
Submitted:
22 March 2025
Posted:
25 March 2025
You are already at the latest version
Abstract
In the past decades, the uncontrolled growth of industrialization led to a steady increase in heavy metals (HMs) in aquatic environments, especially coastal ones, causing negative impacts on them and serious health risks to humans. Indeed, since HMs are non-degradable compounds, they rapidly accumulate in environmental matrixes, such as sediments, and in marine organisms. Microalgae HM tolerance and sorption ability varies by species and metal type, with higher concentrations adversely affecting physiological processes and impairing cell morphology. Below certain thresholds, they can be considered promising candidates for HM removal, offering a viable alternative to conventional methods - often relying on costly chemicals, and producing hazardous by-products. Indeed, these microorganisms exhibit a high surface-to-volume ratio and consequent sorption capacity, which together with rapid growth rates and high biomass yields make them suitable candidates for bioremediation. HM sequestering can employ either dead biomass, which passively adsorbs metals via cell- wall interactions, or living organisms that actively transport HMs into cells. The present review provides an overview of microalgae-based HM removal capability, and HM-driven stress to which they are subjected in case of serious HM pollution.
Keywords:
heavy metals (HMs)
; microalgae
; HM-driven stress
; biosorption
; compartmentalization
; microalgae-based HM removal
1. Introduction
The term heavy metals (HMs) refers to metals with an atomic number greater than 20 and a density that exceeds 5 g/cm3 [1,2]. HMs are typically found in the Earth’s crust and can derive from natural sources (volcanic eruptions, weathering and erosions of soils and rocky substrates, wind-blown dust), but a dramatic increase of their concentration in the past decades is mainly due to the rise of industrialization and urbanization. Significant high HM loads, indeed, are usually due to anthropogenic impact, as a consequence of their use for industrial, agricultural, pharmaceutical and domestic applications, with the highest contribution generated by mining and smelting activities and waste water and solid waste discharges [3,4,5,6,7]. Figure 1 shows both natural and anthropogenic metal sources.
Natural release routes are due to weathering of metal-containing rocks and rock outcroppings, volcanic eruptions, wind dust particles from desertic regions like the Sahara, forest and prairies wildfires emission of volatile HMs and sea sprays and mists delivery of HMs upon bubble bursting [2,4]. On the other end anthropogenic discharge is due to their use in domestic, industrial and agricultural activities. HMs are present in partially treated or untreated waste waters leaking through sewage outfalls into surface water bodies, organic and inorganic waste, expired/used batteries and medical devices, detergents, cosmetics and pharmaceuticals and in urban run-off as a consequence of their large use in domestic applications [4]. In agriculture, they are largely employed to produce fertilizers, mostly of the phosphate-based type, are constituents in pesticides and fungicides and are contained in sewage sludge and in animal manure; continuous crops irrigation from deep wells can also lead to their build-up [4,6]. The main release route due to industrial applications involves ore mining and metal processing and refining, including dust generated from their transport, metal tailings erosion and runoff, smelting at high temperatures delivering HMs in vapour form, thus subject to dry or wet deposition, finishing, recycling, corrosion, and leaching into soil and water [4,7]. Other industrial activities, such as combustion of fossil fuel in refineries and coal incineration, and processing in plastics, textiles, microelectronics, wood preservation and paper and cosmetic production are also responsible for HM loads in the environment [4,8]. Finally, other sources include incineration of municipal waste and landfills, transportations (particularly fuels and auto vehicles engines and their lubricants), fly ash from coal burning and corrosion of commercial waste products. [4,6,7].
Metals are released both in elemental form and as part of both organic and inorganic compounds, especially prominent in areas of mining: leached out metals are carried from acid waters, and can seep into underground waters bodies or be transported into streams, lake and rivers until they reach the sea in the form of suspended particles or dissolved species, with the latter having the most deleterious effects [4,9]. Eventually, the largest fraction of HM is either deposited onto suspended particulate and immobilized by adsorption, coagulation or flocculation, or co-precipitated with other metals forming insoluble complexes, ultimately being transferred to the bottom of water beds into sediments, while only a small portion of these metal remains in its dissolved form [10]. For this reason, heavy metal content and distribution in sediment adjacent to areas with high urbanization and industrialization can be indicative of the impact of human activities on aquatic ecosystem and highlight potential risks linked to industrial and municipal waste discharge [10].
Most detected HMs include cadmium (Cd), chromium (Cr), copper (Cu), lead (Pb), nickel (Ni) and zinc (Zn) [11,12]; Table 1 lists metal released in association to their specific sources.
Mainly HM-impacted areas are localized along the coasts, with nefarious consequences on marine environments and serious threats to human health. Indeed, in addition to their toxicity, heavy metals are considered persistent pollutants, because they cannot be degraded and therefore can remain for a long time in the environment [13]. Due to this, they rapidly accumulate in environmental matrices, such as sediments [14] and marine organisms: in this latter case they also pose considerable biomagnification risks [15,16].
Sediments at the bottom of river beds or on the sea floor are considered a storage basin for metals [4,17], both acting as a sink and as source, with their availability to organism being influenced by the environmental geochemistry.
Assessment on the level of sediment HM contamination is usually performed by calculating parameters such as enrichment factor (EF), allowing discrimination of metals from natural and anthropogenic sources by normalizing sample HM concentration data to a reference element (Fe) and comparing them to available data indicating metals crustal average (normalized on the same element), better if referring to local background values: EF near 1 denotes the metal natural origin, while significantly higher values indicate human activities impact [18]. HM concentrations above background values were assessed in several studies conducted on sediments from South and South-East Mediterranean Sea, specifically in Tunisia, along Moroccan and Egyptian coasts, along the south-eastern coast of Spain and in South Turkey [19,20,21,22,23]. Along the Italian coasts, HM concentration above background values have been found in sediments from the Adriatic [24] and Tyrrhenian [25,26] seas. Notably, in the Southern Tyrrhenian sea, Sprovieri et al. report significant enrichment in Cu, Pb, Zn and Hg in Naples harbour, with EF values above 3 [27], while values considerably higher than the one reported in literature for other polluted rivers in the Mediterranean Sea are found for several metals (including Cd, Cr, Cu, Hg, Ni, Pb and Zn) in the Sarno River and the surrounding area [28,29], highlighting the significant contribution of this river to metal loads into the Southern Tyrrhenian area and the consequent threat this constitutes for the health of this aquatic environment [29].
HMs in sediments can be absorbed by aquatic microbes, while some benthic organisms even feed directly on it, and are thus subject to HM accumulation while they mobilize these compounds in the process [6,16,17]. In addition to these bioturbation phenomena, at the interface between sediment and water column HMs can rapidly shift from one phase to another due to seasonal dynamics and hydrodynamic conditions, with factors controlling their adsorption and desorption ranging from pH—one of the most critical parameters, as low pH may result in metal release from the matrix since H+ ions compete with metal cations for binding sites—to temperature, salinity, metal redox state and sediment’s organic carbon content and grain size [2,17]. Resuspension of HMs in water can be of natural origin, as consequence of tidal action, storms or seismic events, but most often has anthropogenic causes, due to dredging, shipping traffic and industrial discharges [30,31,32,33,34]. The National Oceanic and Atmospheric Administration (NOAA) established sediment quality guidelines (SQGs), that are aimed, trough the evaluation of established parameters, at assessing the risk posed to the aquatic ecosystem by metals available in sediments. Those parameters are the Effects Range Low (ERL) and the Effects Range Medium (ERM), corresponding to the concentrations at which metal exposure causes detrimental effects on either a small percentage or a greater proportion of biota in aquatic environments [18,35]. During their assessment of metal loads in the Sarno River, Montuori et al. found that, based on ERL/ERM values, HM concentrations in water sampled raised serious concerns for ecosystem integrity [29].
Indeed, metals resuspended in the dissolved phase constitute the bioavailable fraction, capable of interacting with living organism and entering the food chain, growing in concentration as HMs move up to the higher trophic levels—a process termed “biomagnification”. As a matter of fact, even though HMs can directly enter into some organisms from water or sediments through ingestion or absorption, as in the case of microalgae and filter feeders, organisms tend to accumulate more and metal; in addition, grazing and predatory life-styles associated with progressively higher levels of trophic chain correspond to higher HM concentration in the organisms’ body, so that larger top predators are the most impacted [6,16]. Danovaro et al., for example, showed that tissue concentration of As, Cd and Cr of marine nematodes with predator lifestyles was higher than members of the same species living as microalgal grazers and deposit feeders, ultimately assessing that these metals, in their experimental conditions, were indeed subject to biomagnification [16]. This not only constitutes a serious issue for aquatic ecosystems, but it is also a threat to human health, as some known bioaccumulators are predatory fishes or filter feeders commonly included in our diets. In a study conducted by De Mora, et al. [36], two predatory fish species of commercial importance had high Cd levels, while Mytilus galloprovincialis, commonly known as Mediterranean mussel, is a known metal bioaccumulator and has been often used as indicator of environmental contamination of Cd, Cr, Cu, Hg, Mn, Ni, Pb and Zn [37,38,39,40,41]. As highlighted by Joksimović, Castelli, Perošević, Djurović and Stanković [38], HM concentration detected in Mytilus galloprovincialis’ soft tissues seem to be correlated with the ones present in the sediments.
Despite their perceived toxicity, at low concentrations some metals are actually central to cell metabolism and functions; they are therefore classified as essential and are also sometimes referred to as “micronutrients” [42,43]. In plants, among others, cobalt (Co), Cu, iron (Fe), manganese (Mn), molybdenum (Mo), Ni and Zn are known to participate in growth, biosynthesis, membrane integrity and metabolites production [42]. Cu, for example, is a constituent in various oxidases and plastocyanins and plays a role in oxidation, photosynthesis, protein and carbohydrate metabolism and cell wall metabolism, Co and Mo are part of enzymes involved in N2 fixation, Mn is present in many enzymes in charge of photoproduction of oxygen in chloroplasts, Fe is important for photosynthesis and N2 fixation, and Zn participates in carbohydrate, lipid and nucleic acids metabolism [44]. Nevertheless, either level of said essential elements above certain thresholds or other metals that have no known biological role and are therefore considered non-essential to organisms, can have toxic effects in plants [4,45,46], animals and on the health of humans [47,48], and, ultimately, on the environment [49,50].
Metal toxicity mechanisms include 1) triggering of oxidative stress status in cells, resulting in depletion of ROS-scavenging compounds and consequent lipid peroxidation, leading to membrane damages, proteins oxidation, damaging and oxidation of nucleic acids, and failure of DNA repair mechanisms, ultimately leading to cell death [51]; 2) competition with other necessary metal ions and inhibition of their uptake by the cell, thus causing deficiency of essential metals and disruption of the processes they participate in (e.g., photosynthesis in plants [45,46]); 3) substitution of metal co-factor or direct interference on enzymes and subsequent impairments of cellular processes [6] (e.g in plants, beside photosynthesis dysregulation, reduced efficiency of oxidative stress response and of carbon fixation and energy production, leading to impaired cell growth [46].
2. Conventional Methods for HM Removal
Conventional methods for HM removal include chemical precipitation, coagulation/flocculation, ion exchange, membrane technologies and electrochemical technologies [52,53,54,55,56].
Chemical precipitation is still largely employed to treat industrial effluents, as it is effective, relatively cheap and does not involve complicated procedures, relying on reaction of metals with different compounds, mostly involving hydroxides or sulfides, to produce insoluble precipitates that can be subsequently filtered, centrifuged or subject to sedimentation in order to separate them from water [57]. Precipitation with hydroxides, with lime as the preferred base, is particularly easy to perform by using appropriate pH range to attain low solubility of different metal hydroxides and remove them, but it produces low density sludge, that can be difficult to dispose, and the final obtained metal concentration is still not very low [54]. On the other end, solubility of sulfide metal precipitates is lower in comparison and as those compounds, unlike metal hydroxides, are not amphoteric, there’s no need to tightly control the pH since the risk of releasing one metal species when trying to solubilize another in effluents containing metal mixtures is minimal, but this method can present some risks, since it can lead to the production of toxic fumes. In addition, separation of metal sulfides from water can be problematic, since precipitates are colloidal and thus not prone to agglomeration and precipitation. Finally, precipitation methods are applicable only to matrices with very high metal contamination and do not allow for the recovery of metal and its potential reuse [52,54].
On the other hand, coagulation/flocculation is commonly employed in heavy metal contaminated wastewater treatment, as it is able to remove soluble compounds, colloidal particles and fine solid suspensions through destabilization of forces that keep particles apart via coagulants and subsequent formation of flocs [52,53]. Coagulants, usually including aluminium, ferrous sulphate and ferric chloride, neutralize particle charges and lead to precipitate formation; polymers like polyacrylamide or polyferric sulfate are usually added to form link between flocs and promote formation of larger particle agglomerates, i.e., flocculation. The actual separation is subsequently carried out by sedimentation or filtration. This method is not entirely suitable to treat HMs wastewater, as flocculants used in removal of other compounds are not suited for this purpose and new flocculants are being investigated. In the meantime, this method is used in combination with others [52,53].
Ion-exchange instead, along with high treatment capacity, exhibits high efficiency as well, as it is suitable for the selective removal of metal ions and the re-use of the produced material. This is achieved with the use of ion-exchange synthetic or natural resins , with the former being preferred for the removal of a wide range of metals, which can exchange metal cations in solution with their own [52]. The preferred ion-exchange materials are sulfide-based, as these compounds are known to form strong covalent bonds with heavy metals through sulfonic acid groups (-SO3H), or weakly acid resins possessing carboxylic acids groups (-COOH): their H+ ions can be exchanged with metal cations as the solution from which they need to be removed passes through the column [55,57]. Even tough highly efficient and selective, also this method has some drawbacks, since it is influenced by a plethora of parameters (like initial metal concentration, ionic charge, contact time, temperature and, most importantly, pH) which need to be tightly controlled and, in addition, is quite expensive as fully saturated resins need to be regenerated by chemical reactions, generating secondary pollution and limiting its possible application for the treatment of large quantities of industrial wastewaters [52,55].
Membrane filtration technologies are easy to use, offer the possibility of scale up applications, exhibit high separation efficiency and selectivity and have low resources requirements: they rely on various kinds of membranes as selective barriers, allowing for selection of retained material [52,56]. Among membrane technologies, ultrafiltration (UF) uses low pressure applied through the membrane to remove dissolved and colloidal material through size exclusion [56,57]. This method would be normally unsuitable to remove HMs, as the pore size is larger than the hydrated dissolved forms of HMs, so either surfactants with opposite electrical charge as sodium dodecyl sulfate (SDS) or water-soluble polymers are added to aggregate metal ions into micelles or complexes with high molecular weight in order to exceed membrane pore size and be subsequently removed: surfactants and polymers are often recovered to avoid excessive costs and secondary pollution, and metal can also be recovered [57]. Reverse Osmosis (RO) relies on solution diffusion thanks to a semi-permeable membrane allowing passage of liquid to be purified while retaining contaminants, and is efficiently used in desalination and contaminant removal, but has the limitation of high energy requirements to generate pressure across the membrane and costs associated with membrane regeneration [57]. Reverse osmosis is often used in combination with nanofiltration, a process in between RO and UF with relatively lower energy requirements and high removal abilities, to achieve efficient separation of both charged and uncharged solutes [56]. Finally, electrodialysis (ED) exploits a charged membrane, using an electric potential difference as a driving force instead of pressure to separate metal ions from one solution to another. The mechanism is anion or cation exchange, depending on membrane’s type and purpose, as it has been used in the past for water desalination but has also shown promising abilities in heavy metal wastewater treatment [56,57].
Finally, electrochemical technologies provide an environmentally friendly alternative to remove and recover valuable metals in their elemental state from waste waters, while generating minimum associated solid waste compared to other traditional metal remediation methods. These techniques allow separation of metal ions exploiting a cathode surface, and comprise electrocoagulation, electro flotation and electro deposition [57,58]. In electrocoagulation (EC), aluminium or Fe electrodes are electrically dissolved at the anode into their ions to generate coagulants directly in the waste water to be treated and hydrogen gas released from the cathode aids in retrieving the particles by inducing their flotation, while in electroflotation (EF) simply exploits electrolysis of hydrogen/oxygen gases produced by electrolysis of water to induce contaminant flotation are retrieval by separation of solid and liquid phase [57]. Electrodeposition exploits an electrochemical cell with electrical current flow for the reduction and deposition of dissolved metal cations on the electrode, and can be used for retrieval of metals present in wastewaters also containing complexing chelating agents like EDTA and many others [57,58]. Electrochemical technologies are considered environmentally friendly techniques compared to other traditional methods and have the added benefit of granting valuable metal retrieval, even though their relatively high costs linked to considerable electric energy supply has traditionally limited their application on large industrial scales [57].
All in all, very different conventional methods for the removal of heavy metals from waste waters are available, each with their own advantages and limitations: chemical precipitation is simple, inexpensive and easily automated, but generates toxic sludges and is efficient only when metal concentration is very high; coagulation-flocculation is also economic and easy to use but requires the use of chemicals and generate sludges, even if comparatively easier to remove due to better settling and dewatering, and metal cannot be removed completely; ion exchange is highly efficient, fast and effective and does not generate sludge, but is suitable only for low metal concentrations, and high costs and secondary pollution are associated to the regeneration of the used resins; membrane filtration also avoids the use of chemicals and exhibits high removal efficiency but has high costs due to energy requirements and membrane fouling and requires a comparatively complex process; electrochemical technologies are simple, rapid, require few chemical and generate limited waste products but are ineffective when metal ion concentration in waters is too low, and high costs are associated to required equipment and electric supply [7,57].
As conventional chemical-physical treatments are quite effective but subject to the abovementioned drawbacks, several non-conventional approaches have been implemented recently to remove toxic metal from wastewaters, including microbial fuel cells, Fenton-like reactions, nanotechnologies and variegate adsorption-based techniques [7,57,59,60,61].
Microbial fuel cells (MFCs) use selected microorganisms to catalyse electric current production starting from organic matter dissolved in effluents to be treated which, in turn, prompts attraction of metal ions, possessing higher redox potential then the anode, to the cathode and thus their capture and recovery. The system essentially exploits organic matter decomposition in anaerobic conditions by microbes positioned on the anode to generate protons and electrons passing either through a special proton exchange membrane or along an external wire respectively to reach the cathode at the other side, where they react with the oxygen present reducing water and closing the circuit, allowing for generation of a continuous flow of electricity, ultimately powering metal recovery from waste water with clean energy, even though this method is still restricted to small-scale application only due to its low production rates and limited efficiencies [7,59]. Nanotechnology-based remediation methods are a broad class which entails the use of special composites, called “nanomaterials”, possessing peculiar high surface-to-volume ratio and distinctive electronic, magnetic and optic properties; they comprise, among others, nanocarbon (in the form of carbon nanotubes, graphene and other carbon derivatives), 2-D single layer materials and nanoparticles [7]. In the last years, they have been successfully applied for the removal of HMs from contaminated industrial waste waters with high efficiency and contained costs, but raising concerns regarding their potential, largely unexplored toxicity limits their use. In some cases, those methods are applied in combination with others to tackle this issue: for example, carbon nanotubes can be immobilized in environmental-friendly adsorbents like calcium alginate to avoid their accidental discharge in waters [7,57,62].
Indeed, adsorption is regarded as one of the most promising method, as it is economic, can be designed with flexibility to suit a wide variety of settings and is highly effective, in some cases offering the added benefit of being a reversible process and thus suitable for adsorbent regeneration through desorption [7,57]. Adsorbent generally used have a large specific surface area, are resistant and stable in a range of temperatures and do not generate secondary pollution through regeneration or processing, ultimately making this method efficient and able to remove significant quantities of metal with high selectivity with relatively low costs and environmental impact [7]. Several materials are employed as adsorbents, including activated carbon (AC), the most widely used due to its high adsorption capacities, synthetic or natural polymer-based materials, low-cost adsorbents obtained from agricultural and industrial residues and many more [52]. AC expensive production process and its need for frequent reactivation after usage due to rapid saturation makes this adsorbent quite costly, and generates secondary pollution [7]. Some synthetic polymeric materials also generate secondary pollution, so often natural polymers like alginate, chitosan, cellulose, lignin and silk are preferred [7,61,63,64]. For example, composites based on alginate, an anionic polysaccharide component of algae’s cell wall, have been showed to remove efficiently multivalent metal cations due to free hydroxyl and carboxyl groups present along the chain of its major component, alginic acid. In addition, alginate is able to form non-toxic, bio-compatible strong and stable matrices, sometimes used to embed other types of adsorbents for enhanced removal efficiency [63]. In general, polymeric material are biodegradable and biocompatible, economic, can be easily adapted to different approaches and show high adsorption abilities, even if they are often unstable under high temperatures and varying pH conditions and have been thus yet scarcely applied in industrial settings [7].
Among the plethora of available adsorbent material, biosorbents are gaining increasing attention as they are easily available in great quantity, low-cost, environmental friendly and highly effective alternatives for the removal of heavy metals, particularly at the low concentrations they show in dilute waste waters [52,61]. For this purpose, a variegate range of biomasses can be used, encompassing non-living biomass waste from other sources and algal and microbial biomass [57]. The main conventional, validated methods for HM removal are reported in Table 2.
In summary, to tackle the issue of heavy metals harmfulness, toxicity and persistence in the environment, several approaches have been implemented for their remediation. To overcome disadvantages associated with conventional methods (chemical precipitation, coagulation/flocculation, ion exchange, membrane technologies and electrochemical technologies), such as high costs and energy demands, excessive use of chemicals, high amount of toxic by-products, incomplete HM removal or low efficiency for metal concentrations below certain thresholds, several non-conventional methods have been proposed, including microbial fuel cells, Fenton-like reactions, nanotechnologies and adsorption. Under the ever-growing demand for efficient, cost-effective yet sustainable environmental-friendly new methods to face heavy metal pollution, bioremediation—remediation of contaminant mediated by living or dead biomasses—is considered a promising field [65].
3. Microalgae as Sentinel Species for HM Pollution
Microalgae are a diverse group of unicellular eukaryotes capable of colonizing a wide range of aquatic ecosystems, and have a capital ecological role. Being photosynthetic organisms, they fix CO2 from the atmosphere [66,67], releasing O2 in the process, to produce a myriad of different organic compounds. In addition to their role as primary producers, they constitute a food source for several organisms, like zooplankton [68] and filter-feeders [69] forming, in turn, the base of food webs in aquatic environments [70,71].
Nowadays, microalgae possess a constantly increasing commercial value, as they are employed in a wide variety of applications [67]. Their metabolites can be used in the pharmaceutical and cosmeceutical field and in the food industry [72,73,74], while microalgae themselves can be employed for bioremediation of contaminants [75,76] or as bio-factory for simultaneous carbon capture and production of bioenergy, including several biofuel applications, and other added-value compound production [73,74,76,77,78]. Due to of their high surface-to-volume ratio and consequent uptake efficiency, microalgae can be easily cultivated in controlled conditions, and some species exhibit quite high growth rates with relatively good biomass yields versus minimal space and nutrient requirements compared to other organisms, making them attractive from a biotechnological point of view. Additionally, they display high variability among groups adapted to survive in a spectrum of different conditions producing, in turn, a broad variety of compounds and can be used in a bio-refinery approach [73,79]. In light of the ever-growing demands of eco-friendly approaches to face the issue of pollution generated by human activities, researches spanning the last decades have showed great interest in microalgae: these organisms have been extensively studied as effective candidates to rid aquatic environments of organic and inorganic contaminants, including nano- and microplastics, pesticides and heavy metals [67,79,80,81,82,83,84,85].
In addition, living microalgal biomasses exposed to heavy metals typically show several recognizable features of stress generated by these contaminants in a dose-dependent [86], species-specific [87] pattern, and may be thus proposed as markers for rapid detection of environmental perturbation. Indeed, to study contamination phenomena, a good bioindicator should show, among other characteristics, a clear dose response, a certain sensitivity and relevance in the ecological network under exam [88].
Impact of heavy metals on microalgal cells has been evaluated in several studies through assessment of alterations in: 1) dynamics of cell division, usually observed as a reduction in their growth rates [86,87,89,90,91,92]; 2) cell’s energy status, either measured by photosynthetic efficiency [92,93,94], also evaluated by chlorophyll content [90,91,92,95,96] or measured by ATP content [93]; 3) cellular and organelles morphology, often observed as impaired cell/organelle size, shape, cell wall’s architecture, loss of membrane integrity and increased vacuolization [87,89,90,91,92] and lipid bodies formation [93]; 4) cell homeostasis and redox balance, usually observed as generation of ROS species [92,95], often assessed by antioxidant enzymes activity [86,89,91,96,97,98] or non-enzymatic pathways [86,90,96,97,98], lipid peroxidation products [86,89,93,95,96,97], protein oxidation [97] and changes in expression profiles of several genes, included the ones involved in HM stress detoxification [93,99,100]; 5) biochemical profile and composition, often assessed trough evaluation of protein and lipid content [90,91,96,101].
When heavy metals first enter in contact with microalgal cells they interact with functional groups (like, for example, -COOH, -NH2, -OH) laying on cell wall [102,103,104] and are adsorbed through a passive process occurring in both living and dead cells, mediated by electrostatic interactions and therefore independent of cell metabolism. Subsequently, an additional process may take place in living microalgae, leading to active metal sequestering inside the cells aided by ion pores, channel or transporters [105,106]. HM accumulation in microalgal cells can elicit stress through various mechanisms, summarized in Figure 2.
Mechanisms proposed to explain hallmarks of stress upon heavy metal accumulation, summarized in Figure 2. are:
1) Competition of metal ions with other essential metal ions for binding sites, resulting in the inhibition of their uptake, through mimicry or usage and blocking of dedicated channels, leading to essential metal deficiency [99,107,108];
2) In the case of redox-active metal, direct generation of reactive oxygen species. Redox-inactive metal instead could bind to microalgal molecules in charge of ROS scavenging (like glutathione i.e., GSH, ascorbic acid, tocopherol, carotenoids, flavonoids, phycocyanin and many more) or enzymes tasked with ROS response—like ascorbate and glutathione peroxidases (APX and GPX), catalase (CAT), Fe or Cu/Zn superoxide dismutases (Fe-SOD and Cu/Zn-SOD) and glutathione reductase (GR)—through their sulfhydryl groups thus resulting in their depletion and consequent accumulation of ROS normally produced during cell metabolism. Ironically, some oxidative stress detoxification enzymes are metalloproteins (like SOD), so essential metal deficiency can also impair their function. ROS, in turn, can oxidize nucleic acids, resulting in DNA damage and failures of its repair mechanisms, and can cause lipid peroxidation, with consequent alteration of cell and/or organelles membrane permeability alteration and damage, and protein oxidation and loss of function [99,108,109,110];
3) Direct binding to proteins exhibiting sulfhydryl groups, leading either to impairment of ROS scavenging system, as explained above, or displacing other metal ions which act as cofactor for certain proteins:
4) Essential metal co-factors deficiency or displacement by internalized HMs ultimately leads to the inhibition of the processes where these are involved. As an example, soluble proteins as Fe-containing cytochrome 6 and Cu-containing plastocyanin participate in photosynthetic electron transport in thylakoids and electron transport chain in the mitochondria, and their inhibition can result in disruption of energy harvesting and ATP synthesis. Moreover, Mg2+, necessary for chlorophyll functioning, can often be substituted by several other divalent cations, like Ni2+, Cu2+, Zn2+, Cd2+, Hg2+ and Pb2+. Energy available for several cellular processes is consequently depleted [108,111,112];
5) Microalgal cells in stressed condition sometimes exhibit vacuolized cytoplasm or organelles, often in an attempt to sequester the damaging agents in order to limit their detrimental effects; lipid bodies can be formed as storage of useful substances number of starch granules can increase, cell division rate decreases due to reduced energy availability [87,112,113,114].
Table 3 summarizes the main effects of HMs on microalgae described in literature.
In literature, several works are based on experiments of exposure of microalgae to HMs [144]; the table above represents, to the best of our knowledge, the most significant studies in which the detrimental effects of HMs on microalgae have been clearly described and not only hypothesized on the basis of macroscopic effects (such as, for example, algal growth inhibition and/or death).
As previously mentioned, particularly high concentrations of HMs can be found in water and sediments as a result of human activities, and their accumulation poses a formidable threat to aquatic environments, therefore their presence and toxic effects on the organisms that inhabit them need to be closely monitored. Commonly used and long-established methods to detect the presence of heavy metals can be based on electrochemistry [145], spectroscopy or chromatography. Techniques based on the latter two can deliver excellent specificity, accuracy and sensitivity but demand highly-trained operators and extensive pre-processing procedures. In addition, methodologies like fluorescence spectroscopy [146], atomic absorption spectrometry [147], colorimetry [148], and high-performance liquid chromatography (HPLC) [149] require specific lab setting, are time-consuming and cost-ineffective, resulting thus unsuitable to regularly monitor areas subjected to heavy metal pollution by carrying out quick on-site measurements. As regulatory standards establishing the permitted metal levels are constantly reviewed worldwide, the demand for miniaturized, portable analytical devices to ensure such limits are being met has increased leading to the design of bio- and biomimetic electrochemical sensors [150] and to various easily-transported compact implementations based on electrochemistry [151], Surface-Enhanced Raman Scattering spectroscopy (SERS) [152], integrable microfluidics [153] and easily carried colorimetry apparatuses [154]. Still, these devices often have limitations, being forced to rely on compound like dyes or nanoparticles to relay the signal. An interesting attempt at a sustainable implementation of this concept has been made by [155], where carbon dots used as HM sensors in aquatic medium are produced from biochar derived from microalgae biofuel.
In any case, it should be underlined that all the analytic systems reported focus on the detection of the metal itself and on its quantification, but this is not always direct indication of the stress that such contaminant imposes on the ecological network of the aquatic environment. These methods fail to reveal the effects that HMs may trigger on the organisms exposed to it, brought about by its bio-available component. Toxicity of a metal for aquatic life is influenced by various factors: in some cases, complexation with organic ligands present in water can reduce the amount of bio-available metal, i.e., the fraction able to contact and/or enter living cells [156]. In other cases some organisms, like microalgae, developed a set of strategies to cope with their toxic effects and limit consequent damages, thus resulting in different species exhibiting distinct sensitivities for various metals [157]. For example, at nanomolar concentrations, Cu is essential for growth, metabolism and survival of several organisms, including microalgae [156,158], and it becomes harmful for living being only above certain thresholds, potentially leading to abnormal phenotypes across a broad spectrum of freshwater and marine organisms [159,160]. When exceeding certain concentrations, this metal has also been shown to impact microalgal cells [161], perturbing growth and photosynthetic efficiency [162], compromising membrane integrity by altering its permeability [89] and giving rise to alterations in cell and organelles shape, possibly due to metal adsorption and/or internalization [87]. For such reasons, evaluation of metal loads in aquatic environments should have a particular focus on investigating their effects on organisms responsive to their concentration. As an example of this field gaining traction in recent years, in [163] several biomarkers are reviewed as indicators of heavy metal-induced environmental perturbation.
Taking this into consideration microalgae could be considered interesting candidates as biosensors to study HM contamination. In depth analysis of metal-triggered morphological alterations in microalgal cell and organelles are traditionally performed with advanced techniques like Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM). SEM, for example was exploited in experiments where Skeletonema spp. cells underwent metal treatments, even if it was specifically used to study metal particles distribution in the media rather than microalgal morphological alterations [131]. TEM was employed to assess ultracellular alterations arising in three microalgal species, P. tricornutum, Tetraselmis sp. and D. tertiolecta, exposed to Cu concentrations close to the EC50 value each specie showed for the metal, uncovering an increased number and size of vacuoles in the latter two species due to the treatment; moreover, in P. tricornutum cell swelling and clumping was also observed [87]. SEM and TEM can be hindered by issues like cells clumping, possibly as a result of metal treatment and its sorption to cell wall [164,165]; even tackling this problem by diluting the sample is a partial solution, since this could mean spacing too much the cells out and thus ending up imaging a non-statistically significant number of objects. For example, TEM imaging of P. tricornutum diatoms exposed to Cu in [89] only includes 2-4 diatoms per image. Finally, electron microscopy techniques are often time-consuming, needing several steps for sample preparation, including fixation with the use of toxic agents and multiple dehydration steps, and expert personnel. If microalgae are to be exploited as bioindicators for the monitoring of heavy metal polluted sites advanced yet faster, non-invasive techniques are needed to image cell and organelles morphological alternations.
4. The Role of Microalgae in HM Removal
4.1. Microalgae for HM Removal: Current Technologies, Mechanisms and Advantages
HM-bioremediation employs microorganisms or plants to sequester and remove heavy metals from the environment, by adsorption, immobilization and/or internalization [65,166,167]. Among several organisms that have been investigated for their bioremediation potential, microalgae are gaining growing attention [167].
Most microalgae possess a rigid cell wall, mainly composed of cellulose enriched with polysaccharides. Specifically, it is usually structured in two layers: the external one has polysaccharides comprising, to various extent for different species, pectin, agar, alginate or algenan polymers; often these polysaccharides are sulfonated (fucoidans). The internal layer instead, contacting the phospholipidic plasma membrane, is built by a structural matrix of cellulose microfibrils engulfing hemicellulose, glycoproteins and pectin or fucans [168,169,170,171]. The carbohydrate composition often significatively varies between different species: for example, cell walls of microalgae in the Chlorophyta division usually possess cellulose or hemicellulose, while diatoms, a particularly large and diverse microalgal group in the Bacillariophyceae class exhibit peculiar siliceous encasing named frustules [172].
Cell wall components like carbohydrates, lipids and proteins can offer negatively charged groups acting as binding sites for positively charged metal cations, as they contain hydroxyl (-OH), carboxyl (-COOH), phosphate (-PO4), amine (-NH2) and sulfhydryl (-SH) groups: these can bind metal ions through electrostatic and covalent interactions or coordination bonds, with high affinity for oxygen and thiol groups [102,173,174]. This first, passive adsorption onto cell wall (termed “biosorption”) is a rapid process—with equilibrium being reached after ca. 30 minutes—and occurs both in living cells and in dead microalgal biomasses, as it is metabolism independent [104,175,176]. In some cases, interaction of living microalgae with metals also results in a second, slower stage of uptake from the media, termed “bioaccumulation” which actively sequesters them into the cell: from there metals diffuse into the cytoplasm and can be bound by dedicated molecules and sequestered into specific compartment or organelles [177,178].
Biosorption actually occurs by attracting heavy metals to cells surface through different mechanisms, including mainly ion exchange between protons/positively charged ions (e.g., lighter metals, like K+, Na+, Ca2+ and Mg2+) on the cell surface and metals in solution [179,180,181], but also physical adsorption due to electrostatic (stronger) attraction and Van der Waals (weaker) forces [179], and, finally, coordination bonds, where heavy metals can form covalent bonds with polarized functional groups like, for example, carboxyl groups contained in polysaccharides or amino groups of chitin and chitosan [107,174]. Metal complexation onto cell wall can also lead to its precipitation into less soluble forms [133]. Finally, some species can produce extracellular polymeric substances (EPS), mainly consisting of modified polysaccharides, and secrete them in the outer layer of cell wall for protection against stress, including adsorption of HMs thanks to charged hydrophobic groups: this, tough considered a biosorption mechanism as well, is a metabolic-dependent process [133,182,183,184].
A high fraction of the total removal of metal in solution by microalgae is hypothesized to be due to active metabolic uptake and bioaccumulation [185]. Unlike biosorption, this second stage of cell-mediated heavy metal sequestering is substantially slow and—most importantly—not an equilibrium process, as it is mediated by specific transporters or carrier molecules and thus actively regulated and directed by living cells [184,186,187]. Several metal transporters families have been characterized, some unique for microalgae and some shared with other organisms, responsible for HM transport across cell membrane into cytosol and, in some cases, into vacuoles, and are functionally divided into two groups [187,188]. Assimilative transporters drive metal towards the cytosol from extracellular or vacuolar spaces, and include natural resistance-associated macrophage proteins (NRAMP), exploiting proton gradient to transport divalent cations across the membrane, and passive Zn and Fe regulated transporters (ZRT and IRT), Zrt-Irt like proteins (ZIP), Fe and Cu transporters (FTR and CTR) and P-type ATPases [99,187,188]. It is worth noticing that some transporters require metal to be in a specific oxidation state, obtained through dedicated enzyme like reductases: for example Cu(II), the soluble cupric Cu form available in marine environments, needs to be reduced to Cu(I) cuprous ion, which is also more reactive, prior to transport [182,188]. In addition, microalgae possess distributive transporters in charge of decreasing HMs cytosolic concentration, via metal trafficking and efflux of metallic ions and metal-ligand complexes toward cells’ exterior or organelles as mechanisms to prevent excessive metal concentrations build up. Distributive transporters include cation diffusion facilitators (CDF) and P1B-type ATPases, relying on antiport of proton or potassium ions or on ATP hydrolysis respectively, FerroPortin (FPN), mithochondrial solute carriers (MSC), Ccc1/VIT1, a Ca2+ sensitive vacuolar Fe transporter, metal tolerance (MTP) and multidrug and toxic compound extrusion (MATE) proteins [99,187,188].
In addition to transporters regulation, metal concentrations inside the microalgal cells are tightly controlled by regulating their trafficking through metal chaperones and chelators, i.e., a wide spectrum of ligands with metal-binding properties (usually due to thiol groups). These can link and shuttle metals across various cellular compartments or, in some cases, even immobilize metal ions extracellularly so as to prevent their interaction with microalgal cells and uptake [99]. Among metal-binding poly-peptides produced by microalgae, we find metallothioneins (MCs) and phytochelatins (PCs) [189]. MCs are genetically-encoded small proteins rich in cysteine residues, possessing a partial negative charge and therefore able to form coordination bonds with metallic cations; even though the gene encoding for such proteins have been predicted only for a few microalgal genera, they are largely known in plants and ciliates, and likely to play an important role in microalgal binding, sequestering and detoxification of metal ions as well as reactive oxygen species (ROS) scavenging [189]. PCs on the other end, are biosynthetically produced polypeptides largely known to be involved in metal binding and transport to the vacuole in microalgae, plants and other organisms [189]. PCs are synthesized by phytochelatin synthase (PCS) using GSH, a thiol-containing tripeptide, as a substrate: GSH is normally employed as the main redox-buffer in oxidative stress mitigation, but is also capable of HM binding and scavenging through its cysteine residues. When multiple GSH units are joined to form PCs, these residues can form disulfide bonds and thus chelate metal ions trough the thiol groups present in the side chains, forming HMs-PCs complexes that are stable in the cytoplasm but can be subsequently cleaved by lower pH in cellular vacuoles [99,189]. In general, metal chelators with various affinities can be produced both to handle their intracellular trafficking and to limit damages due to HM excess. As a matter of fact, some microalgae have been observed to release various kinds of ligands in the extracellular space in response to HM stress in addition, in some cases, to production of exudates with high lipid and polysaccharide content and surfactant properties [190]. Ultimately, HM-chelation and complexation can either aid its transport inside the cell, its efflux or prevent its entrance in the cell.
All these inter-connected mechanisms employed by microalgae to sequester HMs and detoxify their stress are schematically summarized in Figure 3.
Microalgae are generally characterized by fast growth rates (compared to terrestrial plants and macroalgae), and can be easily maintained in open and enclosed systems, ensuring high year-round biomass production and productivities. Living cultures only require a minimum amount of nutrients, which can be also derived from wastewater to optimize process costs [191], while dead biomasses do not require neither nutrients, nor particular conditions of oxygenation/carbon dioxide sparging, and can be re-used several times after some HM desorption steps with washing agents.
The use of some freshwater microalgal species in phycoremediation (i.e., bioremediation employing algae) has been currently implemented for a few devices exploiting non-living biomass to remove pollutants, particularly heavy metals, from waste water: these biosorbents are based on natural or synthetic matrices to embed either individual species or consortia, effectively eliminating the necessity of biomass harvesting (which is the more expensive and tedious step) while simultaneously providing efficient HM sorption. An example is the AlgaSORB system, which consists of a silica gel polymer decorated with immobilized non-living Chlorella vulgaris cells, employed for effective removal of metallic ions from aqueous solutions [192,193]. On the other end, a consortium of different organisms, including algae, yeasts, bacteria and sea weeds on a high density polysulfone matrix is exploited by the BIO-FIX sorption system [193,194]. Finally, B.V. SORBEX company developed a sorption device based on a variety of biomaterials, including Chlorella vulgaris and five macroalgal species, to remove different metal ions across a wide pH range [193,195].
Of course, as stated above, the primary benefit of the use of dead microalgal biomasses as biosorbents is their potential for multiple reusage, but it should be noted that, as detailed in previous sections, these cells cannot internalize HMs after surface adsorption, with the possible risk of releasing the contaminant back into the environment. Furthermore, while non-viable microalgae systems can rapidly remove HMs through biosorption their overall sorption capacity, especially for lower metal concentrations, can be slightly lower compared to live microalgae. The latter, in addition to adsorption, can exploit a broader range of removal mechanisms, including active transport; they may thus sequester metals more stably within cell compartments and, consequently, be employed with greater success in long-term sequestration strategies [196]. Wang and Chen [197] predict that the future of biosorption technologies will likely integrate living microalgal cells in hybrid systems to enhance pollutant removal. Live biomass could result more versatile in removing HMs through mechanisms like precipitation, adsorption, and bioaccumulation. However, one challenge is that viable microalgae can be sensitive to high concentrations of heavy metals, which can affect their health and efficiency [198]: it is therefore crucial to identify metal-tolerant strains in order to optimize bioremediation efforts. Finally, in light of possible in situ applications an added concern should be taken into account: avoiding the introduction of non-native species in the aquatic environment where remediation of the pollutant is carried out is of capital importance. This highlights the relevance of researching local microalgal strains that are naturally adapted to specific environments, making them better suited for bioremediation in those areas.
4.2. The Influence of Abiotic Factors on HM Removal
Abiotic factors like pH, time of exposure, temperature, metal initial concentration and characteristics (ionic size, atomic weight or reduction potential), presence of organic substances or inorganic ligands in solution among others influence HM removal [178].
The main players in biosorption processes efficiency are pH and contact time. Notably, increasing pH means higher adsorption, as protonation level is critical to shift the equilibrium either toward metal complexation or release and determines which functional groups will be available for binding: it has been suggested that carboxyl groups are the predominant available site at pH range 2-5, phosphate groups follow between pH values 5-9 and finally in the pH interval 9-12 groups available also comprise hydroxyl (or amine) groups [173]. Essentially, at low pH the majority of functional groups are blocked by association with hydronium ions (H3O+), while at higher pH values more sites are available for binding [102,185,186]. In addition to cell binding sites availability, pH also affects metal speciation: most metals tend to precipitate in more basic solutions, impairing HM removal efficiency, meaning that this parameters needs to be finely tuned in bioremediation applications [107,178].
Moreover, temperature (even though there’s no real consensus on its influence, probably more related to the kinetics than the capacity) plays a key role, being connected to the solubility of metal ions in aqueous media, their stability in addition to stability of ligands and of the formed complexes, as most chemical reactions are sensitive to temperature ranges. As a matter of fact, higher temperatures lead to greater metal ion solubility, unfavouring the biosorption process [102,185,186,199].
Ionic strength, a measure of the concentration of ions in solution, is also to be considered when using microalgal biomasses for remediation purposes, as high concentration of ions, such as monovalent cations like Na+ and K+, decreases sites available for metal binding via competition for the functional groups, resulting in lower efficiency [178].
Additionally, metal concentration and speciation (whether metals are free ions or complexed with organic/inorganic ligands) ultimately reflect the concentration of free metal ions, which are the species able to bind microalgal cells; the presence of multiple metals also influences co-ions adsorption through antagonistic or synergistic processes, even though non-interactive or additive outcomes are also possible [133,180,199]. Indeed several amino acids as well as humic acids, fulvic acid, EDTA, NTA and various kinds of organic matter cannot interact with HMs forming complexes, thus reducing their bioavailability to microalgae [178,200,201].
As an example, influence of pH, time of exposure, metal concentration and initial cell density on hexavalent Cr biosorption in aqueous solution by Dunaniella salina was investigated by Kaushic and co-workers [202]: they found that highest biosorption efficiency (66.4%) was attained at pH 8.6 and 10% v/v inoculum size within 120 hours. Schiariti et al. reported that, in C. kessleri and S. acutus cultures exposed to Cu, recovered growth rates and reduction in bleaching and lipid peroxidation was observed following acetate addition, thus effectively reducing concentration of free Cu ion [201].
4.3. The Influence of Biotic Factors on HM Removal
In addition to abiotic factors, HM removal is influenced by biotic factors including organism species, cell concentration, division rate and nutrients uptake and size and surface area and, finally, sensitivity [178].
Other crucial factors to explain HM tolerance and adsorption in microalgae are the specific remediation skills of the microalgal species used as biosorbent, and the concentration of the biomass employed—which may be also considered regardless of cell viability in biosorption applications, as more biomass offers an higher surface area and thus more binding sites for the reaction [178]. Indeed, high cell density and, in general, biomass concentration are reportedly associated with higher metal removal, possibly due simply to a larger pool of available binding sites [185]. However, biomass concentration is a parameter that needs to be carefully tuned for bioremediation applications—especially in the case of live biomass—as very high initial cell density could also lead to formation of cell aggregates or screen effect, hindering HM removal by blocking binding sites [178,203]. Microalgal division rates are also related to the size of the species considered, as smaller microalgae exhibit higher specific growth rates due to higher rate of photosynthesis and more efficient transport of nutrients per unit of biomass: the smaller the size, the more efficient is N and P uptake [178,204]. It has also been suggested that sensitivity to metal might be higher for smaller cells, showing an higher surface area to volume ratio and accumulation rates, with respect to larger species [205,206].
Levy and co-workers investigated cell size and walls ultrastructure influence on HM sensitivity and removal from the media: in their study they correlate cell surface area with its tolerance and Cu content for 11 different microalgal species, observing that the three more sensitive species (Minutocellus polymorphus, Micromonas pusilla and Isochrysis sp.) were among the smallest considered; however the relationship was not strictly linear—tolerance of the smallest species of all (M. pusilla) was twice than the one for M. polymorphus, while the largest species tested (Heterocapsa niei) was one of the most sensitive species amongst the ones under exam. In addition, Cu partitioning between cells and media was found to be correlated to cell surface area rather than to sensitivity to the metal—also in this case though, no clearly linear relationship could be elucidated [114]. Indeed, metal sensitivity varies broadly among different species influencing their uptake and removal capacity, even though it is difficult to pinpoint an exact quantitative correlation between the two. Levy et al. also report different IC50 values for three microalgae species exposed to Cu, specifically 8 μg/L for P. tricornutum, 47 μg/L for Tetraselmis sp. and 530 μg/L for D. tertiolecta, while also highlighting that at these specific value for each species Tetraselmis sp. exhibited the highest intracellular Cu content (ca. 0.197 pg Cu/ cell). Notably tough, even if at IC50 doses D. tertiolecta internalized more Cu than P. tricornutum if exposed at the same Cu concentration of 50 μg/L, the former showed slower internalization rate and lower internal metal concentration than the latter, suggesting that these two factor are indeed not the only ones concurring in determination of species sensitivity to a certain metal, and the contribution of microalgal detoxification mechanism should also be taken into account [87].
5. Conclusions
The present paper reports the state of the art of the current knowledge of HM-driven stress in microalgae, and of microalgae-based bioremediation skills for HM removal.
Indeed, we described in detail two pivotal functions of microalgae exposed to HMs: i) their role as sentinel species, showing different signs of stress, such as ROS generation, growth rates and chlorophyll content reduction, cell and organelles impairments, and increase of HM-binding protein synthesis; ii) their contribution to HM removal by mechanisms of passive adsorption onto the cell wall and- in metabolically active cells—through HM internal sequestration in specific comparts. Conventional methods currently available for HM removal involve the use of chemicals, resins or electrochemical technologies, with the main disadvantages of high costs and the possible production of toxic by-products. To overcome these drawbacks, and considering the increased anthropogenic emissions of HMs in aquatic environments, uncovering innovative, ecosustainable technologies based on the employment of aquatic organisms to remove HMs is desirable. Although current algae-based technologies are mostly based on dead biomasses, which main advantage do not require nutrient input and can be also re-used after HM desorption cycles, current knowledge and further studies on the role of living microalgae could assure a more stable and quantitative removal, combining both adsorption and compartmentalization mechanisms.
Author Contributions
E.C.: original draft and figure preparation; I.O and G.R.: critical review; AS: conceptualization, supervision, critical review and editing.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Duffus, J.H. “Heavy metals” a meaningless term? (IUPAC Technical Report). Pure and Applied Chemistry 2002, 74, 793–807. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E. What are heavy metals? Long-standing controversy over the scientific use of the term ‘heavy metals’— proposal of a comprehensive definition. Toxicological & Environmental Chemistry 2018, 100, 6–19. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, N.; Li, Y.; Ren, B.; Ding, X.; Bian, H.; Yao, X. Total concentrations and sources of heavy metal pollution in global river and lake water bodies from 1972 to 2017. Global Ecology and Conservation 2020, 22, e00925. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: a review. Environmental Chemistry Letters 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Aziz, K.H.H.; Mustafa, F.S.; Omer, K.M.; Hama, S.; Hamarawf, R.F.; Rahman, K.O. Heavy metal pollution in the aquatic environment: efficient and low-cost removal approaches to eliminate their toxicity: a review. RSC advances 2023, 13, 17595–17610. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. Journal of Chemistry 2019, 2019, 6730305. [Google Scholar] [CrossRef]
- Zamora-Ledezma, C.; Negrete-Bolagay, D.; Figueroa, F.; Zamora-Ledezma, E.; Ni, M.; Alexis, F.; Guerrero, V.H. Heavy metal water pollution: A fresh look about hazards, novel and conventional remediation methods. Environmental Technology & Innovation 2021, 22, 101504. [Google Scholar] [CrossRef]
- Hama Aziz, K.H.; Mustafa, F.S.; Omer, K.M.; Hama, S.; Hamarawf, R.F.; Rahman, K.O. Heavy metal pollution in the aquatic environment: efficient and low-cost removal approaches to eliminate their toxicity: a review. RSC advances 2023, 13, 17595–17610. [Google Scholar] [CrossRef]
- Vélez-Pérez, L.S.; Ramirez-Nava, J.; Hernández-Flores, G.; Talavera-Mendoza, O.; Escamilla-Alvarado, C.; Poggi-Varaldo, H.M.; Solorza-Feria, O.; López-Díaz, J.A. Industrial acid mine drainage and municipal wastewater co-treatment by dual-chamber microbial fuel cells. International Journal of Hydrogen Energy 2020, 45, 13757–13766. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, Z.-g.; Zeng, G.-m.; Jiang, M.; Yang, Z.-z.; Cui, F.; Zhu, M.-y.; Shen, L.-q.; Hu, L. Effects of sediment geochemical properties on heavy metal bioavailability. Environment International 2014, 73, 270–281. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip Toxicol 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Muhammad, S.; Khan, B.; Khan, H. Investigating the levels of selected heavy metals in surface water of Shah Alam River (A tributary of River Kabul, Khyber Pakhtunkhwa). Journal of Himalayan Earth Sciences 2011, 44, 71–79. [Google Scholar]
- Asiminicesei, D.-M.; Vasilachi, I.C.; Gavrilescu, M. Heavy Metal Contamination of Medicinal Plants and Potential Implications on Human Health. Revista de Chimie 2020, 71, 16–36. [Google Scholar] [CrossRef]
- Verde, R.; Vigliotti, M.; Prevedello, L.; Sprovieri, M.; Ruberti, D. An integrated approach to environmental quality assessment in a coastal setting in Campania (Southern Italy). Environmental Earth Sciences 2013, 70, 407–424. [Google Scholar] [CrossRef]
- Lu, R.; Rong, S.; Wu, J.; Yue, W.; Li, Q. Pollution Assessment and SSD-Based Ecological Assessment of Heavy Metals in Multimedia in the Coast of Southeast China. International Journal of Environmental Research and Public Health 2022, 19, 16022. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Cocozza di Montanara, A.; Corinaldesi, C.; Dell’Anno, A.; Illuminati, S.; Willis, T.J.; Gambi, C. Bioaccumulation and biomagnification of heavy metals in marine micro-predators. Commun Biol 2023, 6, 1206. [Google Scholar] [CrossRef]
- Ahlf, W.; Drost, W.; Heise, S. Incorporation of metal bioavailability into regulatory frameworks—metal exposure in water and sediment. Journal of Soils and Sediments 2009, 9, 411–419. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.; Rahman, H.; Ashekuzzaman, S.; Islam, M.M.; Chowdhury, S.R.; Hossain, M.S. Heavy metals accumulation in coastal sediments. Environmental remediation technologies for metal-contaminated soils.
- Saddik, M.; Fadili, A.; Makan, A. Assessment of heavy metal contamination in surface sediments along the Mediterranean coast of Morocco. Environmental Monitoring and Assessment 2019, 191. [Google Scholar] [CrossRef]
- Ozmen, S. Ecological assesment of Akkuyu nuclear power plant site marine sediments in terms of radionuclide and metal accumulation. Journal of Radioanalytical and Nuclear Chemistry 2020, 325, 133–145. [Google Scholar] [CrossRef]
- Abdallah, M.; Badr-ElDin, A. Ecological risk assessment of surficial sediment by heavy metals from a submerged archaeology harbor, South Mediterranean Sea, Egypt. Acta Geochimica 2020, 39, 226–235. [Google Scholar] [CrossRef]
- Ben Amor, R.; Yahyaoui, A.; Abidi, M.; Chouba, L.; Gueddari, M. Bioavailability and Assessment of Metal Contamination in Surface Sediments of Rades-Hamam Lif Coast, around Meliane River (Gulf of Tunis, Tunisia, Mediterranean Sea). Journal of Chemistry 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Gambi, C.; Dell’Anno, A.; Corinaldesi, C.; Lo Martire, M.; Musco, L.; Da Ros, Z.; Armiento, G.; Danovaro, R. Impact of historical contamination on meiofaunal assemblages: The case study of the Bagnoli-Coroglio Bay (southern Tyrrhenian Sea). Marine Environmental Research 2020, 104907. [Google Scholar] [CrossRef] [PubMed]
- Surricchio, G.; Pompilio, L.; Novelli, A.A.; Scamosci, E.; Marinangeli, L.; Tonucci, L.; d’Alessandro, N.; Tangari, A.C. Evaluation of heavy metals background in the Adriatic Sea sediments of Abruzzo region, Italy. Science of the Total Environment 2019, 684, 445–457. [Google Scholar] [CrossRef]
- Pitacco, V.; Mistri, M.; Granata, T.; Moruzzi, L.; Meloni, M.L.; Massara, F.; Sfriso, A.; Sfriso, A.A.; Munari, C. Sediment contamination by heavy metals and PAH in the Piombino channel (Tyrrhenian Sea). Water 2021, 13, 1487. [Google Scholar] [CrossRef]
- Piazzolla, D.; Scanu, S.; Frattarelli, F.M.; Mancini, E.; Tiralongo, F.; Brundo, M.V.; Tibullo, D.; Pecoraro, R.; Copat, C.; Ferrante, M. Trace-metal enrichment and pollution in coastal sediments in the Northern Tyrrhenian Sea, Italy. Archives of environmental contamination and toxicology 2015, 69, 470–481. [Google Scholar] [CrossRef]
- Sprovieri, M.; Feo, M.L.; Prevedello, L.; Manta, D.S.; Sammartino, S.; Tamburrino, S.; Marsella, E. Heavy metals, polycyclic aromatic hydrocarbons and polychlorinated biphenyls in surface sediments of the Naples harbour (southern Italy). Chemosphere 2007, 67, 998–1009. [Google Scholar] [CrossRef]
- De Pippo, T.; Donadio, C.; Guida, M.; Petrosino, C. The Case of Sarno River (Southern Italy). Effects of geomorphology on the environmental impacts (8 pp). Environmental Science and Pollution Research—International 2006, 13, 184–191. [Google Scholar] [CrossRef]
- Montuori, P.; Lama, P.; Aurino, S.; Naviglio, D.; Triassi, M. Metals loads into the Mediterranean Sea: estimate of Sarno River inputs and ecological risk. Ecotoxicology 2013, 22, 295–307. [Google Scholar] [CrossRef]
- Breuer, E.; Stevenson, A.G.; Howe, J.A.; Carroll, J.; Shimmield, G.B. Drill cutting accumulations in the Northern and Central North Sea: a review of environmental interactions and chemical fate. Marine Pollution Bulletin 2004, 48, 12–25. [Google Scholar] [CrossRef]
- Fichet, D.; Radenac, G.; Miramand, P. Experimental studies of impacts of harbour sediments resuspension to marine invertebrates larvae: Bioavailability of Cd, Cu, Pb and Zn and toxicity. Marine Pollution Bulletin 1998, 36, 509–518. [Google Scholar] [CrossRef]
- Liu, X.; Huang, W. Modeling sediment resuspension and transport induced by storm wind in Apalachicola Bay, USA. Environmental Modelling & Software 2009, 24, 1302–1313. [Google Scholar]
- Rial, D.; Beiras, R. Prospective ecological risk assessment of sediment resuspension in an estuary. Journal of Environmental Monitoring 2012, 14, 2137–2144. [Google Scholar] [CrossRef]
- Webster, T.; Lemckert, C. Sediment resuspension within a microtidal estuary/embayment and the implication to channel management. Journal of Coastal Research 2002, 753–759. [Google Scholar] [CrossRef]
- Long, E.R.; Macdonald, D.D.; Smith, S.L.; Calder, F.D. Incidence of adverse biological effects within ranges of chemical concentrations in marine and estuarine sediments. Environmental Management 1995, 19, 81–97. [Google Scholar] [CrossRef]
- De Mora, S.; Fowler, S.W.; Wyse, E.; Azemard, S. Distribution of heavy metals in marine bivalves, fish and coastal sediments in the Gulf and Gulf of Oman. Mar Pollut Bull 2004, 49, 410–424. [Google Scholar] [CrossRef]
- Mejdoub, Z.; Zaid, Y.; Hmimid, F.; Kabine, M. Assessment of metals bioaccumulation and bioavailability in mussels Mytilus galloprovincialis exposed to outfalls pollution in coastal areas of Casablanca. J Trace Elem Med Biol 2018, 48, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Joksimović, D.; Castelli, A.; Perošević, A.; Djurović, D.; Stanković, S. Determination of trace metals in Mytilus galloprovincialis along the Boka Kotorska Bay, Montenegrin coast. Journal of Trace Elements in Medicine and Biology 2018, 50, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Capelli, R.; Contardi, V.; Fassone, B.; Zanicchi, G. Heavy metals in mussels (Mytilus galloprovincialis) from the gulf of la spezia and from the promontory of Portofino, Italy. Marine Chemistry 1978, 6, 179–185. [Google Scholar] [CrossRef]
- Türk Çulha, S.; Çulha, M.; Karayücel, İ.; Çelik, M.Y.; Işler, Y. Heavy metals in Mytilus galloprovincialis, suspended particulate matter and sediment from offshore submerged longline system, Black Sea. International Journal of Environmental Science and Technology 2017, 14, 385–396. [Google Scholar] [CrossRef]
- Ndiaye, B.; Ndiaye, M.; Cid, B.P.; Diop, A.; Diagne, I.; Cissé, D.; Dione, C.T.; Hanne, M. Trace Metals in Mussels Mytilus galloprovincialis from Dakar Coast (Senegal). American Journal of Analytical Chemistry 2020, 11, 137–145. [Google Scholar] [CrossRef]
- Appenroth, K.J. Definition of “Heavy Metals” and Their Role in Biological Systems. 2010.
- Mehri, A. Trace Elements in Human Nutrition (II)—An Update. Int J Prev Med 2020, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A. Trace elements in soils and plants; CRC press: 2000.
- Kalaivanan, D.; Ganeshamurthy, A. Mechanisms of Heavy Metal Toxicity in Plants. 2015; pp. 85-102.
- Küpper, H.; Andresen, E. Mechanisms of metal toxicity in plants. Metallomics 2016, 8, 269–285. [Google Scholar] [CrossRef]
- Kozlowski, H.; Kolkowska, P.; Watly, J.; Krzywoszynska, K.; Potocki, S. General aspects of metal toxicity. Curr Med Chem 2014, 21, 3721–3740. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, Y.J.; Seo, Y.R. An Overview of Carcinogenic Heavy Metal: Molecular Toxicity Mechanism and Prevention. J Cancer Prev 2015, 20, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A review of toxicity and mechanisms of individual and mixtures of heavy metals in the environment. Environ Sci Pollut Res Int 2016, 23, 8244–8259. [Google Scholar] [CrossRef]
- Sharma, M.; Kant, R.; Sharma, A.; Sharma, A. Exploring the impact of heavy metals toxicity in the aquatic ecosystem. International Journal of Energy and Water Resources 2024. [Google Scholar] [CrossRef]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress part I: mechanisms involved in metal-induced oxidative damage. Curr Top Med Chem 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: A review. Journal of Environmental Management 2011, 92, 407–418. [Google Scholar] [CrossRef]
- Pang, F.M.; Teng, S.P.; Teng, T.T.; Omar, A.M. Heavy metals removal by hydroxide precipitation and coagulation-flocculation methods from aqueous solutions. Water Quality Research Journal 2009, 44, 174–182. [Google Scholar] [CrossRef]
- Lewis, A. Precipitation of Heavy Metals. In Sustainable Heavy Metal Remediation: Volume 1: Principles and Processes, Rene, E.R., Sahinkaya, E., Lewis, A., Lens, P.N.L., Eds.; Springer International Publishing: Cham, 2017; pp. 101–120. [Google Scholar]
- Bashir, A.; Malik, L.A.; Ahad, S.; Manzoor, T.; Bhat, M.A.; Dar, G.; Pandith, A.H. Removal of heavy metal ions from aqueous system by ion-exchange and biosorption methods. Environmental Chemistry Letters 2019, 17, 729–754. [Google Scholar] [CrossRef]
- Buonomenna, M.G.; Mousavi, S.M.; Hashemi, S.A.; Lai, C.W. Water Cleaning Adsorptive Membranes for Efficient Removal of Heavy Metals and Metalloids. Water 2022, 14, 2718. [Google Scholar] [CrossRef]
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: a review. Journal of environmental management 2011, 92, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Maarof, H.I.; Daud, W.M.A.W.; Aroua, M.K. Recent trends in removal and recovery of heavy metals from wastewater by electrochemical technologies. Reviews in Chemical Engineering 2017, 33, 359–386. [Google Scholar] [CrossRef]
- Ezziat, L.; Elabed, A.; Ibnsouda, S.; El Abed, S. Challenges of Microbial Fuel Cell Architecture on Heavy Metal Recovery and Removal From Wastewater. Frontiers in Energy Research 2019, 7. [Google Scholar] [CrossRef]
- Wang, S. A Comparative study of Fenton and Fenton-like reaction kinetics in decolourisation of wastewater. Dyes and Pigments 2008, 76, 714–720. [Google Scholar] [CrossRef]
- Zhao, M.; Xu, Y.; Zhang, C.; Rong, H.; Zeng, G. New trends in removing heavy metals from wastewater. Applied microbiology and biotechnology 2016, 100, 6509–6518. [Google Scholar] [CrossRef]
- Li, Y.; Liu, F.; Xia, B.; Du, Q.; Zhang, P.; Wang, D.; Wang, Z.; Xia, Y. Removal of copper from aqueous solution by carbon nanotube/calcium alginate composites. Journal of Hazardous Materials 2010, 177, 876–880. [Google Scholar] [CrossRef]
- Wang, B.; Wan, Y.; Zheng, Y.; Lee, X.; Liu, T.; Yu, Z.; Huang, J.; Ok, Y.S.; Chen, J.; Gao, B. Alginate-based composites for environmental applications: a critical review. Critical reviews in environmental science and technology 2019, 49, 318–356. [Google Scholar] [CrossRef]
- Naseer, A.; Jamshaid, A.; Hamid, A.; Muhammad, N.; Ghauri, M.; Iqbal, J.; Rafiq, S.; Khuram, S.; Shah, N.S. Lignin and lignin based materials for the removal of heavy metals from waste water-an overview. Zeitschrift für Physikalische Chemie 2019, 233, 315–345. [Google Scholar] [CrossRef]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques–classification based on site of application: principles, advantages, limitations and prospects. World Journal of Microbiology and Biotechnology 2016, 32, 180. [Google Scholar] [CrossRef]
- Benoiston, A.S.; Ibarbalz, F.M.; Bittner, L.; Guidi, L.; Jahn, O.; Dutkiewicz, S.; Bowler, C. The evolution of diatoms and their biogeochemical functions. Philos Trans R Soc Lond B Biol Sci 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the potential of microalgae for new biotechnology applications and beyond: A review. Renewable and Sustainable Energy Reviews 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Vargas, C.A.; Martínez, R.A.; Escribano, R.; Lagos, N.A. Seasonal relative influence of food quantity, quality, and feeding behaviour on zooplankton growth regulation in coastal food webs. Journal of the Marine Biological Association of the United Kingdom 2010, 90, 1189–1201. [Google Scholar] [CrossRef]
- Lauringson, V.; Kotta, J.; Orav-Kotta, H.; Kaljurand, K. Diet of mussels Mytilus trossulus and Dreissena polymorpha in a brackish nontidal environment. Marine Ecology 2013, 35. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Algal Physiology and Large-Scale Outdoor Cultures of Microalgae. In The Physiology of Microalgae. In The Physiology of Microalgae, Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Cham, 2016; pp. 601–652. [Google Scholar]
- Metting, F.B. Biodiversity and application of microalgae. Journal of Industrial Microbiology 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Prinsen, P.; Raheem, A.; Luque, R.; Zhao, M. Microalgae cultivation and metabolites production: a comprehensive review. Biofuels, Bioproducts and Biorefining 2018, 12, 304–324. [Google Scholar] [CrossRef]
- Katiyar, R.; Gurjar, B.R.; Biswas, S.; Pruthi, V.; Kumar, N.; Kumar, P. Microalgae: An emerging source of energy based bio-products and a solution for environmental issues. Renewable and Sustainable Energy Reviews 2017, 72, 1083–1093. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. Journal of Bioscience and Bioengineering 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- SureshKumar, P.; Thomas, J.; Poornima, V. Structural insights on bioremediation of polycyclic aromatic hydrocarbons using microalgae: a modelling-based computational study. Environmental Monitoring and Assessment 2018, 190, 92. [Google Scholar] [CrossRef]
- Nayak, J.K.; Ghosh, U.K. Post treatment of microalgae treated pharmaceutical wastewater in photosynthetic microbial fuel cell (PMFC) and biodiesel production. Biomass and Bioenergy 2019, 131, 105415. [Google Scholar] [CrossRef]
- Singh, J.; Dhar, D.W. Overview of Carbon Capture Technology: Microalgal Biorefinery Concept and State-of-the-Art. Frontiers in Marine Science 2019, 6. [Google Scholar] [CrossRef]
- Lü, J.; Sheahan, C.; Fu, P. Metabolic engineering of algae for fourth generation biofuels production. Energy & Environmental Science 2011, 4, 2451–2466. [Google Scholar] [CrossRef]
- Pessôa, L.C.; Deamici, K.M.; Pontes, L.A.M.; Druzian, J.I.; Assis, D.d.J. Technological prospection of microalgae-based biorefinery approach for effluent treatment. Algal Research 2021, 60, 102504. [Google Scholar] [CrossRef]
- Abomohra, A.; Hanelt, D. Recent Advances in Micro-/Nanoplastic (MNPs) Removal by Microalgae and Possible Integrated Routes of Energy Recovery. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Barra, L.; Sardo, A.; Moros, M.; Smerilli, A.; Chiaiese, P.; Percopo, I.; Cavalletti, E.; Castro-Hinojosa, C.; Balzano, S. Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation. Microorganisms 2022, 10, 2445. [Google Scholar] [CrossRef]
- Cheng, Y.R.; Wang, H.Y. Highly effective removal of microplastics by microalgae <i>Scenedesmus abundans</i>. Chemical Engineering Journal 2022, 435. [Google Scholar] [CrossRef]
- Goh, P.S.; Lau, W.J.; Ismail, A.F.; Samawati, Z.; Liang, Y.Y.; Kanakaraju, D. Microalgae-Enabled Wastewater Treatment: A Sustainable Strategy for Bioremediation of Pesticides. Water 2023, 15. [Google Scholar] [CrossRef]
- Matamoros, V.; Rodríguez, Y. Batch vs continuous-feeding operational mode for the removal of pesticides from agricultural run-off by microalgae systems: A laboratory scale study. Journal of Hazardous Materials 2016, 309, 126–132. [Google Scholar] [CrossRef]
- Webster, L.J.; Villa-Gomez, D.; Brown, R.; Clarke, W.; Schenk, P.M. A synthetic biology approach for the treatment of pollutants with microalgae. Frontiers in Bioengineering and Biotechnology 2024, 12. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghachtouli, N.; El Baouchi, A.; El Arroussi, H. Heavy metals phycoremediation using tolerant green microalgae: Enzymatic and non-enzymatic antioxidant systems for the management of oxidative stress. Journal of Environmental Chemical Engineering 2020, 8, 104460. [Google Scholar] [CrossRef]
- Levy, J.L.; Angel, B.M.; Stauber, J.L.; Poon, W.L.; Simpson, S.L.; Cheng, S.H.; Jolley, D.F. Uptake and internalisation of copper by three marine microalgae: Comparison of copper-sensitive and copper-tolerant species. Aquatic Toxicology 2008, 89, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The role of biomarkers in the assessment of aquatic ecosystem health. Integr Environ Assess Manag 2014, 10, 327–341. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, B. Toxic effects of fluoranthene and copper on marine diatom Phaeodactylum tricornutum. Journal of environmental sciences (China) 2008, 20, 1363–1372. [Google Scholar] [CrossRef]
- Shivaji, S.; VL Dronamaraju, S. Scenedesnus rotundus isolated from the petroleum effluent employs alternate mechanisms of tolerance to elevated levels of cadmium and zinc. Scientific Reports 2019, 9, 8485. [Google Scholar] [CrossRef] [PubMed]
- Manimaran, K.; Karthikeyan, P.; Ashokkumar, S.; Ashok Prabu, V.; Sampathkumar, P. Effect of copper on growth and enzyme activities of marine diatom, Odontella mobiliensis. Bull Environ Contam Toxicol 2012, 88, 30–37. [Google Scholar] [CrossRef]
- Sendra, M.; Yeste, M.P.; Gatica, J.M.; Moreno-Garrido, I.; Blasco, J. Direct and indirect effects of silver nanoparticles on freshwater and marine microalgae (Chlamydomonas reinhardtii and Phaeodactylum tricornutum). Chemosphere 2017, 179, 279–289. [Google Scholar] [CrossRef]
- Pillai, S.; Behra, R.; Nestler, H.; Suter, M.J.-F.; Sigg, L.; Schirmer, K. Linking toxicity and adaptive responses across the transcriptome, proteome, and phenotype of <i>Chlamydomonas reinhardtii</i> exposed to silver. Proceedings of the National Academy of Sciences 2014, 111, 3490–3495. [Google Scholar] [CrossRef]
- Dewez, D.; Oukarroum, A. Silver nanoparticles toxicity effect on photosystem II photochemistry of the green alga Chlamydomonas reinhardtii treated in light and dark conditions. Toxicological & Environmental Chemistry 2012, 94, 1536–1546. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bras, S.; Perreault, F.; Popovic, R. Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Ecotoxicology and Environmental Safety 2012, 78, 80–85. [Google Scholar] [CrossRef]
- Sabatini, S.E.; Juarez, A.B.; Eppis, M.R.; Bianchi, L.; Luquet, C.M.; de Molina, M.d.C.R. Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicology and environmental safety 2009, 72, 1200–1206. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Pinto, E.; Latorre, L.; Bechara, E.J.H.; Colepicolo, P. Antioxidant modulation in response to metal-induced oxidative stress in algal chloroplasts. Archives of Environmental Contamination and Toxicology 2001, 40, 18–24. [Google Scholar] [PubMed]
- Strejckova, A.; Dvorak, M.; Klejdus, B.; Krystofova, O.; Hedbavny, J.; Adam, V.; Huska, D. The strong reaction of simple phenolic acids during oxidative stress caused by nickel, cadmium and copper in the microalga Scenedesmus quadricauda. New biotechnology 2019, 48, 66–75. [Google Scholar] [CrossRef]
- Tripathi, S.; Poluri, K.M. Heavy metal detoxification mechanisms by microalgae: Insights from transcriptomics analysis. Environmental Pollution 2021, 285, 117443. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, A.; Wei, Y.Y.; Meng, Q.; Zheng, Q.; Yang, Z.M. Mercury-induced oxidative stress and impact on antioxidant enzymes in Chlamydomonas reinhardtii. Ecotoxicology 2010, 19, 1285–1293. [Google Scholar] [CrossRef]
- Yang, J.; Cao, J.; Xing, G.; Yuan, H. Lipid production combined with biosorption and bioaccumulation of cadmium, copper, manganese and zinc by oleaginous microalgae Chlorella minutissima UTEX2341. Bioresource technology 2015, 175, 537–544. [Google Scholar] [CrossRef]
- Spain, O.; Plöhn, M.; Funk, C. The cell wall of green microalgae and its role in heavy metal removal. Physiologia Plantarum 2021, 173, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, V.; Alavi, S.A.; Zilouei, H. Mechanisms of heavy metal removal using microorganisms as biosorbent. Water Science and Technology 2014, 69, 1775–1787. [Google Scholar] [CrossRef] [PubMed]
- Pavithra, K.G.; Kumar, P.S.; Jaikumar, V.; Vardhan, K.H.; SundarRajan, P. Microalgae for biofuel production and removal of heavy metals: a review. Environmental Chemistry Letters 2020, 18, 1905–1923. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- García-García, J.D.; Sánchez-Thomas, R.; Moreno-Sánchez, R. Bio-recovery of non-essential heavy metals by intra- and extracellular mechanisms in free-living microorganisms. Biotechnology Advances 2016, 34, 859–873. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.L.; Malcata, F.X. Metal uptake by microalgae: Underlying mechanisms and practical applications. Biotechnology Progress 2012, 28, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Sigaud-Kutner, T.; Zajac, M.A.; Okamoto, O.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in algae. Journal of Phycology 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghatchouli, N.; Arroussi, H. Overview of the management of heavy metals toxicity by microalgae. Journal of Applied Phycology 2022, 34, 475–488. [Google Scholar] [CrossRef]
- Ugya, A.Y.; Imam, T.S.; Li, A.; Ma, J.; Hua, X. Antioxidant response mechanism of freshwater microalgae species to reactive oxygen species production: a mini review. Chemistry and Ecology 2020, 36, 174–193. [Google Scholar] [CrossRef]
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, W.; Jin, M.; Zhang, L.; Qin, L.; Geng, W. Responses and tolerance mechanisms of microalgae to heavy metal stress: A review. Mar Environ Res 2023, 183, 105805. [Google Scholar] [CrossRef]
- Arora, N.; Gulati, K.; Patel, A.; Pruthi, P.A.; Poluri, K.M.; Pruthi, V. A hybrid approach integrating arsenic detoxification with biodiesel production using oleaginous microalgae. Algal Research 2017, 24, 29–39. [Google Scholar] [CrossRef]
- Levy, J.L.; Stauber, J.L.; Jolley, D.F. Sensitivity of marine microalgae to copper: the effect of biotic factors on copper adsorption and toxicity. Sci Total Environ 2007, 387, 141–154. [Google Scholar] [CrossRef]
- Díaz, S.; De Francisco, P.; Olsson, S.; Aguilera, Á.; González-Toril, E.; Martín-González, A. Toxicity, Physiological, and Ultrastructural Effects of Arsenic and Cadmium on the Extremophilic Microalga Chlamydomonas acidophila. International Journal of Environmental Research and Public Health 2020, 17, 1650. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Giráldez, I.; Vega, J.M.; Vigara, J. Impact of heavy metals in the microalga Chlorella sorokiniana and assessment of its potential use in cadmium bioremediation. Aquatic Toxicology 2021, 239, 105941. [Google Scholar] [CrossRef]
- Gillet, S.; Decottignies, P.; Chardonnet, S.; Le Maréchal, P. Cadmium response and redoxin targets in Chlamydomonas reinhardtii: a proteomic approach. Photosynthesis Research 2006, 89, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Arora, N.; Pruthi, V.; Poluri, K.M. Elucidating the bioremediation mechanism of Scenedesmus sp. IITRIND2 under cadmium stress. Chemosphere 2021, 283, 131196. [Google Scholar] [CrossRef]
- Desai, S.R.; Verlecar, X.N.; Nagarajappa; Goswami, U. Genotoxicity of cadmium in marine diatom Chaetoceros tenuissimus using the alkaline Comet assay. Ecotoxicology 2006, 15, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Wang, P.; Qian, W.; Tang, X.; Zhu, X.; Wang, Z.; Cai, Z.; Wang, J. Mitigation effects of CO2-driven ocean acidification on Cd toxicity to the marine diatom Skeletonema costatum. Environmental Pollution 2020, 259, 113850. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chai, X.; Shao, Y.; Hu, L.; Xie, Q.; Wu, H. Toxicity of copper, lead, and cadmium on the motility of two marine microalgae Isochrysis galbana and Tetraselmis chui. Journal of Environmental Sciences 2011, 23, 330–335. [Google Scholar] [CrossRef]
- Novosel, N.; Kasum, D.; Žutinić, P.; Legović, T.; Ivošević DeNardis, N. Short-term effect of cadmium on the motility of three flagellated algal species. Journal of Applied Phycology 2020, 32, 4057–4067. [Google Scholar] [CrossRef]
- Wang, M.-J.; Wang, W.-X. Cadmium sensitivity, uptake, subcellular distribution and thiol induction in a marine diatom: exposure to cadmium. Aquatic toxicology 2011, 101, 377–386. [Google Scholar] [CrossRef]
- Lu, M.-M.; Gao, F.; Li, C.; Yang, H.-L. Response of microalgae Chlorella vulgaris to Cr stress and continuous Cr removal in a membrane photobioreactor. Chemosphere 2021, 262, 128422. [Google Scholar] [CrossRef]
- Arun, N.; Vidyalaxmi; Singh, D. P. Chromium (VI) induced oxidative stress in halotolerant alga Dunaliella salina and D. tertiolecta isolated from sambhar salt lake of Rajasthan (India). Cell Mol Biol (Noisy-le-grand) 2014, 60, 90–96. [Google Scholar]
- Takami, R.; Almeida, J.V.; Vardaris, C.V.; Colepicolo, P.; Barros, M.P. The interplay between thiol-compounds against chromium (VI) in the freshwater green alga Monoraphidium convolutum: Toxicology, photosynthesis, and oxidative stress at a glance. Aquatic Toxicology 2012, 118-119, 80–87. [Google Scholar] [CrossRef]
- Li, N.; Qin, L.; Jin, M.; Zhang, L.; Geng, W.; Xiao, X. Extracellular adsorption, intracellular accumulation and tolerance mechanisms of Cyclotella sp. to Cr(VI) stress. Chemosphere 2021, 270, 128662. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Scarano, G. Copper-induced changes of non-protein thiols and antioxidant enzymes in the marine microalga Phaeodactylum tricornutum. Plant Science 2004, 167, 289–296. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhu, Y.; Hu, Z.; Lei, A.; Wang, J. Towards elucidation of the toxic mechanism of copper on the model green alga Chlamydomonas reinhardtii. Ecotoxicology 2016, 25, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, Q.; Hu, C.-w.; Chen, L.; Liu, Z.-l.; Kong, Z.-m. Cobalt and manganese stress in the microalga Pavlova viridis (Prymnesiophyceae): Effects on lipid peroxidation and antioxidant enzymes. Journal of Environmental Sciences 2007, 19, 1330–1335. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Tan, L.; Wang, J. Toxicity of Co nanoparticles on three species of marine microalgae. Environ Pollut 2018, 236, 454–461. [Google Scholar] [CrossRef]
- Zada, S.; Lu, H.; Khan, S.; Iqbal, A.; Ahmad, A.; Ahmad, A.; Ali, H.; Fu, P.; Dong, H.; Zhang, X. Biosorption of iron ions through microalgae from wastewater and soil: Optimization and comparative study. Chemosphere 2021, 265, 129172. [Google Scholar] [CrossRef] [PubMed]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7, e07609. [Google Scholar] [CrossRef]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Talarek, M.; Bralska, M.; Zambrzycka, E. The effect of lead on the growth, content of primary metabolites, and antioxidant response of green alga Acutodesmus obliquus (Chlorophyceae). Environmental Science and Pollution Research 2015, 22, 19112–19123. [Google Scholar] [CrossRef]
- Ajitha, V.; Sreevidya, C.P.; Sarasan, M.; Park, J.C.; Mohandas, A.; Singh, I.S.B.; Puthumana, J.; Lee, J.-S. Effects of zinc and mercury on ROS-mediated oxidative stress-induced physiological impairments and antioxidant responses in the microalga Chlorella vulgaris. Environmental Science and Pollution Research 2021, 28, 32475–32492. [Google Scholar] [CrossRef]
- Kováčik, J.; Rotková, G.; Bujdoš, M.; Babula, P.; Peterková, V.; Matúš, P. Ascorbic acid protects Coccomyxa subellipsoidea against metal toxicity through modulation of ROS/NO balance and metal uptake. Journal of Hazardous Materials 2017, 339, 200–207. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, D.; Zheng, X.; Sheng, K.; Luo, Y.; Chen, B.; Huang, B.; Yang, X.; Pan, X. The ecotoxicity of single-layer molybdenum disulfide nanosheets on freshwater microalgae Selenastrum capricornutum. Process Safety and Environmental Protection 2023, 175, 426–436. [Google Scholar] [CrossRef]
- Luo, S.-W.; Alimujiang, A.; Balamurugan, S.; Zheng, J.-W.; Wang, X.; Yang, W.-D.; Cui, J.; Li, H.-Y. Physiological and molecular responses in halotolerant Dunaliella salina exposed to molybdenum disulfide nanoparticles. Journal of Hazardous Materials 2021, 404, 124014. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Cheng, Z.Z.; Yang, Z.M. HISN3 Mediates Adaptive Response of Chlamydomonas reinhardtii to Excess Nickel. Plant and Cell Physiology 2013, 54, 1951–1962. [Google Scholar] [CrossRef]
- Li, H.; Yao, J.; Duran, R.; Liu, J.; Min, N.; Chen, Z.; Zhu, X.; Zhao, C.; Ma, B.; Pang, W.; et al. Toxic response of the freshwater green algae Chlorella pyrenoidosa to combined effect of flotation reagent butyl xanthate and nickel. Environmental Pollution 2021, 286, 117285. [Google Scholar] [CrossRef]
- Dahmen-Ben Moussa, I.; Athmouni, K.; Chtourou, H.; Ayadi, H.; Sayadi, S.; Dhouib, A. Phycoremediation potential, physiological, and biochemical response of Amphora subtropica and Dunaliella sp. to nickel pollution. Journal of Applied Phycology 2018, 30, 931–941. [Google Scholar] [CrossRef]
- Kalehjahi, H.A.; Kosari-Nasab, M.; Amini, M.; Movafeghi, A. Defense responses of the green microalgae to the vanadium pentoxide nanoparticles. Oceanological and Hydrobiological Studies 2023, 52, 446–460. [Google Scholar] [CrossRef]
- Hamed, S.M.; Zinta, G.; Klöck, G.; Asard, H.; Selim, S.; AbdElgawad, H. Zinc-induced differential oxidative stress and antioxidant responses in Chlorella sorokiniana and Scenedesmus acuminatus. Ecotoxicology and Environmental Safety 2017, 140, 256–263. [Google Scholar] [CrossRef]
- Nowicka, B. Heavy metal–induced stress in eukaryotic algae—mechanisms of heavy metal toxicity and tolerance with particular emphasis on oxidative stress in exposed cells and the role of antioxidant response. Environmental Science and Pollution Research 2022, 29, 16860–16911. [Google Scholar] [CrossRef]
- Ding, Q.; Li, C.; Wang, H.; Xu, C.; Kuang, H. Electrochemical detection of heavy metal ions in water. Chemical communications 2021, 57, 7215–7231. [Google Scholar] [CrossRef]
- Kim, H.N.; Ren, W.X.; Kim, J.S.; Yoon, J. Fluorescent and colorimetric sensors for detection of lead, cadmium, and mercury ions. Chemical Society Reviews 2012, 41, 3210–3244. [Google Scholar] [CrossRef]
- Perelonia, K.B.S.; Benitez, K.C.D.; Banicod, R.J.S.; Tadifa, G.C.; Cambia, F.D.; Montojo, U.M. Validation of an analytical method for the determination of cadmium, lead and mercury in fish and fishery resources by graphite furnace and Cold Vapor Atomic Absorption Spectrometry. Food Control 2021, 130, 108363. [Google Scholar] [CrossRef]
- Knecht, M.R.; Sethi, M. Bio-inspired colorimetric detection of Hg2+ and Pb2+ heavy metal ions using Au nanoparticles. Anal Bioanal Chem 2009, 394, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Ichinoki, S.; Kitahata, N.; Fujii, Y. Selective Determination of Mercury(II) Ion in Water by Solvent Extraction Followed by Reversed-Phase HPLC. Journal of Liquid Chromatography & Related Technologies—J LIQ CHROMATOGR RELAT TECHNO 2004, 27, 1785–1798. [Google Scholar] [CrossRef]
- Stortini, A.M.; Baldo, M.A.; Moro, G.; Polo, F.; Moretto, L.M. Bio- and Biomimetic Receptors for Electrochemical Sensing of Heavy Metal Ions. Sensors (Basel) 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, Z.; Wang, L.; Dong, J.; Xue, J.; Yu, J.; Wang, Y.; Hua, X.; Wang, M.; Zhang, C.; et al. SERS Assay for Copper(II) Ions Based on Dual Hot-Spot Model Coupling with MarR Protein: New Cu(2+)-Specific Biorecognition Element. Anal Chem 2017, 89, 6392–6398. [Google Scholar] [CrossRef]
- García-Miranda Ferrari, A.; Carrington, P.; Rowley-Neale, S.J.; Banks, C.E. Recent advances in portable heavy metal electrochemical sensing platforms. Environmental Science: Water Research & Technology 2020, 6, 2676–2690. [Google Scholar] [CrossRef]
- Lace, A.; Cleary, J. A Review of Microfluidic Detection Strategies for Heavy Metals in Water. Chemosensors 2021, 9, 60. [Google Scholar] [CrossRef]
- Wei, Q.; Nagi, R.; Sadeghi, K.; Feng, S.; Yan, E.; Ki, S.J.; Caire, R.; Tseng, D.; Ozcan, A. Detection and spatial mapping of mercury contamination in water samples using a smart-phone. ACS Nano 2014, 8, 1121–1129. [Google Scholar] [CrossRef]
- Plácido, J.; Bustamante-López, S.; Meissner, K.E.; Kelly, D.E.; Kelly, S.L. Microalgae biochar-derived carbon dots and their application in heavy metal sensing in aqueous systems. Sci Total Environ 2019, 656, 531–539. [Google Scholar] [CrossRef]
- Quigg, A. Micronutrients. The physiology of microalgae 2016, 211–231. [Google Scholar]
- Hamed, S.M.; Selim, S.; Klöck, G.; AbdElgawad, H. Sensitivity of two green microalgae to copper stress: Growth, oxidative and antioxidants analyses. Ecotoxicology and Environmental Safety 2017, 144, 19–25. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals: Proceedings — 1st Conference on Culture of Marine Invertebrate Animals Greenport, Smith, W.L., Chanley, M.H., Eds.; Springer US: Boston, MA, 1975; pp. 29–60. [Google Scholar]
- Gregorin, C.; Albarano, L.; Somma, E.; Costantini, M.; Zupo, V. Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods. Water 2021, 13, 711. [Google Scholar] [CrossRef]
- Stauber, J.L.; Davies, C.M. Use and limitations of microbial bioassays for assessing copper bioavailability in the aquatic environment. Environmental Reviews 2000, 8, 255–301. [Google Scholar] [CrossRef]
- Serra, A.; Corcoll, N.; Guasch, H. Copper accumulation and toxicity in fluvial periphyton: the influence of exposure history. Chemosphere 2009, 74, 633–641. [Google Scholar] [CrossRef]
- Belghith, T.; Athmouni, K.; Bellassoued, K.; El Feki, A.; Ayadi, H. Physiological and biochemical response of Dunaliella salina to cadmium pollution. Journal of Applied Phycology 2016, 28, 991–999. [Google Scholar] [CrossRef]
- Kadim, M.K.; Risjani, Y. Biomarker for monitoring heavy metal pollution in aquatic environment: An overview toward molecular perspectives. Emerging Contaminants 2022, 8, 195–205. [Google Scholar] [CrossRef]
- Sharan, A.; Nara, S. Exposure-based ecotoxicity assessment of Co(3)O(4) nanoparticles in marine microalgae. Environ Sci Pollut Res Int 2021, 28, 54802–54810. [Google Scholar] [CrossRef]
- Rajalakshmi, A.M.; Silambarasan, T.; Dhandapani, R. Small scale photo bioreactor treatment of tannery wastewater, heavy metal biosorption and CO2 sequestration using microalga Chlorella sp.: a biodegradation approach. Applied Water Science 2021, 11, 108. [Google Scholar] [CrossRef]
- Dixit, R.; Wasiullah, X.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K. Bioremediation of heavy metals from soil and aquatic environment: an overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef]
- Manikandan, A.; Suresh Babu, P.; Shyamalagowri, S.; Kamaraj, M.; Muthukumaran, P.; Aravind, J. Emerging role of microalgae in heavy metal bioremediation. J Basic Microbiol 2022, 62, 330–347. [Google Scholar] [CrossRef]
- Zabed, H.M.; Akter, S.; Yun, J.; Zhang, G.; Awad, F.N.; Qi, X.; Sahu, J.N. Recent advances in biological pretreatment of microalgae and lignocellulosic biomass for biofuel production. Renewable and Sustainable Energy Reviews 2019, 105, 105–128. [Google Scholar] [CrossRef]
- Velazquez-Lucio, J.; Rodríguez-Jasso, R.M.; Colla, L.M.; Sáenz-Galindo, A.; Cervantes-Cisneros, D.E.; Aguilar, C.N.; Fernandes, B.D.; Ruiz, H.A. Microalgal biomass pretreatment for bioethanol production: a review. Biofuel Research Journal 2018, 5, 780–791. [Google Scholar] [CrossRef]
- Kloareg, B.; Quatrano, R. Structure of the cell walls of marine algae and ecophysiological functions of the matrix polysaccharides. Oceanography And Marine Biology: An Annual Review. 1988, 26, 259–315. [Google Scholar]
- Scholz, M.J.; Weiss, T.L.; Jinkerson, R.E.; Jing, J.; Roth, R.; Goodenough, U.; Posewitz, M.C.; Gerken, H.G. Ultrastructure and Composition of the Nannochloropsis gaditana Cell Wall. Eukaryotic Cell 2014, 13, 1450–1464. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Zhao, X.-Q.; Yen, H.-W.; Ho, S.-H.; Cheng, C.-L.; Lee, D.-J.; Bai, F.-W.; Chang, J.-S. Microalgae-based carbohydrates for biofuel production. Biochemical Engineering Journal 2013, 78, 1–10. [Google Scholar] [CrossRef]
- Chojnacka, K.; Chojnacki, A.; Górecka, H. Biosorption of Cr3+, Cd2+ and Cu2+ ions by blue–green algae Spirulina sp.: kinetics, equilibrium and the mechanism of the process. Chemosphere 2005, 59, 75–84. [Google Scholar] [CrossRef]
- Priyadarshini, E.; Priyadarshini, S.S.; Pradhan, N. Heavy metal resistance in algae and its application for metal nanoparticle synthesis. Applied Microbiology and Biotechnology 2019, 103, 3297–3316. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Chang, J.S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresour Technol 2020, 303, 122886. [Google Scholar] [CrossRef]
- Das, N.; Vimala, R.; Karthika, P. Biosorption of heavy metals–an overview. 2008.
- Leong, Y.K.; Chang, J.-S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresource technology 2020, 303, 122886. [Google Scholar] [CrossRef]
- Kumar, K.S.; Dahms, H.-U.; Won, E.-J.; Lee, J.-S.; Shin, K.-H. Microalgae–a promising tool for heavy metal remediation. Ecotoxicology and environmental safety 2015, 113, 329–352. [Google Scholar] [CrossRef]
- Nateras-Ramírez, O.; Martínez-Macias, M.R.; Sánchez-Machado, D.I.; López-Cervantes, J.; Aguilar-Ruiz, R.J. An overview of microalgae for Cd2+ and Pb2+ biosorption from wastewater. Bioresource Technology Reports 2022, 17, 100932. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.; Malcata, F.X. Metal uptake by microalgae: underlying mechanisms and practical applications. Biotechnology progress 2012, 28, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.R.F.; Veloso, T.; Macário, I.P.E.; Pereira, J.L.; Ventura, S.P.M.; Passos, H.; Coutinho, J.A.P. The role of biomass elemental composition and ion-exchange in metal sorption by algae. Chemosphere 2023, 314, 137675. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, B.; Bhuyan, P.P.; Nayak, R.; Patra, S.; Behera, C.; Ki, J.-S.; Ragusa, A.; Lukatkin, A.S.; Jena, M. Microalgal Phycoremediation: A Glimpse into a Sustainable Environment. Toxics 2022, 10, 525. [Google Scholar] [CrossRef]
- Mahlangu, D.; Mphahlele, K.; De Paola, F.; Mthombeni, N.H. Microalgae-Mediated Biosorption for Effective Heavy Metals Removal from Wastewater: A Review. Water 2024, 16, 718. [Google Scholar] [CrossRef]
- Ranjbar, S.; Malcata, F.X. Is Genetic Engineering a Route to Enhance Microalgae-Mediated Bioremediation of Heavy Metal-Containing Effluents? Molecules 2022, 27, 1473. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.; Malcata, F.X. Metal uptake by microalgae: underlying mechanisms and practical applications. Biotechnol Prog 2012, 28, 299–311. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Shah, M.P.; Verma, P. Bioremediation of heavy metals from wastewater: a current perspective on microalgae-based future. Letters in Applied Microbiology 2022, 75, 701–717. [Google Scholar] [CrossRef]
- Mantzorou, A.; Navakoudis, E.; Paschalidis, K.; Ververidis, F. Microalgae: a potential tool for remediating aquatic environments from toxic metals. International Journal of Environmental Science and Technology 2018, 15, 1815–1830. [Google Scholar] [CrossRef]
- Blaby-Haas, C.E.; Merchant, S.S. The ins and outs of algal metal transport. Biochim Biophys Acta 2012, 1823, 1531–1552. [Google Scholar] [CrossRef]
- Balzano, S.; Sardo, A.; Blasio, M.; Chahine, T.B.; Dell’Anno, F.; Sansone, C.; Brunet, C. Microalgal Metallothioneins and Phytochelatins and Their Potential Use in Bioremediation. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Croot, P.L.; Moffett, J.W.; Brand, L.E. Production of extracellular Cu complexing ligands by eucaryotic phytoplankton in response to Cu stress. Limnology and Oceanography 2000, 45, 619–627. [Google Scholar] [CrossRef]
- Hussain, F.; Shah, S.Z.; Ahmad, H.; Abubshait, S.A.; Abubshait, H.A.; Laref, A.; Manikandan, A.; Kusuma, H.S.; Iqbal, M. Microalgae an ecofriendly and sustainable wastewater treatment option: Biomass application in biofuel and bio-fertilizer production. A review. Renewable and Sustainable Energy Reviews 2021, 137, 110603. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Palmieri, M.C.; Garcia, O. Biosorption of Metals: State of the Art, General Features, and Potential Applications for Environmental and Technological Processes; 2011; pp. 151-176.
- Kumar, K.S.; Dahms, H.U.; Won, E.J.; Lee, J.S.; Shin, K.H. Microalgae—A promising tool for heavy metal remediation. Ecotoxicology and Environmental Safety 2015, 113, 329–352. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraghavan, K.; Yun, Y.S. Bacterial biosorbents and biosorption. Biotechnology Advances 2008, 26, 266–291. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnology Advances 2009, 27, 195–226. [Google Scholar] [CrossRef] [PubMed]
- Arica, Y.; Tüzün, İ.; Yalcin, E.; İnce, Ö.; Bayramoglu, G. Utilisation of Native, Heat and Acid-Treated Microalgae Chlamydomonas reinhardtii Preparations for Biosorption of Cr(VI) Ions. Process Biochemistry 2005, 40, 2351–2358. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnology Advances 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.L.; Malcata, F.X. Microalga-Mediated Bioremediation of Heavy Metal–Contaminated Surface Waters. In Biomanagement of Metal-Contaminated Soils, Khan, M.S., Zaidi, A., Goel, R., Musarrat, J., Eds.; Springer Netherlands: Dordrecht, 2011; pp. 365–385. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rai, L.; Gaur, J.; Kumar, H. Phycology and heavy-metal pollution. Biological reviews 1981, 56, 99–151. [Google Scholar] [CrossRef]
- Schiariti, A.; Juárez, A.B.; Rodriguez, M.C. Effects of sublethal concentrations of copper on three strains of green microalgae under autotrophic and mixotrophic culture conditions. Algological Studies 2004, 114, 143–157. [Google Scholar] [CrossRef]
- Kaushik, G.; Raza, K. Potential of novel Dunaliella salina from sambhar salt lake, India, for bioremediation of hexavalent chromium from aqueous effluents: An optimized green approach. Ecotoxicology and environmental safety 2019, 180, 430–438. [Google Scholar]
- Bishnoi, N.R.; Anju Pant, A.P.; Garima, G. Biosorption of copper from aqueous solution using algal biomass. 2004.
- Hein, M.; Pedersen, M.F.; Sand-Jensen, K. Size-dependent nitrogen uptake in micro-and macroalgae. Marine ecology progress series. Oldendorf 1995, 118, 247–253. [Google Scholar] [CrossRef]
- Quigg, A.; Reinfelder, J.R.; Fisher, N.S. Copper uptake kinetics in diverse marine phytoplankton. Limnology and oceanography 2006, 51, 893–899. [Google Scholar] [CrossRef]
- Khoshmanesh, A.; Lawson, F.; Prince, I.G. Cell surface area as a major parameter in the uptake of cadmium by unicellular green microalgae. The Chemical Engineering Journal and the Biochemical Engineering Journal 1997, 65, 13–19. [Google Scholar] [CrossRef]
Figure 1.
Natural and anthropogenic sources of heavy metal release into the environment. Created in BioRender. Cavalletti, E. (2024) BioRender.com/s82v761.
Figure 1.
Natural and anthropogenic sources of heavy metal release into the environment. Created in BioRender. Cavalletti, E. (2024) BioRender.com/s82v761.
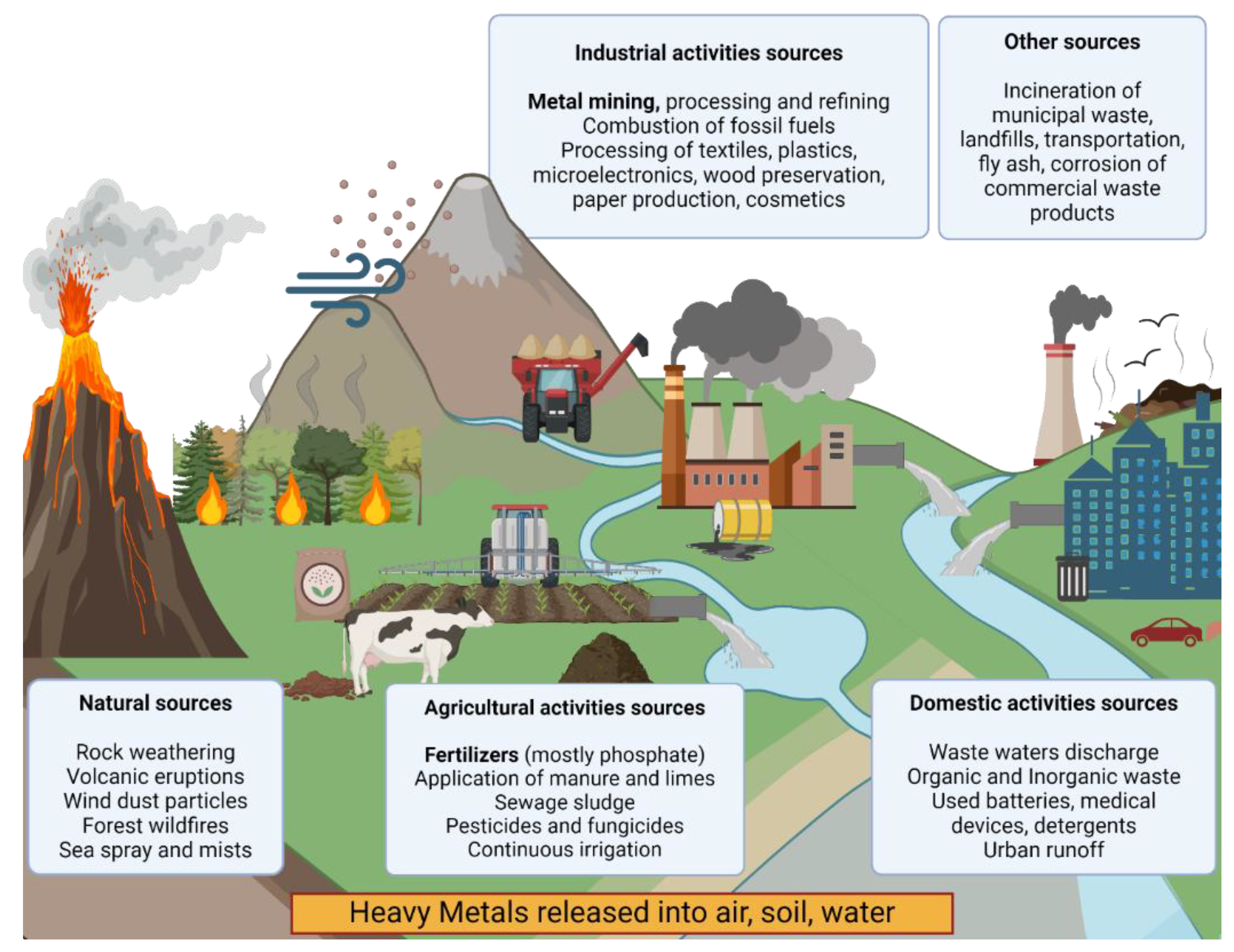
Figure 2.
Mechanisms of HMs triggered stress on microalgal cells explain the typical hallmarks of HM stress. Created in BioRender. Cavalletti, E. (2024) BioRender.com/e05z367.
Figure 2.
Mechanisms of HMs triggered stress on microalgal cells explain the typical hallmarks of HM stress. Created in BioRender. Cavalletti, E. (2024) BioRender.com/e05z367.
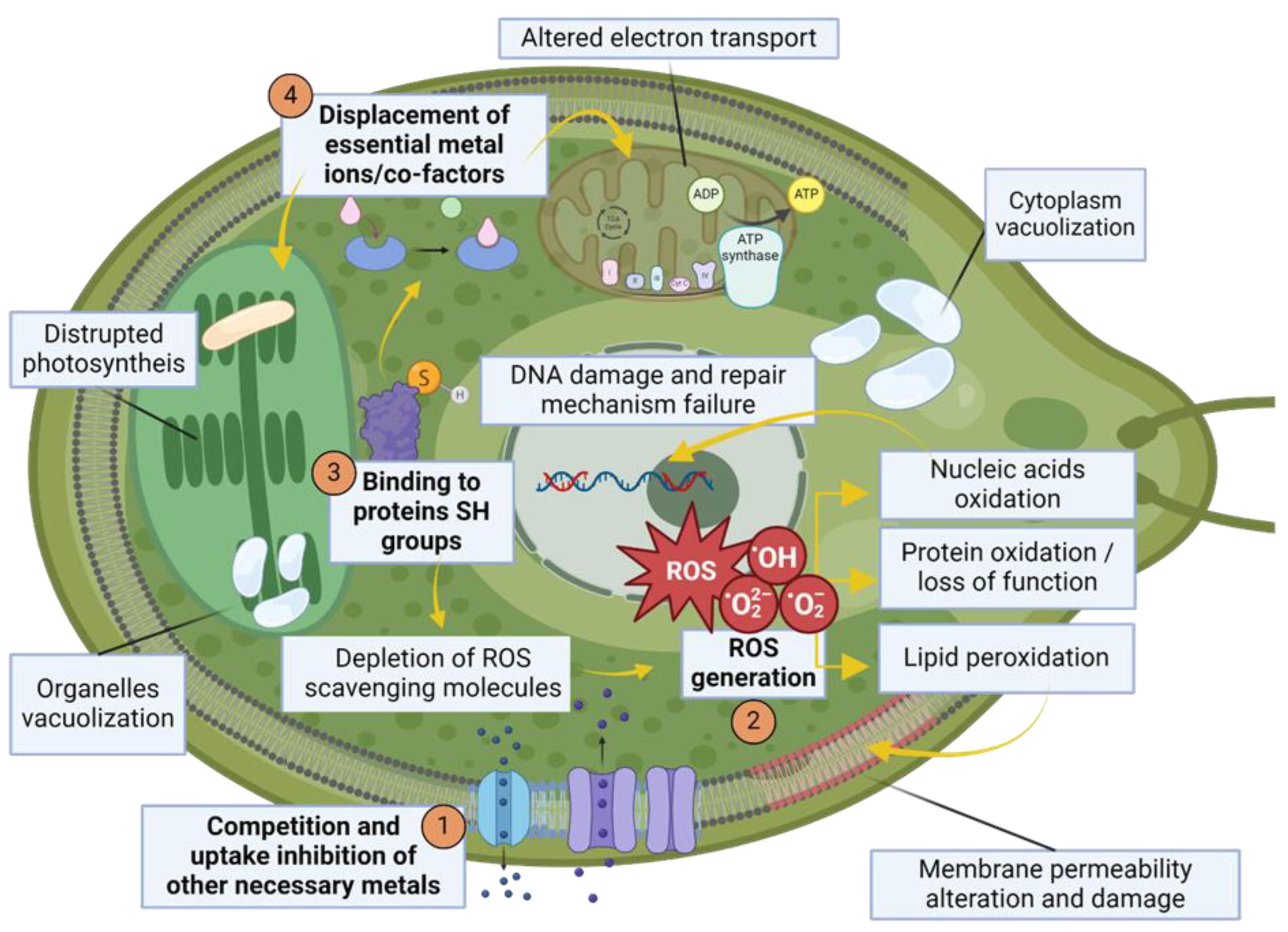
Figure 3.
Schematic representation of microalgae-mediated removal and detoxification mechanisms. 1) The adsorption onto cell wall is due to interaction with its functional groups 2) Active uptake is mediated by ion pores, channels or transporters 3) Excessive ROS production due to HM action is controlled by antioxidant enzymes and ROS-scavenging molecules 4) HMs are sequestered into organelles or vacuoles via metallo-chaperones, chelating compounds and specific transporters 5) Metal excess is effluxed from the cell via specific pumps 6) Microalgae can secrete extracellular ligands to bind metal outside the cell and prevent their entrance. Created in BioRender. Cavalletti, E. (2024) BioRender.com/t46g432.
Figure 3.
Schematic representation of microalgae-mediated removal and detoxification mechanisms. 1) The adsorption onto cell wall is due to interaction with its functional groups 2) Active uptake is mediated by ion pores, channels or transporters 3) Excessive ROS production due to HM action is controlled by antioxidant enzymes and ROS-scavenging molecules 4) HMs are sequestered into organelles or vacuoles via metallo-chaperones, chelating compounds and specific transporters 5) Metal excess is effluxed from the cell via specific pumps 6) Microalgae can secrete extracellular ligands to bind metal outside the cell and prevent their entrance. Created in BioRender. Cavalletti, E. (2024) BioRender.com/t46g432.
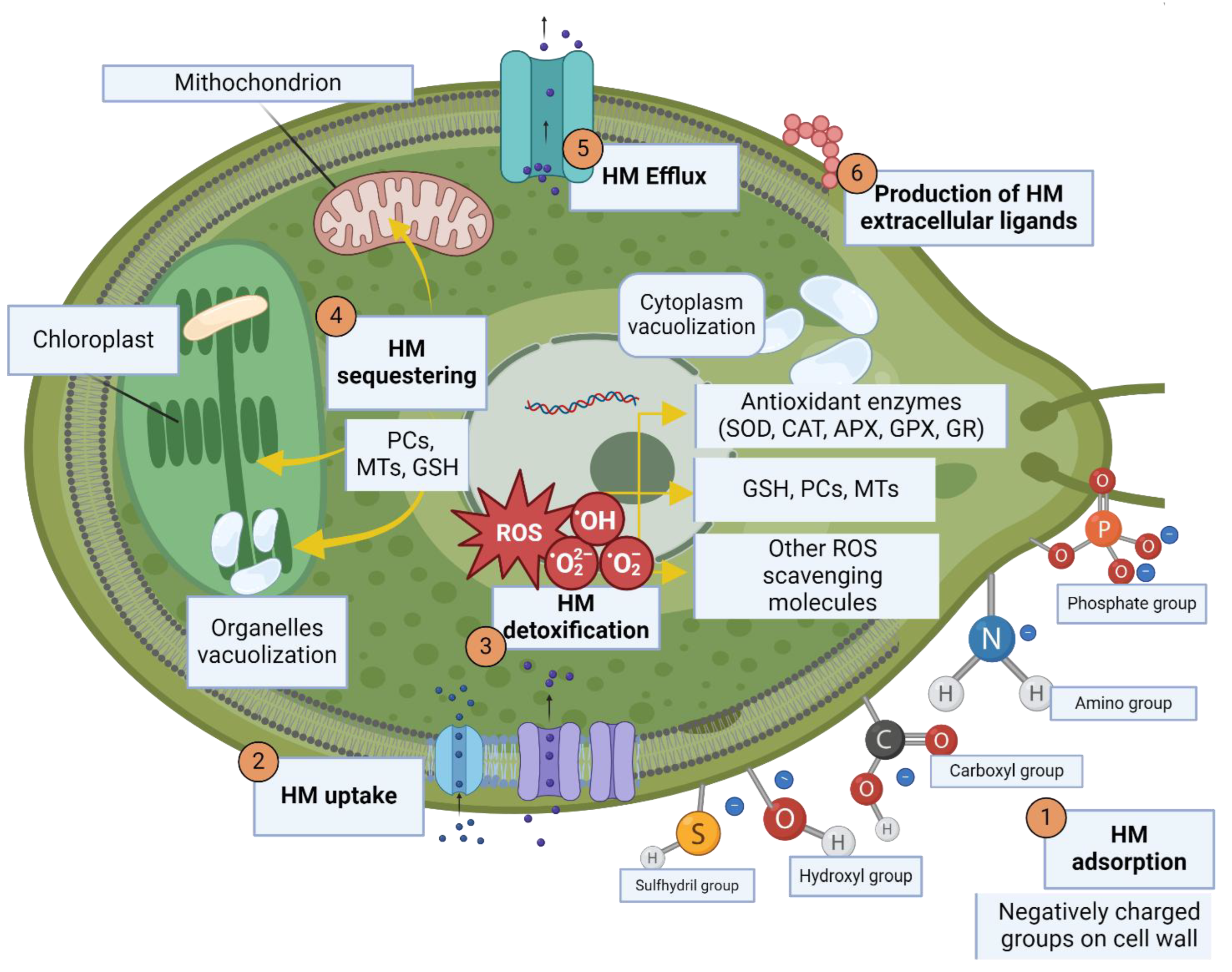

| Source type | HM release type | HMs released |
| Natural | Weathering of geologic parent material and rock outcroppings | Depends on rock composition; generally high Cr, Mn, Co Ni, Cu, Zn, Cd, Sn, Hg, Pb |
| Volcanic eruptions | High Zn, Mn, Pb, Ni, Cu, Cd, Hg. Particulate deposited contains high Cd and Hg | |
| Wind dust particles from desertic regions like Sahara | High Fe; Mn, Zn, Cr, Ni, Pb to a lesser extent | |
| Forest wild fires and prairie fires | Volatile HMs (Hg) | |
| Sea spray and mists | Mostly Cu and Zn from bubble bursting, in addition Cd, Ni, Pb from salt particles | |
| Anthropic | Industrial | |
| Mining ores | Coal mines: Cd, Fe. Gold mines: Hg | |
| Metal processing and refinement | Cu, Cd, Pb, Sn aerosols (smelting and dry/wet deposition), Cd (leachates in soil/water: Zn and Pb refining) | |
| Combustion of fossil fuels in refineries | Cd, Pb and Hg released from coal burning and petroleum combustion | |
| Agricultural | ||
| Organic and inorganic fertilizers to improve crop yield | Cd (high level from phosphate-based type), Cr, Ni, Pb, Zn | |
| Application of manure and limes | Cd, Mn, Zn, Cu and Co | |
| Sewage sludge | Cd, Zn, Cr, Pb, Ni and Cu | |
| Prolonged water irrigation | Cd and Pb from deep wells water sources | |
| Pesticides and fungicides | Cu, Pb, Zn, Fe, Mn and Hg | |
| Domestic | ||
| Urban runoff | Pb, Cu leaching | |
| Detergents | Fe, Mn, Cr, Co, Zn | |
| Used batteries | Pb, Zn, Cd, Ni, Hg | |
| Electronics | Pb, Cd, Hg, Pt, Au, Cr, Ni | |
| Others | ||
| Refuse incineration of municipal waste | Zn, Pb, Al, Sn, Fe, Cu | |
| Transportation (automobiles, diesel-powered vehicles, aircraft) | Ni, Zn, Cd (in engines and Cd from lubricants), Ni and Zn (aerosols emissions), Pb (leaded gasoline) | |
| Corrosion of commercial waste products | Cr, Cu, Pb from galvanized metals, Zn | |
| Fly ash from coal burning | Cd, Hg, Mn, Ni, Fe | |
Table 2.
Main conventional methods for HM removal, with indication of their strengths and constraints.
Table 2.
Main conventional methods for HM removal, with indication of their strengths and constraints.
| Technique | Methodology/Principle | Metals | Advantages | Bottlenecks |
| Chemical precipitation | Reaction of metals with different compounds, mostly involving hydroxides or sulfides, to produce insoluble precipitates | Cu, Cd, Pb, Zn, Mg, Mn | Effective, relatively cheap | Toxic by-products (H2S fumes) |
| Coagulation/Flocculation | Removal of soluble compounds, destabilization of colloids, flocs formation | Zn, Cd, Mn, Ni, Mg | Excellent HM removal under specific conditions | Toxicity of some reagents, strong influence of pH that limits, in some cases, coagulation |
| Ion-exchange | Ion-exchange resin, either synthetic or natural solid resin, has the specific ability to exchange its cations with the metals in the wastewater |
Ce, Fe, Pb, Co, Ni, Cr | High removal efficiency and fast kinetics | Influence of initial metal concentration, ionic charge, contact time, temperature, pH; Resins must be regenerated by expensive procedures |
| Membrane filtration | Use of various kinds of membranes as selective barriers, allowing for selection of retained material | Cd, Cu, Ni, Pb, Zn, Cr | High efficiency, easy operation and space saving | Use of surfactants or water-soluble polymers to aggregate metal ions into micelles or complexes with high molecular weight is mandatory to exceed membrane pore size |
| Electrochemical technologies | Plating-out of metal ions on a cathode surface |
Zn, Cu, Ni, Ag, Cr | Minimum solid waste | Large capital investment and expensive electricity supply |
Table 3.
Effects on microalgae of various heavy metal(loid)s, as reported from scientific literature from the last 20 years.
Table 3.
Effects on microalgae of various heavy metal(loid)s, as reported from scientific literature from the last 20 years.
| HM | Microalgal species | Side-effect | References |
| Arsenic (As) | Chlamydomonas acidophila | As(V) and As(III) (higher extent) 10 and 20 μM generate superoxide anions. TEM underlined ultrastructural changes in As(V) (stigma severely affected but mitochondria preserved, chloroplast intact but thylakoid structure sometimes disrupted, increment in starch granules, vacuolization, excretion of EPS with entrapped As nanoparticles, chromatin masses but no nucleus detection) and As(III) (intense disorganization in stigma and thylakoids, damaged mitochondria, increased lipid droplets and starch granules, nucleolus invisible, formation of nanoparticles, excretion of EPS) | [115] |
| Chlorella sorokiniana | 1 mM As(III) and 5 mM As(V) generated significant inhibition in ammonium consumption (proxy for photosynthetic assimilation of nitrogen), increase in APX and CAT gene expression and activity (higher in As(III), more toxic). | [116] | |
| Cadmium (Cd) | Chlamydomonas reinhardtii | Exposure for 4-5 days 150 μM Cd increases APX MnSOD and GST activity, decreases FeSOD activity | [117] |
| Chlamydomonas acidophila | No significant ROS generation, but severe ultrastructural alteration observed by TEM at 0.05 μM (increase in starch reserves around pyrenoid, cytoplasmic vacuoles with Cd particles later expelled, excretion of EPS, alteration in chromatin structure in nucleus) | [115] | |
| Chlorella sorokiniana | No inhibition of photosynthetic assimilation of nitrogen at 500 μM, APX increase in gene expression but decrease in enzymatic activity, sign of hyperaccumulator | [116] | |
| Scenedesmus sp. | Formation of ROS and lipid peroxidation, increase in APX, GR and CAT activity, increase in lipid content and in saturated and polyunsaturated fatty acids, SEM shown no variation in cell size but surface aberration (rough and ridged) due to Cd adsorption, changes in chlorophyll a, b and carotenoid | [118] | |
| Chaetoceros tenuissimus | DNA damages (detected by Comet assay) increased at increasing Cd concentrations: more than 50% apoptotic cells at day 8 at 10 mg/L Cd | [119] | |
| Skeletonema costatum | 1.2 mg/L Cd increases lipid peroxidation and SOD activity, deregulates genes involved in photosynthesis, increases PC synthesis and upregulates gens involved in peroxisome formation | [120] | |
| Tetraselmis chui, Isochrysis galbana | Reduction in motility: EC50 based on % of motile cells was 121.6 μM for I. galbana and 37,8 μM for T. chui | [121] | |
| Dunaliella tertiolecta, Rhodomonas maculata, Tetraselmis suecica | Cd 1 mg/L after 1 hour results in significant decrease in swimming speed and search radius for all observed species: movement was partly restored after 3 hours | [122] | |
| Thalassiosira nordenskioeldii | Cd exposure for long period influences synthesis of different PC oligomers: PC2 triggered at day 1 but constant afterwards. PC3 and PC4 are produced increasingly in time. Intra-Cd/PC-SH ratio increased with Cd dose and duration suggesting decreased detoxification ability | [123] | |
| Chromium (Cr) | Chlorella vulgaris | Cr(VI) lead to decrease in chlorophyll a content and increase of MDA (lipid peroxidation), which peaks at 2 mg/L. SOD and CAT increase at 0,5 mg/L peaking at 1 mg/L then gradually decrease at increasing Cr(VI) | [124] |
| Dunaliella salina and Dunaliella tertiolecta | Cr(VI) in D. tertiolecta decreased photosynthetic pigments, protein and carbohydrate, lead to higher increase in lipid peroxidation and H2O2 production, increase in rate of RNO bleaching, loss of pigments and thiol (-SH) group than D. salina. SEM with 40 ppm Cr(VI) show damages in both, higher in D. tertiolecta. | [125] | |
| Monoraphidium convolutum | Cr(VI) decreases photosynthetic efficiency, triggers thiols antioxidant response and lipid peroxidation, increases GSH and activity of GR and APX | [126] | |
| Cyclotella sp. | Cr(VI) 5.0 mg/L significantly decreases chlorophyll a, soluble protein content decreases and lipid peroxidation increased with time, SOD and CAT increased with concentration, peaking at 0.5 and 1.0 mg/L, and then decreased. Dissolution of cell membrane sand loss of cellular structure at Cr(VI) 2.0 mg/L observed by TEM | [127] | |
| Copper (Cu) | Phaeodactylum tricornutum, Dunaniella tertiolecta, Tetraselmis sp. | P. tricornutum increases in size and volume upon Cu 10 μg/L and after 72 h at Cu 15 μg/L swells and form cell clumps. D. tertiolecta exposed to Cu 500 μg/L produces exudates. | [87] |
| Phaeodactylum tricornutum | Cu 10 μM: rapid synthesis of PCs of various degree of polymerization, initial decrease in GHS pool, formation of Cu-PC complexes after 1 h, SOD and CAT rapid increase. GR first inhibited and then enhanced. Prolonged Cu exposure increased lipid peroxidation. | [128] | |
| Odontella mobiliensis | After 72 h: chlorophyll a decreased with dose, at Cu 574 μg/L increase of cell length and absent spines, at Cu 926 μg/L structure completely damaged. After 7 days: increase of CAT and peroxidase activities, increase in lipid peroxidation, decrease in nitrate reductase activity at Cu 21.5 μg/L | [91] | |
| Isochrysis galbana, Tetraselmis chui | Significant reduction in motility: EC50 based on % of motile cells was 1.3 μM for I. galbana and 31.4 μM L for T. chui | [121] | |
| Scenedesmus vacuolatus, Chlorella kessleri | S. vacuolatus increases protein and MDA content, and decreases chlorophyll a/b ratio. Increased CAT activity at 210 μM Cu and SOD activity and GSH content at 414 μM Cu | [96] | |
| Chlamydomonas reinhardtii | Photochemical efficiency decreased at 250 μM Cu, lipid peroxidation at its highest after 3 days, GSH content increased at 100 μM and GPX, SOD, glutathione S-transferase induced by Cu 100 and 250 μM | [129] | |
| Chlorella sorokiniana | 1 mM Cu generated significant inhibition in ammonium consumption (proxy for photosynthetic assimilation of nitrogen), APX increase in gene expression but decrease in enzymatic activity, sign of hyperaccumulator | [116] | |
| Cobalt (Co) | Pavlova viridis | Lipid peroxidation, deregulated SOD activity, increase in CAT activity and in and GSH, GPX activity markedly increased | [130] |
| Platymonas subcordiforus, Chaetoceros curvisetus and Skeletonema costatum | Co nanoparticles released Co2+ in solution, aggregated to microalgal cells as observed by SEM. Decrease in chlorophyll a and photosynthetic efficiency observed in S. costatum, TEM images showed increased agglomerates compared to other species due to smaller size and rougher surface. | [131] | |
| Iron (Fe) | Scenedesmus obliquus, Chlorella fusca, Chlorella saccharophila, A. braunii | 50 ppm Fe2+ induced nucleic acid leakage due to cell lysis and cellular ultrastructure changes observed by SEM: rougher surface and cell shrinkage | [132] |
| Lead (Pb) | Scenedesmus obliquus | Pb (141 ppm) caused increase in H2O2 and lipid peroxidation, increase in enzymatic activity of SOD, CAT, APX, GR, guaiacol peroxidase and increase in non-enzymatic ROS response (proline and polyphenols) | [133] |
| Acutodesmus obliquus | Decrease in protein content and chlorosis due to loss of chlorophyll a and b at 500 μM at day 6 and 7, increasing lipid peroxidation with dose. Pb above 10 μM decreased ascorbate and GSH. Low Pb dose (0,1 μM ) increased the activity of SOD, CAT, APX, and GR and higher concentration inhibited activities of all enzymes | [134] | |
| Isochrysis galbana, Tetraselmis chui | Reduction in motility: EC50 based on % of motile cells was 37,8 μM for I. galbana and 10,9 μM for T. chui | [121] | |
| Manganese (Mn) | Pavlova viridis | increase in CAT activity and in and GSH, GPX decreased at increasing concentrations, but at concentrations higher 50 μmol/L the trend was inverted, and it increased rapidly | [130] |
| Mercury (Hg) | Chlamydomonas reinhardtii | Chlorophyll content decrease at 4 μM Hg, lipid peroxidation at increasing Hg concentrations, activity of CAT, APX and SOD increased up to 6 μM but decreased at higher concentrations | [100] |
| Chlorella vulgaris | SEM imaging shows cell shrinkage and structural damages from 2.5% Zn treatment, decreased photosynthetic pigments, concentration-dependent reduction in protein content, ROS increasing with increasing doses, increase of CAT, initial increase of SOD and decrease over time for highest concentrations tested | [135] | |
| Coccomyxa subellipsoidea | 100 μM Hg decrease pigments, potassium ions, soluble proteins, ascorbic acid, non-protein thiols and nitric oxide signal and increased ROS production and CAT and SOD activity | [136] | |
| Molybdenum (Mn) | Selenastrum capricornutum | MoS2 nanosheets at low concentrations (≤ 1.0 mg/L) increase photosynthesis, at concentrations > 1 mg/L induce oxidative stress and membrane damages, EPS production. MoS2 nanosheets aggregates on cell and damaging/penetrating the membrane observed by TEM. | [137] |
| Dunaliella salina | MoS2 NPs enhance photosynthetic efficiency, increases protein and carbohydrate content, induce activity of CATand SOD | [138] | |
| Nickel (Ni) | Chlamydomonas reinhardtii | Ni 90 μM or higher induced generation of ROS and lipid peroxidation, proline and other non-protein thiols were increased while ascorbate was decreased | [139] |
| Chlorella pyrenoidosa | Decrease in chlorophyll a content, formation of ROS and lipid peroxidation, increased SOD and CAT activity | [140] | |
| Dunaliella sp., Amphora subtropica | Reduction in carbohydrate and protein increased total lipids, increase in phenolic compounds and carotenoid content. Increased lipid peroxidation and of SOD, CAT, GPX activity. Reductions in total chlorophyll and chlorophyll a and b at higher metal concentrations, increase in carotenoids | [141] | |
| Silver (Ag) | Dunaliella tertiolecta, Chlorella vulgaris | ROS formation and lipid peroxidation in the presence of AgNPs (1-10 mg/L) | [95] |
| Phaeodactylum tricornutum, Chlamydomonas reinhardtii | Growth rate reduction, ROS generation, membrane damages in the presence of AgCl (aq,) AgCl- and Ag+ | [92] | |
| Vanadium (V) | Chlorella sorokiniana | V2O5 NPs 200 mg/L caused plasmolysis and cell shrinkage observed by SEM. Decrease in photosynthetic pigments and phenolic content and increase of SOD, CAT and APX activity were also observed. | [142] |
| Zinc (Zn) | Chlorella sorokiniana, Scenedesmus acuminatus | Sublethal doses trigger oxidative stress in both species (H2O2 and lipid peroxidation). C. sorokiniana has more efficient antioxidant defense (accumulates more flavonoids, polyphenols, tocopherols, GSH, and ascorbate and GST, GR, SOD, POX, and APX enzyme activity higher) | [143] |
| Chlorella vulgaris | SEM imaging shows cell shrinkage and structural damages from 2,5% Zn treatment, decreased photosynthetic pigments, concentration-dependent reduction in protein content, ROS increasing with increasing doses, increase of CAT, initial increase of SOD and decrease over time for highest concentrations tested | [135] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



