cells is significantly higher than in the crypt region, and the villus region generates numerous free radicals, which may contribute to epithelial cell damage and promote epithelial cell differentiation and migration[40]. Studies have shown that weaning induces oxidative stress in piglets[41], characterized by increased ROS production, decreased antioxidant capacity, reduced villus height, deepened crypts, and decreased digestive enzyme activity[42]. Early weaning may inhibit the p65 and Nrf2 signaling pathways, affecting the expression of antioxidant genes and antioxidant system development[43].
Oxidative stress caused by weaning may result from significant changes in lamb food structure post-weaning, leading to increased basal metabolic rate in the gut and consequent generation of ROS. Our previous research demonstrated that after weaning, solid feed intake rapidly increases while both crude protein and crude fat digestion decrease, with blood biochemical indicators suggesting reduced nitrogen deposition[7]. Compared with liquid milk, early and heavy intake of relatively indigestible plant-based feed likely increases the intestinal burden, leading to oxidative stress. Our previous findings also suggest that antioxidant capacity and ATPase activity in lamb intestinal tissues are closely related to growth performance and intestinal tissue morphology[44]. Our transcriptome analysis results indicate that among the most significant DEGs, NOS2, OXSR1, HSPA1L, and ND6 are all related to oxidative stress and ROS production, further confirming that weaning leads to oxidative damage in intestinal tissues.
Transcriptome analysis also provided insights into the underlying molecular mechanisms. At 49 days of age (28 days post-weaning), 320 DEGs were identified, compared to just 29 DEGs at 26 days of age (5 days post-weaning). This indicates that the effects of weaning on gene expression are not immediate, but rather evolve over time. KEGG enrichment analysis indicated that at 49 days of age, DEGs between weaned and control groups were mainly enriched in pathways related to fat digestion and absorption, cytokine-cytokine receptor interaction, tight junctions, chemokine signaling pathways, and cholesterol metabolism. Our previous research showed that weaning reduces fat digestibility in lambs[7], and enrichment of DEGs in these intestinal pathways confirms this effect. Additionally, the DEGs suggest that the impact of weaning on fat digestion is associated with the expression of apolipoproteins such as apolipoprotein A-IV (APOA4). Notably, APOA4 emerged as the most significantly upregulated gene, which is involved in lipid absorption, transport, and metabolism[45]. Its upregulation suggests that lambs are compensating for decreased fat intake and digestibility following weaning. Weaning is known to affect the gut fat metabolism pathway and apolipoprotein expression in lambs[31]. Furthermore, the enriched pathways indicate that the effects of weaning on transcriptional regulation in the gut are mainly related to immune function regulation and gut barrier integrity.
The DEGs in immune and gut barrier-related pathways include various interleukins, TNF-α superfamily members, and tight junction proteins. Other studies have shown that weaning upregulates immune function-related pathways in calves, and DEGs may be associated with the recovery of gut mucosal immune responses and reduced mucosal thickness during the weaning transition[46], consistent with our findings. This underscores the significant impact of increased immune responses and gut barrier damage on gut health in lambs post-weaning.
Integrating PPI network and KEGG pathway enrichment results provides insights into the molecular mechanisms underlying the observed biological processes and their connections to phenotypic changes. The most significantly enriched KEGG pathways, including those related to immune processes, metabolic pathways, and lipid absorption and metabolism, were well represented within the PPI network. Interestingly, the top 20 most significant DEGs were clustered tightly in the network, particularly for pathways associated with metabolic processes and lipid metabolism. Among these, APOA4 emerged as the most significantly upregulated gene. As a key player in lipid metabolism[45], its upregulation may represent a compensatory response to the reduced fat intake and digestibility observed after weaning. Other genes involved in oxidative stress and immune regulation, such as NOS2, OXSR1, ND6, and COX1, were enriched in metabolic pathways, indicating altered energy metabolism and mitochondrial function, both critical for maintaining gut health and immune responses.
In particular, NOS2, involved in nitric oxide synthesis[47], interacts with IDO1, a key regulator of tryptophan metabolism, which in turn interacts with CXCL11, forming a cascade of molecular interactions connecting metabolic regulation with immune responses. IDO1, a key regulator of tryptophan metabolism, modulates immune responses by influencing the balance between pro-inflammatory and anti-inflammatory pathways[48]. CXCL11, a chemokine involved in recruiting immune cells[49], implies activation of immune pathways in response to metabolic stress. These interactions, together with the immune-related pathways enriched in the PPI network, highlight the connection between metabolic stress and immune function. Activation of inflammatory pathways, driven by these genes, likely contributes to increased gut permeability, further exacerbating damage to the gut epithelium.
Conclusion
This study examined the effects of early weaning on gut health, metabolism, and immune function in lambs. While short-term psychological stress had minimal long-term effects, nutritional stress led to significant changes in gut morphology, including reduced villus height and increased crypt depth, as well as oxidative stress. Transcriptome analysis revealed that weaning altered gene expression in pathways related to immune function, fat digestion, and metabolism. Genes such as APOA4, NOS2, and COX1 were differentially expressed, indicating both metabolic adjustments and immune dysregulation. NOS2 interacts with IDO1 and CXCL11, linking metabolic changes to immune responses. The findings emphasize the complex relationship between metabolic stress, immune function, and gut health during weaning. Future research should focus on validating the molecular mechanisms identified, such as the role of NOS2, IDO1, and associated pathways in metabolic adjustments and immune responses.
Funding
This work was supported by the Central Government's Guiding Fund for Local Science and Technology Development under grant no. 23ZYQC0304, and the Science and Technology Project of Gansu Province - Special Project for Cooperation between the Eastern and Western Regions under grant no. 22CX8NA044, and Fuxi Young Talents Program of Gansu Agricultural University under grant no. Gaufx-04Y01.
Acknowledgments
We would like to thank the native English-speaking scientists of Elixigen Company (Huntington Beach, California) for editing our manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Schichowski C, Moors E, Gauly M. Influence of weaning age and an experimental Haemonchus contortus infection on behaviour and growth rates of lambs. Appl Anim Behav Sci 2010;125:103-8. [CrossRef]
- Fazio E, Medica P, Cravana C, Ferlazzo A. Short- and Long-term Effects of Weaning on Adrenocortical and Functional Response of Lambs. Acta Sci Vet 2014;42.
- Mialon MM, Boivin X, Durand D, et al. Short- and mid-term effects on performance, health and qualitative behavioural assessment of Romane lambs in different milk feeding conditions. Animal 2021;15:100157. [CrossRef]
- Beard SC, Schmied JD, Hodgins DC, Mallard BA. The effects of timing of high immune response phenotyping in relation to weaning on immune responses of crossbred beef calves. J Anim Sci 2023;101. [CrossRef]
- Lambertz C, Farke-Rover A, Gauly M. Effects of sex and age on behavior and weight gain in beef calves after abrupt weaning. Anim Sci J 2015;86:345-50.
- Napolitano F, Annicchiarico G, Caroprese M, et al. Lambs prevented from suckling their mothers display behavioral, immune and endocrine disturbances. Physiol Behav 2003;78:81-9. [CrossRef]
- Li C, Wang G, Zhang Q, et al. Developmental changes of nutrient digestion in young lambs are influenced by weaning and associated with intestinal microbiota. Anim Biotechnol 2023;34:1362-76. [CrossRef]
- Han C, Li M, Li F, et al. Temporary sensory separation of lamb groups from ewes affects behaviors and serum levels of stress-related indicators of small-tailed Han lambs. Physiol Behav 2024;277:114504. [CrossRef]
- Hickey MC, Drennan M, Earley B. The effect of abrupt weaning of suckler calves on the plasma concentrations of cortisol, catecholamines, leukocytes, acute-phase proteins and in vitro interferon-gamma production. J Anim Sci 2003;81:2847-55. [CrossRef]
- Nicolaides NC, Kyratzi E, Lamprokostopoulou A, Chrousos GP, Charmandari E. Stress, the stress system and the role of glucocorticoids. Neuroimmunomodulat 2015;22:6-19. [CrossRef]
- Gao X, Cao Q, Cheng Y, et al. Chronic stress promotes colitis by disturbing the gut microbiota and triggering immune system response. Proc Natl Acad Sci U S A 2018;115:E2960-e9. [CrossRef]
- He Y, Liu N, Ji Y, Tso P, Wu Z. Weaning Stress in Piglets Alters the Expression of Intestinal Proteins Involved in Fat Absorption. J Nutr 2022;152:2387-95. [CrossRef]
- Wang X, Niu L, Wang Y, et al. Combining 16S rRNA Sequencing and Metabolomics Data to Decipher the Interactions between Gut Microbiota, Host Immunity, and Metabolites in Diarrheic Young Small Ruminants. Int J Mol Sci 2023;24. [CrossRef]
- Zhong T, Wang Y, Wang X, et al. Diarrhea in suckling lambs is associated with changes in gut microbiota, serum immunological and biochemical parameters in an intensive production system. Front Microbiol 2022;13:1020657. [CrossRef]
- Han X, Hu X, Jin W, Liu G. Dietary nutrition, intestinal microbiota dysbiosis and post-weaning diarrhea in piglets. Anim Nutr 2024;17:188-207. [CrossRef]
- Upadhaya SD, Kim IH. The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets-A Review. Animals (Basel) 2021;11. [CrossRef]
- Zhuang Y, Wu H, Wang X, et al. Resveratrol Attenuates Oxidative Stress-Induced Intestinal Barrier Injury through PI3K/Akt-Mediated Nrf2 Signaling Pathway. Oxid Med Cell Longev 2019;2019:7591840. [CrossRef]
- Frazee AC, Pertea G, Jaffe AE, et al. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat Biotechnol 2015;33:243-6. [CrossRef]
- Hulbert LE, Moisa SJ. Stress, immunity, and the management of calves. J Dairy Sci 2016. [CrossRef]
- O'Connor DB, Thayer JF, Vedhara K. Stress and Health: A Review of Psychobiological Processes. Annu Rev Psychol 2021;72:663-88. [CrossRef]
- Ledezma-Torres RA, Sanchez-Davila F, Rodriguez-Miranda DA, et al. Sexual performance and semen quality of pubertal lambs treated with different weaning methods. Arch Anim Breed 2022;65:259-65. [CrossRef]
- Millington, GW. The role of proopiomelanocortin (POMC) neurones in feeding behaviour. Nutr Metab (Lond) 2007;4:18. [CrossRef]
- Lynch EM, Earley B, McGee M, Doyle S. Effect of abrupt weaning at housing on leukocyte distribution, functional activity of neutrophils, and acute phase protein response of beef calves. Bmc Vet Res 2010;6:39. [CrossRef]
- Carroll JA, Arthington JD, Chase CC, Jr. Early weaning alters the acute-phase reaction to an endotoxin challenge in beef calves. J Anim Sci 2009;87:4167-72. [CrossRef]
- O'Loughlin A, McGee M, Doyle S, Earley B. Biomarker responses to weaning stress in beef calves. Res Vet Sci 2014;97:458-63.
- Ceja G, Boerman JP, Neves RC, Jorgensen MW, Johnson JS. l-Glutamine supplementation reduces gastrointestinal permeability and biomarkers of physiological stress in preweaning Holstein heifer calves. J Dairy Sci 2023;106:9663-76. [CrossRef]
- Chi H, Pepper M, Thomas PG. Principles and therapeutic applications of adaptive immunity. Cell 2024;187:2052-78. [CrossRef]
- Boivin X, Nowak R, Garcia AT. The presence of the dam affects the efficiency of gentling and feeding on the early establishment of the stockperson-lamb relationship. Appl Anim Behav Sci 2001;72:89-103.
- McCoard SA, Cristobal-Carballo O, Knol FW, et al. Impact of early weaning on small intestine, metabolic, immune and endocrine system development, growth and body composition in artificially reared lambs. J Anim Sci 2020;98. [CrossRef]
- Hu CH, Xiao K, Luan ZS, Song J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J Anim Sci 2013;91:1094-101.
- Han L, Tao H, Kang L, et al. Transcriptome and iTRAQ-Based Proteome Reveal the Molecular Mechanism of Intestinal Injury Induced by Weaning Ewe's Milk in Lambs. Front Vet Sci 2022;9:809188. [CrossRef]
- Barker N, van de Wetering M, Clevers H. The intestinal stem cell. Genes Dev 2008;22:1856-64. [CrossRef]
- Zhu MH, Sung TS, Kurahashi M, et al. Na+-K+-Cl- cotransporter (NKCC) maintains the chloride gradient to sustain pacemaker activity in interstitial cells of Cajal. Am J Physiol Gastrointest Liver Physiol 2016;311:G1037-G46. [CrossRef]
- Shyer AE, Huycke TR, Lee C, Mahadevan L, Tabin CJ. Bending gradients: how the intestinal stem cell gets its home. Cell 2015;161:569-80. [CrossRef]
- Wood KM, Palmer SI, Steele MA, Metcalf JA, Penner GB. The influence of age and weaning on permeability of the gastrointestinal tract in Holstein bull calves. J Dairy Sci 2015;98:7226-37. [CrossRef]
- Dunière L, Ruiz P, Lebbaoui Y, et al. Effects of rearing mode on gastro-intestinal microbiota and development, immunocompetence, sanitary status and growth performance of lambs from birth to two months of age. Anim Microbiome 2023;5:34. [CrossRef]
- Fu ZL, Yang Y, Ma L, et al. Dynamics of oxidative stress and immune responses in neonatal calves during diarrhea. J Dairy Sci 2024;107:1286-98. [CrossRef]
- Pi J, Zhang Q, Fu J, et al. ROS signaling, oxidative stress and Nrf2 in pancreatic beta-cell function. Toxicol Appl Pharmacol 2010;244:77-83. [CrossRef]
- Reuter S, Gupta SC, Chaturvedi MM, Aggarwal BB. Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med 2010;49:1603-16. [CrossRef]
- Dantzer R, O'Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci 2008;9:46-56. [CrossRef]
- Liu M, Ma J, Xu J, et al. Fecal microbiota transplantation alleviates intestinal inflammatory diarrhea caused by oxidative stress and pyroptosis via reducing gut microbiota-derived lipopolysaccharides. Int J Biol Macromol 2024;261:129696. [CrossRef]
- Zhu LH, Zhao KL, Chen XL, Xu JX. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J Anim Sci 2012;90:2581-9.
- Yin J, Wu MM, Xiao H, et al. Development of an antioxidant system after early weaning in piglets. J Anim Sci 2014;92:612-9. [CrossRef]
- Chen Z, Wang G, Wang W, et al. Relationship between jejunum ATPase activity and antioxidant function on the growth performance, feed conversion efficiency, and jejunum microbiota in Hu sheep (Ovis aries). Bmc Vet Res 2024;20:242. [CrossRef]
- Wang F, Kohan AB, Lo C-M, et al. Apolipoprotein A-IV: a protein intimately involved in metabolism. J Lipid Res 2015;56:1403-18. [CrossRef]
- Nishihara K, van Niekerk J, He Z, et al. Reduction in mucosa thickness is associated with changes in immune function in the colon mucosa during the weaning transition in Holstein bull dairy calves. Genomics 2023;115:110680. [CrossRef]
- Sorokin, A. Nitric Oxide Synthase and Cyclooxygenase Pathways: A Complex Interplay in Cellular Signaling. Curr Med Chem 2016;23:2559-78. [CrossRef]
- Romani L, Fallarino F, De Luca A, et al. Defective tryptophan catabolism underlies inflammation in mouse chronic granulomatous disease. Nature 2008;451:211-5. [CrossRef]
- Tokunaga R, Zhang W, Naseem M, et al. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation - A target for novel cancer therapy. Cancer Treat Rev 2018;63:40-7. [CrossRef]
Figure 1.
Effects of weaning on apoptotic cell distribution in ileal samples revealed by TUNEL staining. Nuclei of apoptotic cells are brown and those of non-apoptotic cells are blue. (A) Control group sampled at day 49. (B) Weaning group sampled at day 49. (C) Control group sampled at day 26. (D) Weaning group sampled at day 26.
Figure 1.
Effects of weaning on apoptotic cell distribution in ileal samples revealed by TUNEL staining. Nuclei of apoptotic cells are brown and those of non-apoptotic cells are blue. (A) Control group sampled at day 49. (B) Weaning group sampled at day 49. (C) Control group sampled at day 26. (D) Weaning group sampled at day 26.
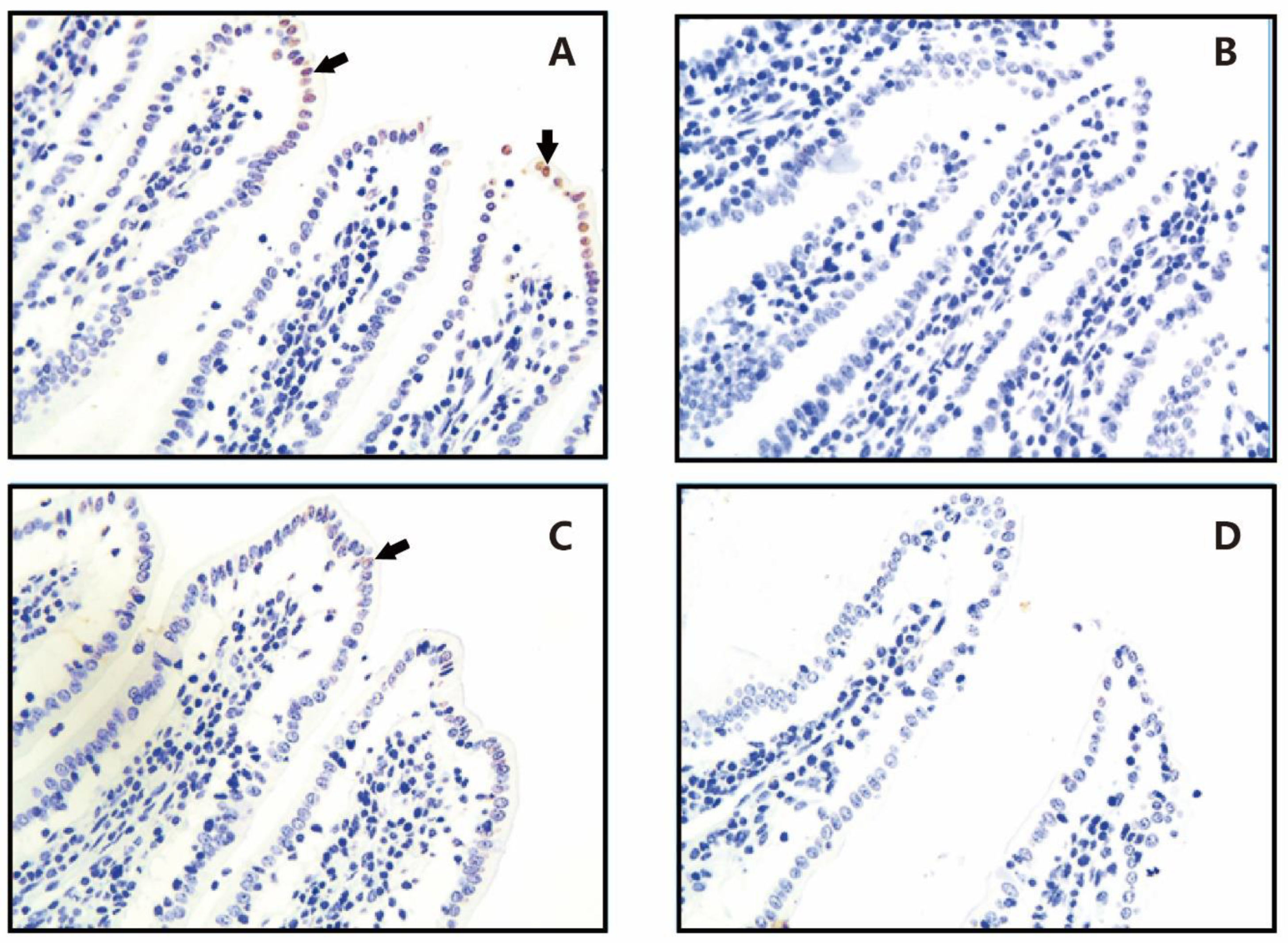
Figure 2.
Number of differentially expressed genes (DEGs). CON26, control group sampled at day 26; EW26, weaning group sampled at day 26; CON49, control group sampled at day 49; EW49, weaning group sampled at day 49.
Figure 2.
Number of differentially expressed genes (DEGs). CON26, control group sampled at day 26; EW26, weaning group sampled at day 26; CON49, control group sampled at day 49; EW49, weaning group sampled at day 49.
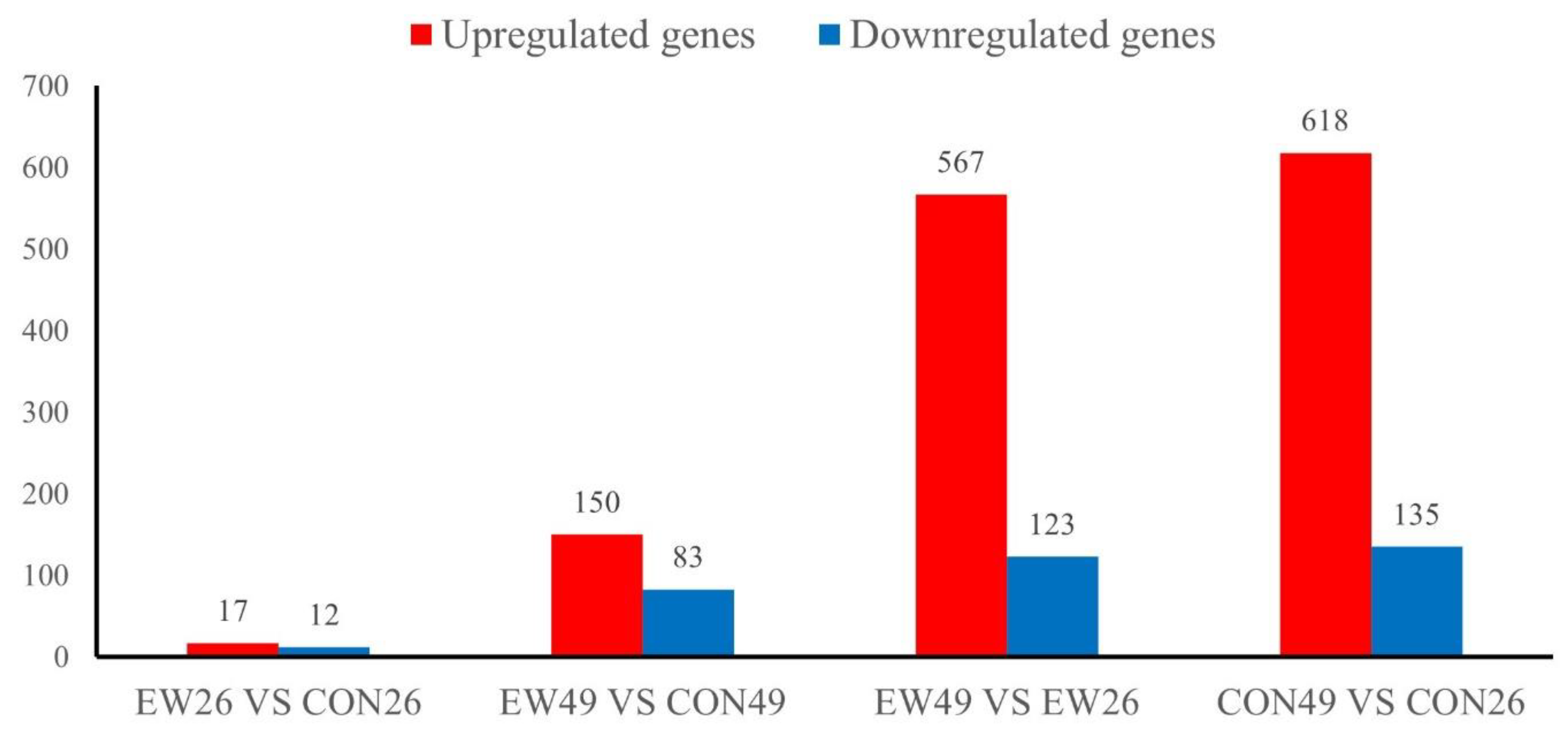
Figure 3.
Results of KEGG enrichment analysis. (A) EW49 vs. CON49. (B) EW26 vs. CON26. The x-axis represents enrichment scores and the y-axis represents pathway terms. Circle color indicates false discovery rate (FDR) based on P-value, and circle size indicates the number of DEGs. CON49, control group sampled at day 49; EW49, weaning group sampled at day 49; CON26, control group sampled at day 26; EW26, weaning group sampled at day 26.
Figure 3.
Results of KEGG enrichment analysis. (A) EW49 vs. CON49. (B) EW26 vs. CON26. The x-axis represents enrichment scores and the y-axis represents pathway terms. Circle color indicates false discovery rate (FDR) based on P-value, and circle size indicates the number of DEGs. CON49, control group sampled at day 49; EW49, weaning group sampled at day 49; CON26, control group sampled at day 26; EW26, weaning group sampled at day 26.
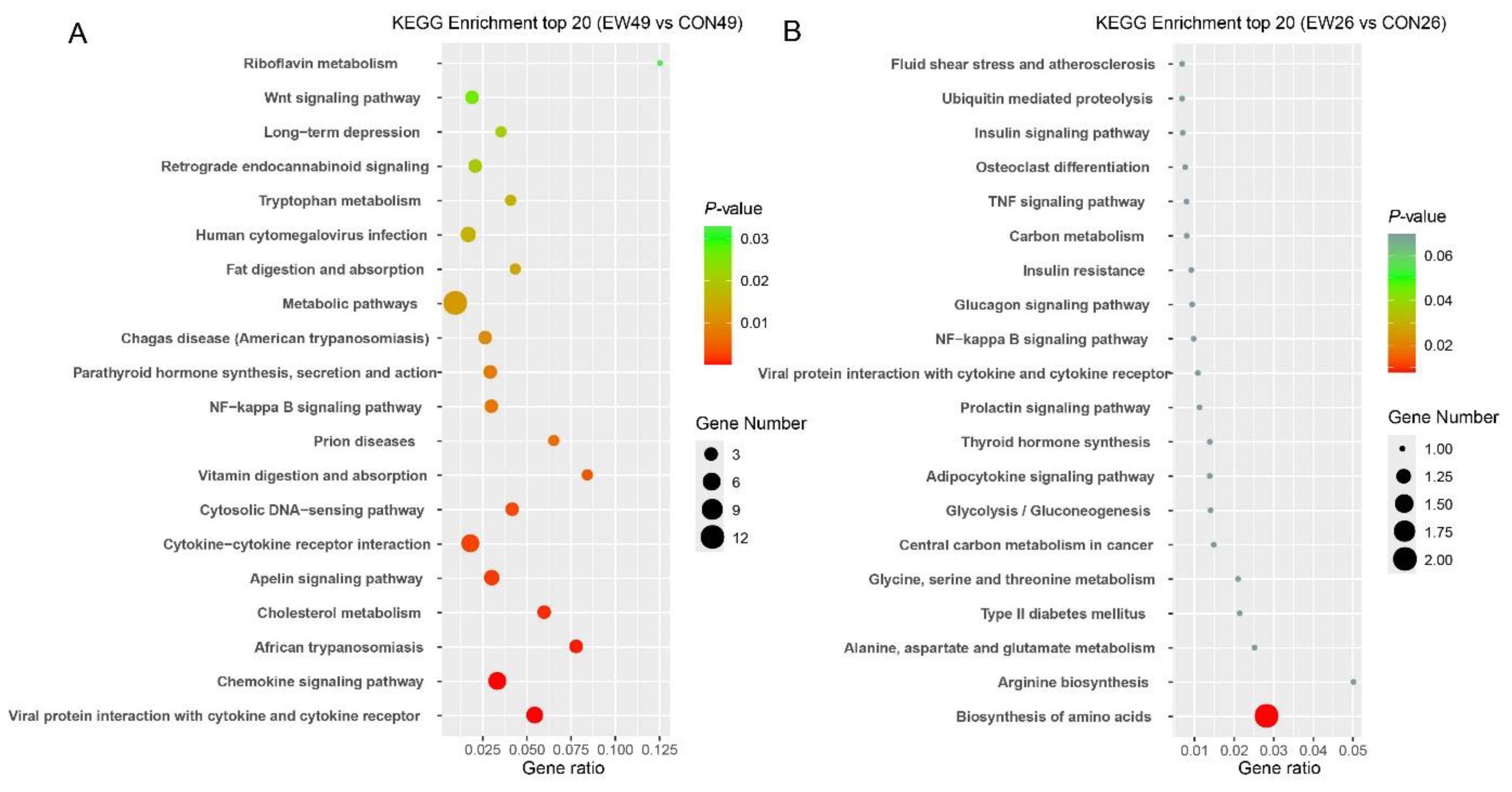
Figure 4.
Protein-protein interaction network of DEGs at 49 days of age.
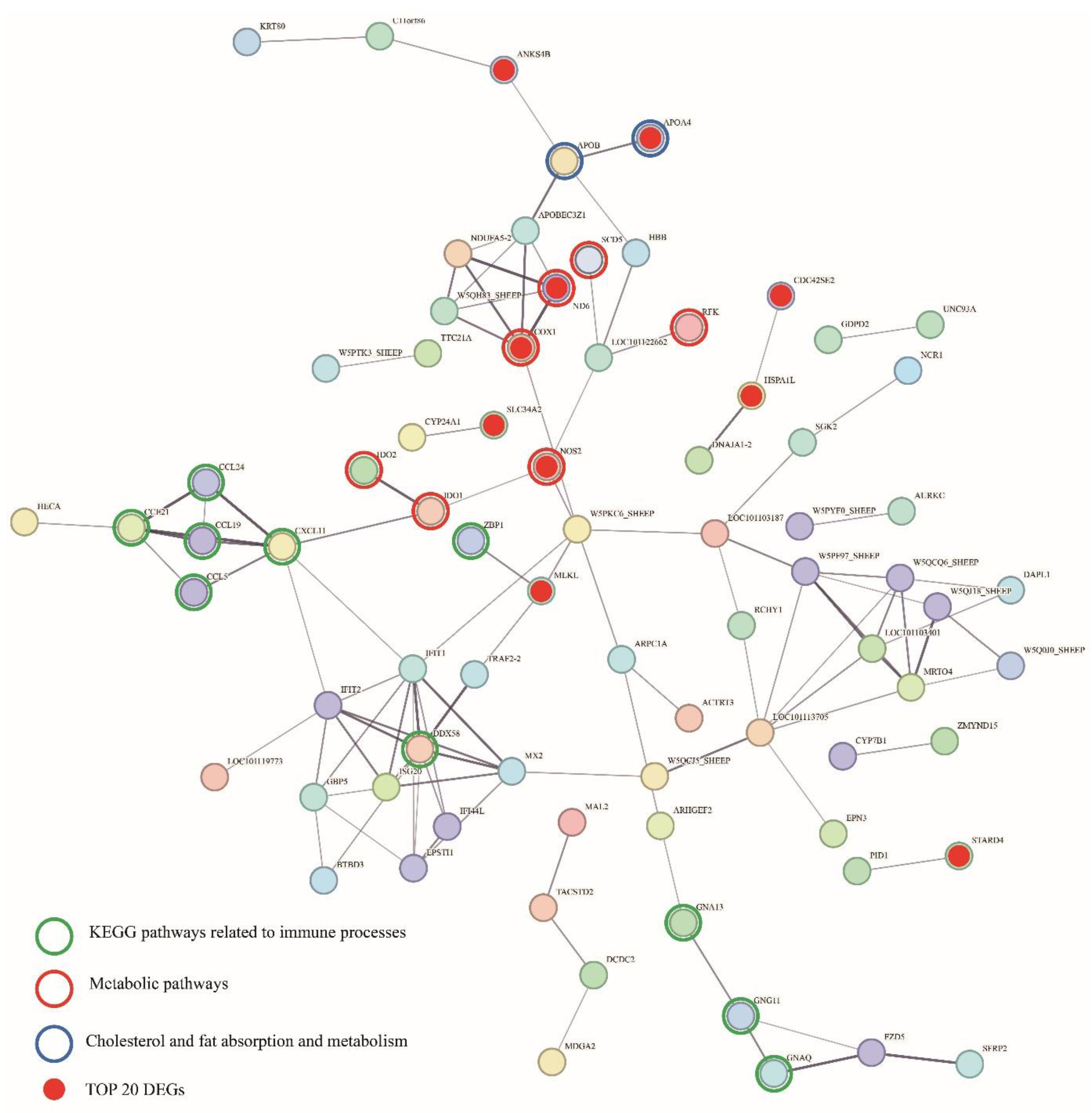

Table 1.
Effect of weaning and age on hematological responses.
| Items | Treatment | Days post-weaning (d) | SEM | P value | ||||||||
| 0 | 1 | 2 | 3 | 7 | 14 | 28 | Weaning | Age | Weaning × Age | |||
| WBC (×109cells/L) | CON | 8.29 | 8.59 | 9.03 | 8.31 | 9.20 | 8.28 | 9.86 | 0.223 | 0.024 | 0.328 | 0.464 |
| EW | 7.95b | 10.92a* | 10.15ab | 10.66a* | 9.79ab | 9.53ab | 9.68ab | |||||
| LYM(×109cells/L) | CON | 3.52b | 3.64b | 3.62b | 3.32b | 4.01b | 4.32b | 5.47a | 0.108 | 0.041 | 0.004 | 0.528 |
| EW | 3.47b | 4.45ab | 4.37ab | 4.46ab* | 4.53ab | 4.69ab | 5.06a | |||||
| NEU(×109cells/L) | CON | 3.49 | 3.71 | 4.06 | 3.69 | 3.68 | 2.75 | 3.30 | 0.150 | 0.145 | 0.097 | 0.642 |
| EW | 3.24b | 5.18a* | 4.41ab | 4.69ab | 3.75ab | 3.35b | 3.14b | |||||
| NEU/LYM | CON | 1.02ab | 1.04ab | 1.21a | 1.12ab | 1.03ab | 0.66ab | 0.61b | 0.043 | 0.945 | 0.006 | 0.858 |
| EW | 1.00ab | 1.31a | 1.10ab | 1.11ab | 0.88ab | 0.74ab | 0.61 b | |||||
| RBC(×109cells/L) | CON | 8.47b | 8.00b | 8.02b | 7.91b | 7.94b | 8.37b | 9.22a | 0.068 | 0.432 | 0.004 | 0.449 |
| EW | 8.04b | 8.49ab | 8.34ab | 8.23ab | 8.25ab | 8.25ab | 9.08a | |||||
| Hb (g/L) | CON | 122.64ab | 113.86ab | 111.83ab | 109.79b | 111.94ab | 116.67ab | 127.43a | 1.066 | 0.586 | 0.024 | 0.293 |
| EW | 113.08 | 120.45 | 115.50 | 113.29 | 109.93 | 112.25 | 121.50 | |||||
Note: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). WBC: white blood cells, LYM: lymphocytes, NEU: neutrophils, RBC: red blood cells, Hb: hemoglobin. * p < 0.05 indicates a significant difference between groups at the same age.. a-c Different lowercase letters within a row indicate significant differences (P < 0.05).
Table 2.
Effect of weaning on plasma stress-related hormones, haptoglobin and TNFα.
| Items | Treatment | Days post-weaning (d) | SEM | P value | ||||||
| 0 | 1 | 2 | 3 | 7 | Weaning | Age | Weaning×Age | |||
| CORT (µg/mL) | CON | 116.43 | 114.62 | 115.92 | 125.68 | 115.82 | 1.297 | 0.225 | 0.383 | 0.232 |
| EW | 118.22 | 129.29* | 124.04 | 122.82 | 123.79 | |||||
| HPT (µg/mL) | CON | 51.02 | 50.75 | 52.02 | 49.15 | 44.94 | 0.826 | 0.391 | 0.109 | 0.960 |
| EW | 51.55 | 54.16* | 53.39 | 50.54 | 48.37 | |||||
| NE (µg/mL) | CON | 1468.23 | 1488.63 | 1508.55 | 1544.86 | 1456.08 | 16.074 | 0.378 | 0.049 | 0.320 |
| EW | 1417.75b | 1542.85ab | 1670.66a* | 1555.05ab | 1458.10b | |||||
| TNF-α (µg/mL) | CON | 97.46 | 94.74 | 94.24 | 95.93 | 90.75 | 0.999 | 0.454 | 0.202 | 0.382 |
| EW | 97.37ab | 103.94a* | 93.15b | 95.45ab | 94.93ab | |||||
Note: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). CORT: cortisol, HPT: haptoglobin, NE: norepinephrine, TNF-α: tumor necrosis factor-alpha. * indicates a significant difference between groups at the same age. a-c Different lowercase letters within a row indicate significant differences (P < 0.05).
Table 3.
Effect of weaning on the intestinal morphology of lambs.
| Items | Day 26 | Day 49 | P value | ||||||
| CON | EW | CON | EW | SEM | Weaning | Age | Weaning × Age | ||
| Duodenum | Villi height (μm) | 210.68 | 238.47 | 452.96 | 437.60 | 8.376 | 0.714 | <0.001 | 0.211 |
| Villi width (μm) | 75.88 | 81.21 | 173.27 | 155.40 | 3.717 | 0.818 | <0.001 | 0.115 | |
| Crypt depth (μm) | 120.00 | 149.87* | 220.67 | 212.29 | 5.020 | 0.296 | <0.001 | 0.070 | |
| Muscle layer thickness (μm) | 97.52 | 116.00* | 112.89 | 136.13* | 4.752 | 4.752 | 0.075 | 0.804 | |
| Jejunum | Villi height (μm) | 430.20 | 412.50 | 447.90 | 413.95 | 11.172 | 0.269 | 0.675 | 0.722 |
| Villi width (μm) | 93.08 | 101.92 | 114.62 | 127.15 | 3.437 | 0.144 | 0.005 | 0.793 | |
| Crypt depth (μm) | 126.95 | 195.33* | 159.18 | 181.18 | 7.699 | 0.012 | 0.567 | 0.156 | |
| Muscle layer thickness (μm) | 99.58 | 87.90 | 93.93 | 86.20 | 5.041 | 0.248 | 0.909 | 0.664 | |
| Ileum | Villi height (μm) | 458.10* | 397.97 | 427.54* | 372.47 | 11.047 | 0.028 | 0.151 | 0.737 |
| Villi width (μm) | 103.74 | 108.57 | 114.86 | 114.07 | 4.240 | 0.814 | 0.339 | 0.744 | |
| Crypt depth (μm) | 123.16 | 189.65* | 173.68 | 192.76 | 7.160 | 0.008 | 0.077 | 0.114 | |
| Muscle layer thickness (μm) | 112.20 | 103.55 | 93.86 | 88.27 | 8.142 | 0.306 | 0.116 | 0.926 | |
| Colon | Villi height (μm) | 429.80 | 464.20 | 480.06 | 478.86 | 11.77 | 0.491 | 0.188 | 0.461 |
| Villi width (μm) | 43.80 | 48.94 | 50.86 | 48.38 | 1.654 | 0.694 | 0.342 | 0.267 | |
| Crypt depth (μm) | 63.46 | 71.48 | 74.94 | 77.44 | 7.067 | 0.715 | 0.546 | 0.848 | |
| Muscle layer thickness (μm) | 174.82 | 187.52 | 202.28 | 189.24 | 10.187 | 0.993 | 0.484 | 0.537 | |
Note: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). * indicates a significant difference between groups at the same age.
Table 4.
Effects of weaning on antioxidant and immune Indices in the ileum.
| Items | Day 26 | Day 49 | P value | |||||
| CON | EW | CON | EW | SEM | Weaning | Age | Weaning×Age | |
| SOD (U/mg) | 0.12 | 0.10 | 0.11 | 0.11 | 0.006 | 0.382 | 0.961 | 0.732 |
| GSH-Px (U/mg) | 1.23 | 1.23 | 1.12 | 1.18 | 0.043 | 0.763 | 0364 | 0.716 |
| MDA (nmol/g) | 8.53 | 8.29 | 10.13 | 22.30* | 1.735 | 0.098 | 0.036 | 0.089 |
| IgA (mg/g) | 1.31* | 0.85 | 1.18* | 0.99 | 0.060 | 0.014 | 0.904 | 0.268 |
Note: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). * indicates a significant difference between groups at the same age. SOD: superoxide dismutase; GSH-Px: glutathione peroxidase; MDA: malondialdehyde; IgA: immunoglobulin A. SEM: standard error of the mean.
Table 5.
List of top 20 DEGs with the lowest p-value between EW49 and CON49.
| Gene ID | Gene Symbol | Description | Group | log2FC | P value | Q value | |
| CON | EW | ||||||
| MSTRG.7028 | APOA4 | Apolipoprotein A4 | 6.67 | 37.14 | 2.45 | <0.001 | <0.001 |
| MSTRG.2698 | SLC10A2 | Solute carrier family 10 member 2 | 5.90 | 18.23 | 1.60 | <0.001 | <0.001 |
| MSTRG.12128 | HSPA1L | Heat shock 70 kda protein 1-like | 0.69 | <0.01 | -9.98 | <0.001 | <0.001 |
| MSTRG.5151 | PRNP | Major prion protein | 0.17 | <0.01 | -8.91 | <0.001 | 0.010 |
| MSTRG.9443 | OXSR1 | Serine/threonine-protein kinase OSR1 | <0.01 | 4.77 | 11.29 | <0.001 | 0.015 |
| MSTRG.2150 | PDE9A | Phosphodiesterase 9A | 11.77 | 27.12 | 1.18 | <0.001 | 0.015 |
| MSTRG.2965 | NOS2 | Nitric oxide synthase 2 | 0.74 | 4.74 | 2.64 | <0.001 | 0.019 |
| MSTRG.19179 | CDC42SE2 | CDC42 small effector 2 | 8.52 | 20.11 | 1.21 | <0.001 | 0.019 |
| MSTRG.22642 | COX1 | Cytochrome c oxidase subunit 1 | 1195.86 | 6.55 | -7.53 | <0.001 | 0.020 |
| MSTRG.12142 | C6orf47 | Uncharacterized protein c6orf47 | 1.04 | <0.01 | -10.56 | <0.001 | 0.020 |
| MSTRG.2916 | ZNF830 | Zinc finger protein 830 | <0.01 | 0.64 | 8.68 | <0.001 | 0.032 |
| MSTRG.19825 | STARD4 | StAR related lipid transfer domain containing 4 | 0.81 | 2.71 | 1.71 | <0.001 | 0.032 |
| MSTRG.20048 | SLC34A2 | Solute carrier family 34 member 2 | 14.21 | 33.44 | 1.21 | <0.001 | 0.032 |
| MSTRG.12237 | TRIM15 | Tripartite motif containing 15 | 0.79 | 2.33 | 1.52 | <0.001 | 0.047 |
| MSTRG.22661 | ND6 | NADH-ubiquinone oxidoreductase chain 6 | <0.01 | 4.74 | 8.59 | <0.001 | 0.049 |
| MSTRG.16608 | SOAT2 | Sterol O-acyltransferase 2 | 0.38 | 1.59 | 2.03 | <0.001 | 0.052 |
| MSTRG.12735 | MS4A18 | Membrane spanning 4-domains A18 | 1.69 | 5.27 | 1.61 | <0.001 | 0.069 |
| MSTRG.9443 | AREG | Amphiregulin | 1.21 | 0.85 | -2.09 | 0.000 | 0.070 |
| MSTRG.14252 | ANKS4B | Ankyrin repeat and sterile alpha motif domain containing 4B | 1.04 | 3.51 | 1.72 | 0.001 | 0.154 |
| MSTRG.5689 | MLKL | Mixed lineage kinase domain like pseudokinase | 2.80 | 5.71 | 1.00 | 0.001 | 0.154 |
Notes: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). DEGs: differentially expressed genes, FC: fold change.
Table 6.
List of top 20 DEGs with the lowest p-value between EW26 and CON26.
| Gene ID | Gene Symbol | Description | Group | log2FC | P value | Q value | |
| CON | EW | ||||||
| MSTRG.24683 | ZBTB33 | Zinc finger and BTB domain containing 33 | 3.31 | 1.7 | -2.08 | <0.001 | 0.922 |
| MSTRG.16036 | RAB11FIP1 | RAB11 family interacting protein 1 | 2.99 | 1.17 | -1.66 | <0.001 | 0.922 |
| MSTRG.21722 | AGGF1 | Angiogenic factor with G-patch and FHA domains 1 | 1.57 | 5.03 | 1.58 | 0.001 | 0.922 |
| MSTRG.4224 | GNA13 | G protein subunit alpha 13 | 50.13 | 31.05 | -4.00 | 0.009 | 0.922 |
| MSTRG.6324 | CMTR2 | CAP methyltransferase 2 | 3.73 | 2.67 | -1.53 | 0.010 | 0.922 |
| MSTRG.12043 | NYAP2 | Neuronal tyrosine-phosphorylated phosphoinositide-3-kinase adaptor 2 | 1.1 | 3.04 | 1.31 | 0.014 | 0.922 |
| MSTRG.10771 | CCL19 | C-C motif chemokine ligand 19 | 5.33 | 10.22 | 1.05 | 0.017 | 0.922 |
| MSTRG.19058 | PRR15 | Proline rich 15 | 3.57 | 2.02 | -1.05 | 0.018 | 0.922 |
| MSTRG.16341 | ASS1 | Argininosuccinate synthase 1 | 4.96 | 13.22 | 1.32 | 0.026 | 0.922 |
| MSTRG.6232 | CES2 | Carboxylesterase 2 | 21.13 | 13.63 | -1.02 | 0.027 | 0.922 |
| MSTRG.18314 | CCND2 | Cyclin D2 | 6.25 | <0.001 | -1.99 | 0.029 | 0.922 |
| MSTRG.22347 | DUOXA2 | Dual oxidase maturation factor 2 | 1.19 | 9.26 | 1.77 | 0.033 | 0.922 |
| MSTRG.22348 | DUOX2 | Dual oxidase 2 | 1.7 | 12.07 | 1.63 | 0.038 | 0.922 |
| MSTRG.5089 | PGAM1 | Phosphoglycerate mutase 1 | 17.72 | 34.79 | 2.36 | 0.041 | 0.922 |
| MSTRG.3081 | CCL8 | Phosphoglycerate mutase 1 | 1.79 | 6.12 | 1.09 | 0.043 | 0.922 |
| MSTRG.21722 | CRAMP1 | CAMP-regulated antimicrobial peptide 1 | 2.77 | 2.05 | -1.02 | 0.044 | 0.922 |
| MSTRG.24683 | SOCS3 | Suppressor of cytokine signaling 3 | 2.32 | 7.51 | 1.06 | 0.046 | 0.922 |
| MSTRG.16036 | RBM15 | Suppressor of cytokine signaling 3 | 4.78 | 2.46 | -1.39 | 0.046 | 0.922 |
| MSTRG.4224 | ISG20 | Interferon-stimulated exonuclease gene 20 | 2.47 | 7.96 | 1.14 | 0.049 | 0.922 |
Notes: CON: Non-weaned group. EW: Early-weaned group (weaned on day 21). DEGs: differentially expressed genes, FC: fold change.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



