
Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
In restricted regions of the rodent brain, neurogenesis persists throughout life, hinting that perhaps similar phenomena may exist in humans. Neural stem cells (NSCs) that reside within the ventricular-subventricular zone (V-SVZ) continually produce functional cells, including neurons that integrate into the olfactory bulb circuitry. The ability to achieve this feat is based on genetically encoded transcriptional programs that are controlled by environmentally regulated post-transcriptional signaling pathways. One such pathway that molds V-SVZ neurogenesis is the mTOR pathway. This pathway integrates nutrient sufficiency with growth factor signaling to control distinct steps of neurogenesis. Alterations in mTOR pathway signaling occur in numerous neurodevelopmental disorders. Here, we review the role of the mTOR pathway in this process and discuss the use of this region to study the mTOR pathway in both health and disease.
Keywords:
neurogenesis
; mTOR
; TSC
; mTORC1
; TSC1
; TSC2
1. Introduction
Multipotent neural stem cells (NSCs) generate vast quantities and types of neurons within the brain. NSCs produce most neurons embryonically however, they persist at least until the end of infancy in discrete brain regions. Joseph Altman was the first to find groups of dividing cells in the ventricular-subventricular zone (V-SVZ), which surrounds the lateral ventricles (LVs) of the postnatal brain1. These V-SVZ cells were labeled by the DNA precursor thymidine which was passed to cells that mature into GABAergic granule cell neurons (GCs) of the olfactory bulb (OB)2–4 (Figure 1). Others identified that these V-SVZ cells behave as NSCs and generate transit amplifying cells (TACs), which are highly proliferative progenitors5. TACs produce neuroblasts that migrate anteriorly along the rostral migratory stream (RMS) into the OB6. This seminal finding pointed toward the idea that a pool of cells and factors might facilitate neurogenesis in the adult brain. While the extent to which neurogenesis persists postnatally in humans is still unclear, genetic diversity, including pathogenic variants, could prolong neurogenesis in humans. The theoretical benefit of this cellular plasticity is the molding of neuronal circuitry to adapt to environmental conditions. In theory, the tradeoff of continued cell division is the genetic vulnerability of having an increased mutational burden and susceptibility to errors in building circuits. Thus, intrinsic genetically encoded mechanisms and environmental signals must carefully balance the costs and benefits of ongoing neurogenesis. The presence of NSCs within the V-SVZ provides a unique opportunity to study signaling pathways and to model diseases. This opportunity is derived from the fact that neonatal LVs are readily accessible for the delivery of molecules, proteins, plasmids, and viruses to manipulate NSCs. Moreover, no surgery is required. Owing to their sessile nature and anatomically restricted positions, specific pools of NSCs can be manipulated to study normal physiological and pathophysiological responses. Here, we discuss the cytoarchitectonic map of the V-SVZ, the cell types generated by NSCs, and the effect of altering a critical regulator of cell and circuit development, the mTOR signaling pathway within different cell types.
2. Heterogeneity in the V-SVZ
The V-SVZ is a three-dimensional topographic compartment having a cytoarchitecture that is patterned and arealized along a fourth dimension, time7. Patterning of the V-SVZ is likely the remnant of the morphogenic gradients and transcriptional identities of NSCs that exist during embryogenesis8. The result is that different regions of the postnatal V-SVZ generate different cell types. For example, dorsal radial glia are a type of NSC in the embryonic brain that generate cortical excitatory neurons starting around embryonic day (E) 12 in mice9. However, in the perinatal brain, they produce glia10,11. Later, dorsal NSCs in the V-SVZ produce dopaminergic periglomerular (PG) neurons in the OB12. This appears directly related to precise transcriptional programs dictated by the expression of the transcription factors Pax6 and Dlx212. Dorsal NSCs may also generate some excitatory glutamatergic PGs that express the transcription factor Tbr113. In comparison, NSCs at the medial-septal wall produce calretinin positive PGs and GCs14. On the other hand, the lateral wall and dorsolateral corner of the ventricles produce OB GCs and dopaminergic PGs14. There are now recognized V-SVZ microdomains that are responsible for generating cells having unique GC laminar positioning and morphologies15. For example, a small ventrally located NKX2.1 microdomain derived from the embryonic medial ganglionic eminence generates postnatal GCs16. The lateral ganglionic eminence (LGE) also contains microdomains that produce striatal glia and OB GCs17.While both populations of NSCs express Dlx5/6 in the LGE, Isl1+ cells migrate to the striatum whereas Er81+ cells generate OB GCs. Transplantation experiments further support the likelihood that the potential and capacity to generate specific cell types is eventually independent of postnatal location and mostly specified by transcriptional programs established embryonically14. This is analogous to the cell intrinsic mechanism of NSC specification in the cortex18. On the other hand, the programs that regulate V-SVZ NSC pools have yet to be fully recognized. For example, PDGFRβ-positive stem cells within the septal and dorso-septal wall generate oligodendrocyte progenitor cells (OPCs)19. In addition, a newer intraventricular localized pool of stem cells within the lumen of the lateral ventricles was identified in the same study19. These findings indicate that diverse NSCs are found along the ventricular walls and produce different cell progeny.
3. Targets of Postnatal Neurogenesis
It takes nearly 1 week for neuroblasts to migrate from the V-SVZ to the OB20,21. There, 94% of neuroblasts immediately begin to mature into GCs as evidenced by the production of basal dendrites that form synapses with centrifugal fibers and receive GABAergic input within 14 days22–24. GC apical dendrites project into the external plexiform layer, where reciprocal synapses start by 21 days, and glutamatergic input occurs approximately one month after GCs have been generated in the V-SVZ22,23,25. GCs are found within the GC layer, mitral/tufted cell layer, and the external plexiform layer15. The newly born GCs have distinct basal and apical dendrite morphologies and laminar positions, albeit the developmental relationship between the morphologies of these cells has not been completely characterized14,15. For example, many of the morphologies overlap at different developmental periods, shorter dendrites may reflect that these “different GCs” represent different states of the same cell, such as stages of development. However, these cells also express select proteins including calretinin and calbindin indicating that both the morphological and molecular characteristics change.
GCs form dendrodendritic synapses within the external plexiform layer that lies beneath glomeruli26,27. There, the dendrites of mitral cells synapse with the dendrites of GCs28. Most GCs are axon-less inhibitory neurons26. The release of glutamate from mitral/tufted cell dendrites caused by orthodromic retrograde backpropagating potential activates GCs that subsequently release GABA from their dendrites28. GC dendrite tiling prevents signaling between nearby mitral cells from different glomeruli. The lateral inhibition of mitral cells mediated by GC therefore increases contrast between signals. The addition of new GCs and dendrites to the circuitry optimizes olfactory processing24. The significance of this is that continued addition of GCs in the OB is a form of structural plasticity that facilitates perceptual learning29.
Postnatal V-SVZ NSCs may also produce additional groups of functional neurons. For example, a seminal finding found that lateral adult V-SVZ NSCs generate striatal GABAergic neurons in humans and this is reduced in Huntington’s disease30. The ability of V-SVZ NSCs to produce striatal neurons is evolutionarily conserved, including in rodents and rabbits. In rats, the production of GABAergic striatal neurons that are parvalbumin positive starts around postnatal (P) 9 and progressively increases for an additional 2-3 weeks whereas the production of calretinin neurons peaks at P531. Ventral V-SVZ NSCs also continually produce neurons within the nucleus accumbens of mice32. Striatal neurogenesis is subject to environmental regulation, including pathological insults. For example, middle cerebral artery occlusion causes neuroblasts to migrate from the SVZ into the adult striatum33. The mechanism by which new striatal neurons are added in the mouse may differ under normal physiological versus pathological conditions.
The human V-SVZ also appears to produce neurons until ~18 months34,35. An evolutionarily novel bifurcation of the RMS called the medial migratory stream (MMS) pours into the ventromedial prefrontal cortex34. Details regarding the integration, maturation, and function of MMS-derived neurons are unknown. While rodents have additional postnatal waves generating GABAergic neurons that leave the RMS to populate the lower cortical layers and integrate into medial prefrontal, cingulated, and infralimbic cortices, their significance remains unclear36. It appears that dorsal V-SVZ NSCs in rodents can also produce cortical astrocytes37.
4. Techniques for Studying V-SVZ NSCs
We will briefly summarize the use of various methods to manipulate NSCs and daughter cells such as, components injected in the LVs including drugs or extracellular vesicles (EVs), neonatal electroporation, and viruses. Postnatal V-SVZ NSCs have a basal fiber that projects to the vasculature and an apical projection that interdigitates between ependyma and the ventricular lumen forming the hub of a pinwheel structure38. This ventricular contact allows one to manipulate NSCs. Injection of different components (e.g., drugs or plasmids) into the ventricles allows the determination of their effects on the behavior of NSCs and the production of cellular progeny. For example, the intraventricular injection of soluble factors into CSF within the LVs has been used to test the effect of growth factors on neurogenesis39. Lipophilic dyes and fluorescent beads have also been injected into the ventricle to label NCSs and study neurogenesis40. EVs have also been injected into the LVs and were taken up by microglia. The EVs were engineered to contain specific microRNAs acting as a microglia morphogen41. Consistent with this, it was demonstrated that NSCs release EVs that are taken up by microglia41. However, there are some limitations to intraventricular injections. First, the adult LVs are surrounded by multi-ciliated ependymal cells, which may prevent efficient uptake of some components. Second, the flow of CSF throughout the ventricular system results in the diffusion of injected components, which can affect many cell types in other brain regions. An example of an affected cell type are choroid plexus epithelial cells, which generate CSF and are found within the LV. Injection of CRE recombinase fused to a TAT peptide into the lateral ventricles demonstrated the selective uptake into choroid plexus epithelial cells42,43. Thus, injection into the LVs has the capacity to affect cells besides NSCs. Another approach that has been extensively used is the neonatal electroporation of plasmid DNA into the ventricles13,44,45. While this approach lacks specificity in uptake besides being restricted to cells with ventricular contact (i.e., ependymal cells versus NCS), the use of specific promoters or the introduction of inducible plasmids into mice having CRE expression within NSCs can overcome this46. Another option is to sort cells or nuclei using markers to study specific cell populations but this requires generating transgenic mice and large numbers of mice due to the small size of the V-SVZ47. An important limitation of using episomal plasmids is that they are diluted in dividing cells following electroporation48,49. Some NSCs may retain the plasmids long-term especially if they become quiescent. Electroporation of CRE recombinase or transposases can alter genomic DNA and prevents this dilution issue50,51. In addition, dilution may be an advantage. For example, the electroporation of tamoxifen-inducible CRE-ERT2 and a conditional plasmid into NSCs is accompanied by rapid dilution from actively dividing NSCs leading to plasmids expression only in the first cohort of daughter cells, including OB GCs. Injection of tamoxifen weeks later can allow for selective recombination in mature GCs allowing to manipulate them and distinguish their roles in the OB circuit from other cell types52. Another limitation of electroporation is that the injection of plasmid DNA into the LVs may cause immune reactions of intraventricular epiplexus immune cells53. The LVs, choroid plexus, and V-SVZ are enriched in immune cells during the perinatal period41. Another way to manipulate NSCs and their progeny postnatally is to use promoter driven tamoxifen inducible CRE-ERT2 mice. A popular example is the use of mice containing the rat nestin promoter driven CRE-ERT254 since nestin is expressed in NSCs. Finally, a last approach to manipulate NSCs and neurogenesis is the use of viral targeting40. In mature mice, there is not extensive proliferation besides in the neurogenic zones allowing for the use of retroviruses. Replication deficient retroviruses can be used to genomically modify NSCs55. This in theory will not lead to targeting quiescent NSCs, which is a limitation for studying the mechanisms that push quiescent NSCs to become active. Other types of viruses such as adenovirus were injected into the parenchyma of the V-SVZ to label NSCs and demonstrate that specific microdomains generate different types of neurons14. ,
5. The mTOR Pathway
Rapamycin was originally discovered in 1965 as a compound derived from Streptomyces hygroscopicus, a bacterium found on Easter Island (also known as Rapa Nui). Initially, it was reported that rapamycin had antifungal properties and was used an antifungal agent. 56. Later on, it was shown that rapamycin works by causing the FK506-binding protein (FKBP12) to bind to and inhibit the mammalian target of rapamycin (mTOR)57. mTOR is a catalytic subunit of two heteromeric kinases termed mTOR complex (mTORC) 1 and 258. Rapamycin causes immediate and partial inhibition of mTORC1, however, at greater doses and over time may inactivate mTORC257,59–64. Each mTORC contains two mTOR kinase domains containing complex specific adapters65–67. The mTORC1 adapter is RAPTOR whereas RICTOR is the mTORC2 adapter68,69.
mTORC1 stimulates cell growth by inducing cap-dependent mRNA translation. This is achieved by mTORC1 inhibitory phosphorylation of eukaryotic initiation factor 4E (eIF4E) binding protein (4EBP)70,71 and phosphorylation of p70S6 kinase (p70S6K), which activates the ribosomal protein S6 (S6)60,72. mTORC1 is activated by GTP bound RHEB73–76. The GTPase RHEB is inhibited by tuberin (TSC2) and hamartin (TSC1), which are encoded by TSC1 and TSC273–76. Tuberin has a catalytic GAP activity that causes RHEB to hydrolzye GTP75. Hamartin and TBC1D7 seem to stabilize Tuberin77. Loss of hamartin or tuberin can cause abnormal brain development as discussed below.
6. Tuberous Sclerosis Complex (TSC)
Patients who are afflicted with TSC are frequently born with malformations of cortical development as well as abnormal growths along the LVs that were both originally called hamartomas78,79. The growths along the V-SVZ are subependymal nodules (SEN), which are benign slow growing tumors. Once these nodules reach a specific size, they are called subependymal giant cell astrocytomas (SEGAs)80. The underlying cause of TSC is inactivating mutations in the TSC1 or TSC2 genes found on chromosomes 9q34 and 16p13.3, respectively81,82. These mutations, including nonsense and missense mutations, deletions, and large rearrangements, cause the loss of TSC1 or TSC283. and uncontrolled mTORC1 activity83,84. Although the events causing V-SVZ SEN and SEGA remain unclear, some mechanisms can be appreciated from studying the role of mTOR in neurogenesis.
7. The TSC-mTORC1 Pathway in V-SVZ Neurogenesis
During development, cellular identity is tightly linked to the expression of distinct transcription factors that promote the availability of mRNAs. Indeed, single cell and single nuclei sequencing have consistently demonstrated the importance of transcriptional programs for establishing identity. This is no different in the postnatal V-SVZ85–87. Since growth factors such as EGF can titrate neurogenesis, environmental signals appear to be overlayed onto genetically encoded programs39. Thus, post-transcriptional mechanisms, including regulation of translation could in theory mold neurogenesis. The uncoupling of mRNA availability from translation occurs as human NSC differentiate into neurons88. This is important because cells might need to rapidly stop translating NSC mRNAs. Many of the differentially translated mRNAs are regulated by mTORC1. Indeed mTORC1 controls mRNA translation at different phases of V-SVZ neurogenesis89. For example, translation of the stem cell factor Sox2 mRNA is regulated by mTORC189. Persistent activation of mTORC1 causes aberrant Sox2 translation90. Therefore, controlling mTORC1-dependent mRNA translation occurs during neurogenesis.
The ability of extracellular factors such as EGF to regulate neurogenesis spurred investigations that identified mTORC1 hyperactivity in dividing Ki67+ cells and Mash1+ TACs produced by NSCs91. Rapamycin treatment reduced the density of TACs and dividing cells. In culture, rapamycin also induced quiescence in neurospheres and prevented neural differentiation91. It is important to mention that rapamycin does not completely inhibit mTORC1 phosphorylation of all substrates92. Thus, how mTORC1 inhibition affects these events is unclear. Despite this, it was known that quiescent NSCs become proliferative when they express EGFR. In fact, introducing mutant active EGFR pushes quiescent NSCs (qNSCs) to divide93. However, tumors are not generated by mutant active EGFR expression alone94. Indeed, EGF infusion into the ventricle only generates V-SVZ (subependymal) polyps 39. Subsequent experiments demonstrated that as NSCs go from a quiescent to an activated state, they activate mTORC146. This is consistent with the fact that tuberin is inhibited by CDK4/6 during the cell cycle and mTORC1 is activated95. Rapamycin also blocked NSC production of TACs and 4E-BP phosphorylation46. This study also demonstrated that RHEB knockdown prevents NSCs from generating TACs and neurons whereas as phospho (mTOR)-resistant 4E-BP1 enhanced NSC self-renewal46. The interpretation of this data is that RHEB activation of mTORC1 and phosphorylation of 4E-BP1 allows for NSCs to undergo symmetric terminal division producing highly proliferative TACs and neurons. This occurs at the expense of self-renewing divisions and maintenance of the NSC pool. RHEB activation of mTORC1 and phosphorylation of 4E-BP1 does not cause NSCs to divide but rather differentiate. These results indicate that mTORC1 is necessary for balancing NSC self-renewal and production of TACs. Excessive mTORC1 activity might in theory cause NSCs to become depleted. Consistent with this hypothesis, Tsc1 deletion from V-SVZ NSCs reduces the size of the OB GC layer over time96.
Tsc1 in qNSCs does not appear to push them to divide as reported with expression of a constitutively active RHEB97. Removal of Tsc1 from V-SVZ NSCs does however cause widespread alterations, including V-SVZ SEN and heterotopic nodules along the RMS, in the OB, and in the striatum96,98. Mice also had ventrally located nodules and developed hydrocephalus96. Ventral and dorsal V-SVZ NSCs vary in mTORC1 activity99. Considering this, the effect of loss of Tsc2 in ventral or dorsal NSCs was examined using transgenic mice expressing Nkx2-1 or Emx1 promoter-driven CRE. This study found that Nkx2-1 deletion of Tsc2 preferentially caused nodules to form along the lateral V-SVZ99. Emx1 is expressed in NSCs that generate excitatory cortical neurons and in postnatal dorsolateral V-SVZ NSCs that generate OB GCs100,101. As expected, Emx1-CRE Tsc1 deletion expanded the V-SVZ and RMS and caused a disorganized OB GC layer which was more spread out102. More Tsc1 mutant dorsolateral NSCs retained BrdU or were Ki67 at early stages and this effect was reduced at later ages. This is also consistent with the effects of mTORC1 hyperactivity on balancing self-renewing cell divisions with exhausting terminal cell divisions. Likewise, loss of Tsc2 by CRE electroporation (in conditional Tsc2 mice) also caused SENs, including those that protrude or are found within the ventricles103. One relevant finding from this study was that striatal astrocytes that are typically generated from the V-SVZ NSCs did not fully differentiate in the striatum and spuriously translated mRNAs associated with stemness following Tsc2 deletion103. These immature cells appeared to aberrantly produce neurons having a striatal-like morphology. The aberrant expression of immature markers in cortical tubers is also documented in TSC patients104. These results further support the need to tightly regulate mTORC1 activity at precise steps of development to prevent the expression of immature proteins in differentiated cells. In addition to loss of TSC genes, simultaneous Tsc1/Pten deletion has been performed using Nestin-CRE-ERT2 mice102. Mice injected with tamoxifen at P10 had nodular/bulbous nodules along the ventricles ~30 days later and often died likely due to cerebellar developmental complications105. To circumvent this issue, tamoxifen was injected later (P15 or P24) followed by examination of the brains months later. Upon inspection, SENs reminiscent of those in TSC patients had developed along the caudate nucleus of the striatum105.
The neuroblasts that are generated from V-SVZ NSCs have low mTORC1 activity and mRNA translation46,90,91. The presence of heterotopic clusters of neurons within the RMS following removal of Tsc1 from V-SVZ NSCs supports a role in migration and mirrors defects of cortical lamination caused by Tsc1 deletion from cortical radial glia48,98. Tsc1 null neuroblasts seemed to migrate slower than heterozygous neuroblasts but in fact likely lost direction due to the abnormal formation of multiple processes98. The migratory defects were mimicked by neuroblasts expressing constitutively active RHEB which also led to ectopic clusters of neurons106. At the molecular level, in vitro analysis of Tsc1 null neuroblasts demonstrated that there was a reduction in autophagic flux and impaired nuclear import of the transcription factor TFEB 107. Restoring TFEB activation in Tsc1 null neuroblasts rescues the migratory defect107.
Many of the neuroblasts that did not reach the OB became GCs in the RMS and their somas become enlarged following Tsc1 or Tsc2 removal or expression of a constitutively active RHEB using neonatal electroporation52,98,106,108 These changes all increased mTORC1 signaling as determined by phosphorylated S6 staining. The involvement of mTOR was demonstrated by neonatal CRE electroporation of mice having conditional mTOR leading to reduction of soma sizes109. While removal of mTOR does not reveal which mTORC is responsible for this effect, rapamycin treatment phenocopies the reduction in size suggesting a role of mTORC1109.
Since neurogenesis is ongoing, it is unclear whether affecting the mTORC1 pathway in mature GCs would have the same effect as changing it in NSCs. In one study, inducible CRE-ERT2 and GFP plasmids were electroporated into neonatal V-SVZ NSCs of Tsc2 conditional mice. The plasmid DNA (indicated by GFP) was diluted from actively dividing cells and ~30 days later, mice were injected with tamoxifen leading to Tsc2 deletion and RFP expression upon recombination in mature GCs. Tsc2 removal had a smaller effect on mTORC1, soma size, and dendrite arbors than when recombination was initiated in neonatal V-SVZ NSCs52. This is consistent with the idea that there may be discrete periods in which Tsc1/2 play more prominent roles110. What signals are turning Tsc1/2 or mTORC1 on and off during these periods is unclear. One possibility is that amino acids such as leucine, which is transported by Slc7a5 and additional transporters, activate mTORC1 and are critical, as their loss leads to OB GC death108,111.
These studies also demonstrated that loss of Tsc1, Tsc2, or increasing Rheb activity, enhanced dendrite arbors of OB GCs52,98,112. mTOR complex components raptor and rictor are well-known to be required for the development of GC dendrite arbors109. mTOR inhibition by rapamycin rescues the increased growth caused by expressing constitutively active Rheb113. Thus, the connectivity of mutant GCs might also be changed when the TSC-mTORC1 pathway is altered. Indeed, overactive Rheb also increased the frequency of inhibitory postsynaptic currents in ectopic GCs106, suggesting an increase in GABAergic synaptic connections. While the electrophysiological properties of action potentials in these mutant OB GCs did not change, the resting membrane potential was hyperpolarized. Further experiments are needed to determine how OB neuron firing, network activity, and olfaction are affected in these models, which could yield insight into the sensory changes that occur in patients.
8. Conclusions
In conclusion, the V-SVZ contains a mosaic of NSCs that produce different cell types owing to their embryonic ontogeny. NSCs may be easily altered, including through genetic manipulations to study molecules that may be relevant in normal developmental processes or in specific diseases. This is exemplified by studies that have examined the role of the TSC-mTORC1 pathway in the context of V-SVZ neurogenesis.
Acknowledgments
DMF has received support from the Tuberous Sclerosis Complex Alliance and the United States of America Department of Defense U.S. Army Medical Research Activity Award Congressionally Directed Medical Research Program Tuberous Sclerosis Complex Research Program W81XWH2010447.
References
- Altman, J.; Das, G.D. Post-Natal Origin of Microneurones in the Rat Brain. Nature 1965, 207, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Altman, J. Autoradiographic study of degenerative and regenerative proliferation of neuroglia cells with tritiated thymidine. Exp. Neurol. 1962, 5, 302–318. [Google Scholar] [CrossRef]
- Altman, J.; Das, G.D. Postnatal Neurogenesis in the Guinea-pig. Nature 1967, 214, 1098–1101. [Google Scholar] [CrossRef]
- Altman, J. Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J. Comp. Neurol. 1969, 137, 433–457. [Google Scholar] [CrossRef] [PubMed]
- Doetsch, F.; García-Verdugo, J.M.; Alvarez-Buylla, A. Cellular Composition and Three-Dimensional Organization of the Subventricular Germinal Zone in the Adult Mammalian Brain. J. Neurosci. 1997, 17, 5046–5061. [Google Scholar] [CrossRef]
- Luskin, M.B. Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 1993, 11, 173–189. [Google Scholar] [CrossRef]
- Ihrie, R.A.; Álvarez-Buylla, A. Lake-Front Property: A Unique Germinal Niche by the Lateral Ventricles of the Adult Brain. Neuron 2011, 70, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Casingal, C.R.; Descant, K.D.; Anton, E. Coordinating cerebral cortical construction and connectivity: Unifying influence of radial progenitors. Neuron 2022, 110, 1100–1115. [Google Scholar] [CrossRef]
- Kwan, K.Y.; Šestan, N.; Anton, E.S. Transcriptional co-regulation of neuronal migration and laminar identity in the neocortex. Development 2012, 139, 1535–1546. [Google Scholar] [CrossRef]
- Marshall, C.A.; Suzuki, S.O.; Goldman, J.E. Gliogenic and neurogenic progenitors of the subventricular zone: Who are they, where did they come from, and where are they going? Glia 2003, 43, 52–61. [Google Scholar] [CrossRef]
- Kriegstein, A.; Alvarez-Buylla, A. The Glial Nature of Embryonic and Adult Neural Stem Cells. Annu. Rev. Neurosci. 2009, 32, 149–184. [Google Scholar] [CrossRef] [PubMed]
- de Chevigny, A.; Core, N.; Follert, P.; Wild, S.; Bosio, A.; Yoshikawa, K.; Cremer, H.; Beclin, C. Dynamic expression of the pro-dopaminergic transcription factors Pax6 and Dlx2 during postnatal olfactory bulb neurogenesis. Front. Cell. Neurosci. 2012, 6, 6. [Google Scholar] [CrossRef]
- Winpenny, E.; Lebel-Potter, M.; E Fernandez, M.; Brill, M.S.; Götz, M.; Guillemot, F.; Raineteau, O. Sequential generation of olfactory bulb glutamatergic neurons by Neurog2-expressing precursor cells. Neural Dev. 2011, 6, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Merkle, F.T.; Mirzadeh, Z.; Alvarez-Buylla, A. Mosaic Organization of Neural Stem Cells in the Adult Brain. Science 2007, 317, 381–384. [Google Scholar] [CrossRef]
- Merkle, F.T.; Fuentealba, L.C.; A Sanders, T.; Magno, L.; Kessaris, N.; Alvarez-Buylla, A. Adult neural stem cells in distinct microdomains generate previously unknown interneuron types. Nat. Neurosci. 2014, 17, 207–214. [Google Scholar] [CrossRef]
- Delgado, R.N.; Lim, D.A. Embryonic Nkx2.1-expressing neural precursor cells contribute to the regional heterogeneity of adult V–SVZ neural stem cells. Dev. Biol. 2015, 407, 265–274. [Google Scholar] [CrossRef]
- Stenman, J.; Toresson, H.; Campbell, K. Identification of Two Distinct Progenitor Populations in the Lateral Ganglionic Eminence: Implications for Striatal and Olfactory Bulb Neurogenesis. J. Neurosci. 2003, 23, 167–174. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, Y.; Dimos, J.T.; A Fasano, C.; Phoenix, T.N.; Lemischka, I.R.; Ivanova, N.B.; Stifani, S.; E Morrisey, E.; Temple, S. The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells. Nat. Neurosci. 2006, 9, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.C.; Maldonado-Soto, A.R.; Silva-Vargas, V.; Mizrak, D.; von Känel, T.; Tan, K.R.; Paul, A.; Madar, A.; Cuervo, H.; Kitajewski, J.; et al. Release of stem cells from quiescence reveals gliogenic domains in the adult mouse brain. Science 2021, 372, 1205–1209. [Google Scholar] [CrossRef]
- Lois, C.; Alvarez-Buylla, A. Long-Distance Neuronal Migration in the Adult Mammalian Brain. Science 1994, 264, 1145–1148. [Google Scholar] [CrossRef]
- Doetsch, F.; Alvarez-Buylla, A. Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. 1996, 93, 14895–14900. [Google Scholar] [CrossRef]
- Whitman, M.C.; Greer, C.A. Synaptic Integration of Adult-Generated Olfactory Bulb Granule Cells: Basal Axodendritic Centrifugal Input Precedes Apical Dendrodendritic Local Circuits. J. Neurosci. 2007, 27, 9951–9961. [Google Scholar] [CrossRef]
- Belluzzi, O.; Benedusi, M.; Ackman, J.; LoTurco, J.J. Electrophysiological Differentiation of New Neurons in the Olfactory Bulb. J. Neurosci. 2003, 23, 10411–10418. [Google Scholar] [CrossRef]
- Sailor, K.A.; Valley, M.T.; Wiechert, M.T.; Riecke, H.; Sun, G.J.; Adams, W.; Dennis, J.C.; Sharafi, S.; Ming, G.-L.; Song, H.; et al. Persistent Structural Plasticity Optimizes Sensory Information Processing in the Olfactory Bulb. Neuron 2016, 91, 384–396. [Google Scholar] [CrossRef]
- Kelsch, W.; Lin, C.-W.; Lois, C. Sequential development of synapses in dendritic domains during adult neurogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 16803–16808. [Google Scholar] [CrossRef]
- Shepherd, G.M.; Chen, W.R.; Willhite, D.; Migliore, M.; Greer, C.A. The olfactory granule cell: From classical enigma to central role in olfactory processing. Brain Res. Rev. 2007, 55, 373–382. [Google Scholar] [CrossRef]
- Rall, W.; Shepherd, G.; Reese, T.; Brightman, M. Dendrodendritic synaptic pathway for inhibition in the olfactory bulb. Exp. Neurol. 1966, 14, 44–56. [Google Scholar] [CrossRef]
- Lepousez, G.; Valley, M.T.; Lledo, P.-M. The Impact of Adult Neurogenesis on Olfactory Bulb Circuits and Computations. Annu. Rev. Physiol. 2013, 75, 339–363. [Google Scholar] [CrossRef]
- Moreno, M.M.; Linster, C.; Escanilla, O.; Sacquet, J.; Didier, A.; Mandairon, N. Olfactory perceptual learning requires adult neurogenesis. Proc. Natl. Acad. Sci. 2009, 106, 17980–17985. [Google Scholar] [CrossRef]
- Ernst, A.; Alkass, K.; Bernard, S.; Salehpour, M.; Perl, S.; Tisdale, J.; Possnert, G.; Druid, H.; Frisén, J. Neurogenesis in the Striatum of the Adult Human Brain. Cell 2014, 156, 1072–1083. [Google Scholar] [CrossRef]
- Klausa, G.; Prime, G.; Bruggencate, G.T.; Schlösser, B. Postnatal development of calretinin- and parvalbumin-positive interneurons in the rat neostriatum: An immunohistochemical study. J. Comp. Neurol. 1999, 405, 185–198. [Google Scholar] [CrossRef]
- García-González, D.; Dumitru, I.; Zuccotti, A.; Yen, T.-Y.; Herranz-Pérez, V.; Tan, L.L.; Neitz, A.; García-Verdugo, J.M.; Kuner, R.; Alfonso, J.; et al. Neurogenesis of medium spiny neurons in the nucleus accumbens continues into adulthood and is enhanced by pathological pain. Mol. Psychiatry 2021, 26, 4616–4632. [Google Scholar] [CrossRef]
- Yamashita, T.; Ninomiya, M.; Acosta, P.H.; García-Verdugo, J.M.; Sunabori, T.; Sakaguchi, M.; Adachi, K.; Kojima, T.; Hirota, Y.; Kawase, T.; et al. Subventricular Zone-Derived Neuroblasts Migrate and Differentiate into Mature Neurons in the Post-Stroke Adult Striatum. J. Neurosci. 2006, 26, 6627–6636. [Google Scholar] [CrossRef]
- Sanai, N.; Nguyen, T.; Ihrie, R.A.; Mirzadeh, Z.; Tsai, H.-H.; Wong, M.; Gupta, N.; Berger, M.S.; Huang, E.; Garcia-Verdugo, J.-M.; et al. Corridors of migrating neurons in the human brain and their decline during infancy. Nature 2011, 478, 382–386. [Google Scholar] [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular–Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018820. [Google Scholar] [CrossRef]
- Inta, D.; Alfonso, J.; von Engelhardt, J.; Kreuzberg, M.M.; Meyer, A.H.; van Hooft, J.A.; Monyer, H. Neurogenesis and widespread forebrain migration of distinct GABAergic neurons from the postnatal subventricular zone. Proc. Natl. Acad. Sci. 2008, 105, 20994–20999. [Google Scholar] [CrossRef]
- Ge, W.-P.; Miyawaki, A.; Gage, F.H.; Jan, Y.N.; Jan, L.Y. Local generation of glia is a major astrocyte source in postnatal cortex. Nature 2012, 484, 376–380. [Google Scholar] [CrossRef]
- Mirzadeh, Z.; Merkle, F.T.; Soriano-Navarro, M.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Neural Stem Cells Confer Unique Pinwheel Architecture to the Ventricular Surface in Neurogenic Regions of the Adult Brain. Cell Stem Cell 2008, 3, 265–278. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Winkler, J.; Kempermann, G.; Thal, L.J.; Gage, F.H. Epidermal Growth Factor and Fibroblast Growth Factor-2 Have Different Effects on Neural Progenitors in the Adult Rat Brain. J. Neurosci. 1997, 17, 5820–5829. [Google Scholar] [CrossRef]
- Doetsch, F.; Caillé, I.; Lim, D.A.; García-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular Zone Astrocytes Are Neural Stem Cells in the Adult Mammalian Brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef]
- Morton, M.C.; Neckles, V.N.; Seluzicki, C.M.; Holmberg, J.C.; Feliciano, D.M. Neonatal Subventricular Zone Neural Stem Cells Release Extracellular Vesicles that Act as a Microglial Morphogen. Cell Rep. 2018, 23, 78–89. [Google Scholar] [CrossRef]
- Peitz, M.; Pfannkuche, K.; Rajewsky, K.; Edenhofer, F. Ability of the hydrophobic FGF and basic TAT peptides to promote cellular uptake of recombinant Cre recombinase: A tool for efficient genetic engineering of mammalian genomes. Proc. Natl. Acad. Sci. USA 2002, 99, 4489–4494. [Google Scholar] [CrossRef]
- Spatazza, J.; Lee, H.H.; Di Nardo, A.A.; Tibaldi, L.; Joliot, A.; Hensch, T.K.; Prochiantz, A. Choroid-Plexus-Derived Otx2 Homeoprotein Constrains Adult Cortical Plasticity. Cell Rep. 2013, 3, 1815–1823. [Google Scholar] [CrossRef]
- Boutin, C.; Diestel, S.; Desoeuvre, A.; Tiveron, M.-C.; Cremer, H. Efficient In Vivo Electroporation of the Postnatal Rodent Forebrain. PLOS ONE 2008, 3, e1883. [Google Scholar] [CrossRef]
- Feliciano, D.M.; Lafourcade, C.A.; Bordey, A. Neonatal Subventricular Zone Electroporation. J. Vis. Exp. 2013, e50197–e50197. [Google Scholar] [CrossRef]
- Hartman, N.W.; Lin, T.V.; Zhang, L.; Paquelet, G.E.; Feliciano, D.M.; Bordey, A. mTORC1 Targets the Translational Repressor 4E-BP2, but Not S6 Kinase 1/2, to Regulate Neural Stem Cell Self-Renewal In Vivo. Cell Rep. 2013, 5, 433–444. [Google Scholar] [CrossRef]
- Holmberg, J.C.; Riley, V.A.; Sokolov, A.M.; Mukherjee, S.; Feliciano, D.M. Protocol for electroporating and isolating murine (sub)ventricular zone cells for single-nuclei omics. STAR Protoc. 2024, 5, 103095. [Google Scholar] [CrossRef]
- Feliciano, D.M.; Su, T.; Lopez, J.; Platel, J.-C.; Bordey, A. Single-cell Tsc1 knockout during corticogenesis generates tuber-like lesions and reduces seizure threshold in mice. J. Clin. Investig. 2011, 121, 1596–1607. [Google Scholar] [CrossRef]
- Platel, J.-C.; Dave, K.A.; Gordon, V.; Lacar, B.; Rubio, M.E.; Bordey, A. NMDA Receptors Activated by Subventricular Zone Astrocytic Glutamate Are Critical for Neuroblast Survival Prior to Entering a Synaptic Network. Neuron 2010, 65, 859–872. [Google Scholar] [CrossRef]
- Siddiqi, F.; Chen, F.; Aron, A.W.; Fiondella, C.G.; Patel, K.; LoTurco, J.J. Fate Mapping by PiggyBac Transposase Reveals That Neocortical GLAST+ Progenitors Generate More Astrocytes Than Nestin+ Progenitors in Rat Neocortex. Cereb. Cortex 2014, 24, 508–520. [Google Scholar] [CrossRef]
- Chen, F.; Rosiene, J.; Che, A.; Becker, A.; LoTurco, J. Tracking and transforming neocortical progenitors by CRISPR/Cas9 gene targeting and PiggyBac transposase lineage labeling. Development 2015, 142, 3601–3611. [Google Scholar] [CrossRef]
- Riley, V.A.; Holmberg, J.C.; Sokolov, A.M.; Feliciano, D.M. Tsc2 shapes olfactory bulb granule cell molecular and morphological characteristics. Front. Mol. Neurosci. 2022, 15, 970357. [Google Scholar] [CrossRef]
- Hattori, Y.; Miyata, T. Embryonic Neocortical Microglia Express Toll-Like Receptor 9 and Respond to Plasmid DNA Injected into the Ventricle: Technical Considerations Regarding Microglial Distribution in Electroporated Brain Walls. eneuro 2018, 5. [Google Scholar] [CrossRef]
- Lagace, D.C.; Whitman, M.C.; Noonan, M.A.; Ables, J.L.; DeCarolis, N.A.; Arguello, A.A.; Donovan, M.H.; Fischer, S.J.; Farnbauch, L.A.; Beech, R.D.; et al. Dynamic Contribution of Nestin-Expressing Stem Cells to Adult Neurogenesis. J. Neurosci. 2007, 27, 12623–12629. [Google Scholar] [CrossRef]
- Levison, S.W.; Goldman, J.E. Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain. Neuron 1993, 10, 201–212. [Google Scholar] [CrossRef]
- Vézina, C. , Kudelski, A. & Sehgal, S. N. Rapamycin (AY-22, 989) a new antifungal antibiotic. J. Antibiot. (Tokyo) 1975, 28, 721–726. [Google Scholar]
- Sabatini, D.M.; Erdjument-Bromage, H.; Lui, M.; Tempst, P.; Snyder, S.H. RAFT1: A mammalian protein that binds to FKBP12 in a rapamycin-dependent fashion and is homologous to yeast TORs. Cell 1994, 78, 35–43. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Burnett, P.E.; Barrow, R.K.; Cohen, N.A.; Snyder, S.H.; Sabatini, D.M. RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc. Natl. Acad. Sci. USA 1998, 95, 1432–1437. [Google Scholar] [CrossRef]
- Chung, J.; Kuo, C.J.; Crabtree, G.R.; Blenis, J. Rapamycin-FKBP specifically blocks growth-dependent activation of and signaling by the 70 kd S6 protein kinases. Cell 1992, 69, 1227–1236. [Google Scholar] [CrossRef]
- Brown, E.J.; Albers, M.W.; Shin, T.B.; Ichikawa, K.; Keith, C.T.; Lane, W.S.; Schreiber, S.L. A mammalian protein targeted by G1-arresting rapamycin–receptor complex. Nature 1994, 369, 756–758. [Google Scholar] [CrossRef]
- Sabers, C.J.; Martin, M.M.; Brunn, G.J.; Williams, J.M.; Dumont, F.J.; Wiederrecht, G.; Abraham, R.T. Isolation of a Protein Target of the FKBP12-Rapamycin Complex in Mammalian Cells. J. Biol. Chem. 1995, 270, 815–822. [Google Scholar] [CrossRef]
- Lamming, D.W.; Ye, L.; Katajisto, P.; Goncalves, M.D.; Saitoh, M.; Stevens, D.M.; Davis, J.G.; Salmon, A.B.; Richardson, A.; Ahima, R.S.; et al. Rapamycin-Induced Insulin Resistance Is Mediated by mTORC2 Loss and Uncoupled from Longevity. Science 2012, 335, 1638–1643. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Ali, S.M.; Sengupta, S.; Sheen, J.-H.; Hsu, P.P.; Bagley, A.F.; Markhard, A.L.; Sabatini, D.M. Prolonged Rapamycin Treatment Inhibits mTORC2 Assembly and Akt/PKB. Mol. Cell 2006, 22, 159–168. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Yip, C.K.; Murata, K.; Walz, T.; Sabatini, D.M.; Kang, S.A. Structure of the Human mTOR Complex I and Its Implications for Rapamycin Inhibition. Mol. Cell 2010, 38, 768–774. [Google Scholar] [CrossRef]
- Aylett, C.H.S.; Sauer, E.; Imseng, S.; Boehringer, D.; Hall, M.N.; Ban, N.; Maier, T. Architecture of human mTOR complex 1. Science 2016, 351, 48–52. [Google Scholar] [CrossRef]
- Hara, K.; Maruki, Y.; Long, X.; Yoshino, K.-I.; Oshiro, N.; Hidayat, S.; Tokunaga, C.; Avruch, J.; Yonezawa, K. Raptor, a Binding Partner of Target of Rapamycin (TOR), Mediates TOR Action. Cell 2002, 110, 177–189. [Google Scholar] [CrossRef]
- Dos, D.S.; et al. Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar]
- Gingras, A.-C.; Gygi, S.P.; Raught, B.; Polakiewicz, R.D.; Abraham, R.T.; Hoekstra, M.F.; Aebersold, R.; Sonenberg, N. Regulation of 4E-BP1 phosphorylation: a novel two-step mechanism. Genes Dev. 1999, 13, 1422–1437. [Google Scholar] [CrossRef]
- Burnett, P.E.; Barrow, R.K.; Cohen, N.A.; Snyder, S.H.; Sabatini, D.M. RAFT1 phosphorylation of the translational regulators p70 S6 kinase and 4E-BP1. Proc. Natl. Acad. Sci. 1998, 95, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Price, D.J.; Grove, J.R.; Calvo, V.; Avruch, J.; Bierer, B.E.; Grove, J.R. Rapamycin-Induced Inhibition of the 70-Kilodalton S6 Protein Kinase. Science 1992, 257, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Li, Y.; Xu, T.; Guan, K.-L. Rheb GTPase is a direct target of TSC2 GAP activity and regulates mTOR signaling. Genes Dev. 2003, 17, 1829–1834. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, X.; Saucedo, L.J.; Ru, B.; Edgar, B.A.; Pan, D. Rheb is a direct target of the tuberous sclerosis tumour suppressor proteins. Nat. Cell Biol. 2003, 5, 578–581. [Google Scholar] [CrossRef]
- Tee, A.R.; Manning, B.D.; Roux, P.P.; Cantley, L.C.; Blenis, J. Tuberous Sclerosis Complex Gene Products, Tuberin and Hamartin, Control mTOR Signaling by Acting as a GTPase-Activating Protein Complex toward Rheb. Curr. Biol. 2003, 13, 1259–1268. [Google Scholar] [CrossRef]
- Garami, A.; Zwartkruis, F.J.; Nobukuni, T.; Joaquin, M.; Roccio, M.; Stocker, H.; Kozma, S.C.; Hafen, E.; Bos, J.L.; Thomas, G. Insulin Activation of Rheb, a Mediator of mTOR/S6K/4E-BP Signaling, Is Inhibited by TSC1 and 2. Mol. Cell 2003, 11, 1457–1466. [Google Scholar] [CrossRef]
- Dibble, C.C.; Elis, W.; Menon, S.; Qin, W.; Klekota, J.; Asara, J.M.; Finan, P.M.; Kwiatkowski, D.J.; Murphy, L.O.; Manning, B.D. TBC1D7 Is a Third Subunit of the TSC1-TSC2 Complex Upstream of mTORC1. Mol. Cell 2012, 47, 535–546. [Google Scholar] [CrossRef]
- Hasbani, D. M.; Crino, P. B. Tuberous sclerosis complex. Handbook of Clinical Neurology 2018, 148, 813–822. [Google Scholar]
- Chan, D.L.; Calder, T.; Lawson, J.A.; Mowat, D.; Kennedy, S.E. The natural history of subependymal giant cell astrocytomas in tuberous sclerosis complex: a review. Prog. Neurobiol. 2017, 29, 295–301. [Google Scholar] [CrossRef]
- Feliciano, D.M. The Neurodevelopmental Pathogenesis of Tuberous Sclerosis Complex (TSC). Front. Neuroanat. 2020, 14, 39. [Google Scholar] [CrossRef]
- Van Slegtenhorst, M.; et al. Identification of the tuberous sclerosis gene TSC1 on chromosome 9q34. Science 277, 805–808 (1997).
- The European Chromosome 16 Tuberous Sclerosis Consortium. Identification and characterization of the tuberous sclerosis gene on chromosome 16. Cell 75, 1305–1315 (1993).
- Henske, E. P. , Jóźwiak, S., Kingswood, J. C., Sampson, J. R. & Thiele, E. A. Tuberous sclerosis complex. Nat. Rev. Dis. Prim. 2, 16035 (2016).
- Feliciano, D.M.; Lin, T.V.; Hartman, N.W.; Bartley, C.M.; Kubera, C.; Hsieh, L.; Lafourcade, C.; O'Keefe, R.A.; Bordey, A. A circuitry and biochemical basis for tuberous sclerosis symptoms: from epilepsy to neurocognitive deficits. Int. J. Dev. Neurosci. 2013, 31, 667–678. [Google Scholar] [CrossRef]
- Zywitza, V.; Misios, A.; Bunatyan, L.; Willnow, T.E.; Rajewsky, N. Single-Cell Transcriptomics Characterizes Cell Types in the Subventricular Zone and Uncovers Molecular Defects Impairing Adult Neurogenesis. Cell Rep. 2018, 25, 2457–2469.e8. [Google Scholar] [CrossRef]
- Basak, O.; Krieger, T.G.; Muraro, M.J.; Wiebrands, K.; Stange, D.E.; Frias-Aldeguer, J.; Rivron, N.C.; van de Wetering, M.; van Es, J.H.; van Oudenaarden, A.; et al. Troy+ brain stem cells cycle through quiescence and regulate their number by sensing niche occupancy. Proc. Natl. Acad. Sci. USA 2018, 115, 201715911–E619. [Google Scholar] [CrossRef]
- Cebrian-Silla, A.; Nascimento, M.A.; A Redmond, S.; Mansky, B.; Wu, D.; Obernier, K.; Rodriguez, R.R.; Gonzalez-Granero, S.; García-Verdugo, J.M.; A Lim, D.; et al. Single-cell analysis of the ventricular-subventricular zone reveals signatures of dorsal and ventral adult neurogenesis. eLife 2021, 10. [Google Scholar] [CrossRef]
- Blair, J.D.; Hockemeyer, D.; Doudna, J.A.; Bateup, H.S.; Floor, S.N. Widespread Translational Remodeling during Human Neuronal Differentiation. Cell Rep. 2017, 21, 2005–2016. [Google Scholar] [CrossRef]
- Baser, A.; Skabkin, M.; Kleber, S.; Dang, Y.; Balta, G.S.G.; Kalamakis, G.; Göpferich, M.; Ibañez, D.C.; Schefzik, R.; Lopez, A.S.; et al. Onset of differentiation is post-transcriptionally controlled in adult neural stem cells. Nature 2019, 566, 100–104. [Google Scholar] [CrossRef]
- Baser, A.; Skabkin, M.; Kleber, S.; Dang, Y.; Balta, G.S.G.; Kalamakis, G.; Göpferich, M.; Ibañez, D.C.; Schefzik, R.; Lopez, A.S.; et al. Onset of differentiation is post-transcriptionally controlled in adult neural stem cells. Nature 2019, 566, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Paliouras, G.N.; Hamilton, L.K.; Aumont, A.; Joppé, S.E.; Barnabé-Heider, F.; Fernandes, K.J.L. Mammalian Target of Rapamycin Signaling Is a Key Regulator of the Transit-Amplifying Progenitor Pool in the Adult and Aging Forebrain. J. Neurosci. 2012, 32, 15012–15026. [Google Scholar] [CrossRef]
- Kang, S.A.; Pacold, M.E.; Cervantes, C.L.; Lim, D.; Lou, H.J.; Ottina, K.; Gray, N.S.; Turk, B.E.; Yaffe, M.B.; Sabatini, D.M. mTORC1 Phosphorylation Sites Encode Their Sensitivity to Starvation and Rapamycin. Science 2013, 341, 364–373. [Google Scholar] [CrossRef]
- Cochard, L.M.; Levros, L.-C.; Joppé, S.E.; Pratesi, F.; Aumont, A.; Fernandes, K.J.L. Manipulation of EGFR-Induced Signaling for the Recruitment of Quiescent Neural Stem Cells in the Adult Mouse Forebrain. Front. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Holland, E.C.; Hively, W.P.; DePinho, R.A.; Varmus, H.E. A constitutively active epidermal growth factor receptor cooperates with disruption of G1 cell-cycle arrest pathways to induce glioma-like lesions in mice. Genes Dev. 1998, 12, 3675–3685. [Google Scholar] [CrossRef]
- Romero-Pozuelo, J.; Figlia, G.; Kaya, O.; Martin-Villalba, A.; Teleman, A.A. Cdk4 and Cdk6 Couple the Cell-Cycle Machinery to Cell Growth via mTORC1. Cell Rep. 2020, 31, 107504. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Shrikhande, G.; Xu, J.; McKay, R.M.; Burns, D.K.; Johnson, J.E.; Parada, L.F. Tsc1 mutant neural stem/progenitor cells exhibit migration deficits and give rise to subependymal lesions in the lateral ventricle. Genes Dev. 2011, 25, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, C.; Feliciano, D.M.; Bordey, A.; Hartman, N.W. Switching on mTORC1 induces neurogenesis but not proliferation in neural stem cells of young mice. Neurosci. Lett. 2016, 614, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, D.M.; Quon, J.L.; Su, T.; Taylor, M.M.; Bordey, A. Postnatal neurogenesis generates heterotopias, olfactory micronodules and cortical infiltration following single-cell Tsc1 deletion. Hum. Mol. Genet. 2012, 21, 799–810. [Google Scholar] [CrossRef]
- Rushing, G.V.; A Brockman, A.; Bollig, M.K.; Leelatian, N.; Mobley, B.C.; Irish, J.M.; Ess, K.C.; Fu, C.; A Ihrie, R. Location-dependent maintenance of intrinsic susceptibility to mTORC1-driven tumorigenesis. Life Sci. Alliance 2019, 2, e201800218. [Google Scholar] [CrossRef]
- Gorski, J.A.; Talley, T.; Qiu, M.; Puelles, L.; Rubenstein, J.L.R.; Jones, K.R. Cortical Excitatory Neurons and Glia, But Not GABAergic Neurons, Are Produced in the Emx1-Expressing Lineage. J. Neurosci. 2002, 22, 6309–6314. [Google Scholar] [CrossRef]
- Kohwi, M.; et al. A subpopulation of olfactory bulb GABAergic interneurons is derived from Emx1- and Dlx5/6-expressing progenitors. J. Neurosci. 27, (2007).
- Magri, L.; Cambiaghi, M.; Cominelli, M.; Alfaro-Cervello, C.; Cursi, M.; Pala, M.; Bulfone, A.; Garcìa-Verdugo, J.M.; Leocani, L.; Minicucci, F.; et al. Sustained Activation of mTOR Pathway in Embryonic Neural Stem Cells Leads to Development of Tuberous Sclerosis Complex-Associated Lesions. Cell Stem Cell 2011, 9, 447–462. [Google Scholar] [CrossRef]
- Riley, V.A.; Shankar, V.; Holmberg, J.C.; Sokolov, A.M.; Neckles, V.N.; Williams, K.; Lyman, R.; Mackay, T.F.; Feliciano, D.M. Tsc2 coordinates neuroprogenitor differentiation. iScience 2023, 26, 108442. [Google Scholar] [CrossRef]
- Crino, P.B.; Trojanowski, J.Q.; Dichter, M.A.; Eberwine, J. Embryonic neuronal markers in tuberous sclerosis: Single-cell molecular pathology. Proc. Natl. Acad. Sci. 1996, 93, 14152–14157. [Google Scholar] [CrossRef]
- Zordan, P.; Cominelli, M.; Cascino, F.; Tratta, E.; Poliani, P.L.; Galli, R. Tuberous sclerosis complex–associated CNS abnormalities depend on hyperactivation of mTORC1 and Akt. J. Clin. Investig. 2018, 128, 1688–1706. [Google Scholar] [CrossRef]
- Lafourcade, C.A.; Lin, T.V.; Feliciano, D.M.; Zhang, L.; Hsieh, L.S.; Bordey, A. Rheb Activation in Subventricular Zone Progenitors Leads to Heterotopia, Ectopic Neuronal Differentiation, and Rapamycin-Sensitive Olfactory Micronodules and Dendrite Hypertrophy of Newborn Neurons. J. Neurosci. 2013, 33, 2419–2431. [Google Scholar] [CrossRef] [PubMed]
- Magini, A.; Polchi, A.; Di Meo, D.; Mariucci, G.; Sagini, K.; De Marco, F.; Cassano, T.; Giovagnoli, S.; Dolcetta, D.; Emiliani, C. TFEB activation restores migration ability to Tsc1-deficient adult neural stem/progenitor cells. Hum. Mol. Genet. 2017, 26, 3303–3312. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.M.; Holmberg, J.C.; Feliciano, D.M. The amino acid transporter Slc7a5 regulates the mTOR pathway and is required for granule cell development. Hum. Mol. Genet. 2020, 29, 3003–3013. [Google Scholar] [CrossRef] [PubMed]
- Skalecka, A.; Liszewska, E.; Bilinski, R.; Gkogkas, C.; Khoutorsky, A.; Malik, A.R.; Sonenberg, N.; Jaworski, J. mTOR kinase is needed for the development and stabilization of dendritic arbors in newly born olfactory bulb neurons. Dev. Neurobiol. 2016, 76, 1308–1327. [Google Scholar] [CrossRef]
- Cox, R.L.; de Anda, F.C.; Mangoubi, T.; Yoshii, A. Multiple Critical Periods for Rapamycin Treatment to Correct Structural Defects in Tsc-1-Suppressed Brain. Front. Mol. Neurosci. 2018, 11, 409. [Google Scholar] [CrossRef]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional Transport of Amino Acids Regulates mTOR and Autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef]
- Sokolov, A.M.; Holmberg, J.C.; Feliciano, D.M. The amino acid transporter Slc7a5 regulates the mTOR pathway and is required for granule cell development. Hum. Mol. Genet. 2020, 29, 3003–3013. [Google Scholar] [CrossRef]
- Lafourcade, C.A.; Lin, T.V.; Feliciano, D.M.; Zhang, L.; Hsieh, L.S.; Bordey, A. Rheb Activation in Subventricular Zone Progenitors Leads to Heterotopia, Ectopic Neuronal Differentiation, and Rapamycin-Sensitive Olfactory Micronodules and Dendrite Hypertrophy of Newborn Neurons. J. Neurosci. 2013, 33, 2419–2431. [Google Scholar] [CrossRef]
Figure 1.
Neonatal Neurogenesis. Numerous cell types are generated from the postnatal?? V-SVZ. NSCs may be quiescent or induced to divide and generate astrocytes, ependyma, NG2, or oligodendrocyte lineage cells. In addition, V-SVZ NSCs generate rapidly dividing transit amplifying cells (TACs), which produce neuroblasts that mature into neurons in the olfactory bulb, namely granule cells.
Figure 1.
Neonatal Neurogenesis. Numerous cell types are generated from the postnatal?? V-SVZ. NSCs may be quiescent or induced to divide and generate astrocytes, ependyma, NG2, or oligodendrocyte lineage cells. In addition, V-SVZ NSCs generate rapidly dividing transit amplifying cells (TACs), which produce neuroblasts that mature into neurons in the olfactory bulb, namely granule cells.

Figure 2.
Origins of Postnatal OB Neurogenesis Diversity.A. Schematic of an embryonic mouse with a blue rectangle indicating where the coronal section of B is taken. B. Coronal section of mouse embryonic having V-SVZ regions labeled including the lateral and medial ganglionic eminences (LGE, MGE). The color corresponds to the cell types and regions that are generated in C and D. C. OB neurons originating from specific regions in the postnatal V-SVZ are color coordinated and originate from those regions indicated in B and D. D. Coronal hemi-section of an adult brain with V-SVZ regions highlighted that generate the neurons in C.
Figure 2.
Origins of Postnatal OB Neurogenesis Diversity.A. Schematic of an embryonic mouse with a blue rectangle indicating where the coronal section of B is taken. B. Coronal section of mouse embryonic having V-SVZ regions labeled including the lateral and medial ganglionic eminences (LGE, MGE). The color corresponds to the cell types and regions that are generated in C and D. C. OB neurons originating from specific regions in the postnatal V-SVZ are color coordinated and originate from those regions indicated in B and D. D. Coronal hemi-section of an adult brain with V-SVZ regions highlighted that generate the neurons in C.
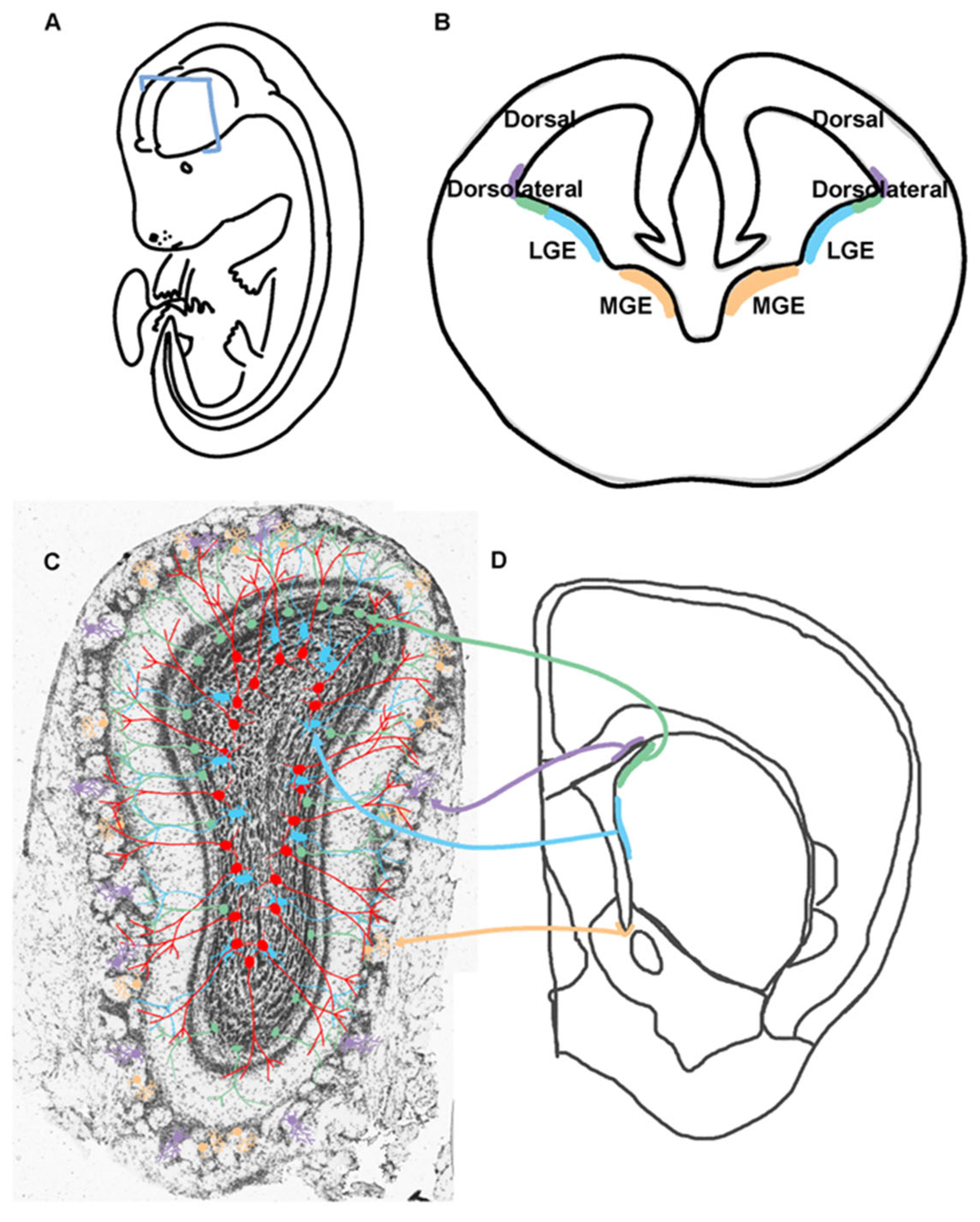
Figure 3.
Olfactory Bulb Circuitry.Olfactory epithelial sensory neurons project axons onto the olfactory bulb. Thes axons form spheres called glomeruli that are surrounded by juxta/periglomerular cells. Stimulation of the sensory neurons can activate mitral cells that contain dendrites. Granule cells in the granule cell layer form dendrodendritic synapses with the mitral cells and release GABA to regulate mitral cell excitation.
Figure 3.
Olfactory Bulb Circuitry.Olfactory epithelial sensory neurons project axons onto the olfactory bulb. Thes axons form spheres called glomeruli that are surrounded by juxta/periglomerular cells. Stimulation of the sensory neurons can activate mitral cells that contain dendrites. Granule cells in the granule cell layer form dendrodendritic synapses with the mitral cells and release GABA to regulate mitral cell excitation.
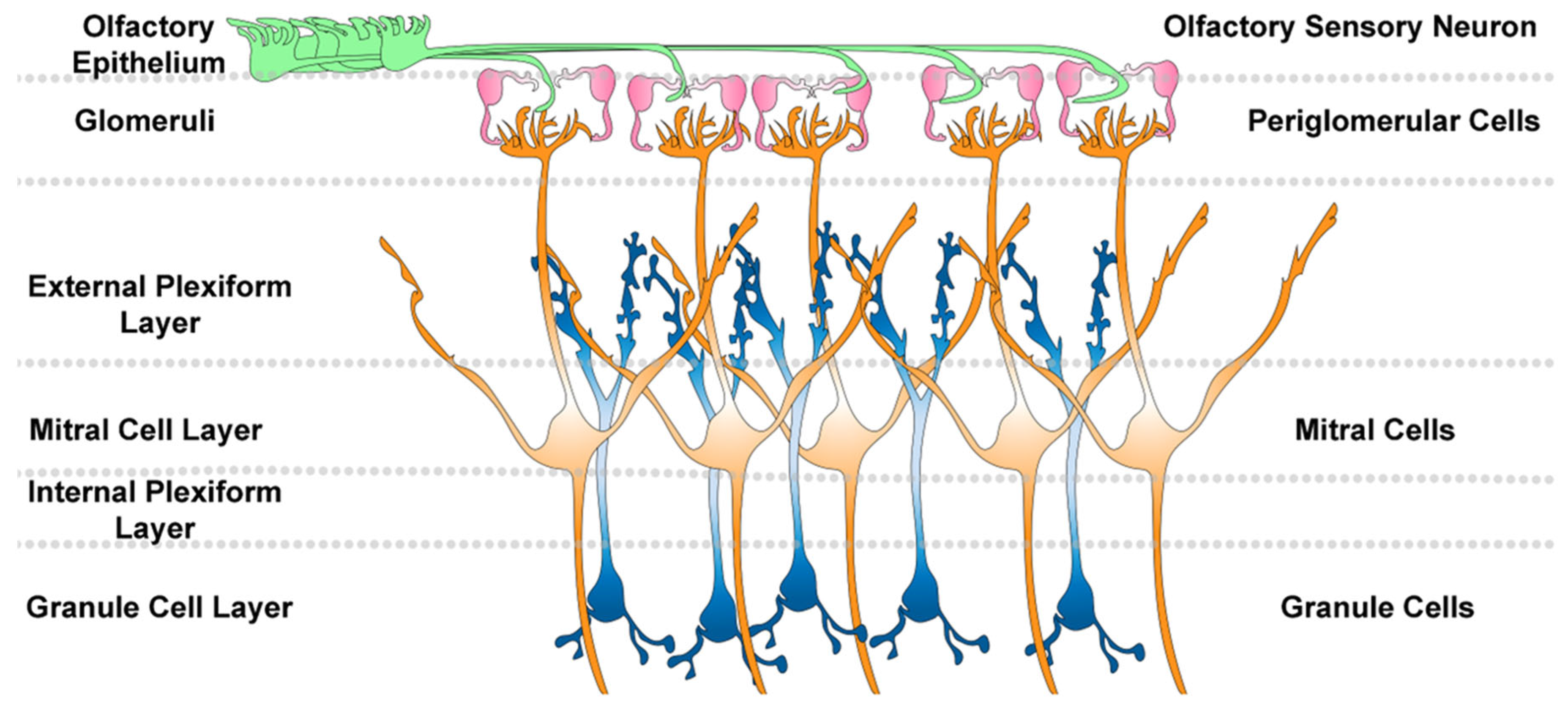
Figure 4.
Techniques for Studying V-SVZ NSCs and Cell Progeny.A, B. Images of a postnatal day 30 olfactory bulb and corresponding V-SVZ cell progeny including GCs and PGs. C. P6 V-SVZ dorsal region following electroporation of RFP plasmid at P0. D. OB demonstrating that neuroblasts from the mouse in C are still migrating to their final destination and do not have the extensive arbors demonstrated in A, B. E. P10 V-SVZ of a nestin-CRE-ERT2 mouse crossed to an inducible RFP mouse demonstrating robust labeling of the V-SVZ. F. P10 OB demonstrating that the vast majority of cells are still migrating into the OB. Note the large number of cells in the core which are DCX positive (green) and Neu-N negative (blue).
Figure 4.
Techniques for Studying V-SVZ NSCs and Cell Progeny.A, B. Images of a postnatal day 30 olfactory bulb and corresponding V-SVZ cell progeny including GCs and PGs. C. P6 V-SVZ dorsal region following electroporation of RFP plasmid at P0. D. OB demonstrating that neuroblasts from the mouse in C are still migrating to their final destination and do not have the extensive arbors demonstrated in A, B. E. P10 V-SVZ of a nestin-CRE-ERT2 mouse crossed to an inducible RFP mouse demonstrating robust labeling of the V-SVZ. F. P10 OB demonstrating that the vast majority of cells are still migrating into the OB. Note the large number of cells in the core which are DCX positive (green) and Neu-N negative (blue).
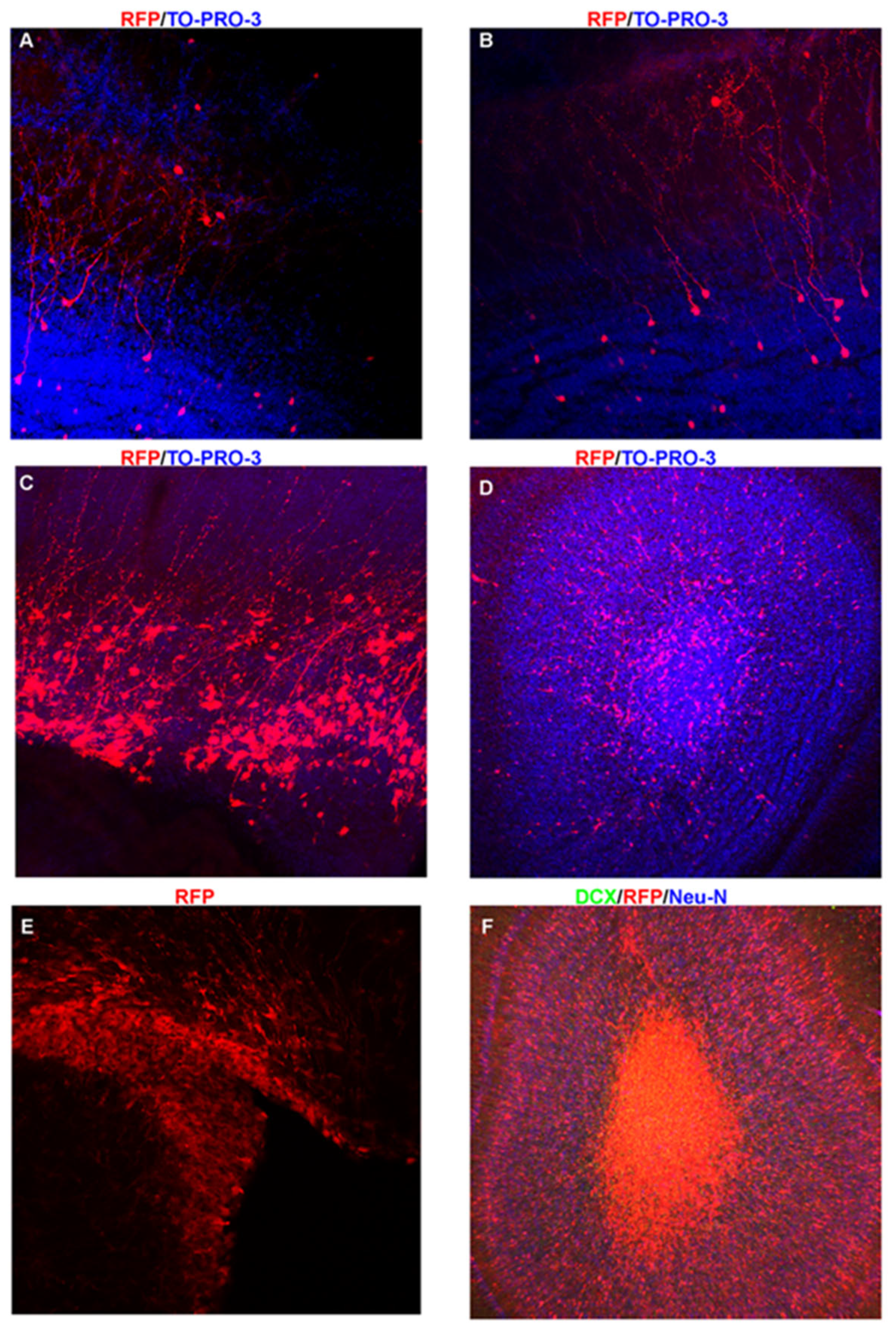
Figure 5.
TSC-mTORC1 Pathway.Ligand (L) activation of receptors (R) leads to inhibition of tuberin bound to hamartin which are encoded by Tsc2 and Tsc1. Normally, tuberin/hamartin inhibit the GTPase Rheb from activating mTORC1. mTORC1 contains mTOR, which is a protein kinase that phosphorylates proteins to stimulate translation of mRNA containing a 5’TOP motif.
Figure 5.
TSC-mTORC1 Pathway.Ligand (L) activation of receptors (R) leads to inhibition of tuberin bound to hamartin which are encoded by Tsc2 and Tsc1. Normally, tuberin/hamartin inhibit the GTPase Rheb from activating mTORC1. mTORC1 contains mTOR, which is a protein kinase that phosphorylates proteins to stimulate translation of mRNA containing a 5’TOP motif.
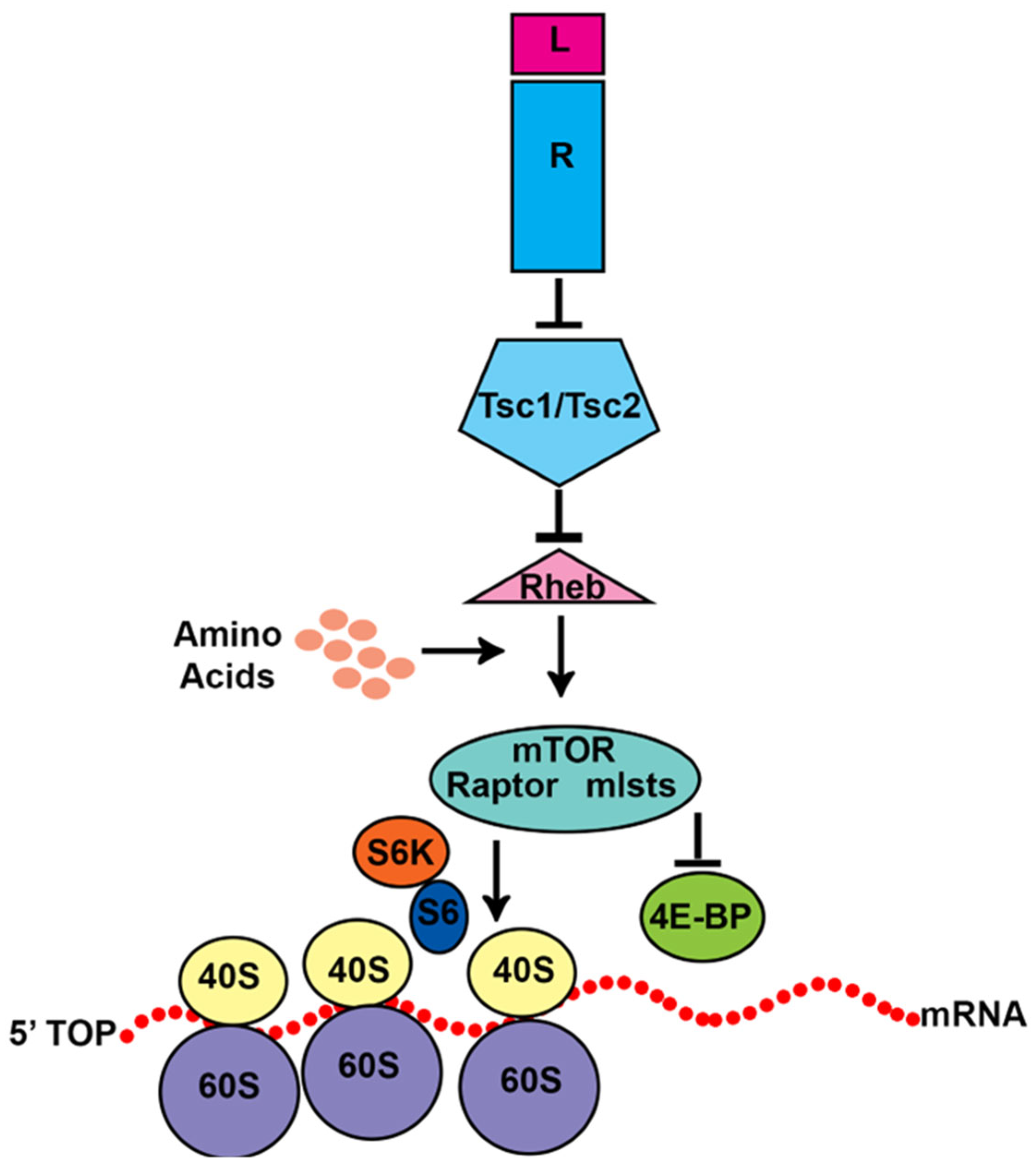
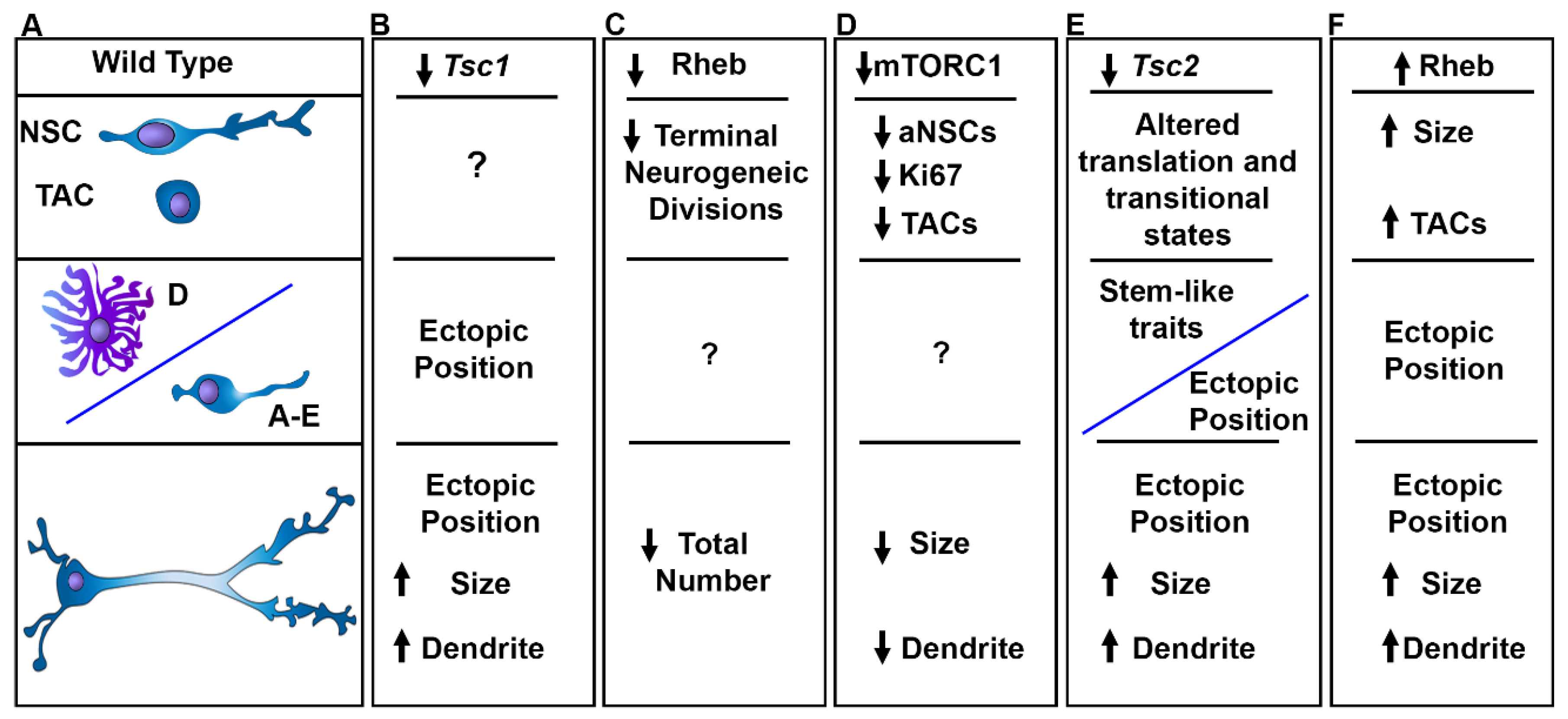
Figure 6.
Alterations in mTORC1 Signaling Affect V-SVZ Neurogenesis. A. Wild type NSCs go from quiescence to an activated state and produce astrocytes and/or transit amplifying cells that generate neuroblasts which mature into granule cells and periglomerular cells. B. Loss of Tsc1 leads to ectopic heterotopias and subependymal nodules as well as fewer GCs in the OB, cytomegaly, and dendrite arborization. C. Reducing Rheb decreases the production of neuroblasts and decreases the total number of neurons produced. D. Decreasing mTORC1 activity by rapamycin reduces the number of dividing NSCs and fewer TACs. Moreover rapamycin and raptor knockdown reduce soma size and dendrite arbors. E. Loss of Tsc2 causes altered transitional states identified by single nuclei RNA sequencing and prevents proper differentiation leading to the production of partially differentiated cells expressing stem cell proteins and causes the formation of V-SVZ hamartomas and SEGA-like lesions. Moreover, OB neurons were cytomegalic and had increased dendrite growth. Ectopic expression of mutant active Rheb increased the size of NSCs and increased the total number of TACs associated with the formation of nodules and ectopic neurons that were cytomegalic and had hypertrophic dendrites.
Figure 6.
Alterations in mTORC1 Signaling Affect V-SVZ Neurogenesis. A. Wild type NSCs go from quiescence to an activated state and produce astrocytes and/or transit amplifying cells that generate neuroblasts which mature into granule cells and periglomerular cells. B. Loss of Tsc1 leads to ectopic heterotopias and subependymal nodules as well as fewer GCs in the OB, cytomegaly, and dendrite arborization. C. Reducing Rheb decreases the production of neuroblasts and decreases the total number of neurons produced. D. Decreasing mTORC1 activity by rapamycin reduces the number of dividing NSCs and fewer TACs. Moreover rapamycin and raptor knockdown reduce soma size and dendrite arbors. E. Loss of Tsc2 causes altered transitional states identified by single nuclei RNA sequencing and prevents proper differentiation leading to the production of partially differentiated cells expressing stem cell proteins and causes the formation of V-SVZ hamartomas and SEGA-like lesions. Moreover, OB neurons were cytomegalic and had increased dendrite growth. Ectopic expression of mutant active Rheb increased the size of NSCs and increased the total number of TACs associated with the formation of nodules and ectopic neurons that were cytomegalic and had hypertrophic dendrites.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



