Submitted:
21 February 2025
Posted:
21 February 2025
You are already at the latest version
Abstract
Long non-coding RNAs (lncRNAs) are a class of RNAs exceeding 200 nucleotides in length, lacking the ability to be translated into proteins. Over the past few decades, an increasing number of publications have established lncRNAs as potent regulators in a broad spectrum of diseases. They modulate the expression of critical genes by affecting transcription, post-transcription, translation, and protein modification. This regulation frequently involves interaction of lncRNAs with other different molecules, such as proteins, RNA and DNA. In recent years, lncRNAs have been identified as regulators of both innate and adaptive immune responses, playing significant roles in the onset and progression of diseases. Additionally, lncRNAs hold potential as biomarkers or therapeutic targets for numerous immune-related diseases. lncRNA nuclear enriched abundant transcript 1 (NEAT1) is a notable example. This review consolidates the latest findings about the role of lncRNA NEAT1 in immune cell function and immune-related diseases.
Keywords:
Long non-coding RNAs
; NEAT1
; Immune cells
; Immune related diseases
; Biomarker
; Therapeutic target
1. Introduction
1.1. Overview of long non-coding RNAs (lncRNAs)
Long non-coding RNAs (lncRNAs) are a group of RNAs longer than 200 nucleotides. LncRNAs lack the capability to translate into full proteins. However, a subset of lncRNAs can translate into small peptides, which have been proven to have biological functions [1,2,3]. Over the last few decades, mounting evidence has demonstrated the role of lncRNAs in various cell processes, including cell proliferation, suppression of apoptosis, invasion, and metastasis [4,5,6,7]. LncRNAs can be classified as antisense, divergent, and intergenic lncRNAs, based on the direction of their transcription and their proximity to neighboring genes [8]. The number of identified lncRNAs has dramatically increased in recent decades. For instance, approximately 95,243 lncRNA genes and 323,950 lncRNA transcripts have been identified in humans from a study in 2023 [9]. Advances in new sequencing techniques and new bioinformatics tools will likely reveal even more lncRNAs [10]. The prevalence of a large amount of lncRNAs underscores their importances in biology and disease. It is also important to note that the expression patterns of lncRNAs exhibits greater tissue specificity and cell-type selectivity compared to coding genes, suggesting their roles in a cell-type-specific manner [11].
The innate and adaptive immune responses are critical for defending the body against pathogens. The innate immune response is the first line of defense and activates adaptive immune responses, leading to the elimination of pathogens. The innate immune response involves immune cells, including macrophages, neutrophils, dendritic cells, natural killer (NK) cells, mast cells, eosinophils, basophils. The adaptive immune response involves immune cells, including B cells, T cells. Within the immune system, various types of immune cells working together to trigger an immune response. Mounting evidence has begun to appreciate the role of lncRNAs in immune cells, including their development, differentiation, activation, and effector functions. Therefore, there are new opportunities to target these critical lncRNAs to modulate immune responses and suppress immune-mediated pathogens, paving the way for innovative therapeutic strategies. Extensive reviews have summarized the role of lncRNAs in immune systems under both physiological and disease conditions [12,13,14]. In this review, we aim to provide a comprehensive overview of one lncRNA involved in multiple immune cell types: Nuclear Enriched Abundant Transcript 1 (NEAT1). (Figure 1). By examining its functions in immune cell regulation and potential as a biomarker and therapeutic target, we can gain insights into its role in immune-related diseases.
1.2. Structure and Function of NEAT1
NEAT1 is conserved across different species and is highly expressed and specifically localized in paraspeckles. Additionally, NEAT1 is essential for the paraspeckle complex [15,16,17]. NEAT1 has two isoforms, NEAT1_1 and NEAT1_2, which are produced through alternative 3’-end processing (Figure 2). The two isoforms share a common promoter, therefore NEAT1_1, the short isoform, completely overlaps with the 5’ end of the longer isoform, NEAT1_2. NEAT1_1 is generated through termination by a polyadenylation signal (PAS) during transcription [18]. In humans, NEAT_1 is approximately 3.7 kilobase (kb) long, and NEAT1_2 is about 23 kb. NEAT1_1 is more abundant than NEAT1_2 in cells. Studies have shown that the two isoforms are localized in diverse structures in the nucleus [18,19,20]. NEAT1_2 is exclusively expressed in nuclear paraspeckles. NEAT1_1 can localize to other structures, such as microspeckles [19]. Furthermore, the two isoforms have diverse functions in biology. NEAT1_2 is essential for paraspeckle assembly and can regulate RNA transcription through regulating the paraspeckles [16,21]. In this review, we will introduce several studies which demonstrate that NEAT1_2 can modulate gene expression through regulating transcription at the targeted site in the paraspeckles. In contrary, NEAT1_1 is nonessential for paraspeckle assembly. NEAT1_1 regulates the expression of the targeted genes by interacting with other proteins in the cells. For instance, It has been found that NEAT1_1 promotes sarcoma metastasis by interacting with a RNA splicing protein, KT-type splicing regulatory protein (KHSRP) [7]. There are other studies further supporting the evidence that two isoforms play a diverse role in the cells. One study showed the relative abundance of NEAT1_1 and NEAT1_2 differs between aggressive and non-aggressive neuroblastoma cancer cells. For instance, high-risk MYCN-amplified aggressive cancer cells express high levels of NEAT1_1 relative to NEAT1_2, while non-aggressive cancer cells with a more differentiated phenotype express high levels of NEAT1_2. They further found that NEAT1_1 acts as an oncogene and NEAT1_2 as a tumor suppressor in neuroblastoma [22]. However, it is noteworthy that the unique feature that NEAT1_1 overlapping with NEAT1_2 makes it difficult to dissect the function of individual isoforms. Therefore, most studies, including those cited in this review, are focused on dissecting the functions of NEAT1 without properly modulating the expression of individual isoforms. Several strategies have been developed to modulate the expression of individual isoforms of NEAT1 for studying the function of individual isoform in biology [19,22,23,24]. Therefore, it is important to carefully choose the right method for exploring which isoform of NEAT1 is important for the observed phenotype.
2. NEAT1 in immune cell function
2.1. NEAT1 regulates innate immune cell
Innate immunity is the first line of defense against invading pathogens, such as virus and microbes. Here, we review on the publications showing the role of NEAT1 in innate immune cell regulation. Hantaan virus (HTNV), commonly found in Asia, is the leading cause of severe hemorrhagic fever with renal syndrome (HFRS), resulting in a high mortality rate. Macrophages are critical for the host innate immune system, serving as the first line of defense against HTNV infection. NEAT1 is upregulated in monocytes and macrophages in mice and human at the early HTNV infection. Additionally, the expression of NEAT1 in patient monocytes is negative correlated with HTNV load and disease progression [25]. In mouse bone marrow-derived macrophages co-cultured with HTNV, knockdown of Neat1 inhibits the activation of inflammatory macrophages, thereby facilitating HTNV infection. Conversely, overexpression of the long isoform of Neat1, Neat1_2, significantly represses viral replication in the same co-culture experiment. Mechanistic study demonstrate that Neat1_2 may upregulate the expression of sterol regulatory element-binding protein 2 (Srebp2). Subsequently, Neat1_2 interacts with Srebp2 to stimulate the activation of inflammatory macrophages, thereby inhibiting HTNV infection [25]. Another study showed that NEAT1 is also significantly upregulated in human umbilical vein endothelial cell (HUVEC) infected with HTNV [26]. Again, they found that NEAT1_2, but not NEAT1_1, is responsible for defending HTNV infection. Depletion of NEAT1_2 in HUVEC dramatically promoted HTNV nucleocapsid protein (NP) production. Transient ectopic expression of NEAT1_2 repressed HTNV NP production, thereby efficiently suppressing HTNV replication. Mechanistic studies showed that NEAT1 relocates splicing factor proline/glutamine-rich (SFPQ) to paraspeckles to derepress its transcriptional inhibitory effect on retinoic acid-inducible gene I (RIG-I) and dexd/h-box helicase 60 (DDX60), thereby modulating the innate immune response against HTNV infection. Interestingly, the mechanism by which NEAT1 can activate transcription by relocating SFPQ has been demonstrated in an earlier study by Imamura K et al [21]. In this study, they found that influenza virus infection, herpes simplex virus infection, or Toll-like receptor 3-p38 pathway-triggered poly I:C stimulation can induce the expression of NEAT1 in cultured stable cell lines. They further determined that upregulated NEAT1_2, but not NEAT1_1, relocated SFPQ to paraspeckles to derepress the expression of interleukin-8 (IL8) gene, resulting in an innate immune response. These studies suggest that the long isoform of NEAT1, NEAT1_2, is critical for defending the virus infection through paraspeckles in an innate immune response.
Morchikh M et al identified another mechanism by which NEAT1 regulates the innate immune response [27]. They showed that NEAT1 is required for the assembly of the HEXIM1-DNA-PK-paraspeckle components-ribonucleoprotein (HDP-RNP) complex, which includes hexamethylene bisacetamide inducible 1 (HEXIM1), the DNA-dependent protein kinase (DNA-PK) complex, SFPQ, and paraspeckle component 1 (PSPC1). Knockdown of HDP-RNP subunits has no impact on poly I:C stimulation but resulted in the loss of interferon (IFN) stimulatory DNA (ISD)-mediated IFNα, IFNβ, and MX dynamin like GTPase 1 (MXA) induction. They further showed that the HDP-RNP complex is a positive regulator of DNA-mediated IFN response gene induction. Cyclic GMP-AMP synthase (cGAS), polyglutamine binding protein 1 (PQBP1), stimulator of interferon genes (STING), interferon regulatory factor 3 (IRF3), the mediators of innate immune activation, all interact with the HDP-RNP complex. The innate immune response caused by ISD can lead to the remodeling of the HDP-RNP complex and activation of both DNA-PKs and IRF3. Using Kaposi’s sarcoma-associated herpesvirus (KSHV) infection of HUVECs as a model, they demonstrated that the HDP-RNP complex is essential for cGAS-dependent KSHV-mediated activation of the innate immune response. It is noteworthy that this study did not show which isoform of NEAT1 is responsible for interacting with the HDP-RNP complex and subsequent function.
The inflammasome plays a critical role in the innate immune response. Inflammasomes are multicomponent signaling platforms that control inflammatory responses. For instance, pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) activate one or more innate pattern recognition receptors (PRRs). Once activated, the sensor proteins bind to and induce the oligomerization of an adaptor protein, apoptosis-associated speck-like protein containing CARD (ASC), resulting in the formation of a single macromolecular aggregate, the ASC speck. Oligomerized ASC then recruits pro-caspase-1, which auto-processes to generate its activated form. Activated caspase-1 induces proteolytic maturation of cytokine interleukin-1β (IL-1β) and interleukin-18 (IL-18), thereby inducing pyroptosis. Inflammasome activation is potent for defending pathogens and damaged cells. However, dysregulated inflammasome activity can lead to autoimmune diseases, cancer, neurodegenerative disorders.
In a mouse model of peritonitis and pneumonia, Zhang P et al showed that Neat1 can interact with multiple proteins involved in inflammasomes to enhance their assembly, resulting in pro-caspase-1 processing [28]. For instance, knockdown of Neat1 in mouse immortalized bone marrow-derived macrophages (iBMDMs) resulted in a significantly reduction in the oligomerization of ASC, indicating impaired assembly of the inflammasome. Conversely, ectopic expression of Neat1 promoted the formation of the NLR family pyrin domain containing 3 (NLRP3) inflammasome, causing enhanced caspase-1 activation and IL-1β maturation. Mechanistically, Neat1 can enhance the activation of NLR family CARD domain containing 4 (NLRC4) and absent in melanoma 2 (AIM2) inflammasomes in an interleukin-6 (IL-6)-independent manner. Once stimulated with inflammasome-activating signals, Neat1 translocates from nuclear paraspeckles to the cytoplasm to modulate inflammasome activation by stabilizing mature caspase-1, promoting IL-1β production and pyroptosis. Lin J et al showed that nickel-cobalt alloy magnetic nanocrystals (NiCo NCs) can inhibit the activation of NLRP3, NLRC4 and AIM2 inflammasomes [29]. Additionally, NiCo NCs inhibited neutrophil recruitment in a mouse model of acute peritonitis and reduced symptoms in another mouse model of colitis. Furthermore, NiCo NCs can reduce the expression of NEAT1, although the study did not show the functional role of NEAT1 in NiCo NCs-mediated inhibition of inflammasomes. It is also important to note that these studies did not specify which isoform of NEAT1 is responsible for regulation the inflammasomes.
Other immune-related diseases involved in the innate immune response are also related to dysregulated expression of NEAT1. Fibrosis is a life-threatening disorder associated with tissue dysfunction caused by excessive deposition of extracellular matrix. Fibrosis is irreversible, and there is no effective treatment for it. Crosstalk between immune cells and other cells may play a critical role in the development of fibrosis. Deletion of RNA-binding motif protein 7 (Rbm7) in nonhematopoietic cells significantly repressed fibrosis in mice. NEAT11 interacts with RBM7. Furthermore, Rbm7 promotes the development of fibrosis by regulating cell death through inducing the dissociation of accumulated Neat1 paraspeckles, thereby preventing the repair of irreversibly damaged cells in a mouse model. Additionally, in vivo delivery of Neat1 siRNA significantly induces fibrosis in a mouse model of Rbm7 knockout. Therefore, this study suggests a functional role of Neat1 in controlling fibrosis [30]. Though the study has not specified which isoform of Neat1 is critical for regulating fibrosis, it may indicate that NEAT1_2 may play a more important role as the paraspeckle is involved in the function.
Several studies have indicated that NEAT1 may play a role in certain immune-related diseases, but no careful mechanistic study has been involved in these publications. For instance, Neat1 expression is significantly upregulated in the serum of mouse with inflammatory bowel disease (IBD). Knockdown of Neat1 reduced the intestinal epithelial barrier and promoted macrophage M1 transformation to M2, thereby reducing the inflammatory reaction [31]. NEAT1 is significantly downregulated in the peripheral blood mononuclear cells (PMBCs), mainly in cluster of differentiation 14 positive (CD14+) monocytes, from early-onset myocardial infarction (MI) patients [32]. Hepatitis B virus (HBV) infection remains a major public health issue, as many people die from HBV-associated liver diseases. Zeng Y et al found that the expression of NEAT1 is significantly downregulated in peripheral blood of patients with chronic HBV infection compared to healthy donors. They suggest that chronic HBV infection (CHB) can repress the innate immune response by downregulating NEAT1 [33]. Neat1 knockout macrophages failed to express a large set of proinflammatory cytokines, chemokines and antimicrobial mediators, leading to the failure of controlling the replication of salmonella enterica serovar Typhimurium or vesicular stomatitis virus [34]. In a mouse model of myocarditis, transfusion with Neat1-low dendritic cells (DCs) reduced the inflammatory response, while in a heart transplantation mode, transfusion with Neat1-low DCs induced immune tolerance. Additional experiment indicates that upregulation of Neat1 can lead to the generation of tolerogenic DCs [35].
Overall, these studies suggest that NEAT1 plays a significant role in regulating the innate immune response. Some studies have showed that the long isoform of NEAT1 plays a significant role in regulating the innate immune response, while many studies did not specify which isoform is critical for defending the disease.
2.2. NEAT1 regulates adaptive immune cell function
Besides the innate immune response, NEAT1 may also regulate the adaptive immune response. NEAT1 expression is significantly upregulated in PBMCs of human immunodeficiency virus 1 (HIV-1) infected patients. The expression of NEAT1 is significantly reduced in HIV-1 infected patients treated with highly active antiretroviral therapy, suggesting that NEAT1 may interact with HIV-1 in vivo [36]. NEAT1 is induced in Jurkat cells infected with HIV-1 [37,38]. Liu H et al found that NEAT1 is significantly decreased following activation of PBMCs and cluster of differentiation 4 positive (CD4+) T cells [39]. Furthermore, HIV-1 replication is enhanced in Jurkat cells with NEAT1 knockout compared to wild-type Jurkat cells, suggesting the potential role of NEAT1 in antiviral responses in adoptive immune cells.
The occurrence and development of rheumatoid arthritis (RA) is mainly caused by T helper 17 (Th17) cells differentiated from CD4+ T cells. NEAT1 is significantly increased in the PBMCs of RA patients and is also significantly upregulated in Th17 cells differentiated from CD4+ T cells in vitro. Knockdown of NEAT1 inhibits CD4+ T cells from differentiating into Th17 cells. NEAT1 upregulates the expression of signal transducer and activator of transcription 3 (STAT3) protein, which is critical for Th17 cell differentiation. In vivo injection of NEAT1 siRNA decreases the number of Th17 cells, thereby reducing arthritis severity in a type II collagen-induced arthritis mouse model [40]. Therefore, this study suggests that NEAT1 is important for CD4+ T cells differentiating into Th17 cells.
Atherosclerosis and myocardial infarction are mainly caused by inflammation. NEAT1 is significantly downregulated in PBMCs of post-MI patients [32]. It is observed that there are more Th cells and T regulatory (Treg) cells in the spleens of Neat1 knockout (Neat1 -/-) mice compared to wild-type mice, suggesting a shift of CD4+ T cell balance towards Th cell proliferation. Additionally, the expression of interferon gamma (IFNγ) is significantly higher in unstimulated splenocytes from Neat1 -/- mice compared with wild-type cells. Therefore, this study suggests that Neat1 is critical for the differentiation and expansion of CD4+ T cells in the adaptive immune system.
Sepsis is a life-threatening disease initiated by cytokine-mediated hyperinflammation. NEAT1 expression is significantly upregulated in the PBMCs of sepsis patients. In a mouse model of sepsis, knockdown of Neat1 accelerates T lymphocyte viability and reduces the apoptosis [41]. Mechanistic studies showed that Neat1 can induce the expression of mast cell-expressed membrane protein 1 (MCEMP1) by sponging microRNA-125 (miR-125) to promote the serum levels of inflammatory factors and T lymphocyte activity and apoptosis, thereby increasing immunity in septic mice.
As a chronic disease, coronary heart disease (CHD) is the most common cardiovascular disease caused by atherosclerosis. The expression of NEAT1 is significantly higher in the plasma of CHD patients compared to normal patients [42]. Additionally, the expression of NEAT1 is positively correlated with tumor necrosis factor alpha (TNFα), IL-1β, IL-6, and interleukin-17 (IL-17) in the plasma of CHD patients. Furthermore, the expression of NEAT1 is also positively correlated with the percentage of Th17 cells, but not T helper 1 (Th1) or T helper 2 (Th2) cells in the plasma of CHD patients, suggesting that NEAT1 may positively regulate CD4+ T cells differentiation into Th17 cells.
As the most abundant allergic disease in the pediatric population, allergic rhinitis (AR) is a hypersensitivity reaction induced by immunoglobulin E (IgG)-mediated inflammatory response, including nasal itching, congestion, rhinorrhea, sneezing. It is reported that the expression of NEAT1 is significantly higher in the PBMCs of AR patients compared to normal patients [43]. Interestingly, the expression of NEAT1 is positively correlated with Th2 cells, but not Th1 cells in AR patients. However, the correlation between NEAT1 and Th17 cells was not examined in the study. Similarly, the expression of NEAT1 is significantly upregulated in CD4+ T cells in peripheral blood of children with asthma [44]. Additionally, knockdown of NEAT1 significantly decreased Th2-related cytokines but had no impact on Th1 cells. These studies suggest that NEAT1 is critical for regulating Th1/Th2 balance in these patients. Another study suggests that NEAT1 could also be critical for regulating Th1/Th2 balance in the patients infected with SARS-CoV-2 (COVID-19) [45].
Overexpression of NEAT1 in CD4+ T cells promotes the levels of Th2-related cytokines interleukin-4 (IL-4), interleukin-5 (IL-5), and interleukin-13 (IL-13) [46]. In contrast, Th1-related cytokine IFN-γ production is reduced by the upregulation of NEAT1 in CD4+ T cells [46]. Mechanistically, NEAT1 promotes signal transducer and activator of transcription 6 (STAT6) protein expression but has no impact on its mRNA level, as NEAT1 interacts with STAT6 protein to prevent its ubiquitination. Therefore, NEAT1 promotes Th2 cell activation through STAT6, supporting the observation that NEAT1 expression is induced in the PBMCs of systemic lupus erythematosus (SLE), which is primarily influenced by a Th2-mediated immune response [46]. In another study, Yan X et al observed significant upregulation of NEAT1 in the CD4+ T cells from PBMCs of children with asthma. Mechanistic studies revealed that knockdown of NEAT1 significantly impaired the production of Th2-related cytokines, thereby inhibiting Th2 differentiation [44]. A shift of CD4+ T cell balance towards T helper cell proliferation was observed both in circulating blood and spleens in Neat1-/- mice compared to Neat1 wild-type mice [32]. This study proposed that NEAT1 is an immunoregulator affecting T cell and monocyte-macrophage lineage differentiation and function.
Autoimmune uveitis (AU) is an immune-related intraocular inflammatory disorder affecting millions of people worldwide. It mainly occurs in young adults and is characterized by inflammatory cell infiltration and ocular damaging, resulting in visual impairment and blindness [47]. Chen S et al identified that NEAT1 is significantly upregulated in the PBMCs of AU patients [48]. In a mouse model of experimental autoimmune uveitis (EAU), they further showed that Neat1 expression is significantly increased in CD4+ T cells in mice with EAU compared to normal mouse. Additionally, Neat1 expression is positively correlated with disease severity. Furthermore, Neat1 is significantly upregulated in Th17 cells compared with Th0 and Th1 cells. They showed that Neat1 induced by interleukin-23 (IL-23)/STAT3 signaling may be critical for the progression of EAU in mice, supported by the evidence that lentivirus-mediated knockdown of Neat1 in mice alleviated EAU compared to control lentivirus-treated mice. They further showed that Neat1 may promote EAU by upregulating the Th17 cell response.
The correlation between NEAT1 expression and B cells is less known. Qiu Y et al conducted single-cell RNA sequencing of CD19+ cells in patients with Waldenström’s macroglobulinemia (WM), a rare lymphoproliferative disorder [49]. They found that NEAT1 expression is significantly upregulated in mature B cells of WM patients. Chattopadhyay P et al found that the expression of NEAT1 is significantly upregulated in the PBMCs of patients infected with COVID-19 [50]. Additionally, they identified that NEAT1 is significantly upregulated in naïve B cells, naïve CD4+ and naïve cluster of differentiation 8 positive (CD8+) T cells in the infected individuals.
Overall, these studies have demonstrated the critical role of NEAT1 in regulating the adaptive immune response, particularly in the differentiation of CD4+ T cells into Th17 cells. However, none of these studies have specified which isoform of NEAT1 is responsible for regulating this differentiation process. Many studies have used Neat1-/- mice, which show significant downregulation of both isoforms of Neat1. The authors who created this knockout mouse have recently generated another mouse that solely expresses the long isoform by using CRISPR/Cas9-mediated deletion of the PAS domain, which is essential for generating the short isoform [23]. This mutant mouse can be used to determine whether the deletion of the short isoform of Neat1 impacts immune cell regulation.
2.3. NEAT1 as a biomarker and therapeutic target in immune-related diseases
As mentioned above, NEAT1 is dysregulated in PBMCs of patients with multiple immune-related diseases. NEAT1 regulates immune cells in response to these diseases. Additionally, NEAT1 can serve as a biomarker for assisting disease management and prognosis. Furthermore, NEAT1 may be a therapeutic target in some immune-related diseases.
Li P et al explored the correlation of NEAT1 with disease severity and recurrence in patients with acute ischemic stroke (AIS) [51]. NEAT1 is significantly upregulated in PBMCs of patients with AIS compared to healthy donors. Additionally, NEAT1 expression positively correlates with the National Institutes of Health Stroke Scale (NIHSS) score. Furthermore, NEAT1 expression is also positive correlated with increased recurrence and death risk, and negatively correlated with recurrence-free survival. Therefore, NEAT1 can serve as a biomarker for AIS. NEAT1 expression is significantly upregulated in sepsis patient samples. There is a significant positive correlation between NEAT1 and the severity of sepsis, indicating that NEAT1 may serve as a potential biomarker for sepsis [52,53,54]. The expression of NEAT1 is significantly high in a subset of PBMCs driven by sepsis. Huang Q et al also showed that NEAT1 expression is significantly upregulated in plasma and serum of sepsis patients. Its expression is significantly associated with higher sepsis risk [53]. Additionally, Higher expression of NEAT1 is associated with good prognosis in sepsis patients. The early diagnosis of behҫet’s disease (BD), a chronic autoimmune disease, is critical to avoid serious and fatal complications. Mohammed SR et al and Mohammed A et al found that NEAT1 expression in serum is significantly correlated with the severity of the disease, suggesting that it can be a biomarker for BD [55,56]. Pediatric immune thrombocytopenic purpura (ITP) is an autoimmune disease. Hamdy SM et al found that NEAT1 expression is significantly upregulated in the sera of children with ITP [57]. Furthermore, they also found that there is a significant upregulation of NEAT1 in non-chronic compared to chronic ITP patients. The platelets under apoptosis that are presented to T lymphocytes with the aid of dendritic cells are important in the pathogenesis of ITP. There is a significant negative correlation between NEAT1 and platelet counts before treatment. Therefore, NEAT1 expression in serum can serve as a potential biomarker for differentiating childhood ITP patients from healthy individuals, in addition to differentiating non-chronic from chronic ITP patients. NEAT1 expression is significantly upregulated in the PBMCs of patients with acute/chronic inflammatory demyelinating polyradiculoneuropathies (AIDP/CIDP) [58]. As the first evidence showing the expression of NEAT1 positively correlates with this disease, further studies are warranted to further elucidate the molecular mechanisms of NEAT1 in AIDP/CIDP. It also suggests that NEAT1 can serve as a biomarker for AIDP/CIDP. Atypical hemolytic uremic syndrome (aHUS) is a rare and fatal thrombotic microangiopathy. However, the biomarker for diagnosing this disease is still unknown. Therefore, Chen I et al conducted single-cell sequencing on PBMCs of aHUS patients and normal controls. They observed significant upregulation of NEAT1 in PBMCs of unstable aHUS patients, providing valuable insight into potential biomarkers for this disease [59]. NEAT1 can potentially be a biomarker for autoimmune disease but also a potential biomarker in subgrouping the disease. For instance, Hamdy SM et al detected that NEAT1 is significantly upregulated in the sera of pediatric immune thrombocytopenic purpura (ITP) patients. Additionally, NEAT1 is further significantly upregulated in the sera of non-chronic in comparison to chronic ITP patients [57]. The expression of NEAT1 is found to be significantly upregulated in the plasma samples of Alzheimer’s disease (AD) patients, suggesting that it can be a promising diagnosis biomarker for this disease [60]. Ballonová L et al showed that NEAT1 is significantly upregulated in monocytes of a group of Hereditary angioedema (HAE) patients. Frailty, an intermediate status of the human aging process, has been correlated with decompensated homeostasis and mortality [61]. Luo OJ et al conducted single-cell analysis in PBMCs from healthy young adults and frail old adults to investigate the immune phenotype of frailty, which is still poorly understood [62]. Interestingly, they identified a specific subset of monocyte exclusively identified in the frailty group. Furthermore, this subset of monocytes exclusively expresses a high level of NEAT1. It is suggested that NEAT1 can also serve as a marker for a subset of monocyte [63]. Multiple studies revealed that NEAT1 expression is highly expressed in PBMCs and saliva of mild and severe patients infected with COVID-19 [64,65,66,67]. Additionally, the expression of NEAT1 positively correlates with the severity of the virus infection risk. Overall, these studies support the evidence that NEAT1 can serve as a biomarker for multiple immune-related diseases.
Upregulated NEAT1 regulates immune response in immune-related diseases, indicating itself as a potential therapeutic target. Inflammatory bowel disease (IBD) is a complex autoimmune disease, including Crohn’s disease and ulcerative colitis. The molecular pathogenesis of IBD remains largely unknown. Liu R et al explored the role of NEAT1 in IBD [31]. Neat1 expression is significantly higher in the serum of IBD mice compared to normal controls. Furthermore, they found that knockdown of Neat1 significantly suppresses the dextran sulfate sodium (DSS)-induced permeability increase in colon tissues from IBD mice, possibly through promoting the transformation of macrophage M1 to M2 and suppressing the inflammatory reaction. Therefore, these results suggest that NEAT1 can be a therapeutic target in IBD. Acute cerebral ischaemia can result in serious consequences, such as uncontrolled reperfusion induced brain injury. Ni X et al dissected the role of NEAT1 in cerebral ischaemia/reperfusion (I/R) injury [68]. NEAT1 expression is significantly upregulated in PBMCs of patients with acute ischaemic stroke (AIS) compared to controls. Knockdown of NEAT1 inhibits apoptosis in N2a cells caused by cerebral I/R injury. Additionally, knockdown of NEAT1 reduces the AKT/STAT3 pathway in BV-2 cells caused by deprivation/reoxygenation (OGD/R) injury. Furthermore, knockdown of NEAT1 suppresses M1 microglial polarization in OGD/R-exposed microglial cells. Therefore, they proposed that NEAT1 can potentially be a therapeutic target in acute cerebral ischaemia. Rheumatoid arthritis (RA) is the most common chronic autoimmune connective tissue disease. NEAT1 is significantly increased in the PBMCs of RA patients. Knock down of NEAT1 inhibits CD4+ T cells from differentiating into Th17 cells by reducing the STAT3 protein. Additionally, In vivo delivery a lentivirus to express a NEAT1 siRNA to target Th17 cells relieves RA in a type II collagen induced mouse model of RA [69]. Another studied confirmed that serum-derived exosomes expressing NEAT1 can promote the occurrence of RA in the same mouse model [40]. Those studies indicate that NEAT1 can be a potential therapeutic target for RA. Ye L et al observed a significant upregulation of NEAT1 in peripheral T cells, including both CD4+ and CD8+ T cells, of patients with primary sjögren’s syndrome (pSS), a systemic autoimmune disease [70]. In addition, the expression of NEAT1 correlates with the severity of pSS. Inhibition of NEAT1 by an antisense increases the expression of C-X-C motif chemokine ligand 8 (CXCL8) and TNF-α in PMA/ionomycin-stimulated Jurkat cells. This study highlights the potential of NEAT1 as a therapeutic target of pSS. Jin F et al identified that NEAT1 is significantly upregulated in the tissues of patients with ischemic stroke [71,72]. Additionally, intraventricular injection of an antisense oligonucleotide to knock down NEAT1 significantly reduced brain damage by reducing activated microglia and proinflammatory cytokines in a mouse model of ischemic stroke, supporting that NEAT1 as a potential therapeutic target for ischemic stroke treatment [71].
Overall, these studies indicate that NEAT1 has the potential to be a therapeutic target for treating multiple immune-related diseases. However, it is important to note that none of these studies have focused on targeting specific isoforms of NEAT1 to modulate these diseases. The antisense oligonucleotide used in the studies targets both isoforms. Therefore, extensive research is needed to determine which isoform should be targeted for therapeutic purposes in future studies.
3. Conclusion
In the past few years, mounting evidence demonstrates the role of lncRNA NEAT1 in immune-related diseases (Table 1), besides cancers. Virus infection directly induced the expression of NEAT1, resulting in immune response to against the virus. Therefore, NEAT1 expression is significantly upregulated in PBMCs and serums of patients infected with various viruses, from COVID-19 [64,65,66,67] to HIV [36,37,38,39]. Additionally, the RNA levels of NEAT1 is often positively correlated with the severity of the infection, such as in sepsis [52,53,54]. All these studies highlight that NEAT1 may serve as a biomarker for virus infection. NEAT1 is also highly expressed in a subset of monocytes, and has been used as biomarkers for this subsets in a single-cell sequencing analysis [63]. NEAT1 can play a role in both innate immunity and adaptive immunity. NEAT1 can induce the activation of inflammasome and activate macrophages. NEAT1 can induce CD4+ T cells to differentiate into Th17 cells and is also critical for regulating Th1/Th2 balance. Furthermore, NEAT1 has a potential as a targeted therapy in multiple preclinical models of immune-related diseases. For instance, knockdown of Neat1 suppresses DSS-induced permeability increase in colon tissues from IBD mice [31]. In vivo delivery a lentivirus expressing Neat1 siRNA relieves RA in a type II collagen-induced mouse model of RA [69]. Intraventricular injection of Neat1 siRNA reduced brain damage in a mouse model of ischemic stroke [71].
Future research on NEAT1 in immune-related disease should focus on dissecting the precise molecular mechanisms by which NEAT1 regulates immune responses and contributes to disease pathogenesis. Investigating the interaction between NEAT1 and different signaling pathways, as well as exploring NEAT1 as a potential biomarker for early detection and therapeutic targeting, could open new avenues for treatment. Additional studies are required to determine which isoform of NEAT1 should be targeted for therapeutic purposes. Several engineering methods have been introduced to modulate the expression of each isoform of NEAT1 in cells [19] and in mice [23]. Those tools should be carefully chosen to determine the role of each NEAT1 isoform in the immune-related diseases and identify the appropriate target. Additionally, advanced techniques like clustered regularly interspaced short palindromic repeats/cas13d (CRISPR/Cas13d) could be potentially employed to modulate the expression of each NEAT1 isoform and assess its effects in preclinical models [73,74]. Overall, a deeper understanding of NEAT1’s functions and regulatory networks may lead to novel therapeutic strategies for managing immune-related diseases.
Author Contributions
J.H., and Z.Z., drafted the first version of the manuscript. All authors edited the manuscript.
Funding
National Institutes of Health 5K22CA248849-03A1.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
This work was supported by the National Cancer institute of the US NIH under award numbers 5 K22 CA248849 (J.H.) and the Providence Portland Medical Foundation. All figure images are created with BioRender.com.
Conflicts of interest: The authors declare no conflict of interest.
References
- Yi, Q.; et al. CircRNA and lncRNA-encoded peptide in diseases, an update review. Mol. Cancer 23, 214 (2024).
- Zhang, Y. LncRNA-encoded peptides in cancer. J. Hematol. Oncol. 17, 66 (2024).
- Tian, H.; Tang, L.; Yang, Z.; Xiang, Y.; Min, Q.; Yin, M.; You, H.; Xiao, Z.; Shen, J. Current understanding of functional peptides encoded by lncRNA in cancer. Cancer Cell Int. 2024, 24, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. <bold>2009</bold>, <italic>136</italic>. [CrossRef]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef]
- He, J.; Tu, C.; Liu, Y. Role of lncRNAs in aging and age-related diseases. Aging Med. 2018, 1, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sachdeva, M.; Xu, E.; Robinson, T.J.; Luo, L.; Ma, Y.; Williams, N.T.; Lopez, O.; Cervia, L.D.; Yuan, F.; et al. The Long Noncoding RNANEAT1Promotes Sarcoma Metastasis by Regulating RNA Splicing Pathways. Mol. Cancer Res. 2020, 18, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.; Feng, C.; Qin, Y.; Xiao, J.; Zhang, Z.; Ma, L. LncBook 2.0: integrating human long non-coding RNAs with multi-omics annotations. Nucleic Acids Res. 2022, 51, D186–D191. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, P.; Kwon, E.; Fitzgerald, K.A.; Weng, Z.; Zhou, C. Flnc: Machine Learning Improves the Identification of Novel Long Noncoding RNAs from Stand-Alone RNA-Seq Data. Non-Coding RNA 2022, 8, 70. [Google Scholar] [CrossRef]
- Sarropoulos, I.; Marin, R.; Cardoso-Moreira, M.; Kaessmann, H. Developmental dynamics of lncRNAs across mammalian organs and species. Nature 2019, 571, 510–514. [Google Scholar] [CrossRef]
- Martinez-Castillo, M. , M Elsayed, A., López-Berestein, G., Amero, P. & Rodríguez-Aguayo, C. An overview of the immune modulatory properties of long non-coding RNAs and their potential use as therapeutic targets in cancer. Noncoding RNA 9, 70 (2023).
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Atianand, M.K.; Caffrey, D.R.; Fitzgerald, K.A. Immunobiology of Long Noncoding RNAs. Annu. Rev. Immunol. 2017, 35, 177–198. [Google Scholar] [CrossRef]
- Chen, L.-L.; Carmichael, G.G. Altered Nuclear Retention of mRNAs Containing Inverted Repeats in Human Embryonic Stem Cells: Functional Role of a Nuclear Noncoding RNA. 2009, 35, 467–478. [CrossRef]
- Clemson, C.M.; Hutchinson, J.N.; Sara, S.A.; Ensminger, A.W.; Fox, A.H.; Chess, A.; Lawrence, J.B. An Architectural Role for a Nuclear Noncoding RNA: NEAT1 RNA Is Essential for the Structure of Paraspeckles. 2009, 33, 717–726. [CrossRef]
- Sasaki, Y. T. F. , Ideue, T., Sano, M., Mituyama, T. & Hirose, T. MENepsilon/beta noncoding RNAs are essential for structural integrity of nuclear paraspeckles. Proc. Natl. Acad. Sci. U. S. A. 106, 2525–2530 (2009).
- Naganuma, T.; Nakagawa, S.; Tanigawa, A.; Sasaki, Y.F.; Goshima, N.; Hirose, T. Alternative 3′-end processing of long noncoding RNA initiates construction of nuclear paraspeckles. EMBO J. 2012, 31, 4020–4034. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Harvey, A.R.; Hodgetts, S.I.; Fox, A.H. Functional dissection of NEAT1 using genome editing reveals substantial localization of the NEAT1_1 isoform outside paraspeckles. RNA 2017, 23, 872–881. [Google Scholar] [CrossRef]
- Yamazaki, T.; Souquere, S.; Chujo, T.; Kobelke, S.; Chong, Y.S.; Fox, A.H.; Bond, C.S.; Nakagawa, S.; Pierron, G.; Hirose, T. Functional Domains of NEAT1 Architectural lncRNA Induce Paraspeckle Assembly through Phase Separation. Mol. Cell 2018, 70, 1038–1053.e7. [Google Scholar] [CrossRef]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. Long Noncoding RNA NEAT1-Dependent SFPQ Relocation from Promoter Region to Paraspeckle Mediates IL8 Expression upon Immune Stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Naveed, A.; et al. NEAT1 polyA-modulating antisense oligonucleotides reveal opposing functions for both long non-coding RNA isoforms in neuroblastoma. Cell. Mol. Life Sci. 78, 2213–2230 (2021).
- Isobe, M.; Toya, H.; Mito, M.; Chiba, T.; Asahara, H.; Hirose, T.; Nakagawa, S. Forced isoform switching of Neat1_1 to Neat1_2 leads to the loss of Neat1_1 and the hyperformation of paraspeckles but does not affect the development and growth of mice. RNA 2019, 26, 251–264. [Google Scholar] [CrossRef]
- Naveed, A. et al. PolyA-modulating antisense oligonucleotides reveal opposing functions for long non-coding RNA NEAT1 isoforms in neuroblastoma. bioRxiv (2020). [CrossRef]
- Yang, Y.; Li, M.; Ma, Y.; Ye, W.; Si, Y.; Zheng, X.; Liu, H.; Cheng, L.; Zhang, L.; Zhang, H.; et al. LncRNA NEAT1 Potentiates SREBP2 Activity to Promote Inflammatory Macrophage Activation and Limit Hantaan Virus Propagation. Front. Microbiol. 2022, 13, 849020. [Google Scholar] [CrossRef]
- Ma, H.; Han, P.; Ye, W.; Chen, H.; Zheng, X.; Cheng, L.; Zhang, L.; Yu, L.; Wu, X.; Xu, Z.; et al. The Long Noncoding RNA NEAT1 Exerts Antihantaviral Effects by Acting as Positive Feedback for RIG-I Signaling. J. Virol. 2017, 91, e02250–16. [Google Scholar] [CrossRef]
- Morchikh, M.; Cribier, A.; Raffel, R.; Amraoui, S.; Cau, J.; Severac, D.; Dubois, E.; Schwartz, O.; Bennasser, Y.; Benkirane, M. HEXIM1 and NEAT1 Long Non-coding RNA Form a Multi-subunit Complex that Regulates DNA-Mediated Innate Immune Response. Mol. Cell 2017, 67, 387–399.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Cao, L.; Zhou, R.; Yang, X.; Wu, M. The lncRNA Neat1 promotes activation of inflammasomes in macrophages. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Lin, J.; Dong, L.; Liu, Y.-M.; Hu, Y.; Jiang, C.; Liu, K.; Liu, L.; Song, Y.-H.; Sun, M.; Xiang, X.-C.; et al. Nickle-cobalt alloy nanocrystals inhibit activation of inflammasomes. Natl. Sci. Rev. 2023, 10, nwad179. [Google Scholar] [CrossRef]
- Fukushima, K.; Satoh, T.; Sugihara, F.; Sato, Y.; Okamoto, T.; Mitsui, Y.; Yoshio, S.; Li, S.; Nojima, S.; Motooka, D.; et al. Dysregulated Expression of the Nuclear Exosome Targeting Complex Component Rbm7 in Nonhematopoietic Cells Licenses the Development of Fibrosis. Immunity 2020, 52, 542–556.e13. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Tang, A.; Wang, X.; Chen, X.; Zhao, L.; Xiao, Z.; Shen, S. Inhibition of lncRNA NEAT1 suppresses the inflammatory response in IBD by modulating the intestinal epithelial barrier and by exosome-mediated polarization of macrophages. Int. J. Mol. Med. 2018, 42, 2903–2913. [Google Scholar] [CrossRef]
- Gast, M.; Rauch, B.H.; Haghikia, A.; Nakagawa, S.; Haas, J.; Stroux, A.; Schmidt, D.; Schumann, P.; Weiss, S.; Jensen, L.; et al. Long noncoding RNA NEAT1 modulates immune cell functions and is suppressed in early onset myocardial infarction patients. Cardiovasc. Res. 2019, 115, 1886–1906. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; et al. Toll-like receptors, long non-coding RNA NEAT1, and RIG-I expression are associated with HBeAg-positive chronic hepatitis B patients in the active phase. J. Clin. Lab. Anal. 33, e22886 (2019).
- Azam, S.; Armijo, K.S.; Weindel, C.G.; Chapman, M.J.; Devigne, A.; Nakagawa, S.; Hirose, T.; Carpenter, S.; Watson, R.O.; Patrick, K.L. The early macrophage response to pathogens requires dynamic regulation of the nuclear paraspeckle. Proc. Natl. Acad. Sci. 2024, 121. [Google Scholar] [CrossRef]
- Zhang, M.; Zheng, Y.; Sun, Y.; Li, S.; Chen, L.; Jin, X.; Hou, X.; Liu, X.; Chen, Q.; Li, J.; et al. Knockdown of NEAT1 induces tolerogenic phenotype in dendritic cells by inhibiting activation of NLRP3 inflammasome. Theranostics 2019, 9, 3425–3442. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Peng, X.; Xie, T.; Lu, X.; Liu, F.; Wu, H.; Yang, Z.; Wang, J.; Cheng, L.; Wu, N. Detection of the long noncoding RNAs nuclear-enriched autosomal transcript 1 (NEAT1) and metastasis associated lung adenocarcinoma transcript 1 in the peripheral blood of HIV-1-infected patients. HIV Med. 2015, 17, 68–72. [Google Scholar] [CrossRef]
- Imam, H.; Bano, A.S.; Patel, P.; Holla, P.; Jameel, S. The lncRNA NRON modulates HIV-1 replication in a NFAT-dependent manner and is differentially regulated by early and late viral proteins. Sci. Rep. 2015, 5, srep08639. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, C.-Y.; Yedavalli, V.S.R.K.; Jeang, K.-T. NEAT1 Long Noncoding RNA and Paraspeckle Bodies Modulate HIV-1 Posttranscriptional Expression. mBio 2013, 4, e00596–12. [Google Scholar] [CrossRef]
- Liu, H.; Hu, P.-W.; Couturier, J.; Lewis, D.E.; Rice, A.P. HIV-1 replication in CD4+ T cells exploits the down-regulation of antiviral NEAT1 long non-coding RNAs following T cell activation. Virology 2018, 522, 193–198. [Google Scholar] [CrossRef]
- Shui, X.; et al. Knockdown of lncRNA NEAT1 inhibits Th17/CD4+ T cell differentiation through reducing the STAT3 protein level. J. Cell. Physiol. 234, 22477–22484 (2019).
- Chen, J.; Xu, X.; Zhang, S. Silence of long noncoding RNA NEAT1 exerts suppressive effects on immunity during sepsis by promoting microRNA-125-dependent MCEMP1 downregulation. IUBMB Life 2019, 71, 956–968. [Google Scholar] [CrossRef]
- Zhu, L.; Lin, X.; Chen, M. LncRNA NEAT1 correlates with Th17 cells and proinflammatory cytokines, also reflects stenosis degree and cholesterol level in coronary heart disease patients. J. Clin. Lab. Anal. 2022, 36, e23975. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhao, S.; Huang, W.; Huang, L.; Huang, M.; Luo, X.; Chang, S. Aberrant expressions of circulating lncRNA NEAT1 and microRNA-125a are linked with Th2 cells and symptom severity in pediatric allergic rhinitis. J. Clin. Lab. Anal. 2022, 36, e24235. [Google Scholar] [CrossRef] [PubMed]
- Yan, X. , Liu, H. & Li, T. Lncrna NEAT1 regulates Th1/Th2 in pediatric asthma by targeting MicroRNA-217/GATA3. Iran. J. Public Health 52, 106–117 (2023).
- Liu, X.; et al. The promotion of humoral immune responses in humans via SOCS1-mediated Th2-bias following SARS-CoV-2 vaccination. Vaccines (Basel) 11, 1730 (2023).
- Huang, S.; Dong, D.; Zhang, Y.; Chen, Z.; Geng, J.; Zhao, Y. Long non-coding RNA nuclear paraspeckle assembly transcript 1 promotes activation of T helper 2 cells via inhibiting STAT6 ubiquitination. Hum. Cell 2021, 34, 800–807. [Google Scholar] [CrossRef] [PubMed]
- de Smet, M. D.; et al. Understanding uveitis: the impact of research on visual outcomes. Prog. Retin. Eye Res. 30, 452–470 (2011).
- Chen, S.; Wang, J.; Zhang, K.; Ma, B.; Li, X.; Wei, R.; Nian, H. LncRNA Neat1 targets NonO and miR-128-3p to promote antigen-specific Th17 cell responses and autoimmune inflammation. Cell Death Dis. 2023, 14, 1–11. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, X.-S.; Yao, Y.; Si, Y.-M.; Wang, X.-Z.; Jia, M.-N.; Zhou, D.-B.; Yu, J.; Cao, X.-X.; Li, J. Single-cell transcriptome analysis reveals stem cell-like subsets in the progression of Waldenström’s macroglobulinemia. Exp. Hematol. Oncol. 2023, 12, 1–6. [Google Scholar] [CrossRef]
- Chattopadhyay, P.; Mehta, P.; Soni, J.; Tardalkar, K.; Joshi, M.; Pandey, R. Cell-specific housekeeping role of lncRNAs in COVID-19-infected and recovered patients. NAR Genom. Bioinform. 2024, 6, lqae023. [Google Scholar] [CrossRef]
- Li, P. , Duan, S. & Fu, A. Long noncoding RNA NEAT1 correlates with higher disease risk, worse disease condition, decreased miR-124 and miR-125a and predicts poor recurrence-free survival of acute ischemic stroke. J. Clin. Lab. Anal. 34, e23056 (2020).
- Shen, J.; Pan, L.; Chen, W.; Wu, Y. Long non-coding RNAs MALAT1, NEAT1 and DSCR4 can be serum biomarkers in predicting urosepsis occurrence and reflect disease severity. Exp. Ther. Med. 2024, 28, 1–12. [Google Scholar] [CrossRef]
- Huang, Q.; Huang, C.; Luo, Y.; He, F.; Zhang, R. Circulating lncRNA NEAT1 correlates with increased risk, elevated severity and unfavorable prognosis in sepsis patients. 2018, 36, 1659–1663. [CrossRef]
- Wen, M.; Cai, G.; Ye, J.; Liu, X.; Ding, H.; Zeng, H. Single-cell transcriptomics reveals the alteration of peripheral blood mononuclear cells driven by sepsis. Ann. Transl. Med. 2020, 8, 125–125. [Google Scholar] [CrossRef]
- Mohammed, S.R.; Abdelaleem, O.O.; Ahmed, F.A.; Abdelaziz, A.A.; Hussein, H.A.; Eid, H.M.; Kamal, M.; Ezzat, M.A.; Ali, M.A. Expression of lncRNAs NEAT1 and lnc-DC in Serum From Patients With Behçet’s Disease Can Be Used as Predictors of Disease. Front. Mol. Biosci. 2022, 8, 797689. [Google Scholar] [CrossRef]
- Mohammed, A.; et al. Association of long non-coding RNAs NEAT1, and MALAT1 expression and pathogenesis of Behçet’s disease among Egyptian patients. Saudi J. Biol. Sci. 29, 103344 (2022).
- Hamdy, S.M.; Ali, M.S.; El-Hmid, R.G.A.; Abdelghaffar, N.K.; Abdelaleem, O.O. Role of Long non Coding RNAs, NEAT1 and Lnc-DC Expression in Pediatric Immune Thrombocytopenic Purpura. Rep. Biochem. Mol. Biol. 2023, 11, 635–643. [Google Scholar] [CrossRef]
- Sadeghpour, S.; Ghafouri-Fard, S.; Mazdeh, M.; Nicknafs, F.; Nazer, N.; Sayad, A.; Taheri, M. Over-Expression of Immune-Related lncRNAs in Inflammatory Demyelinating Polyradiculoneuropathies. J. Mol. Neurosci. 2020, 71, 991–998. [Google Scholar] [CrossRef]
- Chen, I.-R.; Huang, C.-C.; Tu, S.-J.; Wang, G.-J.; Lai, P.-C.; Lee, Y.-T.; Yen, J.-C.; Chang, Y.-S.; Chang, J.-G. Dysregulation of Immune Cell Subpopulations in Atypical Hemolytic Uremic Syndrome. Int. J. Mol. Sci. 2023, 24, 10007. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. et al. NEAT1 involves Alzheimer’s Disease (AD) progression via regulation of glycolysis and P-tau. bioRxiv (2019). [CrossRef]
- Ballonová, L.; Souček, P.; Slanina, P.; Réblová, K.; Zapletal, O.; Vlková, M.; Hakl, R.; Bíly, V.; Grombiříková, H.; Svobodová, E.; et al. Myeloid lineage cells evince distinct steady-state level of certain gene groups in dependence on hereditary angioedema severity. Front. Genet. 2023, 14, 1123914. [Google Scholar] [CrossRef]
- Luo, O.J.; Lei, W.; Zhu, G.; Ren, Z.; Xu, Y.; Xiao, C.; Zhang, H.; Cai, J.; Luo, Z.; Gao, L.; et al. Multidimensional single-cell analysis of human peripheral blood reveals characteristic features of the immune system landscape in aging and frailty. Nat. Aging 2022, 2, 348–364. [Google Scholar] [CrossRef]
- Zhu, Z.; Huang, J.; Zhang, Y.; Hou, W.; Chen, F.; Mo, Y.-Y.; Zhang, Z. Landscape of tumoral ecosystem for enhanced anti-PD-1 immunotherapy by gut Akkermansia muciniphila. Cell Rep. 2024, 43, 114306. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Wang, C.; Vagts, C.; Raguveer, V.; Finn, P.W.; Perkins, D.L. Long non-coding RNAs (lncRNAs) NEAT1 and MALAT1 are differentially expressed in severe COVID-19 patients: An integrated single-cell analysis. PLOS ONE 2022, 17, e0261242. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Adamoski, D.; Genelhould, G.; Zhen, F.; Yamaguto, G.E.; Araujo-Souza, P.S.; Nogueira, M.B.; Raboni, S.M.; Bonatto, A.C.; Gradia, D.F.; et al. NEAT1 and MALAT1 are highly expressed in saliva and nasopharyngeal swab samples of COVID-19 patients. Mol. Oral Microbiol. 2021, 36, 291–294. [Google Scholar] [CrossRef]
- Rahni, Z.; et al. Long non-coding RNAs ANRIL, THRIL, and NEAT1 as potential circulating biomarkers of SARS-CoV-2 infection and disease severity. Virus Res. 336, 199214 (2023).
- Ali, M. A.; et al. Peripheral lncRNA NEAT-1, miR374b-5p, and IL6 panel to guide in COVID-19 patients’ diagnosis and prognosis. PLoS One 19, e0313042 (2024).
- Ni, X.; Su, Q.; Xia, W.; Zhang, Y.; Jia, K.; Su, Z.; Li, G. Knockdown lncRNA NEAT1 regulates the activation of microglia and reduces AKT signaling and neuronal apoptosis after cerebral ischemic reperfusion. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Liu, R.; Jiang, C.; Li, J.; Li, X.; Zhao, L.; Yun, H.; Xu, W.; Fan, W.; Liu, Q.; Dong, H. Serum-derived exosomes containing NEAT1 promote the occurrence of rheumatoid arthritis through regulation of miR-144-3p/ROCK2 axis. Ther. Adv. Chronic Dis. 2021, 12. [Google Scholar] [CrossRef]
- Ye, L.; Shi, H.; Yu, C.; Fu, J.; Chen, C.; Wu, S.; Zhan, T.; Wang, B.; Zheng, L. LncRNA Neat1 positively regulates MAPK signaling and is involved in the pathogenesis of Sjögren's syndrome. Int. Immunopharmacol. 2020, 88, 106992. [Google Scholar] [CrossRef]
- Jin, F.; Ou, W.; Wei, B.; Fan, H.; Wei, C.; Fang, D.; Li, G.; Liu, W.; Liu, J.; Jin, L.; et al. Transcriptome-Wide Analysis to Identify the Inflammatory Role of lncRNA Neat1 in Experimental Ischemic Stroke. J. Inflamm. Res. 2021, ume 14, 2667–2680. [Google Scholar] [CrossRef]
- Li, S.; et al. Construction of lncRNA-mediated ceRNA network for investigating immune pathogenesis of ischemic stroke. Mol. Neurobiol. 58, 4758–4769 (2021).
- Montero, J.J.; Trozzo, R.; Sugden, M.; Öllinger, R.; Belka, A.; Zhigalova, E.; Waetzig, P.; Engleitner, T.; Schmidt-Supprian, M.; Saur, D.; et al. Genome-scale pan-cancer interrogation of lncRNA dependencies using CasRx. Nat. Methods 2024, 21, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Wessels, H.-H.; Méndez-Mancilla, A.; Guo, X.; Legut, M.; Daniloski, Z.; Sanjana, N.E. Massively parallel Cas13 screens reveal principles for guide RNA design. Nat. Biotechnol. 2020, 38, 722–727. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The role of NEAT1 in various immune cells. “Ref” stands for reference.
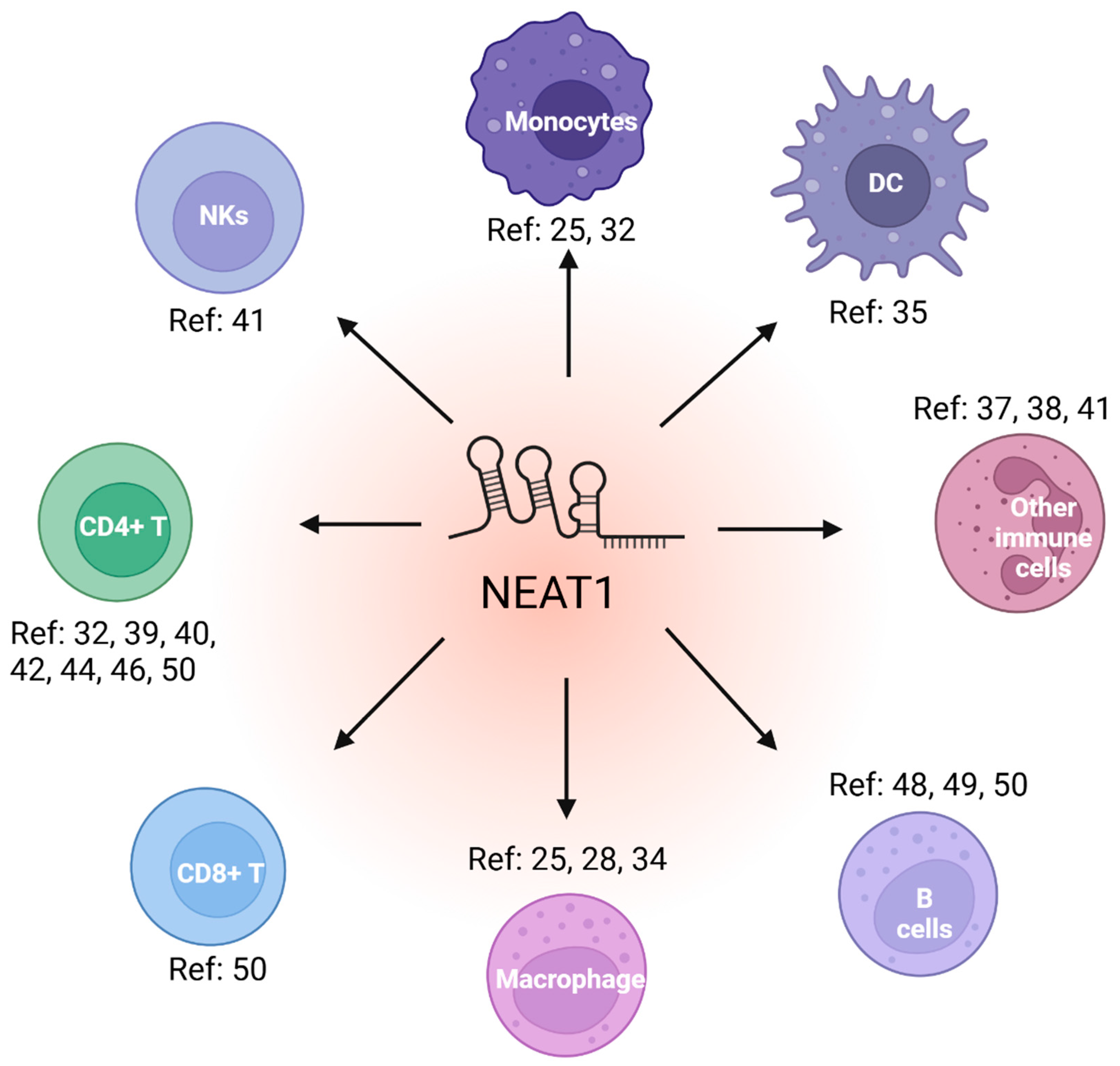
Figure 2.
The structural feature and cellular functions of NEAT1.
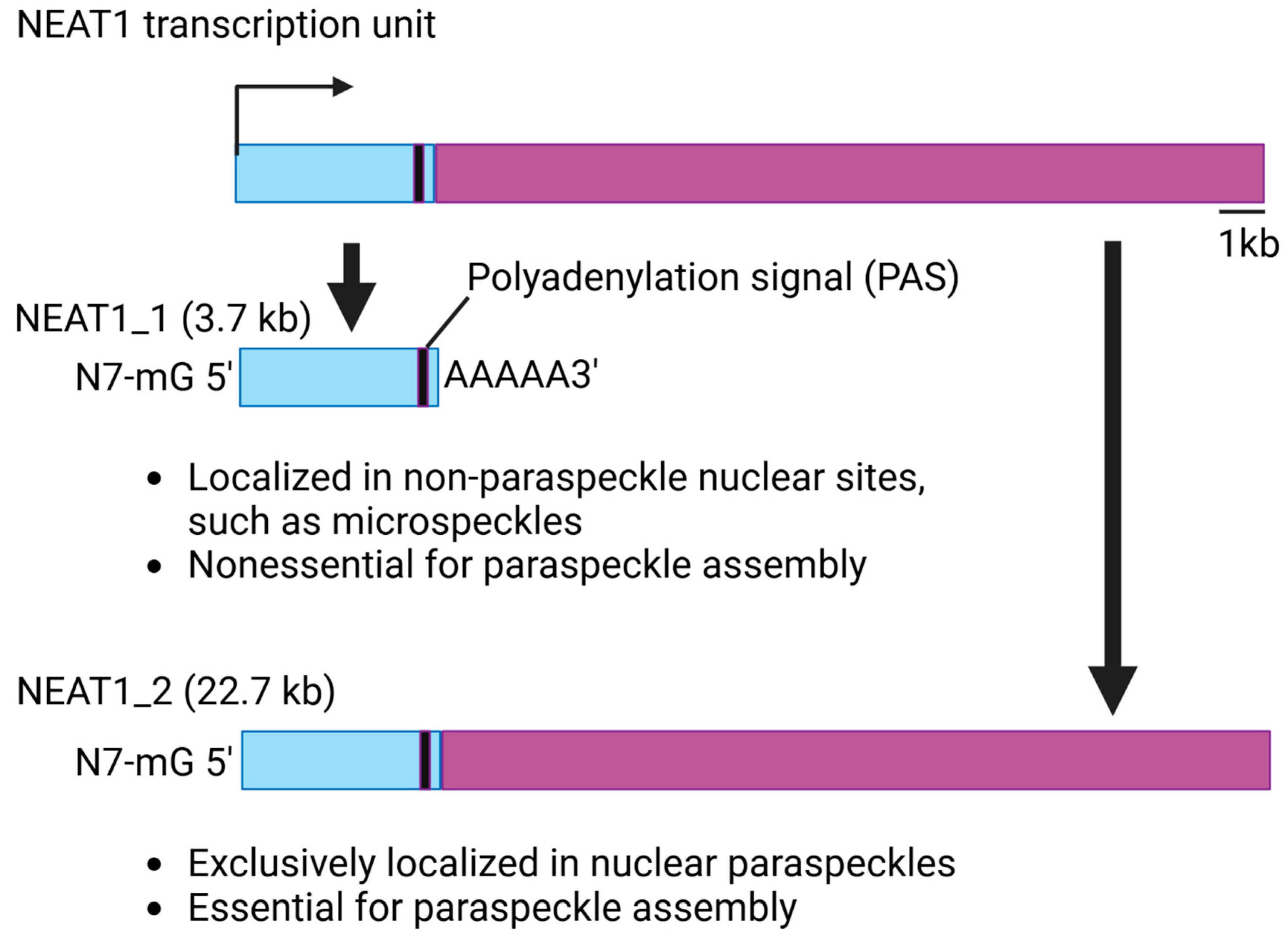

Table 1.
The overview of NEAT1 in different immune-related diseases.
| Disease type | NEAT1 expression | Mechanisms | References (Ref) |
| Hantaan virus (HTNV) | Upregulated | NEAT1 activates inflammatory macrophages through Srebp2 | Ref: [25] |
| Influenza | Upregulated | NEAT1 induces transcription of IL8 | Ref: [21] |
| Kaposi’s sarcoma-associated herpesvirus (KSHV) | N/A | NEAT1 activates the innate immune response through HDP-RNP complex | Ref: [27] |
| Peritonitis and pneumonia | N/A | NEAT1 activates NLRP3, NLRC4 and AIM2 inflammasomes | Ref: [28] |
| Fibrosis | N/A | NEAT1 represses fibrosis through interacting with Rbm7 | Ref: [30] |
| Inflammatory bowel disease (IBD) | N/A | Knockdown of NEAT1 promotes macrophage M2 | Ref: [31] |
| Atherosclerosis and myocardial infarction (MI) | Downregulated | NEAT1 regulate the generation of tolerogenic DCs and CD4+ T cell balance | Ref: [32,35] |
| Hepatitis B virus (HBV) | Downregulated | N/A | Ref: [33] |
| Human immunodeficiency virus 1 (HIV-1) | Upregulated | N/A | Ref: [36,37,38,39] |
| Rheumatoid arthritis (RA) | Upregulated | NEAT1 promotes CD4+ T cells differentiating into Th17 cells | Ref: [40,69] |
| Sepsis | Upregulated | NEAT1 induces MCEMP1 by sponging miR-125 | Ref: [41,52,53,54] |
| Coronary heart disease (CHD) | Upregulated | NEAT1 promotes CD4+ T cells differentiating into Th17 cells | Ref: [42] |
| Allergic rhinitis (AR) | Upregulated | NEAT1 regulates Th1/Th2 balance | Ref: [43] |
| Systemic lupus erythematosus (SLE) | Upregulated | NEAT1 promotes Th2 cell activation through STAT6 | Ref: [46] |
| Asthma | Upregulated | NEAT1 promotes Th2 cell activation | Ref: [44] |
| Autoimmune uveitis (AU) | Upregulated | NEAT1 promotes CD4+ T cells differentiating into Th17 cells | Ref: [47,48] |
| Waldenström’s macroglobulinemia (WM) | Upregulated | N/A | Ref: [49] |
| SARS-CoV-2 (COVID-19) | Upregulated | N/A | Ref: [50,64,65,66,67] |
| Acute ischemic stroke (AIS) | Upregulated | N/A | Ref: [51,71,72] |
| Behҫet’s disease (BD) | Upregulated | N/A | Ref: [55,56] |
| Immune thrombocytopenic purpura (ITP) | Upregulated | N/A | Ref: [57] |
| acute/chronic inflammatory demyelinating polyradiculoneuropathies (AIDP/CIDP) | Upregulated | N/A | Ref: [58] |
| Atypical hemolytic uremic syndrome (aHUS) | Upregulated | N/A | Ref: [59] |
| Alzheimer’s disease (AD) | Upregulated | N/A | Ref: [60] |
| Hereditary angioedema (HAE) | Upregulated | N/A | Ref: [61] |
| Ischaemia/reperfusion (I/R) injury | Upregulated | N/A | Ref: [68] |
| Sjögren’s syndrome (pSS) | Upregulated | N/A | Ref: [70] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



