Submitted:
17 March 2025
Posted:
18 March 2025
You are already at the latest version
Abstract
Background/Objectives: Klebsiella pneumoniae is an important opportunistic pathogen that causes healthcare-associated and community-acquired infections; these include urinary tract infections, bacteremia, pneumonia, and liver abscesses. The rapid spread of multidrug-resistant (MDR) and hy-pervirulent (hvKP) strains of K. pneumoniae, with the ability to adapt to the hospital environment, poses a major challenge for their treatment and control, thereby increasing morbidity and mortality rates globally. In this context, the search for new treatment alternatives is imminent today, with plant-based products being an excellent alternative for use. The aim of this research was to evaluate the antibacterial and an-tibiofilm potential of the alkaloid capsaicin (CAP) against clinical isolates of K. pneumoniae, as well as to explore its possible effect on the cell membrane. Methods/Results: Susceptibility testing using the mi-crodilution method showed antibacterial activity of CAP against all K. pneumoniae isolates tested, with minimum inhibitory concentration (MIC90) values ranging from 28.44 to 1696 µg/mL. Assays to evaluate the effect of CAP against biofilms using the crystal violet technique, showed the ability of CAP to inhibit the formation of biofilms and mature biofilms of K. pneumoniae. Intracellular material leakage experiments (260/280 nm) and Evans blue microscopy showed damage to the cell membrane. Conclusions: This in-dicates that the antibacterial action of CAP could be associated with damage to the integrity of the cell membrane and consequent death of these pathogens. These results serve as a reference for future studies in which the antimicrobial action mechanisms of CAP and its possible synergistic action with other compounds are established.
Keywords:
Klebsiella pneumoniae
; capsaicin
; antibacterial
; antibiofilms
1. Introduction
Klebsiella pneumoniae is an opportunistic pathogen of great global relevance. Over the last decade, it has emerged as a major clinical threat, due to the increasing prevalence of healthcare-associated infections caused by multidrug-resistant (MDR) strains and the emergence of severe community-acquired infections caused by "hypervirulent" strains (hvKP); these pathogens can exceptionally acquire exogenous genetic elements that encode resistance and hypervirulence. The rapid dissemination of these clinical strains, associated with therapeutic failure and high mortality, is alarming. [1,2,3,4,5,6,7]. Klebsiella pneumoniae is a global pathogen with remarkable genetic, phenotypic and pathogenic diversity; there are increasing reports of convergence between these pathotypes and their genetic determinants, raising significant public health concerns. [8]. In the infection process, K. pneumoniae adapts to different environmental conditions, employing the two-component regulatory system (TCS), composed of a sensor histidine kinase and a response regulator, which is an important bacterial regulatory system in response to external stimuli [9]. In recent years, with the widespread use of carbapenems, carbapenem-resistant K. pneumoniae (CRKP) infections have been increasing. Data from the China Antimicrobial Surveillance Network (CHINET) show that the resistance rate of K. pneumoniae to imipenem and meropenem increased eightfold, from 3.0% and 2.9% in 2005 to 25.0% and 26.3% in 2018, respectively. Furthermore, their drug resistance rate to imipenem in four children's hospitals in the report ranged from 32.1% to 45.5%, meaning that K. pneumoniae resistance to carbapenems in the pediatric group deserves more attention [10].
Furthermore, the ability to form biofilms is a crucial virulence trait in the pathogenesis of K. pneumoniae disease, as it promotes increased resistance against environmental stressors and provides a reservoir for dissemination and increased exchange of genes associated with antimicrobial resistance; several virulence factors contribute to biofilm formation by K. pneumoniae, either directly, by promoting increased adherence and/or maturation of the biofilm, or indirectly, by inhibiting biofilm formation by bacterial competitors in its colonizing niche [11,12,13]. Capsule, type 1 and 3 fimbriae (mrkA gene), quorum sensing system type 2 (luxS), D-galactan I synthesis (wbbM), LPS transport (wzm) and polybeta-1,6-N-acetyl-D-glucosamine (pgaA) appear to be involved in K. pneumoniae biofilm [12]. The demonstrated ability of K. pneumoniae to survive and regrow in these biofilms, as well as the high mortality it causes, is of great global concern [14].
In this context, the search and development of novel compounds with potential against K. pneumoniae that are safe, tolerable and effective is urgent today. As is well known, since ancient times products of natural origin, particularly from plants, have made a valuable contribution to pharmacotherapy, especially in infectious diseases and cancer; It is estimated that between 65 and 80% of the world's population in developing countries relies essentially on plants for primary health care due to lack of access to medicine. Furthermore, between 25 and 30% of all drugs available as therapy are derived from natural products, which moves around 20 billion dollars annually in the global pharmaceutical market [15,16]. In this scenario, plants play a primary role as a source of specialized metabolites with recognized medicinal properties [17,18]. Due to their wide chemical diversity, these metabolites can be used directly as bioactive compounds, as drug prototypes or used as pharmacological tools for different targets [19], so they are an excellent alternative to be investigated.
Capsaicinoids (Figure 1) are secondary metabolites of plants, with capsaicin (CAP) (trans-8-methyl-N-vanillyl-6-nonenamide C18H27NO3) being the main responsible for the spiciness of chili peppers; it is biosynthesized through two pathways involved in the metabolism of phenylpropanoids and fatty acids. CAP is used as a food additive and in pharmaceutical applications; it can also act as a cancer preventive agent and shows broad applications against various types of cancer [20]. It has recently attracted attention due to its antimicrobial and antiviral activity. Its antimicrobial potential has been reported against group A hemolytic streptococci [21], Staphylococcus aureus, Listeria monocytogenes [22,23], Vibrio cholerae [23], Acinetobacter baumanii [24], Helicobacter pylori [25], Salmonella typhimurium, Escherichia coli, and Pseudomonas spp., affecting their growth and biofilm formation; likewise, its potential against Candida spp., Toxoplasma gondi and Trypanosoma cruzi has been reported; its action against some viruses has also been documented, whose invasiveness has been affected by the action of this compound [26]. However, the antibacterial potential of this alkaloid against clinical isolates of K. pneumoniae had not been documented, so we hypothesized that CAP could have an effect against K. pneumoniae, and the biofilms formed by these pathogens. The purpose of this research was to evaluate the antibacterial activity of CAP, estimate its capacity to inhibit biofilms and explore its possible effect against the membranes of clinical isolates of K. pneumoniae, contributing to the search for new compounds of natural origin that can serve as adjuvants in the treatment of drug-resistant pathogenic bacteria.
2. Results
2.1. Susceptibility Testing
CAP showed antibacterial activity against all clinical isolates of K. pneumoniae studied; we observed a reduction in the percentage of growth of bacteria treated with CAP, compared to untreated isolates used as a control. Figure 2 shows a similar trend among isolates, with the percentage of growth reduction increasing as the CAP concentration increases. Table 1 shows the MIC values; MIC90 values of CAP were obtained between 28.44 and 1696 µg/mL and it was shown that this effect on K. pneumoniae depended on the CAP concentration, with the percentage of growth reduction increasing as its concentration increased.
We observed that after 24 h the MIC90 values of CAP varied between 28.44 and 1696 µg/mL; as evidenced, the efficacy of CAP was different between strains of the same species.
2.2. Biofilm Reduction
All K. pneumoniae isolates tested produced biofilms on polystyrene microplates as shown in Figure 3a; isolates A10, A42, A38 and A48 were strong biofilm producers while isolates A41 and A49 were moderate biofilm biomass producers. When CAP MIC90 was added to mature biofilms from each isolate, a percentage reduction in biomass in the biofilm ranged from 1.97 to 40.04% after 1 h of exposure to CAP (Figure 3b); while the percentage reduction in biomass formation in the biofilms ranged from 4.26 to 72.48% (Figure 3c). In some cases, these percentages were similar to those obtained with the antibiotics used as control (CIP and GEN) and in others, they were higher, as shown in Table 2 and Table 3.
Table 2 shows the percentages of inhibition of CAP biofilm formation in K. pneumoniae, during 24 hours of exposure.
Table 3 shows the percentages of CAP inhibition against mature K. pneumoniae biofilms after 24 hours of incubation.
2.3. Leakage of Nucleic Acids and Proteins Through the Klebsiella pneumoniae Membrane
The action of CAP on the integrity of K. pneumoniae membranes was evaluated by release assays of intracellular constituents that absorb at 260/280 nm, such as nucleic acids and proteins. These assays were performed at 0, 30, 60 and 120 min after CAP treatment (MIC90 for each isolate). As seen in Figure 4, the OD260/OD280 values in the CAP-treated groups are significantly higher from the start of CAP exposure, compared to the GEN-treated groups, where minimal release and in some cases no leakage of material was observed with this antibiotic with K. pneumoniae A10 (the other isolates showed similar behavior). These results show damage to the cell membrane permeability of K. pneumoniae caused by CAP.
2.4. Effect of CAP on Membrane Integrity
Damage to K. pneumoniae cell membranes by CAP was also evidenced using Evans blue staining. As indicated in Figure 5, when cells were treated with CAP and observed under a light microscope, they stained blue, suggesting damage to cell membrane permeability after 1 h of CAP treatment, thus suggesting that the membrane is a target of the CAP mode of action against clinical isolates of K. pneumoniae.
3. Discussion
Klebsiella pneumoniae infections represent a major challenge in medical science. This pathogen has a large accessory genome of plasmids and chromosomal gene loci; this accessory genome divides K. pneumoniae strains into opportunistic, hypervirulent and multidrug-resistant groups [28], its ability to evade the immune system, its increasing antimicrobial multi-resistance and the emergence of hypervirulent pathotypes have become a great challenge, increasingly limiting the drugs available for its control and increasing morbidity and mortality rates, not only in community infections but also in healthcare-associated infections [1,4,7,29,30,31]. This has sparked our interest in the search for novel, safe and effective compounds with antibacterial potential, particularly against K. pneumoniae. In this context, plant-based compounds are an excellent alternative to be investigated.
In this investigation, we demonstrated that the CAP alkaloid has antibacterial activity against clinical isolates of K. pneumoniae. This effect being concentration dependent, these results are consistent with the results documented [26], indicating that, depending on the concentration and strain of the bacteria, CAP can exert bacteriostatic or even bactericidal effects against a wide range of bacteria. Likewise, the inhibitory effect of CAP against H. pylori strains has been demonstrated, this effect being doce-dependent, at concentrations greater than 10 µg/mL and showing bactericidal activity after 4 hours of exposure [25]. It was also evidenced [21] the effect of CAP against group A Streptococcus, showing MIC values of 64 to 128 µg/mL, and early detection of dead cells in the live/dead cell assay; CAP also inhibited intracellular invasion and hemolytic activity, so it has been suggested that it could thus prevent both the formation of an intracellular reservoir that is difficult to eradicate and the spread of infection to deep tissues. The effect of CAP against bacteria and Candida albicans was documented, showing that L. monocytogenes and S. aureus were more susceptible to the antimicrobial effects of CAP than Salmonella and E. coli O157:H7, while C. albicans were significantly more susceptible than all bacterial species examined [22]. Other studies [24], they have also demonstrated the effect of CAP against colistin-resistant A. baumanii strains, with MIC values > 512 μg/mL and the synergistic effect of CAP, capable of reducing colistin MICs to values below the susceptibility breakpoint. The effect of CAP on the inhibition of cholera toxin production in V. cholerae strains belonging to various serogroups has also been reported, suggesting that CAP could act as a potent repressor to produce this toxin [23]. Alkaloids are widely known for their antibacterial and antifungal potential, the most active ones are mainly planar in structure, amphiphilic, with a molecular mass between 200 and 400 g/mol, and a polar surface of approximately 50 Å2, and they target DNA and/or topoisomerase, as well as the cytoplasmic membrane [32,33].
Biofilms formed by K. pneumoniae represent an important virulence factor that crucially contributes to its pathogenesis and to the multidrug resistance expressed by these pathogens. Natural plant-derived compounds have been shown to have significant antibiofilm properties. This study shows the effect of CAP against biofilm formation and mature biofilms of K. pneumoniae. The effect against mature biofilms was in some cases superior to the effect shown with the evaluated antibiotics CIP and GEN; likewise, the effect shown against mature biofilms was superior to that shown against the formation of these biofilms, in which the effect was in some cases similar to that shown with CIP and GEN. These coincide with reports [26], which indicate that CAP can reduce the pathogenicity of these bacteria, through a variety of mechanisms, such as mitigating the release of toxins or inhibiting the formation of biofilms. On the other hand, the study of natural compounds such as curcumin, eugenol, linoleic acid, chitosan, reserpine and berberine against K. pneumoniae biofilms has been documented [13,34], as well as the effect of thyme and mint essential oils and their active components have been studied as promising antibiofilm agents alone and/or in combination with CIP to inhibit/eradicate K. pneumoniae biofilms [35]. The results of this research are the first report of the antibacterial and antibiofilm effect of CAP against clinical isolates of K. pneumoniae.
This study also explored the effect of CAP on K. pneumoniae membranes, showing that the antibacterial effect of CAP is likely associated with damage to the integrity of K. pneumoniae membranes caused by this alkaloid, as evidenced by the intracellular material efflux experiments at 260/280 nm, which show a significant and early release of intracellular material in K. pneumoniae treated with CAP, compared to untreated cells. This is in agreement with other results [21], which show early detection of dead group A Streptococcus cells when exposed to CAP. Besides, microscopy with Evans blue staining showed blue stained cells when treated with CAP (MIC90), in contrast to untreated cells used as a control, which remained transparent; these results are consistent with documented studies [31] indicating that one of the targets of the antimicrobial action of these compounds is the cell membrane.
In this regard, CAP being a phytochemical with numerous physiological and therapeutic effects, including its important antimicrobial properties and having shown its effect against clinical isolates of K. pneumoniae, further studies are recommended to elucidate the antibacterial action mechanisms of CAP against K. pneumoniae isolates.
4. Materials and Methods
4.1. Reagents
Mueller–Hinton broth (MHB) (Sigma, Mendota Heights, MN, USA) was used for the determination of the MIC and cultures of bacterial isolates. Tryptic soy agar (TSA) and Tryptic soy broth (TSB) (Becton, Dickinson and Company, San Diego, CA, USA), Mueller– Hinton agar (MHA) (Sigma, Mendota Heights, MN, USA), and Brain heart infusion (BHI) broth (Sigma-Aldrich, St. Louis, MO, USA) were also used for the bacterial cultures. The dimethyl sulfoxide (DMSO), phosphate-buffered saline (PBS), crystal violet (CV) and the antibiotics, ciprofloxacin (CIP) y gentamicin (GEN) used in this study were obtained from Sigma-Aldrich, St. Louis, MO, USA. Meanwhile, the glacial acetic acid was obtained from Carlo Erba Reagents, Milan, Italy.
4.2. Capsaicin
Capsaicin was purchased from Sigma-Aldrich Inc, M2028, 8-Methyl-N-vanillyl-trans-6-nonenamide (C18H27NO3) 305.41 g/mol.
4.3. Strains
Eleven clinical isolates of K. pneumoniae (A70, A10, A48, A42, A41, A38, A47, A39, A49, A100, A40) were used in this study. The isolates were cultured from the tracheal aspiration, bronchoalveolar lavage, blood and urine culture samples of patients hospitalized at the Social Health Service S.A.S. in the city of Sincelejo, Colombia. All microorganisms were identified using standard methods: Vitek® 2 Compact. Biomerieux SA (Marcy-l’Étoile,m France). BHI medium was used to maintain the cultures until testing was performed.
4.3. Susceptibility Testing
The MICs of CAP against K. pneumoniae strains were determined by broth microdilution assay using 96-well microtiter plates, following the protocol established in the M07-A9 method of the Clinical and Laboratory Standards Institute (CLSI), [36]. For this purpose, double serial dilutions were made in MHB until reaching final concentrations of the compound (CAP) of 1000, 500, 250, 125, 62.5, 31.25, 15.62, 7.81, 3.90 µg/mL in each reaction well. The assay was performed in a total volume of 200 μL per well, distributed as follows: 100 μL of CAP (at previously described concentrations) and 100 μL of the bacterial inoculum were added at a concentration of 1 × 108 CFU/mL. Each strain was tested in triplicate. The commercial antibiotics CIP and GEN were used as positive controls, and untreated cells as negative controls. Absorbance readings were immediately measured using a ChroMate 4300 ELISA reader at a wavelength of 630 nm, and readings were subsequently taken after 24 hours of incubation at 37 °C. The inhibition percentages were calculated from the following equation:
where ODT24 is the optical density of the test well at 24 h post inoculation; ODT0 the optical density of the test well at 0 h post inoculation; ODGC24 the optical density of the growth control well at 24 h post inoculation and ODGC0: optical density of the growth control well at 0 h post inoculation.
% Inhibition = (1 - (ODT24 - ODT0/ODGC24 - ODGC0)) × 100
4.4. Quantitative Evaluation of Biofilm Inhibition
The effect of CAP on mature K. pneumoniae biofilms was evaluated following the protocol described by [37], with minor modifications. For biofilm formation, bacterial colonies incubated for 24 h in TSA were used, standardizing the bacterial inoculum to 1 × 108 cells/mL. Then, 200 µL of the bacterial inoculum was poured into 96-well polystyrene microplates in each well and incubated at 37 °C for 24 h. The broth was then removed from the microplates and 200 µL of CAP (MIC90 of each isolate) was added in TSB broth and incubated at 37 °C for 1 h. Subsequently, floating cells were removed, and biofilms were washed from the bottom of the wells with deionized water. Excess moisture was then removed by tapping the microplates on sterile napkins and the plates were dried for 5 minutes. Six replicates of each experiment were performed. Cultures without CAP were used as negative control and cultures with CIP and GEN were used as positive control. Biofilm reductions were quantified by staining the wells with 200 µL of 0.1% CV for 20 minutes. Samples were washed with deionized water until excess dye was removed, excess water was carefully dried, and then the CV was solubilized in 250 µL of 30% glacial acetic acid. Absorbance values were measured at 590 nm (OD590) using a SYNERGY LX microplate reader (Biotek). Biofilm production was grouped into the following categories: OD590 < 0.1, non-producers (NP); OD590 0.1–1.0, weak producers (WP); OD590 1.1–3.0, moderate producers (MP); OD590 > 3.0, strong producers (SP). For biofilm formation inhibition assays the standardized bacterial inoculum was incubated simultaneously with the CAP MIC. Biofilm reduction was calculated using the following equation:
where AbsINO: absorbance of the untreated inoculum and AbsCAP: absorbance of the sample treated with CAP
% Biofilm Reduction =(AbsINO-AbsCAP) /AbsCAP × 100
4.5. Leakage of Nucleic Acids and Proteins Through the Cell Membrane
The release of intracellular material was measured according to the methodology proposed by [36], with some modifications. Klebsiella pneumoniae A10 cells grown in nutrient broth were centrifuged at 3000 g for 20 min, washed three times and resuspended in 20 mL of PBS (pH 7.0). The cell suspension was then treated with CAP (MIC90 for each isolate, separate experiments) and incubated at 37 °C for 0, 30, 60 and 120 min. Subsequently, 2 mL of the samples were collected and centrifuged at 3000 g for 20 min. Then, to determine the concentration of the released constituents, 2 mL of the supernatant was used to measure the absorbance at 260/280 nm with a Spectroquant® Prove 300 UV/Vis spectrophotometer (Merck KGaA, Darmstadt, Germany). Samples without CAP and samples with GEN were used as controls. All assays were performed in triplicate.
4.6. Effect of Extracts on Membrane Integrity
To evaluate the effect of CAP on the membrane integrity of K. pneumoniae, staining was also carried out using Evans blue following a protocol described by [36]. Prior to the assay, a 1% solution of Evans blue in phosphate buffered saline (PBS) was prepared. Klebsiella pneumoniae samples were incubated in BHI broth for 24 hours at 37 °C on slides, in triplicate. They were then treated with CAP (CMI90) for 1 h, followed by the addition of Evans blue for 5 min. Untreated cells were used as a control. Samples were then observed under an Olympus CX31 microscope to assess changes in cell membrane integrity.
4.7. Statistical Analysis
The results were analyzed using GraphPad Prism software version 8.0 and Microsoft Excel version 2024. Initially, the Shapiro-Wilk test was used to determine the distribution of the data. Subsequently, Spearman correlation coefficients were used to measure the degree of linearity, the correlation between CAP concentration and the percentage reduction in bacterial growth. To compare the effects of CAP and antibiotics (CIP and GEN) on biofilm reduction, Holm-Sidak's and Games-Howell's tests were used; the Holm-Sidak's test was used to compare the effects of CAP and antibiotics on the leakage of intracellular material through the membrane (260/280 nm).
5. Conclusions
In this study, we investigated the antibacterial potential of the alkaloid CAP against clinical isolates of K. pneumoniae, as well as its role in biofilm inhibition. Furthermore, we explored the action against the cell membrane of these pathogens. We demonstrated the antibacterial action of CAP against K. pneumoniae, this effect being associated with damage to the integrity of the cell membrane, in addition to its action against bacterial biofilms. These studies need to continue, with the aim of elucidating the antibacterial action mechanisms of CAP, and its possible synergistic action with drugs, since it shows promise as an alternative tool for the treatment and control of multi-resistant nosocomial pathogens such as K. pneumoniae.
Author Contributions
Conceptualization, O.I.C.-M., M.P.M.M. and A.A.-O.; methodology, O.I.C.-M., M.P.M.M. and A.A.-O.; validation, O.I.C.-M., M.P.M.M. and A.A.-O.; formal analysis, O.I.C.-M. and A.A.-O.; writing—original draft preparation, O.I.C.-M. and A.A.-O.; writing—review and editing, O.I.C.-M., M.P.M.M. and A.A.-O.; supervision, O.I.C.-M. and A.A.-O.; project administration, A.A.-O.; funding acquisition, A.A.-O. and O.I.C.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by resources from the SI-FCB-01-22 project of the University of Córdoba, Montería, Colombia.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
We would like to thank the University of Córdoba and the Microbiological Interactions Research Group (GIIM).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDR | Multidrug resistance |
| hvKP | Hypervirulent Klebsiella pneumoniae |
| CAP | Capsaicin |
| MIC | Minimum Inhibitory Concentration |
| TCS | Two Component Regulatory System |
| CRKP | Carbapenem-Resistant Klebsiella pneumoniae |
| CHINET | China Antimicrobial Surveillance Network |
| ATBs | Antibiotics |
| CIP | Ciprofloxacin |
| GEN | Gentamicin |
| CLSI | Clinical and Laboratory Standards Institute |
| MHB | Mueller–Hinton Broth |
| TSA | Tryptic Soy Agar |
| TSB | Tryptic Soy Broth |
| MHA | Mueller– Hinton Agar |
| BHI | Brain Heart Infusion |
| DMSO | Dimethyl sulfoxide |
| PBS | Phosphate-buffered saline |
| CV | Crystal violet |
| CFU/mL | Colony forming units/ milliliter |
| OD | Optical density |
References
- García-Cobos, S.; Oteo-Iglesias, J.; Pérez-Vázquez, M. Hypervirulent Klebsiella Pneumoniae: Epidemiology outside Asian countries, antibiotic resistance association, methods of detection and clinical management. Enfermedades Infecciosas y Microbiologia Clinica. 2025. [CrossRef]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The characteristic of virulence, biofilm and antibiotic resistance of Klebsiella Pneumoniae. International Journal of Environmental Research and Public Health. 2020, pp 1–17. [CrossRef]
- Wyres, K. L.; Lam, M. M. C.; Holt, K. E. Population genomics of Klebsiella Pneumoniae. Nature Reviews Microbiology. 2020, pp 344–359. [CrossRef]
- Candan, E. D.; Aksöz, N. Klebsiella Pneumoniae: Characteristics of carbapenem resistance and virulence factors. Acta Biochim Pol 2015, 62 (4), 867–874. [CrossRef]
- Yang, X.; Dong, N.; Chan, E. W. C.; Zhang, R.; Chen, S. Carbapenem resistance-encoding and virulence-encoding conjugative plasmids in Klebsiella Pneumoniae. Trends in Microbiology. 2021, pp 65–83. [CrossRef]
- Wyres, K. L.; Holt, K. E. Klebsiella Pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Current Opinion in Microbiology. 2018, pp 131–139. [CrossRef]
- Van Giang, T.; Anh, N. K.; Phuong, N. Q.; Dat, L. T.; Van Duyet, L. Clinical, antibiotic resistance features, and treatment outcomes of Vietnamese patients with community-acquired sepsis caused by Klebsiella Pneumoniae. IJID Regions 2025, 14. [CrossRef]
- Russo, T. A.; Carlino-MacDonald, U.; Drayer, Z. J.; Davies, C. J.; Alvarado, C. L.; Hutson, A.; Luo, T. L.; Martin, M. J.; McGann, P. T.; Lebreton, F. Deciphering the relative importance of genetic elements in hypervirulent Klebsiella pneumoniae to guide countermeasure development. EBioMedicine 2024, 107. [CrossRef]
- Li, L.; Ma, J.; Cheng, P.; Li, M.; Yu, Z.; Song, X.; Yu, Z.; Sun, H.; Zhang, W.; Wang, Z. Roles of two-component regulatory systems in Klebsiella pneumoniae: regulation of virulence, antibiotic resistance, and stress responses. Microbiological Research. 2023. [CrossRef]
- Luo, K.; Tang, J.; Qu, Y.; Yang, X.; Zhang, L.; Chen, Z.; Kuang, L.; Su, M.; Mu, D. nosocomial infection by Klebsiella pneumoniae among neonates: a molecular epidemiological study. Journal of Hospital Infection 2021, 108, 174–180. [CrossRef]
- Guerra, M. E. S.; Destro, G.; Vieira, B.; Lima, A. S.; Ferraz, L. F. C.; Hakansson, A. P.; Darrieux, M.; Converso, T. R. Klebsiella Pneumoniae biofilms and their role in disease pathogenesis. Frontiers in Cellular and Infection Microbiology. 2022. [CrossRef]
- Vuotto, C.; Longo, F.; Pascolini, C.; Donelli, G.; Balice, M. P.; Libori, M. F.; Tiracchia, V.; Salvia, A.; Varaldo, P. E. Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J Appl Microbiol 2017, 123 (4), 1003–1018. [CrossRef]
- Li, L.; Gao, X.; Li, M.; Liu, Y.; Ma, J.; Wang, X.; Yu, Z.; Cheng, W.; Zhang, W.; Sun, H.; Song, X.; Wang, Z. Relationship between biofilm formation and antibiotic resistance of Klebsiella pneumoniae and updates on antibiofilm therapeutic strategies. Frontiers in Cellular and Infection Microbiology. 2024. [CrossRef]
- Cholley, A. C.; Traoré, O.; Hennequin, C.; Aumeran, C. Klebsiella Pneumoniae survival and regrowth in endoscope channel biofilm exposed to glutaraldehyde and desiccation. European Journal of Clinical Microbiology and Infectious Diseases 2020, 39 (6), 1129–1136. [CrossRef]
- Araldi, R. P.; dos Santos, M. O.; Barbon, F. F.; Manjerona, B. A.; Meirelles, B. R.; de Oliva Neto, P.; da Silva, P. I.; dos Santos, L.; Camargo, I. C. C.; de Souza, E. B. Analysis of antioxidant, cytotoxic and mutagenic potential of Agave sisalana Perrine extracts using Vero cells, human lymphocytes and mice polychromatic erythrocytes. Biomedicine and Pharmacotherapy. 2018, 98, 873–885. [CrossRef]
- Mekonnen Bayisa, Y.; Aga Bullo, T. Optimization and characterization of oil extracted from Croton macrostachyus seed for antimicrobial activity using experimental analysis of variance. Heliyon 2021, 7. [CrossRef]
- Aylate, A.; Agize, M.; Ekero, D.; Kiros, A.; Ayledo, G.; Gendiche, K. In-vitro and in-vivo antibacterial activities of Croton macrostachyus methanol extract against E. coli and S. aureus. Adv Anim Vet Sci 2017, 5, 107–114. [CrossRef]
- Naman, C. B.; Benatrehina, P. A.; Kinghorn, A. D. Pharmaceuticals, Plant Drugs. 2016; Vol. 2. [CrossRef]
- Avato, P. Editorial to the Special Issue – “Natural products and drug discovery". Molecules 2020, 25, 1128. [CrossRef]
- Chapa-Oliver, A. M.; Mejía-Teniente, L. Capsaicin: from plants to a cancer-suppressing agent. Molecules. 2016. [CrossRef]
- Marini, E.; Magi, G.; Mingoia, M.; Pugnaloni, A.; Facinelli, B. Antimicrobial and anti-virulence activity of capsaicin against erythromycin-resistant, cell-invasive group A Streptococci. Front Microbiol 2015, 6. [CrossRef]
- Morrine, A. O.; Zen-Zi, W.; Weih, G. B.; Grant, A. H.; Kamal, D.; David, J. B. Comparative analysis of capsaicin in twenty-nine varieties of unexplored capsicum and its antimicrobial activity against bacterial and fungal pathogens. Journal of Medicinal Plants Research 2018, 12 (29), 544–556. [CrossRef]
- Chatterjee, S.; Asakura, M.; Chowdhury, N.; Neogi, S. B.; Sugimoto, N.; Haldar, S.; Awasthi, S. P.; Hinenoya, A.; Aoki, S.; Yamasaki, S. Capsaicin, a potential inhibitor of cholera toxin production in Vibrio cholerae. FEMS Microbiol Lett 2010, 306 (1), 54–60. [CrossRef]
- Guo, T.; Li, M.; Sun, X.; Wang, Y.; Yang, L.; Jiao, H.; Li, G. Synergistic activity of capsaicin and colistin against colistin-resistant Acinetobacter baumannii: in vitro/vivo efficacy and mode of action. Front Pharmacol 2021, 12. [CrossRef]
- Jones, N. L.; Shabib, S.; Sherman, P. M. Capsaicin as an inhibitor of the growth of the gastric pathogen Helicobacter pylori. FEMS Microbiol Lett 1997, 146 (2), 223–227. [CrossRef]
- Periferakis, A. T.; Periferakis, A.; Periferakis, K.; Caruntu, A.; Badarau, I. A.; Savulescu-Fiedler, I.; Scheau, C.; Caruntu, C. Antimicrobial properties of capsaicin: available data and future research perspectives. Nutrients. 2023. [CrossRef]
- Duelund, L.; Mouritsen, O. Contents of capsaicinoids in chillies grown in Denmark. Food Chemistry 2017, 221, 913–918. [CrossRef]
- Martin, R. M.; Bachman, M. A. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Frontiers in Cellular and Infection Microbiology. 2018. [CrossRef]
- Ballén, V.; Gabasa, Y.; Ratia, C.; Ortega, R.; Tejero, M.; Soto, S. Antibiotic resistance and virulence profiles of Klebsiella pneumoniae strains isolated from different clinical sources. Front Cell Infect Microbiol 2021, 11. [CrossRef]
- Aminul, P.; Anwar, S.; Molla, M. M. A.; Miah, M. R. A. Evaluation of antibiotic resistance patterns in clinical isolates of Klebsiella pneumoniae in Bangladesh. Biosaf Health 2021, 3 (6), 301–306. [CrossRef]
- Beig, M.; Aghamohammad, S.; Majidzadeh, N.; Asforooshani, M. K.; Rezaie, N.; Abed, S.; Khiavi, E. H. G.; Sholeh, M. Antibiotic resistance rates in hypervirulent Klebsiella pneumoniae strains: a systematic review and meta-analysis. Journal of Global Antimicrobial Resistance. 2024, pp 376–388. [CrossRef]
- Sulaiman, M.; Jannat, K.; Nissapatorn, V.; Rahmatullah, M.; Paul, A. K.; de Lourdes Pereira, M.; Rajagopal, M.; Suleiman, M.; Butler, M. S.; Break, M. K. Bin; Weber, J. F.; Wilairatana, P.; Wiart, C. Antibacterial and antifungal alkaloids from asian angiosperms: distribution, mechanisms of action, structure-activity, and clinical potentials. Antibiotics. 2022. [CrossRef]
- Cushnie, T. P. T.; Cushnie, B.; Lamb, A. J. Alkaloids: an overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int J Antimicrob Agents 2014, 44 (5), 377–386. [CrossRef]
- Qian, W.; Sun, Z.; Wang, T.; Yang, M.; Liu, M.; Zhang, J.; Li, Y. Antimicrobial activity of eugenol against carbapenem-resistant Klebsiella pneumoniae and its effect on biofilms. Microb Pathog 2020, 139. [CrossRef]
- Mohamed, S. H.; Mohamed, M. S. M.; Khalil, M. S.; Azmy, M.; Mabrouk, M. I. Combination of essential oil and ciprofloxacin to inhibit/eradicate biofilms in multidrug-resistant Klebsiella pneumoniae. J Appl Microbiol 2018, 125 (1), 84–95. [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Ninth Edition; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32.
- Contreras-Martínez, O.I.; Sierra-Quiroz, D.; Angulo-Ortíz, A. Antibacterial and antibiofilm potential of ethanolic extracts of Duguetia vallicola (Annonaceae) against in-hospital isolates of Pseudomonas aeruginosa. Plants 2024, 13, 1412. [CrossRef]
Figure 1.
Structure of the main capsaicinoids found in species of the genus Capsicum [27]. Capsaicin (1), dihydrocapsaicin (2), nordihydrocapsaicin (3), homocapsaicin (4).
Figure 1.
Structure of the main capsaicinoids found in species of the genus Capsicum [27]. Capsaicin (1), dihydrocapsaicin (2), nordihydrocapsaicin (3), homocapsaicin (4).
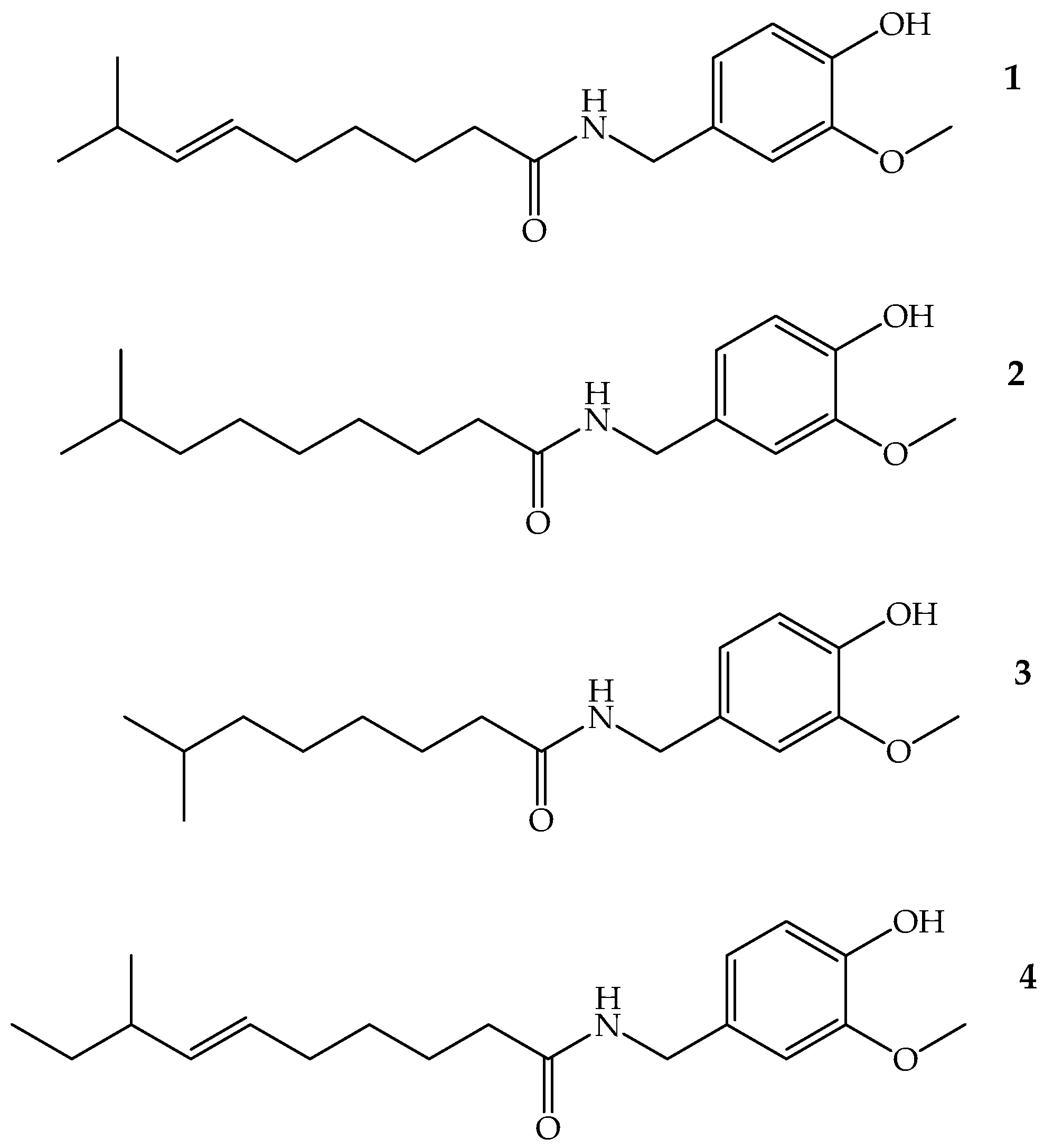
Figure 2.
Growth reduction of K. pneumoniae isolates exposed to CAP (MIC90 of each isolate). A strong and positive linear relationship is observed between the CAP concentration and the percentage of reduction in K. pneumoniae growth, i.e., as the CAP concentration increases, the percentage of reduction in K. pneumoniae growth also increases, which coincides with the Spearman correlation coefficient (0.97 < r < 1) in all isolates. Furthermore, the hypothesis test on the correlation coefficient yields a p value < 0.05, indicating that, with 95% confidence, there is a significant linear relationship.
Figure 2.
Growth reduction of K. pneumoniae isolates exposed to CAP (MIC90 of each isolate). A strong and positive linear relationship is observed between the CAP concentration and the percentage of reduction in K. pneumoniae growth, i.e., as the CAP concentration increases, the percentage of reduction in K. pneumoniae growth also increases, which coincides with the Spearman correlation coefficient (0.97 < r < 1) in all isolates. Furthermore, the hypothesis test on the correlation coefficient yields a p value < 0.05, indicating that, with 95% confidence, there is a significant linear relationship.
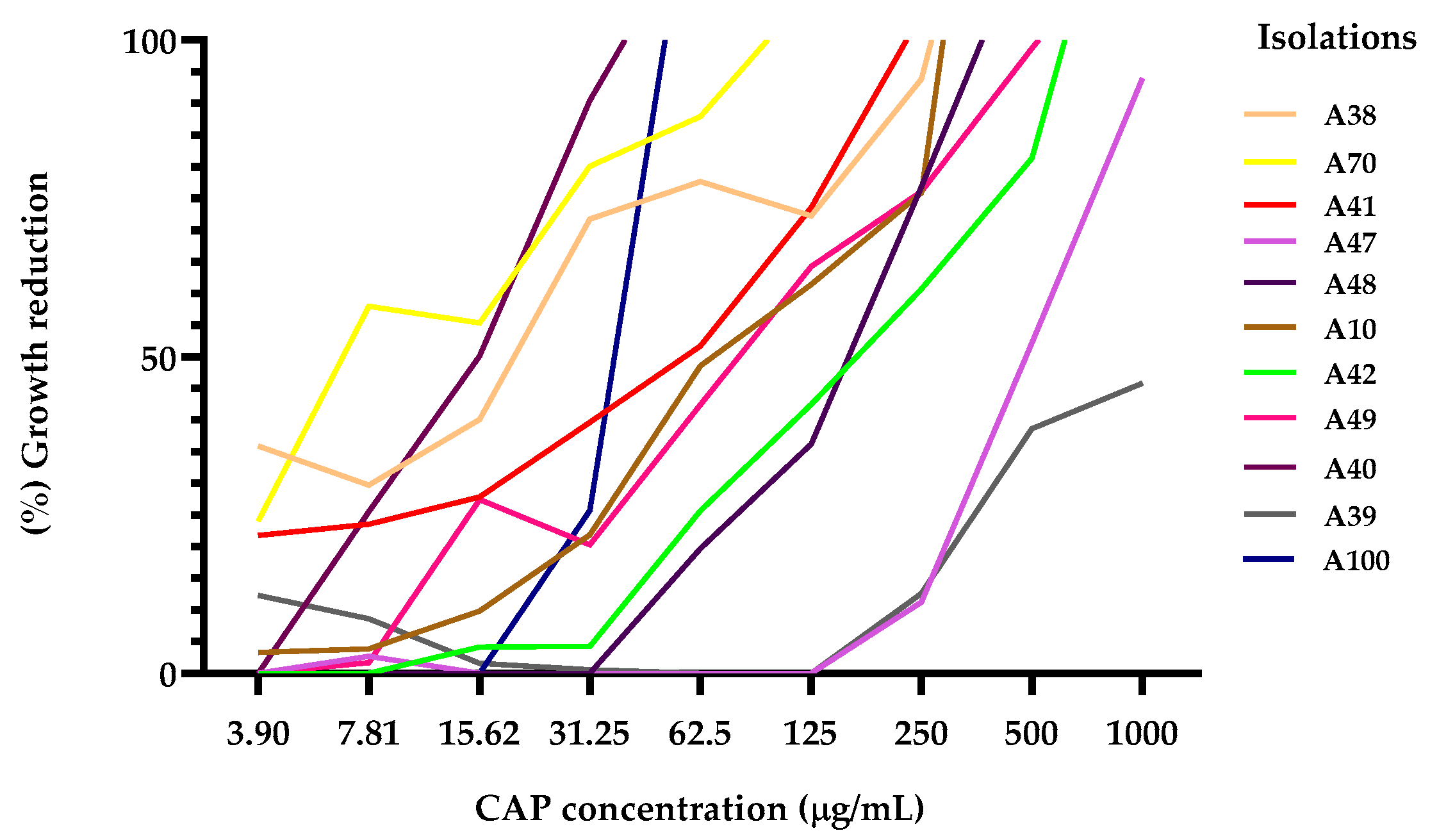
Figure 3.
Action of CAP and ATBs (GEN and CIP) on K. pneumoniae biofilms. (a) Biofilm formation at 37 °C for 24 h, where OD590 > 3 indicates strong biomass production in the biofilms; (b) Percentage reduction of mature biofilms with CAP (MIC90) for each isolate and ATBs (GEN and CIP) (MIC90 µg/mL); (c) Percentage reduction of biofilm formation with CAP (MIC90 for each isolate) and ATBs (GEN and CIP), MIC90 µg/mL. The ANOVA results showing a value of ***p < 0.001 (K. pneumoniae A42) and *p < 0.05 (K. pneumoniae A38, A41 and A48) and the Holm-Sidak's and Games-Howell's tests with a confidence level of 95% indicate that there are significant differences between the effect of CAP and the effect of CIP on the reduction in these isolates.
Figure 3.
Action of CAP and ATBs (GEN and CIP) on K. pneumoniae biofilms. (a) Biofilm formation at 37 °C for 24 h, where OD590 > 3 indicates strong biomass production in the biofilms; (b) Percentage reduction of mature biofilms with CAP (MIC90) for each isolate and ATBs (GEN and CIP) (MIC90 µg/mL); (c) Percentage reduction of biofilm formation with CAP (MIC90 for each isolate) and ATBs (GEN and CIP), MIC90 µg/mL. The ANOVA results showing a value of ***p < 0.001 (K. pneumoniae A42) and *p < 0.05 (K. pneumoniae A38, A41 and A48) and the Holm-Sidak's and Games-Howell's tests with a confidence level of 95% indicate that there are significant differences between the effect of CAP and the effect of CIP on the reduction in these isolates.
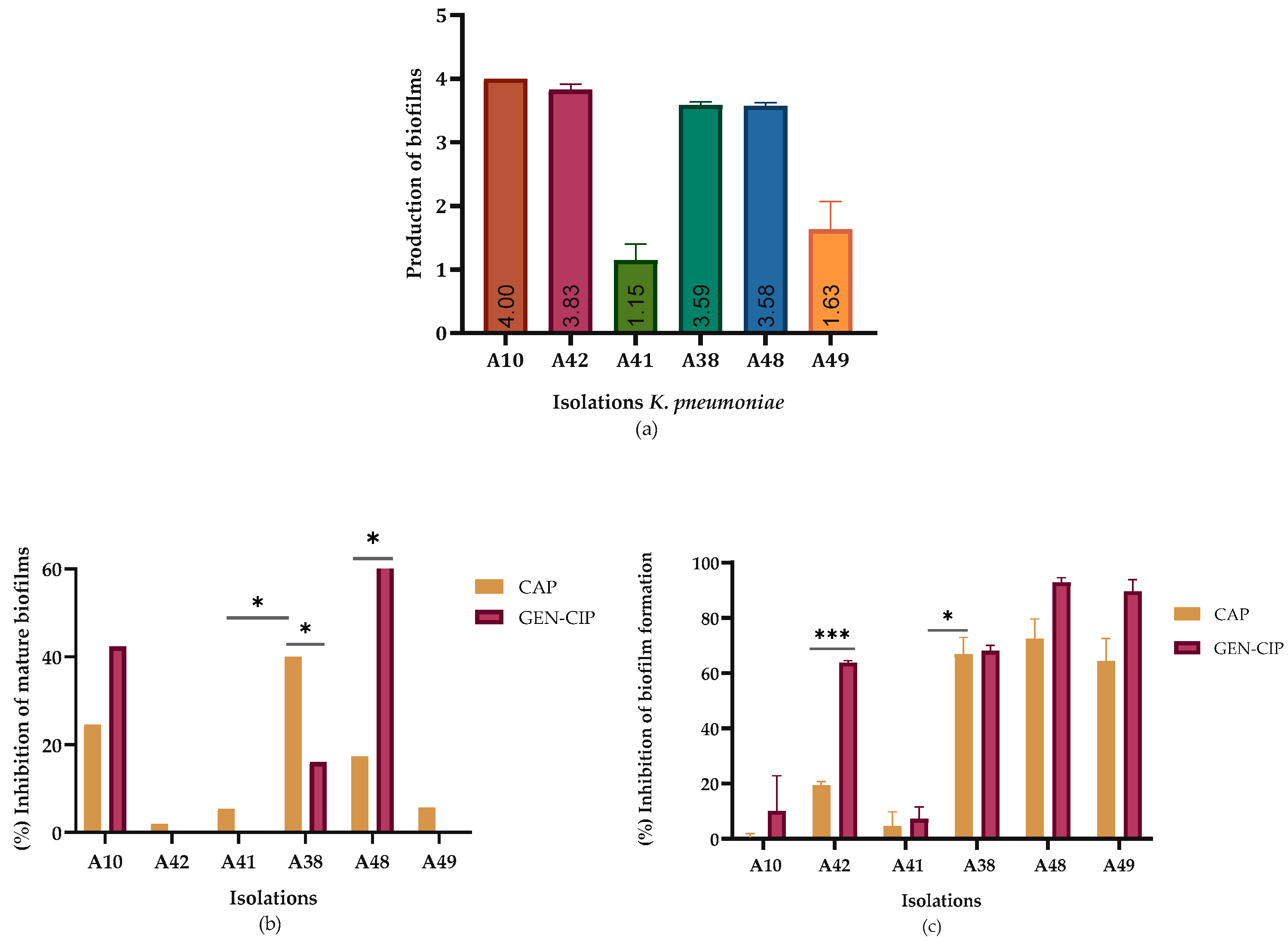
Figure 4.
Time-dependent release of intracellular content at 260/280 nm from K. pneumoniae treated with CAP and GEN (MIC90 µg/mL). The figure shows the OD260/OD280 values of K. pneumoniae A10 treated with CAP and GEN at different times. Results are expressed as the absorbance of the sample (treated with extracts) minus the absorbance of the control (samples without extracts).
Figure 4.
Time-dependent release of intracellular content at 260/280 nm from K. pneumoniae treated with CAP and GEN (MIC90 µg/mL). The figure shows the OD260/OD280 values of K. pneumoniae A10 treated with CAP and GEN at different times. Results are expressed as the absorbance of the sample (treated with extracts) minus the absorbance of the control (samples without extracts).
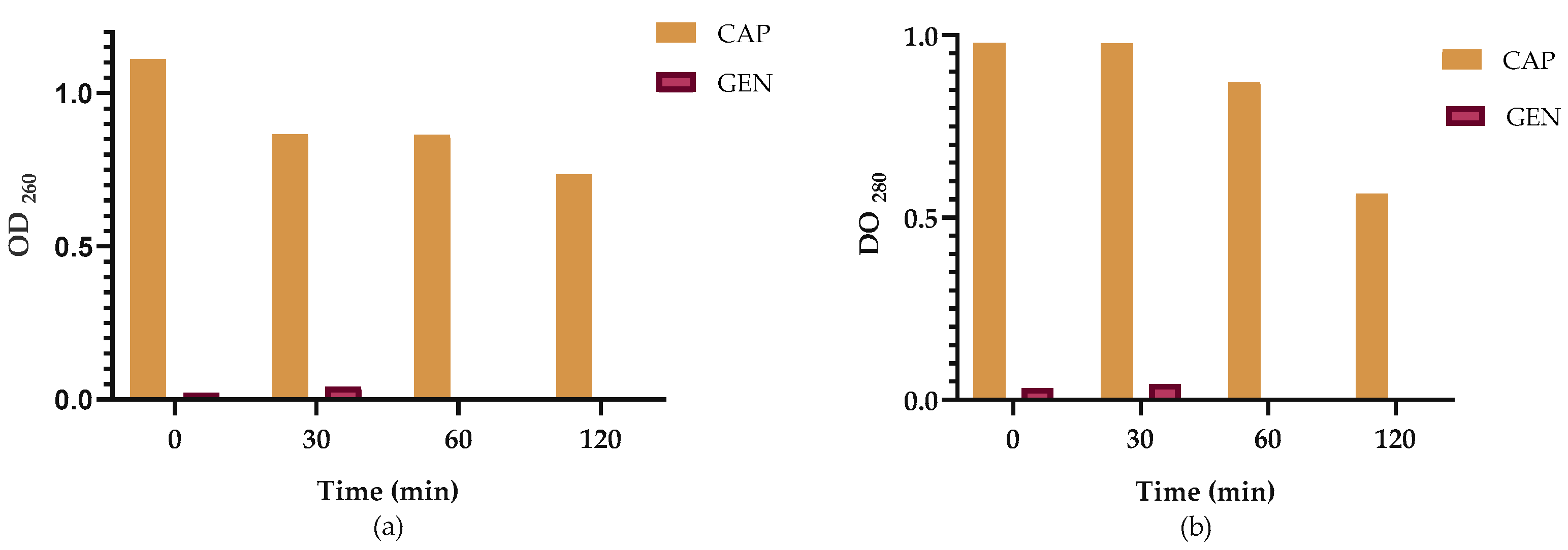
Figure 5.
Microscopic observation (100x) of K. pneumoniae A38, stained with Evans blue, before and after treatment with CAP and CIP; (a) Untreated cells used as control; (b) Cells treated with the MIC90 of CAP; (c) Cells treated with the MIC90 of CIP.
Figure 5.
Microscopic observation (100x) of K. pneumoniae A38, stained with Evans blue, before and after treatment with CAP and CIP; (a) Untreated cells used as control; (b) Cells treated with the MIC90 of CAP; (c) Cells treated with the MIC90 of CIP.


Table 1.
Minimum inhibitory concentration (MIC90) values (µg/mL) of CAP vs antibiotics (ATBs) against K. pneumoniae.
Table 1.
Minimum inhibitory concentration (MIC90) values (µg/mL) of CAP vs antibiotics (ATBs) against K. pneumoniae.
| Isolation of K. pneumoniae | CAP MIC90 | CIP MIC90 | GEN MIC90 |
| A70 | 121.8 | 4.44 | |
| A10* | 276.5 | 0.16 | |
| A48 | 504.1 | 4.55 | |
| A42 | 578.5 | 6.70 | |
| A41 | 191.5 | 1.19 | |
| A38 | 247.8 | 1.49 | |
| A47 | 920.1 | 0.12 | |
| A39 | 1696 | 4.55 | |
| A49 | 385 | 7.19 | |
| A100 | 56.37 | 2.63 | |
| A40 | 28.44 | 2.14 |
ATBs: antibiotics; CAP: capsaicin; CIP: ciprofloxacin; GEN: gentamicin. * Isolate A10 was resistant to CIP, therefore, GEN was used in all experiments with this K. pneumoniae isolate.
Table 2.
Percentages of inhibition of CAP vs ATBs biofilm formation (CIP and GEN) in K. pneumoniae.
| K. pneumoniae isolates | CAP | CIP | GEN |
| A10* | 0.0 | 2.09 | |
| A42 | 19.49 | 63.77 | |
| A41 | 4.26 | 7.40 | |
| A38 | 67.77 | 68.27 | |
| A48 | 72.48 | 92.76 | |
| A49 | 65.26 | 89.96 |
ATBs: antibiotics; CAP: capsaicin; CIP: ciprofloxacin; GEN: gentamicin. *GEN was used with isolate A10 (CIP-resistant).
Table 3.
CAP inhibition percentages against mature biofilms, CAP vs ATBs (CIP and GEN) in K. pneumoniae.
Table 3.
CAP inhibition percentages against mature biofilms, CAP vs ATBs (CIP and GEN) in K. pneumoniae.
| K. pneumoniae isolates | CAP | CIP | GEN |
| A10* | 24.57 | 42.31 | |
| A42 | 1.97 | 0.0 | |
| A41 | 8.45 | 0.00 | |
| A38 | 40.04 | 16.00 | |
| A48 | 16.90 | 63.47 | |
| A49 | 5.69 | 0.0 |
ATBs: antibiotics; CAP: capsaicin; CIP: ciprofloxacin; GEN: gentamicin. *GEN was used with isolate A10 (CIP-resistant).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



