Submitted:
31 March 2025
Posted:
31 March 2025
You are already at the latest version
Abstract
Curcumin, a major phytochemical derived from Curcuma longa, has been shown to enhance the efficacy of chemotherapeutic agents such as doxorubicin, 5-fluorouracil, and cisplatin by overcoming drug resistance, making it a promising adjunct in the treatment of glioblastoma. However, the global gene expression changes triggered by curcumin in glioblastoma remain underexplored. In this study, we investigated the effects of curcumin on human glioblastoma (U87 MG) cells, where it significantly reduced cell viability and proliferation in a dose- and time-dependent manner and induced apoptosis without affecting senescence. Transcriptomic analysis revealed 5,036 differentially expressed genes, with pathway enrichment identifying 13 dysregulated cancer-associated pathways. Notably, curcumin modulated several key regulators involved in MAPK, Ras, TGF-β, Wnt, Cytokine, and TNF signalling pathways. Several apoptosis and cell cycle-associated genes, including PRKCG, GDF7, GDF9, GDF15, GDF5, FZD1, FZD2, FZD8, AIFM3, TP53AIP1, CRD14, NIBAN3, BOK, BCL2L10, BCL2L14, BNIPL, FASLG, GZMM, TNFSF10, TNFSF11, and TNFSF4, were significantly altered. Importantly, RUNX3, a key tumour suppressor, was markedly upregulated following curcumin treatment, emphasizing its potential role in curcumin-mediated anti-tumour effects. This study provides insight into the molecular mechanisms underlying curcumin's action against glioblastoma.
Keywords:
glioblastoma-U87 cells
; natural compound
; curcumin
; apoptosis
; RNAseq
Introduction
Glioblastoma multiforme is the most common primary malignancy of the brain, with an incidence of 3.2 in 100,000 [1]. Glioblastoma remains a formidable challenge, with a 5-year survival rate below 10% despite aggressive treatment. [2]. The hallmarks of glioblastoma include aggressive and diffuse proliferation, and resistance to apoptosis-inducing drugs [3]. Tumor drug resistance is a major cause of treatment failure, driven by cellular heterogeneity, diverse molecular signatures, and variable drug responsiveness [4]. Conventional chemotherapy fails to achieve complete remission and induces toxicity, harming normal cells and causing severe side effects [5].
Plant-derived compounds, known for their pleiotropic anti-cancer effects, have gained interest in overcoming multi-drug resistance [6,7]. Natural products like resveratrol, quercetin, EGCG, and curcumin enhance chemotherapy by sensitizing tumor cells through antioxidant, anti-inflammatory, immune-modulatory, and apoptosis-inducing mechanisms, boosting efficacy without added toxicity [8].
Curcumin, a yellow pigment from Curcuma longa, is a well-studied anti-cancer agent with low toxicity and therapeutic potential [9]. Preclinical studies show that curcumin enhances the efficacy of chemotherapeutic drugs like sulfinosine, 5-FU, doxorubicin, oxaliplatin, and cisplatin, often sensitizing drug-resistant cells [10]. In prostate cancer, curcumin with docetaxel improved anti-proliferative and apoptotic effects[11]. It also boosted 5-FU efficacy in colon, breast, and gastric cancers [12,13]. In a pancreatic cancer xenograft, curcumin with gemcitabine significantly reduced tumor volume by enhancing anti-proliferative and anti-angiogenic pathways[14]. Clinical trials suggest daily curcumin supplementation exerts anti-inflammatory and immune-modulatory effects in various cancers [15].
Curcumin modulates key pathways in tumor initiation, promotion, and progression. It suppresses carcinogenesis by inducing apoptosis, arresting the cell cycle, and inhibiting metastasis, invasion, and angiogenesis [16]. Mechanistically, curcumin regulates genes involved in oncogenic pathways like MAPK/ERK, MMPs, Wnt, STAT3, NF-κB, PI3K/Akt, and mTOR, which cancer cells exploit for growth and survival [17,18,19,20,21].
Although extensive research has revealed curcumin’s anti-carcinogenic properties and its potential to support cancer therapy, studies examining its impact on global gene expression via transcriptome profiling in various cancer cells remain limited. High-throughput RNA sequencing (RNA-seq) of curcumin-treated cells can elucidate the anti-proliferative and cell death pathways associated with curcumin [22]. Recent studies have employed RNA-seq to profile the most significantly up- and down-regulated genes following curcumin treatment, followed by protein–protein interaction analysis and in silico molecular docking to identify novel curcumin targets [23]. Compared to targeted proteomic methods, integrating global pathway enrichment analyses with systemic studies yields a more comprehensive insight into the regulatory and functional dynamics driving curcumin-induced cytotoxicity in cancer, thereby enhancing its therapeutic application [24]. In two breast cancer cell lines, transcriptome profiling showed that curcumin primarily induced cell death via ferroptosis rather than apoptosis, suggesting that ferroptosis may be a more promising therapeutic target [25]. In contrast, in adrenocortical carcinoma cells and their xenograft model, curcumin induced apoptosis mainly through ER stress pathways with accompanying upregulation of p38 and JNK/MAPK signalling [24].
Given the limited number of transcriptome profiling studies on curcumin, further research could enhance our understanding of its cytotoxic mechanisms in cancer [25]. Global gene expression analyses can identify novel genetic targets of curcumin, guiding future research efforts [26]. Accordingly, this study evaluated the impact of curcumin on global gene expression in the U87-MG human glioblastoma cell line using RNAseq, with its cytotoxic effects confirmed via CCK-8 proliferation, apoptosis, and senescence assays.
Materials and Methods
Curcumin
Curcumin (95% purity) [(E, E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione] was obtained from Sigma-Aldrich (St. Louis, MO, USA) and stored as a 5 mM stock solution in ethanol at -20°C, protected from light.
Cell culture
The human glioblastoma cell line (U87 MG) was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were cultured in T75 or T175 flasks (Greiner, Monroe, NC, USA) using high-glucose Dulbecco’s Modified Eagle Medium (DMEM) (ATCC, Manassas, VA, USA) supplemented with 10% fetal bovine serum (Atlas Biologicals, Fort Collins, CO, USA) and 1X antibacterial/antimycotic solution (Gibco, Grand Island, NY, USA). Cultures were maintained in a humidified incubator at 37°C with a gas mixture of 5% CO₂ and 95% air.
Cell Viability Assay
The impact of increasing curcumin concentrations on the viability of U87 MG cells was evaluated. A total of 5 × 10³ cells were seeded per well in a 96-well microplate with 100 μL of media and incubated overnight. Cells were then treated with curcumin at concentrations of 10 μM, 20 μM, 30 μM, 40 μM, and 50 μM, along with ethanol as the vehicle control, for 24, 48, and 72 hours. Cell viability was assessed using the Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan) assay. Briefly, after treatment, 10 μL of CCK-8 solution was added to each well containing 100 μL of treatment media, followed by a 4-hour incubation. Optical density was then measured at 450 nm using the SpectraMax iD3 Multi-Mode Microplate Reader (Molecular Devices, San Jose, CA, USA). The cytotoxicity of curcumin was determined by calculating the IC50 from the dose-response curve obtained through the cell viability assay at 24 hours.
Poly-Caspase Assay
To examine curcumin-induced apoptosis in U87 MG cells, caspase enzyme activity was measured using the Poly Caspase Assay (FAM FLICA, ImmunoChemistry Technologies, Davis, CA, USA). Cells were cultured in T25 flasks with phenol red-free Dulbecco’s Modified Eagle’s Medium (Gibco, Grand Island, NY, USA) supplemented with 10% FBS and 1× antibiotic/antimycotic solution. Once cells reached 80% confluence, they were treated with ethanol (vehicle control), 10 μM, or 20 μM curcumin. Staurosporine (5 µM) (ImmunoChemistry Technologies, Bloomington, MN, USA) served as a positive control. After 24 hours of treatment, cells were collected, stained with FLICA, and incubated for one hour at 37°C. Following washing, a 100 µL aliquot of the cell suspension, at a density of greater than 2 × 10⁶ cells/mL, was transferred to a 96-well flat-bottom microplate. Fluorescence was measured at an excitation wavelength of 488 nm and an emission wavelength of 520 nm using a microplate reader.
SA-Beta Galactosidase Assay
To determine whether curcumin treatment induces or inhibits senescence, β-galactosidase activity—a pH-dependent enzyme marker of senescent cells—was measured using the SA-β-galactosidase assay (Cell Biolabs, San Diego, CA, USA). Cells were cultured in 6-well plates with phenol red-free Dulbecco’s Modified Eagle’s Medium supplemented with 10% FBS and 1× antibiotic/antimycotic solution. Once cells reached 80% confluence, they were treated with ethanol (vehicle control), 10 μM, or 20 μM curcumin. After 24 hours of treatment, cells were collected, stained, and transferred to 96-well flat black-bottom microplates. Fluorescence (optical density) was measured at 360/465 nm (excitation/emission) using a microplate reader, and the results were expressed as Relative Fluorescence Units (RFU).
RNA Isolation, Library Preparation
The effect of curcumin treatment on global gene expression in U87-MG cells was assessed using RNA-seq. Three replicates each of two experimental controls were set up: negative control (vehicle treated cells) and treated cells. Cells were cultured in 60mm x 15mm cell culture dishes (Wuxi NEST Biotechnology Co., Ltd., Jiagnsu, China) until 80-90% confluence and treated with 20 μM curcumin. After incubation for 6 hours, RNA was isolated using the RNeasy Plus Mini Kit (QIAGEN, Germantown, MD, USA) and stored at -20°C until sequencing. Further library preparation we followed protocol [6].
RNA-Seq Analysis
Sequencing reads were processed by trimming adapters and removing low-quality bases (Phred score < 30) using Trimmomatic v0.39 [27]. The filtered reads were then aligned to the human reference genome (GRCh38.p13) using the STAR RNA-Seq aligner v2.7.11a [28], generating BAM alignment files. Read quantification was performed with the HTSeq R package [29] using genome annotations in GFF format. Differential gene expression (DEG) analysis was conducted using DESeq2 [30], considering genes with a log2FoldChange ≥1 and a false discovery rate (FDR) of ≤0.05. Pathway enrichment analysis was carried out using KOBAS [31].
Statistical Analysis
All statistical analyses were conducted using GraphPad Prism v10.4.0.621 for Windows (GraphPad Software, Boston, MA, USA, www.graphpad.com). A p-value of <0.05 was considered statistically significant.
Results
Curcumin Treatment Decreases U87 MG Cell Viability and Proliferation
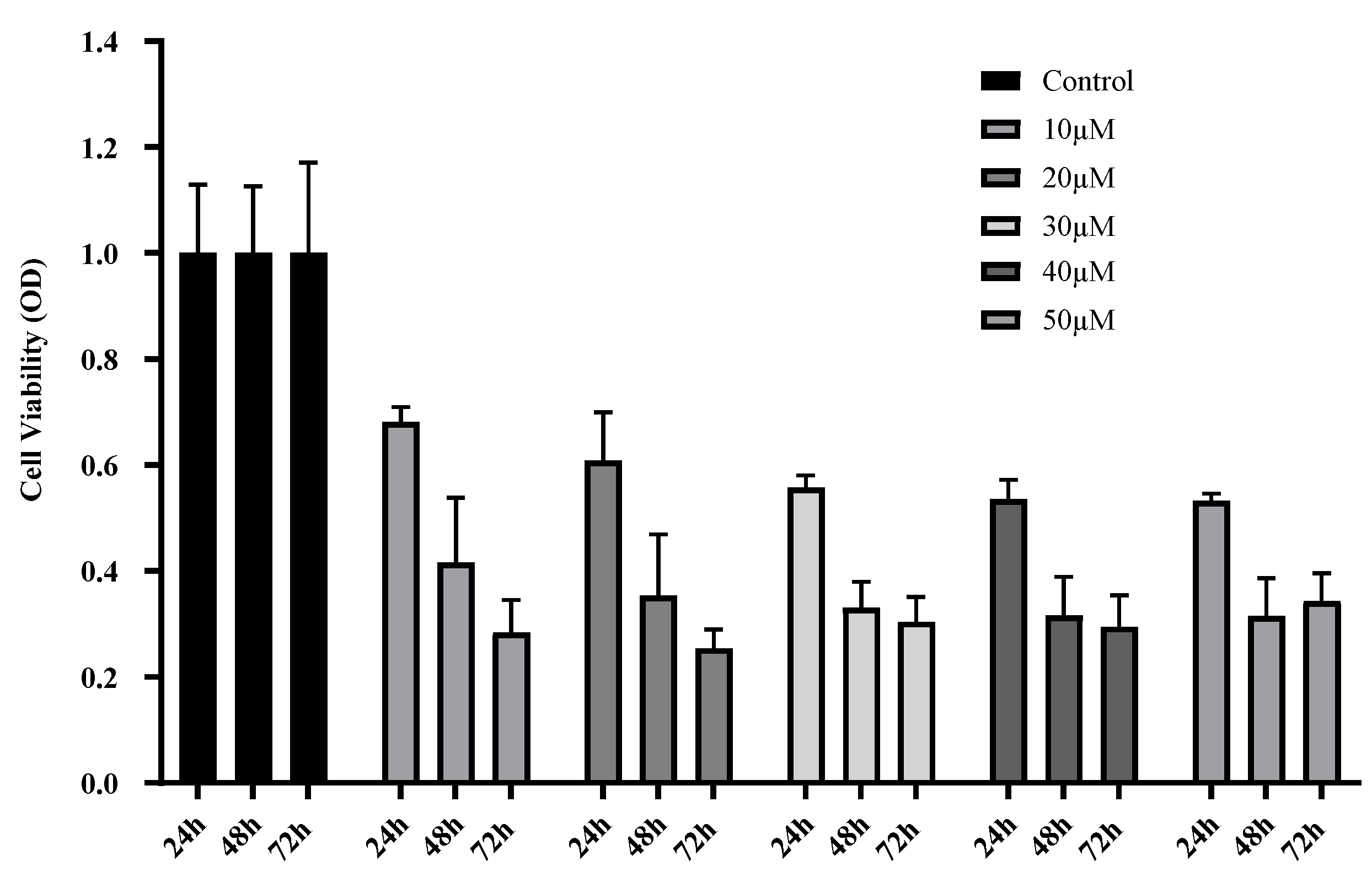
The growth-inhibitory effects of curcumin on human glioblastoma cells were evaluated. Cell viability was assessed using the CCK-8 assay. Results demonstrated that curcumin reduced the viability and proliferation of U87 MG cells in a dose- and time-dependent manner (Figure 1). The median IC50 for curcumin was determined to be 15 µM. Based on this, subsequent experiments were conducted using treatment concentrations of 10 µM and 20 µM.
Figure 1.
Effect of curcumin on glioblastoma cells. The anti-proliferative effects of increasing concentrations of curcumin on viability of U87 MG cells was assessed over a three-day period using the CCK-8 assay. Vehicle treated cells were used as negative control at each interval (100% viability). The viability is presented relative to the negative control. Values are means ± S.D. of three replicates.
Figure 1.
Effect of curcumin on glioblastoma cells. The anti-proliferative effects of increasing concentrations of curcumin on viability of U87 MG cells was assessed over a three-day period using the CCK-8 assay. Vehicle treated cells were used as negative control at each interval (100% viability). The viability is presented relative to the negative control. Values are means ± S.D. of three replicates.
Curcumin Reduced Migration of U87 MG Cells
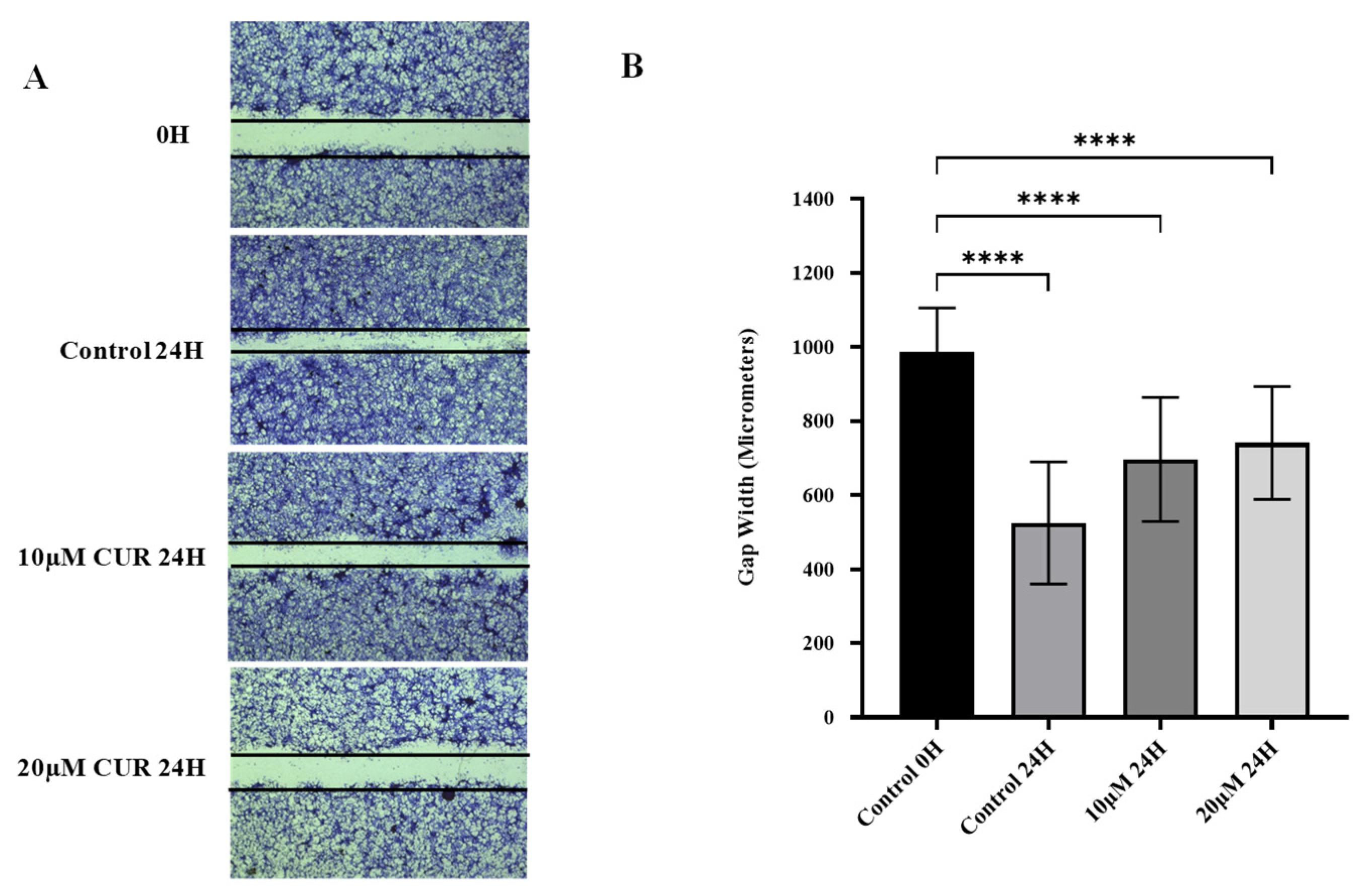
The effects of curcumin treatment on the migratory ability of U87 MG cells were assessed using the scratch wound healing assay. The potential for cells to migrate into the wound in vitro represents their metastatic ability in vivo. A scratch wound was made in the monolayer of U87 MG cells using a 20µL pipette tip, and cells were treated with curcumin or vehicle. Quantitatively, the inhibition of migration was assessed by measuring gap closure after 24 hours of treatment with curcumin or vehicle.
Treatment of the cells with 10 μM and 20 µM curcumin reduced the number of cells that migrated into the wound after 24 hours significantly compared to vehicle-treated cells (p-value <0.0001) (Figure 2). Furthermore, the rate of gap closure was reduced when the curcumin concentration was increased from 10 μM to 20 μM (p-value < 0.01). The results of this study demonstrate that curcumin exhibits anti-migratory effects in U87 MG cells, and this effect is dose-dependent.
Figure 2.
Effect of curcumin on migration of U87 MG cells. (A) The cell monolayer was scratched with a 20 pipette tip, photographed at 0 hours, and treated for 24 hours. (B) Gap closure was quantified by comparing gap width at 0H and 24H post treatment with vehicle or curcumin. Data are mean gap width of six replicates ± SD. One-Way ANOVA was used to examine statistical differences followed by Tukey’s Honestly Significant Test for post hoc testing for differences between treatment groups. ****Significant at p < 0.0001.
Figure 2.
Effect of curcumin on migration of U87 MG cells. (A) The cell monolayer was scratched with a 20 pipette tip, photographed at 0 hours, and treated for 24 hours. (B) Gap closure was quantified by comparing gap width at 0H and 24H post treatment with vehicle or curcumin. Data are mean gap width of six replicates ± SD. One-Way ANOVA was used to examine statistical differences followed by Tukey’s Honestly Significant Test for post hoc testing for differences between treatment groups. ****Significant at p < 0.0001.
Curcumin induced apoptosis but not senescence in U87 MG cells
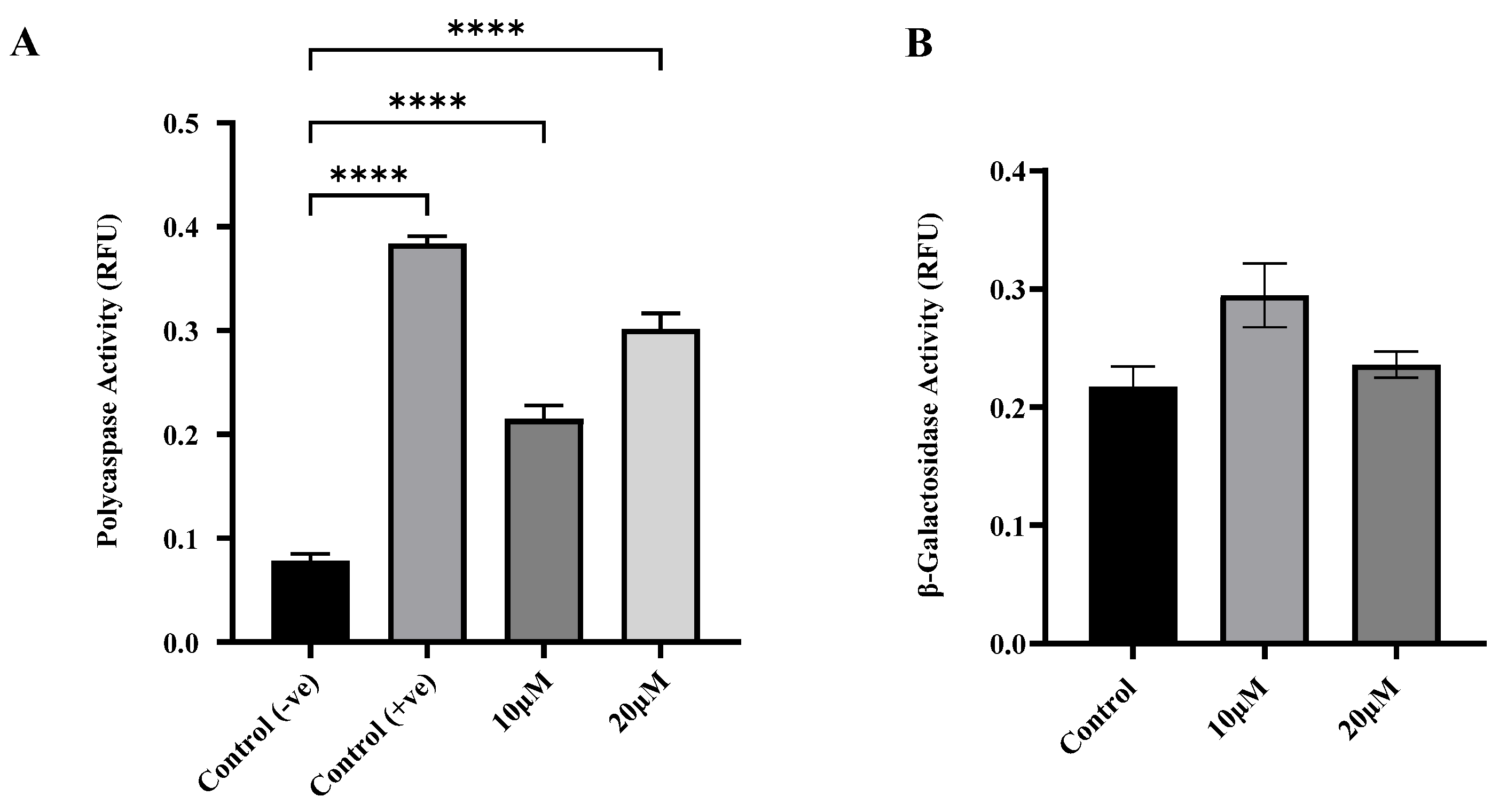
To investigate the effects of curcumin on apoptosis and senescence in human glioblastoma cells, curcumin was administered to cells and cells incubated for 24 hours. Following incubation, apoptotic cells were identified through poly-caspase activity using the FAM-FLICA poly-caspase assay. When compared to untreated cells, curcumin-treated cells had significantly higher poly-caspase activity (p-value < 0.0001) (Figure 3a). Increasing curcumin concentration from 10 to 20 also increased the rate of poly-caspase activity, indicating that curcumin-induced apoptosis in U87 MG cells was dose-dependent.
Senescent cells are identified through elevated lysosomal activity, which can be biochemically assayed through the activity of β-galactosidase enzyme [32]. In this study, curcumin-induced senescence was assayed using the SA-β-Galactosidase assay kit. Curcumin treatment did not significantly induce senescence in U87 MG cells (p > 0.05 when compared to control) (Figure 3b).
Figure 3.
Effects of Curcumin on Apoptosis and Senescence in U87MG Cells. A. Apoptosis induction was determined by polycaspase activation following curcumin treatment for 24h. B. Curcumin-induced senescence was measured using the SA-β-Galactosidase assay. Data are means ± SD of three independent replicates. One-way ANOVA tests were performed to determine the statistical significance of differences between control and treatment means. Tukey’s Honestly Significant Differences tests were used for post hoc analysis of multiple comparisons. ****Significant at P < 0.0001.
Figure 3.
Effects of Curcumin on Apoptosis and Senescence in U87MG Cells. A. Apoptosis induction was determined by polycaspase activation following curcumin treatment for 24h. B. Curcumin-induced senescence was measured using the SA-β-Galactosidase assay. Data are means ± SD of three independent replicates. One-way ANOVA tests were performed to determine the statistical significance of differences between control and treatment means. Tukey’s Honestly Significant Differences tests were used for post hoc analysis of multiple comparisons. ****Significant at P < 0.0001.
Curcumin Induced Differential Gene Expression in U87 MG cells
The impact of curcumin on global gene expression in U87 MG cells was analyzed using RNA sequencing (RNA-seq). Treatment with 20 µM curcumin resulted in the differential expression of 5,036 genes, with 3,418 genes upregulated and 1,618 downregulated. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the top 50 upregulated genes identified enrichment in pathways related to extracellular matrix (ECM)-receptor interaction, focal adhesion, cell adhesion molecules, Notch signaling, NF-κB signaling, Jak-STAT signaling, and choline metabolism in cancer (Figure 4a). Gene Ontology (GO) analysis revealed significant enrichment in molecular functions, including metal ion transmembrane transport, ion channel activity, channel activity, and gated channel activity (Figure 4b).
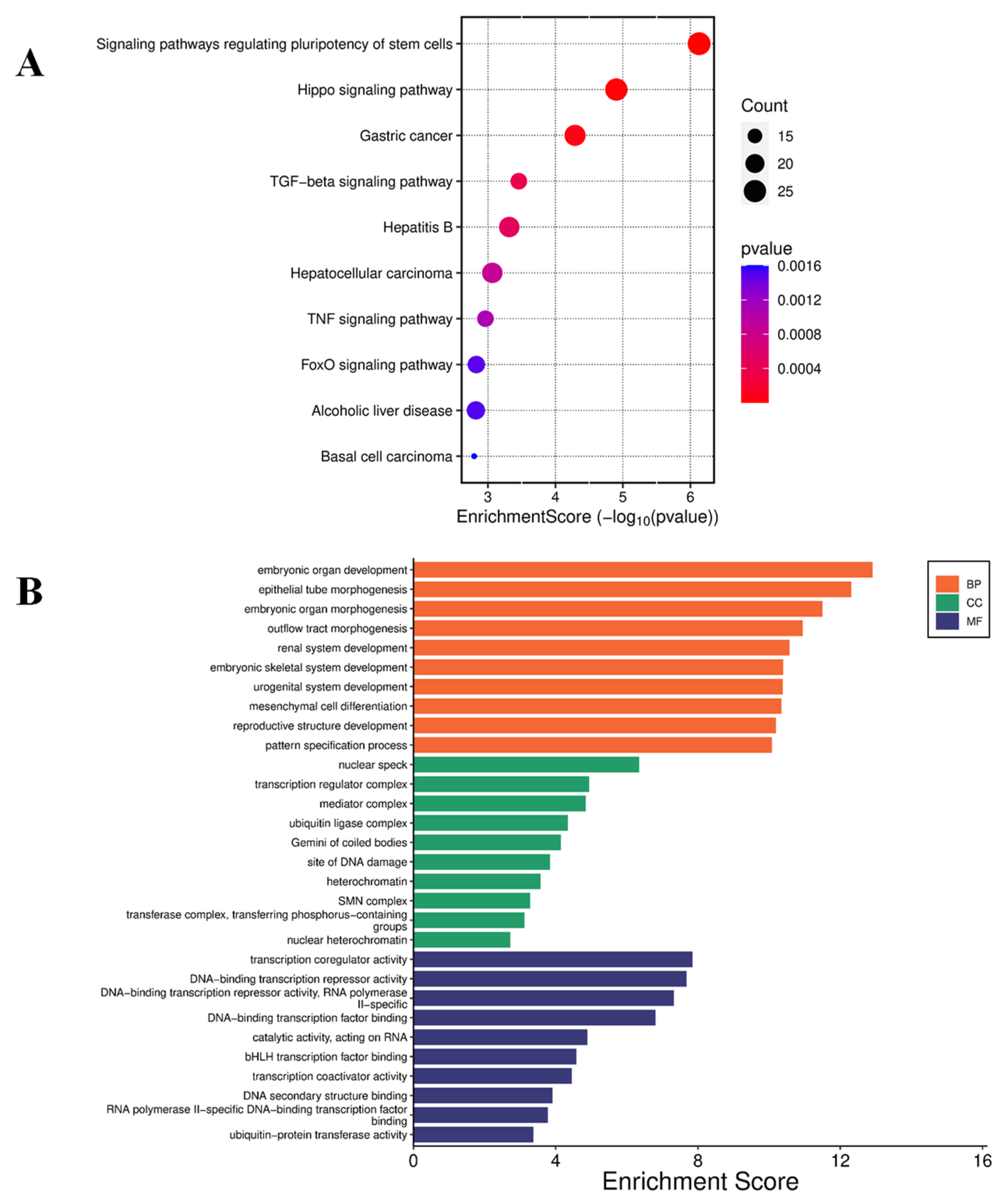
Conversely, KEGG pathway analysis of the top 50 downregulated genes revealed involvement in pathways including O-glycan biosynthesis, Notch signaling, glycine, serine, and threonine metabolism, glycolysis and gluconeogenesis, and cytokine-cytokine receptor interaction (Figure 5a). GO analysis further highlighted enrichment in molecular functions, including DNA-binding transcription repressor activity, DNA-binding transcription factor binding, transcription coactivator activity, and catalytic activity acting on RNA (Figure 5b).
Further analysis of the 50 most significantly altered KEGG pathways identified 13 pathways strongly linked to tumorigenesis, cancer progression, and immune evasion. These included cytokine-cytokine receptor interaction, pathways in cancer, MAPK signaling, regulation of stem cell pluripotency, TGF-β signaling, breast cancer, Ras signaling, IL-17 signaling, PD-L1 expression and PD-1 checkpoint in cancer, proteoglycans in cancer, Wnt signaling, gastric cancer, and viral protein interaction with cytokines and cytokine receptors. Among these, several tumor-suppressive pathways stood out. Genes within MAPK, Ras, TGF-β, Wnt, and cytokine-cytokine receptor signaling, such as PRKCG (+8.81), GDF7 (+7.78), GDF5 (-2.68), FZD1 (+2.62), FZD2 (+2.95), and FZD8 (-3.18), are hypothesized to contribute to curcumin’s anti-tumorigenic effects in this study. Additionally, the well-known tumor suppressor gene RUNX3 was significantly upregulated (+8.93) in curcumin-treated cells, further supporting its potential role in curcumin-mediated tumor suppression.
Figure 4.
Enrichment analysis of up-regulated genes. A. Bubble plot for KEGG pathway enrichment of the top 50 up-regulated genes. Each circle represents an enriched function. The gene enrichment ratio is assigned to the x-axis, and the descriptions of the pathways are assigned to the y-axis. The area of the circles is proportional to the number of genes assigned to the term, and the color to the adjusted P-value. B. GO of three ontologies enriched in curcumin-induced up-regulated genes. The width of each bar represents enrichment score of genes (-log10(P-value)). BP = Biological Process, CC = Cellular Component, MF = Molecular Function.
Figure 4.
Enrichment analysis of up-regulated genes. A. Bubble plot for KEGG pathway enrichment of the top 50 up-regulated genes. Each circle represents an enriched function. The gene enrichment ratio is assigned to the x-axis, and the descriptions of the pathways are assigned to the y-axis. The area of the circles is proportional to the number of genes assigned to the term, and the color to the adjusted P-value. B. GO of three ontologies enriched in curcumin-induced up-regulated genes. The width of each bar represents enrichment score of genes (-log10(P-value)). BP = Biological Process, CC = Cellular Component, MF = Molecular Function.
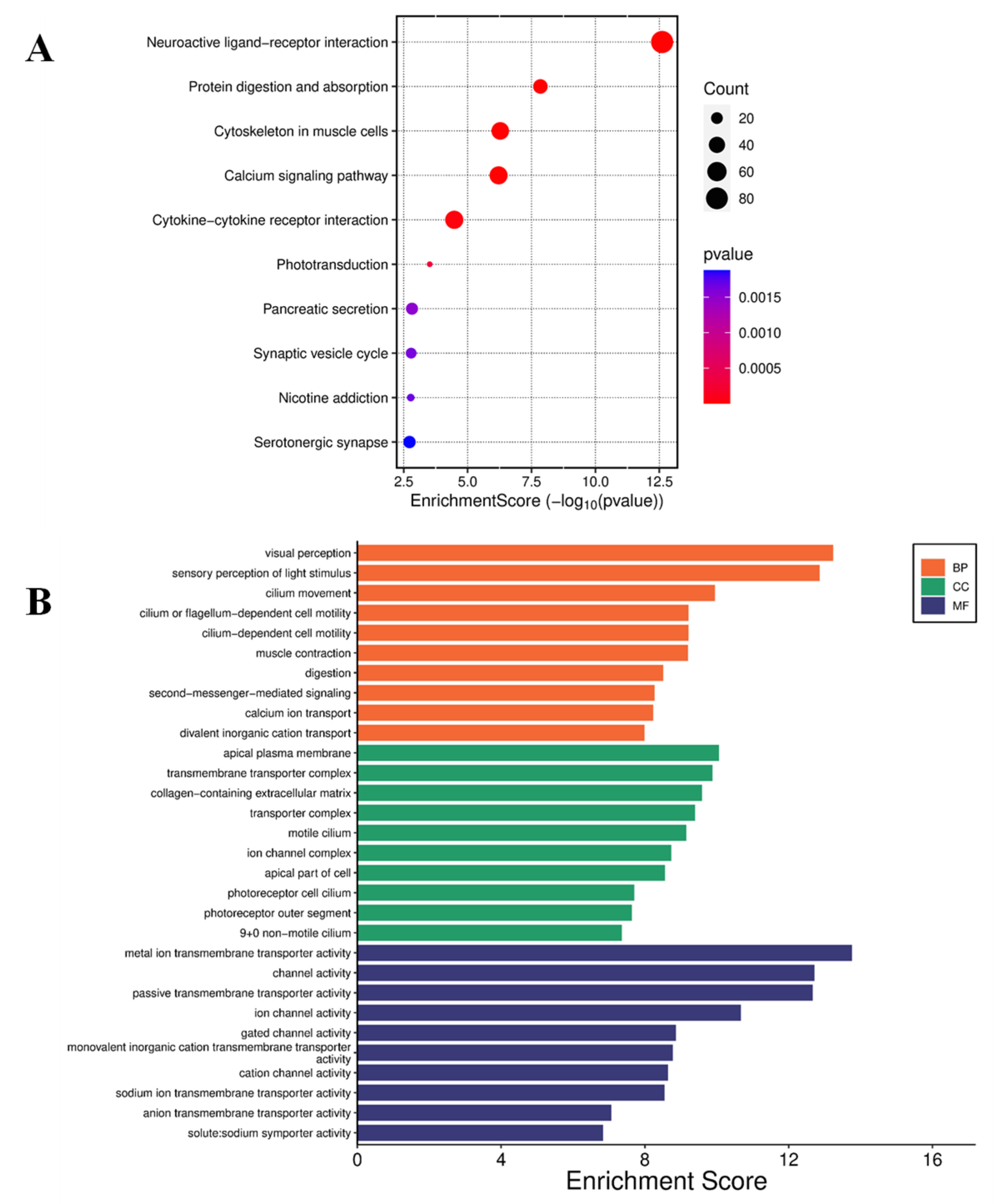
Figure 5.
Enrichment analysis of downregulated genes. A. Bubble plot for KEGG pathway enrichment of the top down-regulated genes. Each circle represents an enriched function. The gene enrichment ratio is assigned to the x-axis and the description of the pathways to the y-axis. The area of the circles is proportional to the number of genes assigned to the term, and the color to the adjusted P-value. B. GO of three ontologies enriched in curcumin-induced down-regulated genes. The width of each bar represents enrichment score of genes (-log10(P-value)). BP = Biological Process, CC = Cellular Component, MF = Molecular Function.
Figure 5.
Enrichment analysis of downregulated genes. A. Bubble plot for KEGG pathway enrichment of the top down-regulated genes. Each circle represents an enriched function. The gene enrichment ratio is assigned to the x-axis and the description of the pathways to the y-axis. The area of the circles is proportional to the number of genes assigned to the term, and the color to the adjusted P-value. B. GO of three ontologies enriched in curcumin-induced down-regulated genes. The width of each bar represents enrichment score of genes (-log10(P-value)). BP = Biological Process, CC = Cellular Component, MF = Molecular Function.
Discussion
Curcumin, a polyphenol from Curcuma longa, is a well-studied natural compound that modulates tumour initiation, progression, and promotion in various cancers [33]. In glioblastoma, curcumin suppresses oncogenic signalling, induces multimodal cell death, and modulates immune components within the tumour microenvironment (TME) [34]. This study examines the impact of curcumin on global gene expression and its effects on proliferation, migration, apoptosis, and senescence in U87MG glioblastoma cells.
Curcumin Suppresses Viability and Migration of U87 MG Cells
Curcumin inhibited U87 MG cell proliferation in a dose- and time-independent manner, aligning with previous studies. RNA sequencing suggests that curcumin suppresses malignancy by inhibiting oncogenic pathways, including Wnt/β-catenin, PI3K/Akt/mTOR, NF-κB, and TGF-β [35,36]. In our study also crucial pathways for cell survival PI3K-Akt, (CDK6 (-1.55) CSF3 (-1.55) LPAR1 (-1.46) EFNA5 (-1.13) PHLPP2(-1.17) PHLPP1 (-1.11) GHR (-1.12) IRS1 (-1.71) KDR (-1.16) KRAS (-1.65) MCL1 (-1.80) PDGFA (-1.59) PDGFRA (-1.66) PIK3R1 (-1.88) PRKAA1 (-1.09) BDNF (-1.85) SGK1 (-1.44 fold) SOS2 (-1.05) TLR2 (-1.03) TLR4 (-2.69) CCNE1 (-1.27) CCNE2 (-1.73)) Wnt, (FRAT2 (-1.12) FZD2 (-2.95) APC (-1.06) SMAD3 (-1.67) WNT5A (-1.82) WNT7B (-1.07) FZD5 (-1.99) FZD1 (-2.62) FZD7 (-1.93) FZD8 (-3.18) ROCK2 (-1.17) mTOR, (FZD2 IRS1 KRAS PIK3R1 PRKAA1 SGK1 SKP2 (-2.19) SOS2 WNT5A WNT7B FZD5 FZD1 FZD7 FZD8), TNF signaling, (TAB1 (-1.13) CEBPB (-1.5) MAPK14 (-1.49) JAG1 (-1.04) IL1B (-1.07) LIF (-1.49) PIK3R1 BCL3 (-1.14) TRAF5 (-1.59) RIPK1 (-1.68) FADD (-2.88) BAG4 (-1.04) and TGF-β signaling (RGMB (-1.99) ID1 (-1.86) ID2 SMAD3 (-1.67) SMAD5 (-1.03) PITX2 (-1.7) SMURF2 (-1.04) BMP2 (-1.62) SP1 (-1.7) TGFBR1 (-1.02) TGFBR2 (-1.98) GDF5 (-2.68) ACVR1B (-1.48) ACVR2A (-1.2) NOG(-3.61) pathways, downregulated which are essential for cancer cell survival, metastasis, and drug resistance. Additionally, curcumin induces G2/M cell cycle arrest by modulating cell cycle regulators, thereby curbing tumor growth both in vitro and in vivo [32, . Current study also cell cycle arrest related cyclin dependent kinase CDK6 (-1.56), CCNE1 (-1.27), CCNE2 (-1.73), KRAS (-1.65), PIK3R1 (-1.88) PDGFA (-1.59), KDR (-1.16) anti-angiogenic effects, TLR4 (-2.4), IRS1 (-1.71) Inflammation reduction. Targeting these pathways effectively induces apoptosis and prevents cancer progression.
Scratch wound assays revealed a significant (p < 0.0001) reduction in U87 MG cell migration following curcumin treatment, with greater inhibition observed at 20 µM (p < 0.05). Curcumin’s anti-migratory effects involve the PI3K/Akt, NF-κB, ERK, and JAK-STAT signaling pathways, which downregulate MMPs, VEGF, and ICAM-1 [18,37,38,39]. In glioblastoma, curcumin inhibited STAT3 phosphorylation, reducing MMP9, Snail, and Twist expression [40], and suppressed MMPs via MAPK inhibition [41]. A study by Abdullah Thani et al. reported decreased levels of MMP2, 9, 14, 15, 16, 17, 24, and 25, further hindering glioblastoma migration [42]. Given glioblastoma’s aggressive migration at diagnosis, these findings underscore the therapeutic potential of curcumin. In our study, JAK STAT (CSF3, GHR, LIF, MCL1, PDGFA, PDGFRA, PIK3R1, SOS2) and Nf-kB (TAB1, IL1B, TLR4, TRAF5, TRAF6, RIPK1) related genes were also down-regulated.
Curcumin Triggers Apoptosis in U87 MG Glioblastoma Cells_
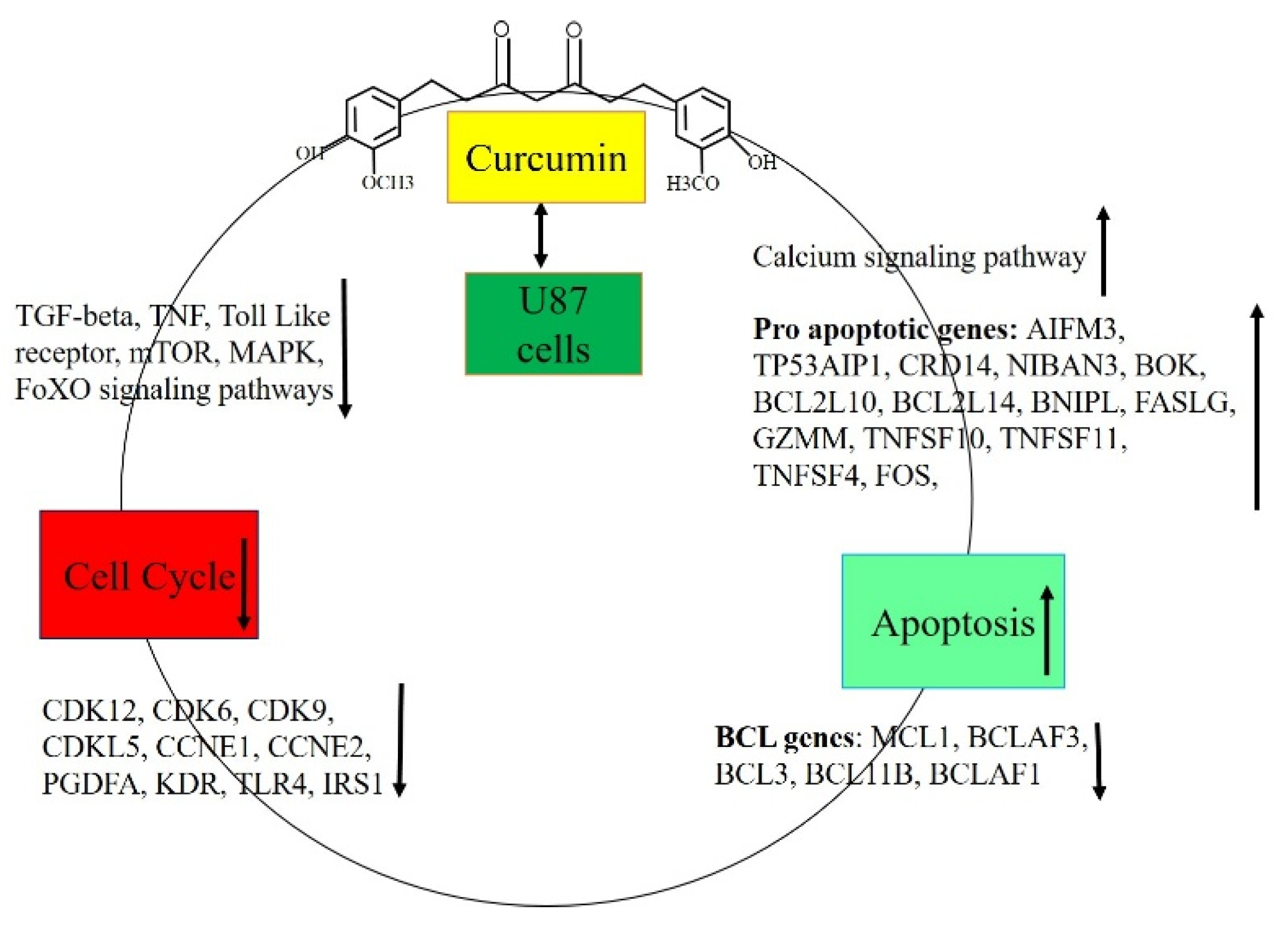
Resistance to apoptosis is a major challenge in glioblastoma treatment, making apoptosis-inducing compounds crucial [36]. Curcumin-induced apoptosis was assessed via poly-caspase activity using the FAM-FLICA probe, which irreversibly binds active caspases 3, 6, 7, 8, 9, and 10. After 24 hours, curcumin significantly increased poly-caspase activity, with higher activation at 20 µM compared to 10 µM, consistent with previous findings [43]. Curcumin promotes apoptosis by modulating apoptosis-related proteins, activating intrinsic (mitochondrial) (AIFM3 (5.3), BOK (2.5), BCL2L10 (, BCL2L14) and extrinsic (death-receptor) (FASLG (4.6), TNFSF10 (2.9), TNFSF4 (1.7) ) pathways, and mediating ER stress (TP53AIP1 (4.2), CRD14, NIBAN3 (3.8), BCL2L14, BNIPL, FASLG (4.6), GZMM, TNFSF11 (6.4)) [36,44,45,46,47] Figure 6. It also enhances cleaved caspase-3, -8, and -9 expression in glioblastoma cells [43,48,49].
Figure 6.
Thematic illustration. Curcumin treatment in U87 glioblastoma cells modulates global gene expression, leading to cell cycle arrest and apoptosis through the coordinated upregulation and downregulation of multiple signalling pathways and their associated genes.
Figure 6.
Thematic illustration. Curcumin treatment in U87 glioblastoma cells modulates global gene expression, leading to cell cycle arrest and apoptosis through the coordinated upregulation and downregulation of multiple signalling pathways and their associated genes.
The effect of curcumin on senescence in U87 MG cells was examined using β-galactosidase activity. While glioma senescence can both suppress and promote malignancy in vivo [32], curcumin (10–20 µM) did not induce senescence (p > 0.05). Prior studies suggest curcumin’s effects depend on concentration—low doses (≤10 µM) induce senescence without cell death, while higher doses trigger apoptosis due to excessive stress [50]. The lack of senescence in this study suggests cytotoxicity at these concentrations, as confirmed by poly-caspase and viability assays.
Curcumin treatment altered gene expression in U87 MG cells
Despite extensive research on curcumin’s role in glioblastoma, its impact on global gene expression remains underexplored. Given its pleiotropic effects in tumorigenesis, studying global expression could reveal novel genetic targets and enhance our understanding of its therapeutic potential. This study highlights the multifaceted role of curcumin in cancer therapy by modulating genes involved in growth, survival, and immune responses. The upregulation of genes like C1orf116 and uncharacterized loci (LOC124902513, LOC107985667, LOC100506358, LOC112268263, LOC105375676) suggests unexplored pathways in cancer regulation. METTL21C (10.29), a methyltransferase, enhances protein modifications related to cell cycle control and apoptosis, aligning with curcumin’s epigenetic modulation [51]. PNLDC1 (10.14) and AMPD1 (10.05-fold), involved in RNA processing and purine metabolism, may disrupt cancer cell homeostasis, impairing growth [52,53]. The upregulation of TBR1 (9.66), a transcription factor involved in neuronal development, suggests that curcumin may influence differentiation pathways—highly relevant in glioblastoma, where differentiation is often impaired [54].
A key finding in this study is the upregulation of RUNX3 (8.81), a runt-related transcription factor with tumor-suppressive roles in multiple cancers. RUNX3 is frequently downregulated in gastric, breast, colorectal, renal, and hepatocellular cancers through point mutations, promoter hypermethylation, and cytoplasmic translocation [55,56]. It also resides in a genomic region often deleted in various tumors, resembling classic tumor suppressor genes [57].
RUNX3 upregulation in curcumin-treated cells likely contributed to reduced viability, migration, and increased apoptosis, aligning with its known tumor-suppressive function in gliomas. Glioma patient studies indicate that RUNX3 is progressively inactivated through promoter hypermethylation, with reduced protein expression associated with disease progression [58]. Mei et al. reported lower RUNX3 expression in glioblastoma tissue compared to normal brain, with re-expression suppressing the migration of U251 and U87 MG cells via MMP2 inhibition [59]. Additionally, RUNX3 overexpression induced Bim-caspase-dependent apoptosis, G0/G1 cell cycle arrest, and suppressed Wnt/β-catenin signaling, thereby improving survival in a xenograft model [60,61].
Protein kinase C gamma (PRKCG), a gene identified in this study as involved in several signaling pathways, was upregulated by a fold change of 8.81 compared to control cells following curcumin treatment. Pathway analysis revealed that PRKCG could be involved in focal adhesion molecules, tyrosine kinase inhibition, and cancer-related pathways, as well as choline metabolism in cancer. PRKCG, is a member of the protein kinase c family of enzymes whose activation mediates various signaling pathways that regulate the balance between cell survival and cell death [62]. This particular iso-enzyme, while previously accepted to be expressed solely in the brain, was found to be aberrantly expressed in some colon and breast cancer cells, where a role for it as a putative tumor suppressor has been suggested [63].
Emerging evidence highlights the tumor-suppressive roles of protein kinase C (PKC) isoenzymes, with most PKC mutations in cancers being not just loss-of-function but also dominant negative [62]. Their frequent inactivation suggests that restoring PKC activity could be a viable cancer treatment strategy. Satow et al. reported the expression of PKCγ (PRKCG) in normal colonic epithelia, with its downregulation linked to poor patient outcomes [64].
In gliomas, PRKCG expression and methylation patterns correlate with tumor progression, with glioblastomas exhibiting the lowest expression and highest methylation [65]. Though PRKCG’s role in glioblastoma remains unclear, its upregulation following curcumin treatment in this study suggests a potential anti-tumorigenic effect, possibly influencing key dysregulated signaling pathways in cancer.
Curcumin has been shown to modulate Wnt signaling, a pathway frequently overactivated in glioblastoma and other cancers [66]. As key receptors for frizzled proteins, transmembrane frizzled receptors drive canonical and non-canonical signaling, influencing proliferation, angiogenesis, migration, and invasion [67]. High expression of FZD1, 2, 5, 6, 7, and 8 correlates with oncogenic mTOR signaling and poor prognosis in gliomas [68].
In this study, curcumin treatment downregulated the expression of five frizzled receptors: FZD2 (2.95), FZD8 (3.18), FZD1 (2.62), FZD7 (1.93), and FZD5 (1.99). Among these, FZD2 is a known prognostic marker for glioma progression, with its expression linked to tumor grade [69]. Ran et al. found that FZD2 suppression reduced glioblastoma cell stemness, proliferation, migration, and invasion, coinciding with the inhibition of the Notch and NF-κB pathways [70].
High expression of FZD1 and FZD8 has been linked to poor overall survival in glioblastoma patients [71,72]. In low-grade glioma, FZD8 was identified as a key driver of tumor recurrence [72]. Interestingly, its repression via bivalent modification in glioma tumorigenesis suggests a potential tumor-suppressive role [73]. While its function in glioblastoma remains unclear, aberrant FZD8 expression has been implicated in gastric, prostate, lung, pancreatic, and renal cancers [74,75]. Additionally, FZD8 activation of Wnt signaling contributes to chemo-resistance in triple-negative breast cancer [76]. Whether FZD downregulation in curcumin-treated cells suppressed tumorigenesis warrants further investigation.
Curcumin treatment influenced TGFβ family protein expression in this study. Growth differentiation factor (GDF) proteins, part of the TGF-β family, regulate cell differentiation, proliferation, and apoptosis [77]. Loss of differentiation, a hallmark of cancer, fuels unchecked proliferation, often driven by cancer stem cells [78]. Inducing differentiation limits proliferation, shifting cells to a therapy-sensitive, less metastatic state [79]. While GDF proteins have both tumorigenic and tumor-suppressive roles, their exact functions in cancer remain unclear [77]. GDF2 suppresses breast and ovarian cancer metastasis [80], while GDF9 (upregulated 3.75-fold by curcumin) exhibits tumor-suppressive effects in bladder cancer [81]. Conversely, elevated GDF15 levels in glioblastoma patients are correlated with a poor prognosis, immune suppression, and increased invasion [82,83]. Interestingly, Kadowaki et al. reported that GDF15 overexpression in glioblastoma cells with low basal levels induced apoptosis [84]. In this study, GDF15 mRNA increased 1.52-fold with curcumin treatment, highlighting the complex and context-dependent roles of GDF and other TGFβ superfamily members.
Curcumin treatment downregulated GDF5 expression (2.68) in this study. While its role in glioblastoma remains unclear, previous findings suggest GDF5 may contribute to cancer progression. It induces cyclin D1 transcription, driving epidermal stem cell proliferation in vitro [85]. TCGA data indicate elevated GDF5 expression in triple-negative breast cancer, linking it to tumor aggressiveness and metastasis [86]. Additionally, GDF5-driven TGF-β1 activity promotes endothelial proliferation and angiogenesis in breast cancer models [87].
Curcumin treatment significantly upregulated GDF7 (7.78), potentially contributing to its anti-tumorigenic effects. Clinically, low GDF7 expression is associated with a poor prognosis and increased recurrence in hepatocellular carcinoma, where it suppresses migration, invasion, and tumor growth in xenograft models [88]. Similarly, lower GDF7 levels predict worse outcomes in breast cancer [41]. In medulloblastoma, GDF7 induces phospho-CREB expression and enhances neuronal differentiation markers [89]. Other upregulated TGF-β family members in this study include NODAL (7.92) and LEFTY2 (6.89-fold). While NODAL drives glioblastoma proliferation, LEFTY2 counteracts its activation of SMAD and ERK signaling [90].
Conclusion
Curcumin is a promising therapeutic agent that modulates key cancer-related processes, including cell proliferation, apoptosis, and the dynamics of cancer stem cells. It exerts its effects by regulating key signaling pathways, including calcium signaling, MAPK, Ras, TGF-β, Wnt, cytokine, and TNF pathways. Curcumin influences the expression of several pivotal genes, including AIFM3, TP53AIP1, CRD14, NIBAN3, BOK, BCL2L10, BCL2L14, BNIPL, FASLG, GZMM, TNFSF10, TNFSF11, TNFSF4, FOS, PRKCG, GDF7, GDF9, GDF15, GDF5, FZD1, FZD2, FZD8, SMAD5, APC, WNT7B, and HOXA1. By modulating these interconnected pathways, curcumin may impair the survival of cancer stem cells, reduce therapy resistance, inhibit metastasis, and sensitize tumor cells to conventional treatments, highlighting its multifaceted role in cancer prevention and therapy.
Author Contributions
Hankins and Reddy conceptualization. Nicole, Iqra and Subbu experimentation, Natarajan RNAseq analysis, Write up Nicole, Subbu, Hankins and Reddy, Editing Subbu and Reddy
Acknowledgments
This material is based upon on the work supported by the National Science Foundation (NSF) under award number 2242771.
Conflicts of Interest
The Authors have no conflicts of interest
Data availability
The raw paired-end Illumina RNA sequencing reads generated in the current study are available in the Sequence Read Archive (SRA) at NCBI under the Bio project accession numbers PRJNA1243709.
References
- Agosti, E.; Zeppieri, M.; De Maria, L.; Tedeschi, C.; Fontanella, M.M.; Panciani, P.P.; Ius, T. Glioblastoma immunotherapy: A systematic review of the present strategies and prospects for advancements. International Journal of Molecular Sciences 2023, 24, 15037. [Google Scholar] [CrossRef]
- Ren, X.; Qin, L. Glioblastoma Management in the Post-COVID-19 Era: Challenges, Strategies, and Adaptations. 2025. [Google Scholar]
- Jackson, G.A.; Adamson, D.C. Similarities in Mechanisms of Ovarian Cancer Metastasis and Brain Glioblastoma Multiforme Invasion Suggest Common Therapeutic Targets. Cells 2025, 14, 171. [Google Scholar] [CrossRef] [PubMed]
- Aleksakhina, S.N.; Kashyap, A.; Imyanitov, E.N. Mechanisms of acquired tumor drug resistance. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 2019, 1872, 188310. [Google Scholar] [CrossRef] [PubMed]
- Srirangan, P.; Sabina, E.P. Protective effects of herbal compounds against cyclophosphamide-induced organ toxicity: a pathway-centered approach. Drug and Chemical Toxicology 2025, 1–43. [Google Scholar] [CrossRef]
- Reddy, C.S.; Natarajan, P.; Nimmakayala, P.; Hankins, G.R.; Reddy, U.K. From Fruit Waste to Medical Insight: The Comprehensive Role of Watermelon Rind Extract on Renal Adenocarcinoma Cellular and Transcriptomic Dynamics. International Journal of Molecular Sciences 2023, 24, 15615. [Google Scholar] [CrossRef] [PubMed]
- Chinreddy, S.R.; Mashozhera, N.T.; Rashrash, B.; Flores-Iga, G.; Nimmakayala, P.; Hankins, G.R.; Harris, R.T.; Reddy, U.K. Unraveling TRPV1’s Role in Cancer: Expression, Modulation, and Therapeutic Opportunities with Capsaicin. Molecules 2024, 29, 4729. [Google Scholar] [CrossRef]
- Datta, S.; Saha, P.; Dey, S.; Sinha, D. Natural Products as Chemosensitizers for Adjunct Therapy in Cancer Management. Pharmacotherapeutic Botanicals for Cancer Chemoprevention 2020, 67–119. [Google Scholar]
- Islam, M.R.; Rauf, A.; Akash, S.; Trisha, S.I.; Nasim, A.H.; Akter, M.; Dhar, P.S.; Ogaly, H.A.; Hemeg, H.A.; Wilairatana, P. Targeted therapies of curcumin focus on its therapeutic benefits in cancers and human health: Molecular signaling pathway-based approaches and future perspectives. Biomedicine & Pharmacotherapy 2024, 170, 116034. [Google Scholar]
- Banerjee, S.; Singh, S.K.; Chowdhury, I.; Lillard Jr, J.W.; Singh, R. Combinatorial effect of curcumin with docetaxel modulates apoptotic and cell survival molecules in prostate cancer. Frontiers in bioscience (Elite edition) 2017, 9, 235. [Google Scholar]
- Tiryaki, S.; Macit, M.; Zemheri, I.E.; Süt, P.A.; Duman, G.; Telci, D. Anticancer Activity of Soy Lecithin-Based Curcumin in Prostate Cancer. Journal of Applied Polymer Science 2025, e56816. [Google Scholar] [CrossRef]
- Du, B.; Jiang, L.; Xia, Q.; Zhong, L. Synergistic inhibitory effects of curcumin and 5-fluorouracil on the growth of the human colon cancer cell line HT-29. Chemotherapy 2005, 52, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Cespedes-Acuña, C.L.; Yang, Z.; Abu Bakar, M.Z.; Chan, K.W.; Deng, X. Plant foods and their bioactives as dietary enhancers for colon cancer treatment with 5-fluorouracil. Food Reviews International 2025, 1–50. [Google Scholar] [CrossRef]
- Casarcia, N.; Rogers, P.; Guld, E.; Iyer, S.; Li, Y.; Burcher, J.T.; Deliberto, L.K.; Banerjee, S.; Bishayee, A. Phytochemicals for the prevention and treatment of pancreatic cancer: Current progress and future prospects. British Journal of Pharmacology 2023. [Google Scholar] [CrossRef]
- Allegra, A.; Mirabile, G.; Ettari, R.; Pioggia, G.; Gangemi, S. The impact of curcumin on immune response: an immunomodulatory strategy to treat sepsis. International journal of molecular sciences 2022, 23, 14710. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Rane, G.; Kanchi, M.M.; Arfuso, F.; Chinnathambi, A.; Zayed, M.; Alharbi, S.A.; Tan, B.K.; Kumar, A.P.; Sethi, G. The multifaceted role of curcumin in cancer prevention and treatment. Molecules 2015, 20, 2728–2769. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, K.M.; Mahesh, V.B.; Brann, D.W. Curcumin suppresses growth and chemoresistance of human glioblastoma cells via AP-1 and NFκB transcription factors. Journal of neurochemistry 2007, 102, 522–538. [Google Scholar] [CrossRef]
- Lin, S.-S.; Lai, K.-C.; Hsu, S.-C.; Yang, J.-S.; Kuo, C.-L.; Lin, J.-P.; Ma, Y.-S.; Wu, C.-C.; Chung, J.-G. Curcumin inhibits the migration and invasion of human A549 lung cancer cells through the inhibition of matrix metalloproteinase-2 and-9 and Vascular Endothelial Growth Factor (VEGF). Cancer letters 2009, 285, 127–133. [Google Scholar] [CrossRef]
- Xu, M.X.; Zhao, L.; Deng, C.; Yang, L.; Wang, Y.; Guo, T.; Li, L.; Lin, J.; Zhang, L. Curcumin suppresses proliferation and induces apoptosis of human hepatocellular carcinoma cells via the wnt signaling pathway. International journal of oncology 2013, 43, 1951–1959. [Google Scholar] [CrossRef]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin inhibits migration and invasion of non-small cell lung cancer cells through up-regulation of miR-206 and suppression of PI3K/AKT/mTOR signaling pathway. Acta pharmaceutica 2020, 70, 399–409. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, K.; Liu, J.; Yang, J.; Tian, Y.; Yang, C.; Li, Y.; Shao, M.; Su, W.; Song, N. Curcumin regulates cancer progression: Focus on ncRNAs and molecular signaling pathways. Frontiers in Oncology 2021, 11, 660712. [Google Scholar] [CrossRef] [PubMed]
- Khazei, K.; Jamali, M.; Sarhadi, S.; Dadashpour, M.; Shokrollahzade, S.; Zarghami, N. Transcriptome profiling of curcumin-treated T47D human breast cancer cells by a system-based approach. Gene Reports 2022, 27, 101556. [Google Scholar] [CrossRef]
- Wang, R.; Yu, H.; Chen, P.; Yuan, T.; Zhang, J. Integrated transcriptome and molecular docking to identify the hub superimposed attenuation targets of curcumin in breast cancer cells. International Journal of Molecular Sciences 2023, 24, 12479. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liang, C.; Yang, H.; Li, X.; Deng, X.; Liang, X.; Li, L.; Huang, Z.; Lu, D.; Ma, Y. Curcumin induces apoptosis and inhibits the growth of adrenocortical carcinoma: Identification of potential candidate genes and pathways by transcriptome analysis. Oncology Letters 2021, 21, 476. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, J.; Zhou, Y.; Gao, Q.; Wang, R.; Fu, Y.; Zheng, L.; Yu, H. Transcriptome investigation and in vitro verification of curcumin-induced HO-1 as a feature of ferroptosis in breast cancer cells. Oxidative Medicine and Cellular Longevity 2020, 2020, 3469840. [Google Scholar] [CrossRef]
- Mo, F.; Xiao, Y.; Zeng, H.; Fan, D.; Song, J.; Liu, X.; Luo, M.; Ma, X. Curcumin-induced global profiling of transcriptomes in small cell lung cancer cells. Frontiers in Cell and Developmental Biology 2021, 8, 588299. [Google Scholar] [CrossRef]
- Bolger, A.; Giorgi, F. Trimmomatic: A flexible read trimming tool for Illumina NGS data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biol 2014, 15, 10–1186. [Google Scholar]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic acids research 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]
- Salam, R.; Saliou, A.; Bielle, F.; Bertrand, M.; Antoniewski, C.; Carpentier, C.; Alentorn, A.; Capelle, L.; Sanson, M.; Huillard, E. Cellular senescence in malignant cells promotes tumor progression in mouse and patient Glioblastoma. Nature communications 2023, 14, 441. [Google Scholar] [CrossRef]
- Giordano, A.; Tommonaro, G. Curcumin and cancer. Nutrients 2019, 11, 2376. [Google Scholar] [CrossRef]
- Wong, S.C.; Kamarudin, M.N.A.; Naidu, R. Anticancer mechanism of curcumin on human glioblastoma. Nutrients 2021, 13, 950. [Google Scholar] [CrossRef]
- Zoi, V.; Kyritsis, A.P.; Galani, V.; Lazari, D.; Sioka, C.; Voulgaris, S.; Alexiou, G.A. The role of curcumin in cancer: a focus on the PI3K/Akt pathway. Cancers 2024, 16, 1554. [Google Scholar] [CrossRef]
- Hesari, A.; Rezaei, M.; Rezaei, M.; Dashtiahangar, M.; Fathi, M.; Rad, J.G.; Momeni, F.; Avan, A.; Ghasemi, F. Effect of curcumin on glioblastoma cells. Journal of cellular physiology 2019, 234, 10281–10288. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-L.; Liu, Y.-Y.; Ma, Y.-G.; Xue, Y.-X.; Liu, D.-G.; Ren, Y.; Liu, X.-B.; Li, Y.; Li, Z. Curcumin blocks small cell lung cancer cells migration, invasion, angiogenesis, cell cycle and neoplasia through Janus kinase-STAT3 signalling pathway. PloS one 2012, 7, e37960. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-y.; Zheng, Y.; Jiao, D.-m.; Chen, F.-y.; Hu, H.-z.; Wu, Y.-q.; Song, J.; Yan, J.; Wu, L.-j.; Lv, G.-y. Curcumin inhibits lung cancer cell migration and invasion through Rac1-dependent signaling pathway. The Journal of nutritional biochemistry 2014, 25, 177–185. [Google Scholar] [CrossRef]
- Li, W.; Sun, L.; Lei, J.; Wu, Z.; Ma, Q.; Wang, Z. Curcumin inhibits pancreatic cancer cell invasion and EMT by interfering with tumor-stromal crosstalk under hypoxic conditions via the IL-6/ERK/NF-κB axis. Oncology reports 2020, 44, 382–392. [Google Scholar] [CrossRef]
- Weissenberger, J.; Priester, M.; Bernreuther, C.; Rakel, S.; Glatzel, M.; Seifert, V.; Kögel, D. Dietary curcumin attenuates glioma growth in a syngeneic mouse model by inhibition of the JAK1, 2/STAT3 signaling pathway. Clinical cancer research 2010, 16, 5781–5795. [Google Scholar] [CrossRef]
- Li, J.; Ye, L.; Parr, C.; Douglas-Jones, A.; Kynaston, H.G.; Mansel, R.E.; Jiang, W.G. The aberrant expression of bone morphogenetic protein 12 (BMP-12) in human breast cancer and its potential prognostic value. Gene Therapy and Molecular Biology 2009, 13, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Abdullah Thani, N.A.; Sallis, B.; Nuttall, R.; Schubert, F.R.; Ahsan, M.; Davies, D.; Purewal, S.; Cooper, A.; Rooprai, H.K. Induction of apoptosis and reduction of MMP gene expression in the U373 cell line by polyphenolics in Aronia melanocarpa and by curcumin. Oncology reports 2012, 28, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Banik, N.L.; Ray, S.K. Curcumin suppressed anti-apoptotic signals and activated cysteine proteases for apoptosis in human malignant glioblastoma U87MG cells. Neurochemical research 2007, 32, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Jiao, J.T.; Qian, Y.; Guo, X.Y.; Huang, J.; Dai, M.C.; Zhang, L.; Ding, X.P.; Zong, D.; Shao, J.F. Curcumin induces G2/M arrest and triggers apoptosis via FoxO1 signaling in U87 human glioma cells. Molecular Medicine Reports 2016, 13, 3763–3770. [Google Scholar] [CrossRef]
- Wang, P.; Hao, X.; Li, X.; Yan, Y.; Tian, W.; Xiao, L.; Wang, Z.; Dong, J. Curcumin inhibits adverse psychological stress-induced proliferation and invasion of glioma cells via down-regulating the ERK/MAPK pathway. Journal of Cellular and Molecular Medicine 2021, 25, 7190–7203. [Google Scholar] [CrossRef]
- Wang, L.; Hu, R.; Dai, A. Curcumin Increased the Sensitivity of Non-Small-Cell Lung Cancer to Cisplatin through the Endoplasmic Reticulum Stress Pathway. Evidence-Based Complementary and Alternative Medicine 2022, 2022, 6886366. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Zhu, D.-J.; Chen, X.-W.; Chen, Q.-K.; Luo, Z.-T.; Liu, C.-C.; Wang, G.-X.; Zhang, W.-J.; Liao, N.-Z. Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget 2017, 8, 40264. [Google Scholar] [CrossRef]
- Su, X.; Chen, S.; Lu, H.; Li, H.; Qin, C. Study on the inhibitory effect of curcumin on GBM and its potential mechanism. Drug Design, Development and Therapy 2781, 2781–2781. [Google Scholar] [CrossRef]
- Huang, T.-Y.; Tsai, T.-H.; Hsu, C.-W.; Hsu, Y.-C. Curcuminoids suppress the growth and induce apoptosis through caspase-3-dependent pathways in glioblastoma multiforme (GBM) 8401 cells. Journal of agricultural and food chemistry 2010, 58, 10639–10645. [Google Scholar] [CrossRef]
- Mosieniak, G.; Adamowicz, M.; Alster, O.; Jaskowiak, H.; Szczepankiewicz, A.A.; Wilczynski, G.M.; Ciechomska, I.A.; Sikora, E. Curcumin induces permanent growth arrest of human colon cancer cells: link between senescence and autophagy. Mechanisms of ageing and development 2012, 133, 444–455. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Yu, Y.; Liu, J.; Luo, T.; Fan, F. Glioblastoma treatment modalities besides surgery. Journal of Cancer 2019, 10, 4793. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zeng, M.; Peng, B.; Li, P.; Zhao, S. Transient receptor potential vanilloid-1 (TRPV1) channels act as suppressors of the growth of glioma. Brain Research Bulletin 2024, 211, 110950. [Google Scholar] [CrossRef] [PubMed]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Malani, N.; Ichikawa, H. Curcumin: the Indian solid gold. The molecular targets and therapeutic uses of curcumin in health and disease 2007, 1–75. [Google Scholar]
- Avci, C.; Dodurga, Y.; Susluer, S.; Sıgva, Z.; Yucebas, M.; Caglar, H.; Akalin, T.; Dalbasti, T.; Oktar, N.; Gunduz, C. Promoter hypermethylation-mediated down-regulation of RUNX3 gene in human brain tumors. Irish journal of medical science 2014, 183, 259–264. [Google Scholar] [CrossRef]
- Chuang, L.; Ito, Y. RUNX3 is multifunctional in carcinogenesis of multiple solid tumors. Oncogene 2010, 29, 2605–2615. [Google Scholar] [CrossRef]
- Li, X.; Zheng, J.; Diao, H.; Liu, Y. RUNX 3 is down-regulated in glioma by Myc-regulated miR-4295. Journal of Cellular and Molecular Medicine 2016, 20, 518–525. [Google Scholar] [CrossRef]
- Steponaitis, G.; Kazlauskas, A.; Vaitkienė, P.; Deltuva, V.P.; Mikuciunas, M.; Skiriutė, D. Oncosuppressive role of RUNX3 in human astrocytomas. Journal of Oncology 2019, 2019, 1232434. [Google Scholar] [CrossRef]
- Mei, P.-J.; Bai, J.; Liu, H.; Li, C.; Wu, Y.-P.; Yu, Z.-Q.; Zheng, J.-N. RUNX3 expression is lost in glioma and its restoration causes drastic suppression of tumor invasion and migration. Journal of cancer research and clinical oncology 2011, 137, 1823–1830. [Google Scholar] [CrossRef]
- Sun, J.; Jia, Z.; Li, B.; Zhang, A.; Wang, G.; Pu, P.; Chen, Z.; Wang, Z.; Yang, W. MiR-19 regulates the proliferation and invasion of glioma by RUNX3 via β-catenin/Tcf-4 signaling. Oncotarget 2017, 8, 110785. [Google Scholar] [CrossRef]
- Sun, J.; Li, B.; Jia, Z.; Zhang, A.; Wang, G.; Chen, Z.; Shang, Z.; Zhang, C.; Cui, J.; Yang, W. RUNX3 inhibits glioma survival and invasion via suppression of the β-catenin/TCF-4 signaling pathway. Journal of neuro-oncology 2018, 140, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C.; Brognard, J. Reversing the paradigm: protein kinase C as a tumor suppressor. Trends in pharmacological sciences 2017, 38, 438–447. [Google Scholar] [CrossRef]
- Pilo, C.A.; Newton, A.C. Two sides of the same coin: protein kinase C γ in cancer and neurodegeneration. Frontiers in cell and developmental biology 2022, 10, 929510. [Google Scholar] [CrossRef] [PubMed]
- Satow, R.; Suzuki, Y.; Asada, S.; Ota, S.; Idogawa, M.; Kubota, S.; Ikeo, N.; Yoneda, A.; Fukami, K. Downregulation of protein kinase C gamma reduces epithelial property and enhances malignant phenotypes in colorectal cancer cells. Iscience 2022, 25. [Google Scholar] [CrossRef]
- Liu, L.; Wang, G.; Wang, L.; Yu, C.; Li, M.; Song, S.; Hao, L.; Ma, L.; Zhang, Z. Computational identification and characterization of glioma candidate biomarkers through multi-omics integrative profiling. Biology Direct 2020, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Wang, X.; Wang, X.-J.; Zheng, B.-Z.; Wang, Y.; Liang, B. Curcumin inhibits the growth via Wnt/β-catenin pathway in non-small-cell lung cancer cells. European Review for Medical & Pharmacological Sciences 2018, 22. [Google Scholar]
- Zhang, X.; Dong, S.; Xu, F. Structural and druggability landscape of frizzled G protein-coupled receptors. Trends in Biochemical Sciences 2018, 43, 1033–1046. [Google Scholar] [CrossRef]
- Huang, K.; Xu, H.; Han, L.; Xu, R.; Xu, Z.; Xie, Y. Identification of therapeutic targets and prognostic biomarkers among frizzled family genes in glioma. Frontiers in Molecular Biosciences 2023, 9, 1054614. [Google Scholar] [CrossRef]
- Hirano, H.; Yonezawa, H.; Yunoue, S.; Habu, M.; Uchida, H.; Yoshioka, T.; Kishida, S.; Kishida, M.; Oyoshi, T.; Fujio, S. Immunoreactivity of Wnt5a, Fzd2, Fzd6, and Ryk in glioblastoma: evaluative methodology for DAB chromogenic immunostaining. Brain tumor pathology 2014, 31, 85–93. [Google Scholar] [CrossRef]
- Ran, Y.; Han, S.; Gao, D.; Chen, X.; Liu, C. Interference of FZD2 suppresses proliferation, vasculogenic mimicry and stemness in glioma cells via blocking the Notch/NF-κB signaling pathway. Experimental and Therapeutic Medicine 2024, 28, 373. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, L.; Jiang, Z.; Tan, G.; Wang, Z. FZD1/KLF10-hsa-miR-4762-5p/miR-224-3p-circular RNAs axis as prognostic biomarkers and therapeutic targets for glioblastoma: a comprehensive report. BMC Medical Genomics 2023, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Guo, Z.; Wang, B.; Yang, M.; Yuan, X.; Ji, B.; Wu, Y.; Chen, S. A Computational Framework to Identify Biomarkers for Glioma Recurrence and Potential Drugs Targeting Them. Frontiers in Genetics 2022, 12, 832627. [Google Scholar] [CrossRef]
- Ohka, F.; Shinjo, K.; Deguchi, S.; Matsui, Y.; Okuno, Y.; Katsushima, K.; Suzuki, M.; Kato, A.; Ogiso, N.; Yamamichi, A. Pathogenic epigenetic consequences of genetic alterations in IDH-wild-type diffuse astrocytic gliomas. Cancer research 2019, 79, 4814–4827. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Z.; Zhang, Y. Expression and prognostic impact of FZDs in pancreatic adenocarcinoma. BMC gastroenterology 2021, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, W.; Zhao, C. Frizzled receptors in tumors, focusing on signaling, roles, modulation mechanisms, and targeted therapies. Oncology Research 2021, 28, 661. [Google Scholar] [CrossRef]
- Yin, S.; Xu, L.; Bonfil, R.D.; Banerjee, S.; Sarkar, F.H.; Sethi, S.; Reddy, K.B. Tumor-initiating cells and FZD8 play a major role in drug resistance in triple-negative breast cancer. Molecular cancer therapeutics 2013, 12, 491–498. [Google Scholar] [CrossRef]
- Caja, L.; Bellomo, C.; Moustakas, A. Transforming growth factor β and bone morphogenetic protein actions in brain tumors. FEBS letters 2015, 589, 1588–1597. [Google Scholar] [CrossRef]
- Tenen, D.G. Disruption of differentiation in human cancer: AML shows the way. Nature reviews cancer 2003, 3, 89–101. [Google Scholar] [CrossRef]
- Hanavadi, S.; Martin, T.; Watkins, G.; Mansel, R.; Jiang, W. The role of growth differentiation factor-9 (GDF-9) and its analog, GDF-9b/BMP-15, in human breast cancer. Annals of surgical oncology 2007, 14, 2159–2166. [Google Scholar] [CrossRef]
- Varadaraj, A.; Patel, P.; Serrao, A.; Bandyopadhay, T.; Lee, N.Y.; Jazaeri, A.A.; Huang, Z.; Murphy, S.K.; Mythreye, K. Epigenetic regulation of GDF2 suppresses anoikis in ovarian and breast epithelia. Neoplasia 2015, 17, 826–838. [Google Scholar] [CrossRef]
- Du, P.; Ye, L.; Li, H.; Ruge, F.; Yang, Y.; Jiang, W.G. Growth differentiation factor-9 expression is inversely correlated with an aggressive behaviour in human bladder cancer cells. International journal of molecular medicine 2012, 29, 428–434. [Google Scholar]
- Codó, P.; Weller, M.; Kaulich, K.; Schraivogel, D.; Silginer, M.; Reifenberger, G.; Meister, G.; Roth, P. Control of glioma cell migration and invasiveness by GDF-15. Oncotarget 2016, 7, 7732. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Li, Z.; Fu, J.; Zhou, R. Growth and differentiation factor 15 regulates PD-L1 expression in glioblastoma. Cancer Management and Research 2019, 2653–2661. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, M.; Yoshioka, H.; Kamitani, H.; Watanabe, T.; Wade, P.A.; Eling, T.E. DNA methylation-mediated silencing of nonsteroidal anti-inflammatory drug-activated gene (NAG-1/GDF15) in glioma cell lines. International journal of cancer 2012, 130, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Bian, R.; Wang, F.; Wang, Y.; Li, X.; Guo, Y.; Zhang, X.; Luo, G.; Zhan, R. GDF-5 promotes epidermal stem cells proliferation via Foxg1-cyclin D1 signaling. Stem cell research & therapy 2021, 12, 1–11. [Google Scholar]
- Liu, M.; Sui, L.; Fang, Z.; Jiang, W.G.; Ye, L. Aberrant expression of bone morphogenetic proteins in the disease progression and metastasis of breast cancer. Frontiers in Oncology 2023, 13, 1166955. [Google Scholar] [CrossRef]
- Margheri, F.; Schiavone, N.; Papucci, L.; Magnelli, L.; Serratì, S.; Chilla, A.; Laurenzana, A.; Bianchini, F.; Calorini, L.; Torre, E. GDF5 regulates TGFß-dependent angiogenesis in breast carcinoma MCF-7 cells: in vitro and in vivo control by anti-TGFß peptides. PloS one 2012, 7, e50342. [Google Scholar] [CrossRef]
- Zhuo, J.; Li, H.; Zhang, P.; He, C.; Shen, W.; Yang, X.; Lin, Z.; Zhuang, R.; Wei, X.; Zheng, S. Growth differentiation factor 7 alleviates the proliferation and metastasis of hepatocellular carcinoma. Liver Research 2024, 8, 259–268. [Google Scholar] [CrossRef]
- Armandari, I.; Zomerman, W.W.; Plasschaert, S.L.; Smit, M.J.; Martini, T.E.; de Camargo Magalhães, E.S.; Hogeling, S.M.; Rozema-Huizinga, G.C.; Lourens, H.J.; Meeuwsen-de Boer, T.G. CREB signaling activity correlates with differentiation and survival in medulloblastoma. Scientific Reports 2021, 11, 16077. [Google Scholar] [CrossRef]
- Nana, A.W.; Yang, P.-M.; Lin, H.-Y. Overview of transforming growth factor β superfamily involvement in glioblastoma initiation and progression. Asian Pacific Journal of Cancer Prevention 2015, 16, 6813–6823. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



