Submitted:
22 March 2025
Posted:
24 March 2025
You are already at the latest version
Abstract
β-glucosidases are a relevant class of enzymes in food industry due to their role in hy-drolyzing different types of glycosidic bonds. This activity allows for formation of vola-tile compounds and release of bioactive aglycone compounds. In addition to endoge-nous β-glucosidase activity present in raw material, the function of β-glucosidases of fermenting microorganisms has been progressively clarified and increasingly appreci-ated. In this regard, several lactic acid bacteria, including Lactiplantibacillus plantarum, showed high β-glucosidase activity, which can be considered as a valid biotechnologi-cal resource in different food sectors. Here, we reviewed the huge literature in which β-glucosidases of L. plantarum was shown to play a role, highlighting how its action re-sults in enhancing the nutritional, sensory and functional properties of fermented foods. To this aim, after a brief introduction of the main functions of these enzymes in the sev-eral kingdoms, we critically analyzed the involvement of L. plantarum β-glucosidases in plant-based foods production, with a particular insight for soy, cassava and olive fer-mented products, as well as in the production of both alcoholic and non-alcoholic bev-erages. We trust, that the reports summarized here can be helpful in planning future re-search and innovative strategies to obtain pleasing, functional, and healthy foods.
Keywords:
lactobacilli
; functional food
; lactic acid bacteria
; glycosides
; aglycones
1. Introduction
β-glucosidases are a class of exoglucosidase enzymes capable of acting on terminal non-reducing β-D-glucosyl residues by hydrolyzing the β-1,4 glycosidic bond of different glycoconjugates including glucosides, oligosaccharides, and 1-O-glucosyl esters, with release of β-D-glucose [1]. On the basis of their amino acid sequences, they are classified in families and clans that share a conserved catalytic mechanism, structure, and active site residues, but may vary in substrate specificity [2,3]. These enzymes are ubiquitous in nature and are found in all domains of living organisms, Archaea, Eubacteria, and Eukaryotes (fungi, plants, and animals, including humans) [4]. In these organisms, β-glucosidases play a significant role in various biological processes and functions including nutritional acquisition and ecological associations. However, most organisms utilize this enzyme for the hydrolysis of oligosaccharides to glucose, the most usable form of carbon. The β-glucosidases fall in the enzyme class EC 3.2.1.21. At present, 133 glycoside hydrolase (GH) families are listed in the frequently updated Carbohydrate Active enZYme (CAZY) database (http://www.cazy.org) [5]. So far, they have been classified into GH1, GH3, GH5, GH9, and GH30. Family GH1 includes β-glucosidases from archaebacteria, plants and mammals, and family GH3 comprises β-glucosidases of some bacterial, mold and yeast origin [6,7,8,9].
In plants, β-glucosidases perform a wide range of biological functions such as pathogen and insect resistance, microbial interactions, lignification, phytohormones activation, signaling mechanisms, cleavage of glycosylated flavonoids, fruit ripening and pigment metabolism [4,10,11,12,13,14].
In humans, three native β-glucosidase enzymes have been identified: glucocerebrosidase, deficiency of which causes Gaucher’s disease; lactase phlorizin hydrolase, deficiency of which causes lactose intolerance; β-glucosidase, a cytosolic enzyme of broad specificity that is abundant in the kidney, liver and small intestine of mammals and plays a crucial role in the transport and/or digestion of dietary sugars [15,16,17].
In insects, β-glucosidases are mainly involved in cellobiose digestion, the breakdown of glucosinolates (glucosylated specialized metabolites) sequestered from host plants to form a dual-component defense system, and communication and recognition among sexual or social interactions. These functions have been found in different groups of insects and they adapt to the system based on the plants they feed on [18,19].
In bacteria and fungi, β-glucosidase is a crucial element of the microbial cellulose multienzyme complex since it is responsible for the regulation of the entire cellulose hydrolysis process by easing cellobiose-mediated suppression and producing the final product glucose [20,21].
The genetic diversity and expression of β-glucosidase-producing microorganisms were studied in different habitats, including food, soil, cow dung and compost, and marine environments [21,22,23,24,25,26,27]. In particular, the β-glucosidase has been identified, purified and characterized by several species belonging to the group of Lactic Acid Bacteria (LAB) [28] and plays an important role in the performance of these microorganisms in food fermentations or during the interaction with their hosts. For example, they have the potential to improve flavor and aroma of alcoholic (e.g. wine and beer) and non-alcoholic beverages (e.g. teas and juices) by releasing aromatic compounds from flavorless glycosides. Microbial β-glucosidase is also used to hydrolyze isoflavonic glycosides (e.g. soybean products) and to reduce bitterness (e.g. juices, table olives) [6].
During the interaction with their hosts, microbes can provide β-glucosidase activities to increase the bioavailability of metabolites that improve the host health, such as plant phenolic compounds, which are usually glycosylated in their dietary format and therefore are less bioavailable than the aglycone forms [29,30,31]. A known example includes the soy isoflavones, which can be released from their glycosylated precursors by some LAB β-glucosidases during the soy fermentation [32]. The β-glucosidase activities from LAB may also have implications for food security. Cassava contains high concentrations of the toxic cyanogenic glucoside linamarin, and LAB contribute to the degradation of linamarin by β-glucosidase activities [6].
On the other hand, it is known that the mycotoxin deoxynivalenol is not toxic in its glycosylated form (deoxynivalenol-3-glucoside) but it can be activated by a LAB β-glucosidase [33].
Given the importance of LAB β-glucosidases, considerable efforts have been focused to increase our knowledge on these enzymes, which usually show a broad specificity [28].
Among the LAB able to produce β-glycosidases, Lactiplantibacillus plantarum (formerly Lactobacillus plantarum) represents an important member. This heterofermentative species is also known for its high adaptability to many different conditions, since it has been isolated from various ecological niches including milk, fruit, cereal crops, vegetables, bee bread, fresh meat [34,35,36,37,38], as well as fermented foods [39,40]. Moreover, this species is widely diffused into the gastro-intestinal tract of animals; several studies, in fact, showed that it colonizes the digestive system of insects [41,42,43], fish (Iorizzo et al., 2021), and mammals, including humans [45]. The inclusion of L. plantarum in both QPS (Qualified Presumption of Safety) and GRAS (Generally Recognised as Safe) lists [46,47], together with the many intrinsic properties of this species, led to the proposal of numerous L. plantarum strains as animal and human probiotics [48,49].
L. plantarum is widely used as a starter culture in the fermentation of raw materials of plant and animal origin, where it contributes to enhancing the sensorial quality and shelf life of fermented products. Some L. plantarum strains also increase the functional properties of various fermented foods by producing a variety of bioactive compounds [50,51].
The present review aims to provide an overview of the role of the activity of β-glycosidase produced by L. plantarum as a valid biotechnological resource in different food sectors.
2. Fermented Plant-Based Foods
The naturally chemicals of fruit includes volatile compounds, both in free and bound form, which occur primarily as glycoconjugates of sugar and an aglycone [52,53]. The sugar moiety includes glucose or a disaccharide, while the aglycone part of glycosides is often represented by monoterpenes, C13-norisoprenoids, benzene derivatives, and long-chain aliphatic alcohols [54,55,56,57]. Therefore, the hydrolysis of odorless glycosylated compounds can make an important contribution to improving the flavor of fruit juices and derived beverages [52]. β-glucosidases are an important class of enzymes because they contribute to the hydrolysis of different types of glycosidic bonds and promotes the formation of free volatile compounds.
Although endogenous β-glucosidases are present in fruits such as grapes, their activity is insufficient due to low stability under juice processing and winemaking conditions. In fact, the optimal pH levels at which plant glucosidases are most active generally range from 4.0 to 6.0; therefore, in the low pH of fruit juices, only limited activity of most glycosidases has been observed [58,59].
Due to this limited action of plant endogenous glycosidases, a large proportion of the aroma compounds in juices remain inactive, in glycosidically-bound form [60]. Research has therefore focused on finding exogenous sources of glucoside hydrolases, which can be used in the production of juices and wines [60,61].
Several procedures can be used to enhance wine aroma by releasing aroma compounds from glycosidic precursors, including acid or enzymatic hydrolysis. Acid hydrolysis causes rearrangements in the aglycone structure with the formation of undesirable flavors, while enzymatic hydrolysis specifically cleaves the glycosidic linkage without altering the aglycone structure [62].
Glycosidic precursors in fruits can be found as D-glucopyranosides in which the volatile aglycone is linked to a single D-glucopyranose by a β-glycoside bond. They can also occur as disaccharides, in which the D-glucopyranose is combined with a second sugar molecule such as α-L-arabinofuranose, α-L-rhamnopyranose or α-L-apiofuranose (Figure 1).
The enzymatic hydrolysis of glycosidically bound aroma compounds occurs in two steps and involves different exoglycosidases depending on the sugar moieties of the substrates. For example, in the presence of rhamnose or apiose: first, a α-L-rhamnosidase or a β-D-apiofuranosidase cleaves the (1-6)-glycosidic linkage, and then, the flavor compounds are liberated from the monoglucosides by the action of a β-glucosidase. A hydrolysis scheme of glycosidic aroma precursors is shown in Figure 1.
Therefore, much attention has been attracted in flavor enhancement of juices or wines through the hydrolysis of the glycoside aroma precursors using microbial β-glucosidases from mould, yeast and LAB [63,64,65,66]
Phenolics, including flavonoids, widely distributed in plants, have received much attention and were recognized as the most abundant antioxidants in the human diet [67]. Increased antioxidative activity in fermented plant-based foods is primarily due to an increase in the amounts of phenolic compounds and flavonoid aglycones during fermentation, which is the result of a microbial hydrolysis activity [68].
Flavonoids are the largest class of polyphenols that can be further categorized into several subgroups including flavonols and anthocyanins, both of which are naturally distributed in plant foods as glycosides containing single or multiple sugar moieties. Flavonoid aglycones are generally more bioavailable than their respective glycosides [69].
Several studies have shown that flavonoid aglycones content in plant-based foods can increase after fermentation due to the microbial β-glucosidase. Therefore, fermentation by LAB possessing this specific enzymatic activity is an effective strategy to increase the bioavailability of natural antioxidants present in fermented plant-based products [70].
2.1. Soymilk and Soybeen Products
Isoflavones, which are produced almost exclusively by plants of the family Fabaceae, most often occur as glycosyl groups in plants. These compounds are found in plant source mainly as O-glycosides, frequently bound to glucose, but also to other sugars such as galactose, rhamnose, arabinose and xylose [71].
The biological activity of isoflavones has been well reporte [72,73]. These compounds, known as phytoestrogens, are known to reduce the incidence of hormone-dependent steroid cancers such as breast, prostate, and colon cancer [74]. In addition, isoflavones have been shown to help prevent and treat several aging-related dysfunctions and diseases, including neurodegenerative disorders, osteoporosis, metabolic and cardiovascular diseases, and menopausal symptoms [75].
Soybeans (Glycine max) are important polyphenol sources in the diet because of their high levels of isoflavones [76]. Major isoflavones in soybean consisting of three aglycones (daidzein, glycitein, genistein) and their β-glycosides (daidzin, glycitin, genistin), acetyl and malonyl-conjugated β-glycosides (6’’-O-acetyl daidzin, acetyl glycitin, acetyl genistin; 6’’O-malonyl daidzin, malonyl glycitin, malonyl genistin) [77]. The O-β-glycosidic bonds of isoflavones are partially hydrolyzed in the gut primarily by microbial β-glycosidases to their aglycones, daidzein, genistein, and glycitein, respectively (Figure 2). Glucoside isoflavones are very poorly absorbed in the small intestine as compared with their aglycones, because of their greater molecular weight and higher hydrophilicity of the glucosides [78,79]. Furthermore, the isoflavone glucosides are known to be less bioactive than their respective aglycones [80].
Several studies have demonstrated that the content of aglycones in soy product was increased after microbial fermentation by LAB, which may be due to the changes of β-glucosidase activity [81,82,83]. Therefore, the use of these bacteria as starters, with the aforementioned enzymatic activity, in soy milk fermentation could contribute to increasing bioavailable isoflavones [84,85], thereby increasing the nutritional values and health benefits of fermented soy products [75].
Aglycones release is due to β-glucosidase activity on β-glucosides, which in most cases are also present as malonylated and acetylated forms. In these latter cases, β-glucosidase is part of a two-step process, which also requires esterase enzyme to remove acetylation (Figure 2).
It has been shown that L. plantarum LP 95 was able to efficiently bio-transform glycosides to their bioactive aglycones, thus could be used as a functional starter culture to increase the antioxidant activity of fermented soymilk products [32,86]. Other studies have confirmed that several L. plantarum strains have great potential to enrich bioactive isoflavones in fermented soy milk products [84,87,88,89]. In recent studies it has been shown that soy milk fermented from L. plantarum 200655 and L. plantarum KU210152 can be used as a prophylactic functional food with neuroprotective effects against oxidative stress [90,91]. Consistently, another study highlighted the increased antioxidant capacity of the L. plantarum Y16 fermented soybean milk, with respect to unfermented one, whose ethanol and water extracts were able to protect HepG2 cells against ABAP oxidative damage; this was reported to be dependent on the activation of Nrf2/Keap1 signaling pathway and the up-regulation of antioxidant systems expression as heme oxygenase-1, superoxide dismutase, catalase, and glutathione peroxidase [92].
Therefore, increased availability of aglycones found in soy milk fermented with L. plantarum may be useful for designing new functional foods.
Moreover, use of selected L. plantarum strains which are more effective in increasing product bioactivity can also significantly increases the quality of a soy-waste product as okara. Ultrasonic treatment L. plantarum BCRC 10357 was applied to induce a biological stress response resulting in a 100 % increase in β-glucosidase activity, with this latter responsible for the biotransformation of isoflavones glycosides to bioactive aglycones (daidzein and genistein) in okara [93]. In addition, the fermentation of enzymatically hydrolyzed okara by L. plantarum UFG169 strain was reported to increase the content of both aglycone isoflavones and vitamin B2, as well as a reduction in off-flavors, thus improving both nutrition and digestibility of this product [94].
2.2. Cassava
Cassava (Manihot esculenta Crantz), also nown as yucca, manioc or mandioca, is a perennial and herbaceous shrub that belongs to the class Malpighiales and Family Euphorbiceae. This crop has great social value and cultural identity and it is now extensively cultivated throughout tropical and subtropical regions, mainly for its edible tubers as a source of carbohydrates, flavonoids, fiber, vitamin C and Minerals [95,96,97,98].
According to Food and Agriculture Organization (FAO), cassava ranks fourth as a food crop in the developing countries, after rice, maize and wheat [99]. Despite the advantages coming from its starchy tubers, other organs of cassava plant, as leaves, can be also used for edible purposes. However, these less noble parts are characterized by a low protein content, rapid post-harvest deterioration and the presence of cyanogenic glucosides as major drawbacks which strongly limit its utilisation as a food [100].
Consumption of improperly processed cassava may constitute a health problem in rural areas of sub-saharan African countries where cassava derived products provide a high percentage of the daily calory intake [101]. In severe cases this may result in acute cyanide intoxication and in chronic paralytic diseases such as konzo and neurological disorders [102,103]. Moreover, cyanogenic glucosides are spread in all parts of the cassava plant, with the highest amounts in the leaves and the root cortex (skin layer) and are present in bound form, mostly 2-(β-D-glucopyranosyloxyl)isobutyronitrile (linamarin) and, to a lesser extent, its derivative 2-(β-D-glucopyranosyloxyl)methylbutyronitrile (lotaustralin) (Figure 3) [104].
These cyanogenic glucosides are not toxic as such because they are absorbed in the gastrointestinal tract and eliminated as such through urination. However, cyanogenic glycosides are hydrolyzed into acetone cyanohydrin by the glycosidases of gut microbiota. The acetone cyanohydrin was degraded spontaneously in the small intestine in which it had alkaline pH conditions. This degradation releases hydrocyanic acid (HCN), which bound to methemoglobin (Figure 3) [105,106] and, as known, exerts its toxicity by inhibiting the cytochrome oxidase, the complex IV of mitochondrial respiratory chain, thus preventing cellular utilization of oxygen [107]. The presence of these cyanogenic glucosides is the major limiting factor to direct utilisation, thereby necessitating its processing prior to consumption. The introduction of new processing methods has helped to reduce cassava’s cyanogenic content and, therefore, exposure levels to its cyanogenic compounds. Cassava is traditionally processed by a wide range of methods, which reduce their toxicity, improve palatability and convert the perishable fresh root into stable products. In most countries, cassava can be processed by boiling, roasting, drying, cold water leaching, or fermentation. The boiling or drying process alone may not detoxify the cyanogenic glycoside. However, fermentation is a decisive step in the detoxification process [97]. Thus, most cassava-based artisanal or marketed products in Africa (e.g. gari, fufu, pupuru, apu, attieke, Ikivunde, chikwangue, Lafun, etc) and in Latin America (e.g. puba, almidon agrio, polvilho azedo) are obtained through spontaneous fermentation. During fermentation, the roots are softened and there is disintegration of the tissue structure which causes linamarin to come into contact with endogenous linamerase, which is found in the cell wall, and microbial linamerase. These enzymatic activities result in subsequent hydrolysis into glucose and acetone cyanohydrin which are easily broken down into acetone and HCN. During the natural drying phase, free HCN evaporates easily having a boiling point of 26 °C.
However, in spontaneous cassava fermentation the activity of β-glycosidase is often not sufficient to break down all cyanogenic glycosides [108]. Moreover, the linamarase elaborated by both cassava plant tissues and fermenting microorganisms has been found to be unstable under high acidic conditions characteristic of the latter part of natural fermentation. Therefore, it is important to review the detoxification methods of cassava and improve their effectiveness for greater consumption of cassava-based foods [109].
The use of exogenous β-glucosidases from microbial sources is suggested, which hydrolyse these cyanogenic glycosides at an elevated level [110,111].
The cassava fermentation process can be carried out naturally (spontaneous fermentation) by relying on the native microbial populations present in the raw materials and in the environment. Fermentation is important for improving the flavor and aroma of the product, as well as safety, especially by reducing its toxic cyanogenic glycosides.
However, the wide range and complexity of the microbiota of spontaneous cassava fermentation are the main factors responsible for the lack of homogeneity and low product quality [112].
L. plantarum and other lactic acid bacteria (LAB) have been reported as the prevalent microorganisms associated with the spontaneous fermentation of cassava [98,113,114,115,116,117,118,119,120]. Some studies have shown that it is possible to significantly degrade cyanogenic glycosides and reduce free HCN in cassava through fermentation using L. plantarum as a single starter or in co-culture with other microorganisms [112,120,121,122,123,124,125,126,127].
Therefore, further studies are desirable for the establishment of new starter cultures that can contribute to the standardization of cassava fermentation conditions, thus ensuring higher quality products and consumer acceptability.
Especially, the selection and use of L. plantarum as a starter may be an effective biotechnological strategy that may allow for greater preservation, flavor enhancement, cyanide reduction, and improved functional properties of fermented cassava-based products [97].
2.3. Olive
Another emerging field of interest for L. plantarum β-glucosidases application is represented by olive production. One of the main problem of olive industry, in fact, is the bitterness of olives which is principally due to the main representative of olive polyphenol glucosides, namely oleuropein [128]. Oleuropein is an O-glycosylated compound constituted by a D-glucose β (1-4) bound to aglycone, which can be hydrolyzed by the β-glucosidase enzyme resulting in D-glucose and aglycone production [129]. At present, the widely used method to debitter olive consists in the alkalyne treatment of drupes by means of NaOH solution; however this methods pones a series of concerns related to both the consumers (treatment-dependent reduction in nutrients) and the environment (wastewaters entriched in toxic NaOH) [130]. At the same time, the olive debittering represents a committed step in the production of table olives giving rise to the seek for alternative NaOH-free methods [131]; in this regard, the activity of microbial β-glucosidases proved to be able in hydrolying oleuropein, thus producing low-molecular-weight phenolic compounds such as hydroxytyrosol and tyrosol [132,133]. Moreover, several papers showed the capability of L. plantarum to hydrolize oleuropein, as well as the occurrence of this microorganism among the spontaneously fermenting species of table olives [40,134,135]. For these reasons, several strains of L. plantarum have been proposed as microbial cultures for table olive fermentation, since their adaptability to fermentation conditions, as well as high β-glucosidase activity makes this species particularly useful in olive debittering [136,137,138,139,140]. More recently, an elegant study [141] proposed three different mechanisms for the coversion of oleuropein into the active compound hydroxytyrosol, which seems to depend on the L. plantarum strain and need, besides β-glucosidases, also the action of esterase activities.
In addition to olive debittering, L. plantarum fermentation has been suggested as a potential approach also for the recovery of valuable bioactive compounds, as hydroxytyrosol and tyrosol, from olive mill wastewater. This species, in fact, shares with the yest Wickerhamomyces anomalus the ability to increase the content of hydroxytyrosol in wastewater phenolic extract, with both microoorganisms proved to be more efficient, than the commercial enzyme, in 2h bioconversion tests [142]. In addition, a very recent study showed other functional properties of L. plantarum present in olive mill wastewater, as the notable acidification capability and the production of antibacterial compounds [143]. These results strongly candidates this species, and its β-glucosidase activity, as a powerful tool in the management of waste and by-products from olive industry.
3. Fermented Beverages
3.1. Alcoholic Beverages
3.1.1. Wine
The wine LAB, naturally present in grape juice, play a significant role in winemaking by guiding a biological process known as malolactic fermentation (MLF).
This process involves the conversion of L-malic acid to L-lactic acid via malate decarboxylase, resulting in a reduction of wine acidity, providing microbiological stabilization and modifications of wine aroma [144].
In the last decades, various papers have shown that LAB metabolism also involves a large array of secondary enzymatic activities capable of generating many volatile secondary compounds [145]. Several studies have demonstrated the presence of β-glycosidase activity in wine LAB, leading to the release of free volatile compounds as terpenes [146,147].
Oenococcus oeni is the main bacterial species responsible for malolactic fermentation, however, in the last two decades, it has been highlighted that other LAB associated with MLF have enormous potential to influence the composition of wine [145,148].
Among all the species, L. plantarum is frequently found on grapes and in wine and is considered as a new generation of MLF starter due to its ability of high ethanol tolerance and good enological characteristics [39,149,150,151,152,153,154,155,156,157]. In addition, L. plantarum has a wide range of enzymes, including β-glucosidase, which can also contribute significantly to the formation of wine aroma during the winemaking process [50,158,159,160].
It is because of these characteristics that some commercial starters belonging to L. plantarum species have been released in the last decade [161].
The hydrolysis of glycosides, previously reported during the malolactic fermentation through selected L. plantarum strains, may be considered as an interesting option to improve the sensorial characteristics of the wines.
Iorizzo et al highlighted that some L. plantarum strains, candidates for MLF, were able to release specific terpenes from odourless grape glycosidic precursors [158]. In another study, L. plantarum M10, used as a malolactic starter after the alcoholic fermentation of Fiano grape juice, caused a significantly higher concentration of linalool in the wine [157].
Other authors found a significant increase in β-citronellol and 2-phenylethyl alcohol amounts after MLF with L. plantarum UNQLp 11 [162]. β-Citronellol is an alcoholic monoterpene that is most abundant in musts and wines and is often found as an odourless glyco-conjugated compound [60]. Thus, β-glucosidase of L. plantarum UNQLp hydrolyzes the monoterpenyl glucoside and releases β-Citronellol. In addition, it has been hypothesized that β-glucosidase activity could also explain the increase in 2-phenylethyl alcohol (an aromatic alcohol that contributes to sweet floral attributes) in wine fermented with UNQLp 11, as previously described for other LAB strains [163].
Several studies have demonstrated that the β-glucosidase activity is mainly affected by pH, temperature, ethanol and sugars [164,165,166,167].
Therefore, screening of L. plantarum strains for their glycosidase activities is important and should be done based on the substrate to be fermented [163,168,169,170].
In another study, the β-glucosidase activity of L. plantarum USC1 was stable between pH 4.5 and 7.5 and with a maximum activity at pH 5.0 and was completely inactivated at pH values below 4.0. The optimum temperature was 45 °C and the enzyme was active against a wide range of aryl b-glucosides and b-linked disaccharides [171].
Brizuela et al analyzed the amount of 1-octanol (mg/mL) obtained by the hydrolysis of the precursor octyl β-D-glucopyranoside in sterile Pinot Noir wine containing 14.5% v/v of ethanol, at different pH values (3.2, 3.5, and 3.8); the results of this study showed that the activity of β-glucosidase is reduced at low pH, but induced in the presence of high ethanol content [172].
3.1.2. Beer
Sour beer is traditionally produced through spontaneous fermentations, involving complex microbial consortia, and is characterized by higher concentrations of organic acids.
While the production of conventional beer is usually limited to yeasts fermentation, the traditional production methods for sour beer, such as Lambic and Geuze beers, originate from Belgium, involve a spontaneous fermentation by multiple microorganisms, including yeasts and bacteria [173,174,175,176,177].
Interest in sour beer has increased substantially in recent decades and research is underway on both spontaneous fermentations and alternative production techniques [178]. Pure-culture fermentations with strains of L. plantarum and S. cerevisiae, in conjunction with the careful application of processing steps, offer a valid alternative to facilitate the production of sour beer. This approach provides a higher level of process control and more rapid fermentation compared to traditional methods [179,180,181].
In a recent study, the co-inoculation of L. plantarum CECT 9567 and S. cerevisiae was applied for the production of a probiotic beer [182]. The authors, as already highlighted by others, attribute the higher polyphenol content observed in beers brewed with co-inoculation to two phenomena: hydrolysis of bound polyphenols and increased free polyphenols. These phenomena are significantly related to β-glucosidase activity [183,184].
3.2. Non-Alcoholic Fermented Fruit Products
Plant-based foods, including fruits and vegetables are naturally rich in minerals, vitamins, dietary fibers, antioxidants, and many other beneficial nutrients that make them essential components of a healthy and balanced diet. Due to new healthy trends, consumption of fruit and vegetable juices have increased in recent years [187].
Two predominant fermentation pathways have been identified in the production of fruit juices: the alcoholic pathway, which involves the utilization of yeast, and the non-alcoholic pathway, which relies on the action of LAB.
Being a traditional food biotechnology, fermentation by LAB is widely used for fruit and vegetable fermentation to convert bioactive components, enhance beneficial properties, extend shelf-life and improve sensory characteristics of final products [188,189].
Fermentation by LAB increasing the content of functional nutrients including polyphenols, flavonoids, organic acids, polysaccharides, amino acids, vitamins, minerals, and other efficacious components, giving the fruit excellent antioxidant, antibacterial, anti-inflammatory, and gut microbiota modulation activities [190,191,192].
Moreover, LAB fermentation can impart distinctive fruity and floral aromas to fruits through the production of esters, ketones, alcohols, terpenes, etc. [193,194,195].
Among the LAB, L. plantarum is quite interesting, as far as its application in the fermentation of a wide range of plant-based substrates is concerned, such as vegetables and fruit juices, since it has genome plasticity and high versatility and flexibility [188].
Several studies have shown that L. plantarum, used as a starter culture, facilitates the enhancement of flavor and aroma in fermented fruit juices through its β-glucosidase activity. Pomegranate juices fermented by L. plantarum POM1 and LP09 were characterized by high levels of terpenes, such as limonene, β-myrcene, γ-terpinene, α-terpinene, α-terpinolene, and p-cymene [196]. Monoterpenes are present in pomegranate juice as either free or glycosidically conjugated precursors and the release of glycosidically bound aromatic compounds has been shown to result in modification or enhancement of the characteristic flavor [197]. In a recent study, it has been found that Sabre mango juice fermented with L. plantarum L75 produced higher levels of β-myrcene [198]. A similar increase in β-myrcene was reported in L. plantarum POM1- and LP09-fermented pomegranate juices [196].
The main biological properties recognized to β-myrcene are the following: anxiolytic, antioxidant, anti-aging, anti-inflammatory and analgesic [199].
A recent study showed that the nutritional quality and flavor characteristics of apricot juice can be improved by L. plantarum LP56 fermentation; specifically, after 6 hours, there was a significant increase in the content of volatile compounds, including α-terpineol, nerol, β-pinene, and terpinene, reaching its maximum level [200].
In another study, L. plantarum NCU116 fermentation had a beneficial effect on the physico-chemical properties, bioactive compounds, antioxidant property, and improved the aroma profile of Momordica charantia juice. Among the terpenoids, myrthenol was the main aromatic compound of fermented juice [201].
Myrtenol is a volatile compound belonging to the terpenoid family of monocyclic monoterpenes and contributed to the woody, pine, balsam, sweet and mint notes. In addition, several reports demonstrated the pharmacological properties of myrtenol, including its antioxidant, antibacterial, antifungal, antidiabetic, anxiolytic, and gastroprotective activities [202].
Ricci et al detected, among the terpene and norisoprenoid class, an increase in limonene, β-linalool, β-damascenon, and eugenol in elderberry juice fermented by L. plantarum 285 [203].
The increase of these compounds could be related with the ability of L. plantarum to produce β-glucosidase [158,194,196].
Fruit beverages fermented by L. plantarum, not only are characterized by a pleasant aroma and taste, but show many health-promoting benefits due to their content of metabolites such as vitamins, organic acids, and phenolic compounds [204,205,206,207].
Several studies have shown that the bioavailability of phenolic compounds is enhanced by different LAB after fermentation of different fruit products [193,208,209,210].
According to several studies, L. plantarum produces enzymes such as β-glucosidase during the fermentation process, which is able to hydrolyze phenolic glycosides to the corresponding aglycones, which have radical scavenging properties [184,211,212]. This process results in an increase in the antioxidant activity of the fermented product [213].
The antioxidant activity of phenolics is related to their chemical structure. In general, flavonoid compounds present a stronger antioxidant activity than non-flavonoids, and combined forms such as glycosides present a lower activity than the free forms [214].
In a study conducted by Meng et al. the effect of different L. plantarum strains on the physicochemical characteristics and antioxidant activities of loquat juice was investigated. Results showed that nerolidol production was significantly upregulated in loquat juices fermented by L. plantarum LP2 [215].
Nerolidol, a terpenoid has good anti-inflammatory, antioxidant, neuroprotective and cardioprotective activities [216,217]. Furthermore, after fermentation by L. plantarum LP2 the antioxidant activity and the total flavonoid content in loquat juice significantly increased.
Landete et al. showed that deglycosylation by L. plantarum CECT 748 transformed food aryl glycosides (phloridzin, esculin, daidzin, and salicin) into their corresponding aryl aglycones (phloretin, esculetin, daidzein, and saligenin). Therefore, in addition to the improvement of their bioavailability, the deglycosylation of specific aryl glycosides by L. plantarum CECT 748 increase the antioxidant activity of glycosylated phenolic compounds [169].
Table 1 shows the main positive effects of the enzymatic activity of L. plantarum, mainly attributable to β-glucosidase, in different fermented fruit products. The data refer to articles published in the last 10 years. However, the impact of fermentation on phenolic compounds seems to depend heavily on the bacterial strain used and the starting material.
Li et al reported that LAB fermentation with L. plantarum 90 significantly increased the total phenolic content, while decreasing the total flavonoid content in fermented jujube juices [228].
The same author had found that the fermentation of apple juice by L. plantarum ATCC14917 caused an increase in antioxidant activity while decreasing the total content of phenols and flavonoids [218], attributing, according to Tian, the greater antioxidant captivity detected to other possible mechanisms [241].
In another study, the flavonol glycosides in sea buckthorn as well as anthocyanins in chokeberry remained unaffected by the fermentation with several several L. plantarum strains obtained from DSMZ (Braunschweig, Germany) [242].
Wei et al. [243] reported general decrease in anthocyanins, phenolic acids, flavonols and flavanols in bog bilberry juice fermented with L. plantarum B7 or L. plantarum C8-1.
Therefore, this suggests that careful selection within the L. plantarum species is crucial in order to identify the most suitable strains to be used for each specific biotechnological application aimed at improving the functional properties of the final products [244].
4. Conclusions
In the last decades, the needs of both producers and consumers in the food sector have continuously grown, thus requiring a particular attention not only to organoleptic aspects but also in terms of health and well-being. This review shows how the enzyme β-glucosidase can be considered crucial for the hydrolysis of several glycosides that give added value to the fermented food matrix. In particular, the activity of β-glucosidase during fermentation by L. plantarum can be considered an important biotechnological strategy in order to increase the nutritional, sensory and functional properties of specific fermented foods. The studies cited in this review showed that the optimal conditions for the β-glucosidase activity extensively differs among the L. plantarum strains and is significantly affected by substrate composition and culture conditions. Therefore, it is essential to optimize these conditions to improve this enzymatic activity also according to the production process adopted to obtain each specific fermented food. Considering that lactic fermentation is an important technology to increase functional properties of fermented foods, we believe that the selection of β-glucosidase-producing L. plantarum strains should remain a focal point of interest in future research, since it can be a valid tool for the design of new functional foods.
Author Contributions
Conceptualization, M.I., C.D.M. and G.P.; writing—original draft preparation, M.I. and G.P.; writing—review and editing, M.I., G.P., C.D.M. and F.C.; visualization, C.D.M.; supervision, M.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sengupta, S.; Datta, M.; Datta, S. β-Glucosidase: Structure, Function and Industrial Applications. In Glycoside Hydrolases; Elsevier, 2023; pp. 97–120.
- Henrissat, B. A Classification of Glycosyl Hydrolases Based on Amino Acid Sequence Similarities; 1991; Vol. 280.
- Henrissat, B.; Davies, G. Structural and Sequence-Based Classification of Glycoside Hydrolases. Curr Opin Struct Biol 1997, 7, 637–644. [CrossRef]
- Ketudat Cairns, J.R.; Esen, A. β-Glucosidases. Cellular and Molecular Life Sciences 2010, 67, 3389–3405. [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes Database (CAZy): An Expert Resource for Glycogenomics. Nucleic Acids Res 2009, 37, D233–D238. [CrossRef]
- Kannan, P.; Shafreen M, M.; Achudhan, A.B.; Gupta, A.; Saleena, L.M. A Review on Applications of β-Glucosidase in Food, Brewery, Pharmaceutical and Cosmetic Industries. Carbohydr Res 2023, 530, 108855. [CrossRef]
- Magwaza, B.; Amobonye, A.; Pillai, S. Microbial β-Glucosidases: Recent Advances and Applications. Biochimie 2024, 225, 49–67. [CrossRef]
- Mól, P.C.G.; Júnior, J.C.Q.; Veríssimo, L.A.A.; Boscolo, M.; Gomes, E.; Minim, L.A.; Da Silva, R. β-Glucosidase: An Overview on Immobilization and Some Aspects of Structure, Function, Applications and Cost. Process Biochemistry 2023, 130, 26–39. [CrossRef]
- Singh, G.; Verma, A.K.; Kumar, V. Catalytic Properties, Functional Attributes and Industrial Applications of β-Glucosidases. 3 Biotech 2016, 6, 3. [CrossRef]
- Liu, C.; He, S.; Chen, J.; Wang, M.; Li, Z.; Wei, L.; Chen, Y.; Du, M.; Liu, D.; Li, C.; et al. A Dual-subcellular Localized Β-glucosidase Confers Pathogen and Insect Resistance without a Yield Penalty in Maize. Plant Biotechnol J 2024, 22, 1017–1032. [CrossRef]
- Kotik, M.; Kulik, N.; Valentová, K. Flavonoids as Aglycones in Retaining Glycosidase-Catalyzed Reactions: Prospects for Green Chemistry. J Agric Food Chem 2023, 71, 14890–14910. [CrossRef]
- Xu, P.; Wu, T.; Ali, A.; Wang, J.; Fang, Y.; Qiang, R.; Liu, Y.; Tian, Y.; Liu, S.; Zhang, H.; et al. Rice β-Glucosidase 4 (Os1βGlu4) Regulates the Hull Pigmentation via Accumulation of Salicylic Acid. Int J Mol Sci 2022, 23, 10646. [CrossRef]
- Lee, K.H.; Piao, H.L.; Kim, H.-Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.-J.; Hwang, I. Activation of Glucosidase via Stress-Induced Polymerization Rapidly Increases Active Pools of Abscisic Acid. Cell 2006, 126, 1109–1120. [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; Atanassova, R. The Sugar-Signaling Hub: Overview of Regulators and Interaction with the Hormonal and Metabolic Network. Int J Mol Sci 2018, 19, 2506. [CrossRef]
- Boer, D.E.C.; van Smeden, J.; Bouwstra, J.A.; Aerts, J.M.F.G. Glucocerebrosidase: Functions in and Beyond the Lysosome. J Clin Med 2020, 9, 736. [CrossRef]
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.A.; Williamson, G. Dietary Flavonoid and Isoflavone Glycosides Are Hydrolysed by the Lactase Site of Lactase Phlorizin Hydrolase. FEBS Lett 2000, 468, 166–170. [CrossRef]
- Elferink, H.; Bruekers, J.P.J.; Veeneman, G.H.; Boltje, T.J. A Comprehensive Overview of Substrate Specificity of Glycoside Hydrolases and Transporters in the Small Intestine. Cellular and Molecular Life Sciences 2020, 77, 4799–4826. [CrossRef]
- He, S.; Jiang, B.; Chakraborty, A.; Yu, G. The Evolution of Glycoside Hydrolase Family 1 in Insects Related to Their Adaptation to Plant Utilization. Insects 2022, 13, 786. [CrossRef]
- Friedrichs, J.; Schweiger, R.; Müller, C. Unique Metabolism of Different Glucosinolates in Larvae and Adults of a Leaf Beetle Specialised on Brassicaceae. Sci Rep 2022, 12, 10905. [CrossRef]
- Ahmed, A.; Nasim, F.; Batool, K.; Bibi, A. Microbial β -Glucosidase: Sources, Production and Applications. J Appl Environ Microbiol 2017, 5, 31–46. [CrossRef]
- Bhatia, Y.; Mishra, S.; Bisaria, V.S. Microbial β-Glucosidases: Cloning, Properties, and Applications. Crit Rev Biotechnol 2002, 22, 375–407. [CrossRef]
- Fu, Y.; Yin, Z.; Wu, L.; Yin, C. Diversity of Cultivable β-Glycosidase-Producing Micro-Organisms Isolated from the Soil of a Ginseng Field and Their Ginsenosides-Hydrolysing Activity. Lett Appl Microbiol 2014, 58, 138–144. [CrossRef]
- Tiwari, R.; Kumar, K.; Singh, S.; Nain, L.; Shukla, P. Molecular Detection and Environment-Specific Diversity of Glycosyl Hydrolase Family 1 β-Glucosidase in Different Habitats. Front Microbiol 2016, 7. [CrossRef]
- Su, H.; Xiao, Z.; Yu, K.; Zhang, Q.; Lu, C.; Wang, G.; Wang, Y.; Liang, J.; Huang, W.; Huang, X.; et al. High Diversity of β-Glucosidase-Producing Bacteria and Their Genes Associated with Scleractinian Corals. Int J Mol Sci 2021, 22, 3523. [CrossRef]
- Oladoja, E.; Oyewole, O.; Adamu, B.; Balogun, A.; Musa, O. Microbial β-Glucosidase: Source, Production and Applications. International Journal of Biology Sciences 2019, 1, 14–22. [CrossRef]
- Ouyang, B.; Wang, G.; Zhang, N.; Zuo, J.; Huang, Y.; Zhao, X. Recent Advances in β-Glucosidase Sequence and Structure Engineering: A Brief Review. Molecules 2023, 28, 4990. [CrossRef]
- Zang, X.; Liu, M.; Fan, Y.; Xu, J.; Xu, X.; Li, H. The Structural and Functional Contributions of β-Glucosidase-Producing Microbial Communities to Cellulose Degradation in Composting. Biotechnol Biofuels 2018, 11, 51. [CrossRef]
- Michlmayr, H.; Kneifel, W. β-Glucosidase Activities of Lactic Acid Bacteria: Mechanisms, Impact on Fermented Food and Human Health. FEMS Microbiol Lett 2014, 352, 1–10. [CrossRef]
- Colautti, A.; Camprini, L.; Ginaldi, F.; Comi, G.; Reale, A.; Coppola, F.; Iacumin, L. Safety Traits, Genetic and Technological Characterization of Lacticaseibacillus Rhamnosus Strains. LWT 2024, 207, 116578. [CrossRef]
- Coppola, F.; Abdalrazeq, M.; Fratianni, F.; Ombra, M.N.; Testa, B.; Zengin, G.; Ayala Zavala, J.F.; Nazzaro, F. Rosaceae Honey: Antimicrobial Activity and Prebiotic Properties. Antibiotics 2025, 14, 298. [CrossRef]
- Nazzaro, F.; Ombra, M.N.; Coppola, F.; De Giulio, B.; d’Acierno, A.; Coppola, R.; Fratianni, F. Antibacterial Activity and Prebiotic Properties of Six Types of Lamiaceae Honey. Antibiotics 2024, 13, 868. [CrossRef]
- Letizia, F.; Fusco, G.M.; Fratianni, A.; Gaeta, I.; Carillo, P.; Messia, M.C.; Iorizzo, M. Application of Lactiplantibacillus Plantarum LP95 as a Functional Starter Culture in Fermented Tofu Production. Processes 2024, 12, 1093. [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic Fate of Deoxynivalenol-3-Glucoside during Digestion. Toxicol Lett 2011, 206, 264–267. [CrossRef]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.K.; Rademaker, J.L.W.; Starrenburg, M.J.C.; Kleerebezem, M.; Molenaar, D.; Van Hylckama Vlieg, J.E.T. Phenotypic and Genomic Diversity of Lactobacillus Plantarum Strains Isolated from Various Environmental Niches. Environ Microbiol 2010, 12, 758–773. [CrossRef]
- Martino, M.E.; Bayjanov, J.R.; Caffrey, B.E.; Wels, M.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebezem, M.; van Hijum, S.A.F.T.; Leulier, F. Nomadic Lifestyle of Lactobacillus Plantarum Revealed by Comparative Genomics of 54 Strains Isolated from Different Habitats. Environ Microbiol 2016, 18, 4974–4989. [CrossRef]
- Siezen, R.J.; van Hylckama Vlieg, J.E.T. Genomic Diversity and Versatility of Lactobacillus Plantarum, a Natural Metabolic Engineer. Microb Cell Fact 2011, 10. [CrossRef]
- Letizia, F.; Albanese, G.; Testa, B.; Vergalito, F.; Bagnoli, D.; Di Martino, C.; Carillo, P.; Verrillo, L.; Succi, M.; Sorrentino, E.; et al. In Vitro Assessment of Bio-Functional Properties from Lactiplantibacillus Plantarum Strains. Curr Issues Mol Biol 2022, 44, 2321–2334. [CrossRef]
- Filannino, P.; De Angelis, M.; Di Cagno, R.; Gozzi, G.; Riciputi, Y.; Gobbetti, M. How Lactobacillus Plantarum Shapes Its Transcriptome in Response to Contrasting Habitats. Environ Microbiol 2018, 20, 3700–3716. [CrossRef]
- Testa, B.; Lombardi, S.J.; Tremonte, P.; Succi, M.; Tipaldi, L.; Pannella, G.; Sorrentino, E.; Iorizzo, M.; Coppola, R. Biodiversity of Lactobacillus Plantarum from Traditional Italian Wines. World J Microbiol Biotechnol 2014, 30, 2299–2305. [CrossRef]
- Iorizzo, M.; Lombardi, S.J.; Macciola, V.; Testa, B.; Lustrato, G.; Lopez, F.; De Leonardis, A. Technological Potential of Lactobacillus Strains Isolated from Fermented Green Olives: In Vitro Studies with Emphasis on Oleuropein-Degrading Capability. Scientific World Journal 2016, 2016. [CrossRef]
- Iorizzo, M.; Testa, B.; Ganassi, S.; Lombardi, S.J.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Probiotic Properties and Potentiality of Lactiplantibacillus Plantarum Strains for the Biological Control of Chalkbrood Disease. Journal of Fungi 2021, 7. [CrossRef]
- Chundakkattumalayil, H.C.; Raghavan, K.T. Health Endorsing Potential of Lactobacillus Plantarum MBTU-HK1 and MBTU-HT of Honey Bee Gut Origin. J Appl Biol Biotechnol 2021, 9, 63–68. [CrossRef]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R.; et al. Inter-and Intra-Species Diversity of Lactic Acid Bacteria in Apis Mellifera Ligustica Colonies. Microorganisms 2020, 8, 1–17. [CrossRef]
- Iorizzo, M.; Albanese, G.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; D’andrea, M.; Iaffaldano, N.; Coppola, R. Presence of Lactic Acid Bacteria in the Intestinal Tract of the Mediterranean Trout (Salmo Macrostigma) in Its Natural Environment. Life 2021, 11. [CrossRef]
- Park, S.-Y.; Lim, S.-D. Probiotic Characteristics of Lactobacillus Plantarum FH185 Isolated from Human Feces. Korean J Food Sci Anim Resour 2015, 35, 615–621. [CrossRef]
- Salvetti, E.; O’Toole, P.W. When Regulation Challenges Innovation: The Case of the Genus Lactobacillus. Trends Food Sci Technol 2017, 66, 187–194. [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the List of QPS-recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA 12: Suitability of Taxonomic Units Notified to EFSA until March 2020. EFSA Journal 2020, 18. [CrossRef]
- Iorizzo, M.; Paventi, G.; Di Martino, C. Biosynthesis of Gamma-Aminobutyric Acid (GABA) by Lactiplantibacillus Plantarum in Fermented Food Production. Curr Issues Mol Biol 2023, 46, 200–220. [CrossRef]
- Iorizzo, M.; Di Martino, C.; Letizia, F.; Crawford, T.W.; Paventi, G. Production of Conjugated Linoleic Acid (CLA) by Lactiplantibacillus Plantarum: A Review with Emphasis on Fermented Foods. Foods 2024, 13, 975. [CrossRef]
- Paventi, G.; Di Martino, C.; Crawford Jr, T.W.; Iorizzo, M. Enzymatic Activities of Lactiplantibacillus Plantarum: Technological and Functional Role in Food Processing and Human Nutrition. Food Biosci 2024, 61, 104944. [CrossRef]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus Plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. Biomed Res Int 2018, 2018.
- Caffrey, A.J.; Lerno, L.A.; Zweigenbaum, J.; Ebeler, S.E. Direct Analysis of Glycosidic Aroma Precursors Containing Multiple Aglycone Classes in Vitis Vinifera Berries. J Agric Food Chem 2020, 68, 3817–3833. [CrossRef]
- Liang, Z.; Fang, Z.; Pai, A.; Luo, J.; Gan, R.; Gao, Y.; Lu, J.; Zhang, P. Glycosidically Bound Aroma Precursors in Fruits: A Comprehensive Review. Crit Rev Food Sci Nutr 2022, 62, 215–243. [CrossRef]
- Zhu, N.; Wang, T.; Ge, L.; Li, Y.; Zhang, X.; Bao, H. γ -Amino Butyric Acid (GABA) Synthesis Enabled by Copper-Catalyzed Carboamination of Alkenes. Org Lett 2017, 19, 4718–4721. [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine Aroma Compounds in Grapes: A Critical Review. Crit Rev Food Sci Nutr 2015, 55, 202–218. [CrossRef]
- Maicas, S.; Mateo, J.J. Hydrolysis of Terpenyl Glycosides in Grape Juice and Other Fruit Juices: A Review. Appl Microbiol Biotechnol 2005, 67, 322–335. [CrossRef]
- Krammer, G.; Winterhalter, P.; Schwab, M.; Schreier, P. Glycosidically Bound Aroma Compounds in the Fruits of Prunus Species: Apricot (p. Armeniaca, l.), Peach (p. Persica, l.), Yellow Plum (p. Domestica, l. Ssp. Syriaca). J Agric Food Chem 1991, 39, 778–781. [CrossRef]
- de Morais Souto, B.; Florentino Barbosa, M.; Marinsek Sales, R.M.; Conessa Moura, S.; de Rezende Bastos Araújo, A.; Ferraz Quirino, B. The Potential of β-Glucosidases for Aroma and Flavor Improvement in the Food Industry. The Microbe 2023, 1, 100004. [CrossRef]
- Fernández-Pacheco, P.; García-Béjar, B.; Briones Pérez, A.; Arévalo-Villena, M. Free and Immobilised β-Glucosidases in Oenology: Biotechnological Characterisation and Its Effect on Enhancement of Wine Aroma. Front Microbiol 2021, 12. [CrossRef]
- Sarry, J.; Gunata, Z. Plant and Microbial Glycoside Hydrolases: Volatile Release from Glycosidic Aroma Precursors. Food Chem 2004, 87, 509–521. [CrossRef]
- Wang, Z.; Chen, K.; Liu, C.; Ma, L.; Li, J. Effects of Glycosidase on Glycoside-Bound Aroma Compounds in Grape and Cherry Juice. J Food Sci Technol 2023, 60, 761–771. [CrossRef]
- Dziadas, M.; Jeleń, H.H. Comparison of Enzymatic and Acid Hydrolysis of Bound Flavor Compounds in Model System and Grapes. Food Chem 2016, 190, 412–418. [CrossRef]
- Muradova, M.; Proskura, A.; Canon, F.; Aleksandrova, I.; Schwartz, M.; Heydel, J.-M.; Baranenko, D.; Nadtochii, L.; Neiers, F. Unlocking Flavor Potential Using Microbial β-Glucosidases in Food Processing. Foods 2023, 12, 4484. [CrossRef]
- Maicas, S.; Mateo, J. Microbial Glycosidases for Wine Production. Beverages 2016, 2, 20. [CrossRef]
- Hjelmeland, A.K.; Ebeler, S.E. Glycosidically Bound Volatile Aroma Compounds in Grapes and Wine: A Review. Am J Enol Vitic 2015, 66, 1–11. [CrossRef]
- Wilkowska, A.; Pogorzelski, E. Aroma Enhancement of Cherry Juice and Wine Using Exogenous Glycosidases from Mould, Yeast and Lactic Acid Bacteria. Food Chem 2017, 237, 282–289. [CrossRef]
- Zhang, Z.; Li, X.; Sang, S.; McClements, D.J.; Chen, L.; Long, J.; Jiao, A.; Jin, Z.; Qiu, C. Polyphenols as Plant-Based Nutraceuticals: Health Effects, Encapsulation, Nano-Delivery, and Application. Foods 2022, 11, 2189. [CrossRef]
- Sun, H.; Xue, Y.; Lin, Y. Enhanced Catalytic Efficiency in Quercetin-4′-Glucoside Hydrolysis of Thermotoga Maritima β-Glucosidase A by Site-Directed Mutagenesis. J Agric Food Chem 2014, 62, 6763–6770. [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)Phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects against Chronic Diseases. Antioxid Redox Signal 2013, 18, 1818–1892. [CrossRef]
- Yang, X.; Hong, J.; Wang, L.; Cai, C.; Mo, H.; Wang, J.; Fang, X.; Liao, Z. Effect of Lactic Acid Bacteria Fermentation on Plant-Based Products. Fermentation 2024, 10, 48. [CrossRef]
- Bustamante-Rangel, M.; Delgado-Zamarreño, M.M.; Pérez-Martín, L.; Rodríguez-Gonzalo, E.; Domínguez-Álvarez, J. Analysis of Isoflavones in Foods. Compr Rev Food Sci Food Saf 2018, 17, 391–411. [CrossRef]
- Atlante, A.; Bobba, A.; Paventi, G.; Pizzuto, R.; Passarella, S. Genistein and Daidzein Prevent Low Potassium-Dependent Apoptosis of Cerebellar Granule Cells. Biochem Pharmacol 2010, 79, 758–767.
- Sharma, D.; Singh, V.; Kumar, A.; Singh, T.G. Genistein: A Promising Ally in Combating Neurodegenerative Disorders. Eur J Pharmacol 2025, 991. [CrossRef]
- Zaheer, K.; Humayoun Akhtar, M. An Updated Review of Dietary Isoflavones: Nutrition, Processing, Bioavailability and Impacts on Human Health. Crit Rev Food Sci Nutr 2017, 57, 1280–1293. [CrossRef]
- Jayachandran, M.; Xu, B. An Insight into the Health Benefits of Fermented Soy Products. Food Chem 2019, 271, 362–371. [CrossRef]
- Hu, C.; Wong, W.T.; Wu, R.; Lai, W.F. Biochemistry and Use of Soybean Isoflavones in Functional Food Development. Crit Rev Food Sci Nutr 2020, 60, 2098–2112. [CrossRef]
- Jung, Y.S.; Rha, C.-S.; Baik, M.-Y.; Baek, N.-I.; Kim, D.-O. A Brief History and Spectroscopic Analysis of Soy Isoflavones. Food Sci Biotechnol 2020, 29, 1605–1617. [CrossRef]
- Izumi, T.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kikuchi, M.; Piskula, M.K.; Kubota, Y. Soy Isoflavone Aglycones Are Absorbed Faster and in Higher Amounts than Their Glucosides in Humans. J Nutr 2000, 130, 1695–1699. [CrossRef]
- Liu, H.; Wang, Y.; Zhu, D.; Xu, J.; Xu, X.; Liu, J. Bioaccessibility and Application of Soybean Isoflavones: A Review. Food Reviews International 2022, 39, 5948–5967. [CrossRef]
- Setchell, K.D.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for Lack of Absorption of Soy Isoflavone Glycosides in Humans, Supporting the Crucial Role of Intestinal Metabolism for Bioavailability. Am J Clin Nutr 2002, 76, 447–453. [CrossRef]
- Chien, H.-L.; Huang, H.-Y.; Chou, C.-C. Transformation of Isoflavone Phytoestrogens during the Fermentation of Soymilk with Lactic Acid Bacteria and Bifidobacteria. Food Microbiol 2006, 23, 772–778. [CrossRef]
- Chun, J.; Kim, G.M.; Lee, K.W.; Choi, I.D.; Kwon, G.; Park, J.; Jeong, S.; Kim, J.; Kim, J.H. Conversion of Isoflavone Glucosides to Aglycones in Soymilk by Fermentation with Lactic Acid Bacteria. J Food Sci 2007, 72. [CrossRef]
- Pyo, Y.-H.; Lee, T.-C.; Lee, Y.-C. Enrichment of Bioactive Isoflavones in Soymilk Fermented with β-Glucosidase-Producing Lactic Acid Bacteria. Food Research International 2005, 38, 551–559. [CrossRef]
- Lim, Y.J.; Lim, B.; Kim, H.Y.; Kwon, S.-J.; Eom, S.H. Deglycosylation Patterns of Isoflavones in Soybean Extracts Inoculated with Two Enzymatically Different Strains of Lactobacillus Species. Enzyme Microb Technol 2020, 132, 109394. [CrossRef]
- Tang, A.L.; Shah, N.P.; Wilcox, G.; Walker, K.Z.; Stojanovska, L. Fermentation of Calcium-Fortified Soymilk with Lactobacillus: Effects on Calcium Solubility, Isoflavone Conversion, and Production of Organic Acids. J Food Sci 2007, 72. [CrossRef]
- Letizia, F.; Fratianni, A.; Cofelice, M.; Testa, B.; Albanese, G.; Di Martino, C.; Panfili, G.; Lopez, F.; Iorizzo, M. Antioxidative Properties of Fermented Soymilk Using Lactiplantibacillus Plantarum LP95. Antioxidants 2023, 12. [CrossRef]
- Du, L.; Ro, K.-S.; Zhang, Y.; Tang, Y.-J.; Li, W.; Xie, J.; Wei, D. Effects of Lactiplantibacillus Plantarum X7021 on Physicochemical Properties, Purines, Isoflavones and Volatile Compounds of Fermented Soymilk. Process Biochemistry 2022, 113, 150–157. [CrossRef]
- Hidayati, D.; Soetjipto, S.; Catur Adi, A. Characteristic and Isoflavone Level of Soymilk Fermented by Single and Mixed Culture of Lactobacillus Plantarum and Yoghurt Starter. Journal of Food and Nutrition Research 2021, 9, 55–60. [CrossRef]
- Izaguirre, J.K.; Barañano, L.; Castañón, S.; Alkorta, I.; Quirós, L.M.; Garbisu, C. Optimization of the Bioactivation of Isoflavones in Soymilk by Lactic Acid Bacteria. Processes 2021, 9, 963. [CrossRef]
- Bock, H.-J.; Lee, H.-W.; Lee, N.-K.; Paik, H.-D. Probiotic Lactiplantibacillus Plantarum KU210152 and Its Fermented Soy Milk Attenuates Oxidative Stress in Neuroblastoma Cells. Food Research International 2024, 177, 113868. [CrossRef]
- Choi, G.-H.; Bock, H.-J.; Lee, N.-K.; Paik, H.-D. Soy Yogurt Using Lactobacillus Plantarum 200655 and Fructooligosaccharides: Neuroprotective Effects against Oxidative Stress. J Food Sci Technol 2022, 59, 4870–4879. [CrossRef]
- Wang, A.; Hou, K.; Mu, G.; Ma, C.; Tuo, Y. Antioxidative Effect of Soybean Milk Fermented by Lactobacillus Plantarum Y16 on 2, 2 –Azobis (2-Methylpropionamidine) Dihydrochloride (ABAP)-Damaged HepG2 Cells. Food Biosci 2021, 44. [CrossRef]
- Peng, H.T.; Yang, C.Y.; Fang, T.J. Enhanced β-Glucosidase Activity of Lactobacillus Plantarum by a Strategic Ultrasound Treatment for Biotransformation of Isoflavones in Okara. Food Sci Technol Res 2018, 24, 777–784. [CrossRef]
- Wang, R.; Thakur, K.; Feng, J.-Y.; Zhu, Y.-Y.; Zhang, F.; Russo, P.; Spano, G.; Zhang, J.-G.; Wei, Z.-J. Functionalization of Soy Residue (Okara) by Enzymatic Hydrolysis and LAB Fermentation for B<inf>2</Inf> Bio-Enrichment and Improved in Vitro Digestion. Food Chem 2022, 387. [CrossRef]
- Parmar, A.; Sturm, B.; Hensel, O. Crops That Feed the World: Production and Improvement of Cassava for Food, Feed, and Industrial Uses. Food Secur 2017, 9, 907–927. [CrossRef]
- Malik, A.I.; Kongsil, P.; Nguyễn, V.A.; Ou, W.; Sholihin; Srean, P.; Sheela, M.; Becerra López-Lavalle, L.A.; Utsumi, Y.; Lu, C.; et al. Cassava Breeding and Agronomy in Asia: 50 Years of History and Future Directions. Breed Sci 2020, 70, 145–166. [CrossRef]
- Halake, N.H.; Chinthapalli, B. Fermentation of Traditional African Cassava Based Foods: Microorganisms Role in Nutritional and Safety Value. Journal of Experimental Agriculture International 2020, 56–65. [CrossRef]
- Lacerda, I.; Miranda, R.; Borelli, B.; Nunes, A.; Nardi, R.; Lachance, M.; Rosa, C. Lactic Acid Bacteria and Yeasts Associated with Spontaneous Fermentations during the Production of Sour Cassava Starch in Brazil. Int J Food Microbiol 2005, 105, 213–219. [CrossRef]
- Howeler, R.; Lutaladio, N.; Thomas, G. Save and Grow: Cassava: A Guide to Sustainable Production Intensification; 2013.
- Latif, S.; Müller, J. Potential of Cassava Leaves in Human Nutrition: A Review. Trends Food Sci Technol 2015, 44, 147–158. [CrossRef]
- Adebayo, W.G. Cassava Production in Africa: A Panel Analysis of the Drivers and Trends. Heliyon 2023, 9, e19939. [CrossRef]
- Kashala-Abotnes, E.; Okitundu, D.; Mumba, D.; Boivin, M.J.; Tylleskär, T.; Tshala-Katumbay, D. Konzo: A Distinct Neurological Disease Associated with Food (Cassava) Cyanogenic Poisoning. Brain Res Bull 2019, 145, 87–91. [CrossRef]
- Adamolekun, B. Neurological Disorders Associated with Cassava Diet: A Review of Putative Etiological Mechanisms. Metab Brain Dis 2011, 26, 79–85. [CrossRef]
- Fukushima, A.R.; Nicoletti, M.A.; Rodrigues, A.J.; Pressutti, C.; Almeida, J.; Brandão, T.; Kinue Ito, R.; Bafille Leoni, L.A.; Souza Spinosa, H. De Cassava Flour: Quantification of Cyanide Content. Food Nutr Sci 2016, 07, 592–599. [CrossRef]
- Cressey, P.; Reeve, J. Metabolism of Cyanogenic Glycosides: A Review. Food and Chemical Toxicology 2019, 125, 225–232. [CrossRef]
- Renchinkhand, G.; Park, Y.W.; Cho, S.-H.; Song, G.-Y.; Bae, H.C.; Choi, S.-J.; Nam, M.S. Identification of β-Glucosidase Activity of L Actobacillus Plantarum CRNB22 in Kimchi and Its Potential to Convert Ginsenoside Rb 1 from P Anax Ginseng. J Food Biochem 2015, 39, 155–163. [CrossRef]
- National Research Council Acute Exposure Guideline Levels for Selected Airborne Chemicals; National Academies Press: Washington, D.C., 2002; ISBN 978-0-309-08511-3.
- Oloya, B.; Adaku, C.; Andama, M. The Cyanogenic Potential of Certain Cassava Varieties in Uganda and Their Fermentation-Based Detoxification. In Cassava - Recent Updates on Food, Feed, and Industry; IntechOpen, 2024.
- Brimer, L. Cassava Production and Processing and Impact on Biological Compounds; 2015; ISBN 9780124047099.
- Bouatenin, K.M.J.-P.; Theodore, D.N.; Alfred, K.K.; Hermann, C.W.; Marcellin, D.K. Excretion of β-Glucosidase and Pectinase by Microorganisms Isolated from Cassava Traditional Ferments Used for Attieke Production in Côte d’Ivoire. Biocatal Agric Biotechnol 2019, 20, 101217. [CrossRef]
- Panghal, A.; Munezero, C.; Sharma, P.; Chhikara, N. Cassava Toxicity, Detoxification and Its Food Applications: A Review. Toxin Rev 2021, 40, 1–16. [CrossRef]
- Penido, F.C.L.; Piló, F.B.; Sandes, S.H. de C.; Nunes, Á.C.; Colen, G.; Oliveira, E. de S.; Rosa, C.A.; Lacerda, I.C.A. Selection of Starter Cultures for the Production of Sour Cassava Starch in a Pilot-Scale Fermentation Process. Brazilian Journal of Microbiology 2018, 49, 823–831. [CrossRef]
- Omar, N. Ben; Ampe, F.; Raimbault, M.; Guyot, J.-P.; Tailliez, P. Molecular Diversity of Lactic Acid Bacteria from Cassava Sour Starch (Colombia). Syst Appl Microbiol 2000, 23, 285–291. [CrossRef]
- Bamigbade, G.B.; Sanusi, J.F.O.; Oyelami, O.I.; Daniel, O.M.; Alimi, B.O.; Ampofo, K.A.; Liu, S.-Q.; Shah, N.P.; Ayyash, M. Identification and Characterization of Lactic Acid Bacteria Isolated from Effluents Generated During Cassava Fermentation as Potential Candidates for Probiotics. Food Biotechnol 2023, 37, 413–433. [CrossRef]
- Oyedeji, O.; Ogunbanwo, S.T.; Onilude, A.A. Predominant Lactic Acid Bacteria Involved in the Traditional Fermentation of Fufu and Ogi, Two Nigerian Fermented Food Products. Food Nutr Sci 2013, 04, 40–46. [CrossRef]
- Putri, W.D.R.; Haryadi; Marseno, D.W.; Cahyanto, M.N. Role of Lactic Acid Bacteria on Structural and Physicochemical Properties of Sour Cassava Starch. APCBEE Procedia 2012, 2, 104–109. [CrossRef]
- Crispim, S.M.; Nascimento, A.M.A.; Costa, P.S.; Moreira, J.L.S.; Nunes, A.C.; Nicoli, J.R.; Lima, F.L.; Mota, V.T.; Nardi, R.M.D. Molecular Identification of Lactobacillus Spp. Associated with Puba, a Brazilian Fermented Cassava Food. Brazilian Journal of Microbiology 2013, 44, 15–21. [CrossRef]
- Wilfrid Padonou, S.; Nielsen, D.S.; Hounhouigan, J.D.; Thorsen, L.; Nago, M.C.; Jakobsen, M. The Microbiota of Lafun, an African Traditional Cassava Food Product. Int J Food Microbiol 2009, 133, 22–30. [CrossRef]
- Figueroa, C.; Davila, A.M.; Pourquié, J. Lactic Acid Bacteria of the Sour Cassava Starch Fermentation. Lett Appl Microbiol 1995, 21, 126–130. [CrossRef]
- Kostinek, M.; Specht, I.; Edward, V.A.; Pinto, C.; Egounlety, M.; Sossa, C.; Mbugua, S.; Dortu, C.; Thonart, P.; Taljaard, L.; et al. Characterisation and Biochemical Properties of Predominant Lactic Acid Bacteria from Fermenting Cassava for Selection as Starter Cultures. Int J Food Microbiol 2007, 114, 342–351. [CrossRef]
- Lei, V.; Amoa-Awua, W.K.A.; Brimer, L. Degradation of Cyanogenic Glycosides by Lactobacillus Plantarum Strains from Spontaneous Cassava Fermentation and Other Microorganisms. Int J Food Microbiol 1999, 53, 169–184. [CrossRef]
- Wakil, S.M.; Benjamin, I.B. Starter Developed Pupuru, a Traditional Africa Fermented Food from Cassava (Manihot Esculenta). Int Food Res J 2015, 22, 2565–2570.
- Gunawan, S.; Widjaja, T.; Zullaikah, S.; Ernawati, L.; Istianah, N.; Aparamarta, H.W.; Prasetyoko, D. Effect of Fermenting Cassava with Lactobacillus Plantarum, Saccharomyces Cereviseae, and Rhizopus Oryzae on the Chemical Composition of Their Flour. Int Food Res J 2015, 22, 1280–1287.
- Kostinek, M.; Specht, I.; Edward, V.A.; Schillinger, U.; Hertel, C.; Holzapfel, W.H.; Franz, C.M.A.P. Diversity and Technological Properties of Predominant Lactic Acid Bacteria from Fermented Cassava Used for the Preparation of Gari, a Traditional African Food. Syst Appl Microbiol 2005, 28, 527–540. [CrossRef]
- Tefera, T.; Ameha, K.; Biruhtesfa, A. Cassava Based Foods: Microbial Fermentation by Single Starter Culture towards Cyanide Reduction, Protein Enhancement and Palatability. Int Food Res J 2014, 21, 1751–1756.
- Kimaryo, V.M.; Massawe, G.A.; Olasupo, N.A.; Holzapfel, W.H. The Use of a Starter Culture in the Fermentation of Cassava for the Production of “Kivunde”, a Traditional Tanzanian Food Product. Int J Food Microbiol 2000, 56, 179–190. [CrossRef]
- Damayanti, E.; Kurniadi, M.; Helmi, R.L.; Frediansyah, A. Single Starter Lactobacillus Plantarum for Modified Cassava Flour (Mocaf) Fermentation. In Proceedings of the IOP Conference Series: Earth and Environmental Science; 2020; Vol. 462.
- Charoenprasert, S.; Mitchell, A. Factors Influencing Phenolic Compounds in Table Olives (Olea Europaea). J Agric Food Chem 2012, 60, 7081–7095. [CrossRef]
- Guggenheim, K.G.; Crawford, L.M.; Paradisi, F.; Wang, S.C.; Siegel, J.B. β-Glucosidase Discovery and Design for the Degradation of Oleuropein. ACS Omega 2018, 3, 15754–15762. [CrossRef]
- Rokni, Y.; Abouloifa, H.; Bellaouchi, R.; Gaamouche, S.; Mchiouer, K.; Hasnaoui, I.; Lamzira, Z.; Ghabbour, N.; Asehraou, A. Technological Process of Fermented Olive. Arabian Journal of Chemical and Environmental Research 2017, 07, 63–91.
- Habibi, M.; Golmakani, M.-T.; Farahnaky, A.; Mesbahi, G.; Majzoobi, M. NaOH-Free Debittering of Table Olives Using Power Ultrasound. Food Chem 2016, 192, 775–781. [CrossRef]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of Starter Cultures to Table Olive Fermentation: An Overview on the Experimental Studies. Front Microbiol 2012, 3. [CrossRef]
- Pino, A.; Vaccalluzzo, A.; Solieri, L.; Romeo, F. V.; Todaro, A.; Caggia, C.; Arroyo-López, F.N.; Bautista-Gallego, J.; Randazzo, C.L. Effect of Sequential Inoculum of Beta-Glucosidase Positive and Probiotic Strains on Brine Fermentation to Obtain Low Salt Sicilian Table Olives. Front Microbiol 2019, 10. [CrossRef]
- Ciafardini, G.; Marsilio, V.; Lanza, B.; Pozzi, N. Hydrolysis of Oleuropein by Lactobacillus Plantarum Strains Associated with Olive Fermentation. Appl Environ Microbiol 1994, 60, 4142–4147. [CrossRef]
- Rokni, Y.; Abouloifa, H.; Bellaouchi, R.; Hasnaoui, I.; Gaamouche, S.; Lamzira, Z.; Salah, R.B.E.N.; Saalaoui, E.; Ghabbour, N.; Asehraou, A. Characterization of β-Glucosidase of Lactobacillus Plantarum FSO1 and Candida Pelliculosa L18 Isolated from Traditional Fermented Green Olive. Journal of Genetic Engineering and Biotechnology 2021, 19, 117. [CrossRef]
- De Leonardis, A.; Testa, B.; Macciola, V.; Lombardi, S.J.; Iorizzo, M. Exploring Enzyme and Microbial Technology for the Preparation of Green Table Olives. European Food Research and Technology 2016, 242, 363–370. [CrossRef]
- Ghabbour, N.; Rokni, Y.; Abouloifa, H.; Bellaouchi, R.; Chihib, N.-E.; Ben Salah, R.; Lamzira, Z.; Saalaoui, E.; Asehraou, A. In Vitro Biodegradation of Oleuropein by Lactobacillus Plantarum FSO175 in Stress Conditions (PH, NaCl and Glucose). Journal of microbiology, biotechnology and food sciences 2020, 9, 769–773. [CrossRef]
- Ghabbour, N.; Rokni, Y.; Lamzira, Z.; Thonart, P.; Chihib, N.E.; Peres, C.; Asehraou, A. Controlled Fermentation of Moroccan Picholine Green Olives by Oleuropein-Degrading Lactobacilli Strains. Grasas y Aceites 2016, 67, e138. [CrossRef]
- Vaccalluzzo, A.; Pino, A.; De Angelis, M.; Bautista-Gallego, J.; Romeo, F.V.; Foti, P.; Caggia, C.; Randazzo, C.L. Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactiplantibacillus Plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives. Microorganisms 2020, 8, 1607. [CrossRef]
- Zago, M.; Lanza, B.; Rossetti, L.; Muzzalupo, I.; Carminati, D.; Giraffa, G. Selection of Lactobacillus Plantarum Strains to Use as Starters in Fermented Table Olives: Oleuropeinase Activity and Phage Sensitivity. Food Microbiol 2013, 34, 81–87. [CrossRef]
- Vaccalluzzo, A.; Solieri, L.; Tagliazucchi, D.; Cattivelli, A.; Martini, S.; Pino, A.; Caggia, C.; Randazzo, C.L. Metabolomic and Transcriptional Profiling of Oleuropein Bioconversion into Hydroxytyrosol during Table Olive Fermentation by Lactiplantibacillus Plantarum. Appl Environ Microbiol 2022, 88. [CrossRef]
- Romeo, F. V.; Granuzzo, G.; Foti, P.; Ballistreri, G.; Caggia, C.; Rapisarda, P. Microbial Application to Improve Olive Mill Wastewater Phenolic Extracts. Molecules 2021, 26. [CrossRef]
- Sciurba, L.; Indelicato, S.; Gaglio, R.; Barbera, M.; Marra, F.P.; Bongiorno, D.; Davino, S.; Piazzese, D.; Settanni, L.; Avellone, G. Analysis of Olive Oil Mill Wastewater from Conventionally Farmed Olives: Chemical and Microbiological Safety and Polyphenolic Profile for Possible Use in Food Product Functionalization. Foods 2025, 14, 449. [CrossRef]
- Cappello, M.S.; Zapparoli, G.; Logrieco, A.; Bartowsky, E.J. Linking Wine Lactic Acid Bacteria Diversity with Wine Aroma and Flavour. Int J Food Microbiol 2017, 243, 16–27. [CrossRef]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic Acid Bacteria in Wine: Technological Advances and Evaluation of Their Functional Role. Front Microbiol 2021, 11. [CrossRef]
- Boido, E.; Lloret, A.; Medina, K.; Carrau, F.; Dellacassa, E. Effect of β-Glycosidase Activity of Oenococcus Oeni on the Glycosylated Flavor Precursors of Tannat Wine during Malolactic Fermentation. J Agric Food Chem 2002, 50, 2344–2349. [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and Bacterial Modulation of Wine Aroma and Flavour. Aust J Grape Wine Res 2005, 11, 139–173. [CrossRef]
- Bartowsky, E.J.; Costello, P.J.; Chambers, P.J. Emerging Trends in the Application of Malolactic Fermentation. Aust J Grape Wine Res 2015, 21, 663–669. [CrossRef]
- du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The Next Generation of Malolactic Fermentation Starter Cultures—an Overview. Food Bioproc Tech 2011, 4, 876–906. [CrossRef]
- Krieger-Weber, S.; Heras, J.M.; Suarez, C. Lactobacillus Plantarum, a New Biological Tool to Control Malolactic Fermentation: A Review and an Outlook. Beverages 2020, 6, 23. [CrossRef]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological Properties of Lactobacillus Plantarum Strains Isolated from Grape Must Fermentation. Food Microbiol 2016, 57, 187–194. [CrossRef]
- Succi, M.; Pannella, G.; Tremonte, P.; Tipaldi, L.; Coppola, R.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E. Sub-Optimal PH Preadaptation Improves the Survival of Lactobacillus Plantarum Strains and the Malic Acid Consumption in Wine-like Medium. Front Microbiol 2017, 8. [CrossRef]
- Tufariello, M.; Capozzi, V.; Spano, G.; Cantele, G.; Venerito, P.; Mita, G.; Grieco, F. Effect of Co-Inoculation of Candida Zemplinina, Saccharomyces Cerevisiae and Lactobacillus Plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms 2020, Vol. 8, Page 726 2020, 8, 726. [CrossRef]
- Pannella, G.; Lombardi, S.J.; Coppola, F.; Vergalito, F.; Iorizzo, M.; Succi, M.; Tremonte, P.; Iannini, C.; Sorrentino, E.; Coppola, R. Effect of Biofilm Formation by Lactobacillus Plantarum on the Malolactic Fermentation in Model Wine. Foods 2020, 9. [CrossRef]
- Bravo-Ferrada, B.M.; Hollmann, A.; Delfederico, L.; Valdés La Hens, D.; Caballero, A.; Semorile, L. Patagonian Red Wines: Selection of Lactobacillus Plantarum Isolates as Potential Starter Cultures for Malolactic Fermentation. World J Microbiol Biotechnol 2013, 29, 1537–1549. [CrossRef]
- Balmaseda, A.; Rozès, N.; Bordons, A.; Reguant, C. Characterization of Malolactic Fermentation by Lactiplantibacillus Plantarum in Red Grape Must. LWT 2024, 199, 116070. [CrossRef]
- Lombardi, S.J.; Pannella, G.; Iorizzo, M.; Testa, B.; Succi, M.; Tremonte, P.; Sorrentino, E.; Di Renzo, M.; Strollo, D.; Coppola, R. Inoculum Strategies and Performances of Malolactic Starter Lactobacillus Plantarum M10: Impact on Chemical and Sensorial Characteristics of Fiano Wine. Microorganisms 2020, 8, 516. [CrossRef]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; García-Ruiz, A.; Muñoz-González, C.; Bartolomé, B.; Moreno-Arribas, M.V. Selection and Technological Potential of Lactobacillus Plantarum Bacteria Suitable for Wine Malolactic Fermentation and Grape Aroma Release. LWT 2016, 73, 557–566. [CrossRef]
- Devi, A.; Anu-Appaiah, K.A.; Lin, T.F. Timing of Inoculation of Oenococcus Oeni and Lactobacillus Plantarum in Mixed Malo-Lactic Culture along with Compatible Native Yeast Influences the Polyphenolic, Volatile and Sensory Profile of the Shiraz Wines. LWT 2022, 158, 113130. [CrossRef]
- Testa, B.; Coppola, F.; Iorizzo, M.; Di Renzo, M.; Coppola, R.; Succi, M. Preliminary Characterisation of Metschnikowia Pulcherrima to Be Used as a Starter Culture in Red Winemaking. Beverages 2024, 10, 88. [CrossRef]
- Brizuela, N.; Tymczyszyn, E.E.; Semorile, L.C.; Valdes La Hens, D.; Delfederico, L.; Hollmann, A.; Bravo-Ferrada, B. Lactobacillus Plantarum as a Malolactic Starter Culture in Winemaking: A New (Old) Player? Electronic Journal of Biotechnology 2019, 38, 10–18. [CrossRef]
- Brizuela, N.S.; Franco-Luesma, E.; Bravo-Ferrada, B.M.; Pérez-Jiménez, M.; Semorile, L.; Tymczyszyn, E.E.; Pozo-Bayon, M.A. Influence of Patagonian Lactiplantibacillus Plantarum and Oenococcus Oeni Strains on Sensory Perception of Pinot Noir Wine after Malolactic Fermentation. Aust J Grape Wine Res 2021, 27, 118–127. [CrossRef]
- Grimaldi, A.; Bartowsky, E.; Jiranek, V. Screening of Lactobacillus Spp. and Pediococcus Spp. for Glycosidase Activities That Are Important in Oenology. J Appl Microbiol 2005, 99, 1061–1069. [CrossRef]
- Gouripur, G.; Kaliwal, B. Screening and Optimization of β-Glucosidase Producing Newly Isolated Lactobacillus Plantarum Strain LSP-24 from Colostrum Milk. Biocatal Agric Biotechnol 2017, 11, 89–96. [CrossRef]
- Barbagallo, R.N.; Spagna, G.; Palmeri, R.; Restuccia, C.; Giudici, P. Selection, Characterization and Comparison of β-Glucosidase from Mould and Yeasts Employable for Enological Applications. Enzyme Microb Technol 2004, 35, 58–66. [CrossRef]
- Olguín, N.; Alegret, J.O.; Bordons, A.; Reguant, C. β-Glucosidase Activity and Bgl Gene Expression of Oenococcus Oeni Strains in Model Media and Cabernet Sauvignon Wine. Am J Enol Vitic 2011, 62, 99–105. [CrossRef]
- Fia, G.; Millarini, V.; Granchi, L.; Bucalossi, G.; Guerrini, S.; Zanoni, B.; Rosi, I. Beta-Glucosidase and Esterase Activity from Oenococcus Oeni: Screening and Evaluation during Malolactic Fermentation in Harsh Conditions. LWT 2018, 89, 262–268. [CrossRef]
- Lorn, D.; Nguyen, T.-K.-C.; Ho, P.-H.; Tan, R.; Licandro, H.; Waché, Y. Screening of Lactic Acid Bacteria for Their Potential Use as Aromatic Starters in Fermented Vegetables. Int J Food Microbiol 2021, 350, 109242. [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl Glycosidases from Lactobacillus Plantarum Increase Antioxidant Activity of Phenolic Compounds. J Funct Foods 2014, 7, 322–329. [CrossRef]
- Wang, S.-Y.; Zhu, H.-Z.; Lan, Y.-B.; Liu, R.-J.; Liu, Y.-R.; Zhang, B.-L.; Zhu, B.-Q. Modifications of Phenolic Compounds, Biogenic Amines, and Volatile Compounds in Cabernet Gernishct Wine through Malolactic Fermentation by Lactobacillus Plantarum and Oenococcus Oeni. Fermentation 2020, 6, 15. [CrossRef]
- Sestelo, A.B.F.; Poza, M.; Villa, T.G. β-Glucosidase Activity in a Lactobacillus Plantarum Wine Strain. World J Microbiol Biotechnol 2004, 20, 633–637. [CrossRef]
- Brizuela, N.S.; Arnez-Arancibia, M.; Semorile, L.; Pozo-Bayón, M.Á.; Bravo-Ferrada, B.M.; Elizabeth Tymczyszyn, E. β-Glucosidase Activity of Lactiplantibacillus Plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of PH and Ethanol Content. Fermentation 2021, 7. [CrossRef]
- Van Oevelen, D.; Spaepen, M.; Timmermans, P.; Verachtert, H. MICROBIOLOGICAL ASPECTS OF SPONTANEOUS WORT FERMENTATION IN THE PRODUCTION OF LAMBIC AND GUEUZE. Journal of the Institute of Brewing 1977, 83, 356–360. [CrossRef]
- Dysvik, A.; La Rosa, S.L.; De Rouck, G.; Rukke, E.-O.; Westereng, B.; Wicklund, T. Microbial Dynamics in Traditional and Modern Sour Beer Production. Appl Environ Microbiol 2020, 86. [CrossRef]
- Testa, B.; Coppola, F.; Letizia, F.; Albanese, G.; Karaulli, J.; Ruci, M.; Pistillo, M.; Germinara, G.S.; Messia, M.C.; Succi, M.; et al. Versatility of Saccharomyces Cerevisiae 41CM in the Brewery Sector: Use as a Starter for “Ale” and “Lager” Craft Beer Production. Processes 2022, 10. [CrossRef]
- Karaulli, J.; Xhaferaj, N.; Coppola, F.; Testa, B.; Letizia, F.; Kyçyk, O.; Kongoli, R.; Ruci, M.; Lamçe, F.; Sulaj, K.; et al. Bioprospecting of Metschnikowia Pulcherrima Strains, Isolated from a Vineyard Ecosystem, as Novel Starter Cultures for Craft Beer Production. Fermentation 2024, 10, 513. [CrossRef]
- Iorizzo, M.; Letizia, F.; Albanese, G.; Coppola, F.; Gambuti, A.; Testa, B.; Aversano, R.; Forino, M.; Coppola, R. Potential for Lager Beer Production from Saccharomyces Cerevisiae Strains Isolated from the Vineyard Environment. Processes 2021, 9. [CrossRef]
- Bossaert, S.; Crauwels, S.; Lievens, B.; De Rouck, G. The Power of Sour - A Review: Old Traditions, New Opportunities. BrewingScience 2019, 72, 78–88.
- Mahanta, S.; Sivakumar, P.S.; Parhi, P.; Mohapatra, R.K.; Dey, G.; Panda, S.H.; Sireswar, S.; Panda, S.K. Sour Beer Production in India Using a Coculture of Saccharomyces Pastorianus and Lactobacillus Plantarum: Optimization, Microbiological, and Biochemical Profiling. Brazilian Journal of Microbiology 2022, 53, 947–958. [CrossRef]
- Cai, G.; Cao, Y.; Xiao, J.; Sheng, G.; Lu, J. The Mechanisms of Lactiplantibacillus Plantarum J6-6 against Iso-α-Acid Stress and Its Application in Sour Beer Production. Systems Microbiology and Biomanufacturing 2024, 4, 1018–1027. [CrossRef]
- Nyhan, L.; Sahin, A.W.; Arendt, E.K. Co-Fermentation of Non-Saccharomyces Yeasts with Lactiplantibacillus Plantarum FST 1.7 for the Production of Non-Alcoholic Beer. European Food Research and Technology 2023, 249, 167–181. [CrossRef]
- Domínguez-Tornay, A.; Díaz, A.B.; Lasanta, C.; Durán-Guerrero, E.; Castro, R. Co-Fermentation of Lactic Acid Bacteria and S. Cerevisiae for the Production of a Probiotic Beer: Survival and Sensory and Analytical Characterization. Food Biosci 2024, 57. [CrossRef]
- Das, A.J.; Seth, D.; Miyaji, T.; Deka, S.C. Fermentation Optimization for a Probiotic Local Northeastern Indian Rice Beer and Application to Local Cassava and Plantain Beer Production. Journal of the Institute of Brewing 2015, 121, 273–282. [CrossRef]
- Tang, F.; Wei, B.; Qin, C.; Huang, L.; Xia, N.; Teng, J. Enhancing the Inhibitory Activities of Polyphenols in Passion Fruit Peel on α-Amylase and α-Glucosidase via β-Glucosidase-Producing Lactobacillus Fermentation. Food Biosci 2024, 62, 105005. [CrossRef]
- Aron, P.M.; Shellhammer, T.H. A Discussion of Polyphenols in Beer Physical and Flavour Stability. Journal of the Institute of Brewing 2010, 116, 369–380. [CrossRef]
- De Francesco, G.; Bravi, E.; Sanarica, E.; Marconi, O.; Cappelletti, F.; Perretti, G. Effect of Addition of Different Phenolic-Rich Extracts on Beer Flavour Stability. Foods 2020, 9, 1638. [CrossRef]
- Dasenaki, M.E.; Thomaidis, N.S. Quality and Authenticity Control of Fruit Juices-A Review. Molecules 2019, 24, 1014. [CrossRef]
- Plessas, S. Advancements in the Use of Fermented Fruit Juices by Lactic Acid Bacteria as Functional Foods: Prospects and Challenges of Lactiplantibacillus (Lpb.) Plantarum Subsp. Plantarum Application. Fermentation 2021, 8, 6. [CrossRef]
- Montet, D.; Ray, R.C.; Zakhia-Rozis, N. Lactic Acid Fermentation of Vegetables and Fruits; 2014; ISBN 9781482223095.
- Yuan, X.; Wang, T.; Sun, L.; Qiao, Z.; Pan, H.; Zhong, Y.; Zhuang, Y. Recent Advances of Fermented Fruits: A Review on Strains, Fermentation Strategies, and Functional Activities. Food Chem X 2024, 22, 101482. [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and Vegetables, as a Source of Nutritional Compounds and Phytochemicals: Changes in Bioactive Compounds during Lactic Fermentation. Food Research International 2018, 104, 86–99. [CrossRef]
- Gustaw, K.; Niedźwiedź, I.; Rachwał, K.; Polak-Berecka, M. New Insight into Bacterial Interaction with the Matrix of Plant-Based Fermented Foods. Foods 2021, Vol. 10, Page 1603 2021, 10, 1603. [CrossRef]
- Sevindik, O.; Guclu, G.; Agirman, B.; Selli, S.; Kadiroglu, P.; Bordiga, M.; Capanoglu, E.; Kelebek, H. Impacts of Selected Lactic Acid Bacteria Strains on the Aroma and Bioactive Compositions of Fermented Gilaburu (Viburnum Opulus) Juices. Food Chem 2022, 378, 132079. [CrossRef]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of Vegetables and Fruits through Lactic Acid Fermentation. Food Microbiol 2013, 33, 1–10. [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and Functional Paths of Lactic Acid Bacteria in Plant Foods: Get out of the Labyrinth. Curr Opin Biotechnol 2018, 49, 64–72. [CrossRef]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Lactic Acid Fermentation Drives the Optimal Volatile Flavor-Aroma Profile of Pomegranate Juice. Int J Food Microbiol 2017, 248, 56–62. [CrossRef]
- Tripathi, J.; Chatterjee, S.; Gamre, S.; Chattopadhyay, S.; Variyar, P.S.; Sharma, A. Analysis of Free and Bound Aroma Compounds of Pomegranate (Punica Granatum L.). LWT - Food Science and Technology 2014, 59, 461–466. [CrossRef]
- Cele, N.P.; Akinola, S.A.; Manhivi, V.E.; Shoko, T.; Remize, F.; Sivakumar, D. Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation. Foods 2022, 11, 682. [CrossRef]
- Surendran, S.; Qassadi, F.; Surendran, G.; Lilley, D.; Heinrich, M. Myrcene—What Are the Potential Health Benefits of This Flavouring and Aroma Agent? Front Nutr 2021, 8. [CrossRef]
- Sun, J.; Zhao, C.; Pu, X.; Li, T.; Shi, X.; Wang, B.; Cheng, W. Flavor and Functional Analysis of Lactobacillus Plantarum Fermented Apricot Juice. Fermentation 2022, 8, 533. [CrossRef]
- Gao, H.; Wen, J.-J.; Hu, J.-L.; Nie, Q.-X.; Chen, H.-H.; Nie, S.-P.; Xiong, T.; Xie, M.-Y. Momordica Charantia Juice with Lactobacillus Plantarum Fermentation: Chemical Composition, Antioxidant Properties and Aroma Profile. Food Biosci 2019, 29, 62–72. [CrossRef]
- Mrabti, H.N.; Jaouadi, I.; Zeouk, I.; Ghchime, R.; El Menyiy, N.; Omari, N. El; Balahbib, A.; Al-Mijalli, S.H.; Abdallah, E.M.; El-Shazly, M.; et al. Biological and Pharmacological Properties of Myrtenol: A Review. Curr Pharm Des 2023, 29, 407–414. [CrossRef]
- Ricci, A.; Cirlini, M.; Levante, A.; Dall’Asta, C.; Galaverna, G.; Lazzi, C. Volatile Profile of Elderberry Juice: Effect of Lactic Acid Fermentation Using L. Plantarum, L. Rhamnosus and L. Casei Strains. Food Research International 2018, 105, 412–422. [CrossRef]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; De Angelis, M.; Salmon, J.-C.; Buchin, S.; De Gara, L.; Gobbetti, M. Effect of Autochthonous Lactic Acid Bacteria Starters on Health-Promoting and Sensory Properties of Tomato Juices. Int J Food Microbiol 2009, 128, 473–483. [CrossRef]
- Di Cagno, R.; Surico, R.F.; Minervini, G.; Rizzello, C.G.; Lovino, R.; Servili, M.; Taticchi, A.; Urbani, S.; Gobbetti, M. Exploitation of Sweet Cherry (Prunus Avium L.) Puree Added of Stem Infusion through Fermentation by Selected Autochthonous Lactic Acid Bacteria. Food Microbiol 2011, 28, 900–909. [CrossRef]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic Responses of Lactobacillus Plantarum Strains during Fermentation and Storage of Vegetable and Fruit Juices. Appl Environ Microbiol 2014, 80, 2206–2215. [CrossRef]
- Hashemi, S.M.B.; Mousavi Khaneghah, A.; Barba, F.J.; Nemati, Z.; Sohrabi Shokofti, S.; Alizadeh, F. Fermented Sweet Lemon Juice (Citrus Limetta) Using Lactobacillus Plantarum LS5: Chemical Composition, Antioxidant and Antibacterial Activities. J Funct Foods 2017, 38, 409–414. [CrossRef]
- Wang, H.; He, X.; Li, J.; Wu, J.; Jiang, S.; Xue, H.; Zhang, J.; Jha, R.; Wang, R. Lactic Acid Bacteria Fermentation Improves Physicochemical Properties, Bioactivity, and Metabolic Profiles of Opuntia Ficus-Indica Fruit Juice. Food Chem 2024, 453, 139646. [CrossRef]
- Mashitoa, F.M.; Manhivi, V.E.; Akinola, S.A.; Garcia, C.; Remize, F.; Shoko, T.; Sivakumar, D. Changes in Phenolics and Antioxidant Capacity during Fermentation and Simulated in Vitro Digestion of Mango Puree Fermented with Different Lactic Acid Bacteria. J Food Process Preserv 2021, 45. [CrossRef]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, Metabolism and Bioavailability of Flavonoids: A Review. Crit Rev Food Sci Nutr 2022, 62, 7730–7742. [CrossRef]
- Tang, S.; Cheng, Y.; Wu, T.; Hu, F.; Pan, S.; Xu, X. Effect of Lactobacillus Plantarum-Fermented Mulberry Pomace on Antioxidant Properties and Fecal Microbial Community. LWT 2021, 147, 111651. [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of Fermentation on the Antioxidant Activity in Plant-Based Foods. Food Chem 2014, 160, 346–356. [CrossRef]
- Slámová, K.; Kapešová, J.; Valentová, K. “Sweet Flavonoids”: Glycosidase-Catalyzed Modifications. Int J Mol Sci 2018, 19. [CrossRef]
- Gaur, G.; Gänzle, M.G. Conversion of (Poly)Phenolic Compounds in Food Fermentations by Lactic Acid Bacteria: Novel Insights into Metabolic Pathways and Functional Metabolites. Curr Res Food Sci 2023, 6, 100448. [CrossRef]
- Meng, F.-B.; Lei, Y.-T.; Li, Q.-Z.; Li, Y.-C.; Deng, Y.; Liu, D.-Y. Effect of Lactobacillus Plantarum and Lactobacillus Acidophilus Fermentation on Antioxidant Activity and Metabolomic Profiles of Loquat Juice. LWT 2022, 171, 114104. [CrossRef]
- Arunachalam, S.; Nagoor Meeran, M.F.; Azimullah, S.; Sharma, C.; Goyal, S.N.; Ojha, S. Nerolidol Attenuates Oxidative Stress, Inflammation, and Apoptosis by Modulating Nrf2/MAPK Signaling Pathways in Doxorubicin-Induced Acute Cardiotoxicity in Rats. Antioxidants (Basel) 2021, 10. [CrossRef]
- Iqubal, A.; Syed, M.A.; Ali, J.; Najmi, A.K.; Haque, M.M.; Haque, S.E. Nerolidol Protects the Liver against Cyclophosphamide-Induced Hepatic Inflammation, Apoptosis, and Fibrosis via Modulation of Nrf2, NF-ΚB P65, and Caspase-3 Signaling Molecules in Swiss Albino Mice. Biofactors 2020, 46, 963–973. [CrossRef]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y.; Wang, M. Enhanced Antioxidant Activity for Apple Juice Fermented with Lactobacillus Plantarum ATCC14917. Molecules 2018, 24, 51. [CrossRef]
- Chen, C.; Lu, Y.; Yu, H.; Chen, Z.; Tian, H. Influence of 4 Lactic Acid Bacteria on the Flavor Profile of Fermented Apple Juice. Food Biosci 2019, 27, 30–36. [CrossRef]
- Peng, W.; Meng, D.; Yue, T.; Wang, Z.; Gao, Z. Effect of the Apple Cultivar on Cloudy Apple Juice Fermented by a Mixture of Lactobacillus Acidophilus, Lactobacillus Plantarum, and Lactobacillus Fermentum. Food Chem 2021, 340, 127922. [CrossRef]
- Hashemi, S.M.B.; Jafarpour, D. Fermentation of Bergamot Juice with Lactobacillus Plantarum Strains in Pure and Mixed Fermentations: Chemical Composition, Antioxidant Activity and Sensorial Properties. LWT 2020, 131, 109803. [CrossRef]
- Tkacz, K.; Chmielewska, J.; Turkiewicz, I.P.; Nowicka, P.; Wojdyło, A. Dynamics of Changes in Organic Acids, Sugars and Phenolic Compounds and Antioxidant Activity of Sea Buckthorn and Sea Buckthorn-Apple Juices during Malolactic Fermentation. Food Chem 2020, 332, 127382. [CrossRef]
- Filannino, P.; Cavoski, I.; Thlien, N.; Vincentini, O.; De Angelis, M.; Silano, M.; Gobbetti, M.; Di Cagno, R. Lactic Acid Fermentation of Cactus Cladodes (Opuntia Ficus-Indica L.) Generates Flavonoid Derivatives with Antioxidant and Anti-Inflammatory Properties. PLoS One 2016, 11, e0152575. [CrossRef]
- Wang, J.; Wei, B.-C.; Wang, X.; Zhang, Y.; Gong, Y.-J. Aroma Profiles of Sweet Cherry Juice Fermented by Different Lactic Acid Bacteria Determined through Integrated Analysis of Electronic Nose and Gas Chromatography–Ion Mobility Spectrometry. Front Microbiol 2023, 14. [CrossRef]
- Kuerban, D.; Lu, J.; Huangfu, Z.; Wang, L.; Qin, Y.; Zhang, M. Optimization of Fermentation Conditions and Metabolite Profiling of Grape Juice Fermented with Lactic Acid Bacteria for Improved Flavor and Bioactivity. Foods 2023, 12, 2407. [CrossRef]
- Huang, Y.; Wang, H.; Zhu, C. Effect of Lactic Acid Bacteria Fermentation on Antioxidation and Bioactivity of Hawthorn Pulp. IOP Conf Ser Earth Environ Sci 2019, 267, 062056. [CrossRef]
- Pan, X.; Zhang, S.; Xu, X.; Lao, F.; Wu, J. Volatile and Non-Volatile Profiles in Jujube Pulp Co-Fermented with Lactic Acid Bacteria. LWT 2022, 154, 112772. [CrossRef]
- Li, T.; Jiang, T.; Liu, N.; Wu, C.; Xu, H.; Lei, H. Biotransformation of Phenolic Profiles and Improvement of Antioxidant Capacities in Jujube Juice by Select Lactic Acid Bacteria. Food Chem 2021, 339, 127859. [CrossRef]
- Zhao, M.; Zhang, F.; Zhang, L.; Liu, B.; Meng, X. Mixed Fermentation of Jujube Juice ( Ziziphus Jujuba Mill.) with L. Rhamnosus <scp>GG</Scp> and L. Plantarum -1: Effects on the Quality and Stability. Int J Food Sci Technol 2019, 54, 2624–2631. [CrossRef]
- Wang, D.; Deng, Y.; Chen, X.; Wang, K.; Zhao, L.; Wang, Z.; Liu, X.; Hu, Z. Elucidating the Effects of Lactobacillus Plantarum Fermentation on the Aroma Profiles of Pasteurized Litchi Juice Using Multi-Scale Molecular Sensory Science. Curr Res Food Sci 2023, 6, 100481. [CrossRef]
- Liu, S.; Peng, Y.-J.; He, W.-W.; Song, X.-X.; He, Y.-X.; Hu, X.-Y.; Bian, S.-G.; Li, Y.-H.; Yin, J.-Y.; Nie, S.-P.; et al. Metabolomics-Based Mechanistic Insights into Antioxidant Enhancement in Mango Juice Fermented by Various Lactic Acid Bacteria. Food Chem 2025, 466, 142078. [CrossRef]
- Park, J.B.; Lim, S.H.; Sim, H.S.; Park, J.H.; Kwon, H.J.; Nam, H.S.; Kim, M.D.; Baek, H.H.; Ha, S.J. Changes in Antioxidant Activities and Volatile Compounds of Mixed Berry Juice through Fermentation by Lactic Acid Bacteria. Food Sci Biotechnol 2017, 26, 441–446. [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem 2018, 250, 148–154. [CrossRef]
- Yaqoob, S.; Imtiaz, A.; Awan, K.A.; Murtaza, M.S.; Mubeen, B.; Yinka, A.A.; Boasiako, T.A.; Alsulami, T.; Rehman, A.; Khalifa, I.; et al. Impact of Fermentation through Synergistic Effect of Different Lactic Acid Bacteria (Mono and Co-Cultures) on Metabolic and Sensorial Profile of Mulberry Juice. Journal of Food Measurement and Characterization 2024. [CrossRef]
- de la Fuente, B.; Luz, C.; Puchol, C.; Meca, G.; Barba, F.J. Evaluation of Fermentation Assisted by Lactobacillus Brevis POM, and Lactobacillus Plantarum (TR-7, TR-71, TR-14) on Antioxidant Compounds and Organic Acids of an Orange Juice-Milk Based Beverage. Food Chem 2021, 343, 128414. [CrossRef]
- Dogan, K.; Akman, P.K.; Tornuk, F. Role of Non-thermal Treatments and Fermentation with Probiotic Lactobacillus Plantarum on in Vitro Bioaccessibility of Bioactives from Vegetable Juice. J Sci food Agric 2021, 101, 4779–4788. [CrossRef]
- Fonseca, H.C.; Melo, D. de S.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Lactiplantibacillus Plantarum CCMA 0743 and Lacticaseibacillus Paracasei Subsp. Paracasei LBC-81 Metabolism during the Single and Mixed Fermentation of Tropical Fruit Juices. Brazilian Journal of Microbiology 2021, 52, 2307–2317. [CrossRef]
- Mantzourani, I.; Kazakos, S.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Bekatorou, A.; Plessas, S. Potential of the Probiotic Lactobacillus Plantarum ATCC 14917 Strain to Produce Functional Fermented Pomegranate Juice. Foods 2019, Vol. 8, Page 4 2018, 8, 4. [CrossRef]
- Vivek, K.; Mishra, S.; Pradhan, R.C.; Jayabalan, R. Effect of Probiotification with Lactobacillus Plantarum MCC 2974 on Quality of Sohiong Juice. LWT 2019, 108, 55–60. [CrossRef]
- Liu, S.; He, Y.; He, W.; Song, X.; Peng, Y.; Hu, X.; Bian, S.; Li, Y.; Nie, S.; Yin, J.; et al. Exploring the Biogenic Transformation Mechanism of Polyphenols by Lactobacillus Plantarum NCU137 Fermentation and Its Enhancement of Antioxidant Properties in Wolfberry Juice. J Agric Food Chem 2024, 72, 12752–12761. [CrossRef]
- Tian, Y.; Wang, Y.; Zhang, N.; Xiao, M.; Zhang, J.; Xing, X.; Zhang, Y.; Fan, Y.; Li, X.; Nan, B.; et al. Antioxidant Mechanism of Lactiplantibacillus Plantarum KM1 Under H2O2 Stress by Proteomics Analysis. Front Microbiol 2022, 13. [CrossRef]
- Markkinen, N.; Laaksonen, O.; Nahku, R.; Kuldjärv, R.; Yang, B. Impact of Lactic Acid Fermentation on Acids, Sugars, and Phenolic Compounds in Black Chokeberry and Sea Buckthorn Juices. Food Chem 2019, 286, 204–215. [CrossRef]
- Wei, M.; Wang, S.; Gu, P.; Ouyang, X.; Liu, S.; Li, Y.; Zhang, B.; Zhu, B. Comparison of Physicochemical Indexes, Amino Acids, Phenolic Compounds and Volatile Compounds in Bog Bilberry Juice Fermented by Lactobacillus Plantarum under Different PH Conditions. J Food Sci Technol 2018, 55, 2240–2250. [CrossRef]
- Yilmaz, B.; Bangar, S.P.; Echegaray, N.; Suri, S.; Tomasevic, I.; Lorenzo, J.M.; Melekoglu, E.; Rocha, J.M.; Ozogul, F. The Impacts of Lactiplantibacillus Plantarum on the Functional Properties of Fermented Foods: A Review of Current Knowledge. Microorganisms 2022, 10. [CrossRef]
Figure 1.
Release of free aroma compounds from glycosidically bound precursors by β-glucosidase activity. Representation of enzymatic activities involved in the release of the volatile aglycone component from three different disaccharides-bound aroma compounds.
Figure 1.
Release of free aroma compounds from glycosidically bound precursors by β-glucosidase activity. Representation of enzymatic activities involved in the release of the volatile aglycone component from three different disaccharides-bound aroma compounds.
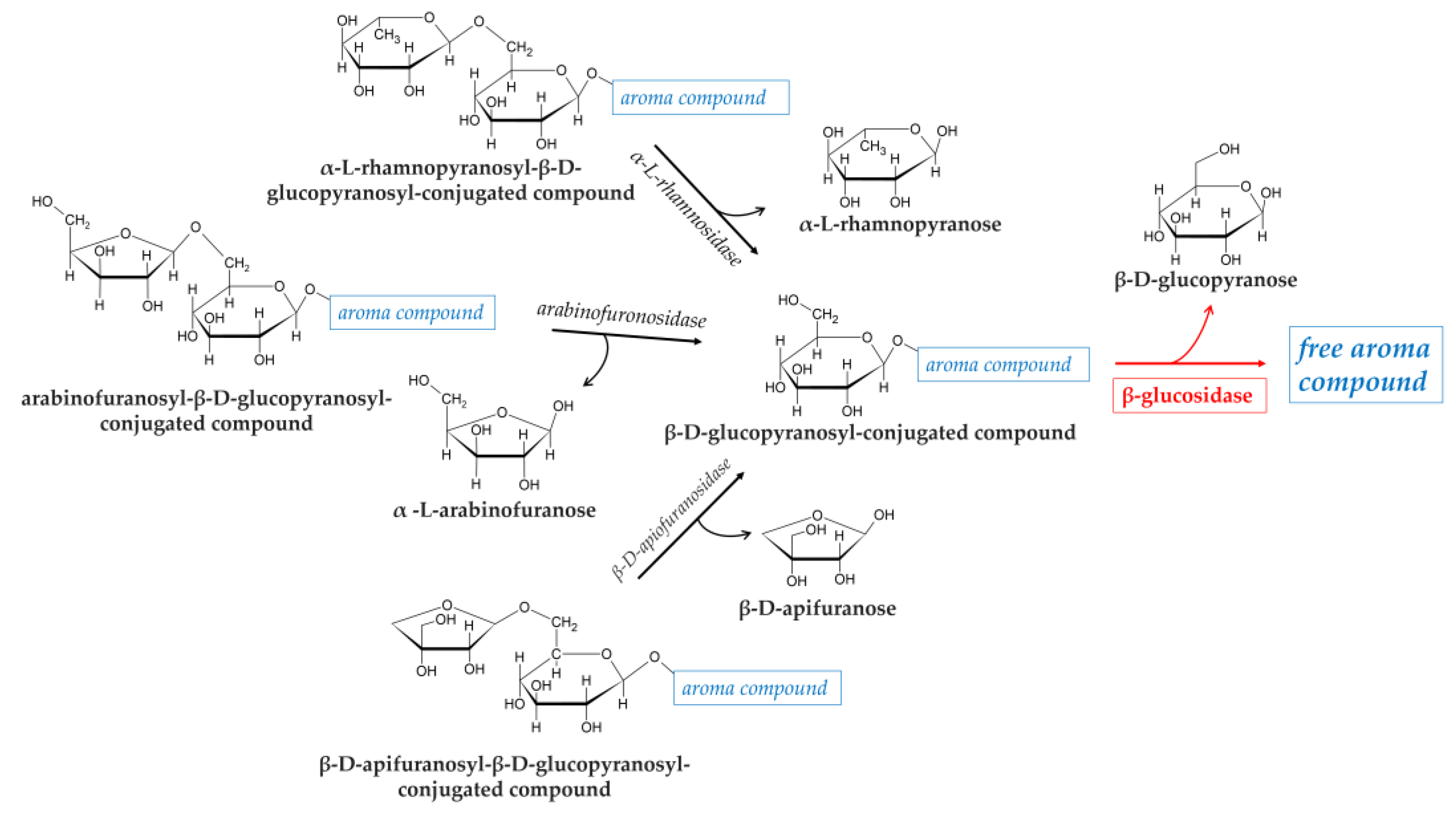
Figure 2.
Release of free isoflavone from the three main soy isoflavones glycosides and their acetylated/malonylated forms. Representation of the two step-process by which the acetyl- and malonyl- forms of soy glycosilated isofavones are hydrolized by esterase and β-glucosidase to release the free form of isoflavone.
Figure 2.
Release of free isoflavone from the three main soy isoflavones glycosides and their acetylated/malonylated forms. Representation of the two step-process by which the acetyl- and malonyl- forms of soy glycosilated isofavones are hydrolized by esterase and β-glucosidase to release the free form of isoflavone.
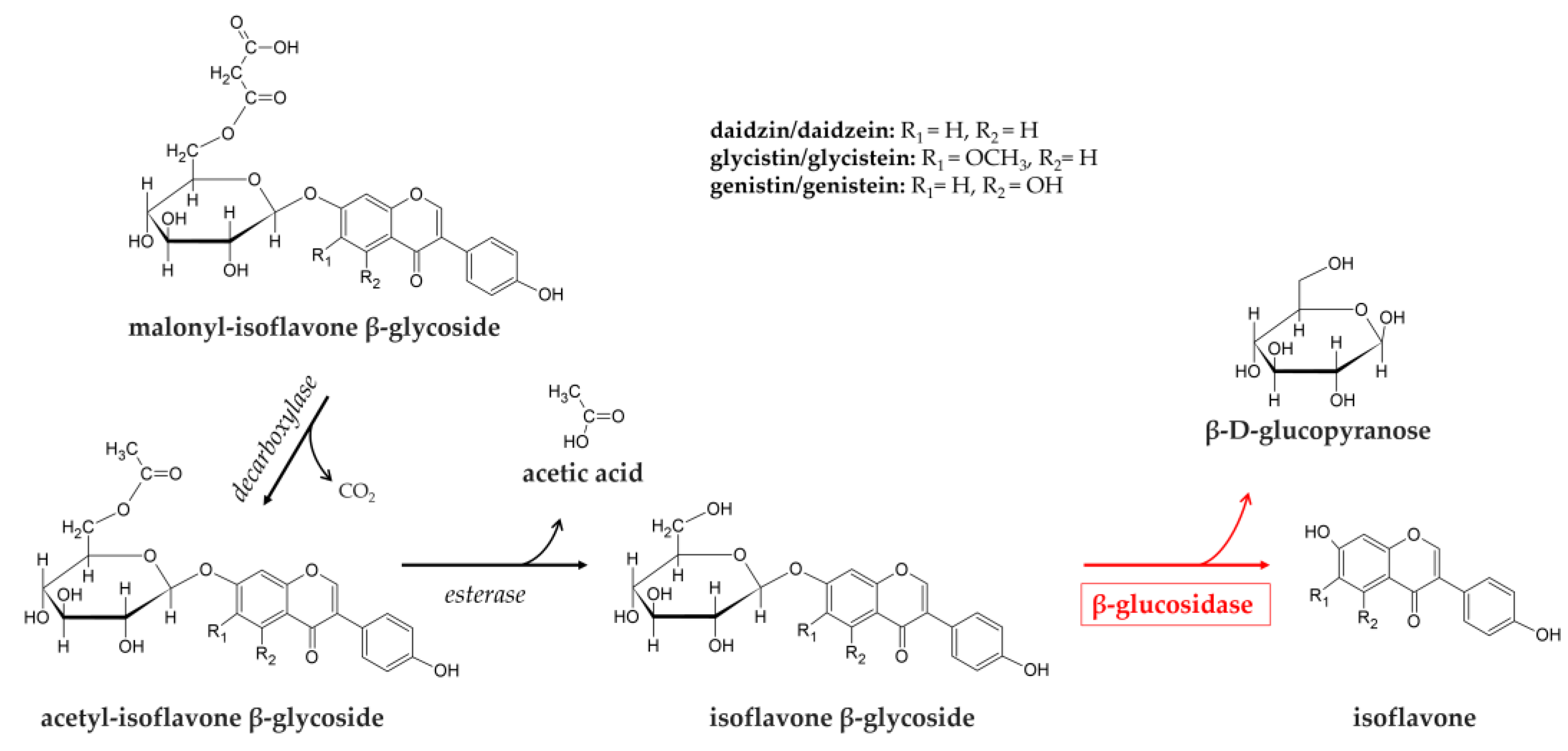
Figure 3.
Cyanidric acid generation by cyanogenic glycosides present in cassava tissues. Degradation of the two principal cyanogenic glycosides of cassava by β-glucosidase activity with production of glucose and subsequent release of cyanidric acid. Targets of this high toxic compound are haemoglobin of erythocytes and cytochrome c oxidase, the complex IV of the respiratory chain embedded in the inner membrane of the mitochondrion. Abbreviations: m.o.m., mitochondrial outer membrane, i.s., intermembrane space; m.i.m., mitochondrial inner membrane.
Figure 3.
Cyanidric acid generation by cyanogenic glycosides present in cassava tissues. Degradation of the two principal cyanogenic glycosides of cassava by β-glucosidase activity with production of glucose and subsequent release of cyanidric acid. Targets of this high toxic compound are haemoglobin of erythocytes and cytochrome c oxidase, the complex IV of the respiratory chain embedded in the inner membrane of the mitochondrion. Abbreviations: m.o.m., mitochondrial outer membrane, i.s., intermembrane space; m.i.m., mitochondrial inner membrane.
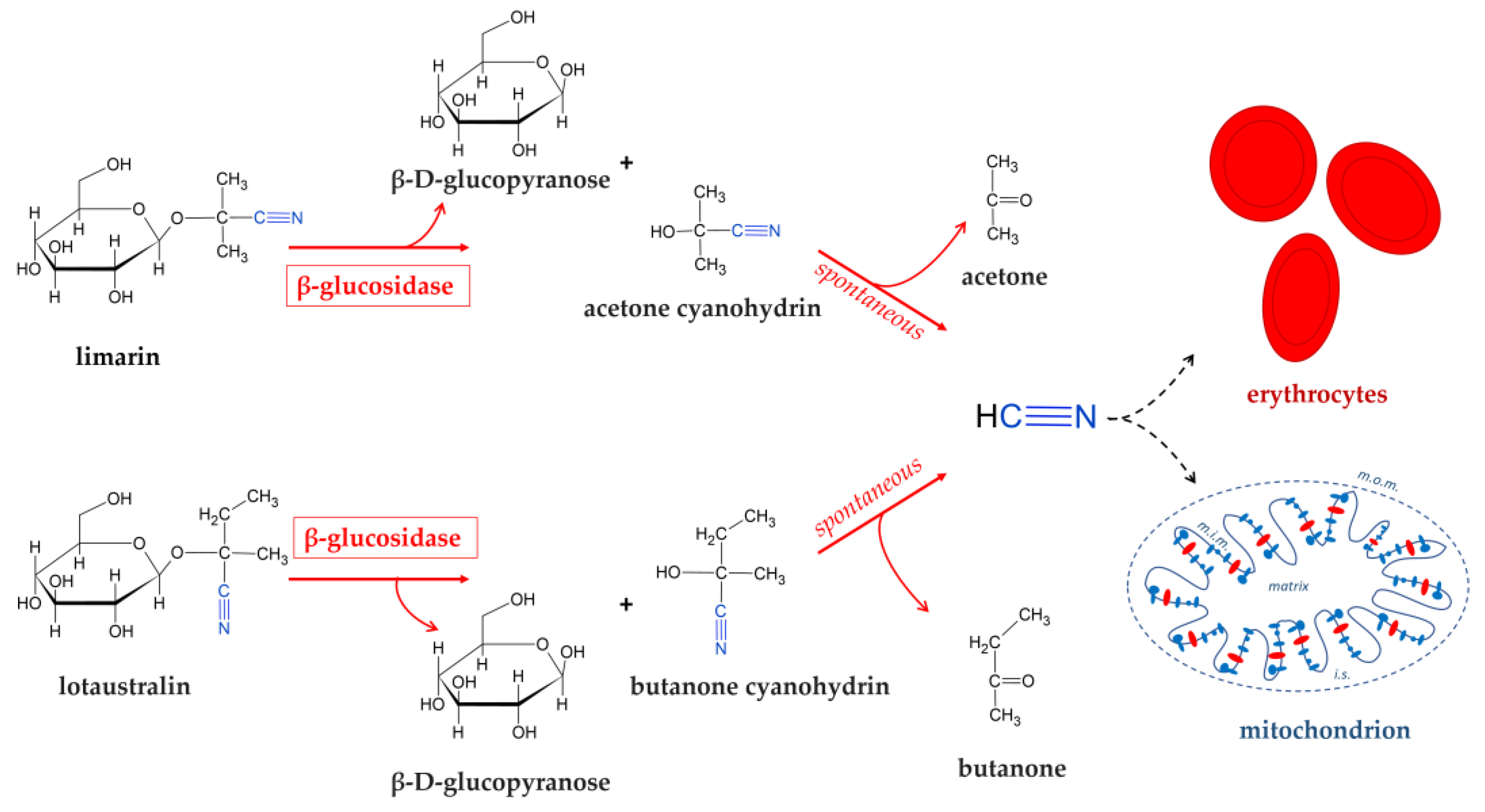

Table 1.
Main positive effects potentially related to β-glucosidase activity of L. plantarum in fermented fruit.
Table 1.
Main positive effects potentially related to β-glucosidase activity of L. plantarum in fermented fruit.
| Fruit processed | Product type | L. plantarum (Lp) strains | Main Positive Effects | Ref |
|---|---|---|---|---|
| Apple | Juice fermented at 37°C for 72 h. | Lp ATCC14917 | Increased antioxidant activity and decreased total phenolics and flavonoid content. | [218] |
| Apple | Juice fermented at 37°C for 80 h | Lp ST-III | Improved flavor profile | [219] |
| Apple | Single Juices from nine apple cultivars fermented at 37°C for 24 h | Lp CICC21805 | Increased terpenes D-limonene and eugenol in some apple cultivars | [220] |
| Apricot | Juice fermented at 37°C for 12 h | Lp LP56 | Increased antioxidant activity and total phenolics; improved flavor profile | [200] |
| Bergamot (Citrus Bergamia Risso) | Juice fermented at 37°C for 72 h | Single and mixed starter: Lp PTCC 1896 Lp AF1 Lp LP3 |
Increased antioxidant activity | [221] |
| Buckthorn berries (Hippophaë rhamnoides L.) |
Juice fermented at 30°C for 72 h |
Lp DSM 10492, Lp DSM 20174 Lp DSM 6872 |
Increased antioxidant activity and flavonoids | [222] |
| Cactus (Opuntia ficus-indica L.) | Cladodes pulp fermented at 30°C for 24 h |
Single starters: Lp CIL6 Lp POM1 Lp 1MR20 |
Increased antioxidant activity and flavonoids (kaemferol and isorhamnetin) | [223] |
| Cherries (Prunus avium L.) | Juice fermented at 37°C for 48 h | Lp JYLP-375 | Improved flavor profile | [224] |
| Cranberrybush/Gilaburu (Viburnum opulus L.) | Juice fermented at 30°C for 12 days | Lp-23 | Increased antioxidant activity and terpenes | [193] |
| Elderberry (Sambucus nigra L.) | Juice fermented at 37°C for 48 h | Single starters: Lp POM1 Lp 1LE1 Lp C1 Lp 1486 Lp 285 |
Increase of terpenes and norisoprenoids (limonene, β-linalool, β-damascenone and eugenol) | [203] |
| Grapes | Juice fermented at 37°C for 32 h | Single and mixed starter: Lp 90 L. helveticus 76 L. casei |
Increased total phenolics and improved flavor profile | [225] |
| Hawthorn (Crataegus pinnatifida) | Pulp fermented at 37°C for 12 h | Mixed starter: Lp, Lactobacillus acidophilus and Lactobacillus casei. | Increased total phenolics and flavonoids | [226] |
| Jujube (Ziziphus jujuba Milll.) | Pulp fermented at 37°C for 24 h | Lp CICC 20265 | Improved flavor profile | [227] |
| Jujube (Zizyphus jujuba Mill.) | Juice fermented at 37°C for 48 h | Lp 90 | Increased antioxidant activity and flavor profile | [228] |
| Jujube (Ziziphus jujuba Milll.) | Juice fermented at 37°C for 28 h | Single and mixed starter: L. rhamnosus GG, Lp-1 Lp-2 L. paracasei 22709 L. mesenteroides 22264 |
Decreased total phenolics and increased total flavonoid content; improved flavor profile |
[229] |
| Lemon (Citrus limetta) | Juice fermented at 37°C for 48 h | Lp LS5 | Increased antioxidant activity | [207] |
| Litchi (Litchi chinensis Sonn. | Juice fermented at 37 °C for 40 h | Single starters: Lp LP28 Lp LP226 Lp LPC2W |
Increased terpenes citronellol, linalool, geraniol, prenol | [230] |
| Loquat (Eriobotrya japonica Lindl.) | Juice fermented at 36°C for 48 h | Lp LZ 2-2 | Increased antioxidant activity, total phenolics and total flavonoids | [215] |
| Mango (Mangifera indica L.) | Juice fermented at 37°C for 48 h | Lp NCU116 | Increased antioxidant activity and total phenolics | [231] |
| Mango (Mangifera indica L.) | Juice fermented at 30°C for 72 h | Single and mixed starter Lp L75, Leuconostoc pseudomesenteroides L 56 |
Increased antioxidant activity and improved flavor profile | [198] |
| Mixed berry (acai berry, aronia, cranberry) | Juice fermented at 37°C for 36 h | Lp LP-115 | Increased antioxidant activity | [232] |
| Momordica charantia L. | Juice fermented at 37°C for 48 h | Lp NCU116 | Increased antioxidant activity, total phenolics and total flavonoids | [201] |
|
Mulberry (Morus nigra) |
Juice fermented at 37°C for 36 h | Lp ATCC SD5209 | Increased antioxidant activity and phenolics (phenolic acids, anthocyanins and flavonols) | [233] |
|
Mulberry (Morus nigra) |
Juice fermented at 37°C for 7 days | Lp CICC 20265 | Increased antioxidant activity | [211] |
|
Mulberry (Morus nigra) |
Juice fermented at 37°C for 48 h | Lp (single colture and/or in co-colture with other LAB) | Improvement of both nutritional and aromatic profile | [234] |
| Orange | Juice-milk fermented at 37 °C for 72 h | Single starters: Lp TR-7 Lp TR-71 Lp TR-14 |
Increased antioxidant activity and total phenolics | [235] |
| Orange, lemon, celery and carrot | Mixed vegetable juice fermented at 37°C for 24 h | Lp HFC8 | Increased antioxidant activity and phenolics (flavonoids, and anthocyanins) | [236] |
| Passion fruit (Passiflora edulis), acerola (Malpighia emarginata), and jelly palm (Butia capitata) | Juice fermented at 37°C for 24 h | Lp CCMA 0743 | Increased flavonoids | [237] |
| Pomegranate (Punica granatum L.) | Juice fermented at 30°C for 24 h | Lp ATCC 14917 | Increased antioxidant activity and total phenolics | [238] |
| Pomegranate (Punica granatum L.) | Juice fermented at 30°C for 120 h | Single starter: Lp C2 Lp POM1 |
Improved flavor profile | [196] |
| Sohiong (Prunus nepalensis) | Juice fermented at 37°C for 72 h | Lp MCC 297 | Increased antioxidant activity, total phenolics, and anthocyanins | [239] |
| Wolfberry | Juice fermented at 37°C for 48 h | Lp NCU137 | Increased antioxidant activity and free phenolics | [240] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



