
Submitted:
24 March 2025
Posted:
26 March 2025
You are already at the latest version
Abstract
Psychosis is one of the leading causes of disability worldwide. Individuals with an early onset, known as early-onset psychosis (EOP), tend to experience a worse prognosis and shorter life expectancy. The etiology of EOP remains unclear, but hypotheses suggest that both environmental and biological factors interact in its development. Epigenetic mechanisms are known to serve as the interface between environmental exposures and biological processes, emerging as potential tools to better understand the etiology of EOP. We characterized the sociodemographic and clinical characteristics, as well as genome-wide epigenetic markers, in Mexican patients with EOP. We found that patients with EOP have a higher epigenetic age using Wu`s clock (p=0.015). In addition, we observed that a reduction in years of schooling is associated with an increase in epigenetic age (Levine clock, =5.07, p=0.001). In our epigenome-wide association study, we identified eight CpGs associated with EOP. Noteworthy, a psychosis-methylation risk score (EOP-MRS) was associated with panic disorder (=1.36, p=0.03), as well as auditory (=1.28, p=0.04) and visual (=1.22, p=0.04) hallucinations. In conclusion, years of education have an impact in epigenetic age. Additionally, our study suggests associations of DNA methylation with EOP. Finally, we developed an MRS that associates clinical manifestations of psychosis.
Keywords:
early-onset psychosis
; epigenome-wide association study
; methylation risk score
; Mexico
; adolescents
1. Introduction
Early-onset psychosis (EOP) is a mental disorder characterized by the onset of psychosis before the age of 18 years [1]. Compared with adult-onset psychosis those affected by EOP have shorter life expectancy, and poor treatment response [2,3]. EOP prevalence is higher than adult-onset psychosis, with an estimated of 8-17% in children and adolescents [4,5,6]. The etiology of EOP is still unknown. Authors have proposed that abnormal neurodevelopment and early neurodegeneration explain the emergence of psychosis in the young population [7,8]. Furthermore, the literature suggests that environmental factors such as bullying, cannabis use, tobacco use, low birth weight, and childhood trauma interact with biological factors in the development of psychosis [3,9,10,11,12]. On the other hand, DNA methylation (DNAm), proposed to be a mediator between environmental exposures and biological effects, is the most studied epigenetic mechanism and could provide a better understanding of biological mechanisms underlying psychotic disorders [13]. New evidence from epigenome-wide association studies (EWAS), a comparison of DNAm sites across the genome, revealed associations with schizophrenia and first episode of psychosis (FEP) [14,15]. Additionally, DNAm is associated with psychotic symptoms in adults and the risk of neuropsychiatric disorders during childhood [8,16]. However, EWAS of clinical defined EOP has not been performed.
Development of novel biomarkers derived from DNAm provides a new approach to understand disease risk, biological and etiological mechanisms, such as aging [17,18,19]. For example, epigenetic clocks are excellent biomarkers to estimate biological age, also referred to as epigenetic age [7]. Limited evidence reveals that epigenetic age correlates with the severity of psychosis, and accelerated biological age is associated with psychotic disorders [20,21,22,23]. Furthermore, the development of methylation risk scores (MRS), representing the sum of an individual epigenetic risks derived from EWAS results, a similar construct of polygenic risk score, have been associated with schizophrenia [24], FEP [17], and neuroimaging changes in individuals with psychosis [8,25]. Studies using MRS showed that could be used to differentiate individuals affected of psychosis and mediation effects of childhood adversity to develop psychosis risk [15]. It is noteworthy that like in many biomedical research areas there is still a lack of diversity in EWAS studies [26].
The current understanding of EOP is still limited and there is scare evidence of epigenetic biomarkers, to advance in these field, the current study aimed to characterize for the first time sociodemographic and clinical characteristics, with a further comprehensive evaluation of genome-wide epigenetic markers (including 11 epigenetic clocks and EOP-MRS) in Mexican children and adolescents with EOP (Figure 1).
2. Results
2.1. Sample Description
We deeply characterized clinical and sociodemographic features of a total of 23 children and adolescents, including 12 psychiatric patients with psychotic symptoms (EOP group) and 11 psychiatric patients without psychotic symptoms (non-EOP group). We observed that the EOP group was older (mean = 15.00, p = 0.030) and had higher years of education (mean = 9.3, p = 0.023) (Table 1), nevertheless was close to the Mexican population mean (9.73 years) [27]. Furthermore, patients with EOP had higher hospital psychiatric admissions (p = 0.027) and higher prevalence of anxiety and stress disorders (p = 0.036). The EOP group had a lower functional score (p = 0.00003), and severe GAF score (global assessment of functioning) (p = 0.017) (Table 1). Our results shows that EOP had a higher comorbidity and lower functionality.
2.2. Epigenetic Age
There is a hypothesis that patients with psychosis have higher biological age [28]. To explore this, we further characterized 11 epigenetic clocks to evaluate whether if patients with EOP have an increased biological age. Our study found that the epigenetic age was higher in the EOP group compared to the non-EOP group (Table 1). We identified a correlation of epigenetic age with socio-demographic and clinical characteristics with the Wu clock, with a lower functionally measured by the GAF scale associated with a higher epigenetic age (Figure 2). Moreover, a higher number of admissions was correlated with an increased epigenetic age. Additionally, the same correlations were observed with the PedBE clock, showing a similar direction of effects. In contrast, the DNAmTL clock showed a negative correlation with the number of admissions (Table 2).
In addition, we observed that children and adolescent patients with psychosis were associated with accelerated epigenetic age (Levine clock, β = 5.07, p = 0.001), Furthermore, clinical characteristics appeared to influence this accelerated epigenetic age. Our findings show that a reduction in schooling (Levine clock, β = -5.01, p = 0.001), a higher number of comorbidities (BLUP clock, β = 0.49, p = 0.041) and more admissions (Wu clock, β = 0.81, p = 0.042) were associated with higher epigenetic age in Mexican patients with EOP (Table 3).
2.3. Epigenome-Wide Association Study
Differential methylation analysis was performed to associate epigenetic risk markers between EOP and non-EOP groups in Mexican patients. We identified 8 differentially methylated CpG sites associated with EOP at nominal significance, CpG sites are distributed across 6 chromosomes (Figure 3). These sites were mapped to 7 genes: ADGRV1 (Adhesion G Protein-coupled Receptor V1), HIST1H2BB (Histone Cluster 1 H2B Family Member B), CEP164 (Centrosomal Protein 164), IRF2BP1 (Interferon Regulatory Factor 2 Binding Protein 1), MAP1B (Microtubule Associated Protein 1B), NAALAD2 (N-Acetylated Alpha-Linked Acidic Dipeptidase 2), and SULT1C4 (Sulfotransferase Family 1C Member 4). The annotation indicated that 4 sites were situated in gene bodies, 2 sites in exons, and 1 site in a transcription start site (TSS200). In terms of their location relative to CpG islands, 25% of the sites were within the island, 25% in the island shores, and 50% in the Open Sea. Besides, 6 CpG sites had lower methylation values, while 2 CpG sites showed higher values in the EOP group compared to the non-EOP group (Table 4). In deep research of CpG methylation sites, there are reports of cg06583549, cg08523325, cg20150189 and cg26028573 in the EWAS catalog (Supplementary sheet).
2.4. Methylation Risk Score
Epigenetic biomarkers could serve as predicting tools for psychosis [29] and currently there is not epigenetic biomarker for EOP. We constructed the EOP-MRS to associate the biomarker with sociodemographic and clinical characteristic in children and adolescent Mexican patients with EOP. Our findings show that panic disorder, auditory hallucinations, and visual hallucinations were associated with a higher MRS using 5 different p-value thresholds: MRS1x10-01 (β=2.10, β=2.26, β=1.93, respectively); MRS1X10-02 (β=2.08, β=2.21, β=1.90, respectively); MRS1x10-03 (β=1.92, β=1.96, β=1.78, respectively); MRS1x10-04 (β=1.91, β=1.73, β=1.54, respectively); and MRS1x10-05 (β=1.36, β=1.28, β=1.22, respectively). Furthermore, a higher MRS was associated with tactile hallucinations (MRS1x10-05, β=1.59), and negative symptoms (MRS1x10-01, β=1.99; MRS1X10-02, β=1.71) (Figure 4). Our results shows that our EOP-MRS may be an indicator for visual and auditive hallucinations in Mexican children and adolescents’ patients.
3. Discussion
This study performed the first characterization of genome-wide epigenetic biomarkers in Mexican children and adolescents with EOP, further explored the correlations and associations between epigenetic age (11 epigenetic clocks), MRS, sociodemographic and clinical characteristics in Mexican children and adolescents with EOP.
The current study demonstrated that lower levels of functioning, admissions, and anxiety-stress disorder were associated with EOP. Our findings are consistent with a study of European patients with FEP (first episode of psychosis), who had lower GAF scores compared to other psychiatric groups [30]. These results support the hypothesis that functional impairment may be influenced by psychiatric comorbidities and admissions, leading to a poor prognosis [31] or premature death [32].
3.1. Years of Schooling Was Associated with Epigenetic Age in EOP
The present study shows that epigenetic age is higher in psychiatric patients with EOP. Consistent with our results, previous work identified that monozygotic twins with psychiatric disorders had an accelerated epigenetic age during early adolescence measured with the Wu clock [19], and in Mexican adults the presence of a mental disorder accelerated Horvath’s epigenetic age in discordant monozygotic twins [33]. In this sense, we suggest that Wu's clock could be used to estimate epigenetic age in the adolescent Mexican population. Furthermore, the epigenetic age was associated with fewer years of schooling in EOP. In accordance with our results, previous work in Mexican population showed that schooling influences epigenetic age [34]. We consider that schooling could influence biological age in Mexican with EOP. Notably, the lack of concordance between different epigenetic clocks could reflect how each clock is calibrated to specific sets of CpGs or tissues. Nevertheless, still further research is needed to explore the effect of other stressors and social determinants of health in the relationship between EOP and epigenetic age [35].
3.2. EWAS Suggested Potential Novel Associations with EOP
Our EWAS analysis identified 8 CpG sites associated with EOP at a nominal level. Nonetheless, a previous meta-analysis reported 95 differentially methylated positions associated with psychosis in adult patients [14]. The MAP1B gene regulates axon growth and synaptic plasticity [36], and its dysregulation leads to disruptions in neurogenesis and synaptic plasticity [37]. The HIST1H2BB gene is involved in cell motility [38] and neurodevelopment [39]. In addition, methylation changes in the CEP164 gene are associated with brainstem malformation [40], and the IRF2BP1 gene is involved in immune suppression [41]. There is evidence that SULT1C4, NAALAD2, and ADGRV1 participate in neurotransmitter regulation [42] glutamate dysregulation in schizophrenia [43] and epilepsy and audiovisual disorders [44,45]. All these findings indicate DNA methylation is implicated in neurodevelopmental disorders, immune-modulatory pathways, neurotransmitter regulation in EOP patients. This suggestion is based on the following findings: (i) a reduced level of methylation is observed at cg24772138, cg05100917, cg06583549, cg20150189, cg08523325, and cg27181762; (ii) a higher level of methylation is observed at cg26028573 and cg13883911.
We identified four CpG sites previously reported in EWAS [46,47,48,49]. Islam et al. (2019) suggest that cg06583549, cg08523325, cg20150189, and cg26028573 present DNAm concordance between buccal epithelial cells and peripheral blood cells in pediatric patients [46]. Furthermore, these sites are associated with children's neurodevelopment [49] and HIV infection in adults [48]. In addition, cg26028573 was associated with alcohol consumption [47]. Notably, this association has been reported in psychotic patients and individuals with developmental disorders [50]. Our findings support evidence that DNAm signature overlap across different pediatric tissues in psychotic patients [51]. Moreover, these sites could reflect shifts across the lifespan [52] and neurodevelopmental disorders [53], suggesting that our results could provide insight into children's neurodevelopment within our population.
3.3. Association Between Clinical Charateristics Associated with Psychosis MRS
Our study showed clinical characteristics associated with EOP-MRS. Previous associations of MRS with clinical characteristics have been identified in other populations. For example, in an adult Australian population diagnosed with schizophrenia, the MRS was associated with clozapine administration [54]. Our EOP-MRS could represent a predictive tool to identify panic disorder and hallucinations in Mexican children and adolescent population. We suggest that EOP-MRS reflects the cumulative effect of multiple epigenetic markers associated with EOP and recapitulates its clinical manifestations.
3.4. Limitations
We considered several limitations in our study: the analysis includes a small sample of psychiatric patients with EOP, which limits our statistical power to detect associations. Additionally, epigenetic studies are not ideal for identifying causal risk factors, as DNA methylation (DNAm) may be influenced by environmental confounders. Moreover, EPIC arrays cover only 3% of CpG sites in the genome, and polymorphisms and mutations were not accounted for in psychosis risk calculations. Furthermore, DNAm profiles focus on peripheral blood, and variations associated with early-onset psychosis may not represent specific brain-tissue biomarkers in the early stages of the disease. Additionally, epigenetic age, EWAS, and psychosis MRS associated with EOP could be biased due to ancestry, pubertal stage, childhood adversity, or environmental exposures not included in the analysis. We did not have a replication cohorts given the difficult of recruitment of EOP. In addition, we should consider pruning, as variability in epigenetic signals may differ across individuals, populations, and ages, particularly during early stages of life. Finally, future research should address these issues including larger samples, DNAm profiles from other tissues, and social determinants of health to replicate our results.
4. Materials and Methods
4.1. Sample Population
Thirty-three patients with previously diagnosed psychiatric disorders were recruited from the Hospital Psiquiátrico Infantil Juan N. Navarro in Mexico City. A psychiatrist assessed all participants according to the DSM-5 (Diagnostic and Statistical Manual of Mental Disorders, 5th Edition). We included patients aged 10 to 18 years of Mexican descent, with the onset of psychotic symptoms before the age of 18. Exclusion criteria included patients with psychosis secondary to a brain infection, neurodegenerative disease, or severe neurodevelopmental disorder.
4.2. Study Design
This was a cross-sectional study with a convenience sample design. Recruitment took place from January 2022 to July 2023.The sample was divided into two groups: the EOP group (patients with psychotic symptoms) and the non-EOP group (patients without psychotic symptoms).
The clinical evaluation of patients was performed using the Kiddie Schedule for Affective Disorders and Schizophrenia Present and Lifetime version DSM-5 (K-SADS PL-5), Spanish edition, validated in the children and adolescent Mexican population [55]. Each individual diagnosis was categorized into one of five groups as follows: 1) mood disorders (disruptive mood dysregulation disorder, dysthymia, major depressive disorder, bipolar disorder); 2) anxiety and stress disorders (separation anxiety disorder, agoraphobia, panic disorder, social phobia, generalized anxiety disorder, post-traumatic stress disorder); 3) conduct disorders (enuresis, encopresis, oppositional defiant disorder, conduct disorder, tic disorder, obsessive-compulsive disorder); 4) neurodevelopmental disorders (attention-deficit/hyperactivity disorder, autism spectrum disorder); 5) eating disorders (bulimia nervosa, other eating disorders).
Clinicians collected sociodemographic information of age, gender, schooling (completed years of education), and clinical data, including weight, height, body mass index (BMI), number of psychiatric hospitalizations (admissions), number of medications (psychiatric treatment), and number of psychiatric comorbidities. BMI values were converted into z-scores with standard deviations. The Global Assessment of Functioning (GAF) was measured on a 100-point scale, where lower scores indicated severe symptoms or impairment [56]. GAF is validated in Spanish and adolescent populations [57]. At the end of the interview, we collected a peripheral venous blood sample (4 ml) from each patient.
4.3. DNA Extraction
Genomic DNA was obtained from whole blood samples using the DNA kit (Qiagen, USA), according to the manufacturer’s instructions. Quality and integrity were evaluated using a NanoDrop spectrophotometer (Thermo Fisher, USA). Sample processing for the methylation study protocol was performed with a genomic DNA concentration of 80 ng/mL. The genomic DNA was bisulfite-converted using the EZ DNA Methylation Kit (Zymo Research, Irvine, CA, USA), according to the manufacturer’s protocol, “Infinium Assay for Methylation Protocol” (Illumina Inc), specific for the “Infinium Methylation EPIC BeadChip Kit.” The Microarray Unit at the National Institute of Genomic Medicine (Mexico City, Mexico) processed the microarray.
4.4. Genomic-Wide Quantification of DNA Methylation
Fluorescence intensities were transformed into IDAT files using GenomeStudio software (Illumina, USA). The IDAT data were imported into the R environment. Beta-normalized methylation levels were obtained following the ENmix (1.22.0) pipeline. Preprocessing included background correction, RELIC dye bias correction, and RCP probe-type bias adjustment. We removed CpG sites with low variability (12,730 CpG sites) and sites that coincided with SNP loci from the analysis. Additionally, 10 samples were eliminated due to sex mismatch, poor quality, and being outliers. Five surrogate variables were estimated to correct for batch effects using `csva()`; and cell proportions (neutrophils, monocytes, natural killer cells, CD8+ T-lymphocytes, CD4+ T-lymphocytes, and B-lymphocytes) were imputed using `estimateCellProp()` (ENmix R library). Finally, 848,643 CpG sites from 23 samples were analyzed to obtain the methylation beta (β) and M values.
4.5. Epigenetic Clocks
Epigenetic age was estimated using β values with two libraries. The methylClock and methylCIPHER packages were chosen for their broad availability of epigenetic calculators The Wu and Pediatric Buccal Epigenetic (PedBE) clocks were included because they are designed to predict age in children and adolescents. The Dunedin Pace of Aging (DunedinPoAm38) and Levine (PhenoAge) clocks were selected for their ability to calculate biological age and predict mortality. DNA methylation-based telomere length (DNAmTL), Hannum, Horvath-1 (multi-tissue), and Zhang (2019) clocks were included because they predict chronological age in blood samples. The Best Linear Unbiased Prediction (BLUP), Elastic Net (EN), and Horvath-2 (skin and blood) clocks were chosen because they were trained on blood, skin, and saliva samples using different arrays (450k and EPIC arrays) [58,59,60,61,62,63,64,65,66].
4.6. Statistical Analysis
Clinical characteristics are presented as means and standard deviations for continuous variables, and as frequencies and percentages for categorical variables. Comparisons between groups were made using the student’s t-test or the Mann-Whitney U test. The associations were assessed with the chi-square test (χ²). In psychiatric patients the correlations between epigenetic age and clinical and sociodemographic characteristics were calculated using corrplot R package. Multiple linear regressions (stepwise regression) were performed to associate sociodemographic and clinical characteristics with epigenetic age (the response variable). Statistical analysis was conducted using R software, version 4.3.3 (https://CRAN.R-project.org, accessed 30 December 2024). The significance level was set at p < 0.05.
4.7. EWAS and Methylation Risk Score
Genome-wide CpG association analysis with EOP was performed using normalized β values. EOP was the response variable, and each individual CpG methylation β value served as the predictor of interest. A logistic model was implemented using the `CpGassoc()` R package [67], with PC1 (principal component 1), PC2, PC3, PC4, PC5, age, and sex included as covariates. A QQ-plot was examined for evidence of genomic inflation (λ = 0.9783, Figure S1). We manually search for previous associations of the CpG sites using the EWAS catalog [68].
CpG sites were stratified by cut-off p-values from the EWAS results to calculate the EOP-MRS (Methylation Risk Score to psychosis): MRS1x10-06 (3 CpG sites, p < 1x10-06), MRS1x10-05 (8 CpG sites, p < 1x10-05), MRS1x10-04 (101 CpG sites, p < 1x10-04), MRS1x10-03 (997 CpG sites, p < 1x10-03), MRS1x10-02 (9,832 CpG sites, p < 1x10-02), MRS1x10-01 (88,916 CpG sites, p < 1x10-01).
Each MRS was calculated as the sum of the individual products of the M values and the Z values (effect size/standard error) for each CpG site (CpG1 + CpG2 + ... + CpGn). The resulting score was then normalized into z-scores.
Finally, linear and logistic regressions were performed to assess associations between EOP-MRS and sociodemographic and clinical characteristics. Sociodemographic and clinical variables served as the response variable, and the EOP-MRS was considered predictor of interest.
5. Conclusions
We found that patients with EOP have lower level of functionality, and a higher epigenetic age. Furthermore, fewer years of education, comorbidity and psychiatric hospital admissions have an impact in epigenetic age. Additionally, our study suggests associations of DNAm with EOP, specifically in genes of immune-modulatory and neurotransmission pathways. Finally, we developed an EOP-MRS that associates clinical manifestations of psychosis and could be used as a risk biomarker for EOP.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Q-Q plot of EWAS of early onset psychosis; Table S1: Clinical characteristics of early-onset psychosis; Sheet S1: Sociodemographic and clinical associations with EOP-MRS, and EWAS catalog; Figure S2: Most frequents psychiatric comorbidities; Table S2: Summary of the stepwise regression between Best Linear Unbiased Prediction`s clock (BLUP), socio-demographic and clinical characteristics: Figure S3 Beta value distribution A) before and B) after quality control; Table S2: Summary of the stepwise regression between DNA methylation-based telomere length (DNAmTL), socio-demographic and clinical characteristics; Figure S4: Distribution of T statistic values of EWAS; Table S4: Summary of the stepwise regression between Dunedin Pace of Aging Methylation (DunedinPoAm38), socio-demographic and clinical characteristics; Table S5: Summary of the stepwise regression between Elastic Net`s clock (EN), socio-demographic and clinical characteristics; Table S6: Summary of the stepwise regression between Hannum´s clock, socio-demographic and clinical characteristics; Table S7: Summary of the stepwise regression between Horvath´s clock (Multi-Tissue), socio-demographic and clinical characteristics; Table S8: Summary of the stepwise regression between Horvath´s clock (Skin & Blood), socio-demographic and clinical characteristics; Table S9: Summary of the stepwise regression between Levine´s clock “PhenoAge”, socio-demographic and clinical characteristics; Table S10: Summary of the stepwise regression between Pediatric-Buccal-Epigenetic´s clock (PedBE), socio-demographic and clinical characteristics; Table S11: Summary of the stepwise regression between Wu´s clock, socio-demographic and clinical characteristics; Table S12: Summary of the stepwise regression between Zhang`s clock, socio-demographic and clinical characteristics; Table S13: Correlations between Best Linear Unbiased Prediction`s clock (BLUP), socio-demographic and clinical characteristics; Table S14: Correlations between DNA methylation-based telomere length (DNAmTL), socio-demographic and clinical characteristics; Table S15. Correlations between Dunedin Pace of Aging Methylation (DunedinPoAm38), socio-demographic and clinical characteristics; Table S16. Correlations between Elastic Net`s clock (EN), socio-demographic and clinical characteristics; Table S17. Correlations between Hannum´s clock, socio-demographic and clinical characteristics; Table S18: Correlations between Horvath´s clock (Multi-Tissue), socio-demographic and clinical characteristics; Table S19: Correlations between Horvath´s clock (Skin & Blood), socio-demographic and clinical characteristics; Table S20: Correlations between Levine´s clock “PhenoAge”, socio-demographic and clinical characteristics; Table S21: Correlations between Pediatric-Buccal-Epigenetic´s clock (PedBE), socio-demographic and clinical characteristics; Table S22: Correlations between Wu´s clock, socio-demographic and clinical characteristics; Table S23: Correlations between Zhang`s clock, socio-demographic and clinical characteristics.
Author Contributions
Conceptualization, H.N. and D.C.G.; methodology, D.R.-R., J.J.M.-M., and G.A.N.-R.; software, D.R.-R., J.J.M.-M., and G.A.N.-R.; validation, I.E.J.-R., H.N, J.L.M.-O. and A.D.G.-M.; formal analysis, D.R.-R., J.J.M.-M., G.A.N.-R. and J.D.C.-C.; investigation, F.S.H., J.C., A.D., P.Z., J.A.Z.-D. and N.L.; resources, F.S.H., J.C., A.D., P.Z., J.A.Z.-D., N.L. and H.N.; data curation, D.R.-R., J.J.M.-M., and I.E.J.-R.; writing—original draft preparation, D.R.-R., J.J.M.-M., I.E.J.-R., G.A.N.-R., and J.D.C.-C.; writing—review and editing, D.R.-R., J.J.M.-M., I.E.J.-R., J.L.M.-O. and A.D.G.-M.; visualization, D.R.-R., J.J.M.-M., I.E.J.-R., J.D.C.-C., H.N. and A.D.G.-M.; supervision, I.E.J.-R., H.N., J.L.M.-O., D.C.G., and A.D.G.-M.; project administration, H.N., D.C.G. and A.D.G.-M.; funding acquisition, H.N., D.C.G. and A.D.G.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Instituto Nacional de Medicina Genómica (INMEGEN).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committees of the “Juan N. Navarro” Psychiatric Hospital and the National Institute of Genomic Medicine (INMEGEN) (HPIJNN-CI-DA-021-2021).
Informed Consent Statement
Parents/tutors received both written and oral information about the study’s ais and provided written informed consent and assent.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
This work was supported by National Institute of Mental Health (NIMH) 1R01MH133621 (DG, HN). This work was supported by the Kavli Insttute for Neuroscience at Yale Unversity Kavli Postdoctoral Award for Academic Diversity to Jose Jaime Martinez-Magaña. This work is supported by the U.S. Department of Veterans Affairs via 1IK2CX002095-01A1 (JLMO), the National Center for PTSD, and NIDA R21DA050160 (JLMO, JJMM). The authors also thank SECIHTI for supporting David Ruiz-Ramos (CVU 888651), and Juan Daniel Cruz-Castillo (CVU 1043199) with the National Scholarship for Postgraduate Studies; they are students of the Doctorate in Biomedical Sciences degree at the Universidad Juárez Autónoma de Tabasco. We want to acknowledge the Unidad de Alta Tecnología para Expresión y Microarreglos of the Instituto Nacional de Medicina Genómica for their technical support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ADGRV1 | Adhesion G Protein-coupled Receptor V1 gene |
| BLUP | Best Linear Unbiased Prediction |
| CEP164 | Centrosomal Protein 164 gene |
| CpG | Cytosine-Guanine site |
| DNA | Deoxyribonucleic Acid |
| DNAm | Deoxyribonucleic Acid methylation |
| DNAmTL | DNA methylation-based telomere length |
| DSM-5 | Diagnostic and Statistical Manual of Mental Disorders, 5th Edition |
| DunedinPoAm38 | Dunedin Pace of Aging |
| EN | Elastic net |
| EOP | Early-onset psychosis |
| EOP-MRS | Psychosis-Methylation Risk Score |
| EWAS | Epigenome-wide association analysis |
| FEP | First episode of pychosis |
| GAF | Global Assessment of Functioning |
| HIST1H2BB | Histone Cluster 1 H2B Family Member B gene |
| IRF2BP1 | Interferon Regulatory Factor 2 Binding Protein 1 gene |
| K-SADS PL-5 | Kiddie Schedule for Affective Disorders and Schizophrenia Present and Lifetime version DSM-5 |
| MAP1B | Microtubule Associated Protein 1B gene |
| MRS | Methylation Risk Score |
| NAALAD2 | N-Acetylated Alpha-Linked Acidic Dipeptidase 2 gene |
| PC | Principal component |
| PedBE | Pediatric Buccal Epigenetic |
| SULT1C4 | Sulfotransferase Family 1C Member 4 gene |
| TSS | Transcription start site |
References
- Salazar de Pablo, G.; Rodriguez, V.; Besana, F.; Civardi, S.C.; Arienti, V.; Maraña Garceo, L.; Andrés-Camazón, P.; Catalan, A.; Rogdaki, M.; Abbott, C.; et al. Umbrella Review: Atlas of the Meta-Analytical Evidence of Early-Onset Psychosis. J Am Acad Child Adolesc Psychiatry 2024, 63, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Fusar-Poli, P.; Salazar de Pablo, G.; Correll, C.U.; Meyer-Lindenberg, A.; Millan, M.J.; Borgwardt, S.; Galderisi, S.; Bechdolf, A.; Pfennig, A.; Kessing, L.V.; et al. Prevention of Psychosis: Advances in Detection, Prognosis, and Intervention. JAMA Psychiatry 2020, 77, 755–765. [Google Scholar] [CrossRef]
- Oliver, D.; Chesney, E.; Cullen, A.E.; Davies, C.; Englund, A.; Gifford, G.; Kerins, S.; Lalousis, P.A.; Logeswaran, Y.; Merritt, K.; et al. Exploring causal mechanisms of psychosis risk. Neurosci Biobehav Rev 2024, 162, 105699. [Google Scholar] [CrossRef]
- Kelleher, I.; Connor, D.; Clarke, M.C.; Devlin, N.; Harley, M.; Cannon, M. Prevalence of psychotic symptoms in childhood and adolescence: a systematic review and meta-analysis of population-based studies. Psychol Med 2012, 42, 1857–1863. [Google Scholar] [CrossRef]
- Driver, D.I.; Thomas, S.; Gogtay, N.; Rapoport, J.L. Childhood-Onset Schizophrenia and Early-onset Schizophrenia Spectrum Disorders: An Update. Child Adolesc Psychiatr Clin N Am 2020, 29, 71–90. [Google Scholar] [CrossRef]
- Zammit, S.; Kounali, D.; Cannon, M.; David, A.S.; Gunnell, D.; Heron, J.; Jones, P.B.; Lewis, S.; Sullivan, S.; Wolke, D.; et al. Psychotic experiences and psychotic disorders at age 18 in relation to psychotic experiences at age 12 in a longitudinal population-based cohort study. Am J Psychiatry 2013, 170, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Iftimovici, A.; Kebir, O.; Jiao, C.; He, Q.; Krebs, M.O.; Chaumette, B. Dysmaturational Longitudinal Epigenetic Aging During Transition to Psychosis. Schizophr Bull Open 2022, 3, sgac030. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Walton, E.; Neumann, A.; Thio, C.H.L.; Felix, J.F.; van, I.M.H.; Pappa, I.; Cecil, C.A.M. DNA methylation at birth and lateral ventricular volume in childhood: a neuroimaging epigenetics study. J Child Psychol Psychiatry 2024, 65, 77–90. [Google Scholar] [CrossRef]
- Patel, P.K.; Leathem, L.D.; Currin, D.L.; Karlsgodt, K.H. Adolescent Neurodevelopment and Vulnerability to Psychosis. Biol Psychiatry 2021, 89, 184–193. [Google Scholar] [CrossRef]
- Rasic, D.; Hajek, T.; Alda, M.; Uher, R. Risk of mental illness in offspring of parents with schizophrenia, bipolar disorder, and major depressive disorder: a meta-analysis of family high-risk studies. Schizophr Bull 2014, 40, 28–38. [Google Scholar] [CrossRef]
- Radua, J.; Ramella-Cravaro, V.; Ioannidis, J.P.A.; Reichenberg, A.; Phiphopthatsanee, N.; Amir, T.; Yenn Thoo, H.; Oliver, D.; Davies, C.; Morgan, C.; et al. What causes psychosis? An umbrella review of risk and protective factors. World Psychiatry 2018, 17, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.J.; Freeman, D.; Lundström, S.; Larsson, H.; Ronald, A. Heritability of Psychotic Experiences in Adolescents and Interaction With Environmental Risk. JAMA Psychiatry 2022, 79, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Kouter, K.; Šalamon Arčan, I.; Videtič Paska, A. Epigenetics in psychiatry: Beyond DNA methylation. World J Psychiatry 2023, 13, 319–330. [Google Scholar] [CrossRef]
- Hannon, E.; Dempster, E.L.; Mansell, G.; Burrage, J.; Bass, N.; Bohlken, M.M.; Corvin, A.; Curtis, C.J.; Dempster, D.; Di Forti, M.; et al. DNA methylation meta-analysis reveals cellular alterations in psychosis and markers of treatment-resistant schizophrenia. Elife 2021, 10. [Google Scholar] [CrossRef]
- Alameda, L.; Liu, Z.; Sham, P.C.; Aas, M.; Trotta, G.; Rodriguez, V.; Di Forti, M.; Stilo, S.A.; Kandaswamy, R.; Arango, C.; et al. Exploring the mediation of DNA methylation across the epigenome between childhood adversity and First Episode of Psychosis-findings from the EU-GEI study. Mol Psychiatry 2023, 28, 2095–2106. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Luo, T.; Yang, C.; Liu, M.; Shen, Y.; Hao, W. Psychotic symptoms associated increased CpG methylation of metabotropic glutamate receptor 8 gene in Chinese Han males with schizophrenia and methamphetamine induced psychotic disorder: a longitudinal study. Schizophrenia (Heidelb) 2024, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Segura, A.G.; Prohens, L.; Julià, L.; Amoretti, S.; M, R.I.; Pino-Camacho, L.; Cano-Escalera, G.; Mane, A.; Rodriguez-Jimenez, R.; Roldan, A.; et al. Methylation profile scores of environmental exposures and risk of relapse after a first episode of schizophrenia. Eur Neuropsychopharmacol 2025, 94, 4–15. [Google Scholar] [CrossRef]
- Kular, L.; Kular, S. Epigenetics applied to psychiatry: Clinical opportunities and future challenges. Psychiatry Clin Neurosci 2018, 72, 195–211. [Google Scholar] [CrossRef]
- Ingram, S.J.; Vazquez, A.Y.; Klump, K.L.; Hyde, L.W.; Burt, S.A.; Clark, S.L. Associations of depression and anxiety symptoms in childhood and adolescence with epigenetic aging. J Affect Disord 2024, 352, 250–258. [Google Scholar] [CrossRef]
- Smigielski, L.; Jagannath, V.; Rössler, W.; Walitza, S.; Grünblatt, E. Epigenetic mechanisms in schizophrenia and other psychotic disorders: a systematic review of empirical human findings. Mol Psychiatry 2020, 25, 1718–1748. [Google Scholar] [CrossRef]
- Yusupov, N.; Dieckmann, L.; Erhart, M.; Sauer, S.; Rex-Haffner, M.; Kopf-Beck, J.; Brückl, T.M.; Czamara, D.; Binder, E.B. Transdiagnostic evaluation of epigenetic age acceleration and burden of psychiatric disorders. Neuropsychopharmacology 2023, 48, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.P.Y.; Abram, S.V.; Loewy, R.L.; Stuart, B.; Fryer, S.L.; Vinogradov, S.; Mathalon, D.H. Brain Age Gap in Early Illness Schizophrenia and the Clinical High-Risk Syndrome: Associations With Experiential Negative Symptoms and Conversion to Psychosis. Schizophr Bull 2024, 50, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Dada, O.; Adanty, C.; Dai, N.; Jeremian, R.; Alli, S.; Gerretsen, P.; Graff, A.; Strauss, J.; De Luca, V. Biological aging in schizophrenia and psychosis severity: DNA methylation analysis. Psychiatry Res 2021, 296, 113646. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.K.; Lin, J.J.; Tseng, H.H.; Wang, X.Y.; Jang, F.L.; Chen, P.S.; Huang, C.C.; Hsieh, S.; Lin, S.H. DNA methylation signature aberration as potential biomarkers in treatment-resistant schizophrenia: Constructing a methylation risk score using a machine learning method. J Psychiatr Res 2023, 157, 57–65. [Google Scholar] [CrossRef]
- Ohi, K.; Shimada, M.; Soda, M.; Nishizawa, D.; Fujikane, D.; Takai, K.; Kuramitsu, A.; Muto, Y.; Sugiyama, S.; Hasegawa, J.; et al. Genome-wide DNA methylation risk scores for schizophrenia derived from blood and brain tissues further explain the genetic risk in patients stratified by polygenic risk scores for schizophrenia and bipolar disorder. BMJ Ment Health 2024, 27. [Google Scholar] [CrossRef]
- Breeze, C.E.; Wong, J.Y.Y.; Beck, S.; Berndt, S.I.; Franceschini, N. Diversity in EWAS: current state, challenges, and solutions. Genome Med 2022, 14, 71. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía. Caracteristicas educativas de la población. Censo de Población y Vivienda 2020. Available online: https://www.inegi.org.mx/app/tabulados/interactivos/?pxq=Educacion_Educacion_05_2f6d2a08-babc-442f-b4e0-25f7d324dfe0&idrt=15&opc=t (accessed on 07 March 2025).
- Chrusciel, J.H.; Orso, R.; de Mattos, B.P.; Fries, G.R.; Kristensen, C.H.; Grassi-Oliveira, R.; Viola, T.W. A systematic review and meta-analysis of epigenetic clocks in schizophrenia. Schizophr Res 2022, 246, 172–174. [Google Scholar] [CrossRef]
- Guo, L.K.; Su, Y.; Zhang, Y.Y.; Yu, H.; Lu, Z.; Li, W.Q.; Yang, Y.F.; Xiao, X.; Yan, H.; Lu, T.L.; et al. Prediction of treatment response to antipsychotic drugs for precision medicine approach to schizophrenia: randomized trials and multiomics analysis. Mil Med Res 2023, 10, 24. [Google Scholar] [CrossRef]
- Engen, M.J.; Vaskinn, A.; Melle, I.; Færden, A.; Lyngstad, S.H.; Flaaten, C.B.; Widing, L.H.; Wold, K.F.; Åsbø, G.; Haatveit, B.; et al. Cognitive and Global Functioning in Patients With First-Episode Psychosis Stratified by Level of Negative Symptoms. A 10-Year Follow-Up Study. Front Psychiatry 2022, 13, 841057. [Google Scholar] [CrossRef]
- Elhassan, N.M.; Elhusein, B.; Al Abdulla, M.; Saad, T.A.; Kumar, R. Sociodemographic and clinical characteristics of patients with recurrent psychiatric readmissions in Qatar. J Int Med Res 2020, 48, 300060520977382. [Google Scholar] [CrossRef]
- Fountoulakis, K.N.; Karakatsoulis, G.N.; Abraham, S.; Adorjan, K.; Ahmed, H.U.; Alarcón, R.D.; Arai, K.; Auwal, S.S.; Berk, M.; Bjedov, S.; et al. Somatic multicomorbidity and disability in patients with psychiatric disorders in comparison to the general population: a quasi-epidemiological investigation in 54,826 subjects from 40 countries (COMET-G study). CNS Spectr 2024, 29, 126–149. [Google Scholar] [CrossRef]
- Martínez-Magaña, J.J.; Genis-Mendoza, A.D.; Santana, D.; Tóvilla-Zárate, C.A.; Lanzagorta, N.; Nicolini, H. [The presence of a psychiatric diagnosis could alter the epigenetic clock in monozygotic twins]. Rev Esp Geriatr Gerontol 2021, 56, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Verjan, J.C.; Esparza-Aguilar, M.; Martín-Martín, V.; Salazar-Perez, C.; Cadena-Trejo, C.; Gutierrez-Robledo, L.M.; Martínez-Magaña, J.J.; Nicolini, H.; Arroyo, P. Years of Schooling Could Reduce Epigenetic Aging: A Study of a Mexican Cohort. Genes (Basel) 2021, 12. [Google Scholar] [CrossRef]
- Raffington, L.; Belsky, D.W. Integrating DNA Methylation Measures of Biological Aging into Social Determinants of Health Research. Curr Environ Health Rep 2022, 9, 196–210. [Google Scholar] [CrossRef]
- Villarroel-Campos, D.; Gonzalez-Billault, C. The MAP1B case: an old MAP that is new again. Dev Neurobiol 2014, 74, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.M.; Reay, W.R.; Geaghan, M.P.; Kiltschewskij, D.J.; Green, M.J.; Weidenhofer, J.; Glatt, S.J.; Cairns, M.J. miRNA cargo in circulating vesicles from neurons is altered in individuals with schizophrenia and associated with severe disease. Sci Adv 2023, 9, eadi4386. [Google Scholar] [CrossRef] [PubMed]
- Valle, B.L.; Rodriguez-Torres, S.; Kuhn, E.; Díaz-Montes, T.; Parrilla-Castellar, E.; Lawson, F.P.; Folawiyo, O.; Ili-Gangas, C.; Brebi-Mieville, P.; Eshleman, J.R.; et al. HIST1H2BB and MAGI2 Methylation and Somatic Mutations as Precision Medicine Biomarkers for Diagnosis and Prognosis of High-grade Serous Ovarian Cancer. Cancer Prev Res (Phila) 2020, 13, 783–794. [Google Scholar] [CrossRef]
- Świtońska, K.; Szlachcic, W.J.; Handschuh, L.; Wojciechowski, P.; Marczak, Ł.; Stelmaszczuk, M.; Figlerowicz, M.; Figiel, M. Identification of Altered Developmental Pathways in Human Juvenile HD iPSC With 71Q and 109Q Using Transcriptome Profiling. Front Cell Neurosci 2018, 12, 528. [Google Scholar] [CrossRef]
- Gana, S.; Serpieri, V.; Valente, E.M. Genotype-phenotype correlates in Joubert syndrome: A review. Am J Med Genet C Semin Med Genet 2022, 190, 72–88. [Google Scholar] [CrossRef]
- Hari, A.; Cruz, S.A.; Qin, Z.; Couture, P.; Vilmundarson, R.O.; Huang, H.; Stewart, A.F.R.; Chen, H.-H. IRF2BP2-deficient microglia block the anxiolytic effect of enhanced postnatal care. Scientific Reports 2017, 7, 9836. [Google Scholar] [CrossRef]
- Dubaisi, S.; Fang, H.; Caruso, J.A.; Gaedigk, R.; Vyhlidal, C.A.; Kocarek, T.A.; Runge-Morris, M. Developmental Expression of SULT1C4 Transcript Variants in Human Liver: Implications for Discordance Between SULT1C4 mRNA and Protein Levels. Drug Metab Dispos 2020, 48, 515–520. [Google Scholar] [CrossRef]
- Carter, C.J. Schizophrenia susceptibility genes converge on interlinked pathways related to glutamatergic transmission and long-term potentiation, oxidative stress and oligodendrocyte viability. Schizophr Res 2006, 86, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Güler, B.E.; Linnert, J.; Wolfrum, U. Monitoring paxillin in astrocytes reveals the significance of the adhesion G protein coupled receptor VLGR1/ADGRV1 for focal adhesion assembly. Basic Clin Pharmacol Toxicol 2023, 133, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Meng, H.; Liang, X.; Lei, X.; Zhang, J.; Bian, W.; He, N.; Lin, Z.; Song, X.; Zhu, W.; et al. ADGRV1 Variants in Febrile Seizures/Epilepsy With Antecedent Febrile Seizures and Their Associations With Audio-Visual Abnormalities. Front Mol Neurosci 2022, 15, 864074. [Google Scholar] [CrossRef]
- Islam, S.A.; Goodman, S.J.; MacIsaac, J.L.; Obradović, J.; Barr, R.G.; Boyce, W.T.; Kobor, M.S. Integration of DNA methylation patterns and genetic variation in human pediatric tissues help inform EWAS design and interpretation. Epigenetics Chromatin 2019, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Dugué, P.A.; Wilson, R.; Lehne, B.; Jayasekara, H.; Wang, X.; Jung, C.H.; Joo, J.E.; Makalic, E.; Schmidt, D.F.; Baglietto, L.; et al. Alcohol consumption is associated with widespread changes in blood DNA methylation: Analysis of cross-sectional and longitudinal data. Addict Biol 2021, 26, e12855. [Google Scholar] [CrossRef]
- Gross, A.M.; Jaeger, P.A.; Kreisberg, J.F.; Licon, K.; Jepsen, K.L.; Khosroheidari, M.; Morsey, B.M.; Swindells, S.; Shen, H.; Ng, C.T.; et al. Methylome-wide Analysis of Chronic HIV Infection Reveals Five-Year Increase in Biological Age and Epigenetic Targeting of HLA. Mol Cell 2016, 62, 157–168. [Google Scholar] [CrossRef]
- Mulder, R.H.; Neumann, A.; Cecil, C.A.M.; Walton, E.; Houtepen, L.C.; Simpkin, A.J.; Rijlaarsdam, J.; Heijmans, B.T.; Gaunt, T.R.; Felix, J.F.; et al. Epigenome-wide change and variation in DNA methylation in childhood: trajectories from birth to late adolescence. Hum Mol Genet 2021, 30, 119–134. [Google Scholar] [CrossRef]
- Qiu, L.; Liang, C.; Kochunov, P.; Hutchison, K.E.; Sui, J.; Jiang, R.; Zhi, D.; Vergara, V.M.; Yang, X.; Zhang, D.; et al. Associations of alcohol and tobacco use with psychotic, depressive and developmental disorders revealed via multimodal neuroimaging. Transl Psychiatry 2024, 14, 326. [Google Scholar] [CrossRef]
- Richetto, J.; Meyer, U. Epigenetic Modifications in Schizophrenia and Related Disorders: Molecular Scars of Environmental Exposures and Source of Phenotypic Variability. Biol Psychiatry 2021, 89, 215–226. [Google Scholar] [CrossRef]
- Short, A.K.; Weber, R.; Kamei, N.; Wilcox Thai, C.; Arora, H.; Mortazavi, A.; Stern, H.S.; Glynn, L.; Baram, T.Z. Individual longitudinal changes in DNA-methylome identify signatures of early-life adversity and correlate with later outcome. Neurobiol Stress 2024, 31, 100652. [Google Scholar] [CrossRef] [PubMed]
- Gui, A.; Jones, E.J.H.; Wong, C.C.Y.; Meaburn, E.; Xia, B.; Pasco, G.; Lloyd-Fox, S.; Charman, T.; Bolton, P.; Johnson, M.H. Leveraging epigenetics to examine differences in developmental trajectories of social attention: A proof-of-principle study of DNA methylation in infants with older siblings with autism. Infant Behav Dev 2020, 60, 101409. [Google Scholar] [CrossRef] [PubMed]
- Kiltschewskij, D.J.; Reay, W.R.; Geaghan, M.P.; Atkins, J.R.; Xavier, A.; Zhang, X.; Watkeys, O.J.; Carr, V.J.; Scott, R.J.; Green, M.J.; et al. Alteration of DNA Methylation and Epigenetic Scores Associated With Features of Schizophrenia and Common Variant Genetic Risk. Biol Psychiatry 2024, 95, 647–661. [Google Scholar] [CrossRef] [PubMed]
- de la Peña, F.R.; Villavicencio, L.R.; Palacio, J.D.; Félix, F.J.; Larraguibel, M.; Viola, L.; Ortiz, S.; Rosetti, M.; Abadi, A.; Montiel, C.; et al. Validity and reliability of the kiddie schedule for affective disorders and schizophrenia present and lifetime version DSM-5 (K-SADS-PL-5) Spanish version. BMC Psychiatry 2018, 18, 193. [Google Scholar] [CrossRef]
- Savić, B.; Jerotić, S.; Ristić, I.; Zebić, M.; Jovanović, N.; Russo, M.; Marić, N.P. Long-Term Benzodiazepine Prescription During Maintenance Therapy of Individuals With Psychosis Spectrum Disorders-Associations With Cognition and Global Functioning. Clin Neuropharmacol 2021, 44, 89–93. [Google Scholar] [CrossRef]
- Gezelius, C.M.E.; Wahlund, B.A.; Wiberg, B.M. Relation between increasing attachment security and weight gain: a clinical study of adolescents and their parents at an outpatient ward. Eat Weight Disord 2023, 28, 82. [Google Scholar] [CrossRef]
- Pelegí-Sisó, D.; de Prado, P.; Ronkainen, J.; Bustamante, M.; González, J.R. methylclock: a Bioconductor package to estimate DNA methylation age. Bioinformatics 2021, 37, 1759–1760. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY) 2018, 10, 573–591. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell 2013, 49, 359–367. [Google Scholar] [CrossRef]
- Zhang, Q.; Vallerga, C.L.; Walker, R.M.; Lin, T.; Henders, A.K.; Montgomery, G.W.; He, J.; Fan, D.; Fowdar, J.; Kennedy, M.; et al. Improved precision of epigenetic clock estimates across tissues and its implication for biological ageing. Genome Med 2019, 11, 54. [Google Scholar] [CrossRef]
- Lu, A.T.; Seeboth, A.; Tsai, P.C.; Sun, D.; Quach, A.; Reiner, A.P.; Kooperberg, C.; Ferrucci, L.; Hou, L.; Baccarelli, A.A.; et al. DNA methylation-based estimator of telomere length. Aging (Albany NY) 2019, 11, 5895–5923. [Google Scholar] [CrossRef] [PubMed]
- McEwen, L.M.; O'Donnell, K.J.; McGill, M.G.; Edgar, R.D.; Jones, M.J.; MacIsaac, J.L.; Lin, D.T.S.; Ramadori, K.; Morin, A.; Gladish, N.; et al. The PedBE clock accurately estimates DNA methylation age in pediatric buccal cells. Proc Natl Acad Sci U S A 2020, 117, 23329–23335. [Google Scholar] [CrossRef] [PubMed]
- Belsky, D.W.; Caspi, A.; Arseneault, L.; Baccarelli, A.; Corcoran, D.L.; Gao, X.; Hannon, E.; Harrington, H.L.; Rasmussen, L.J.; Houts, R.; et al. Quantification of the pace of biological aging in humans through a blood test, the DunedinPoAm DNA methylation algorithm. Elife 2020, 9. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol 2013, 14, R115. [Google Scholar] [CrossRef]
- Horvath, S.; Oshima, J.; Martin, G.M.; Lu, A.T.; Quach, A.; Cohen, H.; Felton, S.; Matsuyama, M.; Lowe, D.; Kabacik, S.; et al. Epigenetic clock for skin and blood cells applied to Hutchinson Gilford Progeria Syndrome and ex vivo studies. Aging (Albany NY) 2018, 10, 1758–1775. [Google Scholar] [CrossRef]
- Ratanatharathorn, A.; Boks, M.P.; Maihofer, A.X.; Aiello, A.E.; Amstadter, A.B.; Ashley-Koch, A.E.; Baker, D.G.; Beckham, J.C.; Bromet, E.; Dennis, M.; et al. Epigenome-wide association of PTSD from heterogeneous cohorts with a common multi-site analysis pipeline. Am J Med Genet B Neuropsychiatr Genet 2017, 174, 619–630. [Google Scholar] [CrossRef]
- Battram, T.; Yousefi, P.; Crawford, G.; Prince, C.; Sheikhali Babaei, M.; Sharp, G.; Hatcher, C.; Vega-Salas, M.J.; Khodabakhsh, S.; Whitehurst, O.; et al. The EWAS Catalog: a database of epigenome-wide association studies. Wellcome Open Res 2022, 7, 41. [Google Scholar] [CrossRef]
Figure 1.
Graphical abstract. Characterizing the social epigenome in Mexican patients with early-onset psychosis.
Figure 1.
Graphical abstract. Characterizing the social epigenome in Mexican patients with early-onset psychosis.
Figure 2.
Heatmap of correlations between socio-demographic, clinical characteristics and epigenetic age in psychiatric patients. The heatmap colors correspond to the significance (p < 0.05): green indicates a positive correlation, and red indicates a negative correlation. Blank spaces indicate no significant correlation. Best Linear Unbiased Prediction clock (BLUP); DNA methylation-based telomere length (DNAmTL); Dunedin Pace of Aging Methylation (DunedinPoAm38); Elastic Net clock (EN); Global Assessment of Functioning (GAF); Horvath clock (Multi-Tissue, Horvath1); Horvath clock (Skin & Blood, Horvath2); Levine clock (PhenoAge); Pediatric-Buccal-Epigenetic clock (PedBE).
Figure 2.
Heatmap of correlations between socio-demographic, clinical characteristics and epigenetic age in psychiatric patients. The heatmap colors correspond to the significance (p < 0.05): green indicates a positive correlation, and red indicates a negative correlation. Blank spaces indicate no significant correlation. Best Linear Unbiased Prediction clock (BLUP); DNA methylation-based telomere length (DNAmTL); Dunedin Pace of Aging Methylation (DunedinPoAm38); Elastic Net clock (EN); Global Assessment of Functioning (GAF); Horvath clock (Multi-Tissue, Horvath1); Horvath clock (Skin & Blood, Horvath2); Levine clock (PhenoAge); Pediatric-Buccal-Epigenetic clock (PedBE).
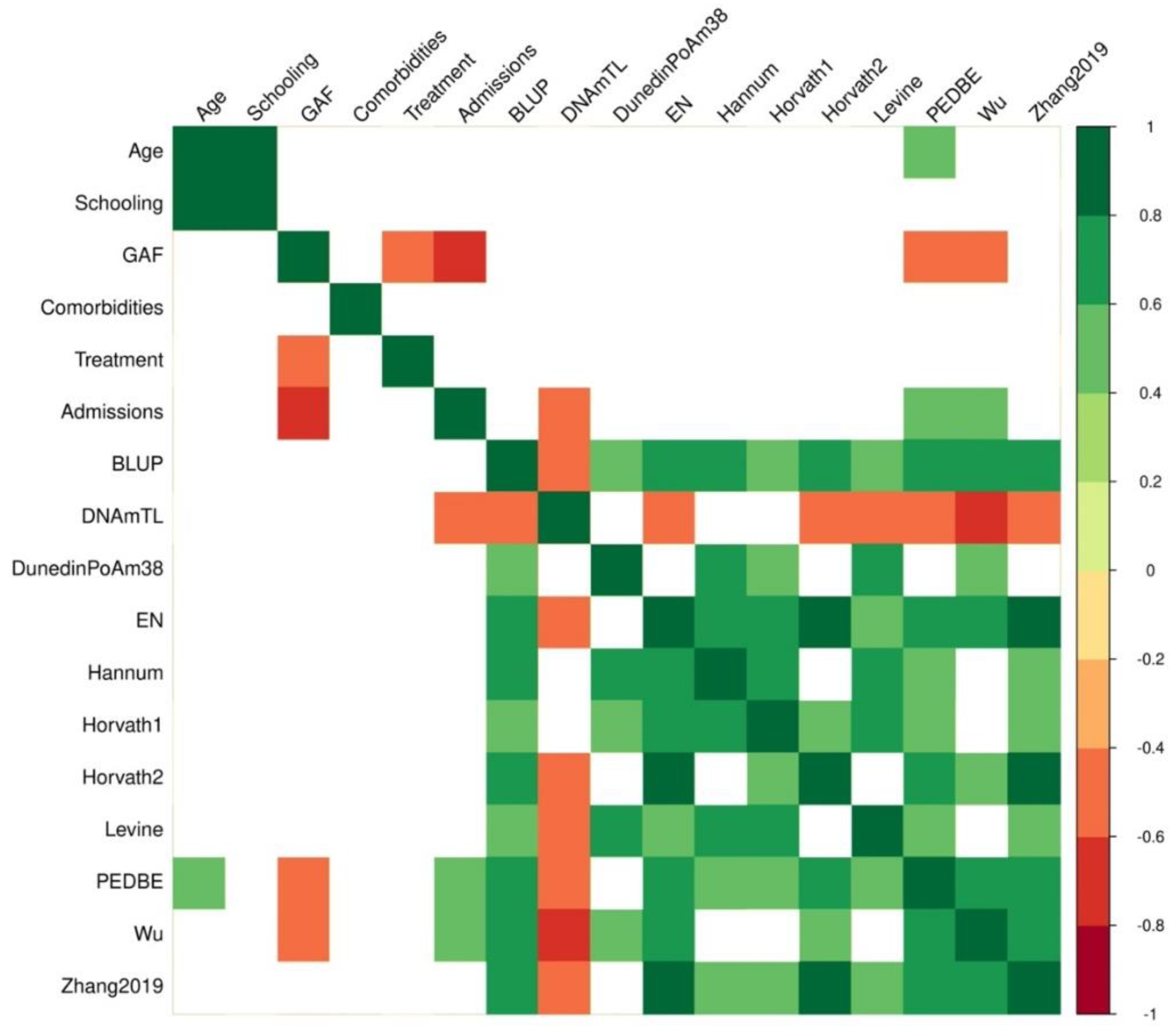
Figure 3.
Manhattan plot of EWAS analysis in EOP. Manhattan plot of CpG sites and p-values. The X axis represents the chromosome position, and the Y axis represents -log10(p). The red horizontal line parallel to the X axis denotes nominal significance (p < 1x10-05). All loci with a p-value < 1x10-05 are annotated to genes according to the human genome assembly (hg19). The association models are adjusted for sex, age, and five surrogate variables. Adhesion G protein-coupled receptor V1 (ADGRV1); Histone Cluster 1 H2B family member B (HIST1H2BB); Centrosomal Protein 164 (CEP164); Interferon Regulatory Factor 2 Binding Protein 1 (IRF2BP1); Microtubule-Associated Protein 1B (MAP1B); N-acetylated alpha-linked acidic dipeptidase 2 (NAALAD2); Sulfotransferase Family 1C Member 4 (SULT1C4). λ = 0.9783.
Figure 3.
Manhattan plot of EWAS analysis in EOP. Manhattan plot of CpG sites and p-values. The X axis represents the chromosome position, and the Y axis represents -log10(p). The red horizontal line parallel to the X axis denotes nominal significance (p < 1x10-05). All loci with a p-value < 1x10-05 are annotated to genes according to the human genome assembly (hg19). The association models are adjusted for sex, age, and five surrogate variables. Adhesion G protein-coupled receptor V1 (ADGRV1); Histone Cluster 1 H2B family member B (HIST1H2BB); Centrosomal Protein 164 (CEP164); Interferon Regulatory Factor 2 Binding Protein 1 (IRF2BP1); Microtubule-Associated Protein 1B (MAP1B); N-acetylated alpha-linked acidic dipeptidase 2 (NAALAD2); Sulfotransferase Family 1C Member 4 (SULT1C4). λ = 0.9783.
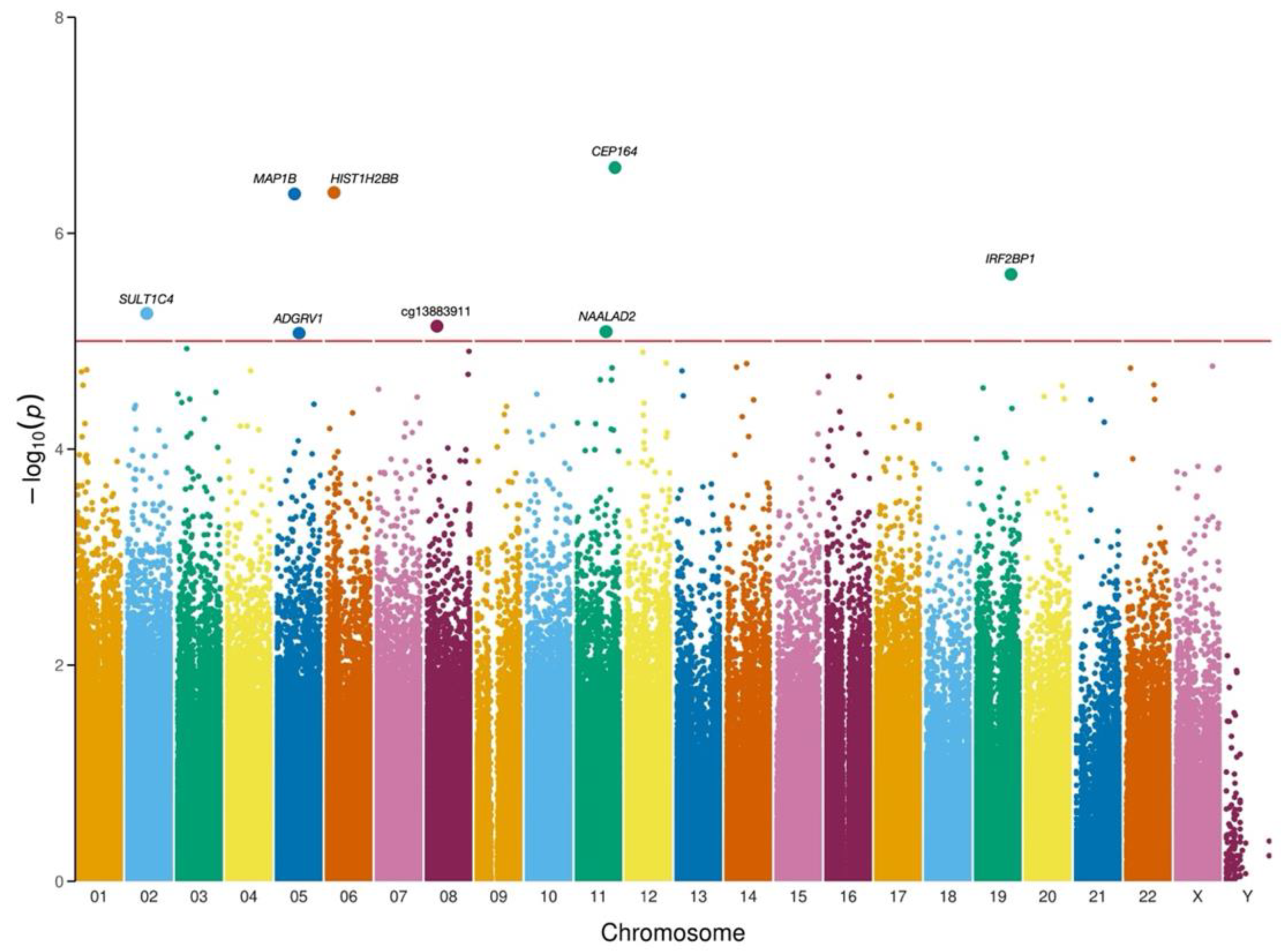
Figure 4.
Association between clinical characteristics and early-onset psychosis methylation risk score (EOP-MRS) in psychiatric patients. The beta values represent the estimated coefficient (β) from logistic regression analysis with a 95% confidence interval (CI95). Significant associations between clinical characteristics and MRS are displayed as solid points (p < 0.05), while non-significant results are displayed as hollow points. Each MRS calculated corresponds to a different cutoff p-value from the EWAS. The forest plot shows only significant associations among the sixty-eight clinical characteristics.
Figure 4.
Association between clinical characteristics and early-onset psychosis methylation risk score (EOP-MRS) in psychiatric patients. The beta values represent the estimated coefficient (β) from logistic regression analysis with a 95% confidence interval (CI95). Significant associations between clinical characteristics and MRS are displayed as solid points (p < 0.05), while non-significant results are displayed as hollow points. Each MRS calculated corresponds to a different cutoff p-value from the EWAS. The forest plot shows only significant associations among the sixty-eight clinical characteristics.
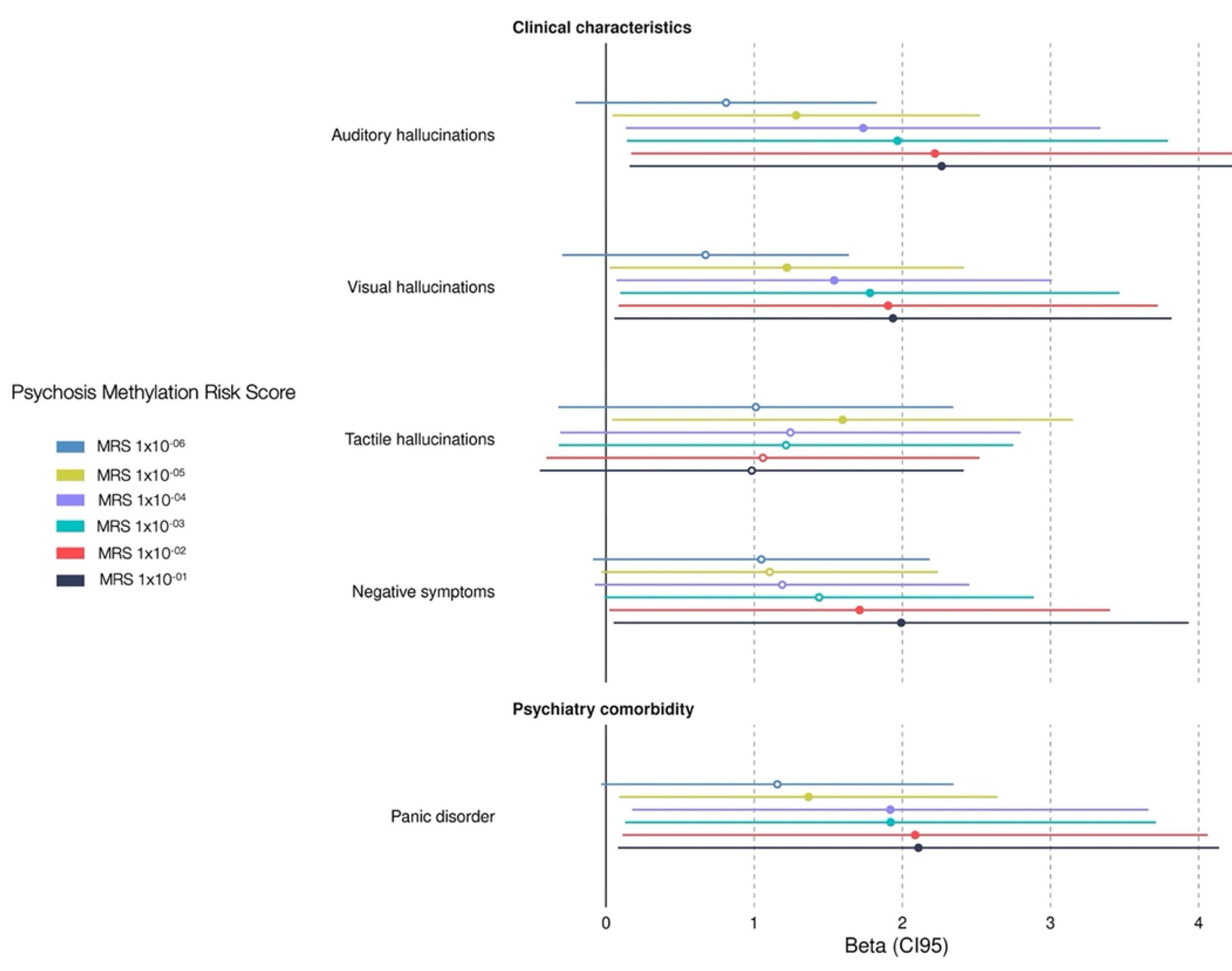

Table 1.
Socio-demographic and clinical characteristics of psychiatric patients.
| Characteristic | EOP (n=12) | Non-EOP (n=11) | p | |
|---|---|---|---|---|
| Age | years ± SD | 15.5 ± 1.56 | 13.36 ± 2.57 | 0.030a |
| Gender | Male, n (%) | 6 (50) | 7 (64) | 0.680c |
| Female, n (%) | 6 (50) | 4 (36) | ||
| Education | years ± SD | 9.3 ± 1.96 | 7.2 ± 2.05 | 0.023a |
| Body Mass Index | z-score, mean ± SD | 1.08 ± 1.15 | 0.67 ± 1.50 | 0.368a |
| Psychiatric admissions | n (%) | 7 (58) | 1 (9) | 0.027c |
| Total, median (min-max) |
1 (0-4) | 0 (0-1) | 0.016b | |
| Psychiatric comorbidity | n (%) | 12(100) | 9 (81) | 0.370c |
| Mood disorders, n (%) | 11 (91) | 8 (72) | 0.316c | |
| Anxiety and stress disorders, n (%) | 10 (83) | 4 (36) | 0.036c | |
| Conduct disorders, n (%) | 5 (41) | 5 (45) | 1c | |
| Neurodevelopment disorders, n (%) | 1 (8) | 4 (36) | 0.155c | |
| Eating disorder, n (%) | 4 (33) | 0 | 0.093c | |
| GAF | Total score, mean ± SD | 43.33 ± 15.14 | 74 ± 6.41 | 0.00003a |
| Minimal, n (%) | 1 (8) | 3 (27) | 0.017c | |
| Mild, n (%) | 1 (8) | 2 (18) | ||
| Moderate, n (%) | 2 (16) | 0 | ||
| Severe, n (%) | 8 (66) | 0 | ||
| Epigenetic age | Wu`s clock, mean ± SD | 11.08 ± 0.89 | 10.29 ± 0.78 | 0.015 |
Abbreviations: BMI = body mass index; GAF = global assessment of functioning; SD = standard deviation; T = Student's t-test; U = U Mann-Whitney test; χ² = chi-square test; a = statistic from a student's t-test; b = statistic from a Mann–Whitney–Wilcoxon test; c = statistic from a χ² test. Notes: Fisher's exact test was applied when values < 5. Bold values denote statistical significance, p < 0.05.
Table 2.
Correlations between socio-demographic, clinical characteristics and epigenetic age in psychiatric patients.
Table 2.
Correlations between socio-demographic, clinical characteristics and epigenetic age in psychiatric patients.
| Epigenetic calculator | Age r, p |
GAF r, p |
Admissions r, p |
|---|---|---|---|
| BLUP | 0.37, 0.079 | -0.40, 0.056 | 0.40, 0.056 |
| DNAmTL | -0.12, 0.555 | 0.27, 0.198 | -0.43, 0.038 |
| DunnedinPoAm38 | 0.26, 0.224 | -0.17, 0.434 | 0.19, 0.368 |
| EN | 0.26, 0.220 | -0.26, 0.217 | 0.40, 0.057 |
| Hannum | 0.25, 0.238 | -0.23, 0.274 | 0.29, 0.174 |
| Horvath-1 | 0.18, 0.398 | 0.13, 0.553 | -0.07, 0.742 |
| Horvath-2 | 0.03, 0.848 | -0.17, 0.427 | 0.34, 0.107 |
| Levine | 0.35, 0.100 | -0.05, 0.806 | 0.09, 0.672 |
| PedBE | 0.41, 0.046 | -0.53, 0.008 | 0.56, 0.005 |
| Wu | 0.30, 0.150 | -0.45, 0.027 | 0.49, 0.015 |
| Zhang | 0.24, 0.253 | -0.21, 0.324 | 0.35, 0.097 |
Abbreviations: R = Pearson´s correlation, Best Linear Unbiased Prediction´s clock (BLUP); DNA methylation-based telomere length (DNAmTL); Dunedin Pace of Aging Methylation (DunedinPoAm38); Elastic Net´s clock (EN); Global Assessment of Functioning (GAF); Horvath´s clock (Multi-Tissue, Horvath-1); Horvath´s clock (Skin & Blood, Horvath-2); Levine´s clock (PhenoAge); Pediatric-Buccal-Epigenetic´s clock (PedBE). Notes: Correlations were made with either Pearson or Spearman. Bold indicates p < 0.05.
Table 3.
Summary of the stepwise regression between epigenetic clocks, socio-demographic, and clinical characteristics in psychiatric patients.
Table 3.
Summary of the stepwise regression between epigenetic clocks, socio-demographic, and clinical characteristics in psychiatric patients.
| Epiclock | Age (years) | Sex | Schooling (years) | Comorbidity | Admissions | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| β (SE) | p | β (SE) | p | β (SE) | p | β (SE) | p | β (SE) | p | |
| BLUP | 1.786 (0.551) | 0.004 | - | - | -1.820 (0.544) | 0.003 | 0.496 (0.225) | 0.041 | - | - |
| DNAmTL | -0.071 (0.023) | 0.006 | - | - | 0.086 (0.022) | 0.001 | -0.134 (0.051) | 0.017 | -0.134 (0.051) | 0.017 |
| EN | 1.365 (0.589) | 0.032 | - | - | -1.475 (0.582) | 0.020 | - | - | - | - |
| Horvath-1 | 1.751 (0.706) | 0.023 | - | - | - | - | - | - | - | - |
| Horvath-2 | 0.604 (0.264) | 0.034 | - | - | -0.903 (0.261) | 0.002 | 0.310 (0.108) | 0.010 | - | - |
| Levine | 5.078 (1.109) | 0.001 | 8.413 (2.377) | 0.002 | -5.017 (1.205) | 0.001 | - | - | - | - |
| PedBE | 0.356 (0.107) | 0.003 | - | - | -0.314 (0.106) | 0.008 | - | - | - | - |
| Wu | - | - | - | - | - | - | - | 0.816 (0.375) | 0.042 | |
| Zhang | 1.938 (0.660) | 0.008 | - | - | -1.810 (0.691) | 0.016 | - | - | - | - |
Abbreviations: β = estimated coefficient; SE = standard error; T = t-value; BLUP = Best Linear Unbiased Prediction clock; DNAmTL = DNA methylation-based telomere length; EN = Elastic Net clock; Horvath-1 = Horvath clock (Multi-Tissue); Horvath-2 = Horvath clock (Skin & Blood); PedBE = Pediatric-Buccal-Epigenetic clock. Bold values denote statistical significance at the p < 0.05 level. Comorbidity values were represented as the number of psychiatric disorders; sex was denoted as a factor (0 = male, 1 = female); admissions were coded as a factor (0 = no, 1 = yes).
Table 4.
Top differentially methylated positions in EOP.
| CPG | Gene anotation | Chr | Position | Relation to Island | β | SE | T | p |
|---|---|---|---|---|---|---|---|---|
| cg24772138 | MAP1B (Body) | 5 | 71405539 | S Shore | -0.4483 | 0.0508 | -8.8233 | 4.30 x10-07 |
| cg26028573 | HIST1H2BB (Exon) | 6 | 26043587 | N Shore | 0.2340 | 0.0264 | 8.8374 | 4.22 x10-07 |
| cg05100917 | CEP164 (TSS200) | 11 | 117198460 | Island | -0.5790 | 0.0627 | -9.2343 | 2.48 x10-07 |
| cg20150189 | SULT1C4 (Body) | 2 | 108999219 | Open Sea | -0.3525 | 0.0498 | -7.0718 | 5.58 x10-06 |
| cg27181762 | ADGRV1 (Body) | 5 | 90195345 | Open Sea | -0.5829 | 0.0860 | -6.7701 | 9.02 x10-06 |
| cg13883911 | - | 8 | 33430120 | Open Sea | 0.3104 | 0.0449 | 6.9051 | 7.26 x10-06 |
| cg08523325 | NAALAD2 (Body) | 11 | 89901450 | Open Sea | -0.3064 | 0.0448 | -6.8337 | 8.14 x10-06 |
| cg06583549 | IRF2BP1 (Exon) | 19 | 46387962 | Island | -0.2669 | 0.0350 | -7.6212 | 2.40 x10-06 |
Abbreviations: Chr = chromosome; β = effect size; TSS = transcription start site; T = t-statistic; SE = standard error. Adhesion G protein-coupled receptor V1 (ADGRV1); Histone Cluster 1 H2B family member B (HIST1H2BB); Centrosomal Protein 164 (CEP164); Interferon Regulatory Factor 2 Binding Protein 1 (IRF2BP1); Microtubule-associated protein 1B (MAP1B); N-acetylated alpha-linked acidic dipeptidase 2 (NAALAD2); Sulfotransferase Family 1C Member 4 (SULT1C4). CpG sites ordered by statistical p-value from EWAS analysis in early onset psychosis. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



