Submitted:
19 March 2025
Posted:
19 March 2025
You are already at the latest version
Abstract
Stone fruits (Prunus spp.) occupy a pivotal position in global fruit production due to their significant nutritional profile and distinctive organoleptic characteristics. Contemporary orchard systems are undergoing transformation through innovative cultivation approaches, notably high-density dwarfing systems, greenhouse cultivation, agri-tech integration, and simplified management. As a crucial agronomic component in modern stone fruit cultivation, rootstock systems confer multi-benefits including enhanced environmental resilience, improved scion productivity, superior fruit quality, controlled vigor and dwarfing capacity. While the majority of european apple orchards have transitioned to dwarfing rootstock systems, achieving substantial gains in productivity and profitability, stone fruits cultivation lag significantly due to the key gaps in prunus rootstock development include genetic complexity, extended evaluation cycles, clonal propagation barriers and limited research programs. Urgent innovation is required to address these challenges in rootstock breeding to meet the demand of sustainable stone fruits production. This review systematically examines strategic breeding objectives and innovative molecular methodologies in Prunus rootstock development, with particular emphasis on marker-assisted selection and genomic prediction technologies. We provide a comprehensive synthesis of breeding achievements across major commercial rootstock cultivars, while proposing forward-looking research strategies incorporating CRISPR-based genome editing and multi-omics approaches. The synthesized insights establish a theoretical pathway for advancing rootstock genetic improvement and sustainable orchard management practices in stone fruit cultivation systems.
Keywords:
stone fruits
; rootstock
; breeding objectives
; breeding techniques
; breeding achievements
; strategies
1. Introduction
Stone fruits, mainly encompassing sweet cherry (Prunus avium), sour cherry (P. cerasus), plums (P. domestica and P. salicina), peaches (P. persica), almonds (P. dulcis), and apricots (P. armeniaca) [1], constitute a vital horticultural group characterized by nutritional richness and organoleptic appeal. Beyond fresh consumption, their transformation into value-added products including jams, juices, dried fruits, and various desserts drives substantial global demand. In 2022, the global export value of stone fruit crops exceeded 10 billion dollars, representing an approximate 47% increase compared to the previous decade [2]. Geospatial production patterns demonstrate concentration within temperate zones between 30° and 40° latitude north latitude, where China maintains dominant producer status with accounting for approximately 50% of the global total output (Figure 1) [2]. Due to strong economic returns, the global production of stone fruits has increased by 15% over the past decade. However, intensive land requirements and escalating labor inputs increasingly compromise grower profitability. In addition, the demand trend in the stone fruit market has shifted from pursuing high yield to focusing on quality. Therefore, this dual pressure necessitates urgent implementation of sustainable intensification strategies through precision horticulture and genetic optimization to reconcile productivity with premium quality standards in stone fruit cultivation systems.
2. Modern Trends in Fruit Tree Cultivation Regime and the Demand for Advanced Rootstock Breeding
The global fruit production industry has undergone transformative changes in cultivation practices over the past century, driven by the need for higher productivity, resource efficiency, and labor optimization. Key trends in modern fruit cultivation regime such as high-density dwarfing systems, protected cultivation, agri-tech integration, and simplified management protocols are reshaping orchard systems [3]. These models achieve efficient fruit cultivation and promote economic development by controlling tree height, regulating the environment, improving soil conditions, introducing mechanized operations, and building diverse ecosystems. Among them, the implementation of dwarfing and dense planting not only directly reduces land use and facilitates manual harvesting but also makes it possible to construct greenhouses and introduce mechanized harvesting equipment. It has become the mainstream model of modern fruit tree cultivation. The dwarfing and dense planting of apples is a successful case, with its core being the M series selected by the East Malling Research Station of dwarfing rootstocks and their derived varieties (such as the P series from Poland, the B series from the Russia, and the G series from the United States). For example, rootstocks like ‘B.9’, ‘M.27’ and ‘M.65’ can reduce tree vigor by 50%, thereby significantly saving land resources and facilitating mechanized management and harvesting [4]. In addition, rootstocks can also eliminate or mitigate the limitations of various soil conditions, enhance tolerance to drought, waterlogging, salinity, and nutrient deficiency, provide resistance to viruses, bacteria, fungi, and pests, and improve yield and fruit quality. Rootstocks have become an indispensable component in the modern cultivation mechanisms of fruit trees. While apple cultivation has achieved remarkable success through rootstock innovation, significant gaps persist in stone fruit rootstock development, reflecting both biological complexities and systemic research challenges. Fortunately, emerging biotechnologies and collaborative frameworks offer unprecedented opportunities to develop next-generation rootstocks. Prioritizing rootstock breeding is critical for climate-resilient, sustainable stone fruit production.
3. Breeding Objectives for Rootstocks of Stone Fruits
In fruit tree cultivation, an ideal rootstock should possess the ability to be easily propagated asexually, have strong environmental adaptability, exhibit strong compatibility with grafting, and effectively improve the growth of scions and enhance fruit quality. These are also the main breeding objectives for rootstocks of stone fruit trees.
3.1. Efficient Clonal Propagation
Clonal rootstocks, due to their identical genotypes, exhibit minimal genetic variation among individuals, which allows for the maximum maintenance of uniform scion traits, thereby stabilizing yield and quality. In production, there are often potential rootstock varieties that possess multiple desirable traits but are unable to be propagated asexually on a large scale due to difficulties in adventitious root formation. Therefore, evaluating the adventitious root-forming ability of rootstocks is an important aspect of rootstock breeding. When breeding stone fruit rootstocks with ease of asexual reproduction as the primary selection trait, screening from a large pool of materials with rich genetic variation is a viable approach. Korkmaz et al. [5] compared the adventitious root formation ability of 79 plum rootstock cuttings, selecting 39 superior materials based on a threshold of root length of 33.50 mm, root number of 3.00, and rooting rate of 30.00%. Evaluating the rooting ability of rootstocks with other desirable traits is a method for further selection. A comparison regarding the adventitious root formation ability and quality of 30 rootstocks potentially tolerant to peach tree short life syndrome (PTSL) showed that ‘GL-ERA-09-33’ (P. persica) had the best rooting efficiency, making it an excellent rootstock for tolerance to PTSL [6]. Furthermore, an evaluation conducted by Rosa et al. [7] on the adventitious root formation ability of semi - hardwood cuttings from four Prunus rootstocks—‘Okinawa’ (P. persica), ‘Tsukuba-1’ (P. persica), ‘Julior’ (P. insititia × P. domestica), and ‘Mirabolano 29C’ (P. cerasifera); it was found that ‘Mirabolano 29-C’ had a rooting rate exceeding 80% with the highest number and quality of adventitious roots. Among five Prunus rootstocks (‘Adafuel’ (P. dulcis × P. persica), ‘Adarcias’ (P. dulcis × P. persica), ‘Cadaman’ (P. persica × P. davidiana), ‘Garnem’ (P. dulcis × P. persica), and ‘GF 677’ (P. dulcis × P. persica)), it was observed that the hardwood cuttings of ‘Garnem’ showed the best adventitious root formation ability, while ‘GF677’ and ‘Cadaman’ were less likely to root [8]. The difficulty in rooting ‘Cadaman’ is associated with a lower ratio of carbohydrates to nitrogen [9], while the difficulty in adventitious root formation for ‘GF677’ may be due to its higher activities of putrescine, polyamine oxidase, and catalase, as well as lower activity of diamine oxidase [10].
3.2. Abiotic Stress Tolerance
Stone fruit trees exhibit a rich diversity of species and demonstrate remarkable adaptability to a wide range of environments. However, they often face various environmental stresses, including drought, waterlogging, soil salinization, and nutrient deficiencies. To mitigate these challenges, breeders have undertaken extensive screening efforts to select rootstocks with enhanced environmental adaptability. This review synthesizes current research progress in these key areas.
Temperature Stress: Temperature is a crucial factor affecting the growth of stone fruits. Differences in temperature due to geographical latitude directly limit the ecological distribution of these fruits. Within suitable habitats, excessively high temperatures in summer or excessively low temperatures in winter can cause growth stagnation or even death of these plants. Therefore, in the context of global climate change, the breeding of heat-tolerant and cold-tolerant rootstocks is of significant importance for maintaining the normal growth of stone fruits and expanding their cultivation areas. The cold tolerance of 48 cherry hybrid rootstocks was tested by Blážková et al. [11], using ‘Colt’ (P. avium × P. pseudocerasus) as a control, and they identified ‘CPHVODÁRNA,’ ‘CPH 43,’ ‘CPH 17,’ ‘CPH 22,’ and ‘CPH 49’ as new cold - tolerant rootstocks. Another study comparing the growth and physiological state of scions grafted onto cherry rootstocks ‘Gisela 5’ (P.cerasus × P.canescens) and ‘Mazzard’ (P. avium) under cold adaptation conditions showed that ‘Mazzard’ was better able to maintain fruit sugar metabolism and had higher soluble sugar content, making it a more cold - tolerant cherry rootstock [12]. In addition, the response of seven Prunus rootstocks (‘Garnem,’ ‘GF 677’, ‘Marianna 2624’ (P. cerasifera × P. musonianna), ‘Myrobolan 29C’ (P. cerasifera), ‘Rootpac 20’ (P. besseyi × P. cerasifera), ‘Rootpac 40’ ((P. amygdalus × P. persica) × (P. amygdalus × P. persica)), and ‘Rootpac R’) to drought, heat shock, and their combination revealed that ‘Rootpac 40’ and ‘Marianna 2624’ consistently showed superior tolerance under stress, exhibiting a stem length increase of up to 30% and a 25% increase in stomatal conductance [13].
Water stress: Water stress encompasses both drought and waterlogging, with the former preventing roots from absorbing adequate water and inhibiting various biological processes, while the latter causes oxygen deprivation in the roots, leading to the loss of normal biological functions. In cherries, a comparison of the root structure and anatomical characteristics of different genotypic rootstocks revealed that ‘SV2-7’ (P. fruticosa) and ‘OV14’ (P. cerasus) are drought tolerant rootstocks [14]. Additionally, ‘Daqingye’ (P. pseudocerasus) and ‘Gisela 6’ (P. cerasus × P. canescens) were found to have higher levels of ROS scavenging ability and were more tolerant to waterlogging [15]. In peaches, studies have shown that the wild almond (P. ramonensis) exhibits higher drought resistance than the common peach rootstocks used in orchards, and is considered a high-quality material for breeding highly resistant rootstocks [16]. The rootstock ‘MP-29’ (P. umbellata × P. persica) is more flood-tolerant than the commonly used rootstock ‘Flordaguard’ (P. persica), making it suitable for promotion in subtropical regions prone to flooding [17]. Comparative studies of peach and plum rootstocks have found that among ‘Mr. S. 2/5’ (P. cerasifera), ‘Monegro’ (P. persica × P. dulcis), and ‘Nemared’ (P. persica × P. davidiana), the plum rootstock ‘Mr. S. 2/5’ has superior root porosity and greater tolerance to flooding [18]. Klumb et al. [19] also found that the plum rootstock ‘Marianna 2624’ has stronger waterlogging tolerance than the peach rootstock ‘NR0170401’ (P. persica) by assessing gas exchange parameters and changes in gene expression involved in glycolysis and ethylene metabolism under flooded conditions.
Salinization: Soil salinization is another factor that affects the growth of root systems in stone fruit rootstocks. Over the past decade or so, there has been some success in the breeding of salt-tolerant rootstocks. A comprehensive salt tolerance analysis of seven Prunus rootstocks was conducted, identifying ‘Mariana 2624’, ‘Garnem’, and ‘Colt’ as having strong salt tolerance, with ‘Mariana 2624’ being recognized as the most salt - tolerant among the plum rootstocks [20]. Among peach and almond rootstocks, ‘Empyrean 1’ (P. persica × P. davidiana), ‘Cornerstone’ (P. persica × P. dulcis), and ‘Bright’s hybrid 5’ (P. dulcis × P. persica) also exhibit strong salt tolerance [21]. Furthermore, the high salt tolerance of the almond rootstock ‘Empyrean 1’ was further confirmed, with suggestions that its mechanism may involve upregulating ion transport proteins and enhancing the deposition of suberin and lignin in the root cortex to cope with salt stress [22].
Nutritional stress: The essential nutrients required by stone fruits include macronutrients such as N, P, K, and Ca, as well as micronutrients like Fe, Mn, and Zn, with both excess and deficiency of these nutrients leading to poor plant growth. Under nutrient-poor soil conditions, rootstocks with efficient nutrient uptake can help reduce the incidence of physiological diseases in the scion and decrease the amount of fertilizer needed. Reports show significant differences in the kinetic parameters of NO3− and NH4+ absorption among different peach rootstocks, with varieties such as ‘Aldrighi’ (P. persica), ‘Tsukuba1’ (P. persica), and ‘Clone 15’ (P. persica) possessing a high-affinity transport system for NH4+, which facilitates nutrient translocation to the scion [23]. Similarly, the peach rootstock ‘Shannong-1’ exhibits higher nitrogen use efficiency compared to ‘Maotao’, a difference that is associated with the differential expression of nitrogen metabolism regulatory genes [24]. Additionally, the peach rootstock ‘Garnem’ may exhibit higher calcium use efficiency than the peach rootstock ‘GF677’ due to its larger cortical cells and secondary enlarged xylem vessels [25]. However, other reports suggest that ‘GF677’ has stronger tolerance to iron chlorosis, which may be attributed to the high expression of genes related to iron ion transport proteins, defense systems, and photosynthesis [26]. Additionally, using tolerant rootstocks to iron chlorosis is the best choice to prevent chlorosis. An evaluation of 17 Prunus rootstocks, conducted by measuring root ferric chelate reductase enzyme activity, leaf SPAD values, and field performance, identified ‘Adesoto’ (P. insititia), ‘Felinem’ (P. dulcis × P. persica), ‘GF 677’, ‘Krymsk 86’ (P. cerasifera × P. persica), and ‘PAC 9921-07’ ((P. besseyi × P. salicina) × P. armeniaca) as having stronger tolerance to iron chlorosis [27]. In soils with excess elements, tolerant rootstocks are also a useful tool for improving scion growth. Studies have found that in soils with high zinc content, the cortical cells of the root systems of peach and plum rootstocks ‘Rigitano’ (P. mume) and ‘Tsukuba-1’ (P. persica) are prone to rupture, while ‘Flordaguard’ shows no significant changes in root tip morphology and anatomy, making it recommended as a rootstock with high zinc tolerance [28].
3.3. Biotic Stress Tolerance
In the production of stone fruit trees, pests and diseases not only directly cause poor growth but also lead to a decline in fruit quality and yield loss. The main pests and diseases affecting stone fruit trees include sharka, crown gall, root rot, and nematodes.
Viral diseases:Plum pox virus (PPV) is one of the most dangerous viruses affecting stone fruit trees, causing host leaves to curl and lose their green color, leading to fruit deformities, altering fruit composition, and even resulting in premature fruit drop [29,30]. PPV typically spreads through infected propagation materials and aphid vectors, and has now been reported in at least 33 countries [30]. To develop rootstocks resistant to PPV, Polák and Oukropec [31] evaluated the resistance of rootstocks ‘Barier’ (P. davidiana × P. persica), ‘Fire’ (P. amygdalus × P. persica), ‘Cadaman’, ‘GF677’, ‘MRS’ (P. cerasifera × P. spinosa), ‘NBS 540-73’ (P. cerasifera × P. holoserica × P. domestica), and ‘Pumiselect’ (P. pumila), finding that ‘GF677’ exhibited high resistance to PPV, while ‘Cadaman’ and ‘Fire’ showed moderate resistance. Subsequently, Polák and Komínek [32] further discovered that ‘GF677’ and ‘Myrobalan BN4Kr’ (P. cerasifera) rootstocks are more suitable for propagating plums, apricots, and peaches in areas affected or threatened by PPV. A recent report also indicates that ‘GF677’ is resistant to a unique PPV strain, PPV-T, found in Turkey, while common rootstocks such as ‘Nemaguard’ (P. persica × P. davidiana), ‘Myrobalan 28B’ (P. cerasifera), and ‘Myrobalan 29C’ (P. cerasifera) are sensitive to PPV-T [33].
Bacterial diseases: Crown gall, caused by Agrobacterium tumefaciens, is one of the most susceptible bacterial diseases for stone fruit saplings such as apricot, peach, and almond. An evaluation was conducted to screen for resistant rootstocks to mitigate the damage of this disease to stone fruit production, assessing the resistance of Prunus rootstocks ‘GF677’, ‘Antafuel’ (P. amygdalus × P. persica), ‘St.Julien GF 655/2’ (P. insititia × P. domestica), and ‘Gisela 5’, and the results showed that ‘St.Julien GF 655/2’ is insensitive to A. tumefaciens and exhibits the strongest resistance [34]. Bacterial canker, caused by Pseudomonas syringae pv syringae (Pss), is a serious disease of stone fruit orchards causing severe yield reductions and the death of entire trees.
Fungal diseases: Root rot mediated by Armillaria mellea or A. tabescens is one of the leading causes of death in stone fruit trees. According to existing reports, the plum rootstock ‘Sharpe’ (P. angustifolia × Prunus spp.) and ‘MP-29’ exhibit resistance to A. tabescens [35,36]. ‘Myrobalan’ (P. cerasifera) and its interspecific hybrids not only show some resistance to A. tabescens but also to A. mellea [37].
Pest damage: Root-knot nematodes (RKN), including Meloidogyne hapla, M. arenaria, M. incognita, and M. javanica, are among the most damaging pests to stone fruit trees. Since 1929, breeders have developed some peach rootstocks resistant to RKN, including ‘Nemaguard’, ‘Hansen’ (P. persica × P. amygdalus), ‘Flordaguard’, ‘Okinawa’, and ‘Nemared’ [38]. Eliwa et al. [39] evaluated that most genotypes of the local Egyptian peach variety ‘Mit-Ghamer’ (P. persica) exhibit resistance or moderate resistance to M. incognita and M. javanica. Unlike peach rootstocks, which are specifically resistant to certain types of RKN, the plum rootstock ‘Myrobalan’ has been found to be completely resistant to over 30 species of RKN, significantly mitigating the damage they cause to plum production [40]. Subsequent research further revealed that the broad-spectrum resistance of ‘Myrobalan’ is primarily attributed to three resistance genes: Mal, Ma2, and Ma3 [41], which may activate the phenylpropanoid and flavonoid metabolic pathways to produce defensive secondary metabolites [42].
3.4. Graft Compatibility
The grafting compatibility between the scion and the rootstock is a crucial factor in determining the performance and longevity of an orchard. Generally, varieties and species with close genetic relationships exhibit good scion-rootstock compatibility, which is specifically manifested by the ability to establish effective vascular connections between the scion and rootstock. Scion-rootstock incompatibility is typically divided into two scenarios: the first is "displacement" graft incompatibility, which manifests in the first year after grafting as growth cessation, leaf drop, and leaf discoloration; the second is "localized" incompatibility, which appears later in development as a disruption in the continuity of vascular and cambial patterns. This also indicates that scion-rootstock incompatibility is delayed and unpredictable, often leading to significant economic losses. Therefore, it is essential to clearly understand scion-rootstock compatibility.
Apricot: When selecting rootstocks for apricot scions, there is a strong universality in choice, as apricots have moderate compatibility with plum rootstocks (P. cerasifera, P. salicina, P. mariana, P. insititia, P. domestica). A comparison of the grafting compatibility of 13 apricot varieties with four plum rootstocks: ‘Marianna 2624’, ‘Miragreen’ (P. cerasifera × P. davidiana), ‘Mirared’ (P. cerasifera × (P. persica × P. davidiana)), and ‘Montclar’ (P. persica), found that over 90% of apricot varieties showed good compatibility with ‘Miragreen’ and ‘Mirared’, making them suitable as universal rootstocks for grafting apricot [43]. The Aula Dei Experimental Station observed through anatomical studies that apricot varieties (‘Búlida’, ‘Canino’, ‘Moniquí’, and ‘Paviot’) all exhibited good grafting compatibility with hexaploid plum rootstock (P. insititia) [44]. Moreover, apricot scions can also successfully graft onto peach, almond, or their hybrid seedlings [45]. Similarly, sand cherry (P. pumila, P. besseyi) and its interspecific hybrids are also suitable as rootstocks for apricots, but apricots should not be grafted onto nanking cherry (P. tomentosa) [45].
Plum: The selection of rootstocks for plum scions also exhibits considerable diversity. The Aula Dei Experimental Station tested the grafting compatibility of five European plum (P. domestica) varieties and six Chinese plum (P. salicina) varieties with 38 hybrid rootstocks. They found that Chinese plums ‘Angeleno’, ‘Black Amber’, ‘Delbarazur’, ‘Songold’, and European plums ‘President’ and ‘Reine Claude Tardive of Chambourcy’ showed perfect grafting compatibility with all tested rootstocks. In contrast, ‘Reine Claude Verte’ exhibited localized incompatibility with ‘Myrobalan B’ and ‘Myrobalan GF 3-1’ in the second year after budding, and ‘Stanley’ was only compatible with plum rootstocks [46].
Cherry: The selection of rootstocks for cherry scions is highly specific. For example, ‘Mazzard’ (P. avium), derived from sweet cherry seedlings, shows good grafting compatibility with all varieties of sweet cherry but exhibits incompatibility with rootstocks from different species, such as P. mahaleb, P. cerasus, P. canescens, P. pseudocerasus, P. fruticosa, and their hybrid progeny [47]. The accumulation of phenolic compounds and increased peroxidase activity may be the reasons for grafting incompatibility between cherry species or within the same species [48,49].
3.5. Dwarfing
Dwarfing rootstocks can control the growth vigor of fruit trees, effectively improve land use efficiency, increase yield per unit area, and facilitate mechanized harvesting and orchard management, thereby reducing labor costs in production.
Cherry: Sweet cherries are the first stone fruit trees to have commercial rootstocks with comprehensive vigor control, and the breeding effects of dwarfing rootstocks are the most systematic and significant. The semi-dwarfing rootstocks, including ‘Gisela 5’, ‘P-HL-C’ (P. avium × P. cerasus), and ‘Weiroot 158’ (P. cerasus), reduce tree height by about 50%; the full-dwarfing rootstocks, such as ‘Gisela 3’ (P. cerasus × P. canescens), ‘Lake’ (P. avium × P. fruticosa), ‘Clare’ (P. avium × (P. cerasus × P. fruticosa)), ‘Cass’ (P. avium × (P. cerasus × P. fruticosa)), ‘Crawford’ (P. cerasus × (P. cerasus × P. canescens)) and ‘Weiroot 53’ (P. cerasus), reduce tree height by about 60% [50].
Peach: Up to now, breeders have also developed peach rootstocks with varying degrees of dwarfing. The ‘Sirio’ rootstock induces trees that are about 40% smaller than those of ‘GF77’ and can improve fruit yield and quality[38]. Compared with the standard rootstock ‘Nemaguard’, ‘Controller 5’ (P. salicina × P. persica) reduces tree vigor by 50-60%, while ‘Controller 9’ (P. salicina × P. persica) reduces it by about 90% [51]. Among ‘Rootpac 20’, ‘IRTA-1’, ‘Adesoto’, ‘Ishtara’, ‘Rootpac R’, and ‘Rootpac 40’, ‘Rootpac 20’ is the most dwarfing rootstock [52]. In addition, a peach dwarf mutant caused by a single nucleotide mutation in the gibberellin receptor GID1 has also been discovered, which has the potential to become an excellent dwarfing rootstock [53].
Plum and apricot: Breeding of dwarfing rootstocks for plums and apricots has been underway for over 20 years, but progress in selection has been relatively slow, with preliminary evaluations of dwarfing rootstocks still ongoing in many regions. A recent study preliminarily selected 13 effective dwarfing rootstocks with strong rooting ability from 79 wild plum germplasms in the Euphrates River region [5]. A 16-year grafting experiment conducted at the Aula Dei experimental station found that the plum rootstock ‘Miral 3278 AD’ (P. cerasifera × P. amygdalus) significantly reduced the growth vigor of two European plum scions (‘Reine Claude of Bavay’ and ‘Reine Claude Tardive of Chambourcy’), while also promoting higher yields, making it a good alternative to the commercial rootstock ‘Myrobalan’ [54]. However, among the current major dwarfing rootstocks for plums and apricots, including ‘Krymsk 1’ (P. tomentosa× P. cerasifera), ‘St. Julien GF655/2’ (P. insititia), ‘Wavit’ (P. domestica), ‘Pixy’ (P. insititia), and ‘Pumiselect’ (P. pumila), most only achieve a semi-dwarfing effect (REF). In contrast, there are reports that two commonly used European plum rootstocks in Italy, ‘Penta’ (P. domestica × P. cerasifera) and ‘Tetra’ (P. domestica), can induce apricot trees to achieve a semi-dwarf to dwarf stature [55].
3.6. Others
Rootstock varieties have functionally differentiated effects on the yield and quality of drupes, with specific rootstocks potentially improving one or several quality indicators of the fruit. In peaches, the rootstock ‘KL-38’ (P. cerasifera) can increase the total phenol, total flavonoid, and total monomeric anthocyanin content to the highest levels, while the rootstocks ‘NG-1’ ((P. davidiana × P. persica) × (P. dulcis × P. persica)) and ‘NGF-14’ ((P. davidiana × P. persica) × P. amygdalus) can enhance the total antioxidant content [56].The rootstocks ‘Adarcias’ and ‘Cadaman’ can promote the accumulation of sugars, phenols, and flavonoids in peaches [57]. The peach rootstock ‘Rootpac 40’ can maintain good levels of soluble solids content and titratable acidity while also increasing fruit size [58].
However, research has found that the effects of rootstocks can be contradictory between fruit yield and quality, and even among different quality indicators. For example, ‘GF677’ significantly improves iron chlorosis tolerance but results in the smallest fruit size, while the rootstocks ‘Barrier1’ (P. persica × P. davidiana) and ‘Citation’ (P. persica × P. salicina) can significantly increase the acidity of fruit but reduce its nutritional value [59]. The peach-plum hybrid rootstock ‘PS’ (P. persica × P. cerasifera) can increase the sugar, total phenol, and ascorbic acid content in nectarines, enhancing their antioxidant activity, but it is sensitive to iron chlorosis [58,60]. Similarly, the sweet cherry rootstock ‘Pikú 3’ (P. pseudocerasus × (P. canescens × P. incisa)) may result in lower yields for the ‘Newstar’ variety, but it produces larger fruits with higher soluble solids content and firmness, showing potential for producing high-quality sweet cherries [61]. Milošević et al. [62] found that rootstocks with stronger growth vigor significantly reduced fruit weight, size, soluble solids content, and titratable acidity, but compared to semi-dwarf and dwarf rootstocks, they increased the content of phenolic compounds.
There is often a significant interaction effect between rootstocks and scions in improving fruit yield and quality. In sweet cherry, it was found that the variety ‘Vera’ (P. avium) grafted onto ‘Egervár’ (P. mahaleb), ‘Carmen’ (P. avium) grafted onto ‘Cemany’ (P. mahaleb), and ‘Petrus’ (P. avium) grafted onto ‘Magyar’ (P. mahaleb) rootstocks achieved the highest yields [63]. For apricot (P. armeniaca) varieties ‘E-101’ and ‘E-404’, grafting onto five different rootstocks revealed that the rootstock ‘PAC 00-08’ ((P. salicina × P. cerasifera) × P. armeniaca) significantly increased the firmness of ‘E-101’ fruits, while ‘PADAC 01-47’ ((P. besseyi × P. armeniaca) × (P. cerasifera × P. armeniaca)) notably enhanced the firmness of ‘E-404’ fruits [64]. Therefore, research on the function of rootstocks and the interaction between rootstocks and scions is crucial for developing effective breeding strategies.
4. Breeding Achievements in Rootstocks of Stone Fruit Trees
To date, in pursuit of diverse breeding objectives, breeding scientists have successfully bred a variety of rootstocks for stone fruit trees, each tailored to meet specific needs. For instance, the easily propagated plum rootstock ‘Myrobalan 29C’ was selected from P. cerasifera seedlings; the iron chlorosis-tolerant rootstock ‘GF677’ was bred from the hybridization of P. dulcis and P. persica; and the fully dwarfing cherry rootstock ‘Gisela 3’ was developed from the crossbreeding of P. cerasus and P. canescens. This paper briefly summarizes the main resistance rootstocks (Figure 2) and growth-controlling rootstocks (Table 1) of stone fruit trees.
5. Molecular Breeding Techniques for Rootstocks of Stone Fruit Trees
Traditional breeding methods include introduction and selection breeding, hybrid breeding, backcross breeding, mutation breeding, distant hybrid breeding, and polyploid breeding. These approaches often have a degree of randomness, and obtaining superior varieties typically requires a long breeding cycle. With the advancement of modern molecular biology, molecular breeding techniques such as marker-assisted selection and genetic engineering have emerged. These technologies allow for targeted improvement of desired traits, significantly reducing the breeding cycle. Molecular breeding is also increasingly being applied to the breeding of rootstocks for stone fruit trees.
5.1. Marker-Assisted Breeding
Marker-Assisted Breeding is a highly effective method for genetic improvement. By developing molecular markers closely linked to target traits, breeders can predict these traits in varieties at an early stage. This significantly enhances selection efficiency, reduces the randomness in the breeding process, and accelerates the breeding program.
In the breeding of rootstocks for stone fruit trees, Marker-Assisted Breeding is mainly used to screen for disease-resistant germplasm. Futhermore, much of the research is still concentrated on developing effective molecular markers. To preemptively combat the effects of PPV on apricots, researchers have identified four resistant varieties—‘Cebir’, ‘Lifos’, ‘Karum’, and ‘Zard’—from a collection of 227 Turkish apricot (P. armeniaca) germplasms, selected them for future breeding programs, and developed a PPV resistance marker, ZP002 [65]. Further research has uncovered a strong correlation between the apricot’s ParPMC2res gene and resistance to PPV, with a five-base pair deletion within the second exon of this gene characterized as a suitable Indel molecular marker for identifying PPV resistance [66]. Additionally, studies have used an F1 population of apricots with PPV resistance and SSR markers to identify the PGS1.23 SSR locus as having the strongest association with PPV resistance, with a detection accuracy of 78.3%, and have selected five apricot germplasms resistant to PPV from 37 samples by using this SSR marker, including ‘Cai Zi Huang’, ‘Yang Ji Yuan’, ‘Chui Zhi Shan Xing’, ‘Da Zao Shu’, and ‘Xi Fu Xing’, providing valuable parental material for future breeding programs aimed at enhancing apricot resistance to PPV [67].
PTSL is a devastating disease in peaches, as it is not easily detectable in the early stages of planting, only becoming apparent after 3 to 5 years, by which time the plants are typically facing death. To accurately predict and select superior germplasms resistant to PTSL, Blenda et al. [68] created an F2 population with varying sensitivity to PTSL by crossing the ‘Guardian 3-17-7’ line (PTSL-tolerant) with ‘Nemaguard’ (PTSL-susceptible) and further self-crossing, and using this population along with 151 AFLP and 21 SSR molecular markers, they constructed a genetic linkage map to identify key molecular markers linked to PTSL resistance.
RKN is a major pest causing poor growth in stone fruit trees. Although several peach rootstocks, such as ‘Nemared’, ‘Nemaguard’, and ‘Flordaguard’, have been developed to resist one or more species of RKN, they are not effective against the newly discovered RKN species, M. floridensis. To select peach rootstocks with stronger and broader resistance to RKN, breeders have utilized an interspecific hybrid population between ‘Flordaguard’ and P. kansuensis, and developed the DP98-025 marker by combining SSR marker analysis, which co-locates with the RKN resistance gene [69]. Duval et al. genotyped and phenotyped individuals of apricot, plum, and peach using established RKN markers, and further developed new resistance-associated molecular markers by analyzing the expression patterns of the RKN resistance gene RMia [70,71].
Additionally, molecular markers have been employed to identify rootstock varieties with dwarfing effects. For instance, a dwarfing-related QTL was localized on chromosome 3 of peach through SLAF-seq and genetic analysis, and subsequent development of SNP markers was carried out [72]. It was also found that the peach dwarfing gene Dw is located at the distal end of linkage group 6, with the second SNP in its coding sequence co-segregating with the dwarf phenotype, a finding confirmed in the dwarfing peach variety ‘Small Sunning’ [73].
5.2. Genetic Engineering Breeding
Plant genetic engineering breeding is an efficient method for obtaining new varieties by utilizing genetic engineering techniques, primarily involving the isolation, modification, chromosomal integration, and in vivo expression of target genes in plant.
Currently, due to the lack of natural virus resistance in stone fruits, obtaining stable virus-resistant varieties through genetic modification is a primary focus in the genetic engineering breeding of stone fruit rootstocks. Ten transgenic lines were successfully generated by infecting the hypocotyls of mature European plum seeds with Agrobacterium rhizogenes, and grafting experiments confirmed that seven of these lines exhibited resistance to PPV [74]. The plum rootstock ‘Elita’ ((P. pumila × P. salicina) × P. cerasifera) was successfully modified by using RNAi technology, resulting in a high-quality rootstock with resistance to PPV [75]. Similarly, a PPV resistance gene was introduced into the plum rootstock ‘Startovaya’ (P. domestica), which remained fully resistant to viral infection for nine years after grafting with scions carrying the PPV virus, yet the transgenic rootstock was unable to provide virus resistance to the scions in this study [76]. Fortunately, recent studies have found that PPV resistance based on RNA silencing can be transferred from transgenic plum rootstocks to non-transgenic apricot scions [77]. PPV translation initiation factors include eIF(iso)4G and eIF(iso)4E. Using RNAi to silence these factors in the sour cherry rootstock ‘146-2’ (P. pumila × P. tomentosa) showed that silencing eIF(iso)4G doesn’t increase resistance, but silencing eIF(iso)4E enhances resistance to PPV, with no signs of infection observed for two years after inoculation, demonstrating the potential to establish PPV-resistant rootstocks by inhibiting translation initiation factors in clonal rootstocks [78].
In addition to improving rootstock resistance, genetic engineering has also been used to enhance the rooting efficiency of stone fruit rootstocks. For example, after the recombinant gene PcMPK3-HA was successfully introduced into the cherry rootstock ‘Gisela 6’, the transgenic lines exhibited a stronger ability to form adventitious roots and positively influenced the root growth of tissue-cultured rootstock seedlings after transplantation [79].
Gene editing, as a method of non-exogenous genome modification, is more widely accepted by the public in terms of food security. Currently, gene editing technology for stone fruits is still in the development stage. Recent reports have shown that the hairy root transformation system combined with CRISPR/Cas9 gene editing technology has successfully achieved targeted editing of two transcription factors, ERF74 and GAI, in almonds [80].
6. Future Perspectives in Stone Fruit Rootstock Research
Modern orchard systems’ success hinges on tailored rootstock solutions-a frontier where apples have set a gold standard. Up to now, breeders have successfully developed a variety of rootstock new varieties with excellent characteristics. However, contemporary horticultural systems face emerging challenges from climate volatility, soil degradation, and evolving pathogen pressures. While biological and economic hurdles persist for stone fruits, emerging biotechnologies and collaborative frameworks offer unprecedented opportunities to develop next-generation rootstocks. This synthesis proposes three strategic research priorities to advance stone fruit rootstock innovation.
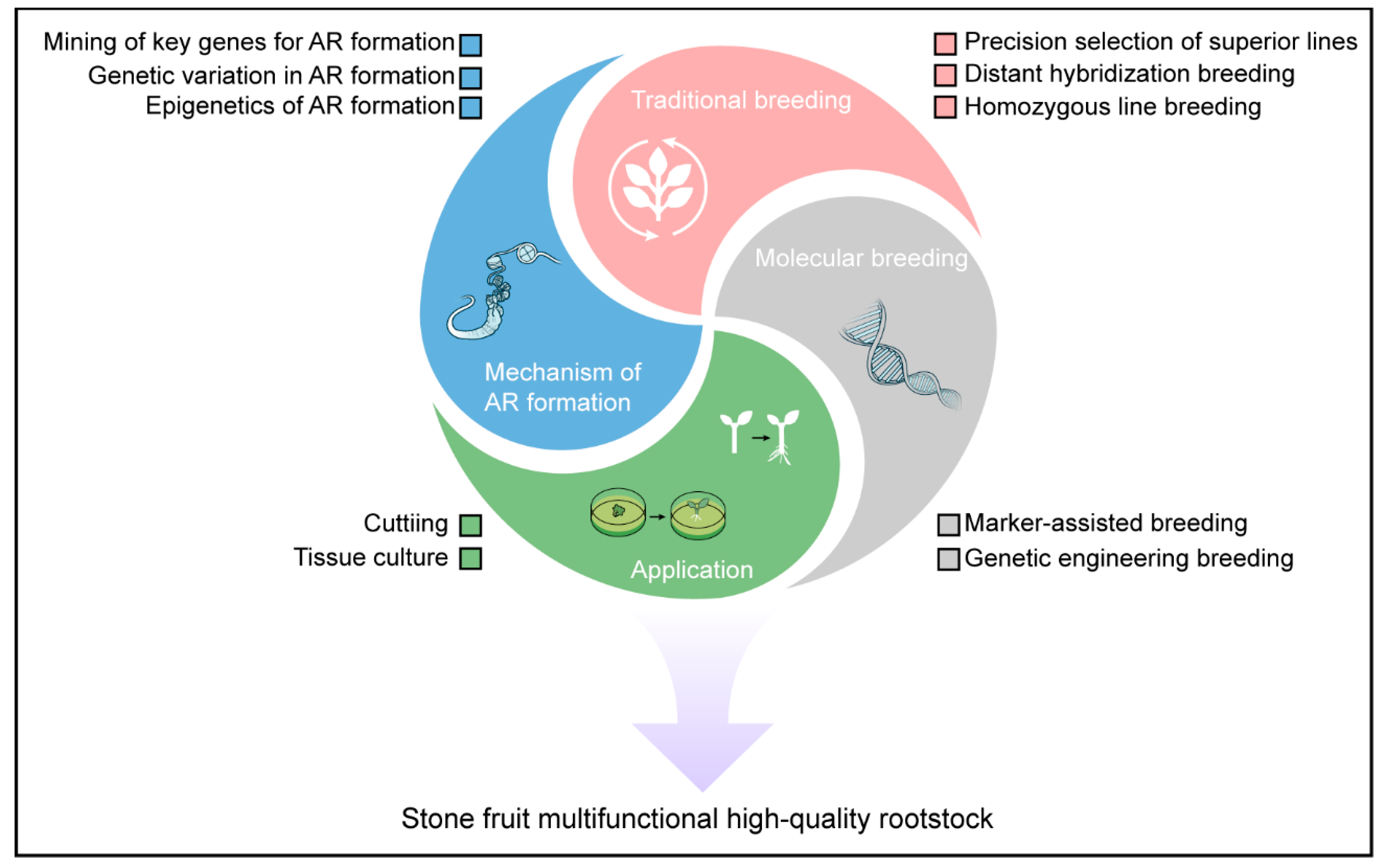
6.1. Revealing the Molecular Mechanism of Adventitious Root Formation in Rootstocks
Currently, many rootstock germplasm resources of stone fruit trees possess excellent traits that can improve the growth vigor or fruit quality of the scions. However, these rootstocks often have weak rooting abilities and can only be used as interstocks. While interstocks can enhance the dwarfing effect and production efficiency of fruit trees to some extent, they cannot ensure the uniformity of scion traits. Therefore, it is essential to identify the key genes that regulate adventitious root formation in stone fruit tree rootstocks. By investigating the molecular mechanisms of adventitious root initiation from multiple perspectives, including epigenetics and molecular regulation, we can fundamentally address the issue of poor rooting in rootstocks. This will pave the way for gradually replacing interstocks with self-rooted rootstocks.
6.2. Developing of Efficient Breeding Strategies and New Techniques for Rootstocks
Although selection breeding and hybrid breeding have relatively low breeding efficiency, they are basic breeding methods that are technically simple and easy to implement. In the future, expanding the scale of germplasm screening and the number of hybrid combinations will have practical significance for the cultivation of high-quality rootstocks. Additionally, creating inbred lines through multiple generations of self-fertilization of self-compatible germplasm, and maximizing heterosis through hybridization of these inbred lines, will also be one of the effective pathways for future rootstock breeding.
Compared to scion cultivar development, stone fruit rootstock breeding programs face significantly extended evaluation cycles, typically requiring 10-15 years or more for commercial validation due to longer juvenile phases and complex trait interactions. These inherent constraints substantially discourage research investment and limit breeder participation in rootstock improvement initiatives. Early molecular marker-assisted breeding has significantly improved breeding efficiency and reduced costs in fruit tree breeding programs. With the evolution of time, biotechnology has been rapidly evolving. The development of novel molecular markers through genome resequencing technology, such as KASP (Kompetitive Allele-Specific PCR), has become a part of modern breeding and will be, or should be, an effective pathway for the future selection and breeding of rootstocks in stone fruit trees. Additionally, elucidating the functions of genes related to important agronomic traits in stone fruit trees and using transgenic or gene-editing technologies to artificially target the creation and improvement of rootstock traits, such as ease of rooting and dwarfing rootstocks, will also be a focus of future rootstock breeding.
6.3. Optimizing Clonal Propagation Techniques for Rootstocks
The clonal propagation of rootstocks is an important factor in ensuring the stable characteristics of scions and achieving high and stable yields. Cutting and tissue culture have long been means of asexual propagation for rootstocks. However, both cutting and tissue culture face genotype-dependent issues, and effective rooting or regeneration methods cannot be widely applied to most high-quality rootstock varieties. Therefore, optimizing the substrate formulations for cuttings and tissue culture and exploring more universal and efficient regeneration and rooting methods will be the direction for the continuous development and improvement of rootstock breeding in stone fruit trees in the future.
Finally, unlike apples’ centralized breeding networks (e.g., Cornell-USDA Geneva program), Prunus rootstock research is dispersed across small public institutes (e.g., INRAE France, CRIOF Italy, and many institutions in China) with limited coordination. Therefore, bridging the rootstock innovation gap in stone fruit also demands international collaboration for Prunus germplasm sharing and phenotyping standardization.
Figure 3.
Research directions and breeding strategies for rootstocks of stone fruit trees. AR: Adventitious roots.
Figure 3.
Research directions and breeding strategies for rootstocks of stone fruit trees. AR: Adventitious roots.
Author Contributions
Conceptualization, J.L., W.Y. and H.S.; data curation, L.Y., J.Z., F.J., M.Z. and Y.W.; writing—original draft preparation, J.L. and W.Y.; writing—review and editing, J.L., W.Y. and H.S.; visualization, J.L. and W.Y.; project administration, H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Beijing Natural Science Foundation (No. 6232011).
Data Availability Statement
Not applicable.
Conflicts of Interest
All authors declare that they have no conflicts of interest.
References
- Riva, S. C.; Opara, U. O.; Fawole, O. A. , Recent developments on postharvest application of edible coatings on stone fruit: A review. Scientia Horticulturae 2020, 262. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations Database (FAOSTAT). In 2022.
- Chen, G.; Boddu, R.; Aadil, R. M. , Study on Double-Layer Stereo Ecological Cultivation Technology of Greenhouse Gardening Fruit Trees. Journal of Food Quality 2022, 2022, 1–8. [Google Scholar] [CrossRef]
- Scalisi, A.; O'Connell, M. G.; Stefanelli, D.; Zhou, S.; Pitt, T.; Graetz, D.; Dodds, K.; Han, L.; De Bei, R.; Stanley, J.; Breen, K.; Goodwin, I. , Narrow orchard systems for pome and stone fruit—a review. Scientia Horticulturae 2024, 338. [Google Scholar] [CrossRef]
- Korkmaz, K.; Bolat, I.; Uzun, A.; Sahin, M.; Kaya, O. , Selection and Molecular Characterization of Promising Plum Rootstocks (Prunus cerasifera L.) among Seedling-Origin Trees. Life, 2023; 13, 1476. [Google Scholar]
- Mayer, N. A.; Ueno, B.; Rickes, T. B.; de Resende, M. V. L. A. , Cloning of rootstock selections and Prunus spp. cultivars by softwood cuttings. Scientia Horticulturae 2020, 273. [Google Scholar] [CrossRef]
- Rosa, G. G. d.; Zanandrea, I.; Mayer, N. A.; Bianchi, V. J., Propagação de porta-enxerto de Prunus spp. por estaquia: efeito do genótipo, do estádio de desenvolvimento do ramo e tipo de estaca. Revista Ceres 2017, 64, 90-97.
- Justamante, M. S.; Mhimdi, M.; Molina-Pérez, M.; Albacete, A.; Moreno, M. Á.; Mataix, I.; Pérez-Pérez, J. M. , Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks. Plants 2022, 11, 913. [Google Scholar] [CrossRef]
- Tsafouros, A.; Frantzeskaki, A.; Assimakopoulou, A.; Roussos, P. A. , Spatial and temporal changes of mineral nutrients and carbohydrates in cuttings of four stone fruit rootstocks and their contribution to rooting potential. Scientia Horticulturae 2019, 253, 227–240. [Google Scholar] [CrossRef]
- Tsafouros, A.; Roussos, P. A. , The possible bottleneck effect of polyamines' catabolic enzymes in efficient adventitious rooting of two stone fruit rootstocks. Journal of Plant Physiology 2020, 244. [Google Scholar] [CrossRef]
- Blažková, J.; Hlušičková, I. , Testing of wood hardiness to winter freezes in selections from progenies of Cerapadus × Prunus avium L. crosses. Horticultural Science 2002, 29, 133–142. [Google Scholar] [CrossRef]
- Turhan, E.; Ergin, S. , Soluble Sugars and Sucrose-Metabolizing Enzymes Related to Cold Acclimation of Sweet Cherry Cultivars Grafted on Different Rootstocks. The Scientific World Journal 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Dogan, M.; Bolat, I.; Turan, M.; Kaya, O. , Elucidating stress responses in Prunus rootstocks through comprehensive evaluation under drought, heat shock and combined stress conditions. Scientia Horticulturae 2025, 339. [Google Scholar] [CrossRef]
- Ljubojević, M.; Zorić, L.; Maksimović, I.; Dulić, J.; Miodragović, M.; Barać, G.; Ognjanov, V. , Anatomically assisted cherry rootstock selection. Scientia Horticulturae 2017, 217, 197–208. [Google Scholar] [CrossRef]
- Jia, L. t.; Qin, X.; Lyu, D. g.; Qin, S. j.; Zhang, P. , ROS production and scavenging in three cherry rootstocks under short-term waterlogging conditions. Scientia Horticulturae 2019, 257, 108647. [Google Scholar] [CrossRef]
- Gerbi, H.; Paudel, I.; Zisovich, A.; Sapir, G.; Ben-Dor, S.; Klein, T. , Physiological drought resistance mechanisms in wild species vs. rootstocks of almond and plum. Trees 2021, 36, 669–683. [Google Scholar] [CrossRef]
- McGee, T.; Schaffer, B.; Shahid, M. A.; Chaparro, J. X.; Sarkhosh, A. , Carbon and nitrogen metabolism in peach trees on different Prunus rootstocks in response to flooding. Plant and Soil 2022, 475, 427–441. [Google Scholar] [CrossRef]
- Ziegler, V. H.; Ploschuk, E.; Weibel, A.; Insausti, P. , Short-term responses to flooding stress of three Prunus rootstocks. Scientia Horticulturae 2017, 224, 135–141. [Google Scholar] [CrossRef]
- Klumb, E. K.; Braga, E. J. B.; Bianchi, V. J. , Differential expression of genes involved in the response of Prunus spp. rootstocks under soil flooding. Scientia Horticulturae 2020, 261. [Google Scholar] [CrossRef]
- Toro, G.; Pimentel, P.; Salvatierra, A. , Effective Categorization of Tolerance to Salt Stress through Clustering Prunus Rootstocks According to Their Physiological Performances. Horticulturae 2021, 7, 542. [Google Scholar] [CrossRef]
- Sandhu, D.; Kaundal, A.; Acharya, B. R.; Forest, T.; Pudussery, M. V.; Liu, X.; Ferreira, J. F. S.; Suarez, D. L. , Linking diverse salinity responses of 14 almond rootstocks with physiological, biochemical, and genetic determinants. Scientific Reports 2020, 10, 21087. [Google Scholar] [CrossRef]
- Shao, Y. h.; Cheng, Y. k.; Pang, H. g.; Chang, M. q.; He, F.; Wang, M. m.; Davis, D. J.; Zhang, S. x.; Betz, O.; Fleck, C.; Dai, T.; Madahhosseini, S.; Wilkop, T.; Jernstedt, J.; Drakakaki, G. , Investigation of Salt Tolerance Mechanisms Across a Root Developmental Gradient in Almond Rootstocks. Frontiers in Plant Science 2021, 11, 595055. [Google Scholar] [CrossRef]
- Paula, B. V. d.; Marques, A. C. R.; Rodrigues, L. A. T.; Souza, R. O. S. d.; Kulmann, M. S. d. S.; Kaminski, J.; Ceretta, C. A.; Melo, G. W. B. d.; Mayer, N. A.; Antunes, L. E.; Ricachenevsky, F. K.; Nicoloso, F. T.; Brunetto, G. , Morphological and kinetic parameters of the uptake of nitrogen forms in clonal peach rootstocks. Scientia Horticulturae 2018, 239, 205–209. [Google Scholar] [CrossRef]
- Chen, Q. j.; Lian, M.; Guo, J.; Zhang, B. b.; Yang, S. k.; Huang, K. x.; Peng, F. t.; Xiao, Y. s. , Comparative Transcriptome Analysis of Two Peach Rootstocks Uncovers the Effect of Gene Differential Expression on Nitrogen Use Efficiency. International Journal of Molecular Sciences 2022, 23, 11144. [Google Scholar] [CrossRef] [PubMed]
- Aras, S.; Keles, H.; Bozkurt, E. , Physiological and histological responses of peach plants grafted onto different rootstocks under calcium deficiency conditions. Scientia Horticulturae 2021, 281, 109967. [Google Scholar] [CrossRef]
- Sun, S. x.; Li, J.; Song, H. y.; Chen, D.; Tu, M. y.; Chen, Q. y.; Jiang, G. l.; Zhou, Z. q. , Comparative transcriptome and physiological analyses reveal key factors in the tolerance of peach rootstocks to iron deficiency chlorosis. 3 Biotech 2022, 12, 38. [Google Scholar] [CrossRef]
- Jiménez, S.; Pinochet, J.; Abadía, A.; Moreno, M. Á.; Gogorcena, Y. , Tolerance Response to Iron Chlorosis of Prunus Selections as Rootstocks. HortScience 2008, 43, 304–309. [Google Scholar] [CrossRef]
- Somavilla, L. M.; Simão, D. G.; Tiecher, T. L.; Hammerschimitt, R. K.; de Oliveira, J. M. S.; Mayer, N. A.; Pavanello, E. P.; Trentin, E.; Belles, S. W.; Brunetto, G. , Structural changes in roots of peach rootstock cultivars grown in soil with high zinc content. Scientia Horticulturae 2018, 237, 1–10. [Google Scholar] [CrossRef]
- Usenik, V.; Marn, M. V. , Sugars and organic acids in plum fruit affected by Plum pox virus. Journal of the Science of Food and Agriculture 2016, 97, 2154–2158. [Google Scholar] [CrossRef]
- Zhou, J.; Xing, F.; Wang, H.; Li, S. , Occurrence, Distribution, and Genomic Characteristics of Plum Pox Virus Isolates from Common Apricot (Prunus armeniaca) and Japanese Apricot (Prunus mume) in China. Plant Disease 2021, 105, 3474–3480. [Google Scholar] [CrossRef]
- Polák, J.; oukroPec, I. , Identification of Interspecific Peach and Prunus sp. Hybrids Resistant to Plum Pox Virus Infection. Plant Protection Science 2010, 46, 139–144. [Google Scholar] [CrossRef]
- Polák, J.; Komínek, P. , Evaluation of rootstocks of stone fruits for resistance to natural Plum pox virus infection. Canadian Journal of Plant Pathology 2014, 36, 116–120. [Google Scholar] [CrossRef]
- Cinar, C. T.; Gazel, M.; Kaya, K.; Olmos, A.; Caglayan, K. , Susceptibility of different prunus rootstocks to natural infection of plum pox virus-Turkey (PPV-T) in Central Anatolia. Physiological and Molecular Plant Pathology 2022, 119. [Google Scholar] [CrossRef]
- T, T.; E, E.; C, T. , Susceptibility of five Prunus rootstocks to Agrobacterium tumefaciens. New Zealand Journal of Crop and Horticultural Science 2005, 33, 343–345. [Google Scholar]
- Beckman, T. G. , ‘Sharpe’, a Clonal Plum Rootstock for Peach. HortScience 2008, 43, 2236–2237. [Google Scholar] [CrossRef]
- Beckman, T. G. , ‘MP-29’, a Clonal Interspecific Hybrid Rootstock for Peach. HortScience 2012, 47, 128–131. [Google Scholar] [CrossRef]
- Baumgartner, K.; Fujiyoshi, P.; Ledbetter, C.; Duncan, R.; Kluepfel, D. A. , Screening Almond Rootstocks for Sources of Resistance to Armillaria Root Disease. HortScience 2018, 53, 4–8. [Google Scholar] [CrossRef]
- Lesmes-Vesga, R. A.; Cano, L. M.; Ritenour, M. A.; Sarkhosh, A.; Chaparro, J. X.; Rossi, L. , Rootstocks for Commercial Peach Production in the Southeastern United States: Current Research, Challenges, and Opportunities. Horticulturae 2022, 8, 602. [Google Scholar] [CrossRef]
- Eliwa, G. I.; Hagag, E. S. , Approach to New peach rootstocks resistant to root-knot nematodes (Meloidogyne species) selected from local Mit-Ghamer peach cultivar. Scientia Horticulturae 2021, 284. [Google Scholar] [CrossRef]
- Rubio-Cabetas, M. J.; Lecouls, A. C.; Salesses, G.; Bonnet, A.; Esmenjaud, D. , Evidence of a new gene for high resistance to Meloidogyne spp.in Myrobalan plum, Prunus cerasifera. Plant Breeding, 1998; 117, 567–571. [Google Scholar]
- Lecouls, A. C.; Rubio-Cabetas, M. J.; Minot, J. C.; Voisin, R.; Bonnet, A.; Salesses, G.; Dirlewanger, E.; Esmenjaud, D. , RAPD and SCAR markers linked to the Ma1root-knot nematode resistance gene in Myrobalan plum (Prunus cerasifera Ehr.). Theoretical & Applied Genetics 1999, 99, 328–335. [Google Scholar]
- Liu, J.; Zhu, J.; Li, H.; Luo, D.; Xie, J.; Li, H.; Liu, S.; Zhang, Y.; Chen, L.; Xie, X.; Wang, D.; Li, K.; Yao, M.; Zhang, G. , A preliminary study on the root-knot nematode resistance of a cherry plum cultivar Mirabolano 29C. Czech Journal of Genetics and Plant Breeding 2023, 59, 133–140. [Google Scholar] [CrossRef]
- Irisarri, P.; Errea, P.; Pina, A. , Physiological and Molecular Characterization of New Apricot Cultivars Grafted on Different Prunus Rootstocks. Agronomy 2021, 11. [Google Scholar] [CrossRef]
- Reig, G.; Zarrouk, O.; Font i Forcada, C.; Moreno, M. Á. , Anatomical graft compatibility study between apricot cultivars and different plum based rootstocks. Scientia Horticulturae 2018, 237, 67–73. [Google Scholar] [CrossRef]
- Mendelné Pászti, E.; Bujdoso, G.; Ercisli, S.; Hrotkó, K.; Mendel, Á. , Apricot Rootstocks with Potential in Hungary. Horticulturae 2023, 9, 720. [Google Scholar] [CrossRef]
- Reig, G.; Salazar, A.; Zarrouk, O.; Forcada, C. F. i.; Val, J.; Moreno, M. Á. , Long-term graft compatibility study of peach-almond hybrid and plum based rootstocks budded with European and Japanese plums. Scientia Horticulturae 2019, 243, 392–400. [Google Scholar] [CrossRef]
- Liu Qing-zhong; Dong-zi, Z.; Jia-wei, W.; Li-si, Z.; Po, H.; Qing-dang, G., The evaluation of sweet cherry rootstocks and their application prospects in the world. Deciduous Fruits 2023, 55, 1–7.
- Skočajić, D.; Gašić, U.; Dabić Zagorac, D.; Nešić, M.; Tešić, Ž.; Meland, M.; Fotirić Akšić, M. , Analysis of Phenolic Compounds for the Determination of Grafts (in) Compatibility Using In Vitro Callus Cultures of Sato-Zakura Cherries. Plants 2021, 10. [Google Scholar] [CrossRef]
- Jalali, A.; Moghaddam, E. G.; Marjani, A. , Early detection of graft incompatibility in sweet cherry by internode association and callus fusion techniques. Plant Cell, Tissue and Organ Culture (PCTOC), 2024; 156. [Google Scholar]
- Iglesias, I.; Botet, R. , The selection of appropriate rootstock and training system towards sustainable production of stone fruits. Italus Hortus 2024, 31. [Google Scholar] [CrossRef]
- Clark, J. R.; Finn, C. E. , Register of New Fruit and Nut Cultivars List 43. HortScience 2006, 41, 1101–1133. [Google Scholar] [CrossRef]
- Lordan, J.; Zazurca, L.; Maldonado, M.; Torguet, L.; Alegre, S.; Miarnau, X. , Horticultural performance of ‘Marinada’ and ‘Vairo’ almond cultivars grown on a genetically diverse set of rootstocks. Scientia Horticulturae 2019, 256. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.; Tan, B.; Jiang, Y.; Zheng, X.; Ye, X.; Guo, Z.; Xiong, T.; Wang, W.; Li, J.; Feng, J. , A single nucleotide mutation in GID1c disrupts its interaction with DELLA1 and causes a GA-insensitive dwarf phenotype in peach. Plant Biotechnology Journal 2019, 17, 1723–1735. [Google Scholar] [CrossRef]
- Reig, G.; Font i Forcada, C.; Mestre, L.; Betrán, J. A.; Moreno, M. Á. , Potential of new Prunus cerasifera based rootstocks for adapting under heavy and calcareous soil conditions. Scientia Horticulturae 2018, 234, 193–200. [Google Scholar] [CrossRef]
- Sottile, F.; Monte, M.; De Michele, A. , Effect of Different Rootstocks on Vegetative Growth of Japanese and European Plum Cultivars in Southern Italy: Preliminary Results. Acta Horticulturae 2007, 375–380. [Google Scholar] [CrossRef]
- Yaman, M.; UĞUr, R.; SÜMbÜL, A.; KeÇE, Y.; GÖNÜLtaŞ, M.; ÜNsal, H. T.; GÜNeŞ, A.; Yildiz, E.; Yilmaz, K. U. Determination of fruit characteristics, nutrients and biochemical contents of Transvalia (Prunus persica L.) peach cultivar grafted on different clonal rootstocks obtained by selection and hybridization. Scientia Horticulturae 2024, 330.
- Font i Forcada, C.; Gogorcena, Y.; Moreno, M. A. , Fruit sugar profile and antioxidants of peach and nectarine cultivars on almond × peach hybrid rootstocks. Scientia Horticulturae 2013, 164, 563–572. [Google Scholar] [CrossRef]
- Reig, G.; Garanto, X.; Mas, N.; Iglesias, I. , Long-term agronomical performance and iron chlorosis susceptibility of several Prunus rootstocks grown under loamy and calcareous soil conditions. Scientia Horticulturae 2020, 262. [Google Scholar] [CrossRef]
- Giorgi, M.; Capocasa, F.; Scalzo, J.; Murri, G.; Battino, M.; Mezzetti, B. The rootstock effects on plant adaptability, production, fruit quality, and nutrition in the peach (cv. ‘Suncrest’). Scientia Horticulturae 2005, 107, 36-42.
- Iglesias, I.; Giné-Bordonaba, J.; Garanto, X.; Reig, G. , Rootstock affects quality and phytochemical composition of ‘Big Top’ nectarine fruits grown under hot climatic conditions. Scientia Horticulturae 2019, 256. [Google Scholar] [CrossRef]
- López-Ortega, G.; García-Montiel, F.; Bayo-Canha, A.; Frutos-Ruiz, C.; Frutos-Tomás, D. Rootstock effects on the growth, yield and fruit quality of sweet cherry cv. ‘Newstar’ in the growing conditions of the Region of Murcia. Scientia Horticulturae 2016, 198, 326-335.
- Milošević, T.; Milošević, N.; Mladenović, J. , Combining fruit quality and main antioxidant attributes in the sour cherry: The role of new clonal rootstock. Scientia Horticulturae 2020, 265. [Google Scholar] [CrossRef]
- Bujdosó, G.; Magyar, L.; Hrotkó, K. Long term evaluation of growth and cropping of sweet cherry (Prunus avium L.) varieties on different rootstocks under Hungarian soil and climatic conditions. Scientia Horticulturae 2019, 256.
- Hernández, F.; Pinochet, J.; Moreno, M. A.; Martínez, J. J.; Legua, P. , Performance of Prunus rootstocks for apricot in Mediterranean conditions. Scientia Horticulturae 2010, 124, 354–359. [Google Scholar] [CrossRef]
- Gürcan, K.; Çetinsağ, N.; Pınar, H.; Macit, T. , Molecular and biological assessment reveals sources of resistance to Plum pox virus - Turkey strain in Turkish apricot (Prunus armeniaca) germplasm. Scientia Horticulturae 2019, 252, 348–353. [Google Scholar] [CrossRef]
- Polo-Oltra, Á.; Romero, C.; López, I.; Badenes, M.; Zuriaga, E. , Cost-Effective and Time-Efficient Molecular Assisted Selection for PPV Resistance in Apricot Based on ParPMC2 Allele-Specific PCR. Agronomy 2020, 10. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, H.; Boris, K.; Yang, L.; Jiang, F.; Zhang, M.; Wang, Y. , RT-PCR Detection of Plum pox virus and the Screening of DNA Markers Linked to PPV-resistance in Apricot. Acta Agriculturae Boreali-Sinica, 2018; 33, 31–38. [Google Scholar]
- Blenda, A. V.; Verde, I.; Georgi, L. L.; Reighard, G. L.; Forrest, S. D.; Muñoz-Torres, M.; Baird, W. V.; Abbott, A. G. , Construction of a genetic linkage map and identification of molecular markers in peach rootstocks for response to peach tree short life syndrome. Tree Genetics & Genomes, 2007; 3, 341–350. [Google Scholar]
- Maquilan, M. A. D.; Olmstead, M. A.; Olmstead, J. W.; Dickson, D. W.; Chaparro, J. X. , Genetic analyses of resistance to the peach root-knot nematode (Meloidogyne floridensis) using microsatellite markers. Tree Genetics & Genomes, 2018; 14. [Google Scholar]
- Duval, H.; Van Ghelder, C.; Portier, U.; Confolent, C.; Meza, P.; Esmenjaud, D. , New Data Completing the Spectrum of the Ma, RMia, and RMja Genes for Resistance to Root-Knot Nematodes (Meloidogyne spp.) in Prunus. Phytopathology, 2019; 109, 615–622. [Google Scholar]
- Duval, H.; Heurtevin, L.; Dlalah, N.; Caravel, C.; Callot, C.; Van Ghelder, C. , Identification and Expression of the peach TNL RMia genes for the Resistance to the Root-knot Nematode Meloidogyne incognita. bioRxiv 2024. [Google Scholar] [CrossRef]
- Lu, Z.; Niu, L.; Chagné, D.; Cui, G.; Pan, L.; Foster, T.; Zhang, R.; Zeng, W.; Wang, Z. , Fine mapping of the temperature-sensitive semi-dwarf (Tssd) locus regulating the internode length in peach (Prunus persica). Molecular Breeding 2016, 36. [Google Scholar] [CrossRef]
- Cantín, C. M.; Arús, P.; Eduardo, I. , Identification of a new allele of the Dw gene causing brachytic dwarfing in peach. BMC Research Notes 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- García-Almodóvar, R. C.; Clemente-Moreno, M. J.; Díaz-Vivancos, P.; Petri, C.; Rubio, M.; Padilla, I. M. G.; Ilardi, V.; Burgos, L. , Greenhouse evaluation confirms in vitro sharka resistance of genetically engineered h-UTR/P1 plum plants. Plant Cell, Tissue and Organ Culture (PCTOC), 2014; 120, 791–796. [Google Scholar]
- Sidorova, T.; Pushin, A.; Miroshnichenko, D.; Dolgov, S. , Generation of Transgenic Rootstock Plum ((Prunus pumila L. × P. salicina Lindl.) × (P. cerasifera Ehrh.)) Using Hairpin-RNA Construct for Resistance to the Plum pox virus. Agronomy, 2017; 8. [Google Scholar]
- Sidorova, T.; Mikhailov, R.; Pushin, A.; Miroshnichenko, D.; Dolgov, S. , Agrobacterium-Mediated Transformation of Russian Commercial Plum cv. “Startovaya” (Prunus domestica L.) With Virus-Derived Hairpin RNA Construct Confers Durable Resistance to PPV Infection in Mature Plants. Frontiers in Plant Science, 2019; 10. [Google Scholar]
- Alburquerque, N.; Pérez-Caselles, C.; Faize, L.; Ilardi, V.; Burgos, L. , Trans-grafting plum pox virus resistance from transgenic plum rootstocks to apricot scions. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Mourenets, L.; Pushin, A.; Timerbaev, V.; Khmelnitskaya, T.; Gribkov, E.; Andreev, N.; Dolgov, S. , Effect of Gene Silencing of Translation Initiation Factors eIF(iso)4G and eIF(iso)4E on Sour Cherry Rootstock Resistance to Sharka Disease. International Journal of Molecular Sciences 2022, 24. [Google Scholar] [CrossRef] [PubMed]
- Zong, X. j.; Xu, L.; Tan, Y.; Wei, H. r. , Development of genetically modified sweet cherry rootstock ‘Gisela 6’ with overexpression of PcMPK3-HA gene by Agrobacterium-mediated genetic transformation. Plant Cell, Tissue and Organ Culture, 2022; 151, 375–384. [Google Scholar]
- Jedličková, V.; Štefková, M.; Sánchez López, J. F.; Grimplet, J.; Rubio Cabetas, M. J.; Robert, H. S. , Genome editing in almond using hairy root transformation system. Plant Cell, Tissue and Organ Culture (PCTOC), 2024; 159. [Google Scholar]
Figure 1.
Production overview of the top 15 global Stone-fruits-producing countries in 2022.
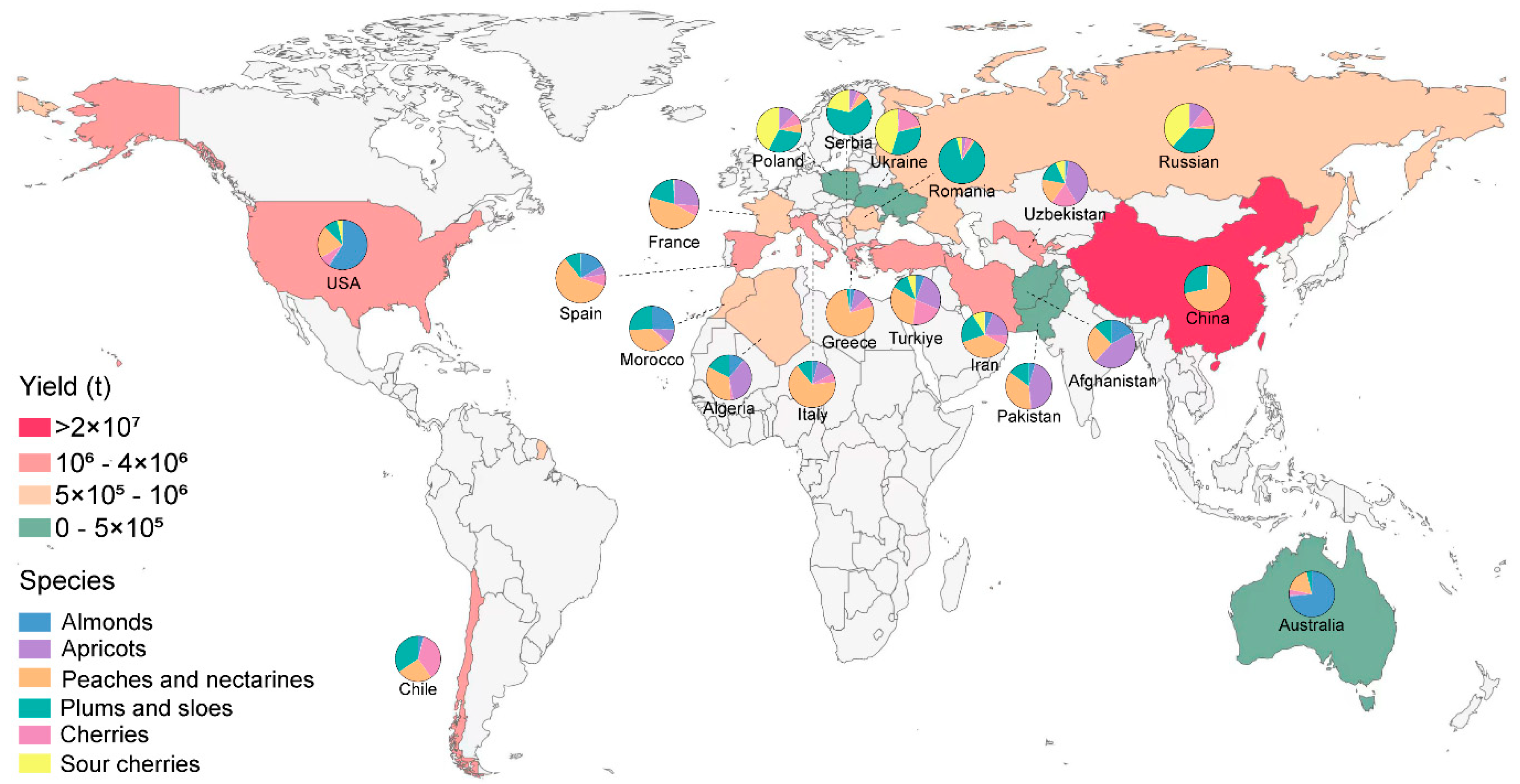
Figure 2.
Common resistant rootstocks for stone fruit trees.Note: The abbreviation in parentheses refers to the bacterial species resisted by the rootstock. PPV: Plum pox virus; AT: Agrobacterium tumefactions; PS: Pseudomonas syringae; AM: Armillaria mellea; Pspp: Phytophthora spp.; CB: Corineum beijerinckii; TP: Tranzschelia. pruni-spinosae; SP: Sphaerotheca pannosa; RN:Rosellinia necatrix.
Figure 2.
Common resistant rootstocks for stone fruit trees.Note: The abbreviation in parentheses refers to the bacterial species resisted by the rootstock. PPV: Plum pox virus; AT: Agrobacterium tumefactions; PS: Pseudomonas syringae; AM: Armillaria mellea; Pspp: Phytophthora spp.; CB: Corineum beijerinckii; TP: Tranzschelia. pruni-spinosae; SP: Sphaerotheca pannosa; RN:Rosellinia necatrix.
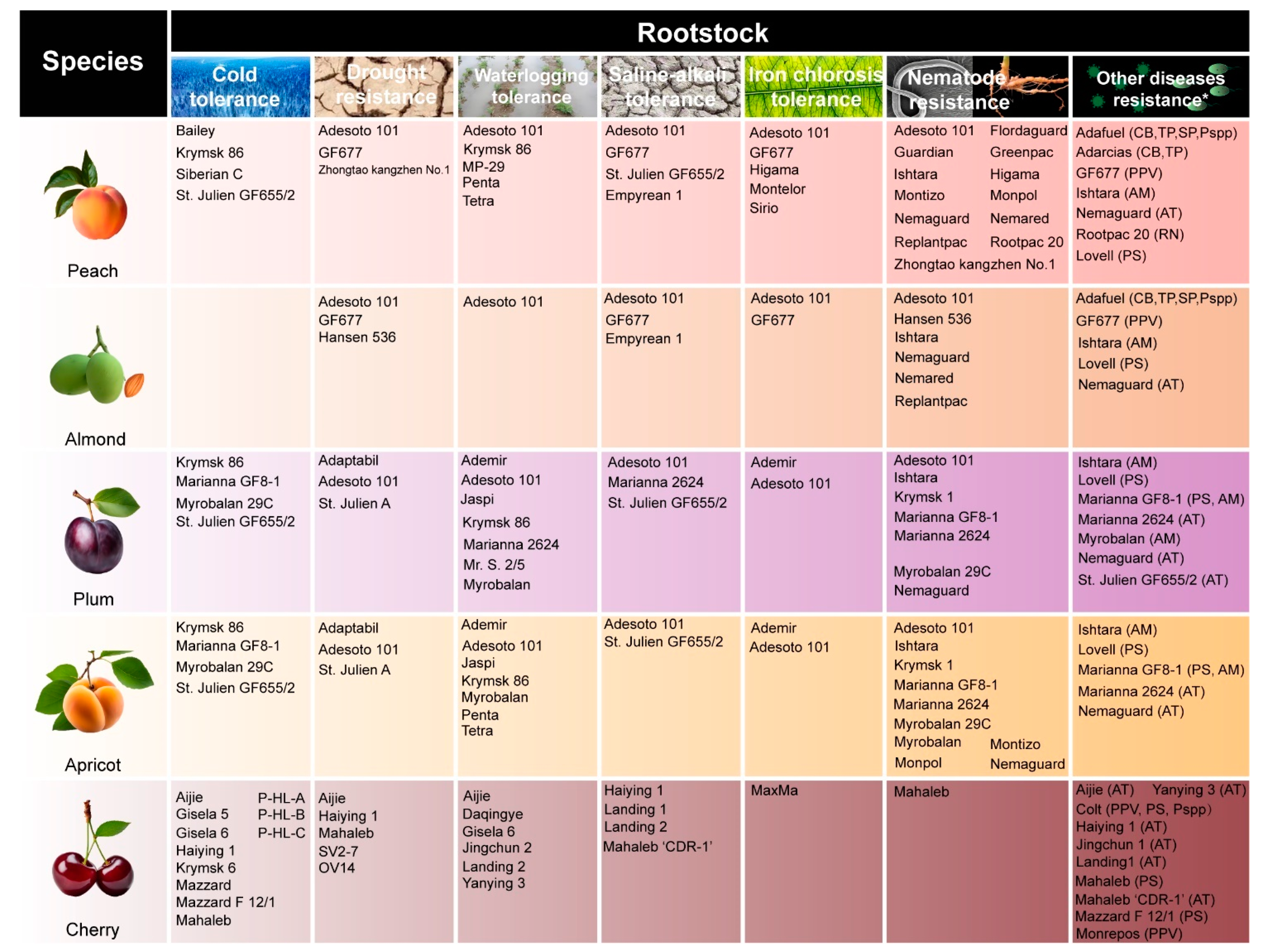

Table 1.
Currently released growth-controlling rootstocks for stone fruit trees.
| Rootstock | Origin | Developed | Compatible scion | Vigour control |
|---|---|---|---|---|
| Hansen 536 | USA | P. dulcis × P. persica | almond | Standard |
| Rootpac 20 | Spain | P. besseyi × P. cerasifera | peach | Dwarf |
| Sirio | Italy | Seedling and open pollinated seedling of GF 557 | peach | Semi-dwarf |
| Siberian C | Canada | P. persica | peach | Semi-vigorous |
| Greenpac | Spain | Cross of Felinem (almond × peach) × Cadaman (peach × P. davidiana) | peach | Semi-vigorous |
| Bailey | USA | Naturalized peach selection | peach | semi-Vigorous |
| Adarcias | Spain | An almond-peach hybrid selected from open pollinated seedling population | peach | Semi-vigorous |
| Higama | France | P. persica | peach | Standard |
| Guardian | USA | P. persica × P. davidiana | peach | very Vigorous |
| Montelor | France | P. spersica | peach | Vigorous |
| Zhong Tao Kang Zhen No. 1 | China | P. spersica × P. kansuensis | peach | vigorous |
| Nemared | USA | P. persica × P. davidiana | peach, almond | Standard |
| GF677 | France | P. dulcis × P. persica | peach, almond | Standard |
| Adafuel | Spain | Seedling population obtained by open pollination of ‘Marcona’ almond cultivars | peach, almond | vigorous |
| Replantpac | Spain | Plum × almond rootstock | peach, almond | Vigorous |
| Penta | Italy | P. domestica × P. cerasifera | peach, apricot | Semi-dwarf |
| Tetra | Italy | P. domestica | peach, apricot | Semi-dwarf |
| Montizo | Spain | Clone selected from Pollizo plum rootstock | peach, apricot | Semi-vigorous |
| Monpol | Spain | Clone selected from Pollizo plum rootstock | peach, apricot | Semi-vigorous |
| Krymsk 1 | Russia | P. tomentosa × P. cerasifera | plum, apricot | Semi-dwarf |
| Jaspi | France | P. salicina × P. spinosa | plum, apricot | Semi-vigorous |
| Adaptabil | Romania | P. besseyi | plum, apricot | Semi-vigorous |
| Ademir | Spain | P. cerasifera | plum, apricot | Semi-vigorous |
| St. Julien A | UK | P. insititia | plum, apricot | Semi-vigorous |
| Myrobalan 29C | USA | P. cerasifera | plum, apricot | standard |
| Marianna GF8-1 | France | P. cerasifera × P.munsoniana | plum, apricot | very Vigorous |
| Marianna 2624 | USA | P. cerasifera × P. musonianna | plum, apricot | very Vigorous |
| Myrobalan | USA | P. cerasifera | plum, apricot | Vigorous |
| St. Julien GF 655/2 | France | P. insititia | plum, apricot, peach, | Semi-dwarf |
| Krymsk 86 | Russia | P. persica × P. cerasifera | plum, apricot, peach, | Standard |
| Ishtara | France | (P. cerasifera × P. salicina) × (P. cerasifera × P. persica) | plum, apricot, peach, almond | Semi-vigorous |
| Adesoto 101 | Spain | P. insititia | plum, apricot, peach, almond | Semi-vigorous |
| Lovell | USA | P. persica | plum, apricot, peach, almond | Standard |
| Nemaguard | USA | P. persica × P. davidiana | plum, apricot, peach, almond | very Vigorous |
| MaxMa | USA | P. Mahaleb × P. avium | sweet cherry | semi-dwarf |
| P-HL-C | Czech Republic | P. avium × P. cerasus | sweet cherry | Semi-dwarf |
| Jingchun 1 | China | P. cerusus × P. pseudocerasus | sweet cherry | Semi-vigorous |
| P-HL-A | Czech Republic | P. avium × P. cerasus | sweet cherry | Semi-vigorous |
| Aijie | China | P. cerasus × P. canescens | sweet cherry | Semi-vigorous |
| Monrepos | Spain | P. cerasifera | sweet cherry | Semi-vigorous |
| Mazzard | USA | P. avium | sweet cherry | very Vigorous |
| P-HL-B | Czech Republic | P. avium × P. cerasus | sweet cherry | Vigorous |
| Mazzard F 12/1 | USA | P. avium | sweet cherry | Vigorous |
| Landing 1 | China | P. avium × P. pseudocerasus | sweet cherry | Vigorous |
| Landing 2 | China | P. avium × P. pseudocerasus | sweet cherry | Vigorous |
| Yanying 3 | China | P. pseudocerasus | sweet cherry | vigorous |
| Haiying 1 | China | P. pseudocerasus | sweet cherry | #N/A |
| Gisela 5 | Germany | P.cerasus × P.canescens | sweet cherry, sour cherry | Semi-dwarf |
| Mahaleb ‘CDR-1’ | USA | P. mahaleb | sweet cherry, sour cherry | Semi-vigorous |
| Gisela 6 | Germany | P.cerasus × P.canescens | sweet cherry, sour cherry | Semi-vigorous |
| Krymsk 6 | Russia | P. cerasus × (P. cerasus × P. maackii) | sweet cherry, sour cherry | Semi-vigorous |
| Mahaleb | USA | P. Mahaleb | sweet cherry, sour cherry | very Vigorous |
| Colt | UK | P. avium × P. pseudocerasus | sweet cherry, sour cherry | vigorous |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



