
Submitted:
07 March 2025
Posted:
10 March 2025
You are already at the latest version
Abstract
Neurocognitive disorders (NCD) are neurodegenerative diseases characterized by decline or loss of cognitive function. Two main risk factors for this condition have been identified: age and the APOE genotype. One of the proposed mechanisms of aging is telomere length (TL), as an association between shorter TL and NCD has been suggested. This study investigated the relationship between TL and the APOE genotype in individuals with neurocognitive impairment (CI). 170 participants aged > 60 years were included. Cognitive function was assessed using MMSE and MoCA tests. Relative telomere quantification and APOE genotyping were performed using RT-PCR. A shorter TL was significantly associated with an increased risk of CI (p < 0.001). The APOE ε4 genotype showed a less significant association, with individuals not carrying the ε4 allele displaying a higher risk of CI (p < 0.05). However, a trend toward shorter TL were observed in individuals carrying the ε4 allele with CI compared to those Non-Cognitive Impairment with subjective memory Complaint. Additionally, the number of years of education was negatively correlated with CI (p < 0.0001). Individuals with shorter TL and fewer years of education demonstrated a higher risk of CI. APOE genotype would be a risk factor for shorter telomeres.
Keywords:
Neurocognitive disorder
; telomere length
; APOE
; cognitive impairment
; dementia
; cognitive reserve
1. Introduction
Neurocognitive disorders (NCD) are multifactorial diseases characterized by a loss of cognitive abilities beyond those expected in normal aging. [1,2,3] The main etiology identified is Alzheimer disease (AD), with 80% of all cases and an incidence of 1.5 million in the world. The second most common cause of NCD is vascular dementia. Regardless of etiology, the main risk factors were age and APOE genotype. [4,5,6]
The APOE genotype is one of the most important markers of risk for Alzheimer’s disease; [3] there are three isoforms that represent higher or lower risk in the development of Alzheimer’s disease: ε2 (protecting allele), ε3 (neutral), and ε4 (highest risk).[7] Enough evidence establishes that the presence of the ε4 allele in this gene confers a high risk of developing NCD. [8] These findings are related to animal models, clinical studies, and GWAS analyses. [9,10]
Originally, the APOE gene was identified by linkage studies that explain–15-20% of the inheritance. In 1991, Namba et al. reported by the first time the reactivity of APOE in amyloid plaques, establishing a relationship between the brain and amyloidogenic diseases.[11] Similarly, in vitro experiments of AB peptide binding to membranes and the association with the APOE ε4 genotype were performed, finding that the ε4 allele is a strong genetic risk factor for AD and amyloid deposition. [7]
Immunohistochemistry assays were developed to identify the presence of amyloid proteins in brain diseases associated with different alleles of the APOE gene. In this manner, the relationship between the deposition of β-amyloid protein, APOE genotype, and Alzheimer’s disease was emphasized, a fact that was later corroborated by larger studies. [12,13]
Currently, there are three largest GWAS studies that identify the APOE locus; in 2013, the international genomic profile of Alzheimer’s disease made a meta-analysis including all GWAS, the most extended study made by this disease until 2018. [10,14] This study included data from the Cohort Consortium for Heart and Aging, European Alzheimer's Disease Initiative, and Alzheimer's Disease Genetic and Environmental Risk Consortium. They discovered 14 genomic regions other than the APOE gene, which they considered the most important risk factor. To date, research on the APOE gene is ongoing, because it is important to conduct assays including different types of populations, since most of the studies were conducted in European or Asian populations. [9,15]
Recently, the study of NCD has taken a multifactorial direction, including research involving an increasing number of regulatory mechanisms inherent to aging [16,17], such as metabolic, cellular, and genetic modifications [18,19,20]. Telomeric shortening is a biomarker of biological aging, and telomeres are repetitive DNA sequences associated with proteins that form a complex called Shelterin. These sequences confer stability to telomeres and can function as markers of biological age and aging, since DNA polymerase cannot complete telomere replication, and telomeres wear out after mitotic divisions. [21]
The decrease in telomere length (TL) in humans is related to senescence (biological aging), oxidative stress, and cytotoxicity accumulation. [22] This process has been associated with vascular and neurodegenerative diseases in the elderly population. [23] Some investigators suggest that telomere shortening could be associated with cognitive impairment; however, the findings are contradictory, since evidence of TL with hippocampal volume and cognitive abilities, including memory and executive function. [24,25] Despite the progress in the study of the association between telomere length and cognitive status, some studies have found no significant evidence, such as the case of Demanelis et al., who found that telomere length is related to events present in aging, but not to cognitive impairment. [26,27]
It is important to consider that aging is a process characterized by interindividual differences, not only at the structural, metabolic, and chemical levels that influence brain changes, but also in the ability to compensate for the loss of functions associated with brain damage. [6,16,28] Understanding the pathology of NCD is crucial, as it involves consideration of influential factors such as age.
2. Methods
Genotyping of the APOE gene polymorphisms (rs7412 and rs429358) was performed by real-time PCR (rt-PCR) using TaqMan® probes (Applied Biosystems, San Francisco, CA, USA). For the genotyping of the rs429358 and rs7412 polymorphisms, assays C_3084793_20 and C_904973_10, respectively, were used. Thermocycling conditions followed the manufacturer's recommendations for each assay. Thermocycling and allelic discrimination were performed using the QuantStudio 6 Flex® real-time instrument (Applied Biosystems, San Francisco, CA, USA).
To determine telomere length, rt-PCR was conducted using the QuantStudio 6 Flex system (Thermo Fisher Scientific, Waltham, MA, USA). Standard curves were generated, and TL data were expressed using the 2-∆∆CT method, based on telomere threshold cycle (TC) values and reference gene signals (∆CT). The control gene used was SDHA (succinate dehydrogenase complex flavoprotein subunit A).
The Mini-Mental State Examination (MMSE) and Montreal Cognitive Assessment (MoCA) tests were used for cognitive assessment. MMSE test excludes emotional or behavioral disorders and evaluates in 11 questions the temporospatial orientation, delayed memory, attention and calculation, language and visuoconstructive drawing ability. The maximum score is 30 points, an indicator of normal cognition; in this study, individuals with scores below 26 points were considered cognitively impaired (CI). [30]
On the other hand, the MoCA test evaluates 6 cognitive domains: memory (5 points), visuospatial ability (4 points), executive function (4 points), attention/concentration/working memory (5 points), language (5 points) and orientation (6 points); the maximum sum of this test is 30 points, with a cut-off point for mild cognitive impairment of less than 26 points. In this test, the final score was adjusted for years of study, adding one point in individuals with less than 12 years of study and 2 points in those with less than 8 years of study. [31]
It is important to consider that scores below 26 on the cognitive tests do not exclude the possibility of a decline in some cognitive domain reported by the participants, so they were classified as individuals without cognitive impairment with subjective memory complaint. (NCI-SMC)
3. Statistical Analysis
Chi-square analysis was used to evaluate the association between APOE genotype and the presence or absence of cognitive impairment, comparing the frequency of the APOE-ε4 allele with those of APOE-ε2 and APOE-ε3. Multivariate linear and logistic regression models were applied to assess the relationship between telomere length, age, and years of education with cognitive impairment. All analyses were performed using RStudio (version 2024.04.1).
4. Results
The mean age observed was 70.5 ± 9.22 years; 44.7% (n = 76) were men, and 55.3% (n = 94) were women. The average number of years of education was 13.14 ± 7.16. Among all individuals, 41.8% (n = 71) had cognitive impairment, while 58.2% (n = 99) had Non cognitive Impairment with subjective memory complaint (NCI-SMC). No statistically significant differences in years of education or age were found between men and women with CI and NCI-SMC.

Table 1.
Socio-demographic and clinical characteristics in subjects with CI and NCI-SMC.
| Mean (n=170) | Male (n=76) | Female (n=94) | p-value | |
|---|---|---|---|---|
| Age a | 70.5 (+/-9.224) | 71.61(+/-8.343) | 69.59(+/-9.83) | p>0.05 |
| Years of study a | 13.135(+/-7.156) | 13.22(+/-6.005) | 13.064(+/-7.997) | p>0.05 |
| CI b | 71 (41.76) | 52 (73%) | 47 (66%) | p<0.01** |
| NCI-SMC b | 99 (58.24) | 24 (24%) | 47 (47%) |
CI: Cognitive impairment, NCI-SMC: Non-Cognitive impairment with subjective memory complaint. aThe mean and standard deviation are shown in parentheses. b Indicates the frequency and percentage in parentheses calculated with the total sample. The p value is shown as a Chi-square function for the comparison of the variables years of study, age and cognitive impairment between men and women.
Several studies have reported that Alzheimer’s disease prevalence is higher in women than in men. [32] However, findings vary, as some authors indicate that this increased incidence is primarily observed in individuals aged 80–85 years or older, with a more pronounced increase in women as age advances.
These findings are complex, as some studies suggest that mild cognitive impairment (MCI) is more prevalent in men than in women, whereas dementia in advanced stages is more common in women. A study by Mutchie et al. (2022) analyzed cognitive status in older adults with hip fractures and determined that men were more likely to have cognitive impairment when assessed with neuropsychological tests rather than clinical evaluations. [33]
It is essential to consider factors influencing cognitive impairment, such as comorbidities (e.g., hypertension, diabetes, hypercholesterolemia, obesity, cardiovascular disease). [34] Psychiatric conditions also play a role. A study analyzing the impact of anxiety and depression on cognitive impairment found that men with anxiety had higher levels of cognitive impairment, while women with anxiety exhibited higher functional performance in daily activities. Conversely, men with depression showed lower cognitive impairment and better performance in daily activities [35]
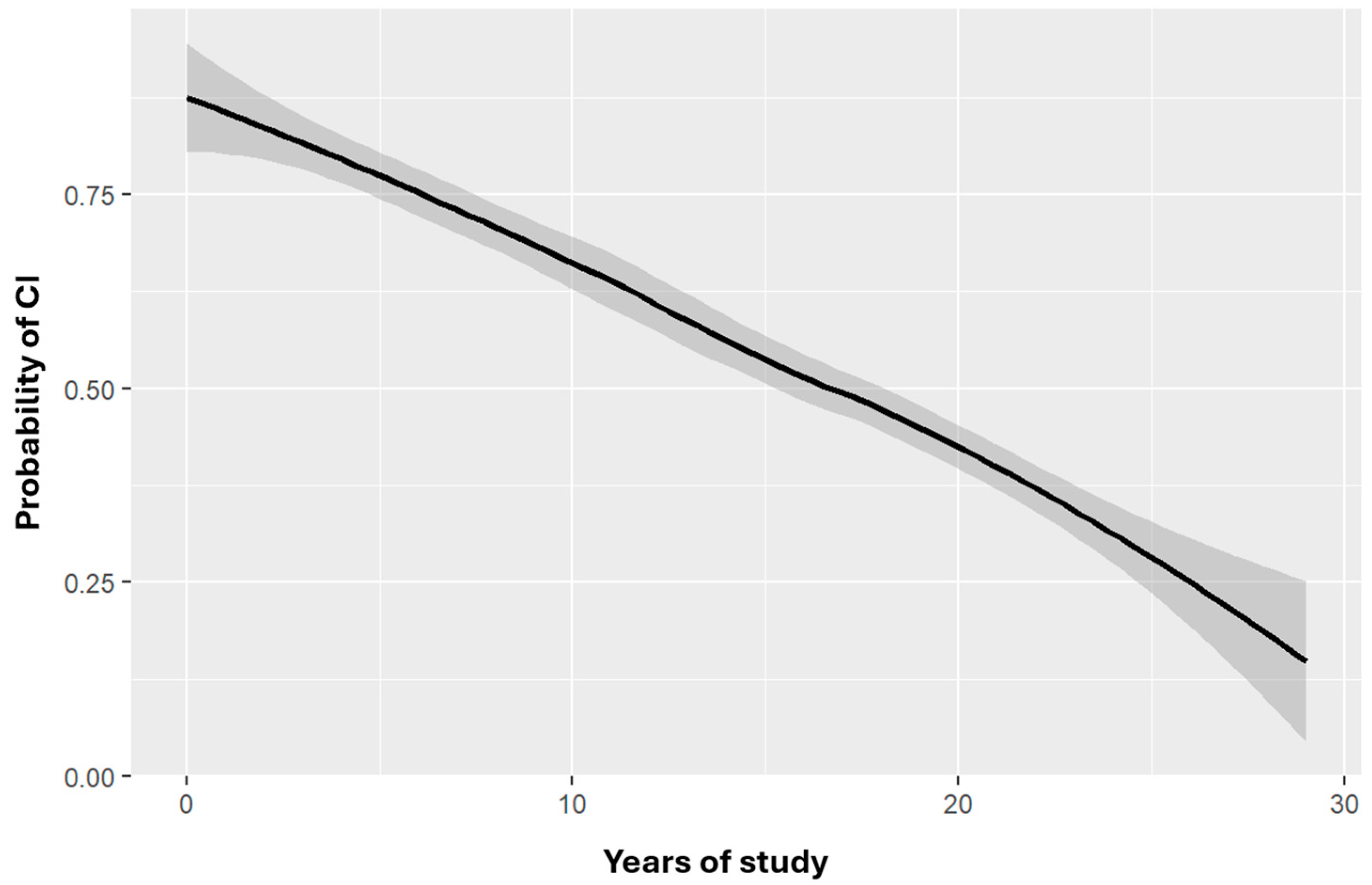
Regarding education, logistic regression analysis showed that for each additional year of education, the risk of cognitive impairment decreased. After adjusting for confounding variables, the statistical significance remained, indicating a negative relationship between years of education and cognitive impairment risk (OR = 0.8456, CI = 0.787–0.901, p < 0.0001) (Figure 2).
Figure 1.
Probability of cognitive impairment by years of study adjusted for age. Risk of cognitive impairment as a function of age-adjusted number of years of study. The shaded area indicates the CI for the function depicted. OR=0.8456 CI=0.787-0.901, p<0.0001***.
Figure 1.
Probability of cognitive impairment by years of study adjusted for age. Risk of cognitive impairment as a function of age-adjusted number of years of study. The shaded area indicates the CI for the function depicted. OR=0.8456 CI=0.787-0.901, p<0.0001***.
The concept of cognitive and brain reserve suggests that differences in individuals’ susceptibility to age-related brain changes or pathological processes are influenced by life experiences [28,36]. Studies have indicated that dementia incidence and prevalence decrease in individuals with larger brain volume, more neurons, and greater synaptic density. [37]
Animal studies support this concept, showing that enriched environments promote neuronal growth and new synaptic connections, enhancing brain plasticity. Years of education, occupation, and cognitive engagement are commonly used as indicators of cognitive reserve, with higher education levels being associated with greater cognitive resilience. [37]
Studies on cognitive reserve in Alzheimer’s patients indicate that higher cognitive reserve scores correlate with better cognitive performance. However, while a higher cognitive reserve may delay disease onset, it does not prevent disease progression. Some studies suggest that individuals with higher cognitive reserve may experience a more rapid cognitive decline once symptoms appear [38]
Although this study focused primarily on years of education, the observed association underscores its potential impact on cognitive decline. A more comprehensive analysis considering additional cognitive reserve factors is necessary.
Age and Cognitive Impairment
Logistic regression analysis found a significant association between age and cognitive impairment, indicating that older individuals have a higher risk of cognitive impairment (OR = 1.149, CI = 1.094–1.21, p = 3.43e-08, p < 0.001).
Figure 2.
Risk of cognitive impairment by age. Risk of CI as a function of number of years of age. Shaded area indicates CI for the depicted function OR=1.149, CI=1.094-1.21, p=3.43e-08 p<0.001**.
Figure 2.
Risk of cognitive impairment by age. Risk of CI as a function of number of years of age. Shaded area indicates CI for the depicted function OR=1.149, CI=1.094-1.21, p=3.43e-08 p<0.001**.
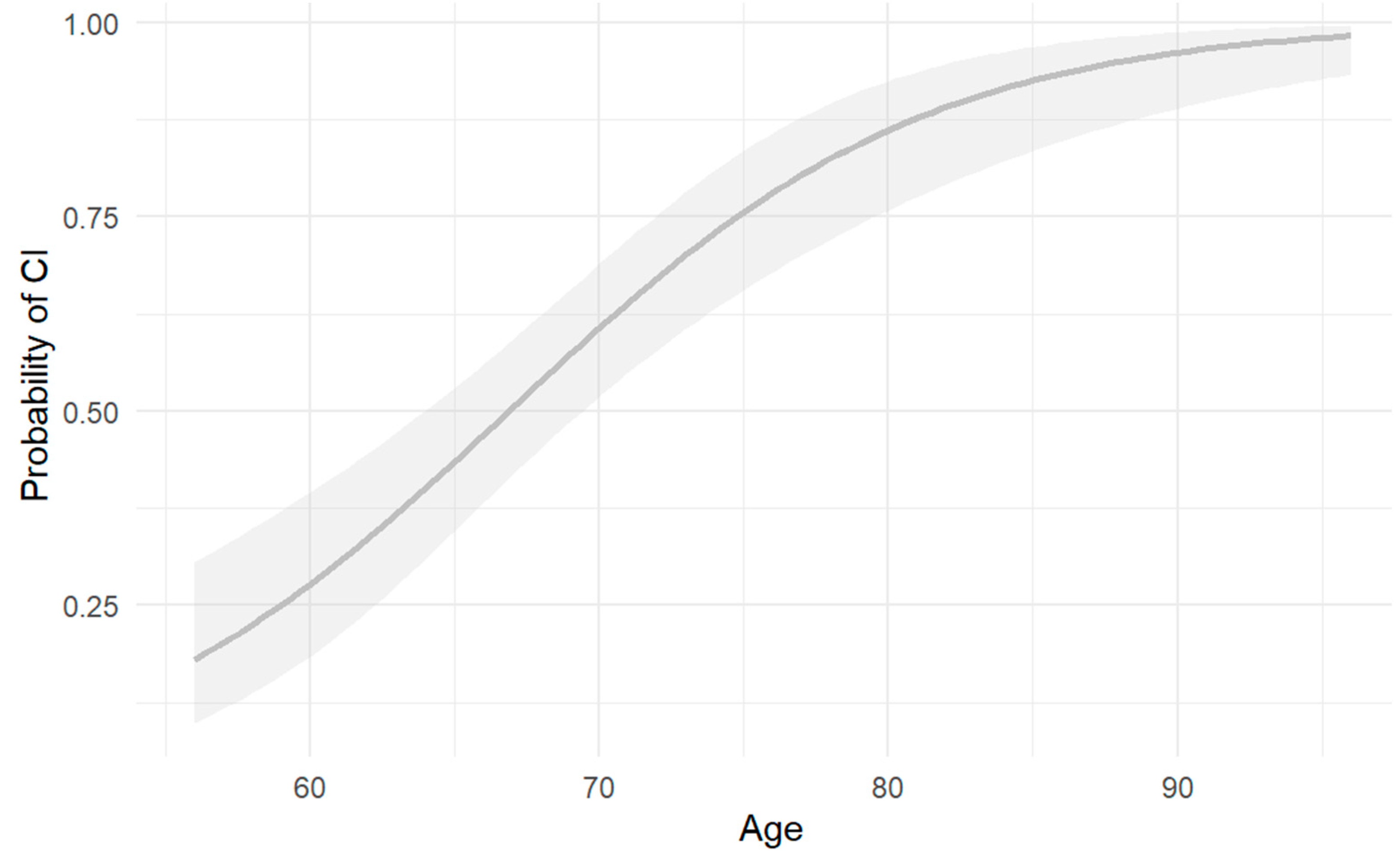
This finding is consistent with prior literature. A European meta-analysis reported that Alzheimer’s disease prevalence increases with age, showing 0.97% prevalence in individuals aged 65–75 years, 7.66% in those aged 75–84 years, and 22.53% in individuals over 85 years. [39]
Regarding genotype, 1.2% (n=2) presented APOEε2/ε3, 76.5% (n=130) APOEε3/ε3, 18.8% (n=32) APOEε3/ε4, 2.9% (n=5) APOEε4/ε4 and only 0.6% (n=1) APOEε2/ε4. Chi-square analysis revealed a statistically significant association between cognitive impairment and APOE genotype (χ² = 4.009, p = 0.045) but the frequencies shows that the absence of APOEε4 confer higher risk of cognitive impairment than presence; this suggestion is not influenced by age or years of study. (Table 2).
Several studies support that the APOEε3 allele is the most common, while the ε4 allele is associated with an increased risk of Alzheimer’s disease. [40] A meta-analysis of clinical studies reported that individuals carrying one copy of ε4 had a 2.6–3.2 times higher risk of developing Alzheimer’s, whereas those with two copies had a 12.9 times higher risk. [29,40]
The risk conferred by the APOE ε4 genotype has been shown in some publications to vary based on gender and age. A study by Molero et al. (2001) found that the presence of at least one ε4 allele was a significant age-stratified risk factor for Alzheimer’s disease only in women, suggesting that both age and gender modulate the APOE ε4-Alzheimer’s disease association. [41]
Beyond metabolic factors, the study population must also be considered, as the influence of the APOEε4 genotype appears to vary depending on ethnic background. For instance, in Asian populations, the ε4 allele and the APOEε2/ε4 genotype were associated with an increased risk of Parkinson’s disease (PD), whereas in Caucasian populations, the APOEε3/ε4 genotype appeared to have a protective effect against PD. [42] These contrasting results emphasize the importance of considering ethnic differences when studying APOE polymorphisms and their health implications.
A study by Campos et al. (2013) found that APOE ε4 frequencies were lower in Hispanic Mexican controls compared to non-Hispanic whites. [43] Similarly, research conducted in a Mexican clinical population found no significant association between the APOEε4 genotype and the presence of Alzheimer’s disease, despite observing higher ε4 frequencies relative to the ε3 allele. [44]
Telomere Length and Cognitive Impairment
Logistic regression analysis found a statistically significant association between shorter telomere length and higher cognitive impairment risk (OR = 0.789, CI = 0.6–1.01, p < 0.05) Numerous studies suggest that telomere shortening is linked to increased risk of cardiovascular disease, Alzheimer’s, and all-cause mortality. A systematic review concluded that individuals at higher risk for AD tend to have shorter telomeres, with Mendelian randomization studies supporting a causal relationship. [45]
Figure 3.
Risk of cognitive impairment as a function of telomeric length. Risk of cognitive impairment as a function of telomeric length number (p<=0.05 OR=0.789 CI=0.6-1.01).
Figure 3.
Risk of cognitive impairment as a function of telomeric length. Risk of cognitive impairment as a function of telomeric length number (p<=0.05 OR=0.789 CI=0.6-1.01).
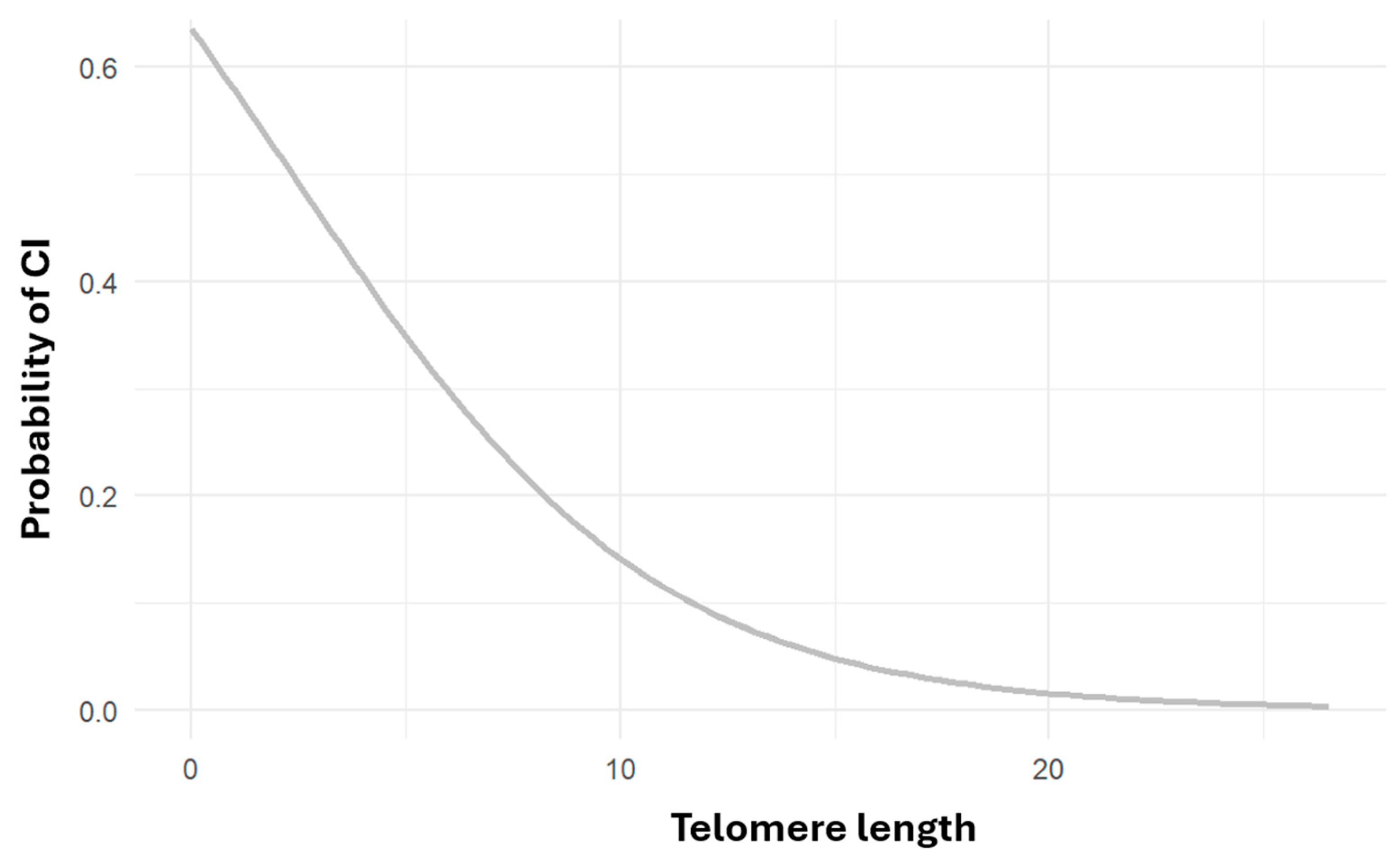
Crocco et al. (2023) found that individuals with Alzheimer’s disease had significantly shorter telomeres, independent of other risk factors such as age, sex, and APOEε4 genotype. [46] Other studies suggest a U-shaped relationship, where both very short and very long telomeres are associated with an increased risk of Alzheimer’s disease. [47]
Telomere Length and APOE genotype.
No statistically significant differences were found when analyzing telomere length relative to sex and years of education. Additionally, no significant association was found between APOEε4 and telomere length (Kruskal-Wallis χ² = 3.2385, df = 3, p = 0.3563). However, among APOEε4 carriers with cognitive impairment, the average telomere length was lower than in those NCI-SMC (Mann-Whitney U = 220.00, p = 0.116, Cohen’s d = 0.232).
Figure 4.
Telomeric length of APOEε4 carriers and cognitive impairment Telomere length. The telomere length of individuals with CI and NCI-SMC carrying the ε4 allele is shown. Bars represent the standard error of the medians of each group. Mann-Whitney 220.00, p=0.116, Cohen's d=0.232.
Figure 4.
Telomeric length of APOEε4 carriers and cognitive impairment Telomere length. The telomere length of individuals with CI and NCI-SMC carrying the ε4 allele is shown. Bars represent the standard error of the medians of each group. Mann-Whitney 220.00, p=0.116, Cohen's d=0.232.
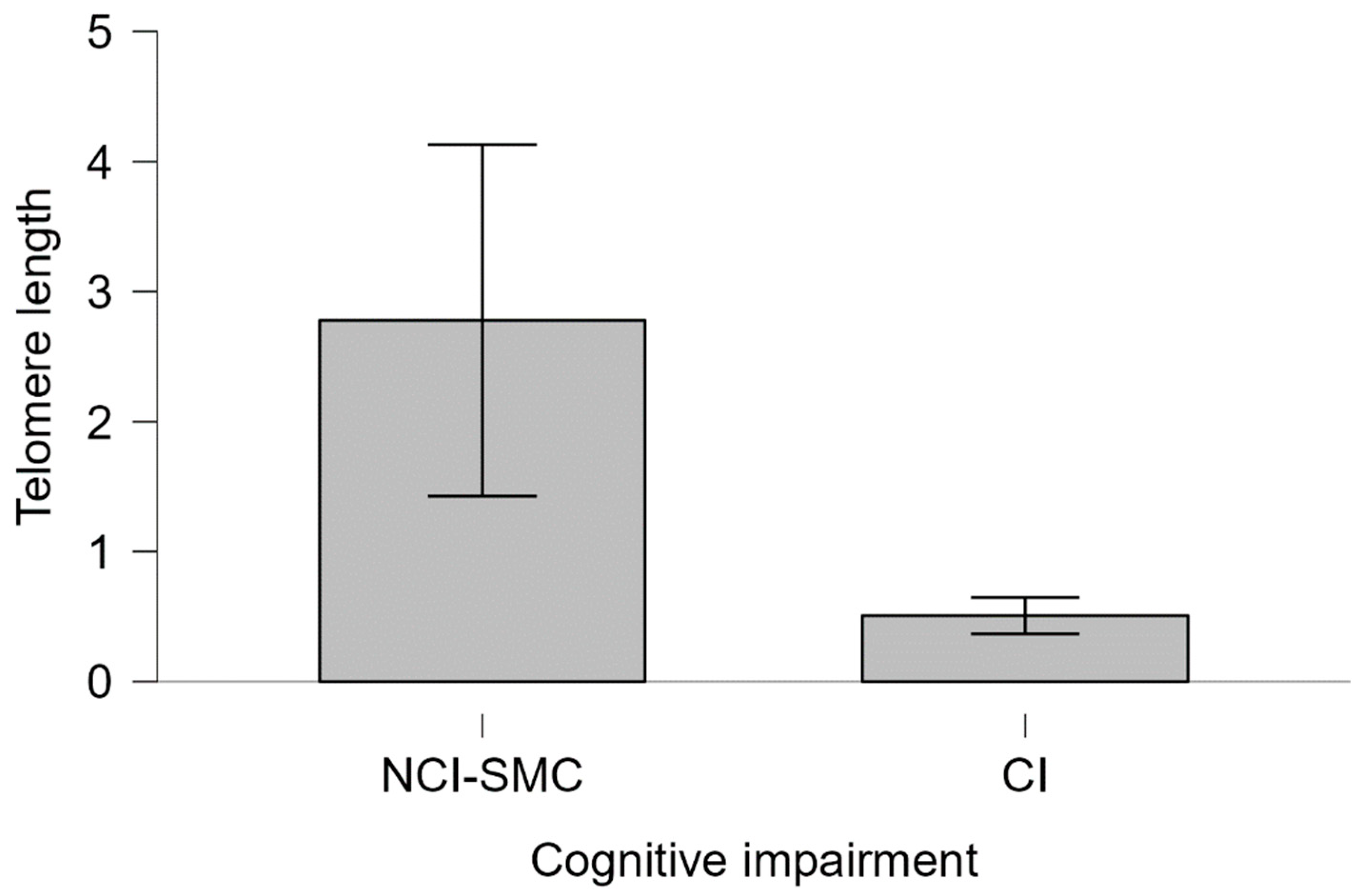
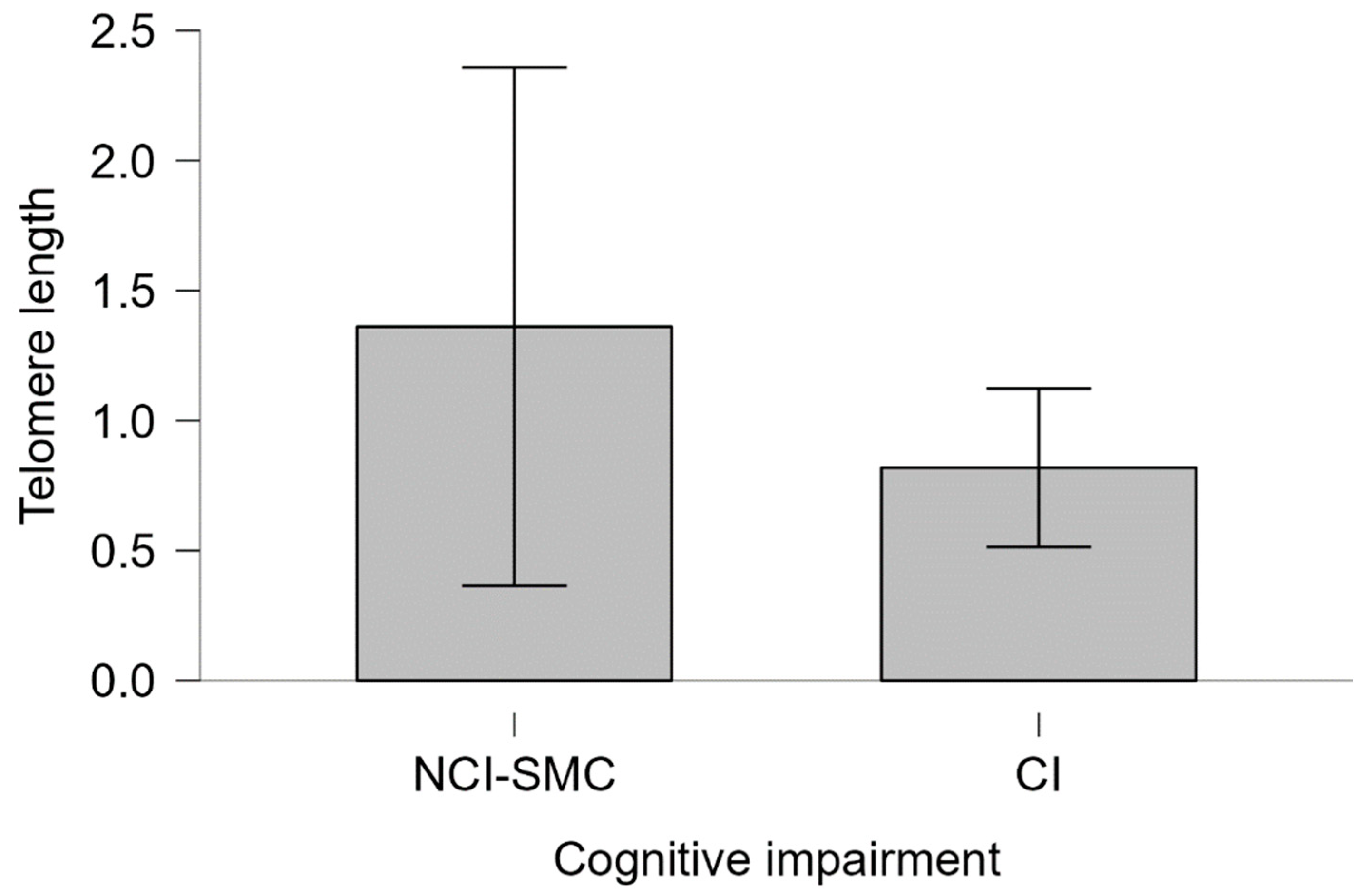
Figure 5.
Telomeric length of APOEε4 noncarriers and cognitive impairment Telomere length. The telomere length of individuals with CI and NCI-SMC not carrying the ε4 allele is plotted. Bars represent the standard error of the medians of each group. Mann-Whitney 2075.5 p=0.453, cohen's d=0.012.
Figure 5.
Telomeric length of APOEε4 noncarriers and cognitive impairment Telomere length. The telomere length of individuals with CI and NCI-SMC not carrying the ε4 allele is plotted. Bars represent the standard error of the medians of each group. Mann-Whitney 2075.5 p=0.453, cohen's d=0.012.
Studies have shown that individuals without APOEε4 tend to have longer telomeres, greater cortical thickness, and lower cerebrospinal fluid tau protein levels. APOE ε4 carriers have a higher risk of Alzheimer’s disease, potentially due to shorter telomere-induced cellular senescence and amyloid accumulation [27,47]
5. Conclusion
Shorter telomere length was associated with an increased risk of cognitive decline. Similarly, fewer years of education emerged as a risk factor for cognitive impairment and the strongest predictor in this study, reinforcing the role of cognitive reserve as a protective factor, independent of genotype, age, and sex. Furthermore, although the APOEε4 genotype was not directly associated with an increased risk of cognitive decline, it may contribute to telomere shortening.
Author Contributions
Conceptualization, P.M.O.; Methodology, P.M.O., J.S.; Software, P.M.O.; Validation, A.G.-M. and R.G.S.; Formal Analysis, P.M.O.; Investigation, P.M.O., S.L.R. and R.R.V.; Resources, H.N., R.G.S. and T.F; Data Curation, P.M.O.; Writing – Original Draft Preparation, P.O.M.; Writing – Review & Editing, A.G.-M. and T.F.; Visualization, P.M.O.; Supervision, A.G.-M.; Project Administration, H.N.; Funding Acquisition H.N. and T.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Instituto Nacional de Medicina Genómica (INMEGEN).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by Ethics Committee of Instituto Nacional de Medicina Genómica, official document number INMG/DI/115/2016 approved
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We would like to express our sincere gratitude to the Universidad Nacional Autónoma de México and the Maestría en Ciencias (Neurobiología) program, as this paper fulfills part of the requirements for obtaining a Master’s degree in Neurobiological Sciences. Additionally, we extend our gratitude to the Secretaria de Ciencias, Humanidades, Tecnología e Innovación (SECIHTI) for its financial support and for providing a graduate scholarship to PMO (CVU 1018978).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- American Psychiatric Association. (2013). Diagnostic and statistical manual of mental disorders (5th ed.). American Psychiatric Publishing.
- Sachdev, P. S., Blacker, D., Blazer, D. G., Ganguli, M., Jeste, D. V., Paulsen, J. S., & Petersen, R. C. (2014). Classifying neurocognitive disorders: The DSM-5 approach. Nature Reviews Neurology, 10(11), 634-642. [CrossRef]
- Castro-Chavira, S. A., Fernandez, T., Nicolini, H., Diaz-Cintra, S., & Prado-Alcala, R. A. (2015). Genetic markers in biological fluids for aging-related major neurocognitive disorder. Current Alzheimer research, 12(3), 200–209. [CrossRef]
- Choreño-Parra, J. A., De la Rosa-Arredondo, T., & Guadarrama-Ortíz, P. (2020). Abordaje diagnóstico del paciente con deterioro cognitivo en el primer nivel de atención. Medicina Interna de México, 36(6). [CrossRef]
- Instituto Nacional de Estadística y Geografía. (2023, 6 de julio). Encuesta Nacional sobre Salud y Envejecimiento en México (ENASEM) 2018. Encuesta Nacional sobre Salud y Envejecimiento en México (ENASEM) y Encuesta de Evaluación Cognitiva, 2021. INEGI. https://www.inegi.org.mx/app/saladeprensa/noticia.html?id=8294.
- Organización Mundial de la Salud. (2024, 1 de octubre). Envejecimiento y salud. https://www.who.int.
- Kim, J., Basak, J. M., & Holtzman, D. M. (2009). The role of apolipoprotein E in Alzheimer's disease. Neuron, 63(3), 287–303. [CrossRef]
- Martínez, S., Ochoa, B., Pérez, M. R., Torrico, F., García, I., & Garcia, C. C. (2022). Apolipoprotein E polymorphisms in adults over 60 years of age with mild cognitive impairment and Alzheimer’s disease in different Venezuelan populations. Polimorfismos del gen de la apolipoproteína E en adultos mayores de 60 años con disminución de la memoria cognitiva y enfermedad de Alzheimer en diferentes poblaciones venezolanas. Biomedica: revista del Instituto Nacional de Salud, 42(Sp. 1), 116–129. [CrossRef]
- Andrews, S. J., Fulton-Howard, B., & Goate, A. (2020). Interpretation of risk loci from genome-wide association studies of Alzheimer's disease. The Lancet. Neurology, 19(4), 326–335. [CrossRef]
- Lambert, J. C., Ibrahim-Verbaas, C. A., Harold, D., Naj, A. C., Sims, R., Bellenguez, C., DeStafano, A. L., Bis, J. C., Beecham, G. W., Grenier-Boley, B., Russo, G., Thorton-Wells, T. A., Jones, N., Smith, A. V., Chouraki, V., Thomas, C., Ikram, M. A., Zelenika, D., Vardarajan, B. N., Kamatani, Y., … Amouyel, P. (2013). Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nature genetics, 45(12), 1452–1458. [CrossRef]
- Namba, Y., Tomonaga, M., Kawasaki, H., Otomo, E., & Ikeda, K. (1991). Apolipoprotein E immunoreactivity in cerebral amyloid deposits and neurofibrillary tangles in Alzheimer's disease and kuru plaque amyloid in Creutzfeldt-Jakob disease. Brain research, 541(1), 163–166. [CrossRef]
- Strittmatter WJ, Saunders AM, Schmechel D, Pericak-Vance M, Enghild J, Salvesen GS, Roses AD. Apolipoprotein E: high-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc Natl Acad Sci U S A. 1993 Mar 1;90(5):1977-81. doi: 10.1073/pnas.90.5.1977. PMID: 8446617; PMCID: PMC46003. Ticinesi A, Tana C, Nouvenne A, Prati B, Lauretani F, Meschi T. Gut microbiota, cognitive frailty and dementia in older individuals: a systematic review. Clin Interv Aging. 2018 Aug 29;13:1497-1511. PMID: 30214170; PMCID: PMC6120508. [CrossRef]
- Pimenova AA, Raj T, Goate AM. Untangling Genetic Risk for Alzheimer's Disease. Biol Psychiatry. 2018 Feb 15;83(4):300-310. Epub 2017 May 22. PMID: 28666525; PMCID: PMC5699970. [CrossRef]
- Coon, K. D., Myers, A. J., Craig, D. W., Webster, J. A., Pearson, J. V., Lince, D. H., Zismann, V. L., Beach, T. G., Leung, D., Bryden, L., Halperin, R. F., Marlowe, L., Kaleem, M., Walker, D. G., Ravid, R., Heward, C. B., Rogers, J., Papassotiropoulos, A., Reiman, E. M., Hardy, J., … Stephan, D. A. (2007). A high-density whole-genome association study reveals that APOE is the major susceptibility gene for sporadic late-onset Alzheimer's disease. The Journal of clinical psychiatry, 68(4), 613–618. [CrossRef]
- Raghavan, N., & Tosto, G. (2017). Genetics of Alzheimer's Disease: the Importance of Polygenic and Epistatic Components. Current neurology and neuroscience reports, 17(10), 78. [CrossRef]
- Ska, B., & Joanette, Y. (2006). Vieillissement normal et cognition [Normal aging and cognition]. Medecine sciences: M/S, 22(3), 284–287. [CrossRef]
- Dziechciaż, M., & Filip, R. (2014). Biological psychological and social determinants of old age: bio-psycho-social aspects of human aging. Annals of agricultural and environmental medicine : AAEM, 21(4), 835–838. [CrossRef]
- 18. Anisimova AS, Alexandrov AI, Makarova NE, Gladyshev VN, Dmitriev SE. Protein synthesis and quality control in aging. Aging (Albany NY). 2018 Dec 18;10(12):4269-4288. PMID: 30562164; PMCID: PMC6326689. [CrossRef]
- da Silva, P. F. L., & Schumacher, B. (2021). Principles of the Molecular and Cellular Mechanisms of Aging. The Journal of investigative dermatology, 141(4S), 951–960. [CrossRef]
- López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2023). Hallmarks of aging: An expanding universe. Cell, 186(2), 243–278. [CrossRef]
- Hernández Fernández, R. A. (2000). Telómeros y telomerasas. Revista Cubana de Investigaciones Biomédicas, 18(2), 121-129. Recuperado de http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-03001999000200009&lng=es&tlng=es.
- Scarabino, D., Broggio, E., Gambina, G., & Corbo, R. M. (2017). Leukocyte telomere length in mild cognitive impairment and Alzheimer's disease patients. Experimental gerontology, 98, 143–147. [CrossRef]
- Gampawar, P., Schmidt, R., & Schmidt, H. (2022). Telomere length and brain aging: A systematic review and meta-analysis. Ageing Research Reviews, 80, 101679. [CrossRef]
- Suchy-Dicey, A. M., Muller, C. J., Madhyastha, T. M., Shibata, D., Cole, S. A., Zhao, J., Longstreth, W. T., Jr, & Buchwald, D. (2018). Telomere Length and Magnetic Resonance Imaging Findings of Vascular Brain Injury and Central Brain Atrophy: The Strong Heart Study. American journal of epidemiology, 187(6), 1231–1239. [CrossRef]
- King, K. S., Kozlitina, J., Rosenberg, R. N., Peshock, R. M., McColl, R. W., & Garcia, C. K. (2014). Effect of leukocyte telomere length on total and regional brain volumes in a large population-based cohort. JAMA neurology, 71(10), 1247–1254. [CrossRef]
- Rodríguez-Fernández, B., Vilor-Tejedor, N., Arenaza-Urquijo, E. M., Sánchez-Benavides, G., Suárez-Calvet, M., Operto, G., Minguillón, C., Fauria, K., Kollmorgen, G., Suridjan, I., de Moura, M. C., Piñeyro, D., Esteller, M., Blennow, K., Zetterberg, H., De Vivo, I., Molinuevo, J. L., Navarro, A., Gispert, J. D., Sala-Vila, A., & Crous-Bou, M.; ALFA study. (2022). Genetically predicted telomere length and Alzheimer's disease endophenotypes: A Mendelian randomization study. Translational Psychiatry, 12(1), 9. [CrossRef]
- Hackenhaar, F. S., Josefsson, M., Adolfsson, A. N., Landfors, M., Kauppi, K., Hultdin, M., Adolfsson, R., Degerman, S., & Pudas, S. (2021). Short leukocyte telomeres predict 25-year Alzheimer's disease incidence in non-APOE ε4-carriers. Alzheimer's research & therapy, 13(1), 130. [CrossRef]
- Wagner, K. H., Cameron-Smith, D., Wessner, B., & Franzke, B. (2016). Biomarkers of Aging: From Function to Molecular Biology. Nutrients, 8(6), 338. [CrossRef]
- Liu, C. C., Liu, C. C., Kanekiyo, T., Xu, H., & Bu, G. (2013). Apolipoprotein E and Alzheimer disease: risk, mechanisms and therapy. Nature reviews. Neurology, 9(2), 106–118. [CrossRef]
- Ciesielska, N., Sokołowski, R., Mazur, E., Podhorecka, M., Polak-Szabela, A., & Kędziora-Kornatowska, K. (2016). Is the Montreal Cognitive Assessment (MoCA) test better suited than the Mini-Mental State Examination (MMSE) in mild cognitive impairment (MCI) detection among people aged over 60? Meta-analysis. Czy test Montreal Cognitive Assessment (MoCA) może być skuteczniejszy od powszechnie stosowanego Mini-Mental State Examination (MMSE) w wykrywaniu łagodnych zaburzeń funkcji poznawczych u osób po 60. roku życia? Metaanaliza. Psychiatria polska, 50(5), 1039–1052. [CrossRef]
- Delgado, C., Araneda, A., & Behrens, M. I. (2019). Validation of the Spanish-language version of the Montreal Cognitive Assessment test in adults older than 60 years. Validación del instrumento Montreal Cognitive Assessment en español en adultos mayores de 60 años. Neurologia, 34(6), 376–385. [CrossRef]
- Au, B., Dale-McGrath, S., & Tierney, M. C. (2017). Sex differences in the prevalence and incidence of mild cognitive impairment: A meta-analysis. Ageing research reviews, 35, 176–199. [CrossRef]
- Mutchie, H. L., Albrecht, J. S., Orwig, D. L., Huang, Y., Boscardin, W. J., Hochberg, M. C., Magaziner, J. S., & Gruber-Baldini, A. L. (2022). Differential misclassification of cognitive impairment by sex among hip fracture patients. Journal of the American Geriatrics Society, 70(3), 838–845. https://doi.org/. [CrossRef]
- Katabathula, S., Davis, P. B., Xu, R., & Alzheimer’s Disease Neuroimaging Initiative (2023). Sex-Specific Heterogeneity of Mild Cognitive Impairment Identified Based on Multi-Modal Data Analysis. Journal of Alzheimer's disease : JAD, 91(1), 233–243. [CrossRef]
- Calatayud, E., Marcén-Román, Y., Rodríguez-Roca, B., Salavera, C., Gasch-Gallen, A., & Gómez-Soria, I. (2023). Sex differences on anxiety and depression in older adults and their relationship with cognitive impairment. Semergen, 49(4), 101923. [CrossRef]
- Stern, Y., Arenaza-Urquijo, E. M., Bartrés-Faz, D., Belleville, S., Cantilon, M., Chetelat, G., Ewers, M., Franzmeier, N., Kempermann, G., Kremen, W. S., Okonkwo, O., Scarmeas, N., Soldan, A., Udeh-Momoh, C., Valenzuela, M., Vemuri, P., Vuoksimaa, E., & the Reserve, Resilience and Protective Factors PIA Empirical Definitions and Conceptual Frameworks Workgroup (2020). Whitepaper: Defining and investigating cognitive reserve, brain reserve, and brain maintenance. Alzheimer's & dementia : the journal of the Alzheimer's Association, 16(9), 1305–1311. [CrossRef]
- Nelson, M. E., Jester, D. J., Petkus, A. J., & Andel, R. (2021). Cognitive Reserve, Alzheimer's Neuropathology, and Risk of Dementia: A Systematic Review and Meta-Analysis. Neuropsychology review, 31(2), 233–250. [CrossRef]
- Soldan, A., Pettigrew, C., Cai, Q., Wang, J., Wang, M. C., Moghekar, A., Miller, M. I., Albert, M., & BIOCARD Research Team (2017). Cognitive reserve and long-term change in cognition in aging and preclinical Alzheimer's disease. Neurobiology of aging, 60, 164–172. [CrossRef]
- Niu, H., Álvarez-Álvarez, I., Guillén-Grima, F., & Aguinaga-Ontoso, I. (2017). Prevalence and incidence of Alzheimer's disease in Europe: A meta-analysis. Prevalencia e incidencia de la enfermedad de Alzheimer en Europa: metaanálisis. Neurologia (Barcelona, Spain), 32(8), 523–532. [CrossRef]
- Baum, L., Chen, L., Ng, H. K., & Pang, C. P. (2000). Apolipoprotein E isoforms in Alzheimer's disease pathology and etiology. Microscopy research and technique, 50(4), 278–281. https://doi.org/10.1002/1097-0029(20000815)50:4<278::AID-JEMT5>3.0.CO;2-T.
- Molero, A. E., Pino-Ramírez, G., & Maestre, G. E. (2001). Modulación por edad y género del riesgo de enfermedad de Alzheimer y demencia vascular asociada al alelo de la apolipoproteína E-ε4 en latinoamericanos: Hallazgos del Estudio de Envejecimiento de Maracaibo. Cartas de Neurociencia, 307(1), 5.
- Li, Y., Li, J., Li, J., Li, X., & Li, Y. (2018). Association between apolipoprotein E polymorphisms and Parkinson's disease risk: A meta-analysis. Frontiers in Aging Neuroscience, 10, 1-9. [CrossRef]
- Campos, M., Edland, S. D., & Peavy, G. M. (2013). Exploratory study of apolipoprotein E ε4 genotype and risk of Alzheimer's disease in Mexican Hispanics. Journal of the American Geriatrics Society, 61(6), 1038–1040. [CrossRef]
- Villalpando-Berumen, J. M., Mejía-Arango, S., Aguilar-Salinas, C. A., Ordoñez-Sánchez, M. L., & Gutiérrez-Robledo, L. M. (2008). Apolipoprotein E epsilon4, Alzheimer's disease, and cognitive performance in elderly Mexican Mestizos. Journal of the American Geriatrics Society, 56(4), 677–682. [CrossRef]
- Piyush, G., Schmidt, R., & Schmidt, H. (2020). Leukocyte telomere length is related to brain parenchymal fraction and attention/speed in the elderly: Results of the Austrian Stroke Prevention Study. Frontiers in Psychiatry, 11, 1-9. [CrossRef]
- Crocco, P., De Rango, F., Dato, S., La Grotta, R., Maletta, R., Bruni, A. C., Passarino, G., & Rose, G. (2023). The shortening of leukocyte telomere length contributes to Alzheimer's disease: Further evidence from late-onset familial and sporadic cases. Biology (Basel), 12(10), 1286. [CrossRef]
- Fani, L., Hilal, S., Sedaghat, S., Broer, L., Licher, S., Arp, P. P., van Meurs, J. B. J., Ikram, M. K., & Ikram, M. A. (2020). Telomere length and the risk of Alzheimer's disease: The Rotterdam Study. Journal of Alzheimer's Disease, 73(2), 707-714. [CrossRef]
Table 2.
Genotype and cognitive impairment.
| APOE ε4 | |||
|---|---|---|---|
| With | Without | p-value | |
| Cognitive impairment | 17 (10) | 83 (48) | p<0.05* |
| Age b | 74.118 (+/-8.49) | 74.195 (+/-9.29) | |
| Years of study b | 7.941 (+/-5.55) | 9.964 (+/-6.01) | |
| NCI-Subjetive memory complaint | 21 (12.35) | 49 (28.8) | |
| Age b | 63.238 (+/-5.04) | 66.26 (+/-6.56) | |
| Años de estudio b | 18.047(+/-5.8) | 18.204 (+/-5.53) | |
| Total | 38 (22.35) | 132 (77.65) | |
| Age | 68.105 (+/-8.67) | 71.189 (+/-9.3) | p=0.966 |
| Years of study | 13.52 (+/-7.58) | 13.023 (+/-7.05) | p=0.347 |
a Frequency is shown and in parentheses the percentage of individuals calculated with the total n. b Mean is shown and in parentheses the standard deviation for each group. The p values were calculated by comparing the variables cognitive impairment, age and years of study between individuals carrying and not carrying the e4 allele.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



