
Submitted:
07 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Freshwater fishes are one of the most threatened animal taxa in North America. In Canada, roughly 30% of the occurring species are currently listed and under protection including redhorses. This review covers Moxostoma spp. ecology, biology, conservation efforts and highlights challenges, to galvanize actions and outcomes through research. Threats to their survival are strongly associated to increasing anthropogenic pressures, superimposed by climate change effects. In Canada, recovery plans for the copper and the black redhorse, and a management plan for the river redhorse are operational. The recovery strategy for the copper redhorse relies on stocking campaigns of 0+ juveniles which was exclusively based on artificial breeding of captured wild broodstock. A conservation aquaculture program that includes genetic diversity and adaptation considerations was recently initiated and areas of refinements identified are: broodstock and early-life rearing protocols and environmental and physical enrichment protocols to improve juveniles post-release fitness. Research through the prism of conservation physiology are proposed in the identification welfare and health bioindicators and adaptability/response to climatic change. Concurrently, aquatic ecosystem protection/restoration, control of invasive species, eDNA detection/telemetry tracking and post-release monitoring efforts should also be reinforced.
Keywords:
Biodiversity
; Catostomidae
; Moxostoma
; Conservation physiology
; Conservation aquaculture
; freshwater fishes
; Moxostoma hubbsi
1. Introduction
Biodiversity and healthy ecosystems are crucial to the well-being of all life forms and the situation has reached high preoccupation levels [1]. North American freshwater fishes represent one of the most severely threatened animal group and exhibit the highest extinction rates [2,3,4]. Strong action plans to protect freshwater faunal biodiversity are required [5,6,7] and many are implemented. The Catostomidae family is ubiquitous in large river systems of North America and can be considered one of the most ecologically important fish groups [8]. Catostomids (or suckers) are generally non-game fish perceived as low value, bearing little interest although they provide valuable ecosystem services [9,10,11]. The family contains over 75 species distributed across 13 genera in North America, of which 35% are facing conservation issues which tend to be less dealt with in comparison to more valued species [12,13], compromising biodiversity through loss of ecological interactions [14].
Twenty-three species of the genus Moxostoma are included in the Catostomidae family [15,16]. Seven Moxostoma species occur in Canada of which five (copper redhorse M. hubbsi (Legendre 1952) silver redhorse M. anisurum (Rafinesque, 1820), river redhorse M. carinatum (Cope 1870), shorthead redhorse M. macrolepidotum (Lesueur 1817) and greater redhorse M. valenciennesi (Jordan 1885)) are known to inhabit sympatrically in the southwestern region of the St Lawrence River and its tributaries [17]. The two species occurring outside of Québec are the black redhorse, M. duquesnei (Lesueur 1817) and the golden redhorse M. erythrurum (Rafinesque 1818). Shorthead and silver redhorses populations are considered healthy [18]. No conservation efforts have been deployed for the greater redhorse, even though some populations are considered vulnerable [19]. Greater, black, golden, silver, shorthead and river redhorse have all been assessed under the International Union for Conservation of Nature (IUCN) Red list of Threatened Species and are listed as Least Concern [20,21,22,23,24,25]. Three species of Moxostoma are registered amongst the 70 fish species listed for protection under Canada’s Species at Risk Act (SARA) [26], the river redhorse (Special Concern), the black redhorse (Threatened) and the copper redhorse (Endangered). The robust redhorse (M. robustrum, Cope 1870), occurring only in the USA is endangered and qualified as declining and severely fragmented [27] (not covered in this review).
The river redhorse is attributed a national Special Concern conservation status (formerly described as “vulnerable” from 1990 to 1999) by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) 2006). A management plan to “maintain self-sustaining populations at current locations and to restore self-sustaining populations in historical range where feasible” is available [28] and a progress report should be available in 2025 [29]. The black redhorse has a limited extent of occurrence, area of occupancy and is found only in a few rivers in southwestern Ontario. Recovery has been determined to be biologically and technically feasible [30]. The copper redhorse is the only freshwater fish species endemic to Québec (Canada), it is restricted to the southern part of the St Lawrence River system and a few of its tributaries and current extent of occurrence and area of occupancy are approximated at ~2,090 km2 and ~70 km2 respectively. In 1999, it became the first animal species to be designated threatened in Québec. The unique population is considered severely fragmented with continuing decline in mature individuals [31]. The recovery plan is conducted by the ministère de l’Environnement, de la Lutte contre les Changements Climatiques, de la Faune et des Parcs (MELCCFP). It is considered the trigger species of one of the 13 Key Biodiversity Areas in Canada, the Richelieu River, their only known spawning area [32]. The value of Moxostoma hubbsi in terms of biodiversity is high since it is one of few lithophilous spawner of North American that rely exclusively on molluscs for feeding [17]. A forceful demonstration of anthropogenic pressure is currently unfolding in the critical habitat of the copper redhorse with the projected construction of a new port terminal [33] which raises major red flags.
An evaluation on the status of North American Catostomidae is available [12] as well as a literature review that includes culture protocols of currently endangered Catostomidae species (no Moxostoma included) [34]. A review published in 2019, aimed at research progress achieved on SARA-listed fish species, included only one Moxostoma species [26]. Mongeau et al. [17] compared the biology of five Moxostoma species based on historical data and a few specimens in 1992. Overall, data on Moxostoma in peer-reviewed journals remains limited. Conservation efforts for imperilled species stem mostly from government initiatives and accordingly, most of the information is available as grey literature and hard to access and often lacks replication or is potentially biased. The time and effort involved in locating, retrieving and sometime translating makes their use challenging but their inclusion is strongly warranted when information is scarce [35,36,37].
Barriers to conservation actions and successful outcomes are the evident inadequacy of the mechanisms that should translate acquired knowledge into effective actions [38]. Given small population sizes at time of listing, delayed protection and insufficient funding of many conservation actions, a strong strategy to generate synergies within a working framework is essential to achieve downlisting of a species [39]. The objective of our review is to provide up-to-date information on the biology, ecology and conservation efforts of Moxostoma spp. in the hope of triggering an uplift of coordinated research efforts notably in the field of conservation aquaculture. Indeed, with a framework that covers genetic, ecological and societal considerations, conservation aquaculture is a powerful tool to assist the recovery of seriously declining aquatic species [40,41]. An example of a successful reintroduction program is the striped bass (Morone saxatilis,Walbaum 1792) which was extirpated from the St Lawrence River in the 1960s. It was successfully reintroduced after a 15-year recovery plan, the backbone of which was the artificial production of juveniles with more than 34.5 million larvae and juveniles stocked [42,43]. The species is now on the waiting list of officially downlisted species, with the last reintroductions carried out in 2020 [44].
2. Methodology
Web of science and SCOPUS were used as abstract and citation databases and employed different combinations of the keywords: “Catostomidae”, “Moxostoma”, “Moxostoma hubbsi” and latin and common names of all species of Moxostoma, “redhorse”, “copper redhorse”, “suckers”, “biology”, “ecology”, “reproduction”, “conservation”, “aquaculture”. Papers evaluating Moxostoma sp. occurring in Canada were prioritized. The research was broadened to articles from the USA and ultimately to other species of Catostomidae. Subsequently, “grey literature” originating from conservation programs, recovery strategies, progress reports, scholar works, or any other relevant publicly available document were also obtained via networking and the free academic search engine (Google Scholar) and assessed as complementary data (over 35) namely provincial and federal government, recovery teams reports and others information issued by NGOs or covered in memoirs or thesis. The Copper Redhorse Recovery Team was consulted. Peer reviewed literature contributed over 120 articles for all Moxostoma species under assessment. The species that yielded the most results was the shorthead redhorse (M. macrolepidotum) whereas the least results were for the greater redhorse (M. valenciannesi).
3. Catostomidae (Suckers): Moxostoma spp.
Catostomidae is a diverse fish group widely spread across North America in nearly all freshwater habitats but especially in large river systems [8,12]. Suckers tend to be large fishes (>500 mm TL) mostly excluded from the pelagic zones, with sub-terminal large lips adapted for benthic feeding. The shape of the lower lip is the best character to discriminate species of the genus. All suckers play a critical role in nutrient transfer from the benthic to the pelagic food webs contributing to the energy cycle of the ecosystem [12]. Because of their trophic level and abundance, suckers in general are good indicators of environmental quality [45]. Figure 1 (A-G) presents the body and mouth apparatus of the seven Moxostoma species occurring in Canada and Figure 2 (A-F) presents distribution range of extinct and resident populations of Moxostoma spp. except M. hubbsi featured at Figure 3 in section 3.7 of this paper. Significant information about the reproductive behaviour of suckers and key similarities amongst river-inhabiting species such as Moxostoma include 1) spring migrations to spawning grounds composed of coarse substrate/ in fast-running waters; 2) aggregations in shallow areas; 3) upstream spawning; 4) trio quiver (two males for one female); 5) egg-burying behaviour, and 6) the absence of parental care (e.g. guarding of eggs) [46]. Environmental cues that trigger spawning have not been thoroughly studied, seasonality, photoperiod, temperature and stream discharge are the main factors [47,48]. Compared to other populations, Moxostoma sp. of the slow warming St Lawrence River system have a late spawning season, usually June-July, when temperatures reach 16 – 19 °C [49]. Spawn occurs generally during the day [48], but night spawning has been observed [49] for M. valenciennesi. Northern species seem to initiate spawning migration when temperatures reach 10 – 13°C for M. valenciennesi; M. duquesnei; M. erythrurum and M. robustrum [47,48,50]. Water velocity and depth at preferred spawning sites for Moxostoma are generally within 1.7 – 4.5 m sec-1 and 0.5 – 1 m respectively [48]. Reproductive timing among closely related Catostomidae species of North America is poorly understood, other than water temperature and flow patterns (discharge) [51].
Redhorses are a good model to evaluate the effect of habitat perturbation as 1) they undergo seasonal migrations for spawning and overwintering [50], and 2) they display interspecific variations in life-history characteristics, habitat preferences and abundance within close range [52,53]. Hybridization between species seems very rare indicating strong acting genetic and behavioural barriers. The only reported hybrid was a M. macrolepidotum × M. valenciennesi [54]. Reid [55] compared scales, pectoral fin rays and opercula for age estimation others developed genetic markers to assist species identification [56,57,58] and more recently metabarcoding initiatives for detection purposes aimed at Moxostoma species, are reported [59,60]. Since redhorses are easy to confuse amongst the Moxostoma genus and with other species of the Catostomidae family, user-friendly identification keys are available [61,62].
3.1. Silver Redhorse, Moxostoma Anisurum (Rafinesque 1820), Chevalier Blanc
The silver redhorse is one of the shortest Moxostoma with an average of 320 mm and has among the longest longevity (> 20 years). The most extensive contribution to describe the species has been carried out by Page and Burr [63]. In summary, the authors noted a “silver color on both sides and a gray to brown back with a slate gray tail, 41-42 rows of scales on both lateral sides, and 14-17 soft rays on the dorsal fin. The rear edge of the protrusible lips has a deep “V” shape, and it is often confused with the black and golden redhorse but has a slightly convex, instead of concave, dorsal fin and is more deep-bodied” (See Figure 1a). A comparative evaluation of the physiology and swimming performance of M. anisurum, M. carinatum and M. macrolepidotum, revealed that the silver redhorse may exhibit the slowest swimming speed, the slowest time to exhaustion and recovery response [64]. It is also believed to have a slower growth rate, lower mortality and higher recruitment variability compared to M. erythrurum and M. macrolepidotum [8]. Spiegel et al. [65] compared food habit, diet overlap and gill raker morphology in M. anisurum, M. erythrurum and M. macrolepidotum. And suggested that gill raker morphology favoured resources partition associated with reduced competitive interactions. Operculum-based estimation of age was compared with scales and pectoral fin rays in M. anisurum, M. carinatum, M. macrolepidotum and M. valenciennesi [55].
3.1.1. Feeding
The silver redhorse consumes a high proportion of benthic organisms such as molluscs (i.e. Sphaeriidae), insects (i.e. Chironomidae), crustaceans, and small amounts of plant material. Vegetal debris found in the stomachs of captured individuals suggesting feeding within or close to grass beds [8,17,54]. A strong diet overlap is described between silver and shorthead redhorse [65]. Vachon [66] described thoroughly the feeding habits of 0+ and 1+ M. anisurum.
3.1.2. Reproduction/Spawning
Youngest spawners reported were 8-years old in the Richelieu River (QC, Canada [17]. Spatial and temporal overlap in spawning conditions between silver and shorthead redhorse has been observed in three Great Lakes tributaries, as well as similar environmental factors associated with spawning [51]. Both species were observed to spawn earlier and at lower temperatures compared to other Moxostoma sp. [17,67]. The water temperature associated with the peak migration window of both species is very similar and is established at 9.6° ± 0.4°C for M. anisurum populations. Males generally arrive at spawning grounds earlier and in greater numbers and even though males typically reach sexual maturity at a younger age, ages of spawners tend to be similar between males and females [67]. Females lay 14 000-29 000 eggs (an estimated 23 450 per 2 kg female) [17] and initial spawning temperature lies between 13-13.8°C (April-late May) [51,64,68].
3.1.3. Distribution/Habitat Preferences
In Canada, the species is native to the Great Lakes and the St Lawrence River system where it is generally the second most abundant redhorse species (after M. macrolepidotum) [17,68] and its habitat preferences include longer and narrow rivers with silty to firm-bottomed pools and lakes. Spawning generally occurs in shallow, fast flowing waters with coarse bed material often located near dams or riffles [17]. See Figure 2a for the distribution range of the extinct and resident populations of silver redhorse.
3.1.4. Cultivation Activities
No data found.
3.1.5. Conservation Status and Threats
Endemic to Canada and the United States, with no conservation status attributed.
3.1.6. Other
M. anisurum has been the animal model for the evaluation of tapeworm speciation to fish host [69]. The silver redhorse was selected along with the golden redhorse and the greater redhorse, in the study of Fraker et al. [70] evaluating the effects of agricultural conservation practices on stream fish communities. Reid [55] provides operculum-based aged estimates for M. anisurum.
3.2. Black Redhorse, Moxostoma Duquesnei (Lesueur 1817), Chevalier Noir
Most of the research efforts have been carried out on specimens from the Grand River in Ontario and focuses mainly on population demographics, structure and genetics [52,53,58,71,72], early-life stage development [73], as well as habitat and spawning [47,74,75,76]. Adult length (TL in cm) ranges from 25.4 to 40.6 and weight from 0.16-0.70 kg with a lifespan of 10-17 years. A population modelling was carried out in Canada in 2014 [77].
Most complete morphological descriptions of the species can be found in Brown [76]. Summarily, “the black redhorse is described as a slender fish with a long and narrow caudal peduncle with a scale count of 12-13, and a lateral line scale count of 44-47. The shape of the lower lip is described as a plicate, with few papillae, and an obtuse angle of approximately 180 degrees”. The species can be differentiated from M. carinatum, M. hubbsi, M. macrolepidotum and M. valenciennesi by its medium-dark gray with hints of blue tail, and from M. anisurum by its transverse grooves in the lips, concave margin of its dorsal fin and pelvic fin ray count (n=9) (see Figure 1b). It resembles most with M. erythrurum has a larger non-overlapping lateral lined with a scale count (44-47 vs 40-42), lower pelvic ray count (9 vs 10), and the absence of head tubercles of the males during spawning. Sexual dimorphism is observed. Near spawning, males will exhibit an orange to pink colour along their sides and black longitudinal stripes [47].
3.2.1. Feeding
The adults use grazing and picking foraging and feed primarily on insect larvae, mainly chironomids, but also on insects [78]. Juveniles are thought to be planktivorous. The spawning habitat, behavior and morphology as isolating mechanisms of the golden redhorse and the black redhorse and are considered occupying the same habitat with a specified geographical area and sometimes competing for resources [47].
3.2.2. Reproduction/Spawning
They are non-guarder, open substratum spawners and lithophiles. Most extensive studies of the breeding and spawning behaviour were carried out [47,79]. In summary, the species moves relatively short distances for spawning compared to other Moxostoma, and spawns on riffles of rubble and gravel in 15-60 cm depths, avoiding fast-running water. Age at maturity appears to vary between 2-6 years and spawning occurs during the spring at 13-21 °C. Females have been observed to lay between 4 100 – 11 550 non-adhesive eggs with diameters varying between 2.6-2.9 mm [79].
3.2.3. Distribution and Habitat Preferences
In Canada, the species is at the northern edge of its native range where it is limited to southwestern Ontario [52]. It is found in the Mississippi and Great Lakes basins where its distribution is large but disjunct in the western part of its range [80]. It is considered the rarest redhorse species of Ontario [81] (Figure 2b). Knowledge of specific habitat preferences is limited, but studies report that the species typically inhabits warm water rivers of moderate size with low turbidity and slow water velocities [67,79]. Clark [82] reports an average 0.22m s-1. According to Bunt et al. [73] larval black redhorse, like adults, are most often located in clean, clear, stable runs with low-moderate flow, over pebble, gravel and cobble substrate, with a mix of sand. It is also suggested that suitable riverbed material for the species include clean coarse substrate, stable channels and well-developed rifles, and that adults rarely occupy submerged aquatic vegetation (SAV) [67]. The persistence of black redhorse populations in Grand River (Ontario), a river known to display poor water quality (e.g. anthropogenic pollution, near lethal concentration oxygen, pH and temperature) to their use of groundwater upwellings as a shelter from poor conditions [74]. According to Bouvier et al. [82] it is vulnerable to drought and the lowering of water levels and has low tolerance to turbidity in comparison to other Moxostoma species present in its distribution area.
3.2.4. Cultivation Activities
No data were found regarding cultivation or stocking activities. However, it has been identified as a research need for the recovery strategy [82].
3.2.5. Conservation Status and Threats
The black redhorse has been designated threatened since 1988, with its status reconfirmed in 2015, and is listed under SARA since 2019 (Reid 2011). M. duquesnei was designated threatened because it has a limited extent of occurrence and area of occupancy. A recovery and action plan were recently proposed1, after recovery was deemed biologically and technically achievable. Main threats are dams and flow regime variations, high turbidity and high siltation rates, habitat destruction, elevated nutrient levels, urban wastewater and agriculture. Other threats include non-indigenous and invasive species (Oncorhynchus mykiss, Salmo trutta, Neogobius melanostomus, Dreissena polymorpha, D. bugensis and Ctenopharyngodon idella) [82]. Population dynamics are particularly sensitive to perturbations that have impacts on the survival of immature individuals (from hatch to age 4), and early adults (age 2-8). A population abundance target of at least 1 700 adults (age 4+), and 3 900 juveniles, requiring 14.5 ha of suitable habitat [77]. Despite longevity of over ten years, Beckman and Howlett [83] studying age structures based on otolith annulus formation, found that they appeared truncated beyond 4 years, indicating that factors affecting recruitment and mortality are at play in some populations. Non-lethal age determination was evaluated [84] and concordance of inter-readers and intra-replicate age interpretations were greater for fin rays than scales. The use of pectoral fin rays for monitoring recruitment and population age structure on sub-samples is recommended.
Dams are identified as one of the main threats as it causes severe population fragmentation which in turn can affect genetic diversity and gene flow [52,53,55] The population is concentrated in a 150 km stretch of the Grand River watershed (drainage of the Lake Erie) which is fragmented in five populations by the presence of four dams without fishway structures. The species may be extirpated from main tributaries of the Lake Huron such as the Sauble River as no specimens have been collected recently, and only few specimens have been reported in other tributaries. It can utilize fish by-pass structures to migrate upstream [85] and more passage structures are needed to aid populations of M. duquesnei. Pollution is another key threat but there is a lack of studies evaluating the impact of pollution, especially where land use is primarily agriculture, and where major urban centers overlap with locations where it is present [86]. Interestingly, an evaluation of endocrine-disrupting compounds (EDC) impacts on reproduction revealed little to no effect on black and golden redhorse species [87].
In summary, research avenues identified by DFO in 2022 as part of the recovery strategy and action plan include 1) the identification of pollutants sources and their effects; 2) the investigation of the impacts of climate change; 3) the assessment of the impacts of invasive and non-native species; 4) human activities impact, 5) the investigation of the potential use of fish ladders and 6) the examination of the suitability of rearing juveniles to restore the declining populations [86].
3.2.6. Other
The black redhorse as a model to study the impacts of rises in water temperature during critical periods and overall fitness based on optimal spawning temperature and summer impairment threshold levels [88]. Strong evidence that otolith annuli provide accurate age is reported [83]. Mitochondrial DNA-based species diagnostic tests have been developed to validate field identifications of the six Ontario redhorse species [58] including the black redhorse.
3.3. Golden Redhorse, Moxostoma Erythrurum (Rafinesque 1818), Chevalier Doré
In Canada, the golden redhorse is endemic to Manitoba and Ontario and the midwestern, southern and eastern regions of the United States [89]. Data on biology, ecology, life-history and reproduction is extremely scarce, especially for Canadian populations, and most information is accessible via network programs or taxon identifiers. The golden redhorse is described as “a smaller-bodied sucker with relatively large scales and short dorsal fin and average size of the species is 300-450 mm and 0.5-0.9 kg. The species also displays olive (light-yellow to bronze) colors and white bellies with grey tail fins and the lateral line scale count is 39-42, and the dorsal fin is slightly concave, while the caudal fin is notched. The scales on the back and sides are without dark spots and it can be differentiated from the silver redhorse by its outer margin on the dorsal fin (i.e. concave) and its ray count (12-13 rays). Its protrusible lips are differentiated by their continuous ridges and the absence of transverse creases (i.e. papillae). The inferior lip also has parallel folds, and the rear margin is V-shaped” (see also Figure 1c) [80]. The life-history and established length conversion factors and body-scale relationships is described [90]. The author observed that most individuals matured at age 3. Larval phases have been described and compared with Catastomus catastomus, a congener species [91].
3.3.1. Feeding
Typically consumes high proportions invertebrates and only small amounts of plant material, like the silver redhorse [54]. Its diet also includes molluscs, aquatic insects and small crustaceans [90]. It has been observed to have a wider food niche as their feeding strategies appear more generalized compared to the shorthead and the silver redhorse. The growth rate of the golden redhorse appears to be the fastest when compared to the silver and the shorthead redhorse [65].
3.3.2. Reproduction/Spawning
The spawning behavior, the habitat, the timing and the conditions is described [75]. The species moves relatively short distances to spawn, which occurs at temperatures of 17-22°C, and the spawning grounds are described as “shallow extensions into the stream channel with low to moderate non turbulent flow over substrates of sand and gravel” [47]. The authors describe the breeding behavior and observe sexual dimorphism. Water velocity and substrate appear to be the environmental cues for spawning sites selection. Slower current and smaller substrate are preferred in comparison to the black redhorse [47,51]. M. erythrurum appears to display aggressive behavior during spawning with the dominant males defending their territories. Although most Moxostoma species typically bury their eggs in gravel, the golden redhorse exhibited no such behavior and deposit their eggs directly on the substrate (5 000-35 000 eggs per female) [47].
3.3.3. Distribution and Habitat Preferences
The species preferences for substrate are more or less defined as it is found in creeks, rivers and streams of various bed material. Information about preferred habitat for feeding is lacking. The native range of the species remains elusive as recent genetic analysis recently revealed the novel presence of the species in the Susquehanna River basin (Pennsylvania, US) [92] (see also Figure 2c). The golden redhorse is thought to be relatively tolerant to elevated turbidity, but highly sensitive to pollution and siltation and its habitat is most associated with that of black and shorthead redhorses. An evaluation of the impact of dams on the distribution of redhorse species in the Grand River revealed that the golden redhorse had the most widespread distribution compared to M. valenciennesi and M. carinatum suggesting better capacities to use fish passage structures [67,85]. Thermal thresholds were evaluated, and the upper lethal incipient limit and the summer impairment threshold are 33.3 and 35.1°C respectively [93].
3.3.4. Cultivation Activities
No data were found regarding cultivation or stocking activities.
3.3.5. Conservation Status and Threats
Populations are considered stable worldwide and in Canada: Least Concern (IUCN) and Not at Risk (COSEWIC).
3.3.6. Other
Temporal trends in macroscopic indicators of fish health were studied [94]. Gill and body abnormalities were visually scored and tested for correlation with seasonal climatic (flow and stream temperature) and environmental factors (pH, dissolved oxygen and turbidity).
3.4. Shorthead Redhorse, Moxostoma Macrolepidotum (Lesueur 1817), Chevalier Rouge
The shorthead redhorse is one of the most studied species, particularly in water contamination and fish parasite studies [45,95,96,97,98,99]. Biological and ecological descriptions cover morphometrics [17,100], larval development [101], and behavior and swimming performance [18,64,68]. Morphology is described as follow: “the shorthead redhorse is one of the most colorful sucker species; its fins range from bright orange to deep red, both lateral sides are of golden-bronze colour, and the belly ranges from fainted yellow to white. Each scale of the back and upper side is of olive-brown color and presents an indistinct crescent-shaped dark spot at the base. Ray count of the dorsal fin and scale count of the lateral line are 12-13 and 41-45 respectively. Pharyngeal teeth have a slightly sharper edge, almost bladelike with no flat surface for grinding, and are quite numerous on the lower half of the first arch. All fins of breeding males, except the dorsal have tubercles, which are most developed on the anal fin and the caudal fin’s inferior lobe. All fins contain only soft rays, and the dorsal fin is considered relatively short and is usually concave” [54,63] (see also Figure 1E).
M. macrolepidotum exhibits most similarities with the silver redhorse as both species have similar colors and overlapping spawning seasons [51,66], display similar spawning behavior [75] and have the same number of scales on the caudal peduncle (n=12) [17]. Distinctive traits include 1) a shorter head, 2) parallel and straight folds on the rear margin of the lower lip (versus V-shaped), 3) thin arch with slender teeth in a comb-like series of the pharyngeal apparatus (versus thick arch with molariform teeth) and 4) a curved dorsal fin. When compared to other species occurring in Québec (Canada), the shorthead redhorse is the shortest (average 500 mm TL) and has the shortest lifespan (average 11 years) and the slowest growth rate [17]. Individuals of southern populations are reported to generally live for up to 6-8 years. Various age estimation techniques were tested. Annuli formation on scales and pectoral fin rays was validated and it was observed that this approach was unsuitable for adult individuals due to annuli crowding [102]. The authors suggest the use of scales for fishes <5 years old (< 400 mmTL); operculum for fishes of 5-15 years (400-450 mm TL); and otoliths for fishes > 15 years. Its name refers to the short and scaleless head that usually represents 17-19% of TL which differentiate it from all other Moxostoma species. Morphometric and meristic characteristics of the larval stage of M. macrolepidotum are described [91,101]. Reid [71] provides information on the size and age structure, growth and mortality of the shorthead.
3.4.1. Feeding
The diet of the shorthead redhorse, according to DeBofsky et al. [97], is amongst the most diverse. It is dominated by Ephemeroptera (53%), followed by Trichoptera (24%), Chironomids (10%), Crustaceans (8%) and sporadically Lepidoptera and Odonates (dragonflies)[17]. Interestingly, in a study by Doyle et al. [99], almost half (46%) of the gastrointestinal tract content was composed of sediment, suggesting its foraging method may be somewhat inefficient, and may constitute a major route for contaminants. Poor efficiency is explained by the smaller size of the palatal organ of M. macrolepidotum limiting food items separation.
3.4.2. Reproduction/Spawning
The species is reported as one of the earliest spawners [66,67]. The water temperature associated with the peak of migration is established at 9.9° ± 0.4°C [66]. Spawning aggregations have been observed [103]. Spawning was initiated when water temperature reached 14°C and could be sustained up to 22°C [51]. Spawning at temperatures as low as 10°C were observed [75] while Reid [67] observed spawning at temperatures of 11-15°C in their northern range. Preferred spawning habitat is coarse substrate of rubble in shallow riffles (15 – 21 cm deep) with moderate to fast flow rates (0.4-1.4 m3 sec-1) [104]. Mongeau et al. [17] reported 12 080-24 170 eggs per female whereas .
3.4.3. Distribution and Habitat Preferences
The shorthead redhorse is amongst the most wide-ranging species of fish endemic to North America and the most widely spread amongst Moxostoma [102] It is native to central and eastern North America but is now also found in Texas (USA). In Canada, it occurs across Québec to Alberta, inhabiting a wide range of bodies of water from small to large rivers and lakes (see also Figure 2d). Populations tend to be denser in medium to large rivers that have strong flow and extensive areas of silt-free sand, gravel and rubble substrates. Feeding and spawning habitats have been described in detail [17,51,75,103,104]. The species has been observed to inhabit depths of 2.4-4.4 m, 3.3-6.7 m, and 1.1-2.3 m in the spring, summer and winter respectively [104]. Critical thermal maximum thresholds were evaluated, and upper lethal incipient limit and critical thermal maximum (CTmax) are 33.3 and 35.1°C respectively similar to the golden redhorse [93]. Hatry et al. [68] confirmed that passage through a fishway structure was feasible, suggesting that additional structure installations in dammed rivers may significantly help natural populations.
3.4.4. Cultivation Activities
No data were found regarding cultivation or stocking activities.
3.4.5. Conservation Status and Threats
Globally, populations of M. macrolepidotum are attributed the status of Least Concern (IUCN, 2015). In Canada, the status of Secure is given to the species at both national and provincial levels.
3.5. Greater Redhorse, Moxostoma Valenciennesi, Chevalier Jaune
The greater redhorse is considered one of the largest of the genus and can measure up to 800 mm TL and considered a long-lived redhorse that requires large, interconnected river systems [105]. Most resemblance is observed with 1) the river redhorse, as they both have red caudal fins, but distinctions can be made with key morphological characteristic and 2) the copper redhorse its most close relative phylogenetically [106] although a more recent phylogenetic relationship suggests reconsiderations [107]. The most extensive description of the species is as follows: “brown-olive colored back with bronze overcast with golden lateral sides and a white belly. Dorsal, caudal, and anal fins are red and anterior bases of scales have an indistinct dark spot. The dorsal fin is slightly convex with often 13-14 fin rays, but sometimes 11-15, and lateral line scale count is on average 42-45. Scale count around the caudal peduncle is 16 on average. The pharyngeal apparatus presents 55 teeth per arch” [105]. Breeding males exhibit tubercles on dorsal and lateral sides of the head and on body scales, whereas breeding females show smaller tubercles on all fins except dorsal and are absent on the body (see also Figure 1e).
3.5.1. Feeding
3.5.2. Reproduction/Spawning
Early descriptions of spawning behavior originate from Jenkins and Jenkins [49]. Most recent and relevant information about spawning habitat and behaviour, fecundity, egg and larval development, and post-spawn movement as well as habitat use are described [48,108,109]. The following is a summary of the description of the reproduction of the greater redhorse found in their work, unless stated otherwise. Earliest age of maturity for males and females were 5 and 6 years respectively, and oldest individuals were 13 years (male) and 11 years old (female). Spawning is initiated at 13°C and typically lasts two weeks and is sustained at water temperatures up to 19.9°C. Selected spawning grounds were characterized as edges and midstream areas of shallow rifles (10 – 63 cm) with average bottom velocity of 0.3m3 sec-1 on pebble, gravel and cobble mixes.
3.5.3. Distribution and Habitat Preferences
The greater redhorse is endemic to north central and northeastern USA as well as Ontario and Québec (Canada). Due to strong resemblance with the river and the shorthead redhorse, distribution records of the greater redhorse are qualified as unreliable [48]. Nonetheless, description of native range is generally accepted, and the species occurs across a wide geographical range from the Mississippi River to the Hudson Bay, and the Great Lakes-St Lawrence system [80] (see also Figure 2e). It is typically found in clear, relatively fast-moving rivers and in both shallow and deep waters of lakes.
3.5.4. Cultivation Activities
To describe the early stages of development of the greater redhorse, and to identify characters to distinguish larvae of this species from other Catostomidae, gametes obtained from maturing specimens captured at spawning locations along the Grand River (Ontario, Canada) were collected and artificially fertilized [108]. Under laboratory conditions, eggs hatched in 6 to 8 days at 16-19C (128-152 degree-days). Fertilized and water-hardened eggs are yellow, 3.0-3.5 mm in diameter, demersal and non-adhesive Larval development was described and compared to Moxostoma species (M. duquesnei, M. erythrurum, M. macrolepidotum and M. hubbsi). Upon hatching, larvae remained on the bottom and became free swimming after five days. Eggs and larvae were reared in an aerated aquaria with gravel as a substrate. Larvae were fed Artemia sp. nauplii and larger larvae were fed copepods and small cladocerans collected from the river of origin. No stocking activities are reported.
3.5.5. Conservation Status and Threats
Globally, populations of M. valenciennesi are considered stable and given the status of Least Concern by IUCN. No monitoring or management programs are in place. Across its range, it occurs in disjunct populations and appears to be stable or increasing in some portions of their range due to pollution control and improved water quality but are rare or declining in the periphery [105].
3.5.6. Other
A habitat suitability index for the greater redhorse to determine the restoration success after the removal of a dam. Results clearly indicate that the positive change in suitability observed is associated with the change in depth of the river [110]. Spawning area of the greater redhorse increased from zero pre-removal to 13 acres post-removal [111].
3.6. River Redhorse, Moxostoma Carinatum, Chevalier de Rivière
Extensive morphological descriptions of M. carinatum are available [16,49,54,112,113], summarily, “both lateral sides of M. carinatum are of brassy or yellowish to green-coppery colour with white undersides and the back is brown to olive-green. Each dorso-lateral scale exhibits a crescent-shaped dark spot and there usually are 12 rows around the caudal peduncle, and 42-47 scales along the lateral line. The species is often mistaken for the greater redhorse but can be differentiated by the number of scales on the caudal peduncle (i.e. greater redhorse: 15-16 scales), and the dorsal fin is straight or slightly concave, whereas the greater redhorse has a convex dorsal fin. The inferior lip is wider than the upper lip and the rear edge is straight, and the caudal fin is forked with the upper lobe longer and pointier than the lower lobe. M. carinatum also exhibits similarities with shorthead redhorse and distinctions can be made from fin colors, size and number of scales, dorsal fin, entirely plicate lips and buccal teeth” (see also Figure 1f). The primary features distinguishing the river redhorse are the pharyngeal arch that supports molariform teeth (also present in the copper redhorse) as well as its moderate-size head, corrugated lips, and the number of scales on the caudal peduncle. It has fewer teeth with a stronger pharyngeal arch than the copper redhorse [114].
3.6.1. Feeding
The river redhorse is also considered molluscivorous and bears enlarged molariform pharyngeal teeth. Molluscs consist of close to 20% of the diet in comparison to 90% for the copper redhorse. Its diet is considered amongst the most varied in Moxostoma and besides bivalves and gastropods, the species was observed to feed on Ephemeroptera (mayflies) and Trichoptera (caddisflies) [17]. The diet of young-of-the-year specimens revealed a variety of food resources such as cladocerans (22%), diatoms (21%), nematodes (15%), copepods (12.5%), protozoans (6%), and chironomid larvae (4%) [66].
3.6.1. Reproduction/Spawning
M. carinatum is a late-maturing species and reach sexual maturity reportedly at 6-20 years compared to 3-5 years at the more southern latitudes [113]. Recorded spawners of the Richelieu River were minimally 500 mm TL and averaged 10 years old. After the copper redhorse, they are second to last spawners amongst Moxostoma sp. to reach spawning grounds [17]. Spawning migrations are initiated at temperatures of 12°C, and spawning lasts two weeks at temperatures of 17-20°C. The water temperature associated with peak migration activity is close to 7°C higher than that of silver and shorthead redhorse and is established at 16.4° ± 0.3°C [67]. Average fecundity for females is estimated to be 14 010 – 31 050 eggs [17]. Nuptial males exhibit tubercles on the snout, caudal and anal fins and their scales become thicker and rougher during spawning (Reid 2003). A dark lateral stripe in the middle of both sides from the snout to the anal region is observed on spawning males, and tubercles on the anal fin were noted on spawning females [16]. Spawning behaviour has been described [67,112]. Excavation of redds prior to spawning is suspected but not confirmed. Such information is crucial as it may reflect spawning habitat requirements. Optimal spawning conditions include 1) riffle-run riverine with cool, clear and fast-flow, 2) < 2 m depth, 3) velocity of 0.6 – 1.2 m s-1, 4) clean, coarse substrate (i.e. boulder, gravel or cobble), 5) limited submerged vegetation, and 6) temperatures between 17 – 24°C [17,54].
3.6.2. Distribution and Habitat Preferences
The species distribution is described as disjunct populations in Alberta, southern Ontario and Québec, and captures revealed sharing of its distribution range with the copper redhorse but is also present in other rivers of Québec such as the Outaouais River, and other streams in Ontario, and is well established in the United States, almost reaching the Gulf of Mexico (see Figure 2f). Main rivers in Québec and Ontario include the Grand, Trent, Thames, Gatineau, Ottawa and Richelieu rivers, as well as the St Lawrence where a drastic decline has been observed [113]. Passive acoustic monitoring and telemetry tags respectively were used to assess spawning migrations and habitat use in relation to various abiotic parameters (i.e. temperature, weather, discharge variation, moonlight, time of day, depth, substrate composition) [116,117]. Preferred spawning habitat is described as moderate-swift current zones (>0.4m/s), where they aggregate in flowing gravel rifles. Outside the spawning season, the species is thought to inhabit deeper pool habitats but has also been reported in moderate-high velocity riffles [112,115]. Young-of-the-year are typically found along vegetated shorelines of average depth of 1.5m with shallow slopes and fine substrate such as silt, clay or sand. Habitat preferences remain poorly understood during non-spawning periods. The limiting factor appears to be water turbidity although it is unclear if the species is most affected by deposition on substrate affecting its benthic feeding, or suspended sediments [113]. The presence of mature adults is linked dense mollusks beds suggesting their use as foraging post-spawning habitats [117].
3.6.3. Cultivation Activities
No data were found regarding cultivation with the exception of a recent trial [118]. Embryos were obtained 2 hours post fertilization, and incubation was terminated just before the complete resorption of the yolk sac after 14 days; survival rate was 86.5%. The governmental facilities of the MELCCFP from 2021 to 2023, conducted artificial spawning and incubation routinely.[119]. No stocking activity is reported.
3.6.4. Conservation Status and Threats
Based on the species’ highly disjunct status, restricted range and disappearance in historical sites, it was designated of special concern by COSEWIC in 2006. It is a SARA-listed fish species, and a management plan is underway to help restore populations in the northern limit of its range [113]. Main threats are: 1) pollution, 2) siltation, 3) habitat destruction, and 4) flow regime variations [113]. The management plan aims to 1) quantify the extent of existing populations, 2) improve knowledge about habitat requirements, biology, and ecology, as well as long-term population and habitat trends, 3) improve habitat conditions, 4) evaluate and mitigate threats, 5) optimise management resources use, and 6) increase public awareness. To ensure its conservation, research efforts are focused on evaluation of quantity and quality of habitat required, especially during non-spawning periods, population dynamics, standardization of sampling protocols, and genetic structures. No publications are yet available.
3.6.5. Other
The species is known to be very sensitive to anesthesia.
3.7. Copper Redhorse, Moxostoma Hubbsi, (Legendre 1952) Chevalier Cuivré
The most cited morphological description of the species originates from Scott and Crossman [16]. Summarily, the name “copper” refers to “the bright coppery to olive colouration of its back, sides, and head, while the belly displays paler shades, and the fins are dusky to coppery as well. The upper lip is narrower than the lower one with indistinct plicae on the inside edge and edges of the inferior lip form a 105° angle. Pharyngeal apparatus displays n= 18- 21 heavy molariform pharyngeal teeth in a single row, soft ray count on dorsal fin is 13-14 rays, and the caudal is forked. Pelvic fins have 9-10 rays; and pectoral fins ray count is 16-17 rays. Caudal peduncle and lateral line scale counts are 15-16, and 45-47, respectively. Spawning males display tubercles on head, body and every fin” (see also Figure 3).
Figure 3.
Current territory of the Copper Redhorse including spawning sites, migratory passages and potential refuge areas. Adapted from DFO [129].
Figure 3.
Current territory of the Copper Redhorse including spawning sites, migratory passages and potential refuge areas. Adapted from DFO [129].
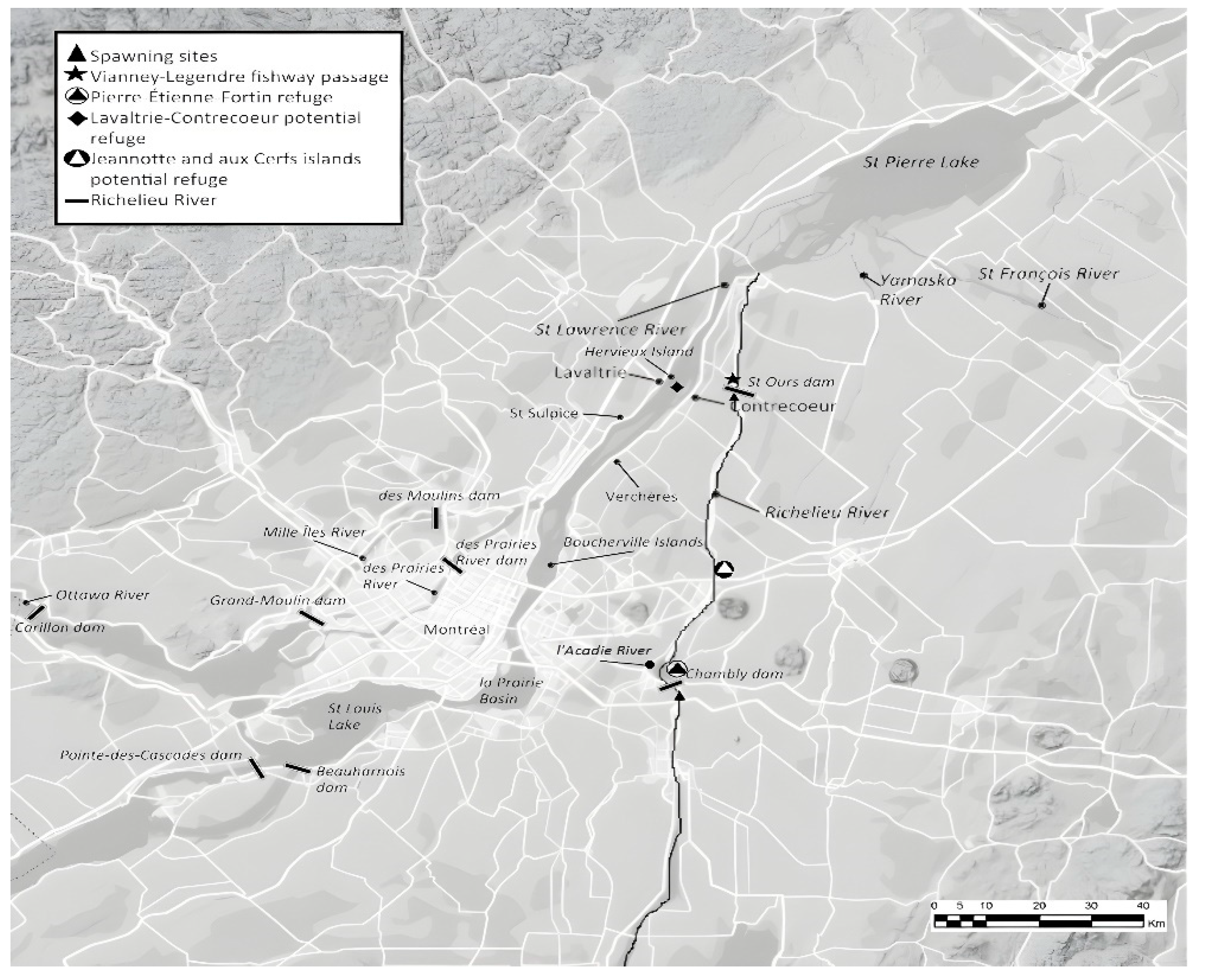
Additional information was published based on fishery inventory data obtained between 1963 and 1973 in Mongeau contributions from 1986 to 1992, and included characterization of growth, feeding behavior, reproduction and habitat preferences. Although based on historical data combined and few specimens, his publications are the most cited. Specific traits that differentiate M. hubbsi from other Moxostoma include 1) larger size (600 – 700 mm TL); 2) superior length growth rate, fecundity and longevity (> 30 years) ; 3) diet; 4) late age maturity (8-10 years); 5) late season breeding (late June- early July), and 6) up to 15-16 rows of scales around the caudal peduncle (vs. 12-13 in other Moxostoma sp.). An identification key is available for the genus Moxostoma (121].
3.7.1. Feeding
Although co-occurring, a high degree of feeding segregation exists between Moxostoma species of Québec [17]. Only M. hubbsi and M. carinatum are considered mollusc feeders with large molariform pharyngeal teeth specialized for crushing shells. The copper redhorse shows the highest degree of specialization, feeding almost exclusively on molluscs (23 taxa within Amnicolidae, Sphaeriidae, Pleuroceridae and Unionidae) [17,122]. Such observations are corroborated by analysis of specific organs associated with feeding. Moxostoma species have chewing pads [123] on the posterior surface of the palatal organ that serves as an obstructing contact surface to coordinate food separation from inorganic debris [124]. Morphology and morphometrics of the palatal organ are valid classification indicators of Moxostoma species as they discriminate for diet and feeding behavior. Morphology comparison amongst 18 species of suckers revealed 1) the broadest range of size and shape variations in M. hubbsi, and 2) highest similarity index between M. hubbsi, M. erythrurum and M. carinatum [124]. The dependency to molluscs as energy source is the highest amongst North American freshwater fish [26] and may limit adaptation capacities to changing environments [125], whereas its rich fatty acid profile may account for greater longevity and fecundity.
3.7.2. Reproduction/Spawning
The copper redhorse typically spawns in more moderate currents and shallow waters (0.4-0.5 m-1 sec, 0.6 – 0.8 m, respectively), but spawning depths may reach 2m [126]. Habitat preferences for spawning have been well described by Mongeau et al. [17] and mostly consist of sections of agitated waters in lowland rivers of medium size with coarse gravel substrate. Spawning ground preferences are shared with all five species currently found in the Richelieu River but M. hubbsi spawns in warmer waters (18 – 26 °C) later in the season, beginning in late June and ending in late July. Age at sexual maturity is considered around 8-10 years and the greatest fecundity counts are reported. A comparison with M. valenciennesi and M. anisurum revealed a 1.26-fold higher egg count (females of 2 kg, averageof 32 750 eggs) [17]. Only two locations in the Richelieu River are known to support reproduction with active spawning sites which consist of the riffle areas in the shallow waters located below the Saint-Ours dam (45°51'46.9"N 73°08'55.2"W) and in the Chambly archipelago (45°26'45.5"N 73°15'38.1"W) within the 70 km section upstream of the St Lawrence River. The capacity of the five (5) Moxostoma species to use the fishway for spawning activities in the Richelieu River has been demonstrated [130]. At both sites, water current is moderately flowing over heterogeneous substrate of coarse gravel, rocks and submerged bedrock fragments in clay, depth is 0.75-2 m. Submerged vegetation areas along the slightly sloped shore (≤20°) in shallow depths (≤1.5m) are important feeding grounds for migrating adults and young-of-the-year juveniles [126]. Female size has been positively correlated with egg diameter ranging from 2.8-3.42 mm. Eggs are non-adhesive, hatch between 89-127 degree-days (i.e. 4.5-6.5 days of incubation at 20°C [131] and the emergent lavae began swimming behaviour between 12-16 days post-fertilization. Juveniles of 80 mm consume small molluscs (<4 mm) and later at 110 mm with a mouth opening of 6.6 mmm could feed on zebra mussels of 8 mm [132].
3.7.3. Distribution and Critical Habitat
The species has a distribution range limited to the southwest of Québec in the St Lawrence River and few secondary rivers, and since its discovery was significantly reduced due to habitat degradation and fragmentation. Two series of telemetry surveys conducted in the spring of 2004 and winter 2007-2008 as part of the MOHU program deployed by a local NGO to develop an atlas of M. hubbsi habitat revealed a greater distribution range than previously estimated (see Figure 3) and reported 1) M. hubbsi still occurs in the Mille-Îles River; 2) first detections in the des Prairies, l’Acadie and St-François rivers, and 3) no detection in Yamaska and Noire rivers; supporting the extirpation theory [127]. Telemetry data was also used to describe summer feeding habitat preferences and key characteristics include 1) aquatic grass beds of medium to elevated density, mainly Vallisneria americana, Potamogeton pectinatus, P. richardsonii, Elodea canadensis and Heterenthera dubia 2) abundance of Unionidae, Sphaeriidae, Amnicolidae and Dreissenidae 3) water velocity below 0.5 m s-1, 4) sand or fine rock substrate, 5) depth inferior to 4 m and 6) summer temperature average of 20°C [126]. Thorough descriptions of critical habitats for young-of-the-year, sub-adults, and adults can also be found in the recovery plan ([128]. Sectors of the St Lawrence that display similar features may be considered as critical habitats for the species and include the Verchères, Saint-Sulpice, Lavaltrie and Contrecoeur areas, as well as the Boucherville islands [126]. Sightings in the submerged vegetation habitat near the Boucherville islands by adult specimens for feeding have been reported. Winter behaviour has not been properly qualified as telemetry data are lacking. More studies are needed to evaluate potential areas for adults to feed in submerged aquatic vegetation (SAV) such as Saint-Louis and Saint-Pierre lakes, and the La Prairie Basin [129]. The Lavaltrie-Contrecoeur area has been suggested as a potential site for the creation of a wildlife refuge and specimens have been observed in the area during spring and fall every year between 1998 and 2004. The shores of Hervieux Island also represent potentially rich aquatic grass beds for feeding based on the capture of mature specimens in 1999 and 2000 [126]. Both sites may offer qualified spawning grounds, but such utilization has not been confirmed.
3.7.4. Cultivation Activities
The only captive copper redhorse population is held at the Baldwin-Coaticook MELCCFP fish farm facility and originates from captures of wild breeders over the years to maintain a founding captive population to supply juveniles for the stock enhancement program [133]. Access to detailed information is not yet available. The first trials of egg incubation for artificial spawning were carried out in 1990 [132]. A recent study briefly reports on the incubation and rearing of eggs of M. hubbsi and M. carinatum [118]. Securing a breeding stock has been an early recommendation for supporting research and stocking activities for M. hubbsi [132]. Hormonal induction, on-site gamete sampling followed by artificial fertilization and egg disinfection procedures to ensure the preservation of wild fitness of stocking fish [134]. All the fertilized eggs are transferred to the MELCCFP facilities for incubation in exterior ponds and indoor tanks pre-stocking. Recently, Lair et al. [135] presented the rearing protocols of juvenile copper redhorse focusing on feeding of juveniles to enhance growth and skeletal malformation. More recently, the use of cryopreserved semen and genotyping was introduced to increase genetic variation and control inbreeding [136]. These activities are in line with the genetic plan for reproduction of the wild population [137,138].
3.7.5. Conservation Status and Threats
The species was designated Threatened by the COSEWIC in 1987; registered for Québec’s Act Respecting Threatened or Vulnerable Species in 1999; designated endangered by the COSEWIC in 2004 (reconfirmed in 2014); listed under the Canadian Species at Risk Act and as Endangered by the IUCN since December 2007; its habitat is protected by two (2) provincial laws (Environment Quality Act and Act respecting Conservation and Development of Wildlife) and one federal law (Canada’s Fisheries Act), and commercial and game fishing are forbidden since 1995 and 1998 respectively. A 10-year recovery program is available (2012-2022) and a new version that covers 2025-2033 is in press.
Biological characteristics contributing to the vulnerability of M. hubbsi include a late sexual maturity, a highly specialized diet, and a late spawning season (i.e. end of June to early of July). The anatomical specialization for feeding on molluscs may increase risk of extinction due to reduced mollusc abundance reported in the SVA in the last decades and bioaccumulation of heavy metals of existing molluscs beds (Woolnough et al. 2020). The longevity (i.e. 30 years) and late sexual maturity extend recovery speed and affect monitoring frequency, hindering more frequent conservation actions (i.e. sampling, stocking and overall assessment). According to the recovery team, the timing of the spawning season is believed to affect reproductive success with observed record high summer temperatures in the last decade that significantly reduce the water depth available at the spawning beds of the Richelieu River, while a peak of aquatic recreational activities in the proximity of the spawning grounds, due to the occurrence of two consecutive national holidays (St-Jean-Baptiste: 24th of June, and Canada Day 1st of July) create harmful waves and subsequent disruptions of reproductive behaviour. Lastly, very little information is available on the physiological threshold limits of the copper redhorse. All threats to the copper redhorse unique population status are enumerated and described in the recovery plan [128] and many are presented below.
- Water contaminants: Sixty-four municipalities (total population 300 000) are located along the Richelieu River and 70% of its watershed is dedicated to agriculture [118,140]. Extensive work has evaluated its water quality and biological integrity with regards to industrial, urban and agricultural pressure, and the impact of wastewater treatment stations installed. Improvements of the global quality of the ecosystem were noted but pollution from municipalities (wastewater, runoff) and industries increases phosphorus, nitrogen, fecal coliforms, suspended matter, turbidity and chlorophyll-a, hindering the physicochemical and bacterial quality index of the Richelieu River. Water quality is deemed good from its point of entry then gradually decreases, becoming severely impacted where it meets the St. Lawrence River. A water management plan addresses habitat degradation (i.e. habitat loss, fragmentation, aquatic vegetation, and invasive species) [141]. Recommendations focus on improving agricultural practices and the enforcing laws for the riparian strip. Main activities directly associated with habitat degradation include urban development, agricultural drainage, riparian strip destruction, erosion, and shoreline artificialization.
Toxic levels of polychlorinated biphenyls (PBCs) and polychlorinated dibenzo dioxins (PCDDs) from industrial activities are reported in the upstream section of the Richelieu River, exceeding protection thresholds for piscivorous terrestrial fauna by 2.2-fold and 25-fold, respectively. Water quality remains an issue as the watershed drains highly developed agricultural areas, contaminating critical habitat with harmful pesticides, herbicides and insecticides [118,142,143]. The phosphorus threshold limit to mitigate eutrophication is set at 0.03 mg L-1 and mean concentrations of 0.037 mg L-1 were observed upstream the Richelieu River (2001-2003), while a sampling campaign (2011-2013) revealed a 2.4-fold excess of the phosphorus quality criterion at mid-river level (St-Jean-sur-Richelieu), with exceeding values upstream near Sorel and in the Lavaltrie-Contrecoeur sector of the St Lawrence River [141].
Endocrine disrupting chemicals (EDC) in aquatic environment can adversely affect growth, development, and reproduction. Difficulties in natural reproduction of the copper redhorse have been associated with peak levels of these chemicals, especially during its late spawning season (June-July) [144]. Toxicological effects on reproductive processes were first evaluated in M. hubbsi and included 1) impaired gamete maturation due to elevated concentrations of agricultural by-product alkylphenol polyethoxylates and, 2) potential olfactory system obstruction by pesticide atrazine, affecting pheromone perception. Neonicotinoids insecticides, such as clothianidin and thiamethoxam, and 23 other pesticides, have been measured at levels above water quality guideline thresholds for chronic (8.3 ng L-1) and acute (200 ng L-1) exposures [142,145]. Following heavy rain events in the Richelieu River, Marchand et al. [118] observed peak concentrations of both clothianidin (89 ng L-1), and thiamethoxam (32 ng L-1) at spawning sites that frequently exceed federal guidelines. The study highlighted eighteen differentially expressed genes involved in immune function, injury response, digestion and nutrient absorption in early-life stages of M. carinatum and M. hubbsi when exposed to the river waters compared to control. After exposure to the river water early hatching was observed in both groups but a 14% decrease in survival rate was observed in M. hubbsi . Nonetheless, certain factors prohibit establishing a clear relationship between contaminants and early-stages performances and further investigation into the effects of complex contaminant mixtures is needed. These contaminants are recurringly found in elevated concentrations in the Richelieu River, particularly during the spawning season. High concentrations of these pesticides have also been reported in the water of tributaries of the Richelieu River during the spawning season and in the sediment of sampling stations along the Saint-Lawrence River [145,146]. Significant work by Maltais et al. [147,148,149] characterized copper and shorthead redhorse vitellogenin (VTG) to develop a non-invasive biomarker to evaluate exposure to EDC. Such techniques propose interesting approaches for monitoring endangered species but have yet to be incorporated in current monitoring of M. hubbsi.
- 2.
- Invasive species: The Great Lakes-St Lawrence River basin contains the most non-native species of any freshwater system, with n= 180 established invasive species [150]. The spread of invasive tench (Tinca tinca) in the St Lawrence River has raised concerns in Québec was reported by the MELCCFP in 2022 especially regarding the redhorse species, as they share habitats, spawning grounds and dietary preferences. Two other benthivorous species, the round goby Neogobius melanostomus and the tubenose goby (Proterorhinus semilunaris) have also been detected and could exert additional pressure on molluscivorous species such as M. hubbsi and M. carinatum due to their shared diet and possible egg and larvae predation [151]. The presence of Asian carp species (Ctenopharyngodon idella, Hypophthalmichtys nobilis, H. molitrix and Mylopharyngodon piceus) has increased in recent years in Canada [152] but only C. idella is confirmed in Québec. Environmental DNA analysis and captures confirmed the presence of grass carp in M. hubbsi habitat. Grass carp poses threats by 1) occurring in its spawning and feeding grounds; 2) competing for food; 3) being highly performing feeders and 4) contributing to habitat degradation [153]. Black carp (M. piceus), native to East Asia present in the United States, has not yet been detected in Québec, but as it is an avid consumer of bivalves, its invasion would impact the eight bivalve species on which M. hubbsi relies on for feeding [154].
Another concern with invasive species is the introduction of foreign pathogens. Tench is a potential carrier of a harmful pathogen to the copper redhorse, V. campylancristrota (Avlijas et al., 2022; Marcogliese et al., 2009). Other worrying parasites include Biacetabulum (Cestoda, Caryophyllidea) which are tapeworms known to preferably select Catostomidae as hosts; M. anisurum, M. carinatum, M. erythrurum have already been listed as hosts in recent studies (Oros et al. 2020; Uhrovič et al. 2021). All of the data was collected for phylogenetic studies of US rivers. The presence of the Asian fish tapeworm (Schyzocotyle acheilognathi) has increasingly been observed in the Great Lakes and the St Lawrence River in recent years (Marcogliese et al., 2016). Grass carp is the native host of the Asian fish tapeworm and contributes to its spreading which poses threats to M. hubbsi [153].
Last but not least, the invasive zebra mussels (Dreissena polymorpha; Quagga mussel, D. bugensis) are also a concern as they 1) reduce the availability of native molluscs species, 2) affect the bioaccumulation process of contaminants, 3) may affect predator growth via potential poor caloric value, and 4) impact overall habitat quality and availability for molluscivorous species [129]. Although aquatic invasive species have been identified as a key threat to the copper redhorse [128], limited efforts have been deployed to monitor and evaluate the impact of invasive and pathogenic species, with main efforts focused on dispersion risk management. Additionally, low survival rates upon release are common in artificial propagation programs, and such rates may be even lower in systems where invasive species occur. The zebra mussel is now considered a prey item in the diet of the copper redhorse (included in the Dreissenaidae family) but with possibly less nutritional values than other main items included in the species diet.
- 3.
- Habitat fragmentation, loss or degradation: A large proportion of the copper redhorse distribution has been modified by anthropogenic activities. Habitat loss, degradation and fragmentation exert major restrictions to the distribution range, causing population decline and recovery more uncertain [128]. Erosion, shoreline hardening, and construction of dams severely impact M. hubbsi. The Richelieu River is of concern as the quality criteria of 13 mg/l of suspended matter can exceed by more than 20% [141]. Despite policies to improve agricultural practices, the absence of windbreaks, livestock presence and altered riparian vegetation all contribute to shoreline erosion. Erosion accelerates siltation, increases turbidity, can disrupt the food chain by altering photosynthesis, and accentuate settling of sediments on gravel in the river interstices, possibly covering spawning grounds and molluscs beds adversely.
Hydroelectric dams and other flow regulation structures in the St Lawrence River system pose a threat to the recovery of the copper redhorse by fragmenting habitat and impeding migration to potential spawning and feeding grounds [128,129]. The St Ours dam forced the copper redhorse (and other Moxostoma sp.) to spawn in its afterbay offering sub-optimal habitat [132]. The Vianney-Legendre fishway passage that was installed at the site now facilitates migration, but successful passage requires lengthy periods of intense activity, and associated exhaustion increases risk to predation for the non-adults, affects overall fitness, and may impact a range of behaviours [130]. The structure was described and energy expenditure of fish negotiating the passage assessed [155]. Wild spawners are regularly capture for artificial reproduction operations at the site.
Loss of SAV reduces recruitment rates and affects overall fish assemblage, as it acts as a refuge for juveniles and habitat for invertebrate preys [156]. Recently, a major industrial development poses serious threats to the copper redhorse. The construction of a new terminal by the Montreal Port Authorities is expected to result in 24.6 ha of permanent loss of fish habitat, and associated dredging is anticipated to result in a permanent loss of 0.8 ha of critical feeding habitat of adult copper redhorse [157]. Considering the obligation of submitting a comprehensive compensation plan prior to final authorization, an advisory expressed beforehand serious concern about the underestimation of potential impacts [158]. The plan presented by the port authorities aims to create new SAV feeding habitats at a pre-selected location to compensate for the loss of habitat that will involve depth and hydraulic modifications of an extended area and plantation techniques involving substrate modifications and submerged pre-vegetated mats (or fascines). Several case-studies of realigned sites (eco-engineered) that include addition of plant material with or without waterbody modification, display ecological relevance according to Taylor et al. [159]: spawning habitat, and Stamps et al. [160]: feeding habitats. However, these authors argued that improved research and more evidence-based studies are needed to increase the certainty of their positive or net-gain outcomes.
3.7.6. On-Going Conservation Efforts and Research
Overall conservation efforts aimed at the recovery of the copper redhorse all fall under the recovery plan major orientations (awareness, research and monitoring and conservation) and main objectives : 1) improve the habitat conditions required for the completion of the life cycle of the copper redhorse in order to ensure its survival and recovery; 2) sustain the population with stock enhancement activities until natural reproduction reaches self-perpetuating status ; 3) encourage research efforts on the subadults component of the natural population; 4) reduce the effects of anthropic pressures on the copper redhorse territory and habitat ; 5) realize regular monitoring of the population status. The viability and the distribution of M. hubbsi was evaluated [161] and the conclusions that 1) the species faced low probability of extinction in the next 100 years but that the probability that the population could drop below the bar of 250 specimens was very high; 2) stocking of 1+ should be favored instead of 0+ (roughly 3-months old individuals) actually used in the assisted reproduction program conducted by the MELCCFP, in order to increase population size and that 3) a closely related species, the greater redhorse (M. valenciennesi) due to its physiological similarity should be included as a valuable proxy in the conservation efforts aimed at M. hubbsi. This section assesses a selection of current conservation and research efforts in line with the recovery plan to restore M. hubbsi to a naturally spawning state.
3.7.6.1. Feeding/Spawning Habitat Creation/Restoration
Submerged aquatic vegetation (SAV) plays a vital ecological role in freshwater ecosystems by 1) improving water quality; 2) providing essential habitat for numerous organisms; 3) contributing to nutrient cycling and 4) buffering wave effects [162]. The creation of SAV can significantly increase fish community diversity. Success relies on coverage extent, light diffusion, plant species, adequate substrate, control of invasive species, and local hydrodynamics [163]. Efficient restoration should rely on native species and extensive knowledge on distribution dynamics and habitat needs [164]. Initiatives to protect aquatic habitats and improve water quality in the Richelieu River watershed have been conducted through the action plan of a consultation and development committee [141] and the Green and Blue Corridor Plan [165]. The latter aims to restore shores using native vegetation, connecting natural areas of the watershed, and encouraging good agricultural and municipal practices regarding waste treatment and compound usage.
Restoration success in vegetated marine and freshwater habitats should consider plant-bivalve interactions, which would also greatly benefit the copper redhorse, a molluscivorous species. Proper co-restoration (plant-bivalve) could increase survival, persistence and resilience of targeted species, promoting the recovery of associated biodiversity and ecosystem services [166]. The MELCCFP is orienting a large part of their efforts in the evaluation, protection and restoration of SVA located in the critical and essential habitat of the copper redhorse and supporting NGO’s and local ZIP committee's actions in fluvial ecosystems protection and rehabilitation.
3.7.8. Genetic Studies (Including Environmental DNA)
A study published in 2006 confirmed the cooper redhorse’s single population theory and observed the persistence of a high genetic diversity possibly due to the long generation time and advanced age at maturity (i.e. 10 years) [138]. In 2024, a new report restates that the population does not suffer from a loss in diversity or alarming inbreeding levels and this despite an important reduction in the global population (60% from 2000-2010 with an observed stability between 2010-2020) [167]. Five recommendations are issued: 1) increase the number of effective broodstock in the wild population; 2) control inbreeding by using individual pedigree and pairing matrix for artificial reproduction efforts; 3) increase the number of broodstock fish participating in artificial reproduction (>30 broodstock fish is a minimum to maintain genetic diversity); 4) reevaluate the stocking program objectives (last update was in 2004) in order to keep artificial contribution > 20% of the global population and 5) maintain a captive population (that should be renewed frequently to avoid domestication risks) until the species achieves recovery. The MELCCFP is systematically gathering genetic information on all breeders captured for artificial reproduction efforts.
Environmental DNA (eDNA) is the genetic material that can be extracted from the environment frequented by a species to provide evidence of occupation of a given habitat. Recent research suggests a relationship between abundance of eDNA and species abundance [168, 169) but the strength of the relationship will depend on environmental and technical parameters. Lacoursière-Roussel et al. [168,170] suggests caution while interpreting both detection and abundance is required in natural environment surveys. The use of eDNA offers the potential to improve detection per unit effort over traditional methods and is increasingly considered a complementary tool for achieving conservation objectives targeting elusive, rare threatened, vulnerable, exotic and invasive species [169]. Environmental DNA metabarcoding (multiple taxa assessment) was used to document spatial distribution in fish communities at a small geographic scale in a large river system (St Lawrence River) by Berger et al. [171]. A total of 67 species were detected among which figured several Catostomidae species including M. macrolepidotum, M. carinatum and M. anisurum and invasive Tinca tinca. Although M. hubbsi remained undetected during the sampling campaign (13-15 Sept. 2017), the technology’s application for monitoring M. hubbsi’s presence is however unquestionable, especially for future more directed sampling activities, i.e. prior/post spawning migrations to Richelieu watershed. In a recent report, MELCCFP detected the presence of M. hubbsi at the mouth of the Yamaska and Noire rivers as well as in areas of confirmed frequentation by the copper redhorse. According to Langlois et al. [172] of the many eDNA detection methodology, real-time polymerase chain reaction (qPCR), is the most accessible and cost-effective. However, considerable variation in data quality, rigor and reliability currently undermine regulatory and policy acceptance of eDNA as a reliable species inventory and monitoring method. Efforts to increase the full mitochondrial genome sequencing of more taxa are needed to support the development of more robust primer/probe test designs. Mitochondrial genome (mt-eDNA) and nuclear genomes (nu-eDNA) have been widely used for eDNA studies, with the former targeted in most studies on animals and plants. McCauley et al. [173] strongly suggest using nu-eDNA and mt-eDNA simultaneously due to differences in persistence in the natural environment. Currently, only nu-eDNA is used for copper redhorse e-DNA studies. Long-read shotgun metagenomic sequencing was recently discussed as a new approach for species identification [173,174] with increased precision for abundance estimations [175].
3.7.9. Telemetry Surveys
Telemetry techniques (radio, acoustic or satellite technology) are used to remotely measure the position of tagged fishes (use of implanted or fixed transmitter), coupled to measurement of biotic and abiotic conditions of the aquatic ecosystems provides invaluable fish-habitat associations at spatial and temporal scales [176]. Telemetry surveys, to locate the habitats frequented by the copper redhorse in the St Lawrence River and its tributaries, was conducted in 2008 [126]. Telemetry data enables interventions, appropriate conservation measures and the suggestion of management methods. Environmental variables associated with habitat selection were used to build a multivariate habitat model to produce maps representing the probability of occurrence of the species in spring, summer and fall [127]. This information was then used to define critical habitat areas for adults in the St Lawrence River. In 2019, six adult fish originating from the artificial reproduction program, were implanted with acoustic transmitters and released in the Richelieu River to provide track movement, behavior and survival data [135].
3.7.10. Ecological River Rehabilitation or Restoration
The global concept of river restoration is the re-establishment of natural physical processes (e.g. variation of flow and sediment transport), features (e.g. particle size and river shape) and physical habitats of a river system. Common river restoration goals include bank stabilization, channel reconfiguration, dam removal/retrofit, fish passage, floodplain reconnection, flow modification, habitat improvement, land acquisition and sometimes species management [177]. Ecological river restoration or rehabilitation activities aim to recover and improve the ecological integrity of degraded watersheds by reestablishing processes that support healthy ecosystems (fish habitat, connectivity, water quality, invasive species eradication etc.) [177,178]. Ultimately these activities may actively support the recovery of targeted species and/or biological communities. Approaches include bioengineering using living plant material, modifying waterbed morphology, and process-based restoration that promotes river function rather than physical characteristics i.e. remeandering, naturalization, restoration, dam removal, erosion and pollution control etc. Protecting and restoring the habitat of the copper redhorse is supported by Québec’s Protection Policy for Lakeshores, Riverbanks, Littoral Zones and Floodplains covering its range but is inconsistently implemented. The MELCCFP offers participative training to municipalities and landowners, but guidelines are not systematically reinforced.
3.7.11. Conservation Aquaculture and Restocking Activities
Reintroduction of imperilled species in their native range through captive-breeding requires research to establish rearing protocols and infrastructure, and outcome (success, failure) and effectiveness indicators remain difficult to identify [35,179]. Reintroduction success criteria are survival, spawning and recruitment, and these variables can strongly be impacted by environmental parameters. Challenges to reintroductions of economically less valuable SARA-listed fish species include 1) limited knowledge on life history and key ecology and biology data for management plans (i.e. fecundity, age structure, mortality, recruitment) and 2) scattered populations with unique sampling requirements [26].
A simple protocol to produce juveniles of M. hubbsi was proposed by Branchaud et al [132]. The artificial reproduction program prior to 2018 was essentially based on wild spawners captures at the Vianney-Legendre structure located at the St-Ours dam. Incubation and on-growing in land-based facilities for restocking in the nursery habitats of the Richelieu River and at the other side of the Vianney-Legendre structure in the summer and fall. Yearly production objectives were 500 000 larvae, 15 000 young-of-the-year originating from 100 families to yield a population of 4 000 mature individuals over a period of 20 years [134] and are currently under review. Actual yields reported by DFO [129] are: 760 000 0+, 99 000 young-of-the-year and 111 1+ in the Richelieu River from 2012-2018. The goal is to rebuild a naturally reproducing population while preserving 90% of initial genetic diversity over a 100-year period. Due to the late age at maturity of the species (10 years), it is too soon to evaluate the impact of stocking activities on the reproductive population status. Captures of sub-adults and adults originating from the earlier stocking efforts have been recorded through genetic analysis, but juvenile survival rates and actual state of the population are unknown.
In 2018, the MELCCFP initiated a structured conservation aquaculture program aimed at juvenile production based on a small and unique population of captive breeders currently held at their facility. The goal is to support juvenile production which has varied increasingly from year to year attributably to yearly capture success of wild breeders. Fish husbandries in laboratory settings are rare for SARA-listed freshwater fish, with only the copper redhorse and the Atlantic whitefish (Coregonus huntsman) benefiting from such installations [26]. At this stage, refinement of cultivation techniques throughout all phases of the production cycle, to ensure numerous and healthy juvenile production to reach sustainable reestablishment of the population, are still on-going at the MELCCFP fish farm facilities. Efforts should also focus on improving fitness of the juveniles to be introduced during the restocking campaigns such as: environmental enrichment, life-skills training, development of “soft release” protocols and disease control and monitoring [180]. Recently, Bannister-Marchand [161] strongly suggested that the 0+ juvenile mostly released during the stock enhancement activities of the MELCCFP should instead target 1+ juveniles to improve their resilience in the natural environment.
3.7.12. Cryopreservation and Cryobanking
Cryopreservation and cryobanking of genetic material from endangered species including fish, will secure the possibility of reestablishing the original populations and preserve their biodiversity value [181]. Research on copper redhorse sperm cryopreservation was initiated in 2012 due to increasing recapture rates of mature males and spawners scarcity. The program objective was to develop and optimize protocols for artificial fertilization with cryopreserved milt, increase the genetic pool by creating families with less-related parents, and counter asynchronism between males and females [136]. Various cryopreservation methods (n=12) using different combinations of diluent at various osmolality, cryoprotectant at different concentrations and various straw size were tested. The selected cryopreservation protocol enabled the creation of a cryobank managed that is currently used by the MELCCFP for their artificial propagation program, with genotyping and matrix pairing assistance to control inbreeding levels. Based on our evaluation further efforts to generate more comprehensive data and optimize the actual cryopreservation protocol are warranted. The other Moxostoma species involved in the development of a cryopreservation technique are the sicklefin and the robust redhorses occurring in the U.S.A [182].
3.8. Future Conservation and Research Orientation for Moxostoma sp.
At the basis of understanding and predicting imperiled freshwater fish species response to environmental changes and stressors, and in our attempt to provide key conservation insights, we wish to underline the many promises held by the field of conservation physiology as presented by Cooke et al. [183]. The increased availabilities of juveniles originating from artificial reproduction efforts should provide resources to facilitate the initiation of physiology research to support recovery efforts aimed at redhorses in particular the copper redhorse. In addition to current initiatives previously covered in this manuscript, we suggest two intertwined and warranted research orientations in conservation physiology: effects of climate change and health status indicators.
3.8.1. Effects of Climate Change
Global climate change is progressively reshaping freshwater ecosystems and affecting fish populations, largely due to species-specific physiological constraints [184]). This research field is insufficiently addressed for many imperiled freshwater fish species, increasing extinction likelihoods due to anthropogenic threats being aggravated by climate change. Carosi et al. [185] analyzed the changes in freshwater fish species distribution due to increasing water temperature and hydrological instability events that occurred during a well-defined period of climate warming. Shifts in upstream-downstream distribution highlighted species-specific dispersal ability and adaptability to climate change. Integrating this approach into redhorses restoration actions should help identify optimal range of habitats, stressor thresholds, cause-and-effect relationships, providing key background information to support crucial management decisions. In addition, invasive species introduction and spreading in novel areas are generally considered to be accelerated due to climate change effectively reducing ecosystem resilience, acting as a threat multiplier for indigenous species and in particular species at risk [186,187]. For example, the critical thermal maxima (CTmax) value helps predict the impacts of global warming on its distribution and adaptation [1]. The CTmax value of the copper redhorse is not currently available. Values for the shorthead and golden redhorses exist (see previous specific sections).
3.8.2. Fish Health
Conservation practices often rely on habitat protection or restoration, but these efforts should be assisted by close monitoring of populations and individuals’ health status. Easily accessible morphometric measurements (size, weight, and condition indexes at different ages) are often included in such programs but may be insufficient to estimate the efficiency of conservation investment. The potential relevance of physiological biomarkers as management tools has only recently been implemented in addressing conservation challenges and should be considered given that fish health is highly sensitive to environmental modifications and an important driver of fitness as reviewed by Mallet et al. [188]. Many markers have been developed to monitor fish health status and accumulated stress at different periods of their life-history. Ideally, biomarkers should be selected according to their ability to discriminate between stress types at the population or individual level. Sequential steps through which stressors impact fish are: 1) health, proceeding to 2) fitness and ultimately to 3) population dynamics. Primary, secondary and tertiary stress responses used as health indicators include physiological, behavioral, morphological and genetic components and include corticosteroids, blood parameters, oxidative stress indicators, inflammatory response, heat shock proteins, osmotic and ion regulation, morphological and organo-somatic changes, behavior and fats. Other indicators include external abnormalities, parasites, disease and genetic responses.
Blood hematological and biochemical markers have been successfully used to monitor stress status of fish according to abiotic factors variations [189,190,191] and may be basis of fish health monitoring toolboxes. Environmental stress, particularly from contaminants, may cause oxidative stress and inflammatory response detrimental to the immune system in fish. Most markers inform on the actual status of individuals but may be less informative on health and stress history in the individual over the last few months or years [192]. Biochemical markers are available and can provide information on the trajectories of fish health and stress status [193,194]. For example, by-products of lipid oxidation will advise on recent stress because of fast lipid turn-over, while oxidized proteins or DNA will accumulate over a much longer period. Modern biochemical and molecular tools are increasingly available to better follow population health status, and managers and stakeholders should prioritize such investments in future conservation strategies. For copper redhorse restoration, initiating health monitoring activities is crucial both on exposed wild population and captive stocks from which juveniles are released for restocking purposes.
4.0. Conclusion
This review underlines the needs for sustained conservation efforts aimed at the genus Moxostoma and more urgently at the critically endangered copper redhorse. In addition to the maintenance and strengthening of the actual actions aimed at Moxostoma sp. conservation conducted nationwide, we suggest an unranked list of recommendations for research:
- Acquire better understanding of microhabitat spawning preferences and develop models to better predict how environmental variables and habitat modification affect reproduction quality, timing and success.
- Explore novel approaches in vegetation engineering aimed at rehabilitation/creation/restoration and protection of SAV habitats.
- Apply more telemetry monitoring efforts to assess the use of essential habitats.
- Increase regulations for the reduction/control of pesticides and other contaminants in essential habitats and document their effects.
- Implement more efforts aimed at the problematic of invasive and exotic species.
- Refine/develop eDNA detection techniques for vulnerable Moxostoma species, molluscs (gasteropods and bivalves) and plant species found in essential and critical habitats.
- Set forth an extensive research program aimed at copper redhorse conservation physiology aimed at species responses to climate change and health status.
- Identification/creation of additional protected areas and protection of essential habitats.
Author Contributions
Conceptualization, N.R.L.F.; methodology, N.R.L.F.; validation, N.R.L.F.; formal analysis, C.D.J.; investigation, NRLF, CDJ, PUB, SL.; resources, N.R.L.F.; data curation, CDJ, NRLF.; writing—original draft preparation, N.R.L.F., C.D.J., P.U.B., H.C.M., S.L. writing—review and editing, N.R.L.F., C.D.J.; visualization, N.R.L.F.; supervision, N.R.L.F.; project administration, N.R.L.F.; funding acquisition, N.R.L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This review was financially supported by a private research contract no. P5101 granted to the first author (NR Le François) and administered by the Institut de Recherche en Biologie Végétale (IRBV).
Acknowledgments
The authors would like to thank H.-F. Ellefsen of the DFO for their constructive remarks. We acknowledge the collaboration of the Administration du Port de Montréal (Environment division) for providing useful information regarding the terminal expansion project.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SAV | Submerged aquatic vegetation |
| COVABAR | Comité de concertation et de valorisation du bassin de la rivière Richelieu |
| SARA | Species at Risk Act (Canada) |
| DFO | Department of Fisheries and Oceans (Canada) |
| IUCN | International Union for Conservation of Nature |
| COSEWIC | Committee on the Status of Endangered Wildlife in Canada |
| MELCCFP | Ministère de l’Environnement, de la Lutte contre les Changements Climatiques, de la Faune et des Parcs |
References
- Desforges, J.E.; Clarke, J.; Harmsen, E.J.; Jardine, A.M.; Robichaud, J.A.; Serré, S.; Chakrabarty, P.; Bennett, J.R.; Hanna, D.E.L.; Smol, J.P.; Rytwinski, T.; et al. The Alarming State of Freshwater Biodiversity in Canada. Can. J. Fish. Aquat. Sci. 2022, 79, 352–365. [Google Scholar] [CrossRef]
- Moyle, P.B.; Leidy, R.A. Loss of Biodiversity in Aquatic Ecosystems: Evidence from Fish Faunas. In Conservation Biology; Fiedler, P.L., Jain, S.K., Eds.; Springer: Boston, MA., USA, 1992; pp. 127–169. [Google Scholar] [CrossRef]
- Cambray, J.A.; Bianco, P.G. 1998. Freshwater Fish in Crisis, a Blue Planet Perspective. Ital. J. Zool. 1998, 65, 345–356. [Google Scholar] [CrossRef]
- Ricciardi, A.; Neves, R.J. and Rasmussen, J.B. Impending Extinctions of North American Freshwater Mussels (Unionoida) following the Zebra Mussel (Dreissena polymorpha) Invasion. J. Animal Ecol. 1998, 67, 613–619. [Google Scholar] [CrossRef]
- Tickner, D.; Opperman, J.J.; Abell, R.; Acreman, M.; Arthington, A.H.; Bunn, S.E.; Cooke, S.J.; Dalton, J.; Darwall, W.; Edwards, G.; et al. Bending the Curve of Global Freshwater Biodiversity Loss: an Emergency Recovery Plan. Biosci. 2020, 70, 330–342. [Google Scholar] [CrossRef]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ Warning to Humanity on the Freshwater Biodiversity Crisis. Ambio 2021, 50, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Sayer, C.A.; Fernando, E.; Jimenez, R.R.; Macfarlane, N.B.W.; Rapacciuolo, G.; Böhm, M.; Brooks, T.M.; Contreras-MacBeath, T.; Cox, N.A.; Harrisson, I.; et al. One-Quarter of Freshwater Fauna Threatened with Extinction. Nature 2025, 638, 138–145. [Google Scholar] [CrossRef]
- Quist, M.C.; Spiegel, J.R. Population Demographics of Catostomids in Large River Ecosystems: Effects of Discharge and Temperature on Recruitment Dynamics and Growth. Riv. Res. Appl. 2012, 28, 1567–1586. [Google Scholar] [CrossRef]
- Rypel, A.L.; Saffarinia, P.; Vaughn, C.C.; Nesper, L.; O’Reilly, K.; Parisek, C.A.; Miller, M.L.; Moyle, P.B.; Fangue, N.A.; Bell-Tilcock, M.; et al. Goodbye to “Rough Fish”: Paradigm Shift in the Conservation of Native Fishes. Fisheries 2021, 46, 605–616. [Google Scholar] [CrossRef]
- Childress, E.S.; McIntyre, P.B. Life History Traits and Spawning Behavior Modulate Ecosystem-Level Effects of Nutrient Subsidies from Fish Migrations. Ecosphere 2016, 7, e01301. [Google Scholar] [CrossRef]
- Booth, M.T.; Hairston, N.G., Jr.; Flecker, A.S. Consumer Movement Dynamics as Hidden Drivers of Stream Habitat Structure: Suckers as Ecosystem Engineers on the Night Shift. Oikois 2020, 129, 194–208. [Google Scholar] [CrossRef]
- Cooke, S.J.; Bunt, C.M.; Hamilton, S.J.; Jennings, C.A.; Pearson, M.P.; Cooperman, M.S.; Markle, D.F. Threats, Conservation Strategies, and Prognosis for Suckers (Catostomidae) in North America: Insights from Regional Case Studies of a Diverse Family of Non-Game Fishes. Biol. Conserv. 2005, 121, 317–331. [Google Scholar] [CrossRef]
- Harris, P.M.; Hubbard, G.; Sandel, M. Catostomidae: Suckers. In Freshwater Fishes of North America: Volume 1: Petromyzontidae to Catostomidae; Warren, M.L., Burr, B.M., Tomerelli, J.R., Eds.; Johns Hopkins University Press: Baltimore, USA, 2014; pp. 451–502. ISBN 978-1421412016. [Google Scholar]
- Ricciardi, A.; Rasmussen, J.B. Extinction rates of North American freshwater fauna. Conserv. Biol. 1999, 13, 1220–1222. [Google Scholar] [CrossRef]
- Wikipedia. Available online: https://en.wikipedia.org/wiki/Moxostoma#:~:text=Teretulus%20Rafinesque%2C%201820-,Species,%2C%201820)%20(Silver%20redhorse) (accessed on 7 March 2025).
- Scott, W.E.; Crossman, E.J. Suckers-Catostomidae. In Freshwater Fishes of Canada; Fisheries Research Board of Canada: Ottawa, Canada, 1973; pp. 563–567. [Google Scholar]
- Mongeau, J.R.; Dumont, P.; Cloutier, L. La Biologie du Suceur Cuivré (Moxostoma hubbsi) Comparée à Celle de Quatre Autres Espèces de Moxostoma (M. anisurum, M. carinatum, M. macrolepidotum et M. valenciennesi). Can. J Zool. 1992, 70, 1354–1363. [Google Scholar] [CrossRef]
- Hatry, C.; Thiem, J.D.; Hatin, D.; Dumont, P.; Smokorowski, K.E.; Cooke, S.J. Fishway Approach Behaviour and Passage of Three Redhorse Species (Moxostoma anisurum, M. carinatum, and M. macrolepidotum) in the Richelieu River, Quebec. Environ. Biol. Fishes, 2016, 99, 249–263. [Google Scholar] [CrossRef]
- Jelks, H.L.; Walsh, S.J.; Burkhead, N.M.; Contreras-Balderas, S.; Diaz-Pardo, E.; Hendrickson, D.A.; Lyons, J.; Mandrak, N.E.; McCormick, F.; Nelson, J.S.; et al. Conservation Status of Imperiled North American Freshwater and Diadromous Fishes. Fisheries 2008, 33, 372–407. [Google Scholar] [CrossRef]
- IUCN Red List. Moxostoma valenciennesi. Available online: https://www.iucnredlist.org/species/202259/18228653 (accessed on 7 March 2025).
- IUCN Red list. Moxostoma duquesnii. Available online: https://www.iucnredlist.org/species/18227213/18234003 (accessed on 7 March 2025).
- UICN Red List. Moxostoma erythrurum. Available online: https://www.iucnredlist.org/species/202165/76571620 (accessed on 7 March 2025).
- UICN Red List. Moxostoma anisurum. Available online: https://www.iucnredlist.org/species/202159/18230929 (accessed on 7 March 2025).
- IUCN Red List. Moxostoma macrolepidotum. Available online: https://www.iucnredlist.org/species/202253/18228990 (accessed on 7 March 2025).
- IUCN Red List. Moxostoma carinatum. Available online: https://www.iucnredlist.org/species/202161/18234655 (accessed on 7 March 2025).
- Lamothe, K.A.; Drake, D.A.R.; Pitcher, T.E.; Broome, J.E.; Dextrase, A.J.; Gillespie, A.; Mandrak, N.E.; Poesch, M.S.; Reid, S.M.; Vachon, N. Reintroduction of Fishes in Canada: A Review of Research Progress for SARA-Listed Species. Environ. Rev. 2019, 27, 575–599. [Google Scholar] [CrossRef]
- IUCN Red List. Moxostoma robustustum. Available online: https://www.iucnredlist.org/species/202257/19033267 (accessed on 7 March 2025).
- Department of Fisheries and Oceans. Management plan for the river redhorse (Moxostoma carinatum) in Canada; Species at Risk Act Management Plan Series; Fisheries and Oceans Canada: Ottawa, Canada, 2018; 48 p. [Google Scholar]
- Boyko, A. (Department of Fisheries and Oceans Canada) Personal communication, 2020.
- DFO. Recovery Strategy and Action Plan for the Black Redhorse (Moxostoma duquesnei) in Canada. Species at Risk Act Recovery Strategy Series. Fisheries and Oceans: Ottawa, Canada, 2022. https://publications.gc.ca/site/eng/9.906661/publication.html. (accessed on 7 March 2025).
- IUCN Red List. Moxostoma hubbsi. Available online: https://www.iucnredlist.org/species/13917/58135857 (accessed on 7 March 2025).
- Tognelli, M.F.; Máiz-Tomé, L.; Kraus, D.; Lepitzki, D.; Mackie, G.; Morris, T.; Carney, J.; Alfonso, N.; Tonn, B.; Cox, N.A.; Smith, K.A. Freshwater Key Biodiversity Areas in Canada. In Informing Species Conservation and Development Planning in Freshwater Ecosystems; IUCN: Arlington, USA, 2017; 42 p. [Google Scholar] [CrossRef]
- Port de Montréal. Available online: https://www.port-montreal.com/en/the-port-of-montreal/projects/terminal-in-contrecoeur (accessed on 7 March 2025).
- Day, J.L.; Jacobs, J.L.; Rasmussen, J. Considerations for the Propagation and Conservation of Endangered Lake Suckers of the Western Lake Suckers of the Western United States J. Wildl. Manag. 2017, 8, 301–312. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Rytwinski, T.; Taylor, J.J.; Bennett, J.R.; Drake, D.A.R.; Martel, A.; Cooke, S.J. Can Conservation Targets for Imperilled Freshwater Fishes and Mussels be Achieved by Captive Breeding and Release Programs? A Systematic Map Protocol to Determine Available Evidence. Environ Evid. 2019, 8, 16. [Google Scholar] [CrossRef]
- Taylor, J.J.; Rytwinski, T.; Bennett, J.R.; Smokorowski, K.E.; Lapointe, N.W.R.; Janusz, R.; Clarke, K.; Tonn, B.; Walsh, J.C.; Cooke, S.J. The Effectiveness of Spawning Habitat Creation or Enhancement for Substrate-Spawning Temperate Fish: a Systematic Review. Environ. Evid. 2019, 8, 19. [Google Scholar] [CrossRef]
- Schmucker, C.M.; Blümle, A.; Schell, L.K.; Schwarzer, G.; Cabrera, L.; von Elm, E.; Briel, M.; Meerpöhl, J.J. Systematic Review Finds that Study Data Not Published in Full Text Article Have Unclear Impact on Meta-Analysis Results in Medical Research. PloS One 2017, 12, e0176210. [Google Scholar] [CrossRef]
- Buxton, R.T.; Bennett, J.R.; Reid, A.J.; Shulman, C.; Cooke, S.J.; Francis, C.M.; Nyboer, E.A.; Pritchard, G.; Binley, A.D.; Avery-Gomm, S.; et al. Key Information Needs to Move from Knowledge to Action for Biodiversity Conservation in Canada. Biol. Conserv. 2021, 256, 108983. [Google Scholar] [CrossRef]
- Eberhard, E.K.; Wilcove, D.S.; Dobson, A.P. Too Few, Too Late: U.S. Endangered Species Act Undermined by Inaction and Inadequate Funding. PloS One 2022, 17, e0275322. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, H.E.; Gentry, R.R.; Halpern, B.S. Conservation Aquaculture: Shifting the Narrative and Paradigm of Aquaculture’s Role in Resource Management. Biol. Conserv. 2017, 215, 162–168. [Google Scholar] [CrossRef]
- Ridlon, A.D.; Wasson, K.; Waters, T.; Adams, J.; Donatuto, J.; Fleener, G.; Froehlich, H.; Govender, R.; Kornbluth, A.; Lorda, J.; et al. Conservation Aquaculture as a Tool for Imperiled Marine Species: Evaluation of Opportunities and Risks for Olympia Oysters, Ostrea lurida. PLoS One 2021, 16, e0252810. [Google Scholar] [CrossRef]
- Robitaille, J.M.; Bérubé, M.; Gosselin, A.; Baril, M.; Beauchamp, J.; Boucher, J.; Dionne, S.; Legault, M.; Mailhot, Y.; Ouellet, B.; et al. Recovery Strategy for the Striped bass (Morone saxatilis), St-Lawrence Estuary Population, Canada; Species at risk act recovery strategy series; Fisheries and Oceans Canada: Ottawa, Canada, 2011; 51p. [Google Scholar]
- Valiquette, E.; Harvey, V.; Pelletier, A.-M. Mise à Jour des Connaissances sur l’Identification, la Description et l’Utilisation Spatio-Temporelle des Habitats du Bar Rayé (Morone saxatilis) de la Population du Fleuve Saint-Laurent, Québec. Secrétariat canadien de consultation scientifique (SCCS), 2017; Doc. Rech. 2292-4272; 2017/005, Région du Québec. https://publications.gc.ca/site/eng/9.833505/publication.html.
- Gaudreau, C.M. (ministère de l’Environnement, de la Lutte contre les Changements Climatique, Faune et Parc MELCCFP) Personal communication, 2024.
- Muttray, A.F.; Muir, D.C.G.; Tetreault, G.R.; McMaster, M.E.; Sherry, J.P. Spatial Trends and Temporal Declines in Tissue Metals/Metalloids in the Context of Wild Fish Health and the St.Clair River Area of Concern. J. Great Lakes Res. 2021, 47, 900–915. [Google Scholar] [CrossRef]
- Page, L.M.; Johnston, C.E. Spawning in the Creek Chubsucker, Erimyzon oblongus, With a Review of Spawning Behavior in Suckers (Catostomidae). Environ. Biol. Fish. 1990, 27, 265–272. [Google Scholar] [CrossRef]
- Kwak, T.J.; Skelly, T.M. Spawning Habitat, Behavior, and Morphology as Isolating Mechanisms of the Golden Redhorse, Moxostoma erythrurum, and the Black Redhorse, M. duquesnei, Two Syntopic Fishes. Environ. Biol. Fish. 1992, 34, 127–137. [Google Scholar] [CrossRef]
- Cooke, S.J.; Bunt, C.M. Spawning and Reproductive Biology of the Greater Redhorse, Moxostoma valenciennesi, in the Grand River, Ontario. Can. Field-Nat. 1999, 113, 497–502. [Google Scholar] [CrossRef]
- Jenkins, R.E.; Jenkins, D.J. 1980. Reproductive Behavior of the Greater Redhorse, Moxostoma valenciennesi, in the Thousand Islands region. Can. Field-Nat. 1980, 94, 426–430. [Google Scholar] [CrossRef]
- Grabowski, T.B.; Isely, J.J. Seasonal and Diel Movements and Habitat Use of Robust Redhorse in the Lower Savannah River, Georgia and South Carolina. Trans. Am. Fish. Soc. 2006, 135, 1145–1155. [Google Scholar] [CrossRef]
- Catalano, M.J.; Bozek, M.A. Influence of Environmental Variables on Catostomid Spawning Phenology in a Warmwater River. Am. Mid. Nat. 2015, 173, 1–16. [Google Scholar] [CrossRef]
- Reid, S.M.; Wilson, C.C.; Carl, L.M.; Zorn, T.G. Species Traits Influence the Genetic Consequences of River Fragmentation of Two Co-Occurring Redhorse (Moxostoma) Species. Can. J. Fish. Aquat. Sci. 2008, 65, 1892–1904. [Google Scholar] [CrossRef]
- Reid, S.M.; Wilson, C.C.; Mandrak, N.E.; Carl, L.M. Population Structure and Genetic Diversity of Black Redhorse (Moxostoma duquesnei) in a Highly Fragmented Watershed. Conserv. Genet. 2008, 9, 531–546. [Google Scholar] [CrossRef]
- Jenkins, R.E.; Burkhead, N.M. . : . 1994; 1084 pp. In Freshwater Fishes of Virginia; American Fisheries Society: Bethesda, MD, USA, 1994; 1084 p. [Google Scholar]
- Reid, S.M. Comparison of Scales, Pectoral Fin Rays and Opercles for Age Estimation of Ontario Redhorse, Moxostoma, Species. Can. Field-Nat. 2007, 121, 29–34. [Google Scholar] [CrossRef]
- Lippé, C.; Dumont, P.; Bernatchez, L. Isolation and Identification of 21 Microsatellite Loci in the Copper Redhorse (Moxostoma hubbsi; Catostomidae) and their Variability in Other Catostomids. Mol. Ecol. Notes 2004, 4, 638–641. [Google Scholar] [CrossRef]
- Lippé, C.; Dumont, P.; Bernatchez, L. High Genetic Diversity and No Inbreeding in the Endangered Copper Redhorse, Moxostoma hubbsi (Catostomidae, Pisces): the Positive Sides of a Long Generation Time. Mol. Ecol. 2006, 15, 1769–1780. [Google Scholar] [CrossRef]
- Reid, S.M.; Wilson, C.C. PCR-RFLP Based Diagnostic Tests for Moxostoma Species in Ontario. Conserv. Genet. 2006, 7, 997–1000. [Google Scholar] [CrossRef]
- Lacoursière-Roussel, A.; Rosabal, M.; Bernatchez, L. Estimating Fish Abundance and Biomass from eDNA Concentrations: Variability Among Capture Methods and Environmental Conditions. Mol. Ecol. Res. 2016, 16, 1401–1414. [Google Scholar] [CrossRef]
- Rider, H.J.; Janosik, A.M. Using Environmental DNA Metabarcoding to Assess the Spatiotemporal Occurrence of the Imperiled River Redhorse (Moxostoma carinatum) in the Escambia-Conecuh River System of Florida and Alabama. USA. Environ. Biol. Fish 2024, 107, 1593–1608. [Google Scholar] [CrossRef]
- MELCCFP. Available online: https://mffp.gouv.qc.ca/documents/faune/GM_catostomides_MFFP.pdf (accessed on 7 March 2025).
- Moxostoma.com. Available online: https://moxostoma.com/redhorse-id-downloads/ (accessed on 7 March 2025).
- Page, L.M.; Burr, B.M. A Field Guide to Freshwater Fishes of North America North of Mexico; Houghton Mifflin Company: Boston., USA, 1991; 432 p. [Google Scholar]
- Hatry, C.; Thiem, J.D.; Binder, T.R.; Hatin, D.; Dumont, P.; Stamplecoskie, K.M.; Molina, J.M.; Smokorowski, K.E.; Cooke, S.J. Comparative Physiology and Relative Swimming Performance of Three Redhorse (Moxostoma spp.) Species: Associations With Fishway Passage Success. Physiol. Biochem. Zool. 2014, 87, 148–159. [Google Scholar] [CrossRef]
- Spiegel, J.R.; Quist, M.C.; Morris, J.E. Trophic Ecology and Gill Raker Morphology of Seven Catostomid Species in Iowa Rivers. J. Appl. Ichthyol. 2011, 27, 1159–1164. [Google Scholar] [CrossRef]
- Vachon, N. Écologie des Juvéniles 0+ et 1+ de Chevalier Cuivré (Moxostoma hubbsi), une Espèce Menacée, Comparée à Celle des Quatre Autres Espèces de Moxostoma (M. anisurum, M. carinatum, M. macrolepidotum, M. valenciennesi) dans le Système de la Rivière Richelieu. MSc thesis, Université du Québec à Montréal, Montréal, Québec, Canada; 1999.
- Reid, S.M. Timing and Demographic Characteristics of Redhorse Spawning Runs in Three Great Lakes Basin Rivers. J. Freshw. Ecol. 2006, 21, 249–258. [Google Scholar] [CrossRef]
- Hatry, C.; Thiem, J.D.; Hatin, D.; Dumont, P.; Smokorowski, K.E.; Cooke, S.J. Fishway Approach Behaviour and Passage of Three Redhorse Species (Moxostoma anisurum, M. carinatum, and M. macrolepidotum) in the Richelieu River, Quebec. Environ. Biol. Fish. 2016, 99, 249–263. [Google Scholar] [CrossRef]
- Uhrovič, D.; Oros, M.; Kudlai, O.; Choudhury, A.; Scholz, T. Molecular Evidence of Three Closely Related Species of Biacetabulum Hunter, 1927 (Cestoda: Caryophyllidea): a Case of Recent Speciation in Different Fish Hosts (Catostomidae)? Parasitology 2021, 148, 1040–1056. [Google Scholar] [CrossRef]
- Fraker, M.E.; Keitzer, S.C.; Sinclair, J.S.; Aloysius, N.R.; Dippold, D.A.; Yen, H.; Arnold, J.G.; Daggupati, P.; Johnson, M.-V.V.; Martin, J.F.; Robertson, D.M.; Sowa, S.P.; White, M.J.; Ludsin, S.A. Projecting the Effects of Agricultural Conservation Practices on Stream Fish Communities in a Changing Climate. Sci. Total Environ. 2020, 747, 141112. [Google Scholar] [CrossRef]
- Reid, S.M. Age, Growth and Mortality of Black Redhorse (Moxostoma duquesnei) and Shorthead Redhorse (M. macrolepidotum) in the Grand River, Ontario. J. Appl. Ichthyol. 2009, 25, 178–183. [Google Scholar] [CrossRef]
- Howlett, D.T. Age, Growth and Population Structure of Black Redhorse (Moxostoma duquesnei) and Golden Redhorse (Moxostoma erythrurum) in Southwest Missouri. MSc thesis Southwest Missouri State University, Springfield, Missouri. 1999. [Google Scholar]
- Bunt, C.M.; Heiman, T.; Mandrak, N.E. Ontogeny of Larval and Juvenile Black Redhorse (Moxostoma duquesnei). Copeia 2013, 121–126. [Google Scholar] [CrossRef]
- Bunt, C.M.; Mandrak, N.E. .; Eddy, D.C.; Choo-Wing, S.A.; Heiman, T.; Taylor, E. Habitat Utilization, Movement and Use of Groundwater Seepages by Larval and Juvenile Black Redhorse Moxostoma duquesnei. Environ. Biol. Fish. 2013, 96, 1281–1287. [Google Scholar] [CrossRef]
- Curry, K.D.; Spacie, A. Differential Use of Stream Habitat by Spawning Catastomids. Am. Mid. Nat. 1984, 111, 267–279. [Google Scholar] [CrossRef]
- Reid, S.M.; Mandrak, N.E.; Carl, L.M.; Wilson, C.C. Influence of Dams and Habitat Conditions on the Distribution of Redhorse (Moxostoma) Species in the Grand River Watershed, Ontario. Environ. Biol. Fish. 2008, 81, 111–125. [Google Scholar] [CrossRef]
- Young, J.A.M.; Koops, M.A. Population modelling of black redhorse (Moxostoma duquesni) in Canada. DFO Can. Sci. Advis. Sec. Res. Doc. 2014: 14pp. Available online: https://publications.gc.ca/collections/collection_2014/mpo-dfo/Fs70-5-2014-020-eng.pdf (accessed on 12 March 2025).
- Brown, B.A. Comparative Life-histories of some species of redhorse, subgenus moxostoma, genus Moxostoma. PhD Thesis, Indiana State University, 1984. Available online: https://scholars.indianastate.edu/etds/2349/.
- Bowman, M.L. Life History of the Black Redhorse, Moxostoma duquesnei (Lesueur), in Missouri. Trans. Am. Fish. Soc. 1970, 99, 546–559. [Google Scholar] [CrossRef]
- Page, L.M.; Burr, B.M. Peterson Field Guide to Freshwater Fishes; Houghton Mifflin Company: Boston, MA, 2011; 663 p. [Google Scholar]
- Holm, E.; Mandrak, N.E.; Burridge, M.E. The ROM Field Guide to Freshwater Fishes of Ontario; Royal Ontario Museum: Toronto, ON, Canada, 2009; 462 p. [Google Scholar]
- Bouvier, L.D.; Burridge, M.E.; Glass, W.R.; Caskenette, A. Information in support of a Recovery Potential Assessment of Black Redhorse (Moxostoma duquesnei) in Canada. DFO Can. Sci. Advis. Sec. Res. Doc. 2021, 38 p. Available online: https://www.dfo-mpo.gc.ca/csas-sccs/Publications/ResDocs-DocRech/2021/2021_021-eng.html (accessed on 12 March 2025).
- Beckman, D.W.; Howlett, D.T. 2013. Otolith Annulus Formation and Growth of Two Redhorse Suckers (Moxostoma: Catastomidae). Copeia 2013, 3, 390–395. [Google Scholar] [CrossRef]
- Reid, S.M.; Glass, W.R. Precision and Comparability of Black Redhorse (Moxostoma duquesnei) Age Estimates Using Scales, pectoral Fin Rays, and Opercle Bones. Can. Manuscr. Rep. Fish. Aquat. Sci. 2014, 9 p. Available online: https://publications.gc.ca/collections/collection_2014/mpo-dfo/Fs97-4-3034-eng.pdf (accessed on 12 March 2025).
- Bunt, C.M.; van Poorten, B.T.; Wong, L. Denil Fishway Utilization Patterns and Passage of Several Warmwater Species Relative to Seasonal, Thermal and Hydraulic Dynamics. Ecol. Freshw. Fish 2001, 10, 212–219. [Google Scholar] [CrossRef]
- Department of Fisheries and Oceans. Black Redhorse (Moxostoma duquesnei): Recovery Strategy and Action Plan 2022. Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/recovery-strategies/black-redhorse-2022.html#toc0 (accessed on 12 March 2025).
- Blazer, V.S.; Iwanowicz, D.D.; Sperry, A.J.; Iwanowicz, L.R.; Alvarez, D.A.; Brightbill, R.A.; Smith, G.; Foreman, W.T.; Manning, R. Reproductive Health Indicators of Fishes from Pennsylvania Watersheds: Association with Chemicals of Emerging Concern. Environ. Monit. Assess. 2014, 186, 6471–6491. [Google Scholar] [CrossRef]
- Rosencranz, J.; Cuddington, K.; Brook, M.; Koops, M.A.; Drake, D.A. Data-Limited Models to Predict River Temperatures for Aquatic Species at Risk. Can. J. Fish. Aquat. Sci. 2020, 78, 1268–1277. [Google Scholar] [CrossRef]
- iNaturalist.org. Available online: https://www.inaturalist.org/taxa/106212-Moxostoma-erythrurum (accessed on 12 March 2025).
- Meyer, W.H. Life History of Three Species of Redhorse (Moxostoma) in the Des Moines River, Iowa. Trans. Am. Fish. Soc. 1962, 91, 412–419. [Google Scholar] [CrossRef]
- Fuiman, L.A.; Witman, D.C. Descriptions and Comparisons of Catostomid Fish Larvae: Catastomus catastomus and Moxostoma erythrurum. Trans. Am. Fish. Soc. 1979, 108, 604–619. [Google Scholar] [CrossRef]
- Moyer, G.R.; Shaw, R.; Wertz, T. Mitochondrial DNA Barcoding Reveals First Documented Evidence of Golden Redhorse (Moxostoma erythrurum) in the Susquehanna River, Pennsylvania. Northeast. Nat. 2021, 28, 149–155. [Google Scholar] [CrossRef]
- Reash, R.J.; Seegert, G.L.; Goodfellow, W.L. Experimentally-Derived Upper Thermal Tolerances for Redhorse Suckers: Revised 316(A) Variance Conditions at Two Generating Facilities in Ohio. Environ. Sci Policy 2000, 3 (Suppl. 1), 191–196. [Google Scholar] [CrossRef]
- Keplinger, B.; Hedrick, J.; Blazer, V.S. Temporal Trends in Macroscopic Indicators of Fish Health in the South Branch Potamac River. North Am. J. Fish. Manag. 2022, 42, 277–294. [Google Scholar] [CrossRef]
- Al-Ansari, A.M.; Saleem, A.; Kimpe, L.E.; Sherry, J.P.; McMaster, M.E.; Trudeau, V.L.; Blais, J.M. Bioaccumulation of the Pharmaceutical 17α-Ethinylestradiol in Shorthead Redhorse Suckers (Moxostoma macrolepidotum) from the St. Clair River, Canada. Environ. Pollut. 2010, 158, 2566–2571. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Nelson, P.A. Lissorchi s macropharynx n. sp. (Digenea: Lissorchiidae) From the Shorthead Redhorse, Moxostoma macrolepidotum (Lesueur)(Osteichthyes: Catostomidae). J. Parasitol. 1998, 84, 1196–1202. [Google Scholar] [CrossRef]
- DeBofsky, A.; Xie, Y.; Jardine, T.D.; Hill, J.E.; Jones, P.D.; Giepsy, J.P. Effects of the Husky Oil Spill on Gut Microbiota of Native Fishes in the North Saskatchewan River, Canada. Aquat. Toxicol. 2020, 229, 105658. [Google Scholar] [CrossRef] [PubMed]
- Moravec, F.; Levron, C.; de Buron, I. Morphology and Taxonomic Status of Two Little-Known Nematode Species Parasitizing North American Fishes. J Parasitol. 2011, 97, 297–304. [Google Scholar] [CrossRef]
- Doyle, J.R.; Al-Ansari, A.M.; Gendron, R.L.; White, P.A.; Blais, J.M. A method to estimate sediment ingestion by fish. Aquat. Toxicol. 2011, 103, 121–127. [Google Scholar] [CrossRef]
- Bister, T.J.; Willis, D.W.; Brown, M.L.; Jordan, S.M.; Newmann, R.M.; Quist, M.C.; Guy, C.S. Proposed Standard Weight (Ws) Equations and Length Categories for 18 Warmwater Nongame and Riverine Fish Species. North Am. Fish. Manag. 2000, 20, 570–574. [Google Scholar] [CrossRef]
- Buynak, G.L.; Mohr, H.W. Larval Development of the Shorthead Redhorse (Moxostoma macrolepidotum) from the Susquehanna River. Trans. Am. Fish. Soc. 1979, 108, 161–165. [Google Scholar] [CrossRef]
- den Haas, T.C.; Bouvier, L.D.; Mandrak, N.E. Identifying and Comparing Annuli across Calcified Structures in Shorthead Redhorse (Moxostoma macrolepidotum). Can. Manuscr. Rep. Fish. Aquat. Sci. 2013, 3023, 10. Available online: https://publications.gc.ca/site/eng/475020/publication.html (accessed on 12 March 2025).
- Coughlan, D.J.; Baker, B.K.; Barwick, D.H.; Garner, A.B.; Doby, W.H. Catastomi Fishes of the Wateree River, South Carolina. Southeastern Naturalist 2007, 6, 305–320. [Google Scholar] [CrossRef]
- Edge, E.N.; Paukert, C.P.; Lobb III, M.D.; Landwer, B.H.P.; Bonnot, T.W. Seasonal Selection Habitat by Spotted Bass and Shorthead Redhorse in a Regulated River in the Midwest, USA. 2000, 36, 1087–1096. River Research and Applications 2000, 36, 1087–1096. [Google Scholar] [CrossRef]
- Healy, B.D. Conservation Assessment for the Greater Redhorse (Moxostoma valenciennesi). USDA Forest Service, Eastern Region, April 2002.
- Harris, P.M.; Mayden, R.L.; Espinosa Pérez, H.S.; Garcia de Leon, F. Phylogenetic Relationships of Moxostoma and Scartomyzon (Catostomidae) Based on Mitochondrial Cytochrome b Sequence Data. J. Fish Biol. 2005, 61, 1433–1452. [Google Scholar] [CrossRef]
- Clements, M.D.; Bart Jr, H.L.; Hurley, D.L. A Different Perspective on the Phylogenetic Relationships of the Moxostomatini (Cypriniformes: Catostomidae) Based on Cytochrome-b and Growth Hormone Intronsequences. Mol. Phylogenet. Evol. 2012, 63, 159–167. [Google Scholar] [CrossRef]
- Bunt, C.M.; Cooke, S.J. Ontogeny of Larval Greater Redhorse (Moxostoma valenciennesi). Am. Mid. Nat. 2004, 151, 93–100. [Google Scholar] [CrossRef]
- Bunt, C.M.; Cooke, S.J. Post-Spawn and Habitat Use by Greater Redhorse, Moxostoma valenciennesi. Ecol. Freshw. Fish 2001, 10, 57–60. [Google Scholar] [CrossRef]
- Tomsic, C.A.; Granata, T.C.; Murphy, R.P.; Livchak, C.J. Using a Coupled Eco-Hydrodynamic Model to Predict Habitat for Target Species Following Dam Removal. Ecol. Eng. 2007, 30, 215–230. [Google Scholar] [CrossRef]
- Cheng, F.; Granata, T. Sediment Transport and Channel Adjustments Associated with Dam Removal: Field Observations. Water Resour. Res. 2007, 43, W03444. [Google Scholar] [CrossRef]
- Parker, B.J. Updated Status of the River Redhorse, Moxostoma carinatum, in Canada. Canadian Field-Naturalist 1988, 102, 140–146. [Google Scholar] [CrossRef]
- DFO Management Plan for the River Redhorse (Moxostoma carinatum) in Canada. Species at Risk Act Management Plan Series. Fisheries and Oceans Canada, Ottawa. 2018. Available online: https://publications.gc.ca/site/eng/9.834906/publication.html (accessed on 12 March 2025).
- French, J.R.P. How Well Can Fishes Prey on Zebra Mussels in Eastern North America. Fisheries. 1993, 18, 13–19. [Google Scholar] [CrossRef]
- Butler, S.E.; Wahl, D.H. Movements and Habitat Use of River Redhorse (Moxostoma carinatum) in the Kankakee River, Illinois. Copeia 2017, 105, 734–742. [Google Scholar] [CrossRef]
- Straight, C.A.; Jakson, C.R.; Freeman, B.J.; Freeman, M.C. Diel Patterns and Temporal Trends in Spawning Activities of Robust Redhorse and River Redhorse in Georgia, Assessed Using Passive Acoustic Monitoring. Trans. Am. Fish. Soc. 2015, 144, 563–576. [Google Scholar] [CrossRef]
- Preville, N.M.; Snyder, E.B.; O’Keefe, D.; Hanshue, S.; Russel, A.; Radecki, J. Habitat Use of the Threatened River Redhorse (Moxostoma carinatum) in the Grand River, MI, USA. Aquat. Sci. 2022, 84, 43. [Google Scholar] [CrossRef]
- Marchand, H.; Barst, B.D.; Boulanger, E.; Vachon, N.; Houde, M.; Xia, J.; Liu, P.; Ewald, J.D.; Bayen, S.; Liu, L.; Head, J.A. 2022. Exposure to contaminated river water is associated with early hatching and dysregulation of gene expression in early life stages of the endangered copper redhorse (Moxostoma hubbsi). Environ. Toxicol. Chem. 2022, 41, 1950–1966. [Google Scholar] [CrossRef] [PubMed]
- Gagnon-Poiré (Ministère de l’Environnement, de la Lutte contre les Changements Climatique, Faune et Parc MELCCFP) Personal communication, 2024.
- Cooke, S.J. University of Waterloo, Ontario, Canada. Personal communication. 2024.
- Vachon, N. Guide et Clé d’Identification des Juvéniles de Chevaliers (genre Moxostoma) du Québec. Société de la faune et des parcs du Québec, Direction de l’aménagement de la faune de Montréal, de Laval et de la Montérégie, Longueuil, Rapport technique 16-14F, 2003 26 p. Available online: https://mffp.gouv.qc.ca/documents/faune/chevalier_guide_cle.pdf (accessed on 12 March 2025).
- McAllister, D.E. The Copper Redhorse, Moxostoma hubbsi A disappearing Canadian endemic. Biodiversity 2011, 2, 27–28. [Google Scholar] [CrossRef]
- Eastman, J.T. The Pharyngeal Bones and Teeth of Catostomid Fishes. Am. Mid. Nat. 1977, 97, 68–88. [Google Scholar] [CrossRef]
- Doosey, M.H.; Bart, H.L. Morphological Variation of the Palatal Organ and Chewing Pad of Catostomidae (Teleostei: Cypriniformes). J. Morphol. 2011, 272, 1092–1108. [Google Scholar] [CrossRef] [PubMed]
- DFO Recovery Strategy for the Copper Redhorse (Moxostoma hubbsi) in Canada. Species at Risk Act Recovery Strategy Series. Fisheries and Oceans Canada, Ottawa, Canada. 2012, 60 pp. Available online: www.registrelep-sararegistry.gc.ca/virtual_sara/files/plans/rs_chevalier_cuivre_copper_redhorse_0312_eng.pdf (accessed on 12 March 2025).
- Vachon, N.; Chagnon, Y. Caractérisation de la population de chevalier cuivré (Moxostoma hubbsi) du fleuve Saint-Laurent (secteur Lavaltrie-Contrecoeur) à partir des captures fortuites d'un pêcheur commercial en 1999, 2000 et 2001. Report prepared for Direction de l'aménagement de la faune de Montréal, de Laval et de la Montérégie, ministère des Ressources naturelles et de la faune et des parcs. 2004. [Google Scholar] [CrossRef]
- Gariépy, S. Déplacements, Domaines Vitaux, Sélection et Caractérisation des Habitats des Chevaliers Cuivrés Adultes dans le Système du Fleuve Saint-Laurent, Québec, Canada. Master thesis-Université du Québec à Rimouski, Rimouski, 2008 https://semaphore.uqar.ca/id/eprint/58/1/Simone_Gariepy_juin2008.pdf.
- Équipe de Rétablissement du Chevalier Cuivré. Plan de Rétablissement du Chevalier Cuivré (Moxostoma hubbsi) au Québec — 2012-2017, Ministère des Ressources naturelles et de la Faune du Québec, Faune Québec. 2012, 55 p. Available online: https://mffp.gouv.qc.ca/documents/faune/plan-retablissement-chevalier.pdf (accessed on 14 March 2025).
- DFO. Report on the Progress of Recovery strategy Implementation for the Copper Redhorse (Moxostoma hubbsi) in Canada for the Period 2012 to 2018. Species at Risk Act Recovery Strategy Report Series. Fisheries and Oceans Canada, Ottawa. 2022, 47 pp. Available online: https://www.canada.ca/content/dam/eccc/documents/pdf/lep-sara/2022/20220526-CopperRedhorseProgressReport-eng.pdf (accessed on 14 March 2025).
- Silva, A.T.; Hatry, C.; Thiem, J.D.; Gutowsky, L.F.G.; Hatin, D.; Zhu, D.Z.; Dawson, J.W.; Katopodis, C.; Cooke, S.J. Behaviour and Locomotor Activity of a Migratory Catostomid during Fishway Passage. PlosOne 2015, 10, e0123051. [Google Scholar] [CrossRef]
- Vachon, N.; Velasquez-Medina, S.; Grondin, P. Motilité des spermatozoïdes du chevalier cuivré dans les différents traitements de cryopréservation en 2013. Ministère des Forêts, de la Faune et des Parcs (MFFP), Direction de la gestion de la faune de l’Estrie, de Montréal, de la Montérégie et de Laval, Secteur de la faune, Rapport technique 2019, 24 p. Available online: https://mffp.gouv.qc.ca/documents/faune/Motilite_spermatozoide_chevalier_cuivre_cryopreservation.pdf (accessed on 14 March 2025).
- Branchaud, A.; Gendron, A.D. 1993. Artificial Spawning and Rearing of the Copper Redhorse, Moxosoma hubbsi (Teleostei: Catostomidae). Can. Field-Nat. 1996, 107, 279–282. Available online: https://www.biodiversitylibrary.org/item/108204#page/295/mode/1up (accessed on 17 March 2025). [CrossRef]
- Couillard, M.-A.; Poiré-Gagnon, R. MELCCFP, personal communications 2024.
- Vachon, N. Reproduction Artificielle, Ensemencements et Suivi de la Population du Chevalier Cuivré (Moxostoma hubbsi) en 2018. Direction de la gestion de la faune de l'Estrie, de Montréal, de la Montérégie et de Laval, Secteur des opérations régionales, ministère des Forêts, de la Faune et des Parcs. 2021, Rapport technique 16-57, 43 p. Available online: https://mffp.gouv.qc.ca/nos-publications/reproduction-artificielle-ensemencements-suivi-population-chevalier-cuivre/ (accessed on 17 March 2025).
- Lair, S.; Vergneau-Grosset, C.; Coulombe, C.; Vachon, N.; Couillard, M.-A. Conservation of the Endangered Copper Redhorse (Moxostoma hubbsi): Implication of the Aquarium du Québec. In Proceedings of the International Association for Aquatic Animal Medecine (IAAAM) annual conference, Salt Lake City, Utah, USA, 21-24 May 2025; Available online: https://www.vin.com/doc/?id=11463515&meta=Generic (accessed on 17 March 2025).
- Vachon, N.; Velasquez-Medina, S.; Grondin, P. Motilité des Spermatozoïdes du Chevalier Cuivré dans les Différents Traitements de Cryopréservation en 2013. Report prepared for Ministère des Forêts, de la Faune et des Parcs (MFFP), Direction de la gestion de la faune de l’Estrie, de Montréal, de la Montérégie et de Laval, Secteur de la faune, Rapport technique 2019, 24 p. Available online: https://mffp.gouv.qc.ca/documents/faune/Motilite_spermatozoide_chevalier_cuivre_cryopreservation.pdf (accessed on 17 March 2025).
- Bernatchez, L. Considérations Génétiques et Protocole de Reproduction Relatifs au Plan de Rétablissement du Chevalier Cuivré (Moxostoma hubbsi). Ministère des Ressources Naturelles, de la Faune et des Parcs, Direction de l’Aménagement de la Faune de Montréal, de Laval et de la Montérégie, Longueuil et Pêches et Océans Canada, région du Québec. 2004, Rapport technique 16-22, 38 p. Available online: https://mffp.gouv.qc.ca/documents/faune/chevalier_cuivre_reproduction.pdf (accessed on 17 March 2025).
- Lippé, C.; Dumont, P.; Bernatchez, L. High Genetic Diversity and No Inbreeding in the Endangered Copper Redhorse Moxostoma hubbsi, (Catostomidae, Pisces): the Positive Sides of a Long Generation Time. Mol. Ecol. 2006, 15, 1769–1780. [Google Scholar] [CrossRef]
- Woolnough, D.A.; Bellamy, A.; Longstaff Hummel, S.; Annis, M. Environmental Exposure of Freshwater Mussels to Contaminants of Emerging Concern: Implications for Species Conservation. J. Great Lakes Res. 2020, 46, 1625–1638. [Google Scholar] [CrossRef]
- Giroux, I.; Simoneau, M. État de l’Écosystème Aquatique du Bassin Versant de la Rivière Nicolet: faits saillants 2004-2006. Ministère du Développement Durable, de l’Environnement et des Parcs, Direction du suivi de l’état de l’environnement, 2008, ISBN 978-2-550-53174-6, 21 p. Available online: https://belsp.uqtr.ca/id/eprint/1372/1/Giroux%20et%20al._2008_Écosystème_aquatique_Nicolet.pdf (accessed on 17 March 2025).
- COVABAR. . Plan Directeur de l’Eau – Plan d’Action 2015-2020 du Bassin Versant de la Rivière Richelieu et de la Zone Saint-Laurent, Beloeil. 2015. Available online: www.covabar.qc.ca/documents/PDE/Portrait/Planaction2015-2020.pdf (accessed on 17 March 2025).
- Montiel-León, J.M.; Munoz, G.; Duy, S.V.; Do, D.T.; Vaudreuil, M.A.; Goeury, K.; Guillemette, F.; Amyot, M.; Sauvé, S. Widespread Occurrence and Spatial Distribution of Glyphosate, Atrazine, and Neonicotinoids Pesticides in the St. Lawrence and Tributary Rivers. Environ. Pollut. 2019, 250, 29–39. [Google Scholar] [CrossRef]
- Patoine, M.; Hébert, S.; D’Auteuil-Potvin, F. Water Quality Trends in the Last Decade for Ten Watersheds Dominated by Diffuse Pollution in Québec, Canada. Water Sci. Technol. 2012, 65, 1095–1101. [Google Scholar] [CrossRef]
- De Lafontaine, Y.; Gilbert, N.; Dumouchel, F.; Brochu, C.; Moore, S.; Pelletier, E.; Dumont, P.; Branchaud, A. Is Chemical Contamination Responsible for the Decline of the Copper Redhorse (Moxostoma hubbsi), an Endangered Fish Species, in Canada? Sci. Total Environ. 2002, 298, 25–44. [Google Scholar] [CrossRef]
- Giroux, I.; Hébert, S.; Berryman, D. Qualité de l’Eau du Saint-Laurent de 2000 à 2014: Paramètres Classiques, Pesticides et Contaminants Émergents. Nat. Can. 2016, 140, 26–34. [Google Scholar] [CrossRef]
- Giroux, I. Pesticides dans les Sédiments de Cours d’Eau au Québec-Échantillonnages Exploratoires de 2018-2021. MELCCFP, Direction générale du suivi de l’état de l’environnement, 2023, 38 p. Available online: https://www.environnement.gouv.qc.ca/eau/flrivlac/pesticides.htm (accessed on 17 March 2025).
- Maltais, D.; Roy, L.R. Purification and Partial Characterization of Vitellogenin from Shorthead Redhorse (Moxostoma macrolepidotum) and Copper Redhorse (Moxostoma hubbsi) and Detection in Plasma and Mucus with a Heterologous Antibody. Fish Physiol. Biochem. 2009, 35, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Maltais, D.; Roy, R.L.; Couillard, C.M. Hybrid ELISAs for Vitellogenins of the Endangered Copper Redhorse Moxostoma hubbsi and the Shorthead Redhorse Moxostoma macrolepidotum (Cypriniformes, Catostomidae). Ecotoxicol. Environ. Saf. 2010, 73, 883–892. [Google Scholar] [CrossRef]
- Maltais, D.; Roy, R.L. Effects of Nonylphenol and Ethinylestradiol on Copper Redhorse (Moxostoma hubbsi), an Endangered Species. Ecotoxicol. Environ. Saf. 2014, 108, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Pagnucco, K.S.; Maynard, G.A.; Fera, S.A.; Yan, N.D.; Nalepa, T.F.; Ricciardi, A. The Future of Species Invasions in the Great Lakes-St. Lawrence River Basin. J. Great Lakes Res. 2015, 4, 96–107. [Google Scholar] [CrossRef]
- Kipp, R.; Ricciardi, A. Impacts of the Eurasian Round Goby (Neogobius melanostomus) on Benthic Communities in the Upper St. Lawrence River. Can. J. Fish. Aquat. Sci. 2012, 69, 469–486. [Google Scholar] [CrossRef]
- Hernandez, C.; Bougas, B.; Perreault-Payette, A.; Simard, A.; Côté, G.; Bernatchez, L. 60 specific eDNA qPCR assays to detect invasive, threatened, and exploited freshwater vertebrates and invertebrates in Eastern Canada. Environ. DNA 2020, 2, 373–386. [Google Scholar] [CrossRef]
- Pouliot, R.; Morissette, O. Risques de Dispersion des Carpes Asiatiques dans les Tributaires du Fleuve Saint-Laurent, Aspects Théoriques, Évaluation Préliminaire de la Franchissabilité des Obstacles pour les Carpes Asiatiques et Actions Pouvant être Mises en Oeuvre pour Contribuer à la Protection des Eaux Intérieures, Ministère des Forêts, de la Faune et des Parcs, Direction de l’expertise sur la faune aquatique, 2019,163 p. Available online: https://belsp.uqtr.ca/id/eprint/1469/1/Pouliot%20%26%20Morissette_risques_dispersion_carpes_asiatiques_St-Laurent.pdf (accessed on 17 March 2025).
- Morrissette, O.; Paradis, Y.; Pouliot, R.; Lecomte, F. Spatio-Temporal Changes in Littoral Fish Community Structure Along the St. Lawrence River (Québec, Canada) Following Round Goby (Neogobius melanostomus) invasion. Aquat. Inv. 2018, 13, 501–512. [Google Scholar] [CrossRef]
- Thiem, J.D.; Binder, T.R.; Dumont, P.; Hatin, D.; Hatry, C.; Katopodis, C.; Stamplecoskie, K.M.; Cooke, S.J. Multispecies Fish Passage Behaviour in a Vertical Slot Fishway on the Richelieu River, Quebec, Canada. River Res. Appl. 2013, 29, 582–592. [Google Scholar] [CrossRef]
- Miller, J.W.; Kocovsky, P.M.; Wiegmann, D.; Miner, J.G. Fish Community Responses to Submerged Aquatic Vegetation in Maumee Bay, Western Lake Erie. N. Am. J. Fish. Manag. 2018, 38, 623–629. [Google Scholar] [CrossRef]
- IAAC-AEIC Canada. Available online: https://iaac-aeic.gc.ca/050/documents/p83969/145788F.pdf (accessed on 20 March 2025).
- SNAP Québec.org. Available online: https://snapquebec.org/wp-content/uploads/2021/04/Science-advice-copper-redhorse-2021.pdf (accessed on 20 March 2025).
- Taylor, J.J.; Rytwinski, T.; Bennett, J.R.; Smokorowski, K.E.; Lapointe, N.W.R.; Janusz, R.; Clarke, K.; Tonn, B.; Walsh, J.C.; Cooke, S.J. The Effectiveness of Spawning Habitat Creation or Enhancement for Substrate-Spawning Temperate Fish: A Systematic Review. Environ. Evid. 2019, 8, 19. [Google Scholar] [CrossRef]
- Stamp, T.; West, E.; Colclough, S.; Plenty, S.; Ciotti, B.; Robbins, T.; Sheehan, E. 2022. Suitability of Compensatory Saltmarsh Habitat for Feeding and Diet of Multiple Estuarine Fish Species. Fish. Manag. Ecol. 2022, 30, 44–55. [Google Scholar] [CrossRef]
- Bannister-Marchand, N. Étude de la viabilité et de la distribution du chevalier cuivré (Moxostoma hubbsi). MSc in Biology, Université de Sherbrook, Sherbrooke, QC Canada. 2023. Available online: https://savoirs.usherbrooke.ca/handle/11143/21327?show=full (accessed on 20 March 2025).
- Botrel, M.; Maranger, R. Global Historical Trends and Drivers of Submerged Aquatic Vegetation Quantities in Lakes. Glob. Chang. Biol. 2023, 29, 2493–2509. [Google Scholar] [CrossRef]
- Looby, A.; Reynolds, L.K.; Reinhardt, C.; Martin, C.W. Submerged Aquatic Vegetation Patch Size Affects Fish Communities in a Turbid-Algal Lake. Front. Conserv. Sci. 2021, 2, 657691. [Google Scholar] [CrossRef]
- Angradi, T.R.; Pearson, M.S.; Bolgrien, D.W.; Bellinger, B.J.; Starry, M.A.; Reschke, C. Predicting Submerged Aquatic Vegetation Cover and Occurrence in a Lake Superior Study. J. Great Lakes Res. 2013, 39, 536–546. [Google Scholar] [CrossRef]
- Plan St-Laurent. Available online: https://www.planstlaurent.qc.ca/en/water-quality/improving-water-quality-2011-2016-projects/nonpoint-source-pollution-2011-2016/brochure-richelieus-corridor-vert-et-bleu.
- Gagnon, K.; Rinde, E.; Bengil, E.G.T.; Carugati, L.; Christianen, M.J.A.; Danovaro, R.; Gambi, C.; Govers, L.L.; Kipson, S.; Meysick, L.; et al. Facilitating Foundation Species: the Potential for Plant-Bivalve Interactions to Improve Habitat Restoration. J. Appl. Ecol. 2020, 57, 1161–1179. [Google Scholar] [CrossRef]
- Couillard, M.-A.; Côté, G.; Garant, D.; Houle, C. État génétique du chevalier cuivré: constats et recommandations pour le plan de production. Ministère de l’Environnement, de la Lutte contre les Changements Climatiques, de la Faune et des Parcs. Gouvernement du Québec. 2024. Available online: https://www.cubiq.ribg.gouv.qc.ca/notice?id=p%3A%3Ausmarcdef_0001330116&locale=fr (accessed on 20 March 2025).
- Lacoursière-Roussel, A.; Rosabal, M.; Bernatchez, L. Estimating Fish Abundance and Biomass From eDNA Concentrations: Variability Among Capture Methods and Environmental Conditions. Mol. Ecol. Resour. 2016, 16, 1401–1414. [Google Scholar] [CrossRef]
- Evans, N.T.; Shirey, P.D.; Wieringa, J.G.; Mahon, A.R.; Lamberti, G.A. Comparative Cost and Effort of Fish Distribution Detection via Environmental DNA Analysis and Electrofishing. Fisheries 2017, 42, 90–99. [Google Scholar] [CrossRef]
- Lacoursière-Roussel, A.; Deiner, K. Environmental DNA is Not the Tool by Itself. Fish Biol. 2019, 98, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.S.; Hernandez, C.; Laporte, M.; Côté, G.; Paradis, Y.; Kameni, T.D.W.; Normandeau, E.; Bernatchez, L. Fine-Scale Environmental Heterogeneity Shapes Fluvial Fish Communities as Revealed by eDNA Metabarcoding. Environ. DNA 2020, 2, 647–666. [Google Scholar] [CrossRef]
- Langlois, V.S.; Allison, M.J.; Bergman, L.C.; To, T.A.; Helbing, C.C. The Need for Robust qPCR-based eDNA Detection Assays in Environmental Monitoring and Species Inventory. Environ. DNA 2020, 3, 519–527. [Google Scholar] [CrossRef]
- McCauley, M.; Koda, S.A.; Loesgen, S.; Duffy, D.J. Multicellular Species Environmental DNA (eDNA) Research Constrained by Overfocus on Mitochondrial DNA. Sci. Tot. Environ. 2024, 912, 169550. [Google Scholar] [CrossRef]
- Koda, S.A.; McCauley, M.; Farrel, J.A.; Duffy, I.J.; Duffy, F.G.; Loesgen, S.; Whilde, J.; Duffy, D.J. A Novel eDNA Approach for Rare Species Monitoring: Application of Long-Read Shotgun Sequencing to Lyns rufus Soil Pawprints. Biol. Conserv. 2023, 287, 110315. [Google Scholar] [CrossRef]
- Portik, D.M.; Brown, C.T.; Pierce-Ward, N.T. Evaluation of Taxonomic Classification and Profiling Methods for Long-Read Shotgun Metagenomic Sequencing Datasets. BM Bioinform. 2022, 23, 541. [Google Scholar] [CrossRef]
- Brownscombe, J.W.; Griffin, L.P.; Brooks, J.L.; Danylchuk, A.J.; Cooke, S.J.; Midwood, J.D. Applications of Telemetry to Fish Habitat Science and Management. Can. J. Fish. Aquat. Sci. 2022, 79, 1347–1359. [Google Scholar] [CrossRef]
- Wohl, E.; Lane, S.N.; Wilcox, A.C. The Science and Practice of River Restoration. Water Resour. Res. 2015, 51, 5974–5997. [Google Scholar] [CrossRef]
- Li, P.; Li, D.; Sun, X.; Chu, Z.; Xia, T.; Zheng, B. Application of Ecological Restoration Technologies for the Improvement of Biodiversity and Ecosystem in the River. Water 2022, 14, 1402. [Google Scholar] [CrossRef]
- Cochran-Biederman, J.L.; Wyman, K.E.; French, W.E.; Loppnow, G.L. Identifying Correlates of Success and Failure of Native Freshwater Fish Reintroductions: Native Freshwater Fish Reintroduction. Conserv. Biol. 2015, 29, 175–186. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, W.; Li, Y.; Yuan, X.; He, X.; Zhao, H.; Mo, J.; Lin, J.; Yang, L.; Liang, B.; et al. Physical Enrichment for Improving Welfare in Fish Aquaculture and Fitness of Stocking Fish: a Review of Fundamentals, Mechanisms and Applications. Aquaculture 2023, 574, 739651. [Google Scholar] [CrossRef]
- Mayer, I.; Psenicka, M. Conservation of Teleost Fishes: Application of Reproductive Technologies. Theriogenology Wild 2024, 100078. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Available online: https://www.fws.gov/project/cryopreservation-lab (accessed on 20 March 2025).
- Cooke, S.J.; Sack, L.; Franklin, C.E.; Farrell, A.P.; Beardall, J.; Wikelski, M.; Chown, S.L. What is Conservation Physiology? Perspectives on an Increasingly Integrated and Essential Science. Conserv. Physiol. 2013, 1, cot001. [Google Scholar] [CrossRef]
- Whitney, J.A.; Al-Chokhachy, R.; Bunnell, D.B.; Caldwell, C.A.; Cooke, S.J.; Elliason, E.J.; Rogers, M.; Lynch, A.J.; Paukert, C.P. Physiological Basis of Climate Change Impacts on North American inland Fishes. Fisheries 2016, 41, 332–345. [Google Scholar] [CrossRef]
- Carosi, A.; Padula, R.; Ghetti, L.; Lorenzoni, M. Endemic Freshwater Fish Range Shifts Related to Global Climate Changes: A Long-Term Study Provides Some Observational Evidence for the Mediterranean Area. Water 2019, 11, 2349. [Google Scholar] [CrossRef]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Chang. Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pysek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Mallet, M.C.; Thiem, J.D.; Butler, G.L.; Kennard, M.J. A Systematic Review of Approaches to Assess Fish Health Responses to Anthropogenic Threats in Freshwater Ecosystems. Conserv. Physiol. 2024, 12, coae022. [Google Scholar] [CrossRef]
- Casanovas, P.; Walker, P.; Johnston, H.; Johnston, C.; Symonds, J.E. Comparative Assessment of Blood Biochemistry and Haematology Normal Ranges Between Chinook Salmon (Oncorhynchus tshawytscha) From Seawater and Freshwater Farms. Aquaculture 2021, 537, 736464. [Google Scholar] [CrossRef]
- Jeong, H.; Kang, M.; Cha, S.-Y.; Byun, J.; Kim, J.; Baek, J.W.; Park, J.J.; Shin, S.R.; Kim, H.J.; Lee, J.S.; Shin, Y.K.; Cheon, Y.-P. Usefulness of Clustering Blood Biochemical Markers to Assess Thermal Stress and Acclimation in Red Seabream, Pagrus major. Aquaculture 2021, 545, 737197. [Google Scholar] [CrossRef]
- Seibel, H.; Baabmann, B.; Rebl, A. Blood will tell: what hematological analyses can reveal about fish. Front. Vet. Sci. 2021, 8, 616955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Q.; Cai, J.; Yang, J.; Shen, Q.; and Xu, S. Chlorpyrifos Exposure in Common Carp (Cyprinus carpio L.) Leads to Oxidative Stress and Immune Responses. Fish Shellfish Immunol. 2017, 67, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.U. Fish Health: An Oxidative Stress Perspective. Fish. Aquacult. J. 2014, 5, e105. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Costantini, D.; Cooke, S.J.; Willmore, W.G. A Comparative and Evolutionary Approach to Oxidative Stress in Fish: A Review. Fish Fish. 2017, 18, 928–942. [Google Scholar] [CrossRef]
Figure 1.
Black and white line drawings of the body and mouth apparatus of the seven Moxostoma species found in Canada including, (A) Silver redhorse, M. anisurum, (B) Black redhorse, M. duquesnei, (C) Golden redhorse, M. erythurum, (D) Shorthead redhorse, M. macrolepidotum, (E) Greater redhorse, M. valenciennesi, (F) River redhorse, M. carinatum, and (G) Copper redhorse, M. hubbsi. © Canadian Museum of Nature.
Figure 1.
Black and white line drawings of the body and mouth apparatus of the seven Moxostoma species found in Canada including, (A) Silver redhorse, M. anisurum, (B) Black redhorse, M. duquesnei, (C) Golden redhorse, M. erythurum, (D) Shorthead redhorse, M. macrolepidotum, (E) Greater redhorse, M. valenciennesi, (F) River redhorse, M. carinatum, and (G) Copper redhorse, M. hubbsi. © Canadian Museum of Nature.

Figure 2.
Black North American distribution range of extinct and resident populations of Moxostoma spp. found in Canada, namely Silver redhorse (A), Black redhorse (B), Golden redhorse (C), Shorthead redhorse (D), Greater redhorse (E) and River redhorse (F). Adapted from the website of IUCN/SSC Red List of Threatened Species (Accessed 17 03 2025).
Figure 2.
Black North American distribution range of extinct and resident populations of Moxostoma spp. found in Canada, namely Silver redhorse (A), Black redhorse (B), Golden redhorse (C), Shorthead redhorse (D), Greater redhorse (E) and River redhorse (F). Adapted from the website of IUCN/SSC Red List of Threatened Species (Accessed 17 03 2025).
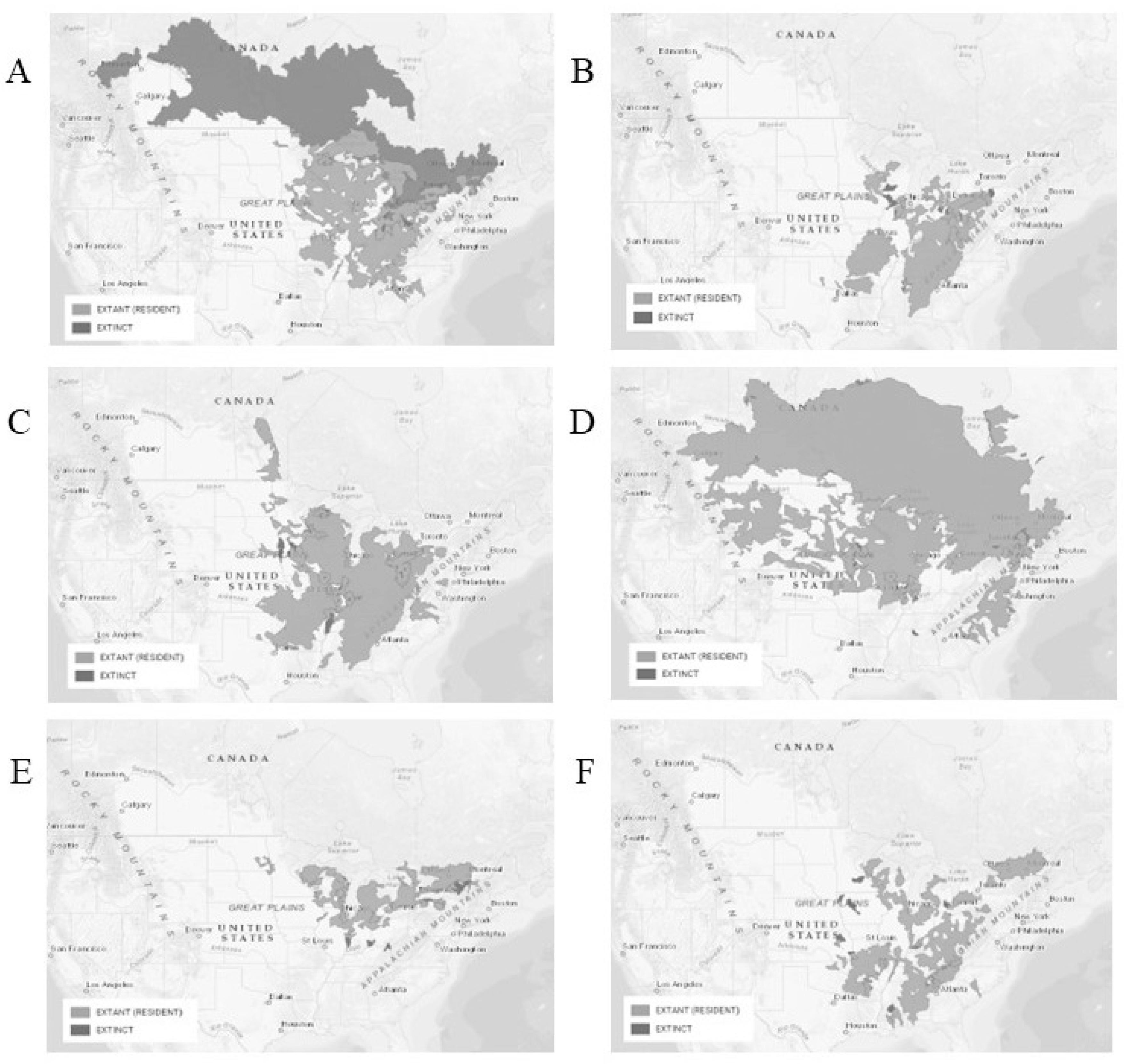
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



