Submitted:
28 March 2025
Posted:
31 March 2025
You are already at the latest version
Abstract
ABC transporters are a large family of proteins that mediate the export or import a variety of molecules, including capsular polysaccharides. These polysaccharide capsules are an important virulence factor that protect bacteria from host immune system attacks, antibiotics, and physicochemical changes in their environment. In Gram-negative pathogenic bacteria, ABC transporter-dependent systems facilitate the export of capsular polysaccharides. These transport systems are composed of three parts: the ABC transporter and the polysaccharide copolymerase protein in the inner membrane and the outer membrane polysaccharide export protein in the outer membrane. The lipid anchor of the capsular polysaccharide binds to a pocket between the two subunits of the ABC transporter transmembrane domain. The three parts of the ABC transporter-dependent system form a tunnel through which the capsular polysaccharide is exported using energy from ATP hydrolysis. Knowledge of the ABC transporter-dependent system and its function is incomplete, requiring further research to better understand the processes of capsular polysaccharide export. This may also allow, in the future, to develop new molecules that inhibit capsular polysaccharide export, which would help the host immune system fight Gram-negative pathogenic bacteria coated with a polysaccharide capsule. This review presents the latest findings on ABC transporter-dependent systems that export polysaccharide capsules in Gram-negative pathogenic bacteria.
Keywords:
ABC transporter
; capsular polysaccharide
; antibiotic resistance
1. Antibiotic-Resistant Bacteria and Their Virulence Factors
Antibiotic-resistant bacteria are a major public health problem. The World Health Organization (WHO) estimates that in 2019, 6 million people died from infections caused by these bacteria[1]. Since 2017, WHO has published a list of priority bacteria to guide the research for new ways to fight these resistant pathogenic bacteria. The list, updated in 2024, includes eight Gram-negative and four Gram-positive bacteria or families of bacteria that cause epidemics, are difficult to treat and are costly for the global health system[1] (Table 1).
To infect an organism, a bacterium has a toolkit that allows it to enter the host, attach to it, protect itself from its immune system, and spread from one organism to another. These tools are virulence factors that can be transmitted between bacteria in two ways: either by horizontal transfer, a binary division of mother bacteria to daughter bacteria, or by vertical transfer, where genetic material is released into the environment upon bacterial death and taken up by other naturally competent bacteria, such as those of the Neisseria group or Haemophilus influenzae. This genetic material itself encodes the virulence factors that the bacteria will express to their advantage[2,3].
Virulence factors can be toxins produced by bacteria[4], adhesins that allow the attachment of bacteria to host cells[5], locomotion systems such as flagella[6], or proteins that "steal" nutrients from the host[7,8]. Another virulence factor, and the focus of this review, is the capsule composed of capsular polysaccharides [5,7,9,10].

Table 1.
WHO list of bacteria to be studied as a priority to fight their resistance to antibiotics[1].
Table 1.
WHO list of bacteria to be studied as a priority to fight their resistance to antibiotics[1].
| Importance | Bacterium | Gram | Presence of a capsule | Capsular transporter |
| Critical | Acinetobacter baumannii | - | Yes[11] | Wzx/Wzy-dependent system |
| Enterobacteria | - | Specie dependent | Wzx/Wzy-dependent system + ABC transporter-dependent system | |
| Mycobacterium tuberculosis | + | Yes[12] | N/A | |
| High | Salmonella Typhi | - | Yes[13] | ABC transporter-dependent system |
| Shigella spp. | - | Specie dependent | Wzx/Wzy-dependent system | |
| Enterococcus faecium | + | No | N/A | |
| Pseudomonas aeruginosa | - | No | N/A | |
| Non-typhoidal Salmonella | - | Yes[14] | Wzx/Wzy-dependent system | |
| Neisseria gonorrhoea | - | No | N/A | |
| Staphylococcus aureus | + | Yes[15] | N/A | |
| Medium | Group A Streptococci | + | Yes[16] | N/A |
| Streptococcus pneumoniae | + | Yes[17] | N/A | |
| Haemophilus influenzae | - | Yes[9] | ABC transporter-dependent system | |
| Group B Streptococci | + | Yes[18] | N/A |
2. Gram-Negative Bacterial Capsule
A capsule is the external envelope of a bacterium. In 3 % of bacteria, such as Bacillus anthracis, it is protein-based, while in 97 % of bacteria, such as Escherichia coli (E. coli) and Neisseria meningitidis, it is composed of polysaccharides[19] (Figure 1). In Gram-negative bacteria, capsular polysaccharides are anchored to the outer membrane by a lipid anchor.
Capsular polysaccharides are homopolymers or heteropolymers of negatively charged polysaccharides whose export is carried out either by an ABC (ATP Binding Cassette) transporter-dependent system or by a Wzx/Wzy-dependent system[20,21]. The composition and structure of capsular polysaccharides are highly diverse. In E. coli, the capsule, visible by electron microscopy, measures between 100 and 400 nm in thickness, with capsular polysaccharides composed of more than 200 saccharide units[22,23]. In pathogenic strains of Neisseria meningitidis, capsular polysaccharides are composed of 130–200 repeating units of amino sugars, sialic acid, glucose and galactose with an alpha linkage[24,25]. Capsular polysaccharides are exported to the outside of the bacteria to form an additional outer layer of protection for the bacteria. All these capsular polysaccharides are virulence factors that help pathogenic bacteria infect a host and protect themselves from its immune system.
2.1. Gram-Negative Pathogenic Bacteria with a Polysaccharide Capsule As a Virulence Factor
Polysaccharide capsules are found in Gram-negative pathogens such as E. coli or Neisseria meningitidis but also in Gram-positive pathogens such as Staphylococcus aureus or Streptococcus pneumoniae[20,26]. They act as virulence factors for several pathogenic bacteria and allow them to survive in different conditions[21,27]. This is the case for Neisseria meningitidis, Haemophilus influenzae, Salmonella enterica serovar Typhi or Campylobacter jejuni[28,29,30,31,32].
These mentioned bacteria express a capsule that is exported by the ABC transporter-dependent system (Table 1). These pathogenic bacteria are the most extensively studied capsule bacteria, and are responsible for a number of diseases in both humans and livestock. Encapsulated E. coli strains cause blood infections, pyelonephritis or meningitis[33]. Strains of Neisseria meningitidis can cause meningitis or septicemia[24]. Strains of Haemophilus influenzae cause respiratory diseases such as pneumonia, of which the encapsulated form is the most virulent[31]. Salmonella enterica serovar Typhi strains are enterobacteria that cause typhoid fever[13]. Strains of Campylobacter jejuni cause bacterial diarrhea with sequelae such as Guillain-Barré syndrome, inflammatory bowel disease, reactive arthritis or irritable bowel syndrome[34]. Pasteurella multocida strains cause many diseases in animals (birds, dogs, cats, cattle, equines) such as chicken cholera, pasteurellosis, pneumonia, atrophic rhinitis, meningitis and hemorrhagic septicemia. They can be transmitted to humans by bites or scratches[35,36].
2.2. Different Types of Polysaccharide Capsules in Gram-Negative Bacteria
E. coli has over 80 different serogroups[37]. These serogroups have allowed to distinguish four different categories of polysaccharide capsules according to their chemical composition, their biosynthetic systems and their export system (Table 2). These four groups are also present in other Gram-negative bacteria.
Group 1 polysaccharide capsules are composed of acidic polysaccharides containing uronic acids and are exported by a Wzx/Wzy-dependent system[38].
Group 2 polysaccharide capsules are linked to the cell via an α-glycerophosphatidic acid residue and, in some bacteria such as E. coli or Neisseria meningitidis, 3-deoxy-D-manno-oct-2-ulosonic acid (Kdo) provides a linker between the lipid and the reducing end of the polysaccharide[37]. Several group 2 capsular polysaccharides resemble vertebrate glycoconjugates, which prevents the vertebrate immune system from recognizing the capsular polysaccharides of bacteria, and therefore eliminating them[38]. Group 2 polysaccharide capsules are exported by the ABC transporter-dependent system.
Group 3 polysaccharide capsules are similarly linked to the cell via an α-glycerophosphatidic acid residue, with Kdo as a linker, and are also exported by an ABC transporter-dependent system[37]. However, group 3 capsular polysaccharides differ from group 2 in the regulation of their expression and the organization of the genes involved in their biosynthesis and export[38].
Group 4 polysaccharide capsules are O-antigen capsules not bound to the outer membrane. Its polysaccharides contain acetamido sugars and are exported by a Wzx/Wzy-dependent system[38].
2.3. Why Target the Polysaccharide Capsule in the Fight Against Antibiotic-Resistant Gram-Negative Pathogenic Bacteria?
Polysaccharide capsules enable bacteria to survive in different environments through bacterial adhesion, resistance to the immune system, adaptation to different physicochemical conditions, and resistance to antibiotics[21,27].
In the case of bacterial adhesion, polysaccharide capsules can have two roles[26]. The first is to allow the formation of biofilms, because the excessive production of capsular polysaccharides allows bacteria to attach to each other, which initiates the formation of the biofilm[40,41]. The second is to allow bacteria to leave the biofilm, by making a polysaccharide capsule that disrupts contacts between bacteria, in order to colonize other environments[26,42,43]. Both mechanisms and their regulation are not well understood.
In case of resistance to the host immune system, polysaccharide capsules can mimic the antigenic composition of some host cells, hide surface antigens that can be recognized by macrophages or inhibit phagocytosis[24,26,44]. The best-known example is the polysaccharide capsule of Neisseria meningitidis serogroup B, whose capsular polysaccharides are structurally identical to a component of the human NCAM (neuronal cell adhesion molecule), allowing it to evade the host immune system. As a result, the immune response is particularly weak against this type of Neisseria meningitidis, making it difficult to develop vaccines targeting these capsular polysaccharides[2,39].
Polysaccharide capsules can protect bacteria from environmental changes such as lower pH, increased temperatures, lower oxygen concentration, variations in osmotic pressures or desiccation[26,37]. Indeed, capsular polysaccharides form a hydrated layer around the bacteria and maintain it in an aqueous environment[45]. In addition, the capsule coats the bacteria and thus gives it better mechanical resistance[46].
In the case of antibiotic resistance, the negatively charged polysaccharide capsules serve to protect the bacteria from the action of antibiotics by preventing chemical molecules like antimicrobial peptides from coming into contact with the bacterial envelope[47,48].
Since the polysaccharide capsule has specific chemical and structural compositions depending on the bacterial strains, it is also interesting to develop vaccines against these Gram-negative pathogenic bacteria to stimulate the immune system of the hosts[49].
2.4. Gene Organization, Expression, and Regulation of Group 2 and 3 Capsular Polysaccharides in Gram-Negative Bacteria
This section describes how the gene organization of Gram-negative antibiotic-resistant pathogenic bacteria intervenes in the biosynthesis and export of group 2 and 3 capsular polysaccharides.
The gene organization is identical in all bacteria that express and export group 2 capsular polysaccharides. This gene cluster is divided into three regions corresponding to proteins with specific functions[29,37,38,50,51,52,53,54] (Figure 2).
Regions 1 and 3 of the gene cluster encode capsular polysaccharide export proteins of the ABC transporter-dependent system and auxiliary proteins useful for this export, such as capsular polysaccharide polymerization proteins[2,21,29,55]. These genes are conserved between bacterial species that express group 2 capsular polysaccharides[27,53].
Region 2 groups together a variable number of genes that encode biosynthesizing proteins of the saccharide units[9,21,38,50,53,56]. The organization and number of these genes is specific to the type of polysaccharide expressed.
The order of the regions may differ depending on the bacterial species. For example, in species of the genus Neisseria, the regions are named A (capsule synthesis), B (capsule export), C (capsule translocation) and the order of the three regions appears to be randomly distributed[57].
The organization of genes involved in the biosynthesis and export of group 3 capsular polysaccharides is diverse[38]. Regions 1 and 3 of the gene cluster have a poorly understood organization. Genes encoding the ABC transporter-dependent system and auxiliary proteins have no specific organization and are mixed. Similar to group 2, region 2 comprises the necessary genes for the biosynthesis of saccharic units[38]. The organization and amount of these genes are specific to each polysaccharide type.
2.5. Polymerization Steps of Group 2 and 3 Capsular Polysaccharides in Gram-Negative Bacteria
Capsular polysaccharides of groups 2 and 3 are polymerized in the cytoplasm and then exported across membranes by an ABC transporter-dependent system.
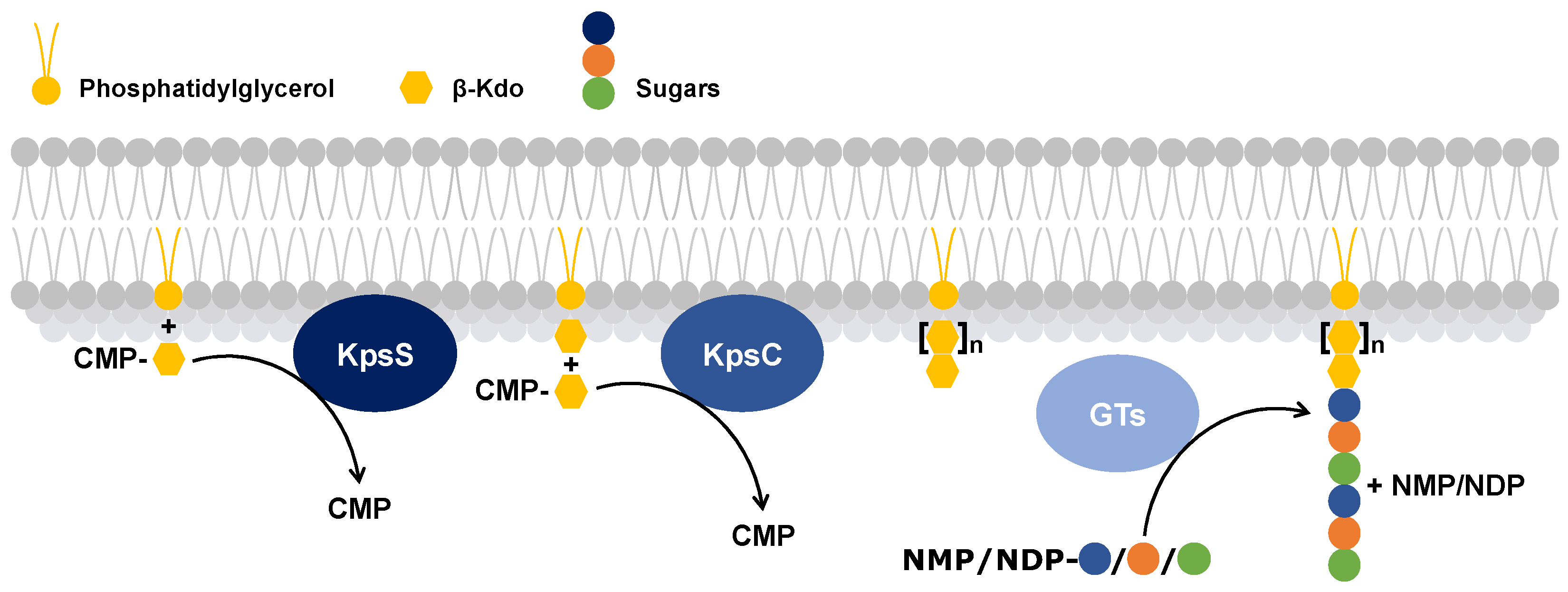
The proteins that polymerize the capsular polysaccharides have retained the same functions; therefore, the polymerization process of capsular polysaccharides has been determined in E. coli. β-Kdo-transferases are a subfamily of glycosyltransferases that play a key role in the polymerization of capsular polysaccharides. These enzymes add a Kdo to a phosphatidylglycerol molecule[59]. For the polymerization, two β-Kdo-transferases are used[59,60,61] (Figure 3):
- The first, KpsS, transfers a β-Kdo residue to a phosphatidylglycerol molecule to form a primer.
- The second, KpsC, extends this primer, using one or more molecules of cytidine 5′-monophosphate-Kdo, which serves as the lipid anchor of the capsular polysaccharide[61].
Figure 3.
Schematic representation of the process of lipid anchor biosynthesis followed by polymerization of a capsular polysaccharide. The yellow lipid corresponds to a phosphatidylglycerol, to which a β-Kdo (yellow hexagon) is added by KpsS and then one or more β-Kdo are added by KpsC. Subsequently, glycosyltransferases (GTs) add specific sugars (blue, orange and green circles) to the lipid anchor to polymerize the capsular polysaccharide. CMP: Cytidine 5′-MonoPhosphate, NMP: Nucleoside MonoPhosphate, NDP: Nucleoside DiPhosphate.
Figure 3.
Schematic representation of the process of lipid anchor biosynthesis followed by polymerization of a capsular polysaccharide. The yellow lipid corresponds to a phosphatidylglycerol, to which a β-Kdo (yellow hexagon) is added by KpsS and then one or more β-Kdo are added by KpsC. Subsequently, glycosyltransferases (GTs) add specific sugars (blue, orange and green circles) to the lipid anchor to polymerize the capsular polysaccharide. CMP: Cytidine 5′-MonoPhosphate, NMP: Nucleoside MonoPhosphate, NDP: Nucleoside DiPhosphate.
3. Export Systems for Capsular Polysaccharides in Gram-Negative Bacteria
Three systems exporting capsular polysaccharides can be found in Gram-negative bacteria: the Wzx/Wzy dependent system, the synthase-dependent system and the ABC dependent system[63] (Figure 4). Some bacteria, such as E. coli, can expressed different capsular polysaccharide groups and therefore possess different export systems[37,38].
3.1. Wzx/Wzy-Dependent System
In group 1 and 4 polysaccharide capsules, export is mediated by the Wzx/Wzy transporter-dependent system. This system is composed of six proteins: a glycosyltransferase (GT), a flippase (Wzx), a polymerase (Wzy), a polysaccharide copolymerase protein (PCP), and an outer membrane polysaccharide export protein (OPX) (Figure 4a).
Gram-negative bacteria such as E. coli use the Wzx/Wzy-dependent system for polymerization of capsular polysaccharides and O-antigens in the periplasm followed by their export from the periplasm to the outer membrane[64,65]. O-antigen polysaccharides will bind to the end of the lipopolysaccharide at the outer membrane to protect bacteria from external attack, increase antibiotic resistance, evade the immune system, and stabilize the outer membrane[66,67]. Capsular polysaccharides will insert themselves into the outer membrane to fulfill their role as virulence factors[21,27].
The polymerization process of capsular polysaccharides and O-antigens occurs in the cytoplasm and begins with a GT that links a sugar residue to a lipid[38]. The flippase Wzx recognizes and transports this activated lipid into the periplasm. Then, the polymerase Wzy adds sugar moieties to the reduced end of the activated lipid, turning it into a polysaccharide. Then, Wzc, a PCP, verifies the length of this polysaccharide and exports it to the outer membrane with the help of Wza, an OPX.
3.2. Synthase-Dependent System
Gram-negative bacteria, such as Pseudomonas aeruginosa, use the synthase-dependent system to synthesize and export exopolysaccharides, such as cellulose and hyaluronic acid, directly from the cytoplasm[71,72]. These polysaccharide homopolymers form a biofilm that protects bacteria. The synthase-dependent system consists of a glycosyltransferase (GT) coupled to a translocase called synthase in the inner membrane and a tetratricopeptide repeat (TPR) protein coupled to a β-barrel exporter protein in the outer membrane (Figure 4b).
3.3. ABC Transporter-Dependent System
For group 2 and 3 polysaccharide capsules, export is done using the ABC transporter-dependent system[21]. This system is composed of four proteins: the nucleotide-binding domain (NBD) and the transmembrane domain (TMD) that constitute the ABC transporter, and the PCP and OPX that make up the ABC transporter-dependent system (Figure 4c).
Gram-negative bacteria, such as Neisseria meningitidis or E. coli, use the ABC transporter-dependent system for the export of capsular polysaccharides to the outer membrane[64,73].
Unlike the Wzx/Wzy transporter-dependent system, polymerization of capsular polysaccharides occurs in the cytoplasm, independently of the ABC transporter-dependent system, with the help of a glycosyltransferase that adds sugar fragments to the lipid anchor. Then, the ABC transporter-dependent system recognizes the capsular polysaccharide by a process that is still poorly understood and exports it across the inner membrane[74]. Then, PCP and OPX export the capsular polysaccharide through the periplasm and outer membrane by a little-known process.
Present in Gram-negative pathogenic bacteria, this ABC transporter-dependent system represents approximately 10 % of polysaccharide capsule transporters[19].
The rest of the capsule transporters are from unknown systems that would allow the export of proteins forming the protein capsules [19].
4. Export of Group 2 and 3 Capsular Polysaccharides by ABC Transporter-Dependent Systems in Gram-Negative Bacteria
4.1. The ABC Transporter and Its Dependent System
ABC transporters are a large family of proteins found in all living organisms[75,76]. They use the energy generated by the hydrolysis of adenosine triphosphate (ATP) to adenosine diphosphate (ADP) to import or export molecules. The ABC transporter-dependent system is composed of four proteins for capsular polysaccharides that span both membranes of Gram-negative pathogenic bacteria[21,73]. The ABC transporter itself is composed of two proteins: the NBD in the cytoplasm and the TMD in the inner membrane. The PCP, which is in the periplasm, connects the OPX in the outer membrane to the ABC transporter. The NBD hydrolyzes ATP to ADP while the TMD, PCP, and OPX form a channel that allows capsular polysaccharides to cross the inner and outer bacterial membranes[62]. Then, the capsular polysaccharide is inserted outside the outer membrane.
ABC transporter-dependent systems have been computationally predicted to have similar functions and folds[62,64,77,78,79]. However, since the different proteins of the ABC transporter-dependent systems share little amino acid sequence identity, they could have different functions. For example, Kps (for Kapsel polysaccharide synthesis) proteins were first detected in E. coli, and then homologous proteins were named similarly[54] (Table 3). Others have a different name such as Ctr, for Neisseria meningitidis, which refers to the Capsule transporter[55,80].
4.2. Structural Description of the ABC Transporter-Dependent System Proteins
4.2.1. NBD and TMD, the Two ABC Transporter Proteins
The ABC transporter superfamily share a basic architecture that includes two homodimers of NBD and TMD.
NBDs share a common fold and similar functions. The NBD is divisible into two constitutive domains: a central catalytic domain containing a Walker A motif (GXXGXGK(S/T)), a Walker B motif (ϕϕϕϕD, where ϕ is a hydrophobic residue), a Q loop, and an H motif (switch region); and a more structurally diverse α-helical domain containing the ABC transporter signature motif (LSGGQ)[81].
TMDs that export polysaccharides share a similar folding (TM-score between 0.61 and 0.81) and similar functions. TMDs are specific to the exported molecule and form the beginning of the export tunnel in the inner membrane[81].
In a recent study, Kuklewicz and Zimmer[74] expressed the ABC transporter-dependent system of Caldimonas thermodepolymerans and the capsule of Pasteurella multocida in an E. coli strain[74]. Then, in the presence of ATP, they were able to obtain three-dimensional structures of KpsMT of Caldimonas thermodepolymerans (Figure 5).
In the absence of ATP and Pasteurella multocida capsular polysaccharides, KpsMT transmembrane channel is closed[74] (Figure 5A). Instead, KpsM subunits (TMD) form a lipid-exposed pocket where the lipid anchor, a Kdo–phosphatidylglycerol (PG) lipid molecule, can bind to and be recognized by KpsM.
In the presence of ATP, the two KpsT (NBD) subunits are closer (Figure 5B). Following ATP hydrolysis in ADP, the space between the KpsM subunits on the periplasmic side increases (Figure 5C). This alters the conformations of the TMDs, which may cause the polysaccharide to move into the tunnel. Binding of the lipid anchor between the two TMDs opens the tunnel.
In the presence of capsular polysaccharides from Pasteurella multocida, the lipid anchor of capsular polysaccharide is fixed in the pocket of the KpsM subunit (Figure 5D). Two structures with different orientations of the lipid anchor were obtained. The first orientation shows that the lipid anchor can be inserted between the KpsM subunits. The second orientation leads to greater space of the KpsM subunits, thus forming a tunnel between these two KpsM subunits.
4.2.2. Polysaccharide Co-Polymerase Protein (PCP)
PCPs are a family of proteins that link a transporter in the internal membrane and the OPX in the external membrane[23,38]. Three PCP subfamilies exist. They have a similar folding but can have supplemental functions (Figure 6)[64,82,83].
PCP-1 and PCP-2, both present in the Wzx/Wzy-dependent system, regulate polysaccharide length, with PCP-2 containing a tyrosine kinase domain. PCP-3 is present in the ABC transporter-dependent system and its function is unknown.
PCPs share less than 10 % amino acid sequence identity, but their three-dimensional structures have similarities[64].
The most conserved structural features in the PCP family are two hydrophobic α helices anchored in the inner membrane, a large periplasmic domain made up of a α/β domain at its base and a helical hairpin at its top (Figure 6). The level of oligomerization depends on experimental conditions and varies between three and twelve polypeptides[64,77,82,84,85].
In the cytoplasmic region, PCP-2 contains a tyrosine kinase domain that PCP-1 and PCP-3 lack[64,77].
In the PCP-3 subfamily, the C-terminal helix can have different contacts with the inner membrane. In KpsE from E. coli, the α-terminal helix lies in the periplasm on the outer part of the inner membrane, whereas the same α-helix is anchored in the inner membrane in CtrB from Neisseria meningitidis[83,86].
In the periplasm, 21 hydrophobic PCP amino acids are conserved in the α/β domain and in the hairpin. The latter consists mainly of residues of leucine, alanine, isoleucine and glycine, which form a leucine zipper[87]. Figure 6 shows that PCP-1 and PCP-3 share a common fold in the periplasmic domain and the PCP-2 periplasmic domain is compacted[88].
4.2.3. Outer Membrane Polysaccharide Export Protein (OPX)
OPX is a family of proteins found in the outer membrane of Gram-negative bacteria[79]. These proteins allow capsular polysaccharides export by the ABC transporter-dependent system across the outer membrane.
In the Protein Data Bank (PDB), the only OPX structure, including the transmembrane domain, is Wza from E. coli (PDB code: 2J58), which is an acetylated lipoprotein (Figure 7)[89]. Wza forms a tunnel with eight polypeptides composed of α-helices and β-sheets. This tunnel is ~18 Å in diameter and can accommodate large molecules such as polysaccharide chains (Figure 7).
Domain 1 of Wza forms a ring that is composed of an antiparallel β-sandwich on the inner part of the tunnel and α-helices on the outer face[90]. The bottom of the domain 1 ring is blocked by eight loops.
Domains 2 and 3 are duplicated and form two rings containing a mixture of β-sheets and α-helices. The ring of domain 3 is larger than that of domain 2.
Domain 4 is anchored to the outer membrane by an amphipathic α-helix barrel[90]. The channel of domain 4 is blocked by eight loops that prevent the passage of unwanted molecules from outside the cell into the periplasm.
In OPXs, very few amino acid residues are identical (approximately 1 %). Despite the low level of sequence identity, OPX structure predictions indicate that these proteins fold in a similar manner[21,41,79,89,90]. Domains 1 and 2 of OPX contain the polysaccharide export sequence (PES, Pfam: 02563) that defines the OPX family[91].
5. Potential Mechanism of Capsular Polysaccharide Transport by the ABC Transporter-Dependent System
According to the experiments of Kuklewicz and Zimmer[74], the export of capsular polysaccharides by the ABC transporter-dependent system appears to begin with the recognition of the lipid anchor by KpsM (TMD), followed by spontaneous binding of the lipid anchor into the pocket of the KpsM subunit[74] (Figure 8). The two structures of KpsMT with the lipid anchor (Figure 5) show the presence of two distinct binding sites in the KpsM subunits or a reorientation of the lipid anchor in the KpsM pocket.
ATP hydrolysis changes the conformation of the KpsM subunits, forming a tunnel[74]. This movement of KpsM subunits could allow the passage of capsular polysaccharide sugars through the tunnel formed by KpsM, KpsE (PCP) and KpsD (OPX)[74] (Figure 8).
It is assumed that the lipid anchor would undergo translocation within the inner membrane from its inner face to its outer face[74] (Figure 8). There is no information on whether the lipid anchor is exported with or without the capsular polysaccharide. However, the hypothesis of a cleavage of the lipid from the lipid anchor has been put forward. This would imply the existence of a process allowing the capsular polysaccharide to be linked to a new lipid in the outer membrane.
6. Discussion
Antibiotic-resistant bacteria were responsible for 6 million deaths in 2019[1]. To reduce this number of deaths, the WHO published a list of antibiotic-resistant bacteria that should be prioritized for research to develop new antibiotics. In this list, seven bacteria or families of bacteria are Gram-negative[1]. To infect a host, these bacteria use many virulence factors, including the polysaccharide capsule[7].
Three transporter-dependent systems (Wzx/Wzy, synthase and ABC) can export capsular polysaccharides. The Wzx/Wzy-dependent system biosynthesizes capsular polysaccharides in the cytoplasm, exports them to the periplasm, then continues the biosynthesis and exports them to the outer membrane[67]. The synthase-dependent system biosynthesizes capsular polysaccharides in the cytoplasm and simultaneously exports them to the outer membrane. The ABC transporter-dependent system exports to the outer membrane only those capsular polysaccharides that have been biosynthesized in the cytoplasm by an independent glycosyltransferase.
The ABC transporter-dependent system is composed of four types of proteins: an NBD and a TMD of the ABC transporter, a PCP-3 and an OPX[67]. This complex allows capsular polysaccharides to cross the inner and outer membranes of bacteria using the hydrolysis energy of ATP.
The structures of the inner membrane-anchored ABC transporter have been solved, showing that the lipid anchor can spontaneously bind into the TMDs (Figure 5) and the PCP-3 complex forms a cage around the TMDs[74].
Although the capsular polysaccharide export mechanism of the ABC transporter-dependent system is not yet fully understood, Kuklewicz and Zimmer provide insight into this mechanism (Figure 8)[74]. For Wzm/Wzt (KpsMT-homologous ABC transporter secreting O-antigen polysaccharides), present in the bacterium Aquifex aeolicus, recognition of O-antigen polysaccharides is achieved by detecting the region of the polysaccharide consisting of a phosphatidylglycerol and a Kdo oligosaccharide[92]. For KpsMT, it is possible that this principle may apply. Indeed, Kuklewicz and Zimmer assume that the lipid anchor (composed of a phosphatidylglycerol and a Kdo oligosaccharide), by binding to a TMD, would initiate the binding of the capsular polysaccharide to the TMD and its export in the ABC transporter, then the hydrolysis of ATP would modify the conformation of the ABC transporter to allow the export of the capsular polysaccharide[74]. Furthermore, for KpsMT, due to a pseudo-double symmetry, the lipid anchor appears to bind in one half of the TMD, KpsM. Therefore, it seems possible that two lipid anchors bind at the same time in both KpsM subunits[74]. This could lead to the export of two capsular polysaccharides by an ABC transporter-dependent system.
The truncated structure of octameric PCP-3 shows that it forms a cage around the ABC transporter composed of hydrophobic α-helices embedded in the inner membrane[74]. This arrangement may create a continuous tunnel from the TMD to the OPX. There are three subfamilies of PCP, the structure of PCP-3 is more similar to the structure of PCP-1 but PCP-3 does not control the length of the capsular polysaccharide[64].
In the ABC transporter-dependent system, OPX can have two types of transmembrane domains, either with an α-helix or a GfcC-like domain[79]. GfcC is a soluble ligand-binding β-protein and is expected to interact with the capsular polysaccharide[93]. The difference in transmembrane domains could indicate that OPXs have different functions.
However, some parts of the mechanism of capsular polysaccharide export by the ABC transporter-dependent system are still not understood:
- Is the lipid anchor lipid exported with the capsular polysaccharide? Kuklewicz and Zimmer hypothesize that the lipid anchor could be cleaved from the capsular polysaccharides when the capsular polysaccharide is engaged in the tunnel[74]. Indeed, there is currently no evidence for lipid transport through the ABC transporter-dependent system. This also implies that another lipid anchor would be available in the outer membrane to bind with a capsular polysaccharide.
- Do PCP and OPX have a function other than that of exporter? Depending on PCP and OPX, this could be possible. Indeed, the C-terminal helix of PCP-3 does not interact in the same way with the inner membrane in different bacterial species[83,86]. This interaction could have an effect on the function of PCP-3. In addition, some OPXs of the ABC transporter-dependent system may have a transmembrane domain that replaces the α-helix[79]. Interaction of this domain with capsular polysaccharides could modify the export process.
A better understanding of the ABC transporter-dependent system would allow the development of capsular polysaccharide export inhibitors to combat Gram-negative bacterial pathogens and to counter antibiotic resistance. A better understanding of OPXs in ABC transporter-dependent systems could also be useful for the development of conjugate vaccines targeting both the polysaccharide capsule and OPXs[94].
Funding
This research received no external funding.
Acknowledgments
We thank Prof. Kristina Djinovic-Carugo for her support, encouragement and advice for the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- WHO Bacterial Priority Pathogens List 2024 Bacterial Pathogens of Public Health Importance, to Guide Research, Development, and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, 2024; ISBN 978-92-4-009346-1.
- Hill, D.J.; Griffiths, N.J.; Borodina, E.; Virji, M. Cellular and Molecular Biology of Neisseria Meningitidis Colonization and Invasive Disease. Clinical Science 2010, 118, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Blokesch, M. Natural Competence for Transformation. Current Biology 2016, 26, R1126–R1130. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, A.; Zhu, J. Pathogenicity and Virulence Regulation of Vibrio Cholerae at the Interface of Host-Gut Microbiome Interactions. Virulence 2020, 11, 1582–1599. [Google Scholar] [CrossRef]
- Bartley, S.N.; Tzeng, Y.L.; Heel, K.; Lee, C.W.; Mowlaboccus, S.; Seemann, T.; Lu, W.; Lin, Y.H.; Ryan, C.S.; Peacock, C.; et al. Attachment and Invasion of Neisseria Meningitidis to Host Cells Is Related to Surface Hydrophobicity, Bacterial Cell Size and Capsule. PLoS One 2013, 8, e55798. [Google Scholar] [CrossRef]
- Livorsi, D.J.; Stenehjem, E.; Stephens, D.S. Virulence Factors of Gram-Negative Bacteria in Sepsis with a Focus on Neisseria Meningitidis. Contrib Microbiol 2011, 17, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L. Virulence Factors and Their Mechanisms of Action: The View from a Damage–Response Framework. Journal of Water and Health 2009, 7, S2–S18. [Google Scholar] [CrossRef]
- Silale, A.; Zhu, Y.; Witwinowski, J.; Smith, R.E.; Newman, K.E.; Bhamidimarri, S.P.; Baslé, A.; Khalid, S.; Beloin, C.; Gribaldo, S.; et al. Dual Function of OmpM as Outer Membrane Tether and Nutrient Uptake Channel in Diderm Firmicutes. Nat Commun 2023, 14, 7152. [Google Scholar] [CrossRef]
- Cifuente, J.O.; Schulze, J.; Bethe, A.; Di Domenico, V.; Litschko, C.; Budde, I.; Eidenberger, L.; Thiesler, H.; Ramón Roth, I.; Berger, M.; et al. A Multi-Enzyme Machine Polymerizes the Haemophilus Influenzae Type b Capsule. Nat Chem Biol 2023, 19, 865–877. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Thomas, V.C.; Hancock, L.E. Capsular Polysaccharide Production in Enterococcus Faecalis and Contribution of CpsF to Capsule Serospecificity. Journal of Bacteriology 2009, 191, 6203–6210. [Google Scholar] [CrossRef]
- Akoolo, L.; Pires, S.; Kim, J.; Parker, D. The Capsule of Acinetobacter Baumannii Protects against the Innate Immune Response. Journal of Innate Immunity 2022, 14, 543–554. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Palacios, A.; Anso, I.; Cifuente, J.; Anguita, J.; Jacobs, W.R., Jr; Guerin, M.E.; Prados-Rosales, R. The Mycobacterium Tuberculosis Capsule: A Cell Structure with Key Implications in Pathogenesis. Biochemical Journal 2019, 476, 1995–2016. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Song, J. Single Missense Mutations in Vi Capsule Synthesis Genes Confer Hypervirulence to Salmonella Typhi. Nat Commun 2024, 15, 5258. [Google Scholar] [CrossRef] [PubMed]
- Perera, S.R.; Sokaribo, A.S.; White, A.P. Polysaccharide Vaccines: A Perspective on Non-Typhoidal Salmonella. Polysaccharides 2021, 2, 691–714. [Google Scholar] [CrossRef]
- O’Riordan, K.; Lee, J.C. Staphylococcus Aureus Capsular Polysaccharides. Clin Microbiol Rev 2004, 17, 218–234. [Google Scholar] [CrossRef]
- Wessels, M.R. Capsular Polysaccharide of Group A Streptococcus. Microbiol Spectr 2019, 7, 7.1.01. [CrossRef]
- Paton, J.C.; Trappetti, C. Streptococcus Pneumoniae Capsular Polysaccharide. Microbiology Spectrum 2019, 7, 10.1128/microbiolspec.gpp3-0019–2018. [CrossRef]
- Vaz, M.J.; Dongas, S.; Ratner, A.J. Capsule Production Promotes Group B Streptococcus Intestinal Colonization. Microbiol Spectr 2023, 11, e02349-23. [Google Scholar] [CrossRef]
- Rendueles, O.; Garcia-Garcerà, M.; Néron, B.; Touchon, M.; Rocha, E.P.C. Abundance and Co-Occurrence of Extracellular Capsules Increase Environmental Breadth: Implications for the Emergence of Pathogens. PLoS Pathog 2017, 13, e1006525. [Google Scholar] [CrossRef]
- Willis, L.M.; Stupak, J.; Richards, M.R.; Lowary, T.L.; Li, J.; Whitfield, C. Conserved Glycolipid Termini in Capsular Polysaccharides Synthesized by ATP-Binding Cassette Transporter-Dependent Pathways in Gram-Negative Pathogens. Proc Natl Acad Sci U S A 2013, 110, 7868–7873. [Google Scholar] [CrossRef]
- Silver, R.P.; Prior, K.; Nsahlai, C.; Wright, L.F. ABC Transporters and the Export of Capsular Polysaccharides from Gram-Negative Bacteria. Research in Microbiology 2001, 152, 357–364. [Google Scholar] [CrossRef]
- Pelkonen, S.; Häyrinen, J.; Finne, J. Polyacrylamide Gel Electrophoresis of the Capsular Polysaccharides of Escherichia Coli K1 and Other Bacteria. J Bacteriol 1988, 170, 2646–2653. [Google Scholar] [CrossRef]
- Kröncke, K.D.; Golecki, J.R.; Jann, K. Further Electron Microscopic Studies on the Expression of Escherichia Coli Group II Capsules. J Bacteriol 1990, 172, 3469–3472. [Google Scholar] [CrossRef]
- Stephens, D.S. Neisseria Meningitidis. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Bennett, J.E.M.D., Dolin, R.M.D., Blaser, M.J.M.D., Eds.; 2020; pp. 2585-2607.e7 ISBN 978-0-323-48255-4.
- Tzeng, Y.-L.; Thomas, J.; Stephens, D.S. Regulation of Capsule in Neisseria Meningitidis. Critical Reviews in Microbiology 2015, 1–14. [Google Scholar] [CrossRef]
- Gao, S.; Jin, W.; Quan, Y.; Li, Y.; Shen, Y.; Yuan, S.; Yi, L.; Wang, Y.; Wang, Y. Bacterial Capsules: Occurrence, Mechanism, and Function. npj Biofilms Microbiomes 2024, 10, 21. [Google Scholar] [CrossRef]
- Sukupolvi-Petty, S.; Grass, S.; StGeme, J.W. The Haemophilus Influenzae Type b hcsA and hcsB Gene Products Facilitate Transport of Capsular Polysaccharide across the Outer Membrane and Are Essential for Virulence. J Bacteriol 2006, 188, 3870–3877. [Google Scholar] [CrossRef]
- Frosch, M.; Edwards, U.; Bousset, K.; Krausse, B.; Weisgerber, C. Evidence for a Common Molecular Origin of the Capsule Gene Loci in Gram-Negative Bacteria Expressing Group II Capsular Polysaccharides. Mol Microbiol 1991, 5, 1251–1263. [Google Scholar] [CrossRef]
- Kroll, J.S.; Loynds, B.; Brophy, L.N.; Moxon, E.R. The Bex Locus in Encapsulated Haemophilus Influenzae : A Chromosomal Region Involved in Capsule Polysaccharide Export. Molecular Microbiology 1990, 4, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Li, N.; Yokoyama, H.; Ezaki, T. Complete Nucleotide Sequence and Molecular Characterization of ViaB Region Encoding Vi Antigen in Salmonella Typhi. J Bacteriol 1993, 175, 4456–4465. [Google Scholar] [CrossRef] [PubMed]
- Moxon, E.R.; Vaughn, K.A. The Type b Capsular Polysaccharide as a Virulence Determinant of Haemophilus Influenzae: Studies Using Clinical Isolates and Laboratory Transformants. Journal of Infectious Diseases 1981, 143, 517–524. [Google Scholar] [CrossRef]
- Keo, T.; Collins, J.; Kunwar, P.; Blaser, M.J.; Iovine, N.M. Campylobacter Capsule and Lipooligosaccharide Confer Resistance to Serum and Cationic Antimicrobials. Virulence 2011, 2, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-Alonso, S.; Blundell-Hunter, G.; Fu, Z.; Gladstone, R.A.; Fillol-Salom, A.; Loraine, J.; Cloutman-Green, E.; Johnsen, P.J.; Samuelsen, Ø.; Pöntinen, A.K.; et al. Evolutionary and Functional History of the Escherichia Coli K1 Capsule. Nat Commun 2023, 14, 3294. [Google Scholar] [CrossRef]
- Guerry, P.; Poly, F.; Riddle, M.; Maue, A.C.; Chen, Y.-H.; Monteiro, M.A. Campylobacter Polysaccharide Capsules: Virulence and Vaccines. Front. Cell. Inf. Microbio. 2012, 2. [Google Scholar] [CrossRef]
- Wilson, B.A.; Ho, M. Pasteurella Multocida: From Zoonosis to Cellular Microbiology. Clin Microbiol Rev 2013, 26, 631–655. [Google Scholar] [CrossRef]
- Hurtado, R.; Maturrano, L.; Azevedo, V.; Aburjaile, F. Pathogenomics Insights for Understanding Pasteurella Multocida Adaptation. International Journal of Medical Microbiology 2020, 310, 151417. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Roberts, I.S. Structure, Assembly and Regulation of Expression of Capsules in Escherichia Coli. Mol Microbiol 1999, 31, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C. Biosynthesis and Assembly of Capsular Polysaccharides in Escherichia Coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef] [PubMed]
- Diaz Romero, J.; Outschoorn, I.M. Current Status of Meningococcal Group B Vaccine Candidates: Capsular or Noncapsular? Clin Microbiol Rev 1994, 7, 559–575. [Google Scholar] [CrossRef]
- Lin, D.; Fan, J.; Wang, J.; Liu, L.; Xu, L.; Li, F.; Yang, J.; Li, B. The Fructose-Specific Phosphotransferase System of Klebsiella Pneumoniae Is Regulated by Global Regulator CRP and Linked to Virulence and Growth. Infect Immun 2018, 86, e00340-18. [Google Scholar] [CrossRef]
- Niu, T.; Guo, L.; Luo, Q.; Zhou, K.; Yu, W.; Chen, Y.; Huang, C.; Xiao, Y. Wza Gene Knockout Decreases Acinetobacter Baumannii Virulence and Affects Wzy-Dependent Capsular Polysaccharide Synthesis. Virulence 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Schembri, M.A.; Dalsgaard, D.; Klemm, P. Capsule Shields the Function of Short Bacterial Adhesins. J Bacteriol 2004, 186, 1249–1257. [Google Scholar] [CrossRef]
- Zafar, M.A.; Hamaguchi, S.; Zangari, T.; Cammer, M.; Weiser, J.N. Capsule Type and Amount Affect Shedding and Transmission of Streptococcus Pneumoniae. mBio 2017, 8, e00989-17. [Google Scholar] [CrossRef]
- Stephens, D.S.; Swartley, J.S.; Kathariou, S.; Morse, S.A. Insertion of Tn916 in Neisseria Meningitidis Resulting in Loss of Group B Capsular Polysaccharide. Infect Immun 1991, 59, 4097–4102. [Google Scholar] [CrossRef]
- Ophir, T.; Gutnick, D.L. A Role for Exopolysaccharides in the Protection of Microorganisms from Desiccation. Appl Environ Microbiol 1994, 60, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Lewis, G.D.; Ashokkumar, M.; Hemar, Y. Inactivation of Microorganisms by Low-Frequency High-Power Ultrasound: 1. Effect of Growth Phase and Capsule Properties of the Bacteria. Ultrasonics Sonochemistry 2014, 21, 446–453. [Google Scholar] [CrossRef]
- Waz, N.T.; Oliveira, S.; Girardello, R.; Lincopan, N.; Barazzone, G.; Parisotto, T.; Hakansson, A.P.; Converso, T.R.; Darrieux, M. Influence of the Polysaccharide Capsule on the Bactericidal Activity of Indolicidin on Streptococcus Pneumoniae. Front. Microbiol. 2022, 13, 898815. [Google Scholar] [CrossRef]
- Llobet, E.; Tomás, J.M.; Bengoechea, J.A. Capsule Polysaccharide Is a Bacterial Decoy for Antimicrobial Peptides. Microbiology 2008, 154, 3877–3886. [Google Scholar] [CrossRef] [PubMed]
- De Smidt, O.; Albertyn, J.; Bragg, R.R.; Van Heerden, E. Genetic Organisation of the Capsule Transport Gene Region from <i>Haemophilus Paragallinarum<i/>. Onderstepoort J Vet Res 2004, 71, 139–152. [Google Scholar] [CrossRef]
- Van Eldere, J.; Brophy, L.; Loynds, B.; Celis, P.; Hancock, I.; Carman, S.; Kroll, J.S.; Moxon, E.R. Region II of the Haemophilus Influenzae Type Be Capsulation Locus Is Involved in Serotype-Specific Polysaccharide Synthesis. Mol Microbiol 1995, 15, 107–118. [Google Scholar] [CrossRef]
- Kroll, J.S.; Zamze, S.; Loynds, B.; Moxon, E.R. Common Organization of Chromosomal Loci for Production of Different Capsular Polysaccharides in Haemophilus Influenzae. J Bacteriol 1989, 171, 3343–3347. [Google Scholar] [CrossRef] [PubMed]
- Bliss, J.M.; Garon, C.F.; Silver, R.P. Polysialic Acid Export in Escherichia Coli K1: The Role of KpsT, the ATP-Binding Component of an ABC Transporter, in Chain Translocation. Glycobiology 1996, 6, 445–452. [Google Scholar] [CrossRef]
- Roberts, I.S. The Biochemistry and Genetics of Capsular Polysaccharide Production in Bacteria. Annu Rev Microbiol 1996, 50, 285–315. [Google Scholar] [CrossRef]
- Vimr, E.; Steenbergen, S.; Cieslewicz, M. Biosynthesis of the Polysialic Acid Capsule inEscherichia Coli K1. Journal of Industrial Microbiology 1995, 15, 352–360. [Google Scholar] [CrossRef]
- Frosch, M.; Weisgerber, C.; Meyer, T.F. Molecular Characterization and Expression in Escherichia Coli of the Gene Complex Encoding the Polysaccharide Capsule of Neisseria Meningitidis Group B. Proc Natl Acad Sci U S A 1989, 86, 1669–1673. [Google Scholar] [CrossRef] [PubMed]
- Karlyshev, A.V.; Champion, O.L.; Churcher, C.; Brisson, J.-R.; Jarrell, H.C.; Gilbert, M.; Brochu, D.; St Michael, F.; Li, J.; Wakarchuk, W.W.; et al. Analysis of Campylobacter Jejuni Capsular Loci Reveals Multiple Mechanisms for the Generation of Structural Diversity and the Ability to Form Complex Heptoses. Mol Microbiol 2005, 55, 90–103. [Google Scholar] [CrossRef]
- Clemence, M.E.A.; Maiden, M.C.J.; Harrison, O.B. Characterization of Capsule Genes in Non-Pathogenic Neisseria Species. Microbial Genomics 2018, 4. [Google Scholar] [CrossRef]
- Cieslewicz, M.; Vimr, E. Thermoregulation of kpsF, the First Region 1 Gene in the Kps Locus for Polysialic Acid Biosynthesis in Escherichia Coli K1. J Bacteriol 1996, 178, 3212–3220. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.; Ovchinnikova, O.G.; Huang, B.-S.; Forrester, T.J.B.; Lowary, T.L.; Kimber, M.S.; Whitfield, C. Mechanism and Linkage Specificities of the Dual Retaining β-Kdo Glycosyltransferase Modules of KpsC from Bacterial Capsule Biosynthesis. Journal of Biological Chemistry 2023, 299. [Google Scholar] [CrossRef] [PubMed]
- Willis, L.M.; Whitfield, C. KpsC and KpsS Are Retaining 3-Deoxy-D-Manno-Oct-2-Ulosonic Acid (Kdo) Transferases Involved in Synthesis of Bacterial Capsules. Proceedings of the National Academy of Sciences 2013, 20753–20758. [Google Scholar] [CrossRef]
- Doyle, L.; Ovchinnikova, O.G.; Myler, K.; Mallette, E.; Huang, B.S.; Lowary, T.L.; Kimber, M.S.; Whitfield, C. Biosynthesis of a Conserved Glycolipid Anchor for Gram-Negative Bacterial Capsules. Nat Chem Biol 2019, 15, 632–640. [Google Scholar] [CrossRef]
- Willis, L.M.; Whitfield, C. Structure, Biosynthesis, and Function of Bacterial Capsular Polysaccharides Synthesized by ABC Transporter-Dependent Pathways. Carbohydr Res 2013, 378, 35–44. [Google Scholar] [CrossRef]
- Caffalette, C.A.; Kuklewicz, J.; Spellmon, N.; Zimmer, J. Biosynthesis and Export of Bacterial Glycolipids. Annu Rev Biochem 2020, 89, 741–768. [Google Scholar] [CrossRef]
- Cuthbertson, L.; Mainprize, I.L.; Naismith, J.H.; Whitfield, C. Pivotal Roles of the Outer Membrane Polysaccharide Export and Polysaccharide Copolymerase Protein Families in Export of Extracellular Polysaccharides in Gram-Negative Bacteria. Microbiol Mol Biol Rev 2009, 73, 155–177. [Google Scholar] [CrossRef]
- Standish, A.J.; Morona, R. Capsule Structure, Synthesis, and Regulation. In Streptococcus Pneumoniae; Elsevier Inc., 2015; pp. 169–179 ISBN 978-0-12-410530-0.
- Perez-Burgos, M.; Garcia-Romero, I.; Jung, J.; Schander, E.; Valvano, M.A.; Sogaard-Andersen, L. Characterization of the Exopolysaccharide Biosynthesis Pathway in Myxococcus Xanthus. J Bacteriol 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Wear, S.S.; Sande, C. Assembly of Bacterial Capsular Polysaccharides and Exopolysaccharides. Annu Rev Microbiol 2020, 74, 521–543. [Google Scholar] [CrossRef]
- Cross, A.S.; Kim, K.S.; Wright, D.C.; Sadoff, J.C.; Gemski, P. Role of Lipopolysaccharide and Capsule in the Serum Resistance of Bacteremic Strains of Escherichia Coli. J Infect Dis 1986, 154, 497–503. [Google Scholar] [CrossRef]
- Hug, I.; Feldman, M.F. Analogies and Homologies in Lipopolysaccharide and Glycoprotein Biosynthesis in Bacteria. Glycobiology 2011, 21, 138–151. [Google Scholar] [CrossRef]
- Huszczynski, S.M.; Coumoundouros, C.; Pham, P.; Lam, J.S.; Khursigara, C.M. Unique Regions of the Polysaccharide Copolymerase Wzz(2) from Pseudomonas Aeruginosa Are Essential for O-Specific Antigen Chain Length Control. J Bacteriol 2019, 201. [Google Scholar] [CrossRef]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial Exopolysaccharides: Biosynthesis Pathways and Engineering Strategies. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Weigel, P.H.; DeAngelis, P.L. Hyaluronan Synthases: A Decade-plus of Novel Glycosyltransferases. J Biol Chem 2007, 282, 36777–36781. [Google Scholar] [CrossRef]
- Cuthbertson, L.; Kos, V.; Whitfield, C. ABC Transporters Involved in Export of Cell Surface Glycoconjugates. Microbiol Mol Biol Rev 2010, 74, 341–362. [Google Scholar] [CrossRef] [PubMed]
- Kuklewicz, J.; Zimmer, J. Molecular Insights into Capsular Polysaccharide Secretion. Nature 2024, 628, 901–909. [Google Scholar] [CrossRef]
- George, A.M. ABC Transporters - 40 Years On; George, A.M., Ed.; 1st ed.; Springer, 2016; ISBN 978-3-319-23475-5.
- Theodoulou, F.L.; Kerr, I.D. ABC Transporter Research: Going Strong 40 Years On. Biochemical Society Transactions 2015, 43, 1033–1040. [Google Scholar] [CrossRef]
- Kalynych, S.; Cherney, M.; Bostina, M.; Rouiller, I.; Cygler, M. Quaternary Structure of WzzB and WzzE Polysaccharide Copolymerases. Protein Sci 2015, 24, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Aller, S.G.; Beis, K.; Carpenter, E.P.; Chang, G.; Chen, L.; Dassa, E.; Dean, M.; Duong Van Hoa, F.; Ekiert, D.; et al. Structural and Functional Diversity Calls for a New Classification of ABC Transporters. FEBS Lett 2020, 594, 3767–3775. [Google Scholar] [CrossRef]
- Sande, C.; Bouwman, C.; Kell, E.; Nickerson, N.N.; Kapadia, S.B.; Whitfield, C. Structural and Functional Variation in Outer Membrane Polysaccharide Export (OPX) Proteins from the Two Major Capsule Assembly Pathways Present in Escherichia Coli. J Bacteriol 2019, 201. [Google Scholar] [CrossRef]
- Harrison, O.B.; Claus, H.; Jiang, Y.; Bennett, J.S.; Bratcher, H.B.; Jolley, K.A.; Corton, C.; Care, R.; Poolman, J.T.; Zollinger, W.D.; et al. Description and Nomenclature of Neisseria Meningitidis Capsule Locus. Emerging Infectious Diseases 2013, 19, 566. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC Transporters: The Power to Change. Nat Rev Mol Cell Biol 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Kalynych, S.; Yao, D.; Magee, J.; Cygler, M. Structural Characterization of Closely Related O-Antigen Lipopolysaccharide (LPS) Chain Length Regulators. J Biol Chem 2012, 287, 15696–15705. [Google Scholar] [CrossRef] [PubMed]
- Larue, K.; Ford, R.C.; Willis, L.M.; Whitfield, C. Functional and Structural Characterization of Polysaccharide Co-Polymerase Proteins Required for Polymer Export in ATP-Binding Cassette Transporter-Dependent Capsule Biosynthesis Pathways. J Biol Chem 2011, 286, 16658–16668. [Google Scholar] [CrossRef]
- Morona, R.; Purins, L.; Tocilj, A.; Matte, A.; Cygler, M. Sequence-Structure Relationships in Polysaccharide Co-Polymerase (PCP) Proteins. Trends Biochem Sci 2009, 34, 78–84. [Google Scholar] [CrossRef]
- Chang, C.W.; Tran, E.N.; Ericsson, D.J.; Casey, L.W.; Lonhienne, T.; Benning, F.; Morona, R.; Kobe, B. Structural and Biochemical Analysis of a Single Amino-Acid Mutant of WzzBSF That Alters Lipopolysaccharide O-Antigen Chain Length in Shigella Flexneri. PLoS One 2015, 10, e0138266. [Google Scholar] [CrossRef]
- Phoenix, D.A.; Brandenburg, K.; Harris, F.; Seydel, U.; Hammerton, T.; Roberts, I.S. An Investigation into the Membrane-Interactive Potential of the Escherichia Coli KpsE C-Terminus. Biochem Biophys Res Commun 2001, 285, 976–980. [Google Scholar] [CrossRef]
- Papadopoulos, M.; Morona, R. Mutagenesis and Chemical Cross-Linking Suggest That Wzz Dimer Stability and Oligomerization Affect Lipopolysaccharide O-Antigen Modal Chain Length Control. J Bacteriol 2010, 192, 3385–3393. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, J.; Clarke, B.R.; Seidel, L.; Bolla, J.R.; Ward, P.N.; Zhang, P.; Robinson, C.V.; Whitfield, C.; Naismith, J.H. The Molecular Basis of Regulation of Bacterial Capsule Assembly by Wzc. Nat Commun 2021, 12, 4349. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Beis, K.; Nesper, J.; Brunkan-Lamontagne, A.L.; Clarke, B.R.; Whitfield, C.; Naismith, J.H. Wza the Translocon for E. Coli Capsular Polysaccharides Defines a New Class of Membrane Protein. Nature 2006, 444, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Beis, K.; Collins, R.F.; Ford, R.C.; Kamis, A.B.; Whitfield, C.; Naismith, J.H. Three-Dimensional Structure of Wza, the Protein Required for Translocation of Group 1 Capsular Polysaccharide across the Outer Membrane of Escherichia Coli. J Biol Chem 2004, 279, 28227–28232. [Google Scholar] [CrossRef]
- Stevenson, G.; Andrianopoulos, K.; Hobbs, M.; Reeves, P.R. Organization of the Escherichia Coli K-12 Gene Cluster Responsible for Production of the Extracellular Polysaccharide Colanic Acid. J Bacteriol 1996, 178, 4885–4893. [Google Scholar] [CrossRef] [PubMed]
- Caffalette, C.A.; Zimmer, J. Cryo-EM Structure of the Full-Length WzmWzt ABC Transporter Required for Lipid-Linked O Antigen Transport. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef]
- Burroughs, A.M.; Balaji, S.; Iyer, L.M.; Aravind, L. A Novel Superfamily Containing the β-Grasp Fold Involved in Binding Diverse Soluble Ligands. Biology Direct 2007, 2, 4. [Google Scholar] [CrossRef]
- Jansen, K.U.; Gruber, W.C.; Simon, R.; Wassil, J.; Anderson, A.S. The Impact of Human Vaccines on Bacterial Antimicrobial Resistance. A Review. Environ Chem Lett 2021, 19, 4031–4062. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the cell envelope of a Gram-negative bacterium. The colored ovals (gray, lilac, purple, red) represent membrane proteins that are inserted into bacterial membranes. The green circles represent a soluble protein that binds to a membrane protein. The blue rounded rectangles ending in blue lines represent the pili. The chains of multi-colored circles attached to the outer membrane by a black line represent the lipopolysaccharides.
Figure 1.
Schematic representation of the cell envelope of a Gram-negative bacterium. The colored ovals (gray, lilac, purple, red) represent membrane proteins that are inserted into bacterial membranes. The green circles represent a soluble protein that binds to a membrane protein. The blue rounded rectangles ending in blue lines represent the pili. The chains of multi-colored circles attached to the outer membrane by a black line represent the lipopolysaccharides.
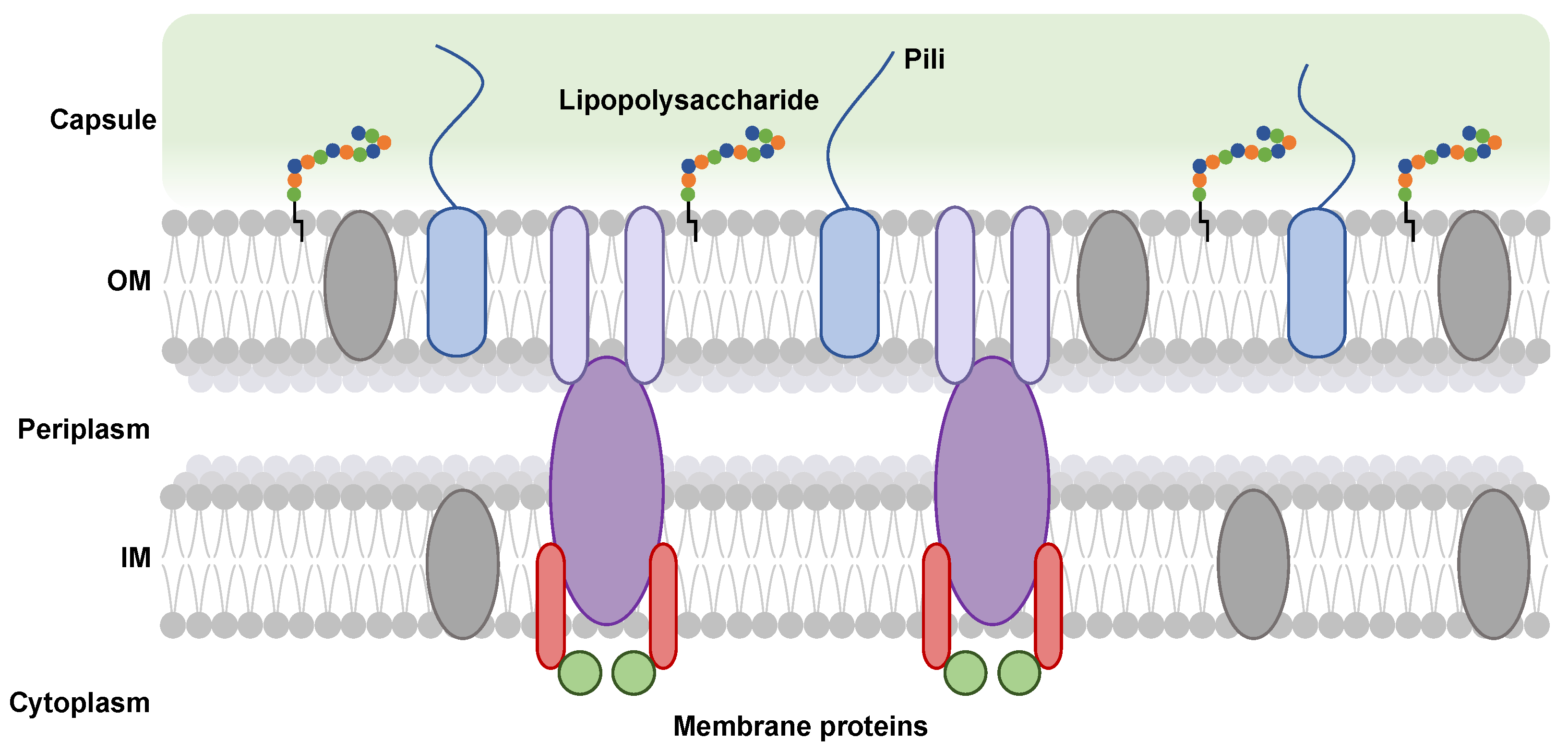
Figure 2.
Gene organization involved in the biosynthesis, polymerization and export of capsular polysaccharides in Gram-negative pathogenic bacteria. NBD: Nucleotide Bing Domain, TMD: TransMembrane Domain, PCP: Polysaccharide Copolymerase Protein, OPX: Outer membrane Polysaccharide eXport protein, β-Kdo: β-Kdo-transferase.
Figure 2.
Gene organization involved in the biosynthesis, polymerization and export of capsular polysaccharides in Gram-negative pathogenic bacteria. NBD: Nucleotide Bing Domain, TMD: TransMembrane Domain, PCP: Polysaccharide Copolymerase Protein, OPX: Outer membrane Polysaccharide eXport protein, β-Kdo: β-Kdo-transferase.
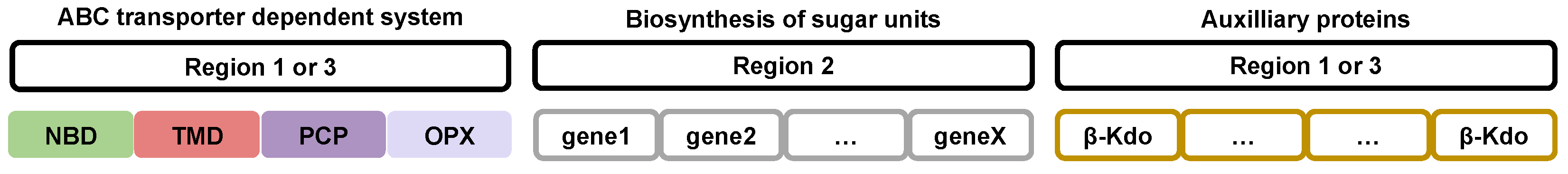
Figure 4.
Polysaccharide export pathways. TPR: TetratricoPeptide Repeat protein, GT: GlycosylTransferase, Wzx: flippase, Wzy: polymerase, PCP: Polysaccharide Co-Polymerase protein, OPX: Outer membrane polysaccharide export protein, TMD: TransMembrane Domain protein, NBD: Nucleotide-Binding Domain protein. Sugar residues are represented by different chains of colored circles and the lipid anchor is represented by a black line attached to these chains. Arrows represent the trajectory of the polysaccharide chain in the complex.
Figure 4.
Polysaccharide export pathways. TPR: TetratricoPeptide Repeat protein, GT: GlycosylTransferase, Wzx: flippase, Wzy: polymerase, PCP: Polysaccharide Co-Polymerase protein, OPX: Outer membrane polysaccharide export protein, TMD: TransMembrane Domain protein, NBD: Nucleotide-Binding Domain protein. Sugar residues are represented by different chains of colored circles and the lipid anchor is represented by a black line attached to these chains. Arrows represent the trajectory of the polysaccharide chain in the complex.
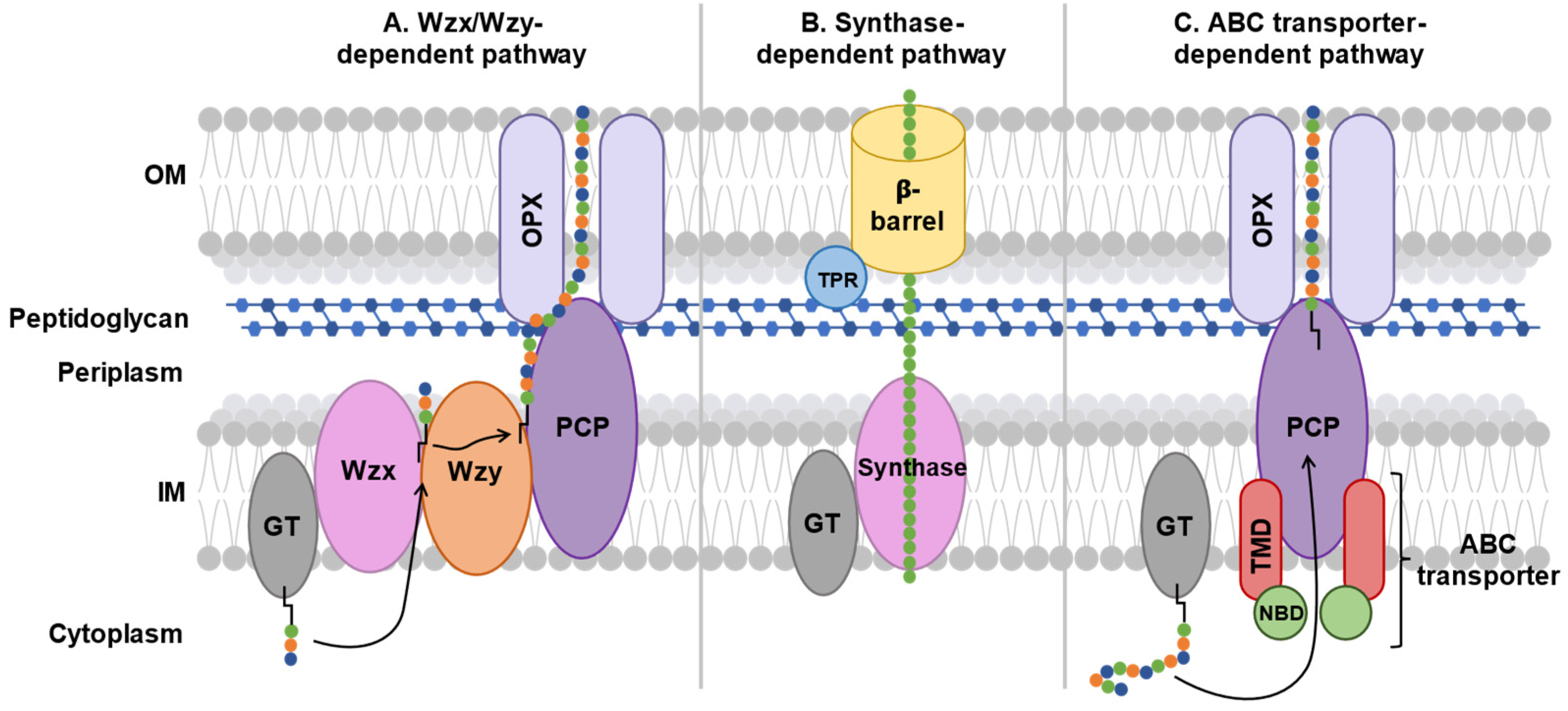
Figure 5.
Five KpsMT structures with different ligands: (A) in Apo form (PDB Code: 8TSW), (B) with two ATP molecules (PDB Code: 8TSH) then (C) hydrolyzed into two ADP molecules (PDB Code: 8TSI), (D) with lipid anchor in two different orientations (+Kdo-PG, PDB Codes: 8TUN and 8TT3). The space between the KpsT subunits (NBD, in green cartoon) varies depending on the type of substrate linked in the KpsT subunits (ATP represented in magenta and ADP spheres represented in pink spheres) and according to the orientations of the lipid anchor (turquoise spheres) in the pocket of the KpsM subunit (TMD, represented on the red surface).
Figure 5.
Five KpsMT structures with different ligands: (A) in Apo form (PDB Code: 8TSW), (B) with two ATP molecules (PDB Code: 8TSH) then (C) hydrolyzed into two ADP molecules (PDB Code: 8TSI), (D) with lipid anchor in two different orientations (+Kdo-PG, PDB Codes: 8TUN and 8TT3). The space between the KpsT subunits (NBD, in green cartoon) varies depending on the type of substrate linked in the KpsT subunits (ATP represented in magenta and ADP spheres represented in pink spheres) and according to the orientations of the lipid anchor (turquoise spheres) in the pocket of the KpsM subunit (TMD, represented on the red surface).
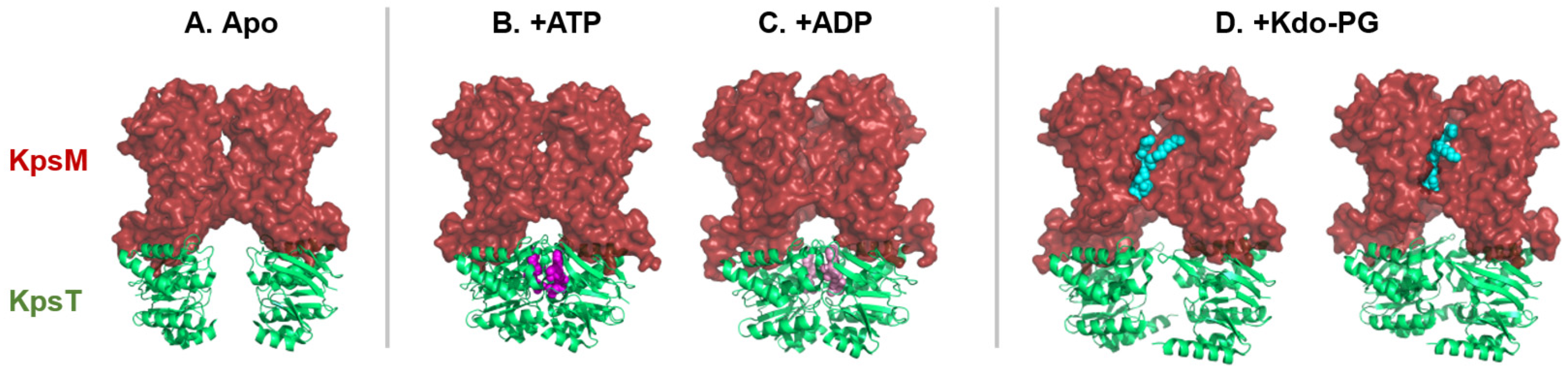
Figure 6.
Cartoon representation of the PCP-1, WzzE from E. coli (PDB Code: 6RBG; orange), PCP-2, Wzc from E. coli (PDB Code: 7NHR; green) and PCP- 3, truncated KpsE from Caldimonas thermodepolymerans (PDB code: 8TSL; purple).
Figure 6.
Cartoon representation of the PCP-1, WzzE from E. coli (PDB Code: 6RBG; orange), PCP-2, Wzc from E. coli (PDB Code: 7NHR; green) and PCP- 3, truncated KpsE from Caldimonas thermodepolymerans (PDB code: 8TSL; purple).
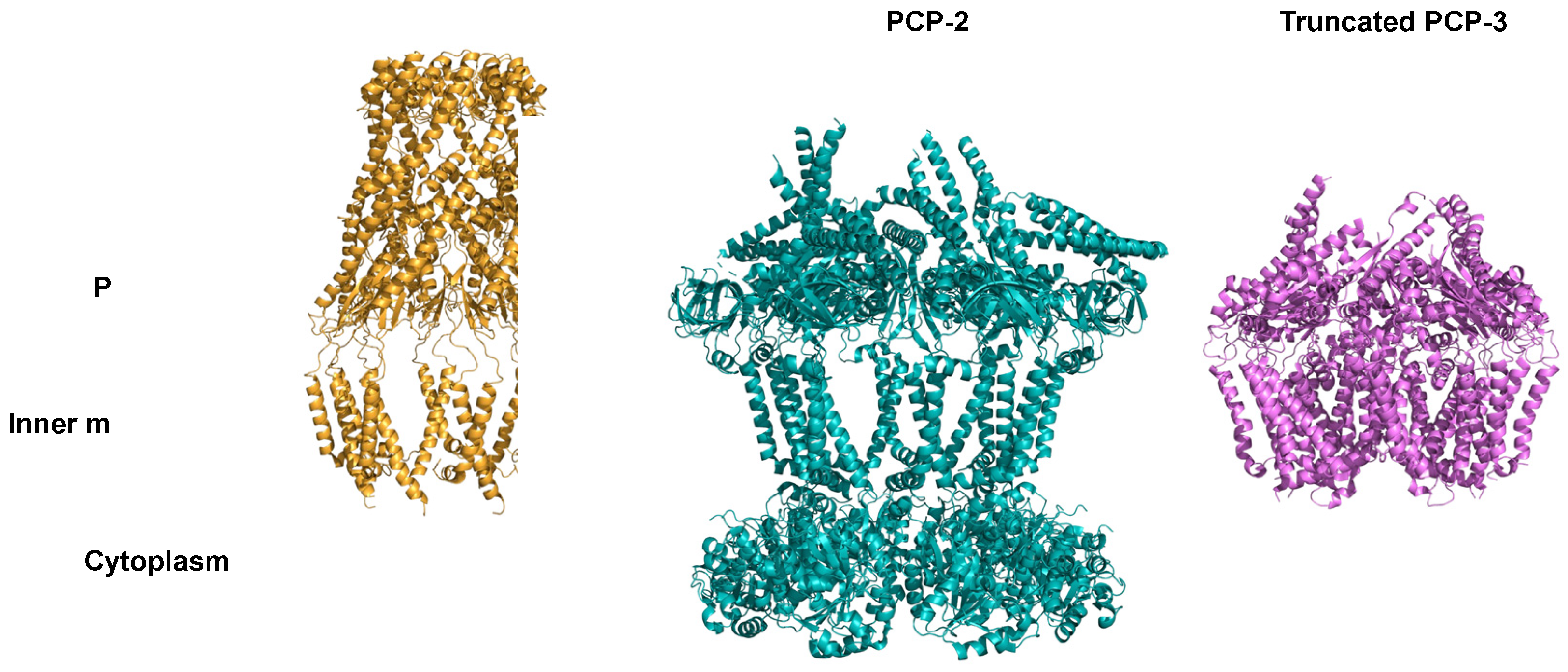
Figure 7.
Cartoon representation of the side view of the OPX from E. coli, Wza (PDB code: 2J58). Each domain of the protein is separated by black dotted lines. The channel is represented by a grey surface.
Figure 7.
Cartoon representation of the side view of the OPX from E. coli, Wza (PDB code: 2J58). Each domain of the protein is separated by black dotted lines. The channel is represented by a grey surface.
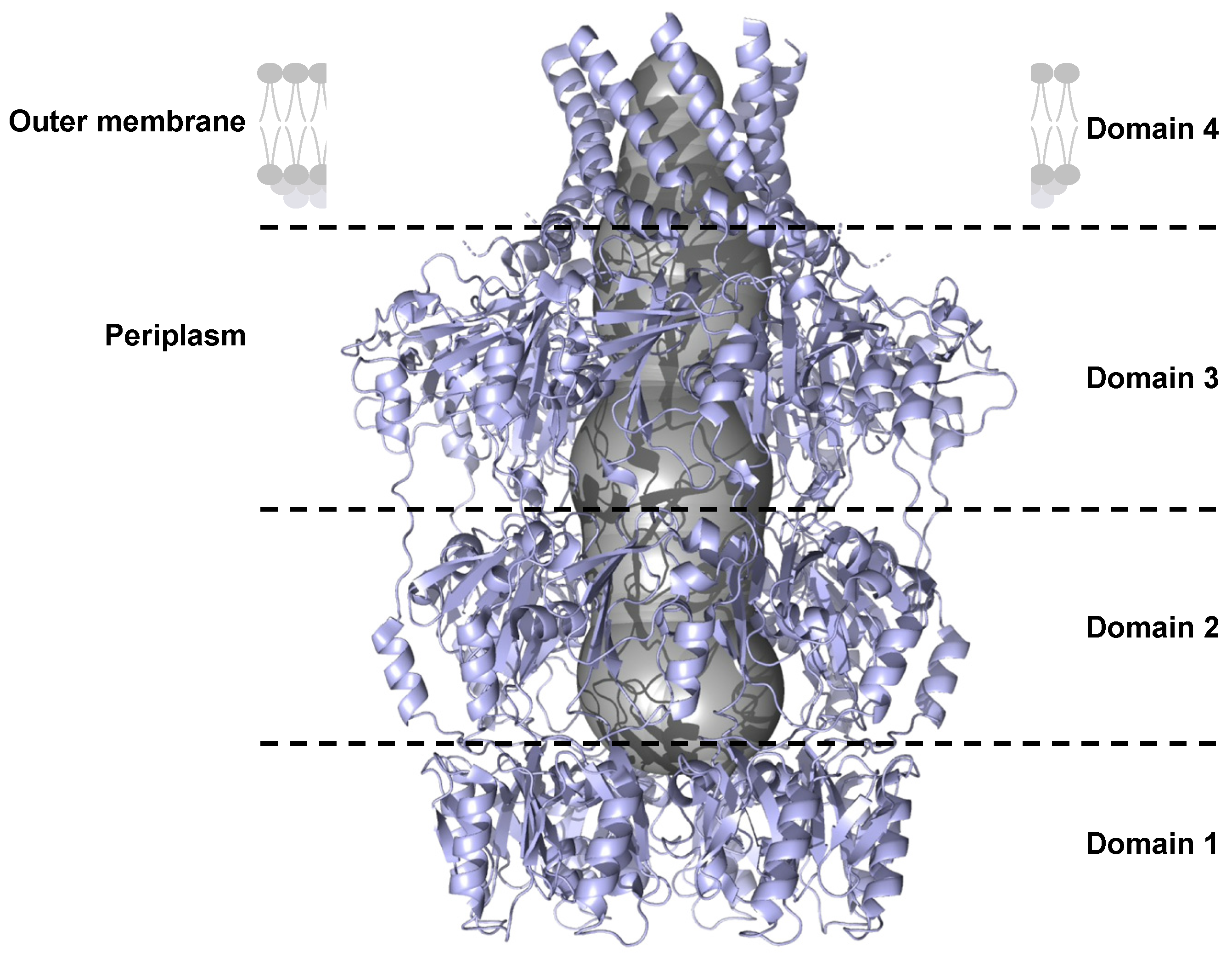
Figure 8.
Potential export mechanism of capsular polysaccharides by an ABC transporter-dependent system. NBDs are represented by green circles, TMDs by red ovals, PCP by a purple oval, and OPX by lilac ovals. The capsular polysaccharide is composed of a lipid anchor represented by a black line and sugars represented by green, orange, and blue colored circles.
Figure 8.
Potential export mechanism of capsular polysaccharides by an ABC transporter-dependent system. NBDs are represented by green circles, TMDs by red ovals, PCP by a purple oval, and OPX by lilac ovals. The capsular polysaccharide is composed of a lipid anchor represented by a black line and sugars represented by green, orange, and blue colored circles.
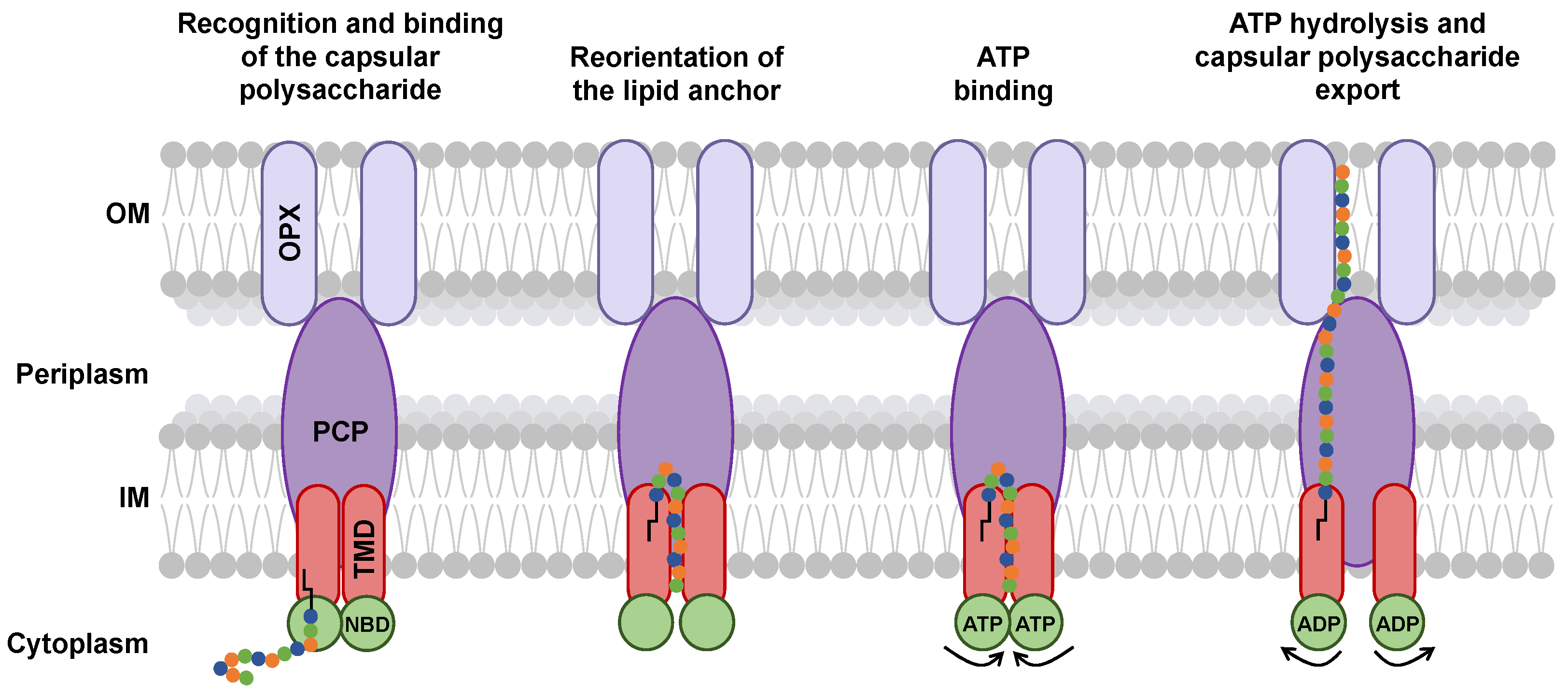
Table 2.
Structure examples for each capsular polysaccharide group.
| Groups | E. coli serotypes | Polysaccharide structures |
| Group 1 | K27 | 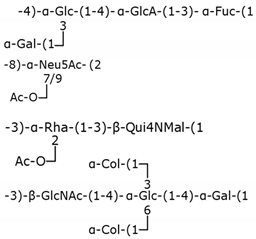 |
| Group 2 | K1 | |
| Group 3 | K10 | |
| Group 4 | O111 |
Glc: glucose, GlcA: glucuronic acid, Fuc: fucose, Gal: galactose, Neu5Ac: N-acetylneuraminic acid, Ac: acetyl group, Rha: rhamnose, Qui4NMal: 4-(2-carboxyacetamido)-4,6-dideoxyglucose, Col: colitose (3,6-dideoxygalactose), GlcNAc: N-acetylglucosamine.
Table 3.
Names of proteins in ABC transporter-dependent systems across different Gram-negative pathogenic bacteria.
Table 3.
Names of proteins in ABC transporter-dependent systems across different Gram-negative pathogenic bacteria.
| Bacterium | ABC transporter | PCP | OPX |
| Escherichia coli | KpsM/KpsT | KpsE | KpsD |
| Neisseria meningitidis | CtrC/CtrD | CtrB | CtrA |
| Haemophilus influenzae | BexB/BexA | BexC | BexD |
| Salmonella enterica serovar Typhi | VexB/VexC | VexD | VexA |
| Campylobacter jejuni | KpsM/KpsT | KpsE | KpsD |
| Pasteurella multocida | KpsM/KpsT | KpsE | KpsD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



