
Submitted:
11 March 2025
Posted:
12 March 2025
You are already at the latest version
Abstract
Human Papillomavirus (HPV) causes anogenital warts and six types of cancer in infected women, men, or children, being a challenge to global public health. HPV capsid is composed of viral type-specific L1 and evolutionarily conserved L2 proteins. Produced in heterologous systems, L1 protein can self-assemble into virus-like particles (VLPs), nanoparticles sizing around 50-60 nm, used as prophylactic vaccines. Devoid of the viral genome, they are safe to users, offering no risk of infection and VLPs are not self-replicating. The immune response induced by VLPs is promoted by conformational viral epitopes, generating effective T- and B-cell reactions. Produced in different cell systems, VLPs can be obtained in large-scale for mass immunization programs. The expression of heterologous proteins was evaluated at various transfection times by transfecting cells with vectors encoding codon-optimized HPV16L1 and HPV16L2 genes. Immunological response induced by chimeric HPV16 L1/L2 VLP was evaluated through preclinical assays by antibody production, suggesting the potential of broad-spectrum protection against HPV, as prophylactic nanovaccine. Considering VLPs serve as platforms to display peptide antigens from other infectious agents or metabolic diseases, such as cancer, preventive and curative strategies are being explored. Assays with small synthetic peptides designed for delivery by HPV16 L1/L2 VLPs as nanocarriers are in progress, which could potentially be a groundbreaking study in the development of HPV chimeric nanovaccines. This review covers the various possibilities for complementary studies to develop potential prophylactic and therapeutic vaccines with broad-spectrum protection, using innovative methodologies to obtain more effective immunobiological in combating HPV and associated diseases.
Keywords:
L1/L2 VLPs
; high-risk HPV
; cervical cancer
; chimeric nanovaccines
; nanocarriers
HIGHLIGHTS
The expression and evaluation of recombinant chimeric HPV16 L1/L2 VLP proteins in different cell line systems have potential both as nanovaccines and as drug carriers, standing out for their versatility and efficacy inducing immune responses and targeted delivery of therapeutic agents.
- Expression efficiency in distinct cell lineages.
- Immunogenicity and efficacy as chimeric nanovaccine.
- Capacity as drug carriers.
1. Introduction
Virus-like particles (VLPs) are a fascinating area in vaccine and drug delivery research. They are considered safe, have good immunogenic potential, and are versatile. Furthermore, it highlights their applications in combating HPV-related diseases, both as a form of prevention against new infections and as nanocarriers to treat already established tumor cells.
Therapeutic vaccines for the treatment of the lower genital tract infections caused by HPV represent an important challenge to be developed. While the structural protein L1 has the capacity for self-assembly, forming VLPs, the protein L2 can anchor and generate greater stability to the L1 complexes [1]. Another interesting characteristic of L2 is the high conservation among the different types of HPV, which may induce a cross immunity for different HPV strains. Despite the unclear durability of immune responses induced by L2, literature data highlight that this protein may be a promising target for developing therapeutic HPV vaccines [2].
VLPs are impressive tools that can be applied to work with viral structures at the atomic level and have become key tools in studies in several areas, such as biology, medicine, and engineering. Thus, the use of VLPs in the development of antiviral vaccines was a natural path to follow. The success of this application paved the way for the development of other types of vaccines and delivery systems based on VLPs, using them as nanomachines capable of delivering drug products directly to their specific targets. The application capabilities of VLPs are promising, leading to studies and investments with the potential for the development of prophylactic and therapeutic vaccines, as a new generation of immunobiological, with higher efficacy and the possibility of significant financial returns [3,4].
VLPs, primarily derived from human and animal viruses, serve as effective vaccines against their specific viruses. Furthermore, VLPs — including those derived from plant and bacterial viruses — can be designed to present foreign peptide antigens from infectious agents or diseases like cancer, facilitating the creation of chimeric VLPs. The primary objective of chimeric VLPs is to enhance the immunogenicity of the displayed foreign peptides, rather than focusing solely on the VLP platforms themselves. [5].
VLPs are composed of viral structural proteins able to self-assemble into particles, spontaneously, being antigenically indistinguishable from infectious virus particles, however, are non-infectious because lack the genetic material required for replication. VLPs are highly attractive as vaccine candidates due to their inherent properties, such as their virus-like size, multimeric antigen presentation, and highly organized, repetitive structure. These features make them particularly effective in eliciting robust and safe humoral and cellular immune responses [5]. To date, several VLP-based vaccines have been approved for both human and veterinary use, with numerous other candidates currently in advanced stages of clinical evaluation [6]. Furthermore, the versatility of VLPs as platforms for displaying foreign antigens has significantly expanded their potential applications. This adaptability allows VLPs to be utilized not only as prophylactic vaccines but also as therapeutic vaccines, paving the way for the development of next-generation vaccine technologies [7,8].
VLPs are the product of the expression of one or more recombinant viral proteins capable of self-assembling into nanoparticles in order 20 to 200 nm in diameter. Due to their lack of viral genome, they are safe for recipients, since they do not cause infection and do not self-replicate. The immune response induced by VLPs is promoted by conformational viral epitopes, which generate an effective T- and B-cell reaction. They can be produced in different systems through genetic engineering, from established cell lines of vertebrates, insects, plants, yeast, and bacteria, allowing large-scale production for mass populational immunization. For human and veterinarian use, prophylactically and/or therapeutically, VLPs provide high immunogenicity as nanovaccines and efficiency as drug nanocarriers [9,10,11].
This review also intends to provide overviews about some aspects of HPV virus-like particles, considering that they come into existence by self-assembly from viral components in the infected cell, through their extracellular stage, until recognized and infecting a new host cell and cease to exist by losing their physical integrity to start a new infectious cycle [12].
We will also present techniques for studying the structure of virus particles and components and some applications of structure-based virus studies.
2. Overview of basic concepts
2.1. Virus-like Particle Vaccines and Platforms for Vaccine Development
Virus-like particle (VLP) vaccines are an innovative and promising approach to vaccine development. They mimic the structure of viruses but do not contain genetic material, making them safe for humans and veterinary use (Figure 1). This characteristic allows VLPs to induce a robust immune response, like that elicited by real viral infections, without the risk of causing disease [3].
The versatility of VLP platforms allows them to create diverse vaccines that can be used against different viruses, bacteria, and even cancer cells. VLPs can be produced using a variety of host cells, such as bacteria, yeast, insect, and mammalian cells. Examples of cell systems used to express VLPs include Escherichia coli, Pichia pastoris, cell insects, Human Embryonic Kidney cells (HEK293 and variant lineages), among others, allowing for scalability and adaptation to the specific needs of each vaccine (Figure 2). For example, Sakauchi et al. (2021) presented an example of scalability using the suspension culture cell line HEK 293F as an expression system for expressing L1 and L2 proteins, resulting in a higher yield of protein compared to expression in HEK 293T, an adherent cell line [13]. Bei et al. (2024) demonstrated an expression platform utilizing SF9 insect cells, transfected with baculovirus, as an effective system for producing VLPs that make up the recombinant 14-valent HPV vaccine, currently in phase III trials [14]. Other cell lineages are also utilized as expression platforms for vaccine production. For instance, the insect cell line Trichoplusia ni is employed to produce the Cervarix vaccine. Similarly, the yeast species Saccharomyces cerevisiae and Pichia pastoris are used to manufacture the Gardasil and WalrinVax vaccines, respectively. Additionally, the bacterium Escherichia coli serves as the expression system for the production of the Cecolin vaccine [15]. VLPs can also be used as platforms for drug delivery, acting as nanocarriers to carry substances directly to target cells, increasing treatment efficacy and reducing side effects [16].
VLP technology has several advantages that can be applied to the development of vaccines and other therapeutics, as we will discuss in the following sections.
2.2. Uses of VLPs
- Preventive Vaccines: VLPs have been used in vaccines against viruses such as HPV (human papillomavirus) and hepatitis B. They induce an effective immune response, helping to prevent infections.
- Oncology Therapies: In addition to vaccines, VLPs are being explored as nanocarriers for cancer therapies, allowing targeted delivery of drugs directly to tumor cells.
- Personalized Vaccines: VLPs' versatility allows them to be adapted to create personalized vaccines for different pathogens or virus variants, responding quickly to emerging outbreaks.
- Delivery Platforms: VLPs can deliver antigens from different pathogens, facilitating the creation of combination vaccines.
2.3. Advantages of VLPs Compared to Other Vaccine Approaches
- Safety: Because VLPs do not contain viral genetic material, they cannot cause infections, making them a safe option for vaccination.
- Immunogenicity: VLPs are highly immunogenic and can induce a strong and long-lasting immune response, often with fewer doses than other vaccines.
- Versatility: The ability to modify VLPs to include different antigens allows them to be used in a wide range of diseases, from viral infections to cancer.
- Efficient Production: VLPs can be produced in mammalian cells, such as HEK cell lines, which are ideal for large-scale production and can quickly be adapted to various antigens.
- Fewer Side Effects: The non-infectious nature of VLPs makes them less likely to cause side effects than the attenuated virus vaccines.
2.4. Structural basis of viral function
Examining viruses' structural composition and fundamental architecture is essential to comprehensively understanding the mechanisms by which they infect host cells and replicate. The following outlines the primary components and characteristics.
2.4.1. Basic Structure of Viruses
- I.
- Capsid: The capsid is a protein coat that surrounds and protects the virus's genetic material. It is composed of proteins called capsomeres, which are arranged in specific patterns. The shape of the capsid can be icosahedral, helical, or complex, depending on the type of virus. For example, the capsid of HPV is composed of two structural proteins, L1 and L2, which assemble to form an icosahedral structure. The major capsid protein, L1, organizes into 72 pentameric capsomeres, arranged in a T=7 symmetric lattice (Figure 3.A). These capsomeres create the outer shell of the capsid, providing the virus with its characteristic spherical shape. The minor capsid protein, L2, is located internally and plays a role in stabilizing the structure and facilitating viral genome packaging. Together, L1 and L2 form a robust and highly organized capsid that protects the viral DNA and mediates host cell entry.
- II.
- Genetic Material: Viruses contain genetic material, either DNA or RNA, which can be single- or double-stranded. This material allows the virus to hijack host cell machinery, replicate, and produce new viral particles. For example, HPV has a double-stranded DNA genome that encodes proteins for replication and assembly, enabling it to infect and spread within host cells. This genetic diversity is key to viruses' ability to infect a wide range of hosts and cause disease.
- III.
- Viral Envelope: Some viruses have a lipid envelope surrounding the capsid. This envelope is derived from the host cell membrane and contains viral proteins that help the virus adhere to and enter the cells. Enveloped viruses, such as HIV and influenza viruses, are generally more sensitive to disinfectants and environmental conditions.
2.4.2. Viral Function
- I.
- Attachment and Entry: Proteins on the surface of the viral capsid or envelope bind to specific receptors on the host cell surface. For example, in HPV, the major capsid protein L1 interacts with heparan sulfate proteoglycans (HSPGs) on the cell surface. This binding trigger structural changes in the virus, exposing other proteins essential for internalization. In HPV, the minor capsid protein L2 becomes exposed after the L1-HSPG interaction. The exposed L2 protein then interacts with additional cell receptors, such as integrins, facilitating clathrin- or caveolin-mediated endocytosis. This process enables the virus to enter the host cell and initiate infection.
- II.
- Replication: Once inside the cell, the virus's genetic material is released and uses the host cell machinery to replicate itself. This may involve transcription and translation of viral genetic material to produce new viral proteins.
- III.
- Assembly: After replication, new viral particles (viruses) are assembled from the genetic material and proteins produced. Depending on the type of virus, this may occur in the nucleus or cytoplasm of the cell.
- IV.
- Release: Finally, the new viruses are released from the host cell. In enveloped viruses, this usually occurs by budding, where the virus acquires its envelope from the host cell membrane. In non-enveloped viruses, the cell may rupture, releasing viral particles.
In conclusion, the structural basis of viral function and the basic architecture of viruses are fundamental to understanding how these infectious agents operate and replicate. The capsid, composed of proteins, protects the virus's genetic material and is essential for its replication and infection. The presence of an envelope in some viruses adds a layer of complexity, allowing specific interactions with host cells [3].
These structures not only define the shape and stability of viruses but also determine their ability to infect cells and spread. The interaction between viral proteins and cellular receptors is crucial for virus adhesion and entry, while replication and assembly depend on the host cellular machinery [12].
To summarize, viral architecture is a determining factor in the efficacy of infection and the pathogenicity of viruses. Understanding these structures and functions is essential for the development of vaccines and antiviral therapies, allowing the creation of effective strategies to combat viral infections and protect public health [12,20].
3. Overview of Specific Concepts
3.1. Prophylactic and Therapeutic DNA Vaccine against HPV and Virus-Associated Cancers
Cervical cancer is the fourth most prevalent type of cancer among women worldwide, with approximately 600,000 new cases registered in 2020. Data indicates that approximately 90% of the 342,000 deaths recorded from this type of cancer occurred in low-income countries [21]. There is also an association between HPV and other types of cancer, such as head, neck, and oropharyngeal carcinomas [22,23,24], anogenital (vaginal, vulvar, penile, and anal) [25,26], and cases of non-melanoma skin cancers [27,28].
The GLOBOCAN 2022 database provides estimates of new cancer cases and deaths worldwide for all cancers combined (ICD-10 codes C00–C97) and for 36 specific cancer types. These include oropharynx, anus, vulva, vagina, cervix uteri, penis, and nonmelanoma skin cancer (NMSC; ICD-10 code C44, excluding basal-cell carcinomas) [29]. The global cancer burden for 2022, based on the latest GLOBOCAN estimates by the International Agency for Research on Cancer (IARC), is accessible through the Global Cancer Observatory's "Cancer Today" platform [30].
Currently, the main form of prevention of cervical cancer is to perform periodic cytological exams, Pap smear tests, and prophylactic vaccination of pre-adolescent girls and boys. In Brazil, the prophylactic vaccine Gardasil® quadrivalent was made available by the Ministry of Health (Ministério da Saúde do Brasil), free of charge through the Unified Health System (Sistema Único de Saúde – SUS) for boys and girls aged 9 to 19 [31], as well as for men and women aged 9 to 26 living with HIV or AIDS, patients who have received organ transplants, bone marrow transplants and people undergoing cancer treatment [32]. Individuals outside the age range covered by the SUS can receive HPV immunization at private vaccination clinics. Two vaccine options are available: the quadrivalent vaccine, recommended for females aged 9 to 45 and males aged 9 to 26, and the nonavalent vaccine, suitable for everyone aged 9 to 45. Furthermore, the vaccines can be used by people who are undergoing treatment or have already had an HPV infection, as they can protect against other types of the HPV virus and prevent the formation of new genital warts and the risk of cancer [33]. Currently, the 9-valent Gardasil is the most effective vaccine, able to promote immunization against nine HR-HPV strains (HPV-6, 11, 16, 18, 31, 33, 45, 52, and 58). Nevertheless, a recombinant 14-valent HR-HPV vaccine (SCT 1000) is in phase III trials in China; this vaccine offers protection against the same strains as Gardasil-9 and five additional strains (HPV-35, 39, 51, 56, and 59) [14] (Table 1).
There is no specific treatment for HPV infections or lesions that have already developed, making prophylactic vaccination and preventive measures essential for reducing infection risk and detecting HPV-related lesions early. Among the proteins expressed by HPV, L2, E7, and E6 are prominent targets for DNA vaccine development. The L2 protein, part of the viral capsid, is well conserved across different HPV types, making it a promising candidate for cross-protective vaccines. E6, an oncogenic protein, plays a key role in the malignant transformation of infected cells, particularly in conjunction with E7. The oncogenes E6 and E7 are essential for malignancy, as they integrate into the host DNA during cancer progression. E6 promotes the degradation of the tumor suppressor protein p53, while E7 inactivates the retinoblastoma protein (pRb), disrupting cell cycle regulation and enabling uncontrolled cell proliferation [34,35,36].
Despite the availability of prophylactic vaccines, HPV infections remain extremely common worldwide. The present invention, summarized, relates to a prophylactic and therapeutic vaccine against HPV and cancers associated with the virus, intended for individuals infected with HPV who have developed cancer [37,38]. It also relates to DNA expression vectors that can efficiently produce the L2 protein of the HPV16 virus capsid, in addition to the viral oncoprotein E6 associated with tumors of the Human Papillomavirus. This invention focuses on producing recombinant fusion or hybrid proteins through gene cloning in expression vectors. By combining regulatory sequences with protein-coding sequences from one or more genes, fused genes are translated into proteins that trigger a long-lasting humoral immune response by stimulating the production of anti-L2 antibodies to prevent infection (prophylactic action). These proteins also activate the cellular immune response, targeting tumor cells by inducing anti-E6 activity and promoting the expression of tumor necrosis factor (TNF) cytokines, which enhance the body’s ability to combat malignancies (therapeutic action). This approach offers a comprehensive solution that integrates prevention and treatment [37,38].
DNA vaccines offer a promising alternative to traditional platforms due to their simplicity, low production costs, thermal stability (eliminating the need for refrigeration), and ability to be administered in repeated doses without eliciting an immune response against the vector. They have the potential to induce both humoral and cellular immunity and are highly tolerable in humans, making them suitable for various applications. However, their full clinical potential is hindered by challenges such as limited delivery efficiency, as only a small fraction of the DNA is internalized and expressed by target cells, and weaker immune responses compared to conventional vaccines, often requiring adjuvants or enhanced delivery methods. Additionally, theoretical safety concerns, including the possibility of genomic integration, though rare, cannot be entirely disregarded. Overcoming these obstacles is essential for DNA vaccines to achieve their transformative potential in modern medicine [39].
3.2. Peptide-Based Vaccine: Current Landscape in the Therapeutic HPV-Related Cancers
Peptides are part of a group of subunit vaccines, i.e., vaccines based on antigenic subunits or epitopes. Due to their ability to precisely alter immune response, small size, and low to medium production costs using simple and well-known methods, these molecules are attracting attention [40]. Peptide epitopes have several advantages in their application as antigens for developing prophylactic and therapeutic vaccines. They can be synthesized either chemically or biologically using several approaches, including solid-phase peptide synthesis (SPPS) [41,42], cell-free systems [43,44], and recombinant production in bacteria, yeast, or mammalian cells [13,44].
The VLP structural proteins may contain several epitopes that are searchable by molecular modeling and in silico assays, identifying high-level immune stimulation peptide sequences [45]. Zhai et al. (2017) observed that peptides from HPV L2-protein (amino acid sequences 17-36, 69-86, and 108-122) could be coated on MS2 bacteriophage VLP to provide complete and cross-protection against HR-HPV types, protecting mice from cervicovaginal infections with HPV pseudoviruses 16, 18, 31, 33, 45, and 58 at levels comparable to those immunized with Gardasil-9 [46]. In turn, Namvar et al. (2019) observed highly conserved peptide sequences between HR-HPV subtypes in the L1 (5 regions) and L2 (4 regions) proteins, which showed a high affinity for MHC-I and MHC-II receptors, suggesting the ability to elicit cellular and humoral immune responses [47].
Research into peptide-based vaccines against HPV goes beyond the structural proteins that make up VLPs, seeking epitopes in the oncoproteins E6 and E7, involved in HPV carcinogenesis by interference in the cell cycle control. E6 interacts with E6AP forming a complex responsible for p53 degradation, causing loss of the cell cycle checkpoint and avoiding apoptosis [48]. E7 causes the release of E2F transcription factors and degradation of pRb, forcing the cells through premature S-phase entry, by transcription of cyclins and tumor suppressor protein p16INK4A [45,49]. Using E6 and E7 epitopes was an approach with promising potential for developing therapeutic vaccines, as a promise for treating pre-existing infections and already established tumors, besides generating protection against new infections [40].
Currently, several vaccines with peptide epitopes of E6 and E7 have been in clinical trials of diverse phases, such as the polypeptide vaccine PepCan-based on four E6 epitope sequences with a new adjuvant of yeast extract (Candin®), which promotes Th1 increasing and anti-E6 responses [50]. In phase II studies, a synthetic long peptide derived from the HPV16 E6 and E7 tumor epitopes (HPV16-SLP) showed a specific T-cell response, establishing a long-lasting immunological memory in individuals with low-grade cervical lesions [51], besides inducing broad IFN-γ-related T-cell responses with well-tolerated in individuals with advanced or recurring HPV16-induced gynecological cancer [52]. In addition to increasing immunogenicity and protecting peptides from degradation, adjuvants can improve molecular delivery and enhance pharmacokinetics and pharmacodynamic benefits [40].
3.3. In Silico Peptides Evaluation Directed to HPV
Computational techniques, including CADD (Computer-Aided Design and Drafting) and in silico assays are essential tools in HPV vaccine development. The main advantage of CADD is that it allows modeled ideas to be explored before physical prototyping is implemented. CADD streamlines the design of multiepitope constructs by predicting interactions between HPV antigens (e.g., L1, L2, E6, and E7) and immune receptors, while in silico assays simulate immunogenicity, toxicity, and structural stability. These methods reduce costs and accelerate candidate selection. However, their reliance on simplified models and incomplete databases requires experimental validation to confirm predictions [53,54].
Diverse treatments have already been tested to combat HPV-associated diseases, including the use of therapeutic vaccines developed on different platforms, such as attenuated vectors (viral or bacterial), peptide/protein vaccines and even cell-based vaccines [47,55,56,57]. Several HPV vaccines with therapeutic potential are in advanced stages of clinical trials. These vaccines utilize various epitopes derived from the oncoproteins E6 and E7, as well as the structural proteins L1 and L2, and are developed on diverse vaccine platforms [55]. Notable examples include HPV16 L1E7 chimeric virus-like particles (CVLPs) [58], the ZYC-101a DNA vaccine (encoding HPV16/18 E6 and E7) [59], the TA-CIN protein vaccine (an E6-E7-L2 tandem fusion protein) [60], TA-HPV (a recombinant vaccinia virus expressing E6 and E7) [61], and the SGN-00101 protein vaccine (a fusion of heat shock protein with HPV16 E7) [62]. While each approach offers distinct advantages, they also present certain limitations. Consequently, identifying potent and safe strategies to enhance the immunogenicity of therapeutic vaccines remains a critical priority. In this context, the use of computational tools, such as in silico studies, is essential for advancing vaccine development [57,63,64].
At this point, immunoinformatic tools can assist scientists in predicting high immunogenic and conserved epitopes, which induce B- or T-cell responses against HPV infection, and the linkage of antigens to HSPs provides a promising strategy to increase the efficiency of vaccine candidates [57].
In silico approaches were employed to design the multiepitope L1-L2-E7 and HSP70-L1-L2-E7 constructs as promising vaccine candidates. The E7 protein, used alone or in combination with other HPV proteins, particularly E6, has been a focus in the development of therapeutic clinical trials [64]. Moreover, vaccine constructs incorporating the E7 protein along with L1 and L2 proteins were evaluated. The expression of both multiepitope DNA constructs was analyzed in a mammalian cell line. Finally, their immunological and anti-tumor effects were assessed in C57BL/6 mice, demonstrating promising results [63].
3.5. Chimeras Designed for New HPV Vaccines
Currently, three licensed vaccines are based on the mixture of L1 VLPs from two, four, or nine different HR-HPVs [66]. Rather than increasing the diversity of L1-based VLPs, emerging strategies focus on developing prophylactic vaccines using chimeric VLPs. These VLPs are formed by recombinant proteins that incorporate highly conserved neutralizing epitopes from the structural protein L2, inserted into the L1 protein. Studies, such as those conducted by Boxus et al. (2016), have demonstrated that chimeric L1/L2 VLPs can effectively display selected L2 epitopes at doses comparable to those of licensed vaccines. Furthermore, these chimeric VLPs induce robust immune and protective responses against multiple HR-HPV types [9]. Such findings suggest that chimeric L1/L2 VLPs represent a promising, cost-effective approach for developing broadly protective vaccines against various HPV types.
The minor capsid protein L2 harbors highly conserved, HPV-neutralizing epitopes that are shared across multiple high-risk HPV types [67]. Recent research has explored the potential of broadening vaccine-mediated protection through AS04-adjuvanted vaccines based on chimeric L1/L2 constructs. These chimeric VLPs were designed by integrating L2 epitopes into the DE loop and/or C-terminus of the L1 protein. One such construct, an HPV18 L1-based chimeric VLP, incorporated HPV33 L2 epitopes (amino acid residues 17–36) into the L1 DE loop and HPV58 L2 epitopes (amino acid residues 56–75) into the L1 C-terminus. This chimeric VLP elicited durable immune responses and demonstrated cross-protection against multiple HPV types, including HPV6, 11, 16, 31, 35, 39, 45, 58, and 59, as evidenced by pseudovirion and quasivirion assays in mouse and rabbit models. In rabbits, the breadth and magnitude of protection were further amplified when the chimeric L1/L2 VLP was administered alongside HPV16/18 L1 VLPs [9]. These findings highlight the potential of the HPV18 L1/L2 chimeric VLP, particularly when formulated with AS04 and combined with HPV16/18 L1 VLPs, as a promising strategy for achieving broad-spectrum protection against diverse HPV types in humans.
Personalized cancer immunotherapies, combined with nanotechnology (nanovaccines), are revolutionizing treatment strategies for HPV-related cancers, complementing advancements in chimeric HPV vaccine studies. Chimeric HPV vaccines, designed to incorporate L1 and L2 epitopes from multiple HPV types, aim to broaden the protective immune response and prevent a wider range of HPV infections [7,9]. While these vaccines address prevention, personalized nanovaccines focus on treating existing cancers by tailoring therapies to individual tumors' unique genetic and molecular profiles [10].
Nanotechnology plays a pivotal role in both approaches. In chimeric HPV vaccines, nanoparticles enhance the stability and immunogenicity of VLPs, improving their ability to elicit broad and durable immune responses. Similarly, in personalized cancer vaccines, nanocarriers such as polymeric nanoparticles and high-density lipoprotein-mimicking nano-discs facilitate efficient delivery of tumor-specific antigens, boosting immune activation and precision in targeting HPV-related malignancies [68].
Recent preclinical studies highlight the potential of chimeric L1/L2 VLPs to provide cross-protection against diverse HPV types beyond those targeted by current prophylactic vaccines. Personalized cancer nanovaccines build on this progress by addressing therapeutic gaps and enabling the treatment of HPV-related cancers such as cervical and oropharyngeal cancers. Additionally, integrating Proteolysis-Targeting Chimeras (PROTACs) with nanotechnology offers a novel approach to selectively degrade oncogenic HPV proteins, enhancing treatment efficacy. The synergy between chimeric HPV vaccines, personalized nanovaccines, and advanced technologies like PROTACs presents a comprehensive strategy to combat HPV-related diseases, overcoming challenges in nanocarrier optimization and safety to provide innovative solutions for both prevention and treatment [69].
4. Viral Structure and Physical Properties
Determining the structure and physical properties of viruses is a complex process that involves several scientific techniques and approaches [70,71,72]. The following outlines present the main methods and factors contributing to this determination.
4.1. Imaging Methods
- Electron Microscopy (EM) is one of the main techniques for visualizing viral morphology. It allows the observation of viruses at high resolution, revealing details about the shape, size, and organization of the capsid and envelope, if present (Figure 4.A and B).
- X-ray Crystallography allows the determination of the three-dimensional structure of viral proteins at the atomic level, providing detailed information about the arrangement of proteins in the capsid.
4.2. Spectroscopy
- Nuclear Magnetic Resonance Spectroscopy (NMR) can be used to study the structure of viral proteins in solutions, helping to understand how proteins interact with each other and with the genetic material.
- Mass Spectrometry (MS) can be used to analyze the protein composition of viruses, identifying the present proteins and their post-translational modifications.
4.3. Biochemical Analysis
- Virus Purification Studies allow the analysis of their physical properties, such as size, shape, and stability. Methods such as density gradient centrifugation help separate viruses from other cellular components.
- Stability Assays Tests under different conditions (pH, temperature, presence of solvents) help determine viruses' physical properties, such as their resistance and sensitivity to disinfectants.
4.4. Computational Modeling
- Molecular Simulations, performed in computational models, can predict the structure and interactions of viral proteins, helping to understand how the structure influences the virus's function and pathogenicity (Figure 3.A and B).
4.5. Genomics and Proteomics
- Virus Genetic Sequencing provides information about the organization and composition of the viral genome, which is essential to understanding the structure and function of the encoded proteins.
- Proteomic Analysis is a fundamental approach to identifying and quantifying viral proteins, improving our understanding of viral biology and interactions with host cells. The following topic discusses how proteomic analysis is crucial in viral studies.
4.6. Highlights in Proteomic Analysis
4.6.1. Viral Protein Identification
The first step in proteomic analysis is the extraction of viral proteins from purified viral particles or infected cells. This may involve cell lysis and purification of viral proteins using methods such as density gradient centrifugation. The extracted proteins are often digested into smaller peptides using enzymes such as trypsin. This facilitates subsequent analysis, as the peptides are easier to separate and identify [74].
Mass Spectrometry (MS) is a technique widely used to identify proteins. The peptides generated by digestion are ionized and analyzed in a mass spectrometer, which provides information about the mass of the peptides. Comparison of the data obtained with protein sequence databases allows the identification of viral proteins [74,75].
4.6.2. Quantification of Viral Proteins
Viral proteins can be quantified using absolute and relative quantification methods. The absolute quantification methods determine the exact amount of a protein in a sample, while the relative quantified methods compare the abundance of proteins in different samples. The relative quantification of samples is often performed using techniques such as tandem mass spectrometry (MS/MS) [76].
Moreover, labeling techniques, such as TMT (Tandem Mass Tag) or iTRAQ (Isobaric Tags for Relative and Absolute Quantitation), allow the comparison of protein abundance in different samples in a single experiment, facilitating the analysis of variations in protein expression in response to conditions, such as viral infection [75].
4.6.3. Functional Analysis
Proteomic analysis can also be applied to study interactions between viral and host cell proteins. Functional analysis of protein-protein interactions (PPIs) is a crucial area of research that helps to understand how viruses interact with host cells and how these interactions influence infection, viral replication, and pathogenicity [77].
PPIs are fundamental to the biology of viruses, playing a crucial role in their life cycle, immune evasion, and regulation of host cellular processes. These interactions are vital for various stages of the viral life cycle, including attachment, entry, replication, assembly, and release. Viruses also exploit PPIs to evade the immune system. Many viral proteins disrupt immune signaling pathways, inhibiting the production of interferons or blocking antigen presentation to avoid detection. Additionally, some viruses interact with host apoptosis-regulating proteins, delaying programmed cell death and prolonging the lifespan of the infected cell, thereby maximizing their replication. Beyond immune evasion, certain viral proteins alter host gene expression to create an environment favorable for replication, often suppressing antiviral responses while enhancing processes beneficial to the virus [74,77].
Studying PPIs provides critical insights into these mechanisms, employing various methods to identify and analyze these interactions. Techniques such as co-immunoprecipitation isolate proteins of interest and their binding partners for analysis using mass spectrometry or Western blotting [75]. The last two-hybrid assays allow the detection of interactions by linking proteins of interest to transcription factor domains, while fluorescence resonance energy transfer (FRET) uses fluorophores to monitor interactions in real time within living cells [78]. Fluorescence microscopy further enhances understanding by visualizing the subcellular localization of viral proteins and their interactions with host components [79].
Functional analysis of PPIs reveals how viruses manipulate host cells, leading to profound biological effects. By engaging with host cell machinery, viral proteins can disrupt normal cellular physiology, such as altering the cell cycle, which can result in uncontrolled proliferation. These interactions also contribute to the pathogenicity of viruses, determining their ability to cause disease. The last few decades have seen significant advancements in understanding these processes, with the COVID-19 pandemic intensifying global research efforts [74,75].
Future virology research is poised to address critical challenges, including cataloging the genomic and phenotypic diversity of viruses, exploring mechanisms of disease emergence, and developing innovative therapies, vaccines, and interventions. Investigating host-virus interactions in greater depth and understanding the role of viruses in chronic diseases are also pivotal areas of focus. Additionally, leveraging viruses as tools in cell biology promises to unlock new scientific frontiers. These endeavors, spanning scientific, social, and political dimensions, will shape the field of virology for decades to come [80].
Acknowledgments
The authors would like to thank the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil) for the postdoctoral fellowship to C.S.O, and Instituto Butantan and Fundação Butantan for the financial support to A.M.C.
References
- Ahmels, M.; Mariz, F.C.; Braspenning-Wesch, I.; Stephan, S.; Huber, B.; Schmidt, G.; Cao, R.; Müller, M.; Kirnbauer, R.; Rösl, F.; et al. Next Generation L2-Based HPV Vaccines Cross-Protect against Cutaneous Papillomavirus Infection and Tumor Development. Front. Immunol. 2022, 13, 1010790. [CrossRef]
- Gardella, B.; Pasquali, M.F.; La Verde, M.; Cianci, S.; Torella, M.; Dominoni, M. The Complex Interplay between Vaginal Microbiota, HPV Infection, and Immunological Microenvironment in Cervical Intraepithelial Neoplasia: A Literature Review. IJMS 2022, 23, 7174. [CrossRef]
- Mohsen, M.O.; Bachmann, M.F. Virus-like Particle Vaccinology, from Bench to Bedside. Cell Mol Immunol 2022, 19, 993–1011. [CrossRef]
- Yan, D.; Wei, Y.-Q.; Guo, H.-C.; Sun, S.-Q. The Application of Virus-like Particles as Vaccines and Biological Vehicles. Appl Microbiol Biotechnol 2015, 99, 10415–10432. [CrossRef]
- Kheirvari, M.; Liu, H.; Tumban, E. Virus-like Particle Vaccines and Platforms for Vaccine Development. Viruses 2023, 15, 1109. [CrossRef]
- Crisci, E.; Bárcena, J.; Montoya, M. Virus-like Particle-Based Vaccines for Animal Viral Infections. Inmunología 2013, 32, 102–116. [CrossRef]
- Bárcena, J.; Blanco, E. Design of Novel Vaccines Based on Virus-Like Particles or Chimeric Virions. In Structure and Physics of Viruses; Mateu, M.G., Ed.; Subcellular Biochemistry; Springer Netherlands: Dordrecht, 2013; Vol. 68, pp. 631–665 ISBN 978-94-007-6551-1.
- González-Domínguez, I.; Lorenzo, E.; Bernier, A.; Cervera, L.; Gòdia, F.; Kamen, A. A Four-Step Purification Process for Gag VLPs: From Culture Supernatant to High-Purity Lyophilized Particles. Vaccines 2021, 9, 1154. [CrossRef]
- Boxus, M.; Fochesato, M.; Miseur, A.; Mertens, E.; Dendouga, N.; Brendle, S.; Balogh, K.K.; Christensen, N.D.; Giannini, S.L. Broad Cross-Protection Is Induced in Preclinical Models by a Human Papillomavirus Vaccine Composed of L1/L2 Chimeric Virus-Like Particles. J Virol 2016, 90, 6314–6325. [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like Particles: Preparation, Immunogenicity and Their Roles as Nanovaccines and Drug Nanocarriers. J Nanobiotechnol 2021, 19, 59. [CrossRef]
- Petrov, G.V.; Galkina, D.A.; Koldina, A.M.; Grebennikova, T.V.; Eliseeva, O.V.; Chernoryzh, Y.Yu.; Lebedeva, V.V.; Syroeshkin, A.V. Controlling the Quality of Nanodrugs According to Their New Property—Radiothermal Emission. Pharmaceutics 2024, 16, 180. [CrossRef]
- Structure and Physics of Viruses: An Integrated Guide; Mateu, M.G., Ed.; Subcellular Biochemistry; Springer Nature Switzerland: Cham, 2024; Vol. 105; ISBN 978-3-031-65186-1.
- Sakauchi, D.; Sasaki, E.A.K.; Anni, F.O.B.; Cianciarullo, A.M. Production of HPV16 Capsid Proteins in Suspension Cultures of Human Epithelial Cells. World J. Adv. Res. Rev. 2021, 9, 258–268. [CrossRef]
- Bei, L.; Gao, S.; Zhao, D.; Kou, Y.; Liang, S.; Wu, Y.; Zhang, X.; Meng, D.; Lu, J.; Luo, C.; et al. Immunogenicity Assessment of a 14-Valent Human Papillomavirus Vaccine Candidate in Mice. Vaccines 2024, 12, 1262. [CrossRef]
- Wang, R.; Huang, H.; Yu, C.; Li, X.; Wang, Y.; Xie, L. Current Status and Future Directions for the Development of Human Papillomavirus Vaccines. Front. Immunol. 2024, 15, 1362770. [CrossRef]
- Gupta, R.; Arora, K.; Roy, S.S.; Joseph, A.; Rastogi, R.; Arora, N.M.; Kundu, P.K. Platforms, Advances, and Technical Challenges in Virus-like Particles-Based Vaccines. Front. Immunol. 2023, 14, 1123805. [CrossRef]
- Nierengarten, M.B. Men Face Substantial Lifelong Risk of Oral HPV Infection: A Study Provides Information on Rates of Newly Acquired Oral HPV Infections and Associated Risk Factors for Acquiring HPV Infections in a Multinational Cohort of More than 3000 Men. Cancer 2025, 131, e35714. [CrossRef]
- Zhao, Q.; Modis, Y.; High, K.; Towne, V.; Meng, Y.; Wang, Y.; Alexandroff, J.; Brown, M.; Carragher, B.; Potter, C.S.; et al. Disassembly and Reassembly of Human Papillomavirus Virus-like Particles Produces More Virion-like Antibody Reactivity. Virol J 2012, 9, 52. [CrossRef]
- Lowe, J.; Panda, D.; Rose, S.; Jensen, T.; Hughes, W.A.; Tso, F.Y.; Angeletti, P.C. Evolutionary and Structural Analyses of Alpha-Papillomavirus Capsid Proteins Yields Novel Insights into L2 Structure and Interaction with L1. Virol J 2008, 5, 150. [CrossRef]
- Zella, D.; Gallo, R.C. Viruses and Bacteria Associated with Cancer: An Overview. Viruses 2021, 13, 1039. [CrossRef]
- Singh, D.; Vignat, J.; Lorenzoni, V.; Eslahi, M.; Ginsburg, O.; Lauby-Secretan, B.; Arbyn, M.; Basu, P.; Bray, F.; Vaccarella, S. Global Estimates of Incidence and Mortality of Cervical Cancer in 2020: A Baseline Analysis of the WHO Global Cervical Cancer Elimination Initiative. The Lancet Global Health 2023, 11, e197–e206. [CrossRef]
- Adelstein, D.J.; Rodriguez, C.P. Human Papillomavirus: Changing Paradigms in Oropharyngeal Cancer. Curr Oncol Rep 2010, 12, 115–120. [CrossRef]
- Pickard, R.K.L.; Xiao, W.; Broutian, T.R.; He, X.; Gillison, M.L. The Prevalence and Incidence of Oral Human Papillomavirus Infection Among Young Men and Women, Aged 18–30 Years. Sexually Transmitted Diseases 2012, 39, 559–566. [CrossRef]
- Andersen, A.S.; Koldjær Sølling, A.S.; Ovesen, T.; Rusan, M. The Interplay between HPV and Host Immunity in Head and Neck Squamous Cell Carcinoma. Intl Journal of Cancer 2014, 134, 2755–2763. [CrossRef]
- De Vuyst, H.; Clifford, G.M.; Nascimento, M.C.; Madeleine, M.M.; Franceschi, S. Prevalence and Type Distribution of Human Papillomavirus in Carcinoma and Intraepithelial Neoplasia of the Vulva, Vagina and Anus: A Meta-analysis. Intl Journal of Cancer 2009, 124, 1626–1636. [CrossRef]
- De Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide Burden of Cancer Attributable to HPV by Site, Country and HPV Type. Intl Journal of Cancer 2017, 141, 664–670. [CrossRef]
- Accardi, R.; Gheit, T. Cutaneous HPV and Skin Cancer. La Presse Médicale 2014, 43, e435–e443. [CrossRef]
- Paolini, F.; Cota, C.; Amantea, A.; Curzio, G.; Venuti, A. Mucosal Alpha-Papillomavirus (HPV89) in a Rare Skin Lesion. Virol J 2015, 12, 105. [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J Clinicians 2024, 74, 229–263. [CrossRef]
- International Agency for Reasearch on Cancer (IARC) Cancer Today.
- Brazil Healthy Ministry NOTA TÉCNICA No 41/2024-CGICI/DPNI/SVSA/MS 2024.
- Cianciarullo, A.M. Revista SODEBRAS. 2014, pp. 134–141.
- INCA, I.N. de C. Prevenção do câncer do colo do útero Available online: https://www.gov.br/inca/pt-br/assuntos/gestor-e-profissional-de-saude/controle-do-cancer-do-colo-do-utero/acoes/prevencao (accessed on 18 January 2025).
- Herrero, R.; González, P.; Markowitz, L.E. Present Status of Human Papillomavirus Vaccine Development and Implementation. The Lancet Oncology 2015, 16, e206–e216. [CrossRef]
- Wang, D.; Li, Z.; Xiao, J.; Wang, J.; Zhang, L.; Liu, Y.; Fan, F.; Xin, L.; Wei, M.; Kong, Z.; et al. Identification of Broad-Genotype HPV L2 Neutralization Site for Pan-HPV Vaccine Development by a Cross-Neutralizing Antibody. PLoS ONE 2015, 10, e0123944. [CrossRef]
- Vande Pol, S.B.; Klingelhutz, A.J. Papillomavirus E6 Oncoproteins. Virology 2013, 445, 115–137. [CrossRef]
- Cianciarullo, A.M.; Sasaki, E.A.K. Process of Producing an Immunological Composition of Prophylatic and Therapeutic DNA against HPV and Cancers Associated with the Virus, Hybrid Protein, Expression Vector, Immunologial Composition, and Its Use. Patent 2022.
- Cianciarullo, A.M.; Sasaki, E.A.K. PROCESSO DE PRODUÇÃO DE UMA COMPOSIÇÃO IMUNOLÓGICA DE DNA PROFILÁTICA E TERAPÊUTICA CONTRA HPV E CÂNCERES ASSOCIADOS AO VÍRUS, PROTEÍNA HÍBRIDA, VETOR DE EXPRESSÃO, COMPOSIÇÃO IMUNOLÓGICA E SEUS USOS. Patent 2019.
- Hancock, G.; Hellner, K.; Dorrell, L. Therapeutic HPV Vaccines. Best Practice & Research Clinical Obstetrics & Gynaecology 2018, 47, 59–72. [CrossRef]
- Hamley, I.W. Peptides for Vaccine Development. ACS Appl. Bio Mater. 2022, 5, 905–944. [CrossRef]
- Jaradat, D. Thirteen Decades of Peptide Synthesis: Key Developments in Solid Phase Peptide Synthesis and Amide Bond Formation Utilized in Peptide Ligation. Amino Acids 2018, 50, 1–30. [CrossRef]
- Merrifield, B. [1] Concept and Early Development of Solid-Phase Peptide Synthesis. In Methods in Enzymology; Elsevier, 1997; Vol. 289, pp. 3–13 ISBN 978-0-12-182190-6.
- El-Baky, N.A.; Elkhawaga, M.A.; Abdelkhalek, E.S.; Sharaf, M.M.; Redwan, E.M.; Kholef, H.R. De Novo Expression and Antibacterial Potential of Four Lactoferricin Peptides in Cell-Free Protein Synthesis System. Biotechnology Reports 2021, 29, e00583. [CrossRef]
- Torres, M.D.T.; Cao, J.; Franco, O.L.; Lu, T.K.; De La Fuente-Nunez, C. Synthetic Biology and Computer-Based Frameworks for Antimicrobial Peptide Discovery. ACS Nano 2021, 15, 2143–2164. [CrossRef]
- Tîrziu, A.; Avram, S.; Mada, L.; Crișan-Vida, M.; Popovici, C.; Popovici, D.; Faur, C.; Duda-Seiman, C.; Păunescu, V.; Vernic, C. Design of a Synthetic Long Peptide Vaccine Targeting HPV-16 and -18 Using Immunoinformatic Methods. Pharmaceutics 2023, 15, 1798. [CrossRef]
- Zhai, L.; Peabody, J.; Pang, Y.-Y.S.; Schiller, J.; Chackerian, B.; Tumban, E. A Novel Candidate HPV Vaccine: MS2 Phage VLP Displaying a Tandem HPV L2 Peptide Offers Similar Protection in Mice to Gardasil-9. Antiviral Research 2017, 147, 116–123. [CrossRef]
- Namvar, A.; Bolhassani, A.; Javadi, G.; Noormohammadi, Z. In Silico/In Vivo Analysis of High-Risk Papillomavirus L1 and L2 Conserved Sequences for Development of Cross-Subtype Prophylactic Vaccine. Sci Rep 2019, 9, 15225. [CrossRef]
- Ronchi, V.P.; Klein, J.M.; Edwards, D.J.; Haas, A.L. The Active Form of E6-Associated Protein (E6AP)/UBE3A Ubiquitin Ligase Is an Oligomer. Journal of Biological Chemistry 2014, 289, 1033–1048. [CrossRef]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2020, 10, 3116. [CrossRef]
- Greenfield, W.W.; Stratton, S.L.; Myrick, R.S.; Vaughn, R.; Donnalley, L.M.; Coleman, H.N.; Mercado, M.; Moerman-Herzog, A.M.; Spencer, H.J.; Andrews-Collins, N.R.; et al. A Phase I Dose-Escalation Clinical Trial of a Peptide-Based Human Papillomavirus Therapeutic Vaccine with Candida Skin Test Reagent as a Novel Vaccine Adjuvant for Treating Women with Biopsy-Proven Cervical Intraepithelial Neoplasia 2/3. OncoImmunology 2015, 4, e1031439. [CrossRef]
- Van Poelgeest, M.I.E.; Welters, M.J.P.; Van Esch, E.M.G.; Stynenbosch, L.F.M.; Kerpershoek, G.; Van Persijn Van Meerten, E.L.; Van Den Hende, M.; Löwik, M.J.G.; Berends-van Der Meer, D.M.A.; Fathers, L.M.; et al. HPV16 Synthetic Long Peptide (HPV16-SLP) Vaccination Therapy of Patients with Advanced or Recurrent HPV16-Induced Gynecological Carcinoma, a Phase II Trial. J Transl Med 2013, 11, 88. [CrossRef]
- De Vos Van Steenwijk, P.J.; Van Poelgeest, M.I.E.; Ramwadhdoebe, T.H.; Löwik, M.J.G.; Berends-van Der Meer, D.M.A.; Van Der Minne, C.E.; Loof, N.M.; Stynenbosch, L.F.M.; Fathers, L.M.; Valentijn, A.R.P.M.; et al. The Long-Term Immune Response after HPV16 Peptide Vaccination in Women with Low-Grade Pre-Malignant Disorders of the Uterine Cervix: A Placebo-Controlled Phase II Study. Cancer Immunol Immunother 2014, 63, 147–160. [CrossRef]
- Gomes, D.; Silvestre, S.; Duarte, A.P.; Venuti, A.; Soares, C.P.; Passarinha, L.; Sousa, Â. In Silico Approaches: A Way to Unveil Novel Therapeutic Drugs for Cervical Cancer Management. Pharmaceuticals 2021, 14, 741. [CrossRef]
- Matos, A.S.; Invenção, M.D.C.V.; Moura, I.A.D.; Freitas, A.C.D.; Batista, M.V.D.A. Immunoinformatics Applications in the Development of Therapeutic Vaccines against Human Papillomavirus-related Infections and Cervical Cancer. Reviews in Medical Virology 2023, 33, e2463. [CrossRef]
- Yang, A.; Farmer, E.; Wu, T.C.; Hung, C.-F. Perspectives for Therapeutic HPV Vaccine Development. J Biomed Sci 2016, 23, 75. [CrossRef]
- Yang, A.; Jeang, J.; Cheng, K.; Cheng, T.; Yang, B.; Wu, T.-C.; Hung, C.-F. Current State in the Development of Candidate Therapeutic HPV Vaccines. Expert Review of Vaccines 2016, 15, 989–1007. [CrossRef]
- Kayyal, M.; Bolhassani, A.; Noormohammadi, Z.; Sadeghizadeh, M. In Silico Design and Immunological Studies of Two Novel Multiepitope DNA-Based Vaccine Candidates Against High-Risk Human Papillomaviruses. Mol Biotechnol 2021, 63, 1192–1222. [CrossRef]
- Kaufmann, A.M.; Nieland, J.D.; Jochmus, I.; Baur, S.; Friese, K.; Gabelsberger, J.; Gieseking, F.; Gissmann, L.; Glasschröder, B.; Grubert, T.; et al. Vaccination Trial with HPV16 L1E7 Chimeric Virus-like Particles in Women Suffering from High Grade Cervical Intraepithelial Neoplasia (CIN 2/3). Intl Journal of Cancer 2007, 121, 2794–2800. [CrossRef]
- Garcia, F.; Petry, K.U.; Muderspach, L.; Gold, M.A.; Braly, P.; Crum, C.P.; Magill, M.; Silverman, M.; Urban, R.G.; Hedley, M.L.; et al. ZYC101a for Treatment of High-Grade Cervical Intraepithelial Neoplasia: A Randomized Controlled Trial. Obstetrics & Gynecology 2004, 103, 317–326. [CrossRef]
- Gaillard, S.; Deery, A.; Fader, A.N.; Huh, W.; Liang, M.; Straughn, M.; Arend, R.; Wu, T.C.; Leath, C.; Roden, R. Safety and Feasibility of an HPV Therapeutic Vaccine (TA-CIN) in Patients with HPV16 Associated Cervical Cancer (458). Gynecologic Oncology 2022, 166, S228–S229. [CrossRef]
- Kaufmann, A.M.; Stern, P.L.; Rankin, E.M.; Sommer, H.; Nuessler, V.; Schneider, A.; Adams, M.; Onon, T.S.; Bauknecht, T.; Wagner, U.; et al. Safety and Immunogenicity of TA-HPV, a Recombinant Vaccinia Virus Expressing Modified Human Papillomavirus (HPV)-16 and HPV-18 E6 and E7 Genes, in Women with Progressive Cervical Cancer. Clin Cancer Res 2002, 8, 3676–3685.
- Roman, L.D.; Wilczynski, S.; Muderspach, L.I.; Burnett, A.F.; O’Meara, A.; Brinkman, J.A.; Kast, W.M.; Facio, G.; Felix, J.C.; Aldana, M.; et al. A Phase II Study of Hsp-7 (SGN-00101) in Women with High-Grade Cervical Intraepithelial Neoplasia. Gynecologic Oncology 2007, 106, 558–566. [CrossRef]
- Kayyal, M.; Bolhassani, A.; Noormohammadi, Z.; Sadeghizadeh, M. Immunological Responses and Anti-Tumor Effects of HPV16/18 L1-L2-E7 Multiepitope Fusion Construct along with Curcumin and Nanocurcumin in C57BL/6 Mouse Model. Life Sciences 2021, 285, 119945. [CrossRef]
- Bahmani, B.; Amini-bayat, Z.; Ranjbar, M.M.; Bakhtiari, N.; Zarnani, A.-H. HPV16-E7 Protein T Cell Epitope Prediction and Global Therapeutic Peptide Vaccine Design Based on Human Leukocyte Antigen Frequency: An In-Silico Study. Int J Pept Res Ther 2021, 27, 365–378. [CrossRef]
- Gupta, S.K.; Singh, A.; Srivastava, M.; Gupta, S.K.; Akhoon, B.A. In Silico DNA Vaccine Designing against Human Papillomavirus (HPV) Causing Cervical Cancer. Vaccine 2009, 28, 120–131. [CrossRef]
- Zhu, F.-C.; Zhong, G.-H.; Huang, W.-J.; Chu, K.; Zhang, L.; Bi, Z.-F.; Zhu, K.-X.; Chen, Q.; Zheng, T.-Q.; Zhang, M.-L.; et al. Head-to-Head Immunogenicity Comparison of an Escherichia Coli-Produced 9-Valent Human Papillomavirus Vaccine and Gardasil 9 in Women Aged 18–26 Years in China: A Randomised Blinded Clinical Trial. The Lancet Infectious Diseases 2023, 23, 1313–1322. [CrossRef]
- Gambhira, R.; Karanam, B.; Jagu, S.; Roberts, J.N.; Buck, C.B.; Bossis, I.; Alphs, H.; Culp, T.; Christensen, N.D.; Roden, R.B.S. A Protective and Broadly Cross-Neutralizing Epitope of Human Papillomavirus L2. J Virol 2007, 81, 13927–13931. [CrossRef]
- Zhou, X.; Lian, H.; Li, H.; Fan, M.; Xu, W.; Jin, Y. Nanotechnology in Cervical Cancer Immunotherapy: Therapeutic Vaccines and Adoptive Cell Therapy. Front. Pharmacol. 2022, 13, 1065793. [CrossRef]
- Mukerjee, N.; Maitra, S.; Gorai, S.; Ghosh, A.; Alexiou, A.; Thorat, N.D. Revolutionizing Human Papillomavirus (HPV)-related Cancer Therapies: Unveiling the Promise of Proteolysis Targeting Chimeras (PROTACs) and Proteolysis Targeting Antibodies (PROTABs) in Cancer Nano-vaccines. Journal of Medical Virology 2023, 95, e29135. [CrossRef]
- Structure and Physics of Viruses: An Integrated Textbook; Mateu, M.G., Ed.; Subcellular Biochemistry; Springer Netherlands: Dordrecht, 2013; Vol. 68; ISBN 978-94-007-6551-1.
- Johnson, J.E.; Speir, J.A. Virus Particle Structure: Principles. In Encyclopedia of Virology; Elsevier, 2008; pp. 393–401 ISBN 978-0-12-374410-4.
- Johnson, J.E. Multi-Disciplinary Studies of Viruses: The Role of Structure in Shaping the Questions and Answers. Journal of Structural Biology 2008, 163, 246–253. [CrossRef]
- Cianciarullo A.M.; Oliveira C.S.; Vicente P.I.; Sakauchi D.; Sasaki E.A.K. Prophylactic and Therapeutic Vaccines against HPV and Associated Cancers; 1st ed.; CRV Publisher, 2024; ISBN 978-65-251-5296-7.
- Lum, K.K.; Cristea, I.M. Proteomic Approaches to Uncovering Virus–Host Protein Interactions during the Progression of Viral Infection. Expert Review of Proteomics 2016, 13, 325–340. [CrossRef]
- Zheng, J.; Tan, B.H.; Sugrue, R.; Tang, K. Current Approaches on Viral Infection: Proteomics and Functional Validations. Front. Microbio. 2012, 3. [CrossRef]
- Balvers, M.; Gordijn, I.F.; Voskamp-Visser, I.A.I.; Schelling, M.F.A.; Schuurman, R.; Heikens, E.; Braakman, R.; Stingl, C.; Van Leeuwen, H.C.; Luider, T.M.; et al. Proteome2virus: Shotgun Mass Spectrometry Data Analysis Pipeline for Virus Identification. Journal of Clinical Virology Plus 2023, 3, 100147. [CrossRef]
- Saha, D.; Iannuccelli, M.; Brun, C.; Zanzoni, A.; Licata, L. The Intricacy of the Viral-Human Protein Interaction Networks: Resources, Data, and Analyses. Front. Microbiol. 2022, 13, 849781. [CrossRef]
- Kocjan, B.J.; Seme, K.; Poljak, M. Detection and Differentiation of Human Papillomavirus Genotypes HPV-6 and HPV-11 by FRET-Based Real-Time PCR. Journal of Virological Methods 2008, 153, 245–249. [CrossRef]
- Castelletto, S.; Boretti, A. Viral Particle Imaging by Super-Resolution Fluorescence Microscopy. Chemical Physics Impact 2021, 2, 100013. [CrossRef]
- Holmes, E.C.; Krammer, F.; Goodrum, F.D. Virology—The next Fifty Years. Cell 2024, 187, 5128–5145. [CrossRef]
Figure 1.
The difference between HPV virus-like particles (VLPs) and the HPV virus is significant. The L1 protein is a structural protein that groups to form L1 pentamers. These L1 pentamers can self-assemble into a structure known as L1 VLP. The L2 protein is another structural protein associated with HPV; however, it cannot self-assemble. Instead, L2 acts as a stabilizer for the L1 pentamers and co-assembles with them to form chimeric L1/L2 VLPs. The HPV viral particle consists of a structural capsid formed by both L1 and L2 proteins, encapsulating the viral genome inside.
Figure 1.
The difference between HPV virus-like particles (VLPs) and the HPV virus is significant. The L1 protein is a structural protein that groups to form L1 pentamers. These L1 pentamers can self-assemble into a structure known as L1 VLP. The L2 protein is another structural protein associated with HPV; however, it cannot self-assemble. Instead, L2 acts as a stabilizer for the L1 pentamers and co-assembles with them to form chimeric L1/L2 VLPs. The HPV viral particle consists of a structural capsid formed by both L1 and L2 proteins, encapsulating the viral genome inside.
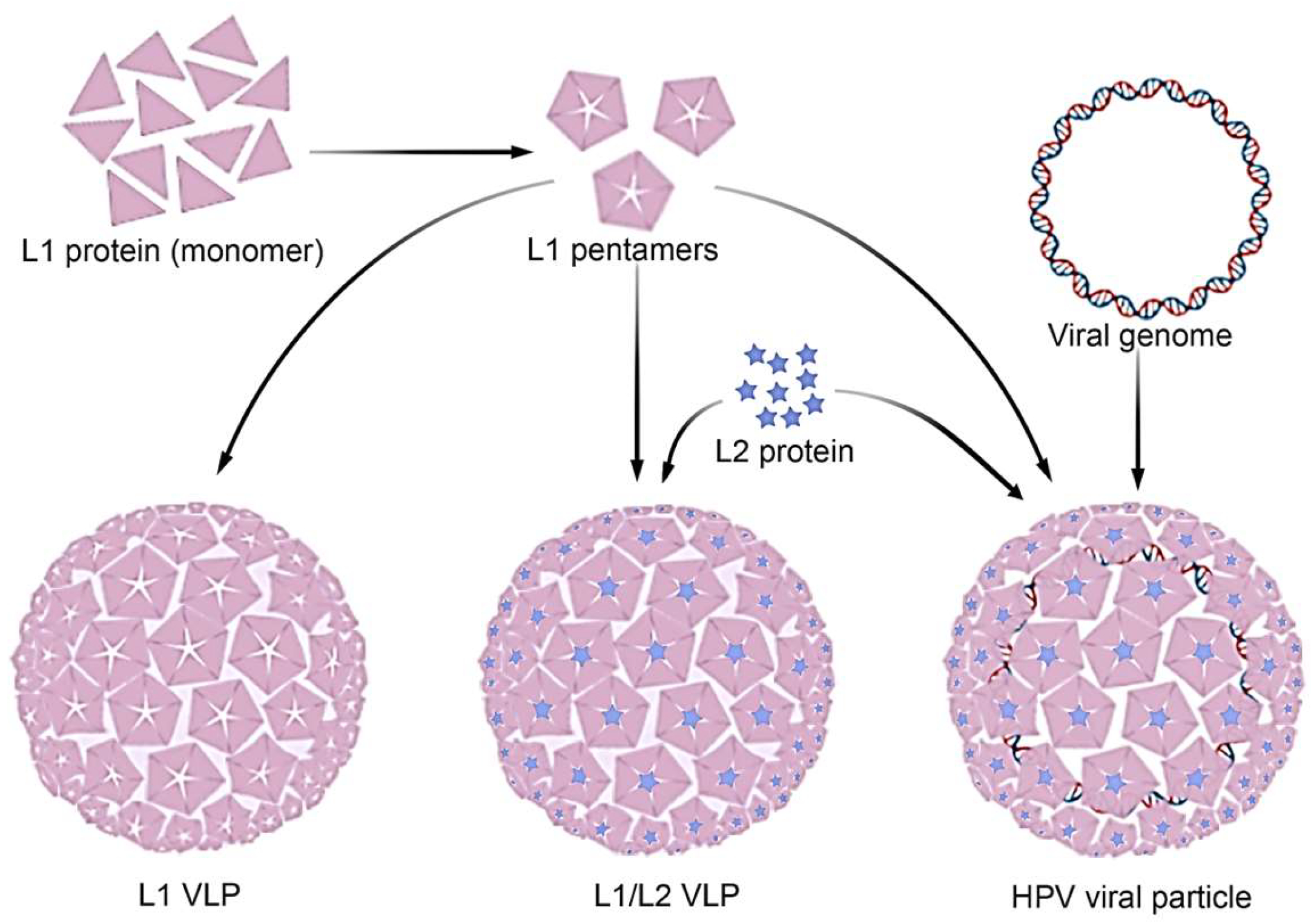
Figure 2.
Schematic steps to obtain VLP: First, the target gene can be inserted into an expression vector, and subsequently, the modified vector can be introduced into a host cell. The protein should then be expressed using the chosen host cell platform. Following expression, the cell products should be processed, followed by VLP maturation and purification (VLP AI model by Nierengarten, 2025 [17]).
Figure 2.
Schematic steps to obtain VLP: First, the target gene can be inserted into an expression vector, and subsequently, the modified vector can be introduced into a host cell. The protein should then be expressed using the chosen host cell platform. Following expression, the cell products should be processed, followed by VLP maturation and purification (VLP AI model by Nierengarten, 2025 [17]).
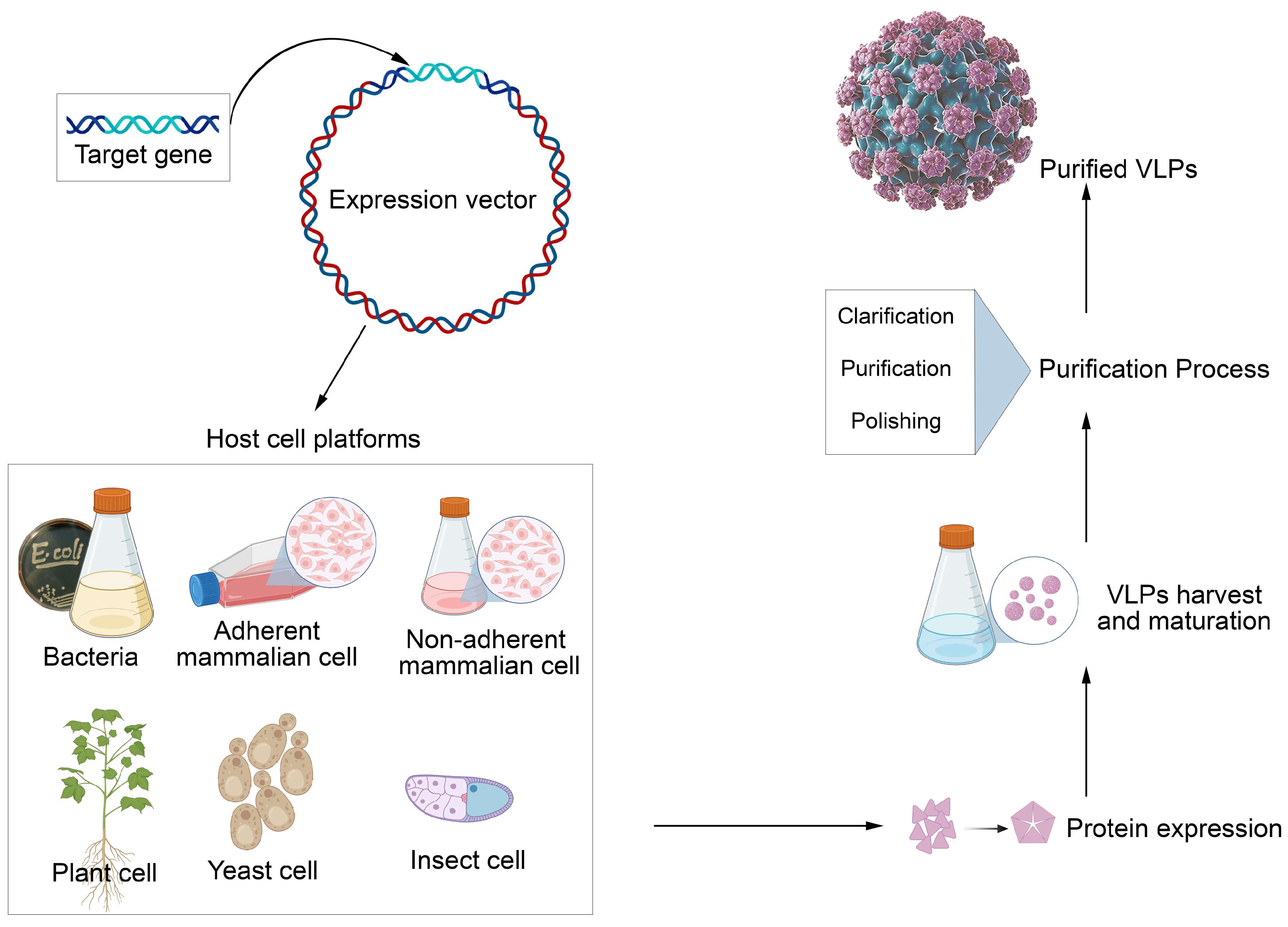
Figure 3.
A) Atomic model showing the T = 7 symmetry of the HPV capsid, highlighting the main capsid protein L1 organized into pentameric capsomeres shown in yellow (Adapted by Zhao et al, 2012 [18]). B) Predicted 3D model of the interaction between L1 and L2 proteins. HPV16 L1 structure was obtained from the RCSB Protein Data Bank and HPV16 L2 protein was predicted using the 3D-Jigsaw and Swiss Modelling Server (Adapted by Lowe et al, 2008 [19]).
Figure 3.
A) Atomic model showing the T = 7 symmetry of the HPV capsid, highlighting the main capsid protein L1 organized into pentameric capsomeres shown in yellow (Adapted by Zhao et al, 2012 [18]). B) Predicted 3D model of the interaction between L1 and L2 proteins. HPV16 L1 structure was obtained from the RCSB Protein Data Bank and HPV16 L2 protein was predicted using the 3D-Jigsaw and Swiss Modelling Server (Adapted by Lowe et al, 2008 [19]).
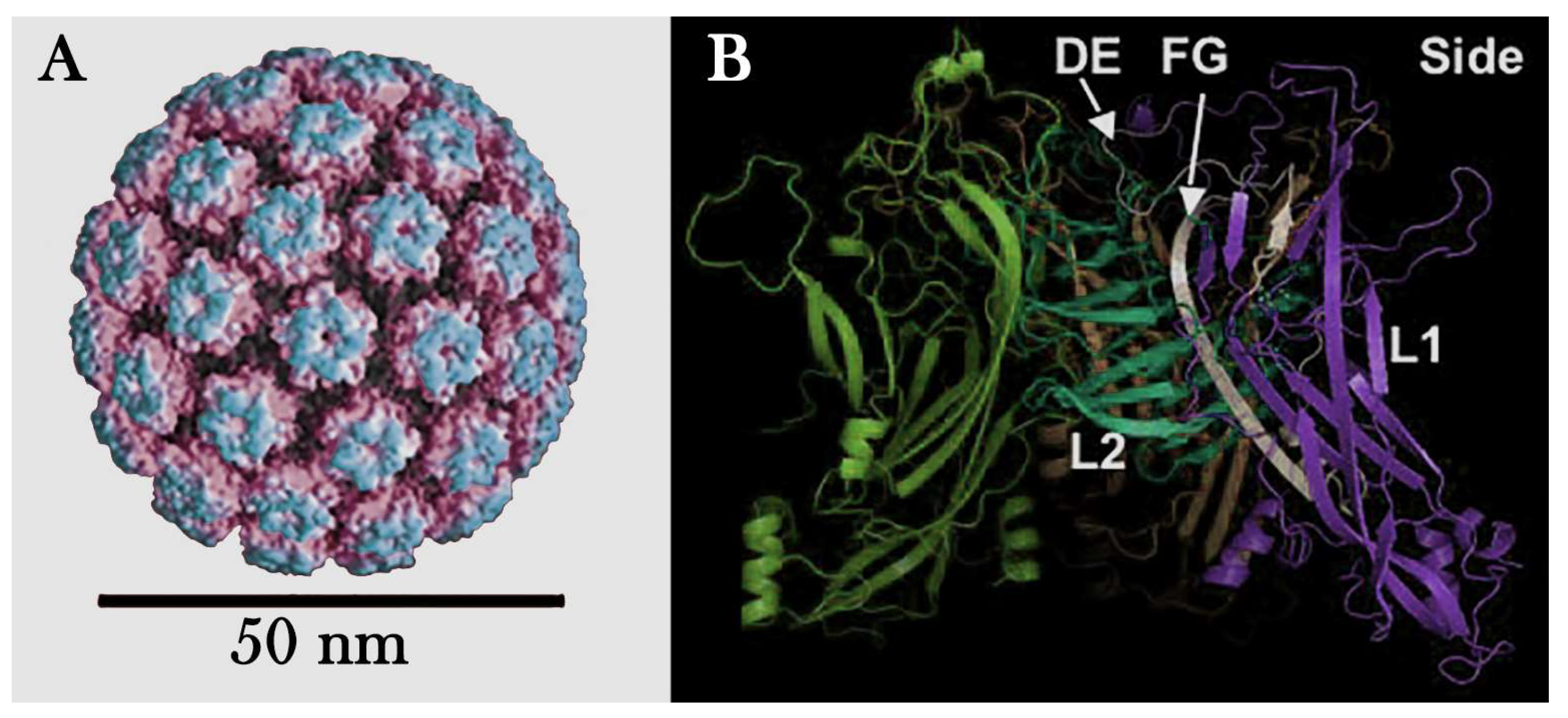
Figure 4.
Chimeric L1/L2 VLPs were produced in a plenty matrix of capsid proteins from HPV16. These proteins were obtained using HEK 293T cells as the host system. A) Transmission electron microscopy images show VLPs in formation, with the larger VLP measuring approximately 30 nm in diameter. Ultrastructural immunocytochemistry detection for L1 and L2 proteins, employing 5 nm (blue arrows) and 10 nm (pink arrows) colloidal gold particles were applied, respectively. B) A highlight showing the L1/L2 VLP of HPV16 in an advanced maturation stage, measuring around 50 nm in diameter. Bars = 50 nm (Images by Cianciarullo et al. 2024 [73]).
Figure 4.
Chimeric L1/L2 VLPs were produced in a plenty matrix of capsid proteins from HPV16. These proteins were obtained using HEK 293T cells as the host system. A) Transmission electron microscopy images show VLPs in formation, with the larger VLP measuring approximately 30 nm in diameter. Ultrastructural immunocytochemistry detection for L1 and L2 proteins, employing 5 nm (blue arrows) and 10 nm (pink arrows) colloidal gold particles were applied, respectively. B) A highlight showing the L1/L2 VLP of HPV16 in an advanced maturation stage, measuring around 50 nm in diameter. Bars = 50 nm (Images by Cianciarullo et al. 2024 [73]).
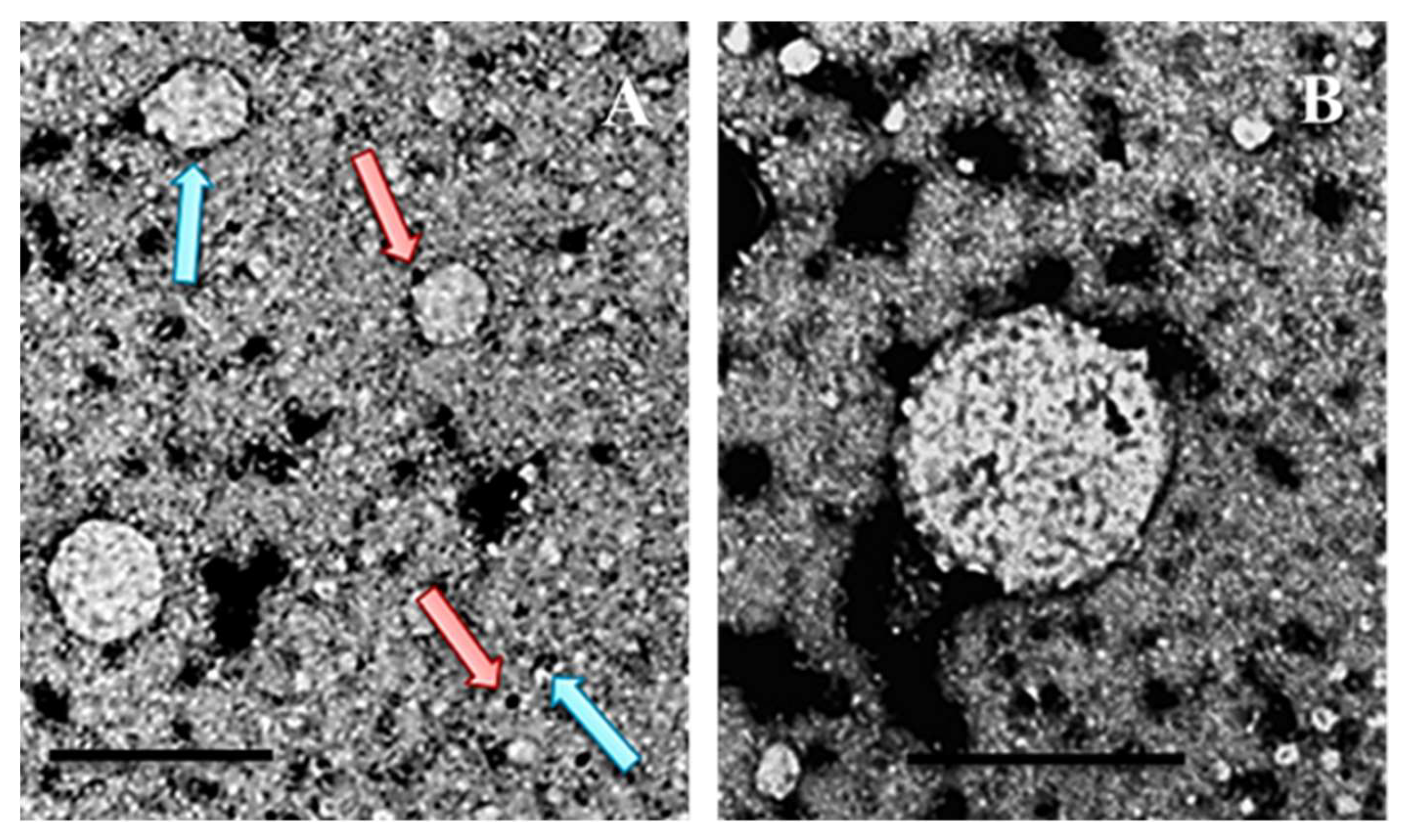

Table 1.
Comparison between prophylactic based-VLP vaccines against HPV.
| Bivalent Cervarix® |
Bivalent Cecolin® |
Bivalent WalrinVax® |
Quadrivalent Gardasil® |
Quadrivalent Cervavac-4* |
Nonavalent Gardasil-9® |
14-valent SCT-1000 |
|
|---|---|---|---|---|---|---|---|
| Approval | 2009 | 2019 | 2022 | 2006 | 2022 | 2014 | Phase III |
| Manufacturer | GlaxoSmith Kline plc |
Xiamen Innovax Co., Ltd | Walvax Co., Ltd |
Merck & Co., Inc | Serum Institute of India | Merck & Co., Inc | SinoCelltech Group |
| VLP HPV Type (protein dose) | HPV-16 (20 µg) HPV-18 (20 µg) |
HPV-16 (40 µg) HPV-18 (20 µg) |
HPV-16 (40 µg) HPV-18 (20 µg) |
HPV-6, 18 (20 µg each) HPV-11, 16 (40 µg each) |
HPV-6, 18 (20 µg each) HPV-11, 16 (40 µg each) |
HPV-6 (30 mg) HPV-16 (60 mg) HPV-11, 18 (40 mg each) HPV-31, 33, 45, 52, 58 (20 mg each) |
HPV-11, 18 (40 µg each) HPV-6 (30 µg) HPV-16 (60 µg) HPV-31, 33, 35, 39, 45, 51, 52, 56, 58, 59 (20 µg each) |
| Expression system | Trichoplusia ni insect cell line | Escherichia coli | Pichia pastoris | Saccharomyces cerevisiae | Hansenula polymorpha | Saccharomyces cerevisiae | SF9 insect cells, transfected with Baculovirus |
| Adjuvant | AS04 | Aluminum hydroxide | Aluminum phosphate | AAHS | Aluminum hydroxide | AAHS | Aluminum phosphate |
* Available only in India.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



