
Submitted:
25 February 2025
Posted:
27 February 2025
You are already at the latest version
Abstract
Anthracnose is a disease caused by phytopathogenic fungi such as Colletotrichum siamense that attacks plants and fruits causing great postharvest losses. Different alternatives for the control of this fungus have been studied, and in the present study we evaluated the in vitro antifungal activity of chitosan mixed with methanolic extracts of Baccharis glutinosa and Jacquinia macrocarpa as well as toxicity in different models. Using the radial growth technique, it was observed that the mycelial development of C. siamense was altered and reduced during exposure to the different treatments evaluated during the first hours of incubation, indicating a fungistatic effect. While the cell viability, by colorimetric assay using the XTT salt, showed alteration since the chitosan reduced proliferation by 50 %, while the plant extracts and their mixtures with chitosan reduced approximately 40 % indicating cell damage, which was confirmed by fluorescence microscopy. In addition, toxicity tests showed that the extract of J. macrocapra affected the germination percentage of Lactuca sativa seeds while radicle length was reduced in all treatments, except with chitosan alone. The larval survival test for Artemia salina with the extracts indicated their potential toxicity by causing up to 60% mortality. The results indicate that the extracts of B. glutinosa and J. macrocarpa mixed with chitosan are a good option for the control of C. siamense, but at the concentrations used they show a toxic effect in the models evaluated.
Keywords:
Chitosan
; Colletotrichum siamense
; acute toxicity
; phytotoxicity
; Baccharis glutinosa
; Jacquinia macrocarpa
1. Introduction
Plant pathogenic fungi are one of the main infectious agents in plants, causing significant crop yield losses worldwide [1]. Genera Colletotrichum, Botrytis, Fusarium and Rhizopus cause the most frequent diseases in fruits and vegetables, the most characteristic symptom being necrosis, which is the structural or functional deterioration of tissues due to the death of their cells [2,3]. In Mexico, 46 species of Colletotrichum spp. have been reported and the interest in studying C. siamense is growing due to numerous cases of anthracnose generated in mango, papaya and avocado [4,5,6]. Anthracnose is controlled with synthetic fungicides whose toxicity makes them a risky practice, since their prolonged use causes harmful effects on the environment, on the biota, on the appearance of resistance by these pathogens, and they are not economically profitable [7]. For this reason, safer, sustainable, non-toxic alternatives and integrated management practices that preserve biodiversity, soil quality, human health and limit the proliferation of these microorganisms and their diseases are being sought [8]. Among these alternatives is the use of natural products with antifungal activity such as plant extracts and chitosan [9,10]. Chitosan is a biopolymer derived from the chitin of the exoskeleton of crustaceans and insects. This polysaccharide is used in the food industry as a preservative for its ability to inhibit the growth of microorganisms, while in agricultural applications it is used as a crop biostimulant, growth promoter and activator of plant immune response defenses [11,12]. Some studies have been reported demonstrating the antifungal effect of chitosan. Li et al. [13], reported the efficacy of chitosan in the control of anthracnose on mango caused by C. gloeosporoides. Additionally, Nascimento et al. [14], found fungistatic and fungicidal effect against C. gloeosporioides, as well as inducing morphological changes in its spores and hyphae. Other alternatives for the control of fungi in food are the extracts of Baccharis glutinosa and Jacquinia macrocarpa, two plants native to the state of Sonora. Rosas-Burgos et al. [15] reported that the methanolic extract of B. glutinosa have antifungal activity against phytopathogenic and toxigenic fungi. On the other hand, methanolic extracts of B. glutinosa and ethanolic of J. macrocarpa showed antifungal activity against F. verticillioides, Aspergillus flavus and A. parasiticus, which indicates that they are a viable alternative for the combat of phytopathogenic fungi [16]. Based on the above, the objective of the present study was to evaluate methanolic extracts of Jacquinia macrocarpa and Baccharis glutinosa alone and in combination with chitosan on the development of Colletotrichum siamense.
2. Materials and Methods
2.1. Materials
Commercial chitosan (CS) of medium molecular weight (153 kDa) was used with a deacetylation degree of 78 % (Sigma Aldrich, CAS. 448877). Lactic acid (J.T. Baker, CAS. 50-21-5) and propidium iodide (>94% purity) (Sigma Aldrich, CAS 25535-16-4).
2.2. Preparation of Methanolic Extract
B. glutinosa plants were collected along the banks of the Sonora River at Aconchi, Sonora (29°49’32.3”N 110°14’07.6”W), while the ones for J. macrocarpa in the area of Los Arrieros, Guaymas, Sonora (28°20’06.2”N 111°08’54.4”W). The leaves of adult plants were taken and placed in plastic bags to be transported to the laboratory. Once there, the stems were removed and the leaves were left to dry at 27 °C for one week. On the eighth day of drying, the leaves of each plant were separately ground to a fine powder for maceration. For this purpose, 60 g of the powder were mixed with 1 L of 70 % methanol and stirred for one week. The mixtures obtained were filtered through Whatman #1 filter paper and placed in a rotary evaporator and then in a fume hood for seven days until complete evaporation. The solubility of the plant extracts was evaluated by mixing 100 mg of each extract with 25 mL of sterile distilled water and only J. macrocarpa was dissolved in it, while B. glutinosa was dissolved in 35% acetone, so they were used in this way in the study.
2.3. Chitosan Solution
A stock solution of 20 g/L was prepared by constant agitation in an aqueous lactic acid solution (1%, v/v) overnight, and the pH was adjusted to 5.3 ± 0.1 using 1 M NaOH. From this stock solution, working solutions of 2 and 4 g/L were prepared for addition to the extracts of B. glutinosa (ExB) and of J. macrocarpa (ExJ).
2.4. Evaluation of Antifungal Activity
A strain of Colletotrichum siamense (H6-1) as used, it was identified by genomic alignment approach. The fungus was inoculated in flasks with enriched v8 medium and potato dextrose agar (BDBioxon) and incubated at 27 ± 2 °C with 12 h light/dark for eight days. Subsequently, a spore suspension was prepared with 0.1 % Tween 20 solution by counting spores in a Neubauer chamber until the concentration was adjusted to 1x105 spores/mL.
2.4.1. Radial Growth Kinetics
The radial growth of the fungus was evaluated in Petri dishes with Czapek agar mixed with the eight treatments: agar control Czapek (Cz), commercial fungicide positive control Innovator® (Inn) (active ingredients: 2- (Ticianometry) benzothiazole and methylbisthiocyanate), acetone 35 % (Ace), chitosan (CS) (4 y 2 mg/mL), ExJ (4 y 2 mg/mL), ExB (4 y 2 mg/mL), ExJ-CS, and ExB-CS. Once the agar was solidified, 1x105 spores/mL were deposited in the center of the plate and incubated at 27 ± 2°C. Measurements were made every 24 h for 7 days, the time it took for growth to reach the edge of the plate in the Cz control. The results were reported in millimeters (mm) and each test was performed in triplicate. To calculate the percentage inhibition, the following formula was used:
where Rc is the radial growth of the Czapek control and Rt is the radial growth of the treatment.
2.4.2. Cell Viability Test
It was performed using XTT salt (Sigma-Aldrich). The tetrazolium salt is negatively charged turns orange when reduced to a soluble formazan dye and the amount of reduced XTT reflects cellular metabolic activity. A 96-well microplate was used for the assay and divided into eight groups, one per treatment, each with three wells. Every well was added 100 μL of inoculum containing 4x106 spores/mL and incubated for four h at 27 ± 2 °C. Then 100 μL of the treatment was added to the respective wells and incubated for four hours. Then 50 μL of XTT solution and 7 μL of menadione was deposited to each and incubated for a further three h. Finally, the absorbance of each well was read at a wavelength of 450 nm in an ELISA spectrophotometer (Modelo iMark, BIO RAD) [17].
2.4.3. Cell Integrity Damage Analysis
A 96-well flat-bottom microplate was used. 100 μL of inoculum (4x106 spores/mL) was added in each well and incubated four h at 27 ± 2 °C. Then 100 μL of the different treatments were added (control Cz, CS at 4 mg/mL, ExB and ExJ at 2 mg/mL) in each well, maintaining the same incubation conditions for 24 h. Subsequently, 5 μL of propidium iodide (PI, 10 μM) was added in each well and incubated 5 h. Then, in each well, the effect of the treatments on the permeability of the fungal membrane was observed with an inverted epifluorescence microscope (model DMi8; Leica Microsystems, Wetzlar, Germany) equipped with fluorescence filter (546/10 excitation filter and 585/40 emission), DFC 450C cooled camera (Leica) and fluorescence overlay software (LAS AF version 3.1.0).
2.5. Phytotoxicity Bioassay
It was evaluated through germination and growth of L. sativa seeds, eight groups of nine-centimeter glass Petri dishes with filter paper at the bottom were used for this purpose. Each was added 2.5 mL of a treatment and 20 seeds and had a water blank control at pH 6 ± 0.3. They were placed in a germinator at 25 ºC, 95% relative humidity and 12 h light/darkness for 120 h. Germinated, non-germinated and abnormal seeds were counted and radicle length was measured. The representative parameters of toxicity were estimated according to García et al. [18]:
where Ri is the average root length of the treatment and Rc the root length of the control. SGi is the number of seeds germinated in the treatment and SGc the number of seeds germinated in the control.
2.6. Acute Toxicity Test in Artemia salina
250 mL of sterile seawater and A. salina eggs were placed in two Erlenmeyer flasks adapted with an aeration system and artificial illumination and incubated for 24 h at 25 ºC for nauplii hatching [17]. Ten of the hatched nauplii were placed in test tubes with 5 mL of sterile seawater and each treated separately, left for 24 h under illumination, the number of survivors in each was counted and reported as percentage survival.
2.7. Statistical Analysis
A completely randomized design was used, and an analysis of variance was performed on the experimental data with a significance level of α = 0.05. Tukey’s multiple range test was performed for comparison of homogeneous groups at a confidence interval of 95 % using the JMP 5.0 software. Results are reported as means ± standard deviation.
3. Results
3.1. Evaluation of Antifungal Activity
3.1.1. Radial Growth Kinetics
One of the most important parameters in the study of filamentous fungi is their mycelial growth. Before mixing the chitosan (CS) with the extracts (ExB and ExJ), an analysis was performed to determine the most effective concentrations of both CS and ExB and ExJ. Table 1 shows that between the CS solution of 4 mg/mL and 2 mg/mL there was a significant difference (P<0.05) in mycelial growth, since the higher the concentration, the greater the inhibition of radial growth. With respect to ExJ and ExB, both extracts at the two concentrations retarded mycelial growth with respect to the Cz control and there was only significant difference (P<0.05) between the type of plant (Table 1). Chávez-Magdaleno et al. [17] reported that 1 % medium molecular weight CS inhibited mycelial growth by 21 % in two species of Colletotrichum close to that obtained in this study. On the contrary, Gálvez-Marroquín et al. [19] observed total inhibition of Colletotrichum spp. with 2.5% low molecular weight CS, which is different from that of our study. Aranaz et al. [11] mentioned that the antimicrobial activity of CS varies depending on the type of microorganism, degree of deacetylation, molecular weight, concentration and exposure time, among others. In addition, the low molecular weight has greater activity as it can penetrate more easily into the interior of the cell. After analyzing the results of radial growth with CS and extracts from both plants, it was decided to use the 4 mg/mL concentration in the subsequent assays.
Figure 1 shows the mycelial development of C. siamense in the different treatments. The control Inn (Innovator®) was shown to be 100% efficient, since the fungus did not develop. It was observed that ExB alone and combined with CS retarded growth by 100 % during the first 72 h. Buitimea-Cantúa et al. [16] found antifungal activity in fractions of B. glutinosa and J. macrocarpa and indicate that they act as enzyme inhibitors in fungi, deforming their cell wall. The behavior of the fungus with CS was as expected, since this compound and its derivatives cause leakage of intracellular contents and inhibit microbial growth by affecting endogenous chitinase activity [10,20]. In addition, Nascimento et al. [14], reported that CS caused, among other damages, spore abnormalities, as observed in our study. In the same vein, Rosas-Burgos et al. [15] reported that ExB inhibited by 60 % the growth of A. flavus, A. parasiticus and F. verticillioides after 14 days of incubation. On the other hand, in the case of J. macrocarpa there are not many reports. García-Sosa et al. [21] mentioned that sakurasosaponin is one of the main metabolites responsible for the antifungal activity in J. flammea as well as the jacquinonic acid detected in other species of Jacquinia.
3.1.2. Cell Viability Test
Radial growth indicated that C. siamense upon being in contact with the treatments adapted and developed slowly compared to the control Cz. Therefore, a viability assay was performed to evaluate the effect of the treatments at the cellular level. The percentage of viable spores in ExJ-CS, ExB and ExB-CS was of 69, 64 and 67 %, respectively. Meanwhile with CS it was 50% (Figure 2). No difference was found between extracts, only with respect to CS (P<0.05). With ExJ it was not possible to quantify it due to interference from the pigments of this plant, which generated intense green tones. The latter was reported by Amiel-Pérez et al. [22] by evaluating plant extracts with this same technique and concluded that methanol solubilizes pigments that are left as residues and affect the reading. There are no other studies reporting these observations, possibly due to the nature of the compounds present in this plant, even though cell viability with XTT has been applied in several fungal species.
3.1.3. Cell Integrity Damage Analysis
The possible damage to the cell membrane of C. siamense was confirmed by microscopic observation in the presence of propidium iodide, which is a nuclear fluorescence marker that intercalates with the double-stranded DNA to form fluorescent complexes. It was observed that both the samples from CS and ExJ treatments showed a higher proportion of damaged cells with respect to the Cz control (Figure 3), which indicates membrane permeability. Furthermore, this is in agreement with the cell proliferation inhibition data found with the XTT method. It may also be related to what was found by Buitimea-Cantúa et al. [16] that these extracts adversely affect chitinase and β,1-3 glucanase enzymes involved in the formation of the cell wall and hyphae in filamentous fungi. Damage to the spore membrane can lead to malfunctioning of organelle-selective properties such as barrier defense, intracellular material flow, among others.
3.2. Phytotoxicity Bioassay
Phytotoxicity is a parameter that indicates possible damage to plant models by determining alteration in the seed and changes in the structure from root elongation. Figure 4 shows that the treatments affected the percentage of germination of the seeds with respect to the control (P<0.05) presenting alterations in the elongation of the roots in the germinated seeds. Rootlets are the first part of the seedling to emerge from the seed during germination and serve for water and nutrient absorption. Germination index in the treatments were low compared to the control (P<0.05), which may be due to the presence of phenolic compounds in the extracts. Dias et al. [23] reported that ethanolic extracts at 10 % of Baccharis dentata, B. uncinella and B. anomala inhibieron about 90 % of the germination rate of L. sativa. It was observed that CS presented the highest germination percentage, possibly due to its growth-promoting properties, which is consistent with Nurliana et al. [24], who reported that CS (0.2 g/L) reduced the stress and promoted the growth of L. sativa in drought conditions. Hernández-Téllez et al. [25] germinated seeds of L. sativa with CS nanoparticles and CS-capsaicin, both at 0.1 mg/mL, achieving only 12 % germination. This low germination was related to the size and structure of the nanoparticles that penetrated the plant cells and induced some kind of stress. The low values in our study indicate that the extracts of both plants have phytotoxic effect on seedling development.
3.3. Acute Toxicity Test in Artemia salina
Survival was 39, 28, 38, 27 and 100% in treatments with ExJ, ExJ-CS, ExB, ExB-CS and CS, respectively. No difference was found between them only with respect to CS (P>0.05) (Figure 5). This, in addition to the effect of the plant components, could also be attributed to a variability in the pH of the water salinity due the nature of the methanolic extracts, since A. salina requires a pH of 7.0 to 8.5 for optimal survival [26]. In a study by Bhoopathy et al. [27], the toxicity of CS nanoparticles on A. salina was evaluated, reporting survival rates of less than 50%. They suggest that the lethality and growth of the Artemia spp. are influenced by the molar mass of the CS, which is consistent with the difference between percentages obtained in both studies.
4. Conclusions
Significant fungistatic activity of these mixtures was found, demonstrating their potential as antifungals against C. siamense as well as to alter its cell viability. Detection of mycelial growth retardation may lead to the development of in vivo treatments to delay the onset of anthracnose. The results also indicate that the extracts of B. glutinosa and J. macrocarpa affect fungal cell integrity, induce death of A. salina and have a phytotoxic effect on lettuce seedling development during germination.
Author Contributions
Conceptualization, A.K.L.-M. and M.O.C.-R.; methodology, A.C.G.-I.; R. I. S.-M, and S.E.M.-S.; investigation, S.d.G.-d.; supervision, M.P.-J. and A.K.L.-M.; writing—original draft preparation, S.d.G.-d; writing—review and editing, A.K.L.-M.; M.O.C.-R. and M.P.-J.; project administration, M.P.-J. and A.K.L.-M.; funding acquisition, M.O.C.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Facultad Interdisciplinaria de Ciencias Biológicas y de Salud of the Universidad de Sonora (project USO313007895). Cortez-Rocha MO has received research support from the Universidad de Sonora.
Institutional Review Board Statement
“Not applicable” for studies not involving humans or animals.
Data Availability Statement
Data can be requested from the corresponding authors.
Acknowledgments
The authors thank to Eber Addí Quintana Obregón PhD of the Centro de Investigación en Alimentación y Desarrollo (CIAD, A.C., Hermosillo) for the donation of the strain of Colletotrichum siamense (H6-1).
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Khan, R.A.; Najeeb, S.; Hussain, S.; Li, Y. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef] [PubMed]
- Mahadevakumar, S.; Sridhar, K.R. Diversity of Pathogenic Fungi in Agricultural Crops. In Plant, Soil and Microbes in Tropical Ecosystems, 1st ed.; Dubey, S.K., Verma, S.K., Eds.; Springer: Singapore, 2021; pp. 101–149. [Google Scholar]
- Rojo-Báez, I.; Álvarez, B.; García, R.S.; León, J.; Sañudo, A.; Allende, R. Situación actual de Colletotrichums spp. en México: Taxonomía, caracterización, patogénesis y control. Rev Mex Fitopatol 2017, 35, 549–570. [Google Scholar] [CrossRef]
- Fuentes-Aragón, D.; Silva-Rojas, H.V.; Guarnaccia, V.; Mora-Aguilera, J.A.; Aranda-Ocampo, S.; Bautista-Martínez, N.; Téliz-Ortíz, D. Colletotrichum species causing anthracnose on avocado fruit in Mexico: Current status. Plant Pathol 2020, 69, 1513–1528. [Google Scholar] [CrossRef]
- García-Estrada, R.S.; Cruz-Lachica, I.; Osuna-García, L.A.; Márquez-Zequera, I. First Report of Papaya (Carica papaya) anthracnose caused by Colletotrichum plurivorum in Mexico. Plant Dis 2020, 104, 589–589. [Google Scholar] [CrossRef]
- Trinidad-Ángel, E.; Ascencio, F.; Ulloa, J.A.; Ramírez, J.C.; Ragazzo, J.A.; Calderón, M.; Bautista, P.U. Identificación y caracterización de Colletotrichum spp. causante de antracnosis en aguacate de Nayarit, México. Rev Mex Cienc Agric 2017, 8, 3953–3964. [Google Scholar] [CrossRef]
- Ciofini, A.; Negrini, F.; Baroncelli, R.; Baraldi, E. Management of post-harvest anthracnose: current approaches and future perspectives. Plants 2022, 11, 1856. [Google Scholar] [CrossRef]
- Azeem, S.; Agha, S.; Jamil, N.; Tabassum, B.; Ahmed, S.; Raheem, A.; Khan, A. Characterization and survival of broad-spectrum biocontrol agents against phytopathogenic fungi. Rev Argent Microbiol 2022, 54, 233–242. [Google Scholar] [CrossRef]
- El Khetabi, A.; Lahlali, R.; Ezrari, S.; Radouane, N.; Lyousfi, N.; Banani, H.; Askarne, L.; Tahiri, A.; El Ghadraoui, L.; Belmalha, S.; Ait Barka, E. Role of plant extracts and essential oils in fighting against postharvest fruit pathogens and extending fruit shelf life: A review. Trends Food Sci Technol 2022, 120, 402–417. [Google Scholar] [CrossRef]
- Poznanski, P.; Hameed, A.; Orczyk, W. Chitosan and chitosan nanoparticles: Parameters enhancing antifungal activity. Molecules 2023, 28, 2996. [Google Scholar] [CrossRef]
- Aranaz, I.; Alcántara, A.R.; Civera, M.C.; Arias, C.; Elorza, B.; Heras-Caballero, A.; Acosta, N. Chitosan: An overview of its properties and applications. Polymers 2021, 13, 3256. [Google Scholar] [CrossRef]
- Pellis, A.; Guebitz, G.M.; Nyanhongo, G.S. Chitosan: Sources, processing and modification techniques. Gels 2022, 8, 393. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bi, X.; Dai, Y.; Ren, R. Enhancing mango anthracnose control and quality maintenance through chitosan and iturin A coating. LWT 2024, 198, 115955. [Google Scholar] [CrossRef]
- Nascimento, J.I.G.; Stamford, T.C.M.; Melo, N.F.C.B.; Nunes, I.D.S.; Lima, M.A.B.; Pintado, M.M.E.; Stamford-Arnaud, T.M.; Stamford, N.P.; Stamford, T.L.M. Chitosan–citric acid edible coating to control Colletotrichum gloeosporioides and maintain quality parameters of fresh-cut guava. Int J Biol Macromol 2020, 163, 1127–1135. [Google Scholar] [CrossRef]
- Rosas-Burgos, E.C.; Cortez-Rocha, M.O.; Cinco-Moroyoqui, F.J.; Robles-Zepeda, R.E.; López-Cervantes, J.; Sánchez-Machado, D.I.; Lares-Villa, F. Antifungal activity in vitro of Baccharis glutinosa and Ambrosia confertiflora extracts on Aspergillus flavus, Aspergillus parasiticus and Fusarium verticillioides. World J Microbiol Biotechnol 2009, 25, 2257–2261. [Google Scholar] [CrossRef]
- Buitimea-Cantúa, G.V.; Rosas-Burgos, E.C.; Cinco-Moroyoqui, F.J.; Burgos-Hernández, A.; Plascencia-Jatomea, M.; Cortez-Rocha, M.O.; Gálvez-Ruiz, J.C. In vitro effect of antifungal fractions from the plants Baccharis glutinosa and Jacquinia macrocarpa on chitin and β-1,3-glucan hydrolysis of maize phytopathogenic fungi and on the fungal β-1,3-glucanase and chitinase activities. J Food Saf 2013, 33, 526–535. [Google Scholar] [CrossRef]
- Chávez-Magdaleno, M.E., Luque-Alcaraz, A.G., Gutiérrez-Martínez, P., Cortez-Rocha, M.O., Burgos-Hernández, A., Lizardi-Mendoza, J., Plascencia-Jatomea, M. Effect of chitosan-pepper tree (Schinus molle) essential oil biocomposites on the growth kinetics, viability and membrane integrity of Colletotrichum gloeosporioides. Rev Mex Ing Quim 2018, 17, 29–45. [CrossRef]
- García, A.; Espinosa, R.; Delgado, L.; Casals, E.; González, E.; Puntes, V.; Barata, C.; Font, X.; Sánchez, A. Acute toxicity of cerium oxide, titanium oxide and iron oxide nanoparticles using standardized tests. Desalination 2011, 269, 136–141. [Google Scholar] [CrossRef]
- Gálvez-Marroquín, L.A.; Martínez-Bolaños, M.; Cruz-Chávez, M.A.; Ariza-Flores, R.; Cruz-López, J.A.; Magaña-Lira, N.; Cruz-de-la-Cruz, L.L.; Ariza-Hernández, F.J. Inhibition of mycelial growth and conidium germination of Colletotrichum sp. for organic and inorganic products. Agro Prod 2022, 15, 25–32. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Asselin, A. Antifungal activity of chitosan on two postharvest pathogens of strawberry fruits. Phytopathology 1992, 82, 398–402. [Google Scholar] [CrossRef]
- García-Sosa, K.; Sánchez-Medina, A.; Álvarez, S.L.; Zacchino, S.; Veitch, N.C.; Simá-Polanco, P.; Peña-Rodríguez, L.M. Antifungal activity of sakurasosaponin from the root extract of Jacquinia flammea. Nat Prod Res 2011, 25, 1185–1189. [Google Scholar] [CrossRef]
- Amiel-Pérez, J.; Fukusaki, A.; Enciso, N.; Altamirano, C.; Mayanga-Herrera, A.; Marcelo, Á.; Marin-Sánchez, O. Interferencia de pigmentos vegetales al aplicar la técnica XTT a extractos de Buddleja globosa, Senecio tephrosiodes Turcz y Equisetum giganteum. Científica 2016, 13, 9–29. [Google Scholar]
- Dias, M.P.; Nozari, R.M.; Santarém, E.R. Herbicidal activity of natural compounds from Baccharis spp. on the germination and seedlings growth of Lactuca sativa and Bidens pilosa. Allelopathy J 2017, 42, 21–36. [Google Scholar] [CrossRef]
- Nurliana, S.; Fachriza, S.; Hemelda, N.M. Chitosan application for maintaining the growth of lettuce (Lactuca sativa) under drought condition. In Proceedings of the 4th International Conference on Food and Agriculture (ICOFA), Jember, Indonesia, 6–7 November 2021; IOP Conf Ser: Earth Environ. Sci. 2022, 980, 012013. [CrossRef]
- Hernández-Téllez, C.N.; Luque-Alcaraz, A.G.; Núñez-Mexía, S.A.; Cortez-Rocha, M.O.; Lizardi-Mendoza, J.; Rosas-Burgos, E.C.; Rosas-Durazo, A.; Parra-Vergara, N.V.; Plascencia-Jatomea, M. Relationship between the antifungal activity of chitosan–capsaicin nanoparticles and the oxidative stress response on Aspergillus parasiticus. Polymers 2022, 14, 2774. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cortés, A.; Almendariz-Tapia, F.; Gómez-Álvarez, A.; Burgos-Hernández, A.; Luque-Alcaraz, A.; Rodríguez-Félix, F.; Plascencia-Jatomea, M. Toxicological assessment of cross-linked beads of chitosan-alginate and Aspergillus australensis biomass, with efficiency as biosorbent for copper removal. Polymers 2019, 11, 222–239. [Google Scholar] [CrossRef]
- Bhoopathy, S.; Inbakandan, D.; Thirugnanasambandam, R.; Kumar, C.; Sampath, P.; Bethunaickan, R.; Raguraman, V.; Vijayakumar, G.K. A comparative study on chitosan nanoparticle synthesis methodologies for application in aquaculture through toxicity studies. IET Nanobiotechnol 2021, 15, 418–426. [Google Scholar] [CrossRef]
Figure 1.
Kinetics of C. siamense radial growth in Czapek agar added with CS, ExB, ExB-CS, ExJ and ExJ-CS incubated at 27 ± 2°C. Controls: Cz=Czapek agar, Inn= Fungicide, Ace=Acetone.
Figure 1.
Kinetics of C. siamense radial growth in Czapek agar added with CS, ExB, ExB-CS, ExJ and ExJ-CS incubated at 27 ± 2°C. Controls: Cz=Czapek agar, Inn= Fungicide, Ace=Acetone.
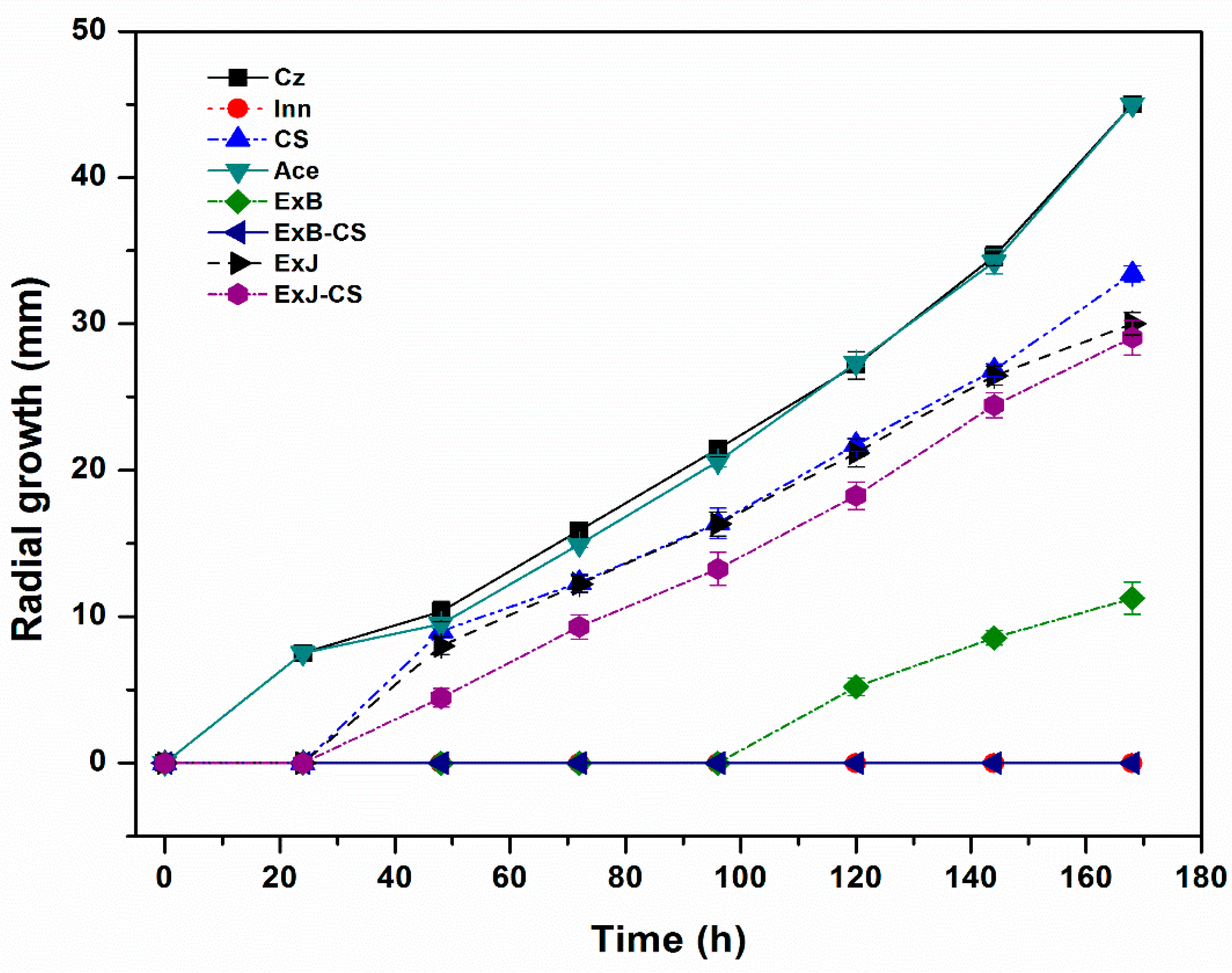
Figure 2.
Cell viability of C. siamense exposed at different treatments incubated at 27 ± 2°C. The bars represent means (n=3) ± standard deviation. Different letter represent a significant difference (P<0.05) between the treatments and control Czapek.
Figure 2.
Cell viability of C. siamense exposed at different treatments incubated at 27 ± 2°C. The bars represent means (n=3) ± standard deviation. Different letter represent a significant difference (P<0.05) between the treatments and control Czapek.
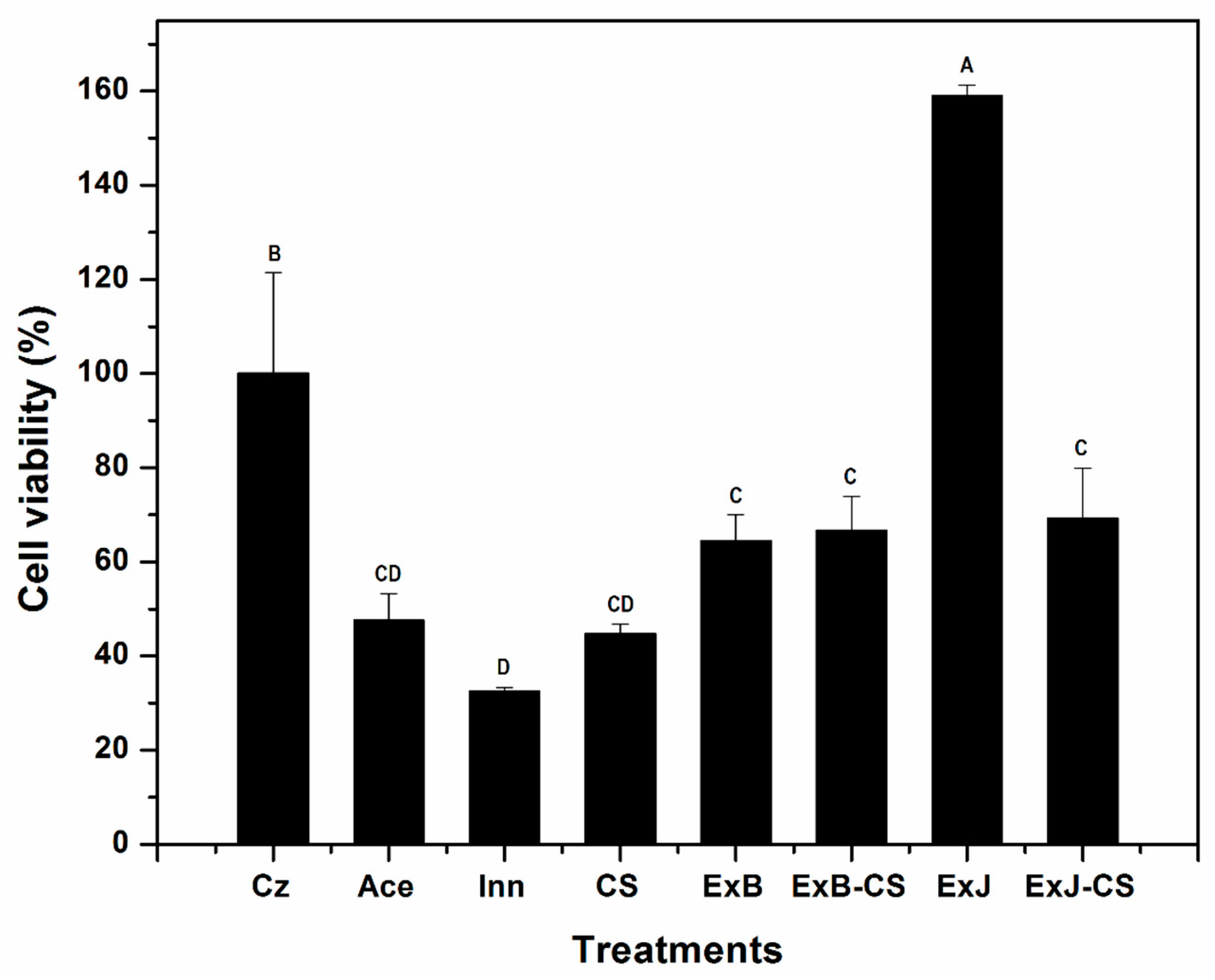
Figure 3.
Propidium iodide staining (Fluorescence microscopy) showing the effect of treatments on membrane permeability of C. siamense: A: Czapek agar, B: CS, C: ExB-CS, D: ExJ-CS.
Figure 3.
Propidium iodide staining (Fluorescence microscopy) showing the effect of treatments on membrane permeability of C. siamense: A: Czapek agar, B: CS, C: ExB-CS, D: ExJ-CS.
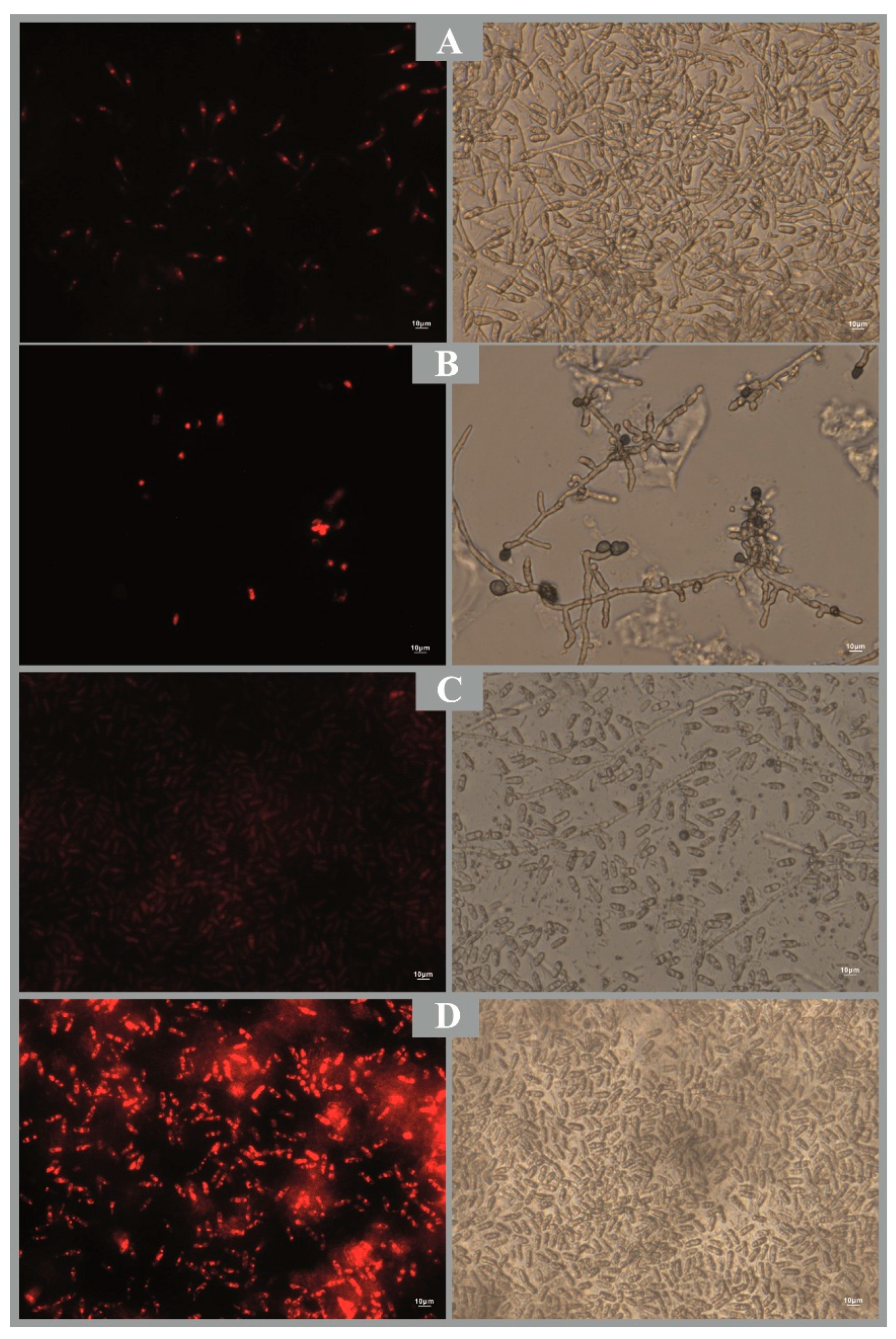
Figure 4.
Phytotoxicity effect in L. sativa seeds (percentage of germination and germination index). The bars represent means (n=3) ± standard deviation. Different capital letters indicate a significant difference in germination index and lower-case letters indicate a significant difference in percentage of germination between treatments (P<0.05).
Figure 4.
Phytotoxicity effect in L. sativa seeds (percentage of germination and germination index). The bars represent means (n=3) ± standard deviation. Different capital letters indicate a significant difference in germination index and lower-case letters indicate a significant difference in percentage of germination between treatments (P<0.05).
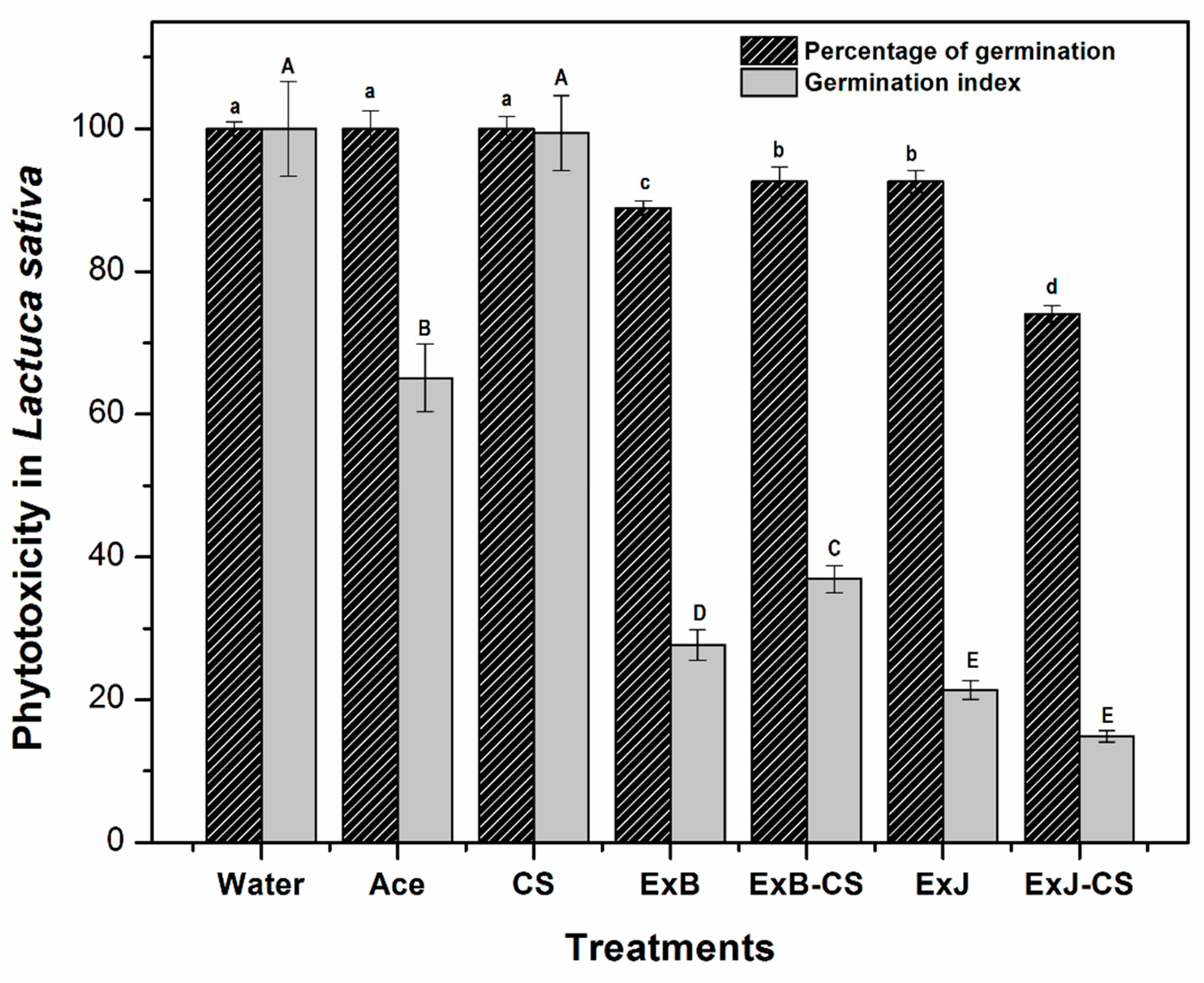
Figure 5.
Survival of Artemia salina exposed to different treatments. The bars represent means (n=3) ± standard deviation. Different letter represent a significant difference (P<0.05) between the treatments and control seawater.
Figure 5.
Survival of Artemia salina exposed to different treatments. The bars represent means (n=3) ± standard deviation. Different letter represent a significant difference (P<0.05) between the treatments and control seawater.
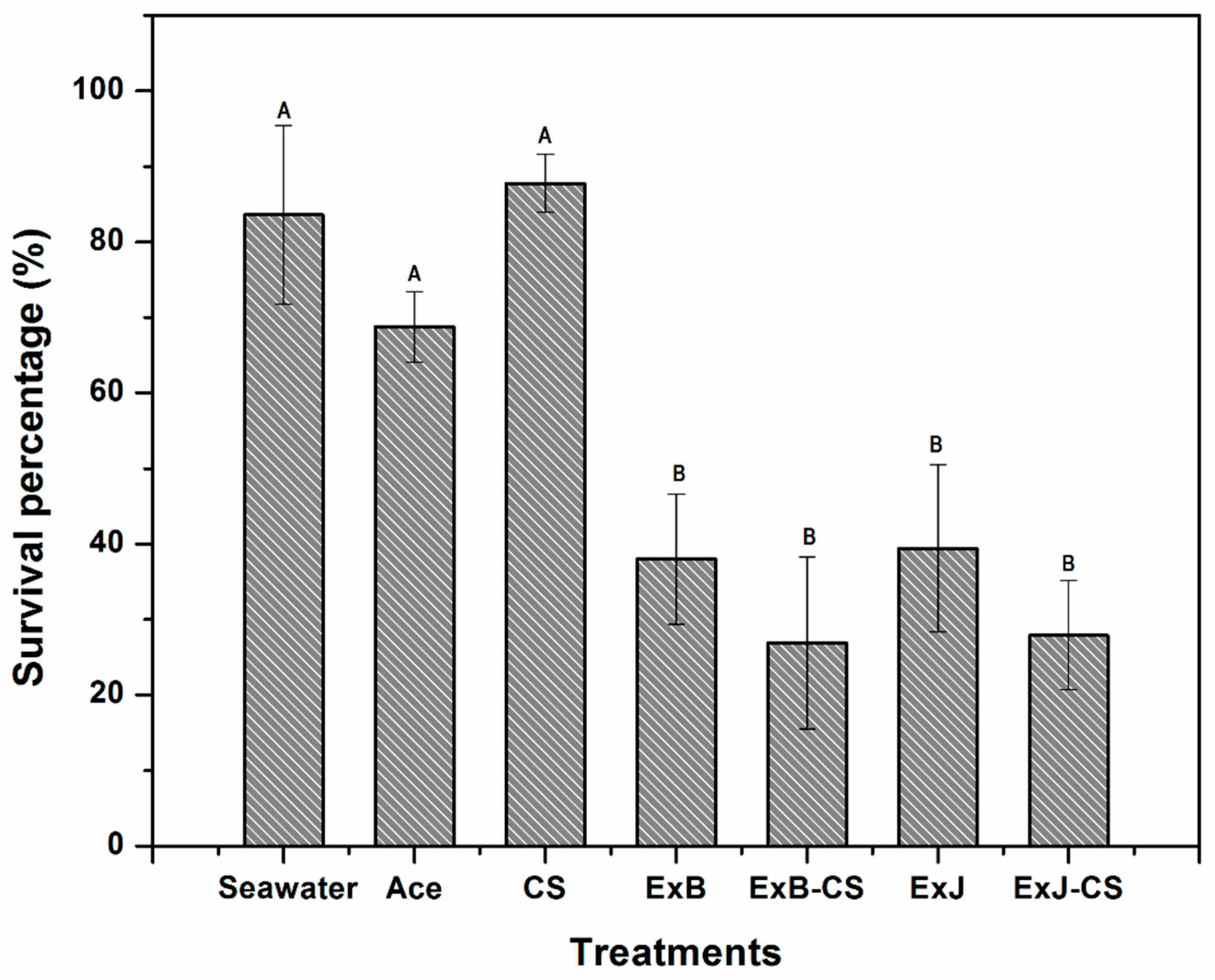

Table 1.
Radial growth inhibition of C. siamense inoculated in Czapek agar added with CS, ExB and ExJ incubated at 27 ± 2°C.
Table 1.
Radial growth inhibition of C. siamense inoculated in Czapek agar added with CS, ExB and ExJ incubated at 27 ± 2°C.
|
Time (h) |
Chitosan (CS) | B. glutinosa (ExB) | J. macrocarpa (ExJ) | |||
| 2 mg/mL | 4 mg/mL | 2 mg/mL | 4 mg/mL | 2 mg/mL | 4 mg/mL | |
| 24 | 100 ± 0.0a | 100 ± 0.0a | 100 ± 0.0a | 100 ± 0.0a | 100 ± 0.0a | 100 ± 0.0a |
| 48 | 60.9 ± 2.7b | 62.4 ± 2.3b | 100 ± 0.0a | 100 ± 0.0a | 43.7 ± 2.3c | 42.1 ± 2.7c |
| 72 | 29.5 ± 0.8d | 47.0 ± 3.7b | 79.6 ± 2.5a | 81.1 ± 1.5a | 45.1 ± 5.9bc | 38.3 ± 1.7c |
| 96 | 26.6 ± 1.4d | 46.3 ± 1.8b | 71.8 ± 0.7a | 72.6 ± 1.8a | 42.3 ± 6.0bc | 33.8 ± 5.6cd |
| 120 | 22.4 ± 1.5d | 42.5 ± 2.6Cb | 63.3 ± 1.5a | 67.2 ± 2.3a | 41.8 ± 3.9b | 33.7 ± 2.6c |
| 144 | 25.7 ± 1.0d | 39.8 ± 11.2c | 62.5 ± 0.8a | 66.8 ± 2.5a | 45.4 ± 1.2b | 38.2 ± 2.3c |
| 168 | 27.6 ± 0.0f | 38.6 ± 0.9e | 63.5 ± 0.6b | 67.6 ± 0.4a | 47.4 ± 0.6c | 43.3 ± 0.4d |
Values are means (n=3) ± standard deviation. The means followed by different letter in the same row are significantly different according to Tukey’s multiple comparison tests (P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



