
Submitted:
28 February 2025
Posted:
03 March 2025
You are already at the latest version
Abstract
Despite the emerging role of transcriptional regulators in schizophrenia as key mo lecular effectors re-sponsible for the dysregulation of multiples biological processes, limited information is available in brain are-as that control higher cognitive functions as the cerebellum. To identify transcription factors that could control a wide panel of altered proteins in the cerebellar cortex in schizophrenia, we analyzed a dataset obtained us-ing one-shot liquid chromatography-tandem mass spectrometry on postmortem human cerebellar cortex in chronic schizophrenia (PXD024937 identifier in ProteomeXchange repository). Our analysis revealed a panel of 11 enriched transcription factors (SP1, KLF7, SP4, EGR1, HNF4A, CTCF, GABPA, NRF1, NFYA, YY1) and MEF2A, that could be controlling 250 altered proteins. The top 3 significantly enriched transcription factors were SP1, YY1, and EGR1 and the transcription factors with the largest number of targets were SP1, KLF7 and SP4 which belong to the Krüppel superfamily. An enrichment in vesicle-mediated transport was found for SP1, KLF7, EGR1, HNF4A, CTCF and MEF2A targets while pathways related to signaling, inflamma-tion/immune response, apoptosis, and energy were found for SP1 and KLF7 targets. EGR1 targets were en-riched in RNA processing and, GABPA and YY1 targets were mainly involved in organelle organization and assembly. This study provides a reduced panel of transcriptional regulators that could be impacting on multi-ple pathways through the control of a number of targets in the cerebellum in chronic schizophrenia. These findings suggest that this panel of transcription factors could be key targets for pharmacological interventions in schizophrenia.
Keywords:
schizophrenia
; cerebellum
; transcription factors
1. Introduction
Schizophrenia (SZ) is a polygenetic psychiatric disorder with heritability up to 80% [1]. The mechanisms underlying this disorder are complex and are not completely understood. However, hypotheses such as neurodevelopmental and cognitive dysmetria have been proposed as a framework for the understanding of this psychiatric disorder. The neurodevelopment hypothesis argues that the genetic predisposition and possible alterations during intrauterine life could lead to altered development of the central nervous system (CNS) which could manifest during the adolescence [2,3,4]. In the last decades, the cerebellum has been suggested to be implicated in this pathophysiology through the cognitive dysmetria hypothesis [5]. This hypothesis states that dysfunction of the cortico-thalamo-cerebellar circuit (CCTC) contributes to symptom emergence in SZ [6,7,8]. In the context of CCTC circuit, the cerebellum innervates through of the thalamus to prefrontal and parietal cortex, areas involved in cognitive functions and altered in SZ [9]. Although the cerebellum is highly organised tissue, this consists of a homogeneous neuronal population with granular cells making up approximately 90% of the population [10]. This feature makes the cerebellum a useful model for proteomic study and finds molecular alterations that could alter internal circuits.
Transcription factors (TFs) control gene networks that are required for the processes of regionalisation and neuronal precursors migrations during cerebellar development[11]. In the context of SZ, is known that several signaling pathways are dysregulated, therefore, is necessary to identify the transcriptional programs that regulate the differentially expressed genes involved in the altered pathways. In this context, studies have associated altered expression of several TFs such as TCF4 with a high risk of SZ [12]. This relationship could be likely explained by the fact that TCF4 during the development is essential for neuronal migration during cortex cerebellar development [13]. Also, it is known that dendritic organisation could be affected in SZ. Altered protein expression in postmortem cerebellum of some members SP/KLF superfamily as Specificity Proteins (SPs) have been related to the altered dendritic organization and neuronal growth in SZ [14,15] and Krüppel like factor (KLF) at neuronal morphogenesis [16,17]. In addition, transcriptional dysregulation of NKX2-1 and EGR1 has been related with altered GABAergic neurotransmission in SZ [18] which could lead to altered synaptic proccess and the poor cognitive function described in SZ. Thus, the accumulative effect of altered expression of these TFs could cause the dysregulation of the transcriptional networks which could compromise the neuronal structure, synaptic efficiency and lead to the dysfunction of signaling pathways in SZ. However, the identification of trancriptional factors that could be modulating large networks of altered genes in the cerebellum in SZ and how these transcription factors impact on specific pathways and biological functions has not been deeply study so far.
Our aim was to identify possible transcriptional regulators in the cerebellum which could be responsible of altered levels of diferent proteins. In addition, we further investigate the biological processes and signaling pathways controlled by transcription factor-dependent altered programs.
2. Results
We analyzed a previous dataset of 250 altered proteins in the human cerebellum cortex in Chronic SZ obtained from the proteomic study using one-shot liquid chromatography-tandem mass [19]. The dataset of the proteomic profile of cerebellum has been deposited in ProteomeXchange repository with PXD024937 identifier.
To carry out the study we performed an experimental design showed in Figure 1, where the 250 altered proteins were used to search for transcription factors that could be controlling them. To find the biological processes and pathways that could be regulating these transcription factors, we performed gene ontology analyses with the protein groups regulated by each transcription factor.
2.1. Putative Transcriptional Programs Responsible of Changes in the Proteomic Profile in the Cerebellum
To investigate the transcriptional program that could be controlling the 250 altered proteins in SZ, we performed an enrichment analysis of TFs. Our enrichment analysis for the transcription factor targets showed 40 significant TFs (p-value<0.05) (Supplementary dataset 1). We generated a list of 11 potential TFs that could be controlling the 250 altered proteins according to the following criteria: the TFs would regulate more than 15% of the target proteins (Figure 2). These TFs were: SP1, KLF7, SP4, EGR1, HNF4A, CTCF, GABPA, NRF1, NFYA, YY1, and MEF2A. This analysis revealed that the top 3 most significant TFs were SP1, EGR1 and YY1, with 125, 60 and 37 targets, respectively (Supplementary dataset 2). Furthermore, the analysis showed that the TFs with the largest percentage of target proteins were SP1 (125 targets), KLF7 (76 targets) and SP4 (66 targets), all of which belong to the Krüppel superfamily.
2.2. Altered Biological Processes Controlled by Transcriptional Programs in the Cerebellum in Chronic Schizophrenia
Our gene ontology analysis in target genes revealed 10 out of 11 TFs have enriched biological processes (FDR<0.05). The most significant biological processes were regulated by SP1, KLF7, EGR1 and GABPA (Figure 3). In this analysis, SP1 and KLF7 target proteins were enriched in functions related to cytoskeleton organization development, cellular and organelle organization and inflammation/immune response. KLF7 target proteins showed significantly enriched processes related to neutrophil-mediated immunity and granulocyte activation. EGR1 targets were enriched in cytoskeleton organization development and RNA processing such as mRNA metabolism and RNA catabolic processes. GABPA and YY1 targets were mainly involved in cellular and organelle organization and assembly. The biological processes involved in synaptic functions were enriched for target proteins of MEF2A, SP1 and KLF7. MEF2A and SP1 target proteins were enriched in regulation of vesicle-mediated transport, while KLF7 together to those of SP1 were also enriched in the regulation of intracellular transport. SP4 target proteins were enriched in some biological processes associated to cellular and organelle organization and assembly functions mainly. In contrast, NRA2A was only implicated in assembly functions.
2.3. Altered Pathway Analysis Controlled by Transcriptional Programs in the Cerebellum in Chronic Schizophrenia
Our results revealed pathways significantly enriched (FDR<0.05) in altered targets of 5 TFs: SP1, KLF7, EGR1, HNF4A and CTCF (Figure 4). The enriched pathways were mainly detected in targets regulated by Krüppel superfamily TFs such as SP1 and KLF7, with 28 and 13 pathways respectively. SP1 targets showed enrichment in all pathways. The vesicle-mediated transport pathway was under the control of targets of 5 TFs. EGR1 altered targets were enriched in pathways involved in transport and signaling. HNF4A altered targets were only enriched in pathways related to vesicle-mediated transport and membrane trafficking pathways. CTCF targets were enriched in pathways involved in transport and processes associated with the Golgi complex. Moreover, SP1 and KLF7 altered targets showed an enrichment in pathways related to signaling, inflammation/immune response, apoptosis and energy (mitochondrial processes and glucose transport mediated by translocation of SLC2A4 (GLUT4) to the plasma membrane).
3. Discussion
Our study identified 11 potential TFs enriched in the cerebellum in chronic SZ that could control the expression of the 250 significantly altered proteins, contributing to dysregulation of several biological processes and pathways in SZ. Several studies have implicated 10 out of these 11 TFs in SZ: SP1 [20,21,22], KLF7 and SP4 [23,24,25,26,27,28], EGR1 [29,30,31], HNF4A [32], CTCF [33,34,35], GABPA [33], NRF1 [36,37], NFYA [38], YY1 [34], and MEF2A [39].
3.1. Transcription Factor Dependent-Enriched Biological Processes
3.1.1. Cytoskeleton and Organelle Organization
The enrichment analysis showed that SP1, KLF7, and SP4 which belong to the SP/KLF superfamily, had the greatest number of target genes. The SP/KLF superfamily is characterized by its binding to GC boxes in promoter regions with almost identical affinity due to the high homology in their DNA-binding domains [40] Our results identified biological processes such as cytoskeleton organization/development, cellular/organelle organization and pathways related to signaling as the most enriched categories for SP1, SP4 and KLF7. The cytoskeleton mediates a large variety of cellular functions, including supporting cellular morphology and cellular activities such as vesicle trafficking, neuronal migration, and neurite outgrowth [41]. SP1 in astrocytes has been implicated in neurite outgrowth and synaptogenesis [42], while SP4 has been associated with dendritic arborization in cerebellum [14,43]. KLF7 has been implicated in enhancing axon growth [44,45], formation of dendritic branching in the hippocampus and altered axon projection in several brain regions [46]. Moreover, KLF7 has been reported to be involved in the maturation of granular neurons in the cerebellum during early postnatal development [46]. In addition, studies performed in the postmortem cerebellum have shown altered protein levels of SP1 and SP4 linked to negative symptoms in chronic SZ. Altered levels of both transcription factors were also found in the hippocampus in these subjects [15] and in the prefrontal cortex, only SP1 protein levels were reduced in these subjects [24] suggesting a region-specific dysregulation of these TFs in SZ. These reports together with our results point to a possible dysregulation of KLF7 in SZ that leads to the alteration of the maturation of granular cells and axon growth, while altered expression of SP1 and SP4 could be related to altered formation of neurites and the dendritic arborization patterns. All these processes could eventually lead to altered cell-cell communication in the inner cerebellar circuits and the connection of the cerebellum with other brain regions.
3.1.2. mRNA Processing and Splicing
Our analysis reports that a protein set involved in biological processes related to mRNA processing could be under the transcriptional control of SP1, EGR1, and KLF7 with SP1 target genes being the only ones enriched in splicing. It has recently been shown that alternative splicing could play a role in SZ [47,48]. Many of the archetypal genes associated with SZ, for example, DISC1 [49] and ERBB4 [50], are aberrantly spliced transcripts. However, the molecular mechanism underpinning this aberrant splicing is unknown. A study in mice showed that Sp1 enhances the transcription of the splicing factor Slu7, while depletion of Sp1 repressed Slu7 expression, thereby affecting alternative splicing processes [51]. Thus, further studies will be needed to test the possibility that SP1-dependent altered splicing could be mediating the generation of aberrant alternative splicing forms in key genes in SZ physiopathology, such as DISC1 and ERBB4.
3.1.3. Synaptic Function
In our study, the most significant enriched process from synaptic function was vesicle transport linked to MEF2A target genes. MEF2A is a transcription factor expressed in adults and implicated in neuronal development, the formation of postsynaptic granule neuron dendritic claws [52,53]. Moreover, the study of Crisafulli et al., found that at least seven single-nucleotide polymorphism in MEF2A could be related to SZ [54,55]. Also, MEF2A has been related as a negative regulator in the AMPA receptors expression, which participates in the memory processes [56] suggesting that this transcription factor could be involved in the cognitive decline in SZ. Therefore, a dysregulation of MEF2A not only could be responsible of altered synaptic morphology in cerebellar granule neurons, but also in the neurotransmitter vesicle transport to the active presynaptic zone in this neurons in SZ.
3.2. Transcription Factor Dependent-Enriched Pathways
3.2.1. Transport and Golgi Complex
Pathways related to transport and the Golgi complex, such as vesicle-mediated and membrane trafficking, were the pathways found to be most enriched for the target proteins of SP1, EGR1, HNF4A, and CTCF. All these pathways are involved in the functioning of the Golgi apparatus. Protein transport from the endoplasmic reticulum to the Golgi complex requires transport vesicles [57]. Recently, it has been proposed that the Golgi phosphoprotein 3 (GOLPH3), which participates in protein trafficking, receptor recycling, and glycosylation in the Golgi, can regulate the transcription of the proinflammatory cytokines such as TNF-α, this regulation could be mediated by the EGR1/ERK pathway [58]. This evidence raises the question of whether EGR1 could be implicated in the inflammatory processes in SZ. Moreover, all the TFs involved in the transport and the Golgi complex such as SP1, EGR1 [59], HNF4A [60] and CTCF [33] have been previously reported to be altered in SZ. However, the role of these TFs in anterograde transport or functions associated with the Golgi apparatus in the context of SZ is unknown.
3.2.2. Immune Response and Inflammatory Processes
Although the neurodevelopmental hypothesis is well accepted, the inflammation, dysregulation of the immune mechanisms and degenerative views have also been suggested as hypotheses which have generated a large debate in the field [61,62,63,64,65,66,67,68,69]. The imbalance in the levels of proinflammatory and anti-inflammatory cytokines has been related to symptoms and cognitive decline in SZ [70,71]. In our study, biological processes and pathways related to the immune response were found to be enriched linked to specific transcriptional programs. The transcriptional control of the targets involved in inflammatory events could be regulated by some members of the Krüppel-like factor family such as SP1 and KLF7. KLF7 has been related to increases in the levels of IL-6, which play a role in both inflammatory and anti-inflammatory responses [72]. KLF7 could promote the increase of IL-6 through PKCζ/NF-κB [73] and TLR4/NF-κB/IL-6 signaling [74]. In addition, studies have reported high levels of IL-6 in SZ subjects [75,76]. A study reported that KLF7 can induce macrophage activation [79,80]. Moreover, several members of the Krüppel-like factor family, such as KLF2, KLF4 and KLF6, have been reported to be involved in the immune system and inflammation [77,78,79] which is in line with our results. Thus, taken together, these findings suggest that KLF7 could have a relevant role in inflammatory processes in SZ.
Another member of Krüppel-like factor family is SP1. SP1 has been associated with the activation of interleukin 21 receptors in T cells [80,81], which mediate the activation of several cell types involved in the immune response [82]. Furthermore, SP1 has been implicated in interleukin 12 (IL-12) expression [83]. IL-12 induces the differentiation of T-helper 1 cells [84] during the adaptive immune response. In this sense altered IL-12 levels have been reported in the plasma of SZ subjects [85,86]. Also, SP1 induces the activation of macrophage inflammatory protein-2 (MIP-2), which is involved in recruiting neutrophils to inflammatory regions [87]. In addition, SP1 has also been implicated in the crosstalk between the interferon regulatory factors and NFκB pathways, thereby contributing to the TLR-dependent antiviral response [88]. In SZ, it has been reported that SP1 could interact with the TLR4-MyD88-IκBα-NFκB pathway, which mediates its interaction with NFκB [89]. Thus, SP1 could be an activator of the immune response. The dysregulation of IL-12 expression due to the altered function of SP1 could lead to dysfunctional differentiation of T-helper cells and an altered adaptive immune response in SZ. Thus, our study suggests the possible participation of SP1 in inflammatory processes in SZ subjects.
3.2.3. Apoptotic Events
Disseminated apoptotic events in the CNS throughout the developmental period and later phases impact on the emergence of SZ and the progression of the disease [90,91]. These apoptotic processes support the neu-rodegenerative hypothesis proposed for SZ [92,93]. However, the transcriptional program involved in this process is unknown. Our analysis revealed that SP1 and KLF7 could participate in mitochondrial apoptosis. While some studies have demonstrated that overexpression of SP1 could induce apoptosis, others have re-ported that the depletion of SP1 increases the sensitivity of cells to DNA damage [94,95,96] and eventually leads to apoptosis. Thus, SP1 could have a dual function in apoptosis. Moreover, it has been reported that depletion of KLF7 increases cell apoptosis in animal models [97]. Although KLF6 has been reported to be regulated of mitochondrial function during apoptosis [98,99], no information is available for KLF7 in this function. However, it has recently been proposed that KLF7 could inhibit the inflammatory and apoptotic processes in cell lines via NRF1/KLF7 [100]. Thus, in the context of SZ, altered expression of SP1 and KLF7 could activate apoptotic signaling pathways in the CNS and contribute to the disseminated apoptosis described in SZ [101].
4. Materials and Methods
4.1. Bioinformatic Analysis
To identify transcription factor enrichment we used FunRich Tool v.3.1.3. To represent the results obtained with FunRich Tool, we used Graph Prism version 7.00 (GraphPad Software, San Diego, CA, USA). To perform non-redundant enriched categories analysis for Gene Ontology, and Pathways we used Webgestalt (WEB-based Gene SeT Analysis Toolking) and the method of Over-Representation Analysis (ORA), supported by Fisher`s exact test [102]. For pathways analysis we used the Reactome database. The enrichment analyses were set to FDR=0.1. To represent the enrichment analysis, we performed a heat map with Perseus software platform (version 1.6.1.3. http://coxdocs.org/doku.php?id=perseus:start#cite)
5. Conclusions
The altered proteins in the cerebellum in schizophrenia are the target genes of just 11 transcription factors: SP1, SP4, EGR1, KLF7, HNF4A, CTCF, MEF2A, GABPA, NRF1, YY1 and NYFA. Our results show that transport-related pathways are enriched for SP1, KLF7, EGR1, HNF4A and CTCF altered targets. The signaling-related pathways are enriched for SP1, KLF7 and EGR1 altered targets. SP1 and KLF7 could contribute to the signaling dysfunction induced by dendritic arborization alterations and to the loss of the maturation of granular cells in the cerebellum respectively. Pathways involving inflammatory/immune responses and apoptosis are enriched with SP1 and KLF7 altered targets. SP1 could participate in the immune response and induce the differentiation of T helper cells and KLF7 could induce the macrophage activation. This suggests that SP1 and KLF7 could play a prominent role in the cerebellum in chronic schizophrenia. Together, all these findings suggest that the altered function of a limited number of transcription factors could have an impact on disseminated pathways involved in different cellular functions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary dataset 1: Transcription factors. Supplementary dataset 2: Proteins targets for each transcription factor.
Author Contributions
Conceptualization, A.V. and B.R.; Formal analysis, A.V and B.R.; Funding acquisition, B.R.; Investigation, A.V., and B.R.; Methodology, A.V; Project administration, B.R.; Resources, B.R.; Software, A.V.; Supervision, B.R.; Writing – original draft, A.V. and B.R.; Writing – review & editing, A.V., and B.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Miguel Servet grant (MS16/00153-CP16/00153 to BR) financed and integrated into the National R + D + I and funded by the Instituto de Salud Carlos III (Spanish Ministry of Health)—General Branch Evaluation and Promotion of Health Research—and the European Regional Development Fund (ERDF). This work was also supported by CONICYT-Doctorado Becas Chile 2015 (72160426 grant number to AV) and Universidad San Sebastián, Chile (USS-FIN-24-PASI-09 to AV), the Institute de Salud Carlos III (Spanish Ministry of Health) (PI18/00213 to BR).
Institutional Review Board Statement
This study was approved by the Institutional Ethics Committee of Parc Sanitari Sant Joan de Déu (code PIC151-16 and date approval 24-November-2016).
Informed Consent Statement
Not applicable for this study.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Rose for the English editing of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| SZ | Schizophrenia |
| CB | Cerebellum |
| CCTC | Cortico-thalamo-cerebellar circuit |
| CNS | Central nervous system |
| TFs | Transcription factors |
| NKX2-1 | Homeobox protein Nkx-2.1 |
| SP1 | Transcription factor SP1 |
| SP4 | Transcription factor SP4 |
| KLF7 | Krüppel-like factor 7 |
| EGR1 | Early growth response protein 1 |
| HNF4A | Hepatocyte nuclear factor 4-alpha |
| CTCF | Transcriptional repressor CTCFL |
| GABPA | GA-binding protein alpha chain |
| NRF1 | Endoplasmic reticulum membrane sensor NFE2L1 |
| NFYA | Nuclear transcription factor Y subunit alpha |
| MEF2A | Myocyte-specific enhancer factor 2A |
| YY1 | Transcriptional repressor protein YY1 |
References
- Murray, R.M.; Bhavsar, V.; Tripoli, G.; Howes, O. 30 Years on: How the Neurodevelopmental Hypothesis of Schizophrenia Morphed Into the Developmental Risk Factor Model of Psychosis. Schizophr. Bull. 2017, 43, 1190–1196. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Eyre, H.; Jacka, F.N.; Dodd, S.; Dean, O.; McEwen, S.; Debnath, M.; McGrath, J.; Maes, M.; Amminger, P.; et al. A review of vulnerability and risks for schizophrenia: Beyond the two hit hypothesis. Neurosci Biobehav Rev. 2016, 65, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, J.L.; Giedd, J.N.; Gogtay, N. Neurodevelopmental model of schizophrenia: update 2012. Mol. Psychiatry 2012, 17, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Guerrin CGJ, Doorduin J, Sommer IE, de Vries EFJ. The dual hit hypothesis of schizophrenia: Evidence from animal models. Vol. 131, Neuroscience and Biobehavioral Reviews. Elsevier Ltd; 2021. p. 1150–68.
- Andreasen NC, Daniel S, Leary SO, Paradiso S. ·" Cognitive of " Cognitive Dysmetria " as an Integrative Theory of Dysfunction in Cortical- Schizophrenia : A Dysfunction Subcortical--Cerebellar Circuitry ? Network. 2018;(March):203–18.
- Andreasen, N.C.; Nopoulos, P.; O'Leary, D.S.; Miller, D.D.; Wassink, T.; Flaum, M. Defining the phenotype of schizophrenia: Cognitive dysmetria and its neural mechanisms. Biol. Psychiatry 1999, 46, 908–920. [Google Scholar] [CrossRef]
- Bernard, J.A.; Orr, J.M.; Mittal, V.A. Cerebello-thalamo-cortical networks predict positive symptom progression in individuals at ultra-high risk for psychosis. NeuroImage: Clin. 2017, 14, 622–628. [Google Scholar] [CrossRef]
- Eun Kim S, Jung S, Sung G, Bang M, Lee SH. Impaired cerebro-cerebellar white matter connectivity and its associations with cognitive function in patients with schizophrenia. 2021. [CrossRef]
- Chen, P.; Ye, E.; Jin, X.; Zhu, Y.; Wang, L. Association between Thalamocortical Functional Connectivity Abnormalities and Cognitive Deficits in Schizophrenia. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Pfaff, DW. Neuroscience in the 21st century: From basic to clinical. Neuroscience in the 21st Century: From Basic to Clinical. 2013;1–3111.
- Leto, K.; Arancillo, M.; Becker, E.B.E.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.B.; Dusart, I.; Haldipur, P.; Hatten, M.E.; et al. Consensus Paper: Cerebellar Development. Cerebellum 2016, 15, 789–828. [Google Scholar] [CrossRef]
- Badowska DM, Brzózka MM, Kannaiyan N, Thomas C, Dibaj P, Chowdhury A, et al. Modulation of cognition and neuronal plasticity in gain-and loss-of-function mouse models of the schizophrenia risk gene Tcf4. 2020. [CrossRef]
- Mesman, S.; Bakker, R.; Smidt, M.P. Tcf4 is required for correct brain development during embryogenesis. Mol. Cell. Neurosci. 2020, 106, 103502. [Google Scholar] [CrossRef]
- Ramos, B.; Gaudillière, B.; Bonni, A.; Gill, G. Transcription factor Sp4 regulates dendritic patterning during cerebellar maturation. Proc. Natl. Acad. Sci. 2007, 104, 9882–9887. [Google Scholar] [CrossRef]
- Pinacho, R.; Valdizán, E.M.; Pilar-Cuellar, F.; Prades, R.; Tarragó, T.; Haro, J.M.; Ferrer, I.; Ramos, B. Increased SP4 and SP1 transcription factor expression in the postmortem hippocampus of chronic schizophrenia. J. Psychiatr. Res. 2014, 58, 189–196. [Google Scholar] [CrossRef]
- Ye, B.; Kim, J.H.; Yang, L.; McLachlan, I.; Younger, S.; Jan, L.Y.; Jan, Y.N. Differential Regulation of Dendritic and Axonal Development by the Novel Krüppel-Like Factor Dar1. J. Neurosci. 2011, 31, 3309–3319. [Google Scholar] [CrossRef] [PubMed]
- Hsueh YP, Hirata Y, Simmen FA, Denver RJ, Ávila-Mendoza J, Subramani A. Krüppel-Like Factors 9 and 13 Block Axon Growth by Transcriptional Repression of Key Components of the cAMP Signaling Pathway. 2020; Available from: www.frontiersin.
- Malt, E.A.; Juhasz, K.; Malt, U.F.; Naumann, T. A Role for the Transcription Factor Nk2 Homeobox 1 in Schizophrenia: Convergent Evidence from Animal and Human Studies. Front. Behav. Neurosci. 2016, 10, 59–59. [Google Scholar] [CrossRef] [PubMed]
- Vera-Montecinos, A.; Rodríguez-Mias, R.; MacDowell, K.S.; García-Bueno, B.; Bris, Á.G.; Caso, J.R.; Villén, J.; Ramos, B. Analysis of Molecular Networks in the Cerebellum in Chronic Schizophrenia: Modulation by Early Postnatal Life Stressors in Murine Models. Int. J. Mol. Sci. 2021, 22, 10076. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shachar, D. The interplay between mitochondrial complex I, dopamine and Sp1 in schizophrenia. J Neural Transm. 2009;116(11):1383–96.
- Fusté, M.; Meléndez-Pérez, I.; Villalta-Gil, V.; Pinacho, R.; Villalmanzo, N.; Cardoner, N.; Menchón, J.M.; Haro, J.M.; Soriano-Mas, C.; Ramos, B. Specificity proteins 1 and 4, hippocampal volume and first-episode psychosis. Br. J. Psychiatry 2016, 208, 591–592. [Google Scholar] [CrossRef]
- Fusté, M.; Pinacho, R.; Meléndez-Pérez, I.; Villalmanzo, N.; Villalta-Gil, V.; Haro, J.M.; Ramos, B. Reduced expression of SP1 and SP4 transcription factors in peripheral blood mononuclear cells in first-episode psychosis. J. Psychiatr. Res. 2013, 47, 1608–1614. [Google Scholar] [CrossRef]
- Chen, J.; He, K.; Wang, Q.; Li, Z.; Shen, J.; Li, T.; Wang, M.; Wen, Z.; Li, W.; Qiang, Y.; et al. Role played by theSP4gene in schizophrenia and major depressive disorder in the Han Chinese population. Br. J. Psychiatry 2016, 208, 441–445. [Google Scholar] [CrossRef]
- Pinacho, R.; Villalmanzo, N.; Roca, M.; Iniesta, R.; Monje, A.; Haro, J.M.; Meana, J.J.; Ferrer, I.; Gill, G.; Ramos, B. Analysis of Sp transcription factors in the postmortem brain of chronic schizophrenia: A pilot study of relationship to negative symptoms. J. Psychiatr. Res. 2013, 47, 926–934. [Google Scholar] [CrossRef]
- Pinacho, R.; Saia, G.; Meana, J.J.; Gill, G.; Ramos, B. Transcription factor SP4 phosphorylation is altered in the postmortem cerebellum of bipolar disorder and schizophrenia subjects. Eur. Neuropsychopharmacol. 2015, 25, 1650–1660. [Google Scholar] [CrossRef]
- Saia, G.; Lalonde, J.; Sun, X.; Ramos, B.; Gill, G. Phosphorylation of the transcription factor Sp4 is reduced by NMDA receptor signaling. J. Neurochem. 2014, 129, 743–752. [Google Scholar] [CrossRef]
- Pinacho, R.; Saia, G.; Fusté, M.; Meléndez-Pérez, I.; Villalta-Gil, V.; Haro, J.M.; Gill, G.; Ramos, B. Phosphorylation of Transcription Factor Specificity Protein 4 Is Increased in Peripheral Blood Mononuclear Cells of First-Episode Psychosis. PLOS ONE 2015, 10, e0125115–e0125115. [Google Scholar] [CrossRef]
- Zhou, X. Over-representation of potential SP4 target genes within schizophrenia-risk genes. Mol. Psychiatry 2021, 27, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.-M.; Chen, S.-J.; Hsu, S.-H.; Cheng, M.-C. Functional analyses and effect of DNA methylation on the EGR1 gene in patients with schizophrenia. Psychiatry Res. 2019, 275, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Ramaker, R.C.; Bowling, K.M.; Lasseigne, B.N.; Hagenauer, M.H.; Hardigan, A.A.; Davis, N.S.; Gertz, J.; Cartagena, P.M.; Walsh, D.M.; Vawter, M.P.; et al. Post-mortem molecular profiling of three psychiatric disorders. Genome Med. 2017, 9, 1–12. [Google Scholar] [CrossRef]
- Iwakura, Y.; Kawahara-Miki, R.; Kida, S.; Sotoyama, H.; Gabdulkhaev, R.; Takahashi, H.; Kunii, Y.; Hino, M.; Nagaoka, A.; Izumi, R.; et al. Elevation of EGR1/zif268, a Neural Activity Marker, in the Auditory Cortex of Patients with Schizophrenia and its Animal Model. Neurochem. Res. 2022, 47, 2715–2727. [Google Scholar] [CrossRef] [PubMed]
- Vawter, M.P.; Mamdani, F.; Macciardi, F. An integrative functional genomics approach for discovering biomarkers in schizophrenia. Briefings Funct. Genom. 2011, 10, 387–399. [Google Scholar] [CrossRef]
- Juraeva, D.; Haenisch, B.; Zapatka, M.; Frank, J.; GROUP Investigators ,; PSYCH-GEMS SCZ working group ,; Witt, S.H.; Mühleisen, T.W.; Treutlein, J.; Strohmaier, J.; et al. Integrated Pathway-Based Approach Identifies Association between Genomic Regions at CTCF and CACNB2 and Schizophrenia. PLOS Genet. 2014, 10, e1004345. [CrossRef]
- Huo, Y.; Li, S.; Liu, J.; Li, X.; Luo, X.-J. Functional genomics reveal gene regulatory mechanisms underlying schizophrenia risk. Nat. Commun. 2019, 10, 1–19. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Liu, J.; Wang, J.; Li, X.; Huo, Y.; Li, Y.; Liu, Y.; Li, M.; Xiao, X.; et al. Regulatory variants at 2q33.1 confer schizophrenia risk by modulating distal gene TYW5 expression. Brain 2021, 145, 770–786. [Google Scholar] [CrossRef]
- McMeekin, L.J.; Lucas, E.K.; Meador-Woodruff, J.H.; McCullumsmith, R.E.; Hendrickson, R.C.; Gamble, K.L.; Cowell, R.M. Cortical PGC-1α-Dependent Transcripts Are Reduced in Postmortem Tissue From Patients With Schizophrenia. Schizophr. Bull. 2016, 42, 1009–1017. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Ma, X.; Long, Q.; Yu, L.; Chen, Y.; Wu, W.; Guo, Z.; Teng, Z.; Zeng, Y. The NRF1/miR-4514/SOCS3 Pathway Is Associated with Schizophrenia Pathogenesis. Clin. Neurol. Neurosci. 2021, 5, 82. [Google Scholar] [CrossRef]
- Smeland, O.B.; Frei, O.; Kauppi, K.; Hill, W.D.; Li, W.; Wang, Y.; Krull, F.; Bettella, F.; Eriksen, J.A.; Witoelar, A.; et al. Identification of Genetic Loci Jointly Influencing Schizophrenia Risk and the Cognitive Traits of Verbal-Numerical Reasoning, Reaction Time, and General Cognitive Function. JAMA Psychiatry 2017, 74, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.C.; Javidfar, B.; Pothula, V.; Ibi, D.; Shen, E.Y.; Peter, C.J.; Bicks, L.K.; Fehr, T.; Jiang, Y.; Brennand, K.J.; et al. MEF2C transcription factor is associated with the genetic and epigenetic risk architecture of schizophrenia and improves cognition in mice. Mol. Psychiatry 2018, 23, 123–132. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, J.; Crofts, L.A.; Quinlan, K.G.; Czolij, R.; Perkins, A.C.; Crossley, M. Human KLF17 is a new member of the Sp/KLF family of transcription factors. Genomics 2006, 87, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xu, J.; Chen, J.; Kim, S.; Reimers, M.; Bacanu, S.-A.; Yu, H.; Liu, C.; Sun, J.; Wang, Q.; et al. Transcriptome sequencing and genome-wide association analyses reveal lysosomal function and actin cytoskeleton remodeling in schizophrenia and bipolar disorder. Mol. Psychiatry 2015, 20, 563–572. [Google Scholar] [CrossRef]
- Hung, C.-Y.; Hsu, T.-I.; Chuang, J.-Y.; Su, T.-P.; Chang, W.-C.; Hung, J.-J. Sp1 in Astrocyte Is Important for Neurite Outgrowth and Synaptogenesis. Mol. Neurobiol. 2020, 57, 261–277. [Google Scholar] [CrossRef]
- Ramos, B.; Valín, A.; Sun, X.; Gill, G. Sp4-dependent repression of neurotrophin-3 limits dendritic branching. Mol. Cell. Neurosci. 2009, 42, 152–159. [Google Scholar] [CrossRef]
- Laub, F.; Lei, L.; Sumiyoshi, H.; Kajimura, D.; Dragomir, C.; Smaldone, S.; Puche, A.C.; Petros, T.J.; Mason, C.; Parada, L.F.; et al. Transcription Factor KLF7 Is Important for Neuronal Morphogenesis in Selected Regions of the Nervous System. Mol. Cell. Biol. 2005, 25, 5699–5711. [Google Scholar] [CrossRef]
- Laub, F.; Aldabe, R.; Friedrich, V.; Ohnishi, S.; Yoshida, T.; Ramirez, F. Developmental Expression of Mouse Krüppel-like Transcription Factor KLF7 Suggests a Potential Role in Neurogenesis. Dev. Biol. 2001, 233, 305–318. [Google Scholar] [CrossRef]
- Laub, F.; Lei, L.; Sumiyoshi, H.; Kajimura, D.; Dragomir, C.; Smaldone, S.; Puche, A.C.; Petros, T.J.; Mason, C.; Parada, L.F.; et al. Transcription Factor KLF7 Is Important for Neuronal Morphogenesis in Selected Regions of the Nervous System. Mol. Cell. Biol. 2005, 25, 5699–5711. [Google Scholar] [CrossRef]
- Barry, G.; A Briggs, J.; Vanichkina, D.P.; Poth, E.M.; Beveridge, N.J.; Ratnu, V.S.; Nayler, S.P.; Nones, K.; Hu, J.; Bredy, T.W.; et al. The long non-coding RNA Gomafu is acutely regulated in response to neuronal activation and involved in schizophrenia-associated alternative splicing. Mol. Psychiatry 2014, 19, 486–494. [Google Scholar] [CrossRef]
- Saia-Cereda, V.M.; Santana, A.G.; Schmitt, A.; Falkai, P.; Martins-De-Souza, D. The Nuclear Proteome of White and Gray Matter from Schizophrenia Postmortem Brains. Complex Psychiatry 2017, 3, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Nakata, K.; Lipska, B.K.; Hyde, T.M.; Ye, T.; Newburn, E.N.; Morita, Y.; Vakkalanka, R.; Barenboim, M.; Sei, Y.; Weinberger, D.R.; et al. DISC1 splice variants are upregulated in schizophrenia and associated with risk polymorphisms. Proc. Natl. Acad. Sci. 2009, 106, 15873–15878. [Google Scholar] [CrossRef]
- Law, A.J.; Kleinman, J.E.; Weinberger, D.R.; Weickert, C.S. Disease-associated intronic variants in the ErbB4 gene are related to altered ErbB4 splice-variant expression in the brain in schizophrenia. Hum. Mol. Genet. 2007, 16, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Alberstein, M.; Amit, M.; Vaknin, K.; O'Donnell, A.; Farhy, C.; Lerenthal, Y.; Shomron, N.; Shaham, O.; Sharrocks, A.D.; Ashery-Padan, R.; et al. Regulation of transcription of the RNA splicing factor hSlu7 by Elk-1 and Sp1 affects alternative splicing. RNA 2007, 13, 1988–1999. [Google Scholar] [CrossRef] [PubMed]
- Shalizi A, Gaudillière B, Yuan Z, Stegmüller J, Shirogane T, Ge Q, et al. A Calcium-Regulated MEF2 Sumoylation Switch Controls Postsynaptic Differentiation. Science (1979). 2006 Feb 17;311(5763):1012 LP – 1017.
- Lisek, M.; Przybyszewski, O.; Zylinska, L.; Guo, F.; Boczek, T. The Role of MEF2 Transcription Factor Family in Neuronal Survival and Degeneration. Int. J. Mol. Sci. 2023, 24, 3120. [Google Scholar] [CrossRef]
- Crisafulli, C.; Drago, A.; Calabrò, M.; Spina, E.; Serretti, A. Progress in Neuro-Psychopharmacology & Biological Psychiatry A molecular pathway analysis informs the genetic background at risk for schizophrenia. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2015, 59, 21–30. [Google Scholar] [CrossRef]
- Thygesen, J.H.; Zambach, S.K.; Ingason, A.; Lundin, P.; Hansen, T.; Bertalan, M.; Rosengren, A.; Bjerre, D.; Ferrero-Miliani, L.; Rasmussen, H.B.; et al. Linkage and whole genome sequencing identify a locus on 6q25–26 for formal thought disorder and implicate MEF2A regulation. Schizophr. Res. 2015, 169, 441–446. [Google Scholar] [CrossRef]
- Carmichael, R.E.; Wilkinson, K.A.; Craig, T.J.; Ashby, M.C.; Henley, J.M. MEF2A regulates mGluR-dependent AMPA receptor trafficking independently of Arc/Arg3.1. Sci. Rep. 2018, 8, 5263. [Google Scholar] [CrossRef]
- Watson P, T DJS. ER-to-Golgi transport : Form and formation of vesicular and tubular carriers. 2005;1744:304–15.
- Qin, F.; Chen, G.; Yu, K.N.; Yang, M.; Cao, W.; Kong, P.; Peng, S.; Sun, M.; Nie, L.; Han, W. Golgi Phosphoprotein 3 Mediates Radiation-Induced Bystander Effect via ERK/EGR1/TNF-α Signal Axis. Antioxidants 2022, 11, 2172. [Google Scholar] [CrossRef]
- Etemadik M, Nia A, Wetter L, Feuk L. Transcriptome analysis of fibroblasts from schizophrenia patients reveals differential expression of schizophrenia-related genes. 2020;1–9.
- Vawter M, Mamdani F, Macciardi F. An integrative functional genomics approach for discovering biomarkers in schizophrenia. 2011;(December).
- Altamura, A.C.; Pozzoli, S.; Fiorentini, A.; Dell'Osso, B. Neurodevelopment and inflammatory patterns in schizophrenia in relation to pathophysiology. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2013, 42, 63–70. [Google Scholar] [CrossRef]
- Meyer, J.M.; McEvoy, J.P.; Davis, V.G.; Goff, D.C.; Nasrallah, H.A.; Davis, S.M.; Hsiao, J.K.; Swartz, M.S.; Stroup, T.S.; Lieberman, J.A. Inflammatory Markers in Schizophrenia: Comparing Antipsychotic Effects in Phase 1 of the Clinical Antipsychotic Trials of Intervention Effectiveness Study. Biol. Psychiatry 2009, 66, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.S.; Phillips, M.R.; Yang, L.H.; Kegeles, L.S.; Susser, E.S.; Lieberman, J.A. Neurodegenerative model of schizophrenia: Growing evidence to support a revisit. Schizophr. Res. 2022, 243, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Comer, A.L.; Carrier, M.; Tremblay, M.È.; Cruz-Martín, A. The Inflamed Brain in Schizophrenia: The Convergence of Genetic and Environmental Risk Factors That Lead to Uncontrolled Neuroinflammation. Front. Cell. Neurosci. 2020, 14, 274. [Google Scholar] [CrossRef]
- Gober, R.; Dallmeier, J.; Davis, D.; Brzostowicki, D.; Vaccari, J.P.d.R.; Cyr, B.; Barreda, A.; Sun, X.; Gultekin, S.H.; Garamszegi, S.; et al. Increased inflammasome protein expression identified in microglia from postmortem brains with schizophrenia. J. Neuropathol. Exp. Neurol. 2024, 83, 951–966. [Google Scholar] [CrossRef]
- Miller, B.J.; Goldsmith, D.R. Evaluating the Hypothesis That Schizophrenia Is an Inflammatory Disorder. FOCUS 2020, 18, 391–401. [Google Scholar] [CrossRef]
- Howes OD, Mccutcheon R. Inflammation and the neural diathesis-stress hypothesis of schizophrenia: a reconceptualization. 2017;7. Available from: www.nature.com/tp.
- Hughes, H.; Ashwood, P. Overlapping evidence of innate immune dysfunction in psychotic and affective disorders. Brain, Behav. Immun. - Heal. 2020, 2, 100038. [Google Scholar] [CrossRef]
- Fond G, Lançon C, Korchia T, Auquier P, Boyer L. The Role of Inflammation in the Treatment of Schizophrenia. Vol. 11, Frontiers in Psychiatry. Frontiers Media S.A.; 2020.
- Carril Pardo C, Oyarce Merino K, Vera-Montecinos A. Neuroinflammatory Loop in Schizophrenia, Is There a Relationship with Symptoms or Cognition Decline? Vol. 26, International Journal of Molecular Sciences. Multidisciplinary Digital Publishing Institute (MDPI); 2025.
- Cui, L.-B.; Wang, X.-Y.; Fu, Y.-F.; Liu, X.-F.; Wei, Y.; Zhao, S.-W.; Gu, Y.-W.; Fan, J.-W.; Wu, W.-J.; Gong, H.; et al. Transcriptional level of inflammation markers associates with short-term brain structural changes in first-episode schizophrenia. BMC Med. 2023, 21, 1–13. [Google Scholar] [CrossRef]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 cytokine: An overview of the immune regulation, immune dysregulation, and therapeutic approach. Int. Immunopharmacol. 2022, 111, 109130. [Google Scholar] [CrossRef]
- Yang X, Liang M, Tang Y, Ma D, Li M, Yuan C, et al. KLF7 promotes adipocyte inflammation and glucose metabolism disorder by activating the PKCζ/NF-κB pathway. FASEB Journal. 2023 Jul 1;37(7).
- Zhang, M.; Wang, C.; Wu, J.; Ha, X.; Deng, Y.; Zhang, X.; Wang, J.; Chen, K.; Feng, J.; Zhu, J.; et al. The Effect and Mechanism of KLF7 in the TLR4/NF-κB/IL-6 Inflammatory Signal Pathway of Adipocytes. Mediat. Inflamm. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Borovac, J.; Bosch, M.; Okamoto, K. Regulation of actin dynamics during structural plasticity of dendritic spines: Signaling messengers and actin-binding proteins. Mol. Cell. Neurosci. 2018, 91, 122–130. [Google Scholar] [CrossRef]
- Zhou, X.; Tian, B.; Han, H.-B. Serum interleukin-6 in schizophrenia: A system review and meta-analysis. Cytokine 2021, 141, 155441. [Google Scholar] [CrossRef] [PubMed]
- Nayak, L.; Goduni, L.; Takami, Y.; Sharma, N.; Kapil, P.; Jain, M.K.; Mahabeleshwar, G.H. Kruppel-Like Factor 2 Is a Transcriptional Regulator of Chronic and Acute Inflammation. 2013, 182, 1696–1704. [CrossRef]
- Luo, W.-W.; Lian, H.; Zhong, B.; Shu, H.-B.; Li, S. Krüppel-like factor 4 negatively regulates cellular antiviral immune response. Cell. Mol. Immunol. 2016, 13, 65–72. [Google Scholar] [CrossRef]
- Date, D.; Das, R.; Narla, G.; Simon, D.I.; Jain, M.K.; Mahabeleshwar, G.H. Kruppel-like Transcription Factor 6 Regulates Inflammatory Macrophage Polarization. J. Biol. Chem. 2014, 289, 10318–10329. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Kim, H.-P.; Xue, H.-H.; Liu, H.; Zhao, K.; Leonard, W.J. Interleukin-21 Receptor Gene Induction in Human T Cells Is Mediated by T-Cell Receptor-Induced Sp1 Activity. Mol. Cell. Biol. 2005, 25, 9741–9752. [Google Scholar] [CrossRef] [PubMed]
- El-Said H, FKMAR; et al. MiR302c, Sp1, and NFATc2 regulate interleukin-21 expression in human CD4+CD45RO+ T lymphocytes. J Cell Physiol. 2019;234:5998–6011.
- Shbeer, A.M.; Robadi, I.A. The role of Interleukin-21 in autoimmune Diseases: Mechanisms, therapeutic Implications, and future directions. Cytokine 2024, 173, 156437. [Google Scholar] [CrossRef]
- Sun, H.-J.; Xu, X.; Wang, X.-L.; Wei, L.; Li, F.; Lu, J.; Huang, B.-Q. Transcription Factors Ets2 and Sp1 Act Synergistically with Histone Acetyltransferase p300 in Activating Human Interleukin-12 p40 Promoter. Acta Biochim. et Biophys. Sin. 2006, 38, 194–200. [Google Scholar] [CrossRef]
- Powell, M.D.; Read, K.A.; Sreekumar, B.K.; Jones, D.M.; Oestreich, K.J. IL-12 signaling drives the differentiation and function of a TH1-derived TFH1-like cell population. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Kim Y k, Suh I b, Kim H, Han C s, Lim C s, Choi S h, et al. The plasma levels of interleukin-12 in schizophrenia, major depression, and bipolar mania : effects of psychotropic drugs. 2002;1107–14.
- Ozbey, U.; Tug, E.; Kara, M.; Namli, M. The value of interleukin-12B (p40) gene promoter polymorphism in patients with schizophrenia in a region of East Turkey. Psychiatry Clin. Neurosci. 2008, 62, 307–312. [Google Scholar] [CrossRef]
- Lee, K.-. .-W.; Lee, Y.; Kwon, H.-.-J.; Kim, D.-.-S. Sp1-associated activation of macrophage inflammatory protein-2 promoter by CpG-oligodeoxynucleotide and lipopolysaccharide. Cell. Mol. Life Sci. 2005, 62, 188–198. [Google Scholar] [CrossRef]
- Iwanaszko, M.; Kimmel, M. NF-κB and IRF pathways: cross-regulation on target genes promoter level. BMC Genom. 2015, 16, 1–8. [Google Scholar] [CrossRef]
- MacDowell, K.S.; Pinacho, R.; Leza, J.C.; Costa, J.; Ramos, B.; García-Bueno, B. Differential regulation of the TLR4 signalling pathway in post-mortem prefrontal cortex and cerebellum in chronic schizophrenia: Relationship with SP transcription factors. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2017, 79, 481–492. [Google Scholar] [CrossRef]
- O’Donnell BF, Faux SF, McCarley RW, Kimble MO, Salisbury DF, Nestor PG, et al. Increased Rate of P300 Latency Prolongation with Age in Schizophrenia: Electrophysiological Evidence for a Neurodegenerative Process. Arch Gen Psychiatry. 1995 Jul 1;52(7):544–9.
- Morén, C.; Treder, N.; Martínez-Pinteño, A.; Rodríguez, N.; Arbelo, N.; Madero, S.; Gómez, M.; Mas, S.; Gassó, P.; Parellada, E. Systematic Review of the Therapeutic Role of Apoptotic Inhibitors in Neurodegeneration and Their Potential Use in Schizophrenia. Antioxidants 2022, 11, 2275. [Google Scholar] [CrossRef] [PubMed]
- Glantz, L.A.; Gilmore, J.H.; Lieberman, J.A.; Jarskog, L.F. Apoptotic mechanisms and the synaptic pathology of schizophrenia. Schizophr. Res. 2006, 81, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Parellada, E.; Gassó, P. Glutamate and microglia activation as a driver of dendritic apoptosis: a core pathophysiological mechanism to understand schizophrenia. Transl. Psychiatry 2021, 11, 271. [Google Scholar] [CrossRef]
- Deniaud, E.; Baguet, J.; Mathieu, A.-L.; Pagès, G.; Marvel, J.; Leverrier, Y. Overexpression of Sp1 transcription factor induces apoptosis. Oncogene 2006, 25, 7096–7105. [Google Scholar] [CrossRef]
- Torabi, B.; Flashner, S.; Beishline, K.; Sowash, A.; Donovan, K.; Bassett, G.; Azizkhan-Clifford, J. Caspase cleavage of transcription factor Sp1 enhances apoptosis. Apoptosis 2018, 23, 65–78. [Google Scholar] [CrossRef]
- Deniaud E, Baguet J, Chalard R, Blanquier B, Brinza L, Meunier J, et al. Overexpression of transcription factor Sp1 leads to gene expression perturbations and cell cycle inhibition. Overexpression of Transcription Factor Sp1 Leads to Gene Expression Perturbations and Cell Cycle Inhibition. PLoS One [Internet]. 2009;4(9). Available from: https://inria.hal.science/hal-00851247.
- Lei, L.; Laub, F.; Lush, M.; Romero, M.; Zhou, J.; Luikart, B.; Klesse, L.; Ramirez, F.; Parada, L.F. The zinc finger transcription factor Klf7 is required for TrkA gene expression and development of nociceptive sensory neurons. Genes Dev. 2005, 19, 1354–1364. [Google Scholar] [CrossRef]
- Mallipattu, S.K.; Horne, S.J.; D’agati, V.; Narla, G.; Liu, R.; Frohman, M.A.; Dickman, K.; Chen, E.Y.; Ma’ayan, A.; Bialkowska, A.B.; et al. Krüppel-like factor 6 regulates mitochondrial function in the kidney. J. Clin. Investig. 2015, 125, 1347–1361. [Google Scholar] [CrossRef]
- Piret, S.E.; Guo, Y.; Attallah, A.A.; Horne, S.J.; Zollman, A.; Owusu, D.; Henein, J.; Sidorenko, V.S.; Revelo, M.P.; Hato, T.; et al. Krüppel-like factor 6–mediated loss of BCAA catabolism contributes to kidney injury in mice and humans. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Xu, Q.; Kong, F.; Zhao, G.; Jin, J.; Feng, S.; Li, M. USP7 alleviates neuronal inflammation and apoptosis in spinal cord injury via deubiquitinating NRF1/KLF7 axis. Neurol. Res. 2024, 46, 1–10. [Google Scholar] [CrossRef]
- Jarskog, L.F.; Glantz, L.A.; Gilmore, J.H.; Lieberman, J.A. Apoptotic mechanisms in the pathophysiology of schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry. 2005, 29, 846–858. [Google Scholar] [CrossRef]
- Wang, J.; Duncan, D.; Shi, Z.; Zhang, B. WEB-based GEne SeT AnaLysis Toolkit (WebGestalt): update 2013. Nucleic Acids Res. 2013, 41, W77–W83. [Google Scholar] [CrossRef]
Figure 1.
Experimental design to identify enriched transcription factors and its dependent-altered biological processes and pathways in the cerebellum in schizophrenia.
Figure 1.
Experimental design to identify enriched transcription factors and its dependent-altered biological processes and pathways in the cerebellum in schizophrenia.
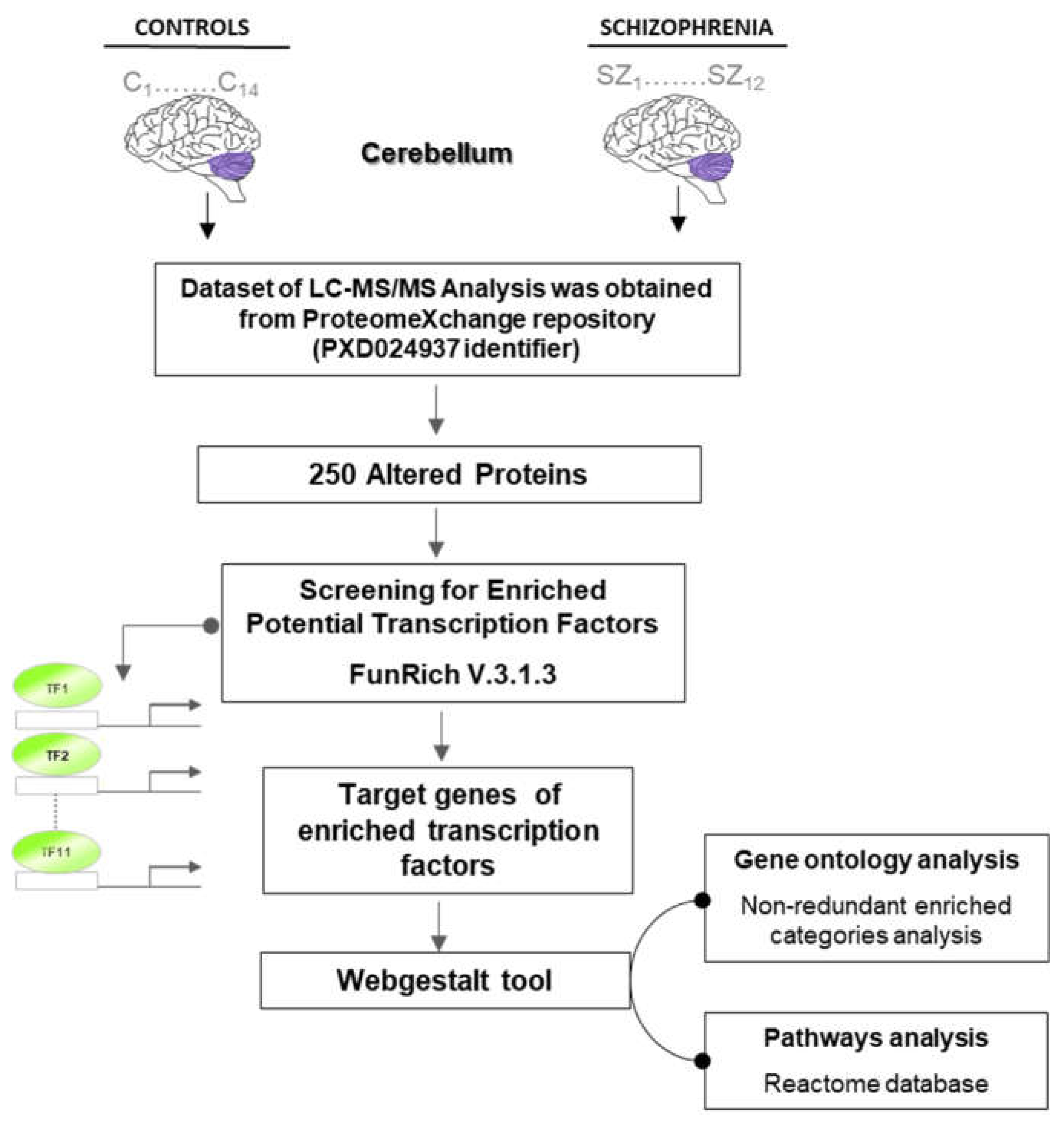
Figure 2.
Potential transcription factors involved in the regulation of the altered proteins in the cerebellum of chronic schizophrenia patients. The X-axes show the percentaje of target genes for each transcription factor. The Y-axes show the –log10 enrichment p-value. The size of the bubble indicates the number of protein targets.
Figure 2.
Potential transcription factors involved in the regulation of the altered proteins in the cerebellum of chronic schizophrenia patients. The X-axes show the percentaje of target genes for each transcription factor. The Y-axes show the –log10 enrichment p-value. The size of the bubble indicates the number of protein targets.
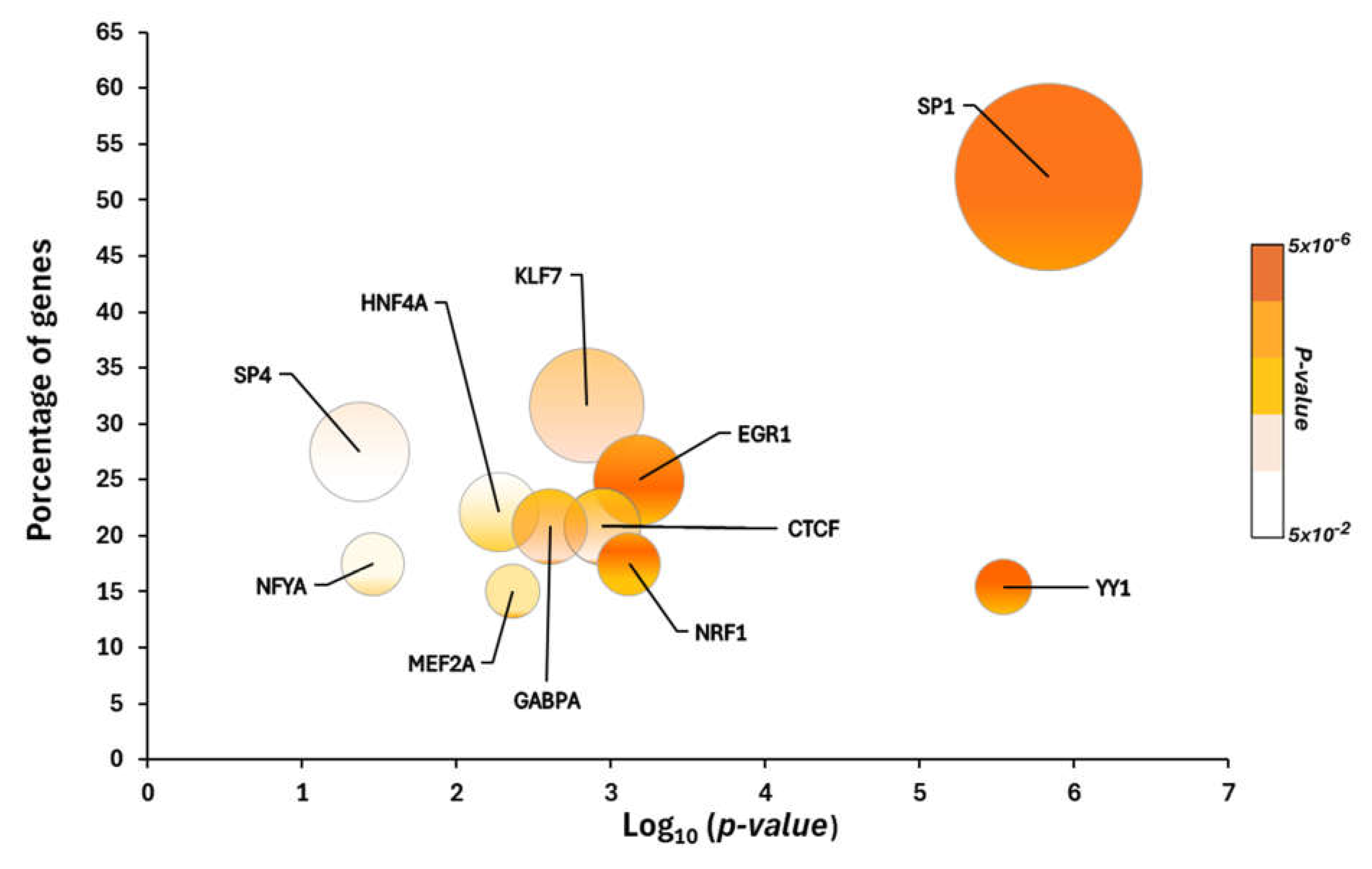
Figure 3.
Non-redundant enriched biological process categories for altered targets of transcription factors. The enrichment analysis was performed using Webgestalt and the heat map visualization of enriched biological process were performed using Perseus software.
Figure 3.
Non-redundant enriched biological process categories for altered targets of transcription factors. The enrichment analysis was performed using Webgestalt and the heat map visualization of enriched biological process were performed using Perseus software.
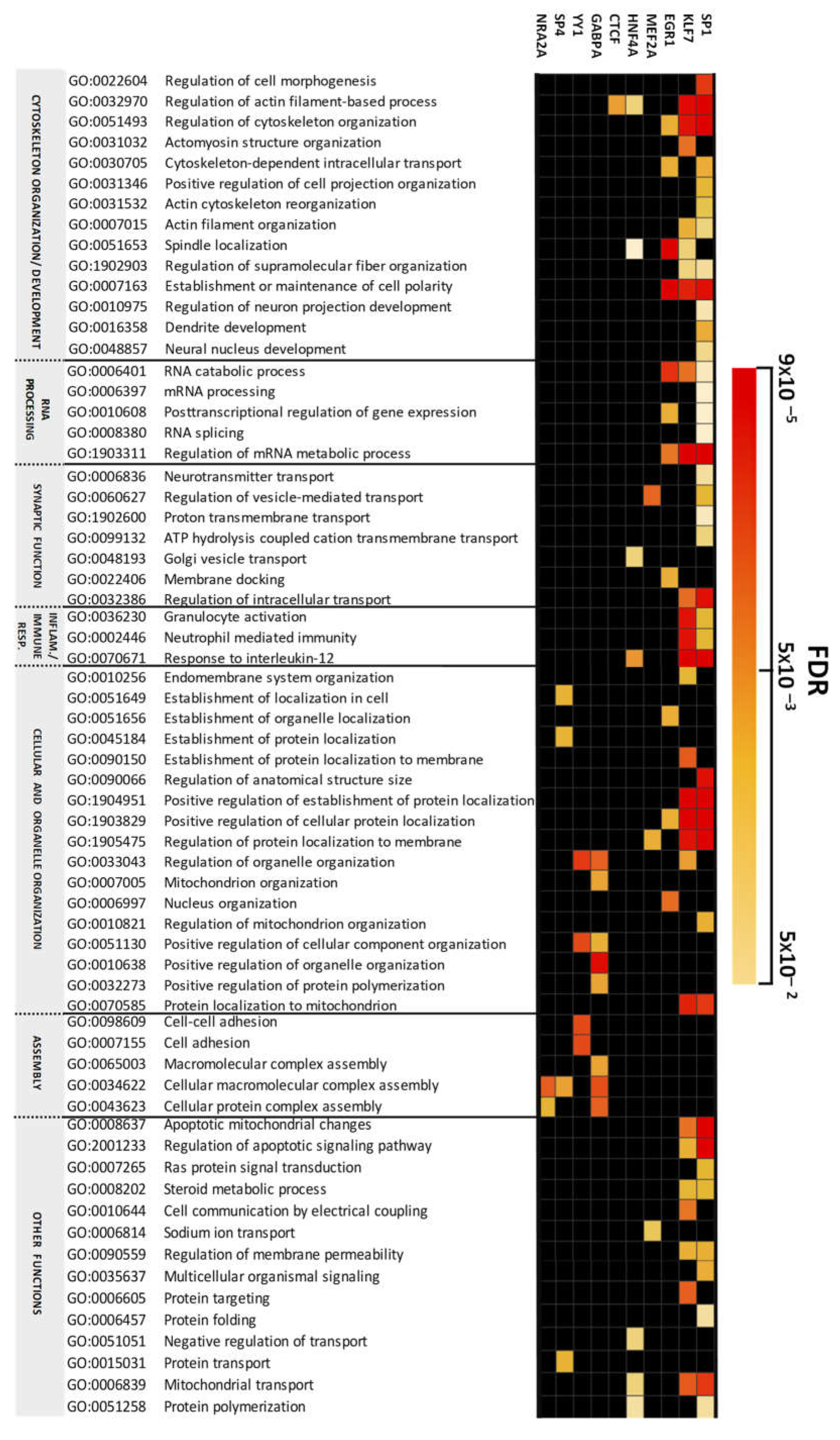
Figure 4.
Non-redundant enriched pathways for altered targets of transcription factors. We used the Reactome database for enrichment pathway analysis and the results are displayed as a heat map created using Perseus software.
Figure 4.
Non-redundant enriched pathways for altered targets of transcription factors. We used the Reactome database for enrichment pathway analysis and the results are displayed as a heat map created using Perseus software.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



