Submitted:
17 February 2025
Posted:
19 February 2025
You are already at the latest version
Abstract
Xylan is the primary hemicellulosic part of plant biomass, xylanase plays significant role for its degradation. The current research work was done to increase xylanase production in wild and mutant form from fungal isolates. Out of 21 soil samples, 6 fungal strains were found to be good xylanse producer in optimized condition mutant form of the strains gave significantly higher level of enzyme production. 21 soil samples were collected, 16 fungal isolates were purified and characterized morphological and molecular basis. Isolated producers were subjected to improve xylanase production under optimized growth condition and fermentation media. From these, 6 fungal isolates were found to be good producer of xylanase.ZGCL22 (KT964482), ZGCL49 (KT964481), ZGCL18 (KT964484), ZGCL44 (KT964483), ZGCL51 (KT964485) and ZGCL12 (KT964480) were good xylanase producer fungal isolates. 30°C temperature and 5.0 pH was found to be optimum for most of the xylanase producer fungal isolates except ZGCL44 strain which was found to be optimum at 9.0 pH for 5 days of fermentation period. Xylanase maximum activity observed at 50°C, at 30 minutes of incubation with 1% substrate concentration. On the basis of 18S rRNA sequencing using ITS primer. Xylanase encoding gene xyn B was amplified in one fungal strain ZGCL22. These local fungal isolates were good producers of xylanase and should be explored for enhanced biological production of xylanases under optimized conditions. Fungal production of xylanase will be cheaper and easier.
Keywords:
Microbial biotechnology
; Xylanase production
; Fungal xylanases
; wild and mutant strains
; xyn B gene
1. Introduction
Xylan may be the primary hemicellulosic part as well as the reason for in relation to 20-35% involving dried weight of seed biomass. It is composed of β-1,4-glycosidically linked xylose residues and is the most abundant hemicellulose as well as a complex polysaccharide [1]. Xylan hydrolysis was an important step for proper utilization of lignocellosic material. For the treatment of lignocellosic waste, microbial enzymes are used, which was considered to be cost effective and efficient method. Xylanase plays a vital role in many biotechnological applications. Xylanases are used in wood Kraft pulp as a bleaching agent which is considered its main industrial application. In recent years, some of the bleaching processes call for a lot of chlorine based chemicals, which are poisonous, persistent and mutagenic as well as causes various harmful effects on biological systems [2,3,4,5]. Various reports got exhibited that the usage of xylanases can lead to improvement of nutritive valuations of feed and performance of animals for example piggery, poultry and fishery. Interest in xylanase has extensively increased due to its wide range of industrial application. It is used in biofuel production, pretreatment process, poultry and bakery products, bio-bleaching and pulping in paper industry, waste treatment, clarification of wines and fruit juices, extraction of oils and pigments [6]. Submerged type fermentation can be used for xylanase production with suitable carbon source, which is a key factor in its production and agricultural wastes serve as cost effective alternative [7,8]. Microorganisms are involved for degradation of xylan in nature. Many fungi and bacteria are used for xylanase production such as Aspergillus, Trichoderma, Phanerochaete, Streptomyces, Chytridiomyces,Clostridium, Fibrobacter, Bacillus and Pichia Different microorganisms have ability to produce different degree of this enzyme according to their activities as well as different xylanase spectrum. Fungi are the major applicable source of xylanase at industrial level. Commercial xylanolytic preparation is prepared through genetically modified Aspergillus or Trichoderma strains. Penicillium and Aspergillus are filamentous fungi that play a crucial role as xylanase producers, as they secrete the enzyme into the medium at higher levels compared to other microorganisms [9,10,11,12,13]. Xylanases produced from Aspergillus niger was first time purified and characterized in 1977. Bacteria, yeast and fungi are the xylanase producing microorganisms, fungi produce greater amount of xylanase than bacteria. Fungal species that are known for xylanase production typically belong to genera including Aspergillus, Fusarium, Penicillium, Trichoderma, and Cunninghamella [14,15,16]. Although microorganisms secrete various metabolites, including enzymes, in small quantities, these amounts are often insufficient for large-scale industrial use. As a result, researchers are enthusiastically seeking methods to enhancement the production of these target metabolites. Strategies such as optimizing medium components and improving strains through mutagenesis and recombinant DNA technology are commonly explored [17,18]. Although mutations (whether chemical or physical) are sometimes seen as detrimental, they often result in microorganisms that are better suited to their environment, leading to enhanced enzyme production. UV radiation is an effective inducer of such mutations [19,20]. Therefore, the current study aim to screen potent fungal strains for xylanase production, optimization of fermentation media and growth conditions to further increase yield, enhance their production through UV mutation, and detect the presence of the XynB gene in both wild and mutant strains.
2. Materials and Methods
2.1. Collection and Isolation of Xylanolytic Fungi
Soil samples were collected from local paper or pulp industry, humus, compost, dung and fruit plants in sterilized and air tight bottles from different sites of district Lahore. Soil sample (1g) was dissolved in sterilized distilled water (100 mL) and then serial dilution were made. Each dilution was spread on the sterilized PDA (potato dextrose agar) plates following incubation for 7 days at 30 °C. Isolated colonies were streak further on fresh PDA plates until pure colonies of fungus were obtained.
2.2. Sampling and Processing of Agricultural Residue as Enzyme Substrate
Agricultural residues were utilized as fermentation substrates in this study, which aimed to produce valuable compounds from cost-effective materials. The substrates were chosen for their high potential to generate hydrolytic enzymes, particularly xylanases. Wheat bran, corn-cob, and rice straw were obtained from local markets in the Lahore district. The agro-residues underwent careful processing before being used as enzyme substrates. To remove dust and impurities, they were washed thoroughly with distilled water, then dried in a desiccator at 60 °C, and ground into a fine, uniform powder with a mesh size of 40 mm. The processed samples were then stored individually in airtight bottles until required for fermentation.
2.3. Screening of Xylanase Producing Fungi
Fungal strains capable of producing xylanase were evaluated for their activity using xylan as a pure substrate. Both qualitative and quantitative analyses were conducted to assess xylanase activity. The screening of fungal isolates was performed to investigate their ability to harvest extracellular xylanase while growing on xylan-rich media (10g/L) with pH adjusted to 5.5 ± 0.05. A fungal spore (1 mL) suspension was inoculated into`` the sterilized xylan medium (50 mL), and the flasks were incubate in a shaking incubator for 5 days at 31±1 °C. The extracellular enzymes were harvested after specific incubation period by filtration with Whatmann paper number 1 filter paper. It was further clarified via centrifugation at 10,000 rpm for 10 minutes. The collected enzyme considered as crude enzyme filtrate [21].
2.3.1. Qualitative Assay
The crude enzyme was utilized for a qualitative assay. Beechwood xylan (10.0 g/L) was dissolved in 100 mM phosphate buffer adapted to various pH levels (4.5, 5.5, and 6.5). Agar (18.0 g/L) was added to the xylan solutions, and the mixture was sterilized via steam sterilization for 15 minutes. After cooling, the solution (25ml) was poured into sterilized 90 mm petri plates. The plates were kept overnight at 31±1°C to check for contamination. Using a sterilized cork borer (10 mm diameter), wells were prepared in the agar. 100 μL of the extracted enzyme was dispensed in each well, while sterilized distilled water served as the control in one well. The plates were kept upright at 30±1°C. The diameter (in mm) of the clear zones was measured and recorded. To further enhance visibility, the zones of clearance were compared by staining the plates with Congo red solution (1%) for 30 min at room temperature. Then plates were de-stained with 1M sodium chloride solution, and the clear zone diameters were remeasured.
2.3.2. Quantitative Analysis of Xylanase Activity (Enzyme Assays)
Xylanase activity was assessed by combining 0.5 mL of 1% (w/v) beechwood xylan, prepared in 50 mM sodium citrate buffer (pH 5.5), with 0.5 mL of the enzyme solution. The mixture was then placed at 50 °C for 30 min. The reaction was terminated by adding 1 mL of 3,5-dinitrosalicylic acid (DNS) reagent, followed by boiling the reaction mixture for 5 minutes. After cooling, the absorbance was measured at 550 nm. The amount of released reducing sugar was quantified in mg/mL by comparing the absorbance values with a standard curve of xylose. One unit of xylanase is defined as the quantity of enzyme required to release 1 millimole of xylose equivalents per minute under the specified assay conditions.
2.4. Mutagenesis of Fungal Strains for Xylanase Enhancement
Mutation of the fungal spores was performed by following Ire et al (2021) with slight modification. The isolates were obtained by serially diluting a 24 h old culture. Under sterile conditions, 0.5 mL of the suspension was inoculated onto 0.5% xylan-PDA agar plates. 10-6 dilution was used and exposed it to the UV radiation for different time intervals (5, 25 and 60 min) at the distance of 10 cm from UV light (30W lamp, 254nm). Afterwards, 1 mL of UV exposed spore suspension was spread on prepared PDA plates under aseptic conditions. All plates were covered with aluminum foil or black paper to avoid photo radiation exposure to stabilize the A=T content of genome. To observe the growth of mutant strains plates were kept at 30 °C for 5 days.
2.5. Screening of Fungal Mutant Strains
Fungal mutant strains were screened to evaluate their xylanase production ability using both qualitative and quantitative assays as mentioned earlier in section 2.3.
Those strains which showed improvement in xylanase production capability after UV exposure were selected for further experimentation.
2.7. Enzyme Production Under Fermentation Conditions
After a 72-h incubation period, 3% of the inoculum was transferred into each 250 mL fermentation flask for extracellular xylanase production. Various culture conditions, such as incubation time, pH, and temperature, were optimized to enhance enzyme yield [22,23,24].
2.7.1. Effect of Carbon Sources on Xylanase Production in Wild and Mutant Fungal Strains
The impact of various carbon sources on xylanase production was studied using both wild and mutant fungal strains. Locally sourced substrates, including wheat bran, corn cobs, and rice straw, were individually used at a 2% concentration. The results provided insights into how carbon source influence xylanase activity, highlighting differences in performance between the wild-type and mutant strains.
2.7.2. Effect of pH on Xylanase Production in Wild and Mutant Fungal Strains
Xylanase biosynthesis was conducted across various pH levels (3, 5, 7 and 9) using wheat bran as fermentation substrate. To determine the optimal pH for enzyme production, pH of fermentation media was maintained using 1M tartaric acid and 1M NaOH solutions. On the fifth day of inoculation, the enzyme was extracted through filtration, followed by centrifugation at 10,000 rpm for 5 minutes.
2.7.3. Effect of Temperature on Xylanase Production in Wild and Mutant Fungal Strains
Xylanase production was investigated at varying temperatures (20°C, 30°C, 40°C, and 50°C) to assess the optimal temperature for enzyme activity. The study compared the performance of wild and mutant fungal strains, revealing how temperature variations influence xylanase production in each strain.
2.7.4. Effect of Incubation Time on Xylanase Production in Wild and Mutant Fungal Strains
To identify the optimal incubation period for maximum xylanase activity, samples were collected at sequential time intervals of 24, 48, 72, 96, 120, 144, and 168 h (1–7 days.
2.8. Isolation of Genomic DNA and from Wild and Mutant Fungal Strains
The organism was identified based on morphology and 18S rRNA sequencing. The genomic DNA from fungal strain were isolated by following Ire et al 2021.
2.8.1. Amplification of 18S rRNA Gene
A thermal cycler was used for PCR and reaction mixture having total volume of 50µl; containing 4µl of 10X taq buffer, 17 µl injection water, 16µl of 2.5mM MgCl2, 2 µl of 200mM dNTPs (deoxyribonucleoside tri phosphate), 2 µl of genomic DNA, 0.8 µl taq polymerase, 4 µl of each forward and reverse primer (ITS1 5' TCCGTAGGTGAACCTGCGG3' and ITS4 5' TCCTCCGCTTATTGATATGC3') was used for PCR. The reaction mixture was prepared in a PCR reaction tubes, kept in a thermal cycler with initial denaturation for 2 min at 94 °C, followed by 30 amplification cycles for 50 seconds at 54 °C, and elongation at 72 °C for 1 min. The final elongation step was performed at 72 °C for 20 min. Sterile double-distilled water was used as a negative control in place of template DNA.
2.8.2. Sequencing of Fungal Strains
PCR product was run on agarose gel (1.5%), at 90 volts for 30 min, using a 1 kb DNA ladder as the standard [25]. Bands were cut and gene clean was performed using Invitrogen gene clean kit, samples were sent for sequencing first base laboratory Malaysia.
2.9. Amplification of Xylanase Gene in Wild and Mutant Fungal Strain
Amplification of xylanase genes of wild and mutated fungal strains of Aspergillus niger (xlnB), was done by using proper conditions of PCR as mentioned above. For xyn B gene detection 63°C annealing temperature was used. 50 µl of reaction mixture was prepared as following; 10X taq buffer 4µl, injection water, 16 µl; 2.5mM MgCl2, 16µl; 200mM dNTPs, 2 µl; genomic DNA, 3 µl; taq polymerase, 0.8 µl and 4 µl of each forward and reverse XynB primers were used. Sequence of Xyn B forward and reverse primers are following:
Forward primer:
5'GGAATTCATGCTCACCAAGAACCTTCTC3'
Reverse Primer:
5'CCGCTCGAGTTACTGAACAGTGATGGAGGAAGA3'
2.9.1. Sequencing of Amplified Gene
The amplification product was resolved on a 1% agarose gel through electrophoresis, conducted at 80 volts for 20 min. The PCR product of amplified genes of wild and mutant Aspergillus niger was sent for sequencing to first base laboratory Malaysia.
3. Results
3.1. Sampling of Natural Sources for Xylanolytic Fungi
For analyzing the xylanase producing fungus, 21 soil samples were collected from paper or pulp industries, poultry dumping soil, local soil, composting material, fish market etc. from district Lahore and nearby areas. The physiological parameters such as temperature and pH of soil sample were also measured.
3.2. Isolation and Purification of Fungal Species
Among 21 soil samples 16 colonies were isolated and then further purified. Pure colonies of fungal isolates were obtained by streak on PDA plates and colony morphology was observed (Table 1).

Table 1.
Colony morphology of fungal isolates.
| Sr. No. |
Sample no. | Colony morphology | |||||
|---|---|---|---|---|---|---|---|
| Colony colour | Colony size | Colony elevation | Colony margin | Colony surface | Colony form | ||
| 1. | ZGCL01 | Green | Small | Flat | Undulate | Smooth | Irregular |
| 2. | ZGCL09 | Green | Small | Sharp | Entire | Smooth | Regular |
| 3. | ZGCL23 | Green | Small | Raised | White filiform | Rough | Irregular |
| 4. | ZGCL05 | Green | Small | Raised | White filiform | Rough | Irregular |
| 5. | ZGCL06 | Dark green | Big | Flat | Undulate | Smooth | Irregular |
| 6. | ZGCL27 | Black | Big | Raised | Filiform | Rough | Filamentous |
| 7. | ZGCL49 | Black | Small | Raised | Entire | Rough | Irregular |
| 8. | ZGCL09 | Dark green | medium | Raised | Filiform | Rough | Filamentous |
| 9. | ZGCL18 | White, green | Small | Raised | Entire | Smooth | Circular |
| 10. | ZGCL47 | Green | Small | Flat | Entire | Smooth | Circular |
| 11. | ZGCL14 | White, green | Medium | Flat | Filiform | Smooth | Circular |
| 12. | ZGCL44 | White | Large | Flat | Undulate | Smooth | Irregular |
| 13. | ZGCL51 | Green | Small | Raised | White entire margin | Smooth | Circular |
| 14. | ZGCL18 | White | Big | Flat | Undulate | Smooth | Irregular |
| 15. | ZGCL19 | Light green | Medium | Raised | Filiform | Rough | Circular |
| 16. | ZGCL12 | Green | Small | Flat | Undulate | Smooth | Circular |
3.3. Screening of Xylanase Producing Fungi
Xylan was used as a pure substance and those fungal strains was selected which gave positive results in this media. Qualitative and quantitative analysis was done to estimate the activity of xylanase.
3.3.1. Qualitative Assay
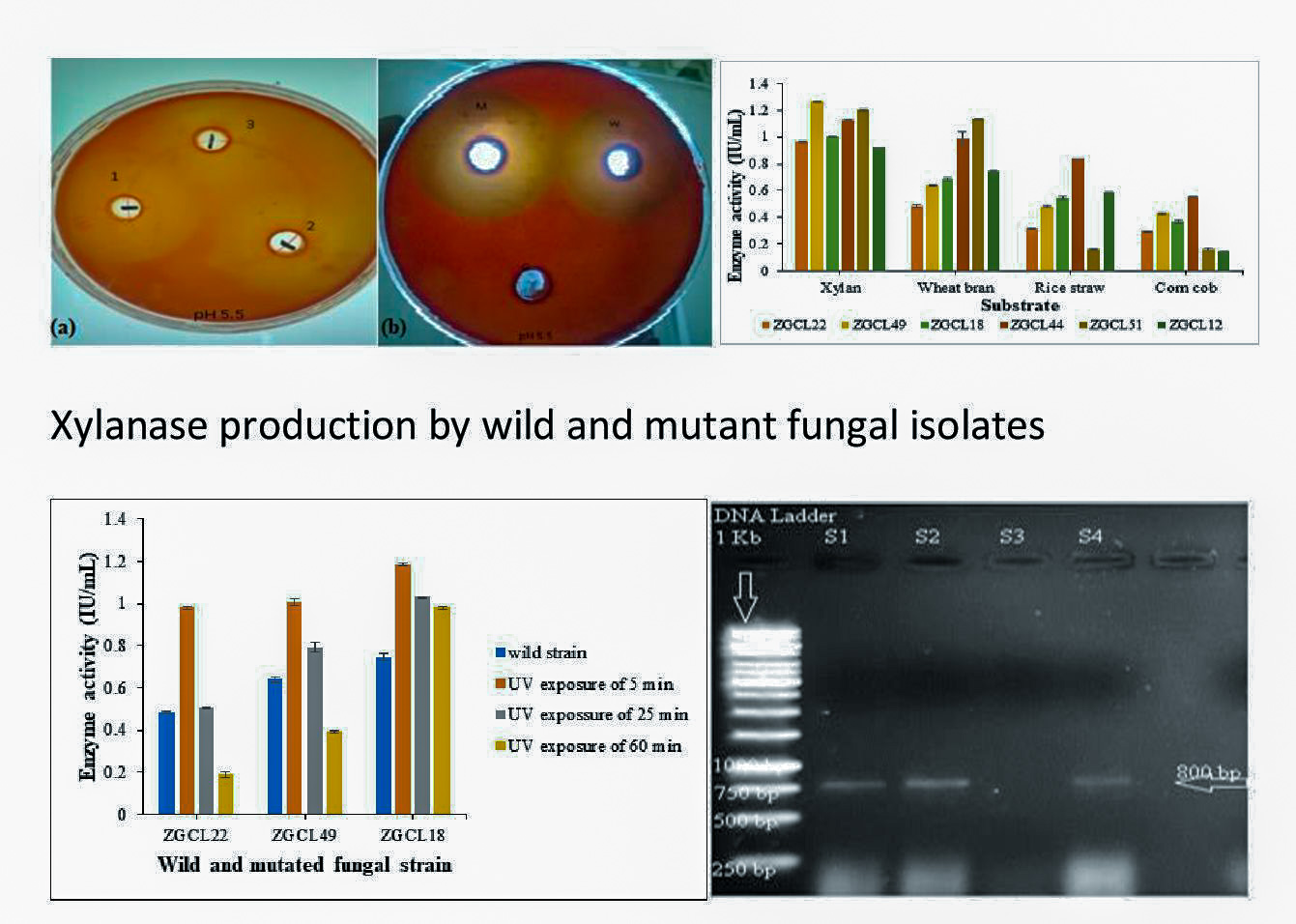
6 out of 16 fungal isolates gave positive results with xylan agar media. Following strains show clear zone around their well after 24 h of incubation at 30 °C (Figure 1a).
Figure 1.
zones showing enzyme activity.
3.3.2. Quantitative Enzyme Assays of Xylanase Activity
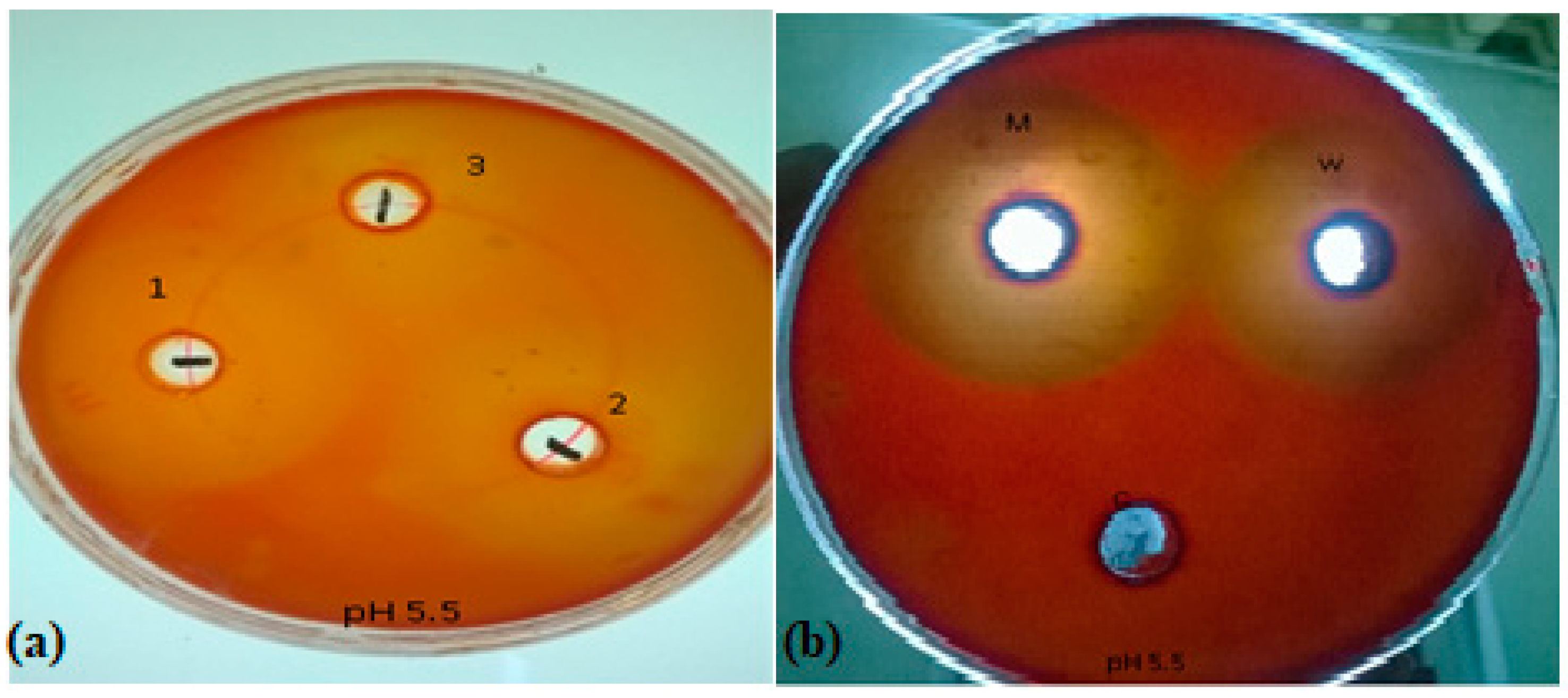
These 6 fungal strains were also tested by DNS method. Following fungal strains were good xylanase producer. Figure 2 show that ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 gave maximum xylanase production at 0.96, 1.26, 1.00, 1.12, 1.19 and 0.92 IU/mL respectively by using xylan as pure carbon source.
Figure 2.
Enzyme activity by different fungal isolates.
3.4. Mutation of the Xylanase Producing Fungal Strains
3 xylanase producing fungal strains were mutated through UV radiation. ZGCL22, ZGCL49, ZGCL12 were mutated for different time interval (5, 25 and 60min).
3.5. Screening of Fungal Mutant Strains
Fungal mutant strains were screened for their ability to produce xylanase using qualitative and quantitative assays.
3.5.1. Qualitative Assay
Qualitative assay of mutated fungal strains was performed at pH5.5 (Figure 1b).
3.5.2. Quantitative Assay
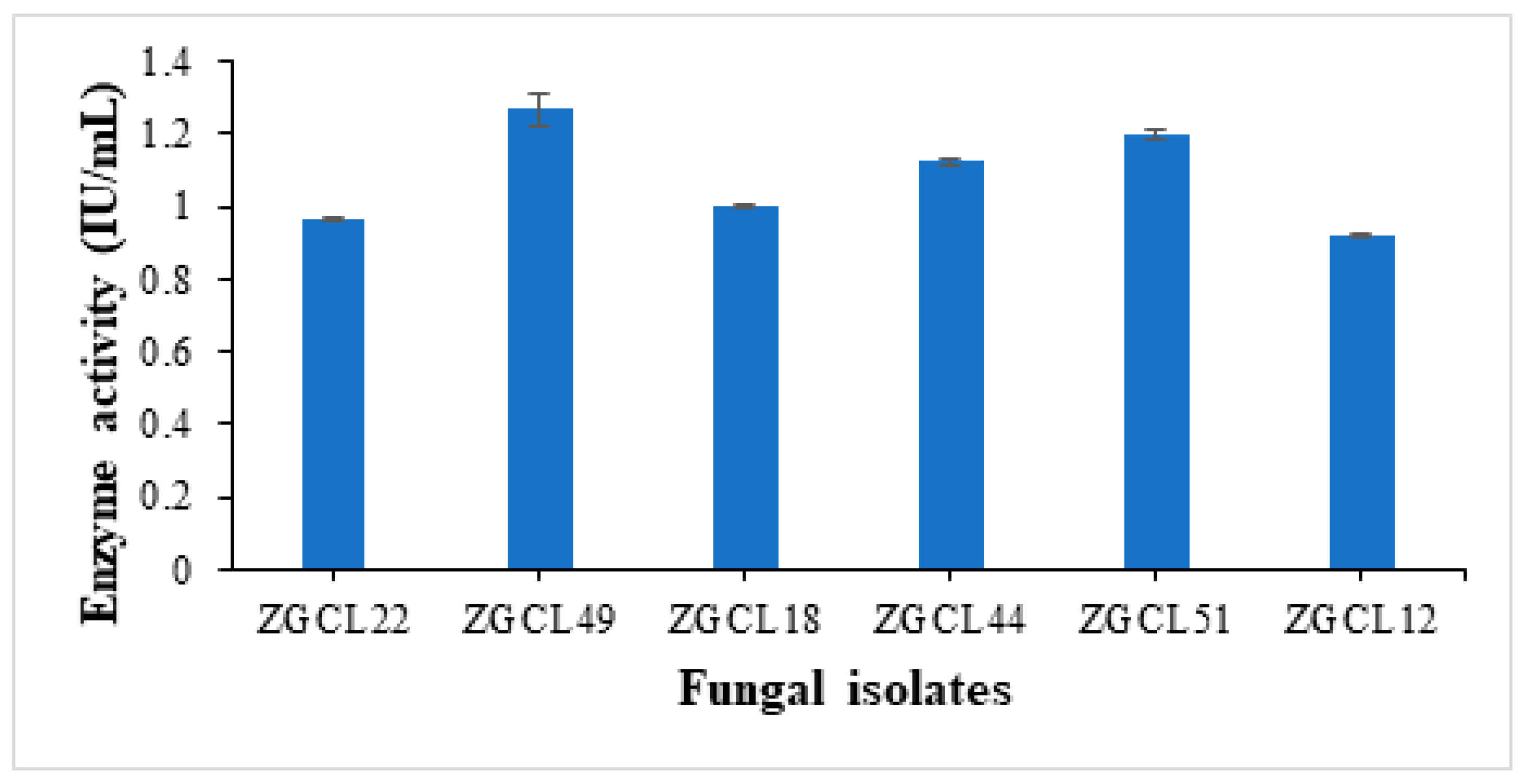
DNS method was applied to the wild and mutated (U.V radiation for 5, 25 and 60 minutes) fungal strains. Figure 3 represented that the ZGCL22, ZGCL49 and ZGCL12 gave 0.48 IU/mL, 0.63IU/mL and 0.74 IU/mL xylanase production respectively. However, at 5 minutes of UV mutation of these strains gave maximum xylanase production i.e. 0.97 IU/mL, 1.01 IU/mL and 1.18 IU/mL respectively. UV exposure of 5 minutes mutated fungal strains (ZGCL22(1), ZGCL49(1), ZGCL12(1)) were selected as they produced xylanase at higher level.
Figure 3.
Enzyme activity by wild and mutant fungal strains.
3.6. Enzyme Production Under Fermentation Conditions
Following 72 h of incubation, 1% of the inoculum used for the production of extracellular enzyme while optimizing various culture conditions including incubation time, temperature and pH.
3.6.1. Optimization of Carbon Source for Enhanced Xylanase Production
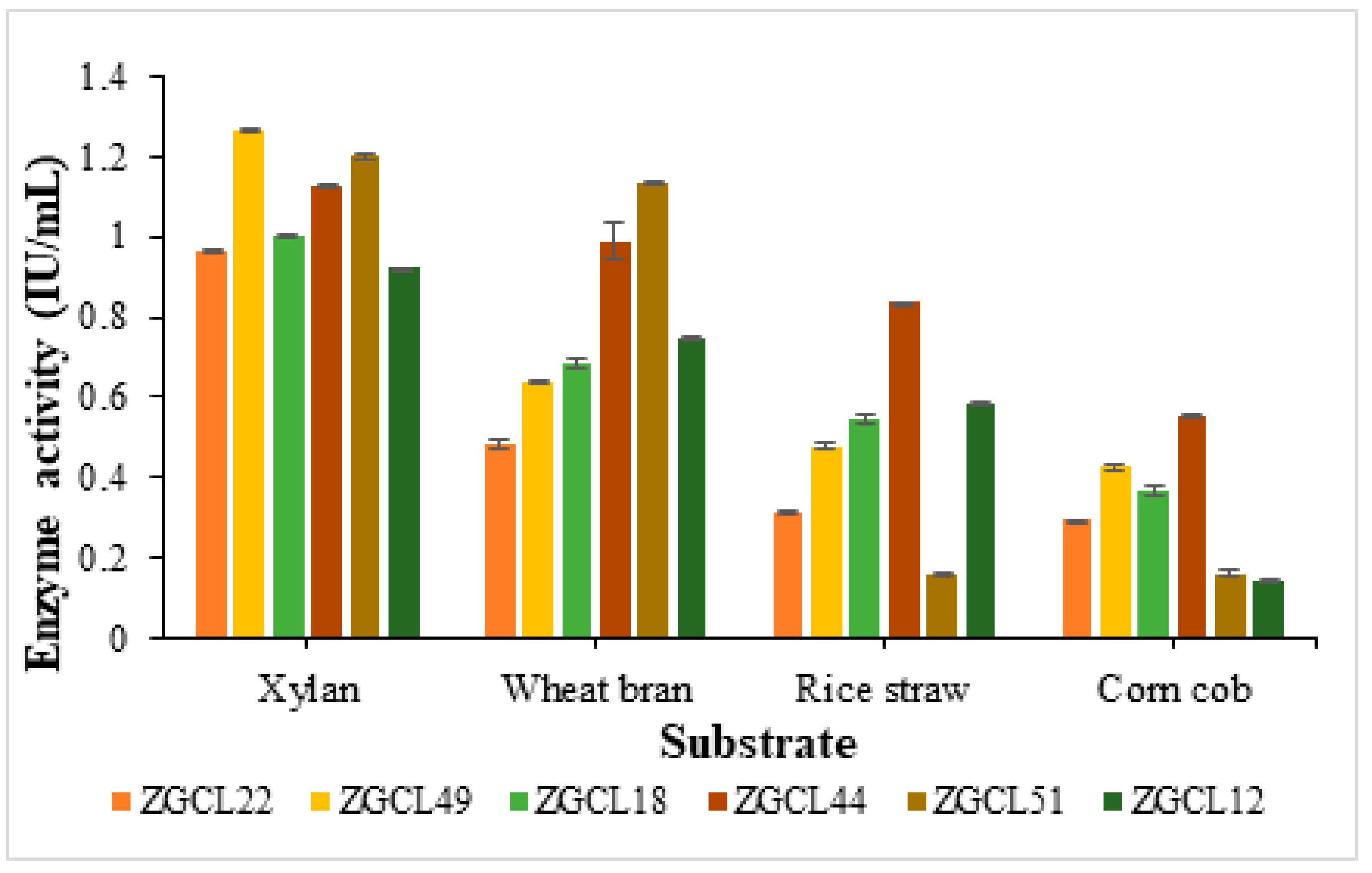
Various locally sourced substrates, including wheat bran, corn cobs, and rice straw, were individually utilized at a 2% concentration for xylanase production. Additionally, xylan was employed as a pure substrate. As depicted in Figure 4, wheat bran was identified as the most effective substrate for xylanase production compared to rice straw and corn cobs. The fermentation process for substrate selection was carried out over a period of five days.
Figure 4.
Enzyme activity by fungal strains in different substrates.
3.6.2. Effect of Different pH on Xylanase Production
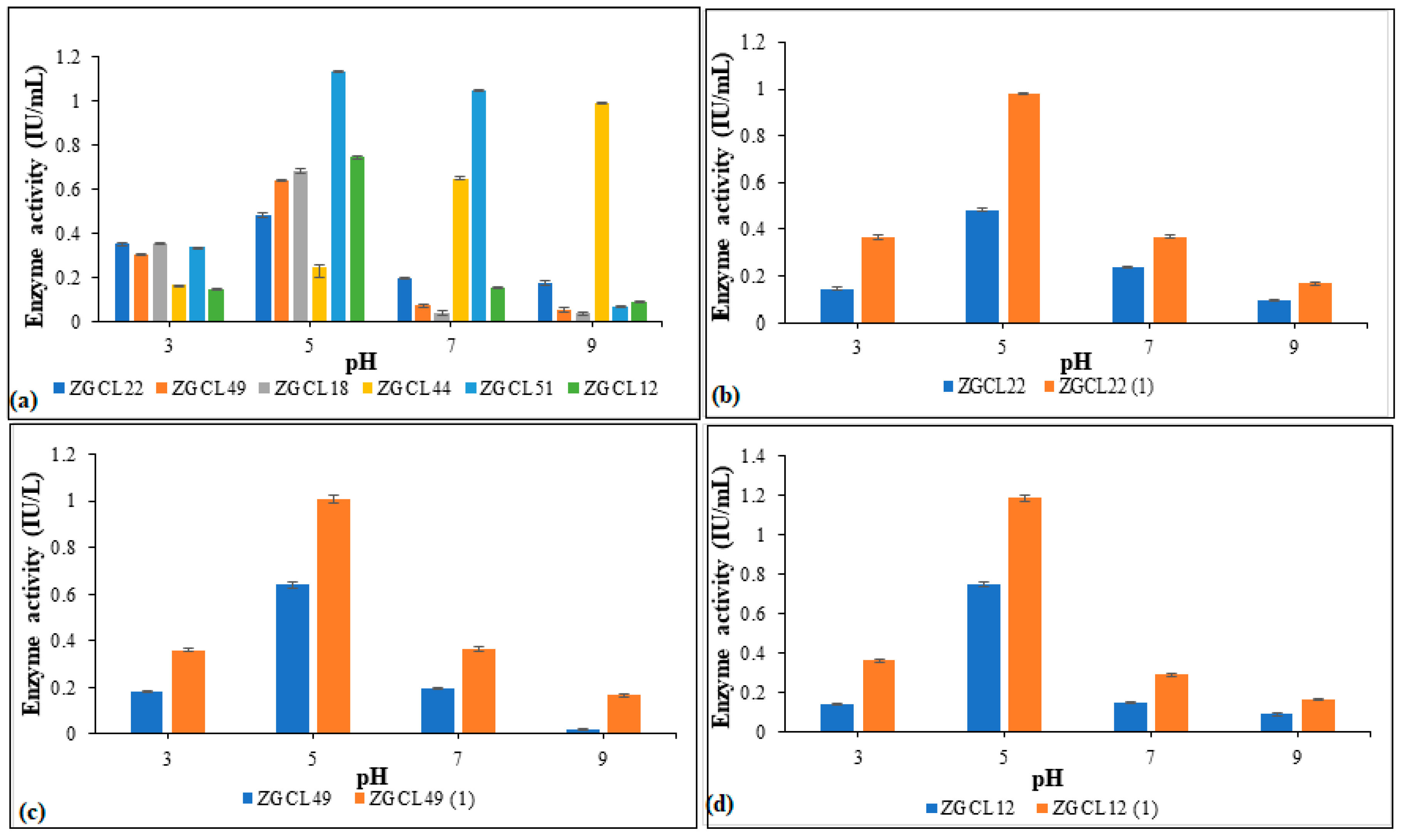
Different pH range 3-9 were tested for fungal xylanase production. At pH 5.0 ZGCL22, ZGCL49, ZGCL18, ZGCL51 and ZGCL12 show maximum xylanase production values such as 0.48, 0.64, 0.68, 1.13 and 0.74 IU/mL respectively. However ZGCL44 show maximum xylanase production value 0.98 IU/mL at pH 9.0 represented by Figure 5a.
Fungal strains ZGCL22, ZGCL49 and ZGCL12 wild and its mutated type were tested for different pH range of 3-9. At pH 5.0 ZGCL22 show 0.48 IU/mL and its mutated form ZGCL22 (1) show 0.97 IU/mL [Figure 5b], ZGCL49 show 0.64 IU/mL and its mutated form ZGCL49 (1), 1.00 IU/mL [Figure 5c]and ZGCL12 show 0.74IU/mL and its mutated form ZGCL12 (1) show 1.18 IU/mL xylanase production [Figure 5d].
Figure 5.
Enzyme activity by fugal strains in given pH.
3.6.3. Temperature
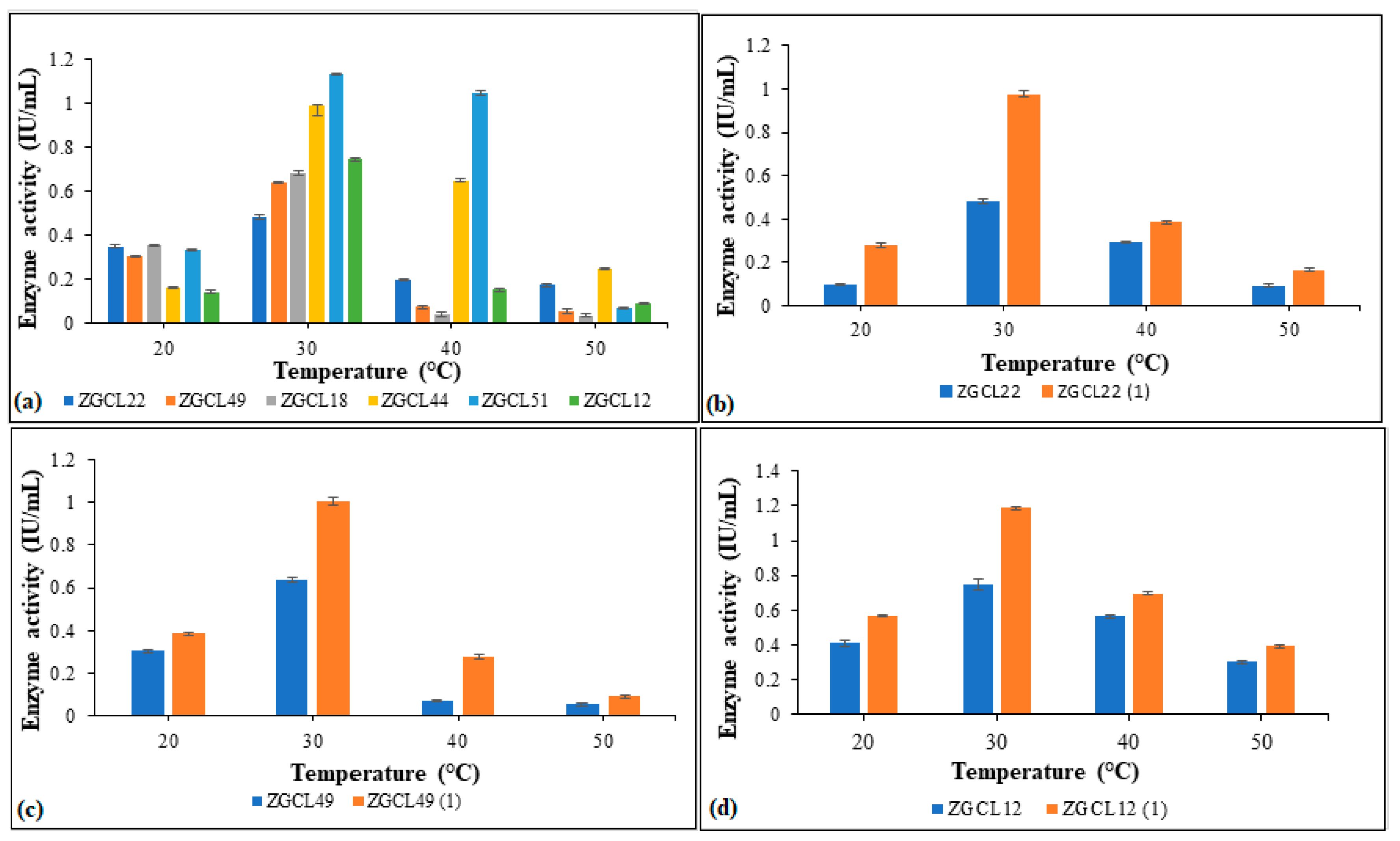
For temperature optimization, xylanase production was evaluated at varying incubation temperatures (20, 30, 40 and 50°C). 30°C was found to be optimum temperature for xylanase production for wild and mutated fungal strains.
ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 show maximum xylanase production values such as 0.48, 0.64, 0.68, 0.98, 1.13 and 0.74 IU/mL respectively at 30°C represented by Figure 6a.
Figure 6.
Enzyme activity by fungalstrains at given temperatures.
3.6.4. Incubation Time
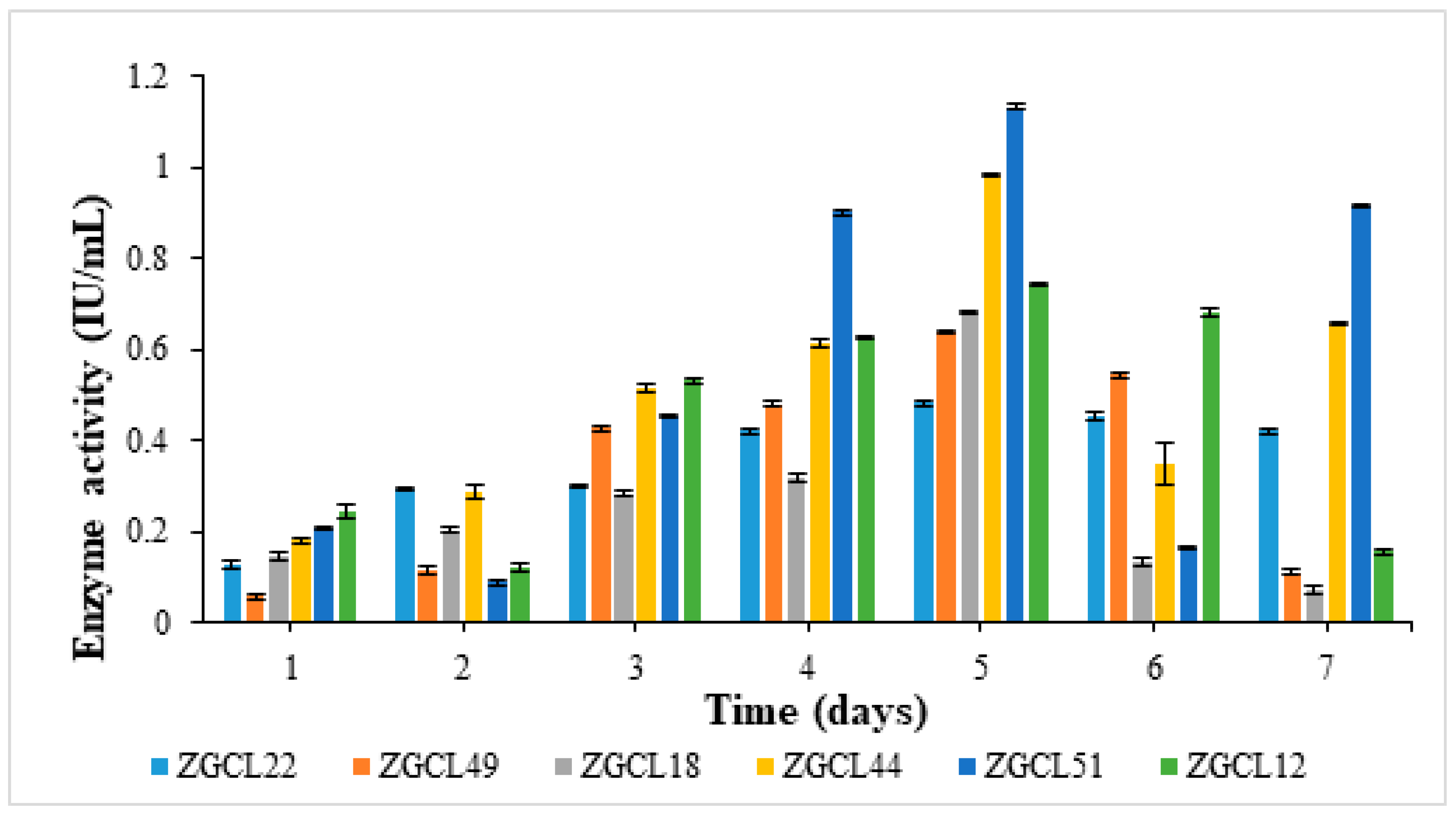
To determine the optimal incubation period for maximum xylanase activity, samples were collected at predefined time intervals of 24, 48, 72, 96, 120, 144, and 168 h (1-7 days). An incubation period of 120 hours was determined to be the optimal time for xylanase production. ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 show maximum xylanase production values such as 0.48, 0.64, 0.68, 0.98, 1.13 and 0.74 IU/mL respectively (Figure 7).
Figure 7.
Enzyme Activity by fungal strains in given days.
3.7. Sequencing of Fungal Strains
On the basis of 18S rRNA sequencing different fungal isolates were identified at species level (Table 2).
Table 2.
Fungal isolates showing percentage homology with standard fungal isolates.
| Sr. # | Isolates | Strain | Homology with | Percentage (%) | Accession number |
|---|---|---|---|---|---|
| 1. | ZGCL22 | Aspergillus niger | Aspergillus niger isolate SUMS0061 FJ011541.1 | 100 | KT964482 |
| 2. | ZGCL49 | Aspergillus flavus | Aspergillus flavus strain 176.1B2 KP784374.1 | 99 | KT964481 |
| 3. | ZGCL18 | Fusarium oxysporum | Fusarium oxysporum isolate FoxySIN8 KJ466113.1 | 100 | KT964484 |
| 4. | ZGCL44 | Pencillium digitatum | Pencillium digitatum, strain FRr 1313 AY373910.1 | 100 | KT964483 |
| 5. | ZGCL51 | Trichoderma harzonium | Trichoderma harzonium strain HEND AF055216.1 | 100 | KT964485 |
| 6. | ZGCL12 | Aspergillus oryzae | Aspergillus oryzae NRRL 506. (AF459735.1 | 100 | KT964480 |
3.8. Characterization of Xylanase Gene in Wild and Mutant Fungal Strain
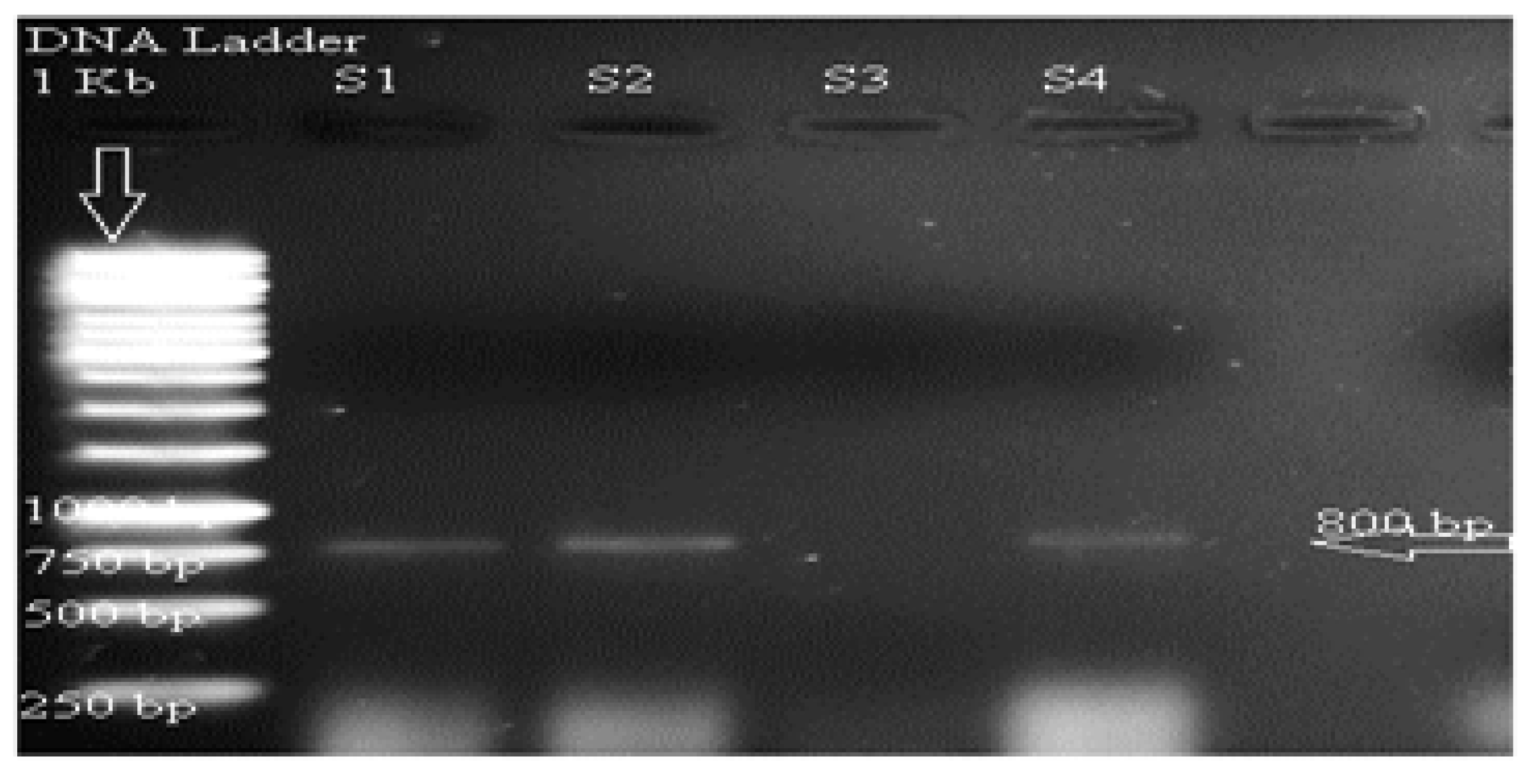
The xynB gene was amplified from the genomic DNA of fungal isolates using specific forward and reverse primers. The resulting PCR product was subsequently visualized on a 0.8% agarose gel alongside a DNA ladder for size reference. (Figure 8).
Figure 8.
PCR product of 800bp size.
3.8.1. Sequencing of Amplified Gene
Sequencing of xynB gene of wild and mutant Aspergillus niger was done through first base laboratory Malaysia.
Sequence of Aspergillus niger (xynB gene) wild strain:
ATGTGACCTACACCAACGGTGACGCTGGCTCGTACACCGTCGAGTGGTCCAACGTTGGCAACTTTGTTGGTGGAAAGGGCTGGAACCCTGGAAGTGCGCAGTAAGTTAACCTCTCTCACGCTGTCCCTGAATCGGGCTCCGTGAAATGAGAGCTCACATAACTACTGGGCCATCTTCTACTGCCTCACCCTCCCATGCAGCGGCTCTAGCTACCTTTCCCGCTATGGGTGGACAACTCACCCCCTGGTCGAATGTACATCCCCGAGACCCACGGGGAATCTACCCAGGTTCATGCCCAGTATTAGCCCAAGCGTTGAAGTGGTTGGAAGTGGTAACAGTTCCTCCAACTCTCTTGTTCTGGCGAACGGACCAGTACTGGGTGAAGGTAGCGGTTCCTTGGATAGAAGCGGCGTTGGAGCGGGTAGCTGTGTAGATATCGTAGACGGATCTATCGGANGTGACGGTGCCCTTGTACGTGCCTCCATTGCCGGGGTTGAAAAACTCTCATGATTGTTCGATACTCTAGGAGTGGTTTAAGCGCTTTACAAAGGCCTAATCCATAACTTAATGGTATCTCTTCCGAGGGTCTGGCATTTATATCACTNATGGGCCAAAGCAACAGCTTCTCTCAAATAGCCTTGTTTTTGTTATCTGCATAAAACGCATGAGGCAGCAGAGGCTCCTTCAGTAACG
Sequence of Aspergillus niger (xynB gene) mutant strain:
CACCAACGGTGACGCTGGCTCGTACACCGTCGAGTGGTCCAACGTTGGCAACTTTGTTGGTGGAAAGGGCTGGAACCCTGGAAGTGCGCAGTAAGTTAACCTCTCTCACGCTGTCCCTGAATCGGGCTCCGTGAGATGAGAGCTCACATAACTACTGGGCCATCTTCTACTGCCTCACCCTCCCATGCTGCGGCTCTAGCTACCTTTCCCGCTATGGGTGGACAACTCACCCCCTGGTCGAATGTACATCCCCGAGACCCACGGGGAATCTACCCAGGTTCATGCCCAGTATTAGGCCAACCGTTGAAGTGGTAGGAAGTGGTAACAGTTCCTCCAACTCTCTTGTTCTGGCGAACGGACCAGTACTGGGTGAAGGTAGCGGTTCCTTGGATAGTAGCGGCGTTGGAGCGGGTAGCTGTGTAGATATCGTAGACGGATCTATCGGANGTGACGGTGCCCTTGTACGTGCCTCCATTGCCGGGGTTGAAAAACTCTCATGATTGTTCGATACTCTAGGAGTGGTTTAAGCGCTTTACAAAGGCCAAATCCATAACTTAATGGTATCTCTTCCGAGGGTCTGGCATTTATATCACTNATGGGCCAAAGCAACAGCTTCTCTCAAATAGCCTTGTTTTTGTTATCTGCATAAAACGCATGAGGCAGCAGAGGCTCC
4. Discussion
Xylanase plays a crucial role in various industries, including animal feed, pulp and paper, textile, and food processing. Additionally, it has potential applications in converting xylan-rich waste from the food and agricultural industries into xylose, as well as in the production of biofuels and chemical feedstocks. Xylanase plays an active role in the lysis of xylan, used in various biotechnological applications. Out of various agro industrial raw material wheat straw, wheat bran and sugarcane baggase are the most suitable for this purpose and they also give high productivity on low cost [26].
The objective of the present work was to screen those fungal isolates which were good producer of xylanase. And then mutate them for the increase yield of xylanase production. Twenty one soil samples were collected from dumping and composting sites to isolate the fungal species which were good producer of xylanase. Sixteen fungal isolates were isolated from twenty one soil samples and on potato dextrose agar medium these strains were purified. The strains were identified by their colony morphological characterization. These sixteen isolates were screened qualitatively using xylan agar medium and quantitatively through DNS method. ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 gave maximum xylanase production at 0.96, 1.26, 1.00, 1.12, 1.19 and 0.92 IU/mL respectively by using xylan as pure carbon source. All the fungal strains were give maximum growth at fifth day of incubation and after this stipulated period of time growth started decline. Only one strain ZGCL44 showed optimum pH at 9.0, rest of 5 strains showed optimum pH at 5.0. All the fungal strains show maximum growth at temperature 30oC and growth started decline before and after that temperature. Wheat bran was used as a carbon source, gave maximum production of enzyme after xylan as compare to rice straw and corn cob. ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 show maximum xylanase production values such as 0.48, 0.64, 0.68, 0.98, 1.13 and 0.74 IU/mL respectively with wheat bran as carbon source. Different fermentation media was used for optimum production of enzyme, four different media was assessed for this purpose. Out of these four media the maximum production of xylanase was observed by M1 medium. The fungal species ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 showed maximum xylanase production i.e. 0.48, 0.64, 0.68, 0.98, 1.13 and 0.74 IU/mL respectively. This might be due to the availability of carbon and nitrogen content in sufficient amount which might be required for their proper growth. The remaining three media might lack sufficient quantities of some of these components as they produce less quantity of enzyme. The highest productivity 0.62 and 0.60 unit/mL occurred on peptone and xylan as organic, carbon and nitrogen sources (Al-Qahtani et al., 2013). The media pH has a great effect on the organism growth as well as on the synthesis of enzyme. ZGCL22, ZGCL49, ZGCL18, ZGCL51 and ZGCL12 show maximum xylanase production values such as 0.48, 0.64, 0.68, 1.13 and 0.74 IU/mL respectively at pH 5.0. However ZGCL44 show 0.98 IU/mL value at pH9.0. The optimum pH for enzyme activity of Aspergillus nigerwas found to be 5.0. Temperature of the media showed great effect on fungus growth and enzyme production. The optimum temperature for the maximum production of the enzyme were found to be 30oC. The enzyme production at this optimum temperature of fungal isolates ZGCL22, ZGCL49, ZGCL18, ZGCL44, ZGCL51 and ZGCL12 were 0.48, 0.64, 0.68, 0.98, 1.13 and 0.74 IU/mL respectively. Time of incubation had a tremendous effect on the enzyme production. 5 days of incubation was found to be optimum time for xylanase production. Maximum production of xylanase was produced by using groundnut shell as a sole carbon source, yeast extract and potassium nitrate (KNO3) as organic and inorganic nitrogen source, at pH 6.0 moisture level 75% for 5 days of fermentation.
The PCR product size was found to be 800 bp of fungal DNA. On the basis of 18S rRNA sequencing using ITS primer, ZGCL22 was 100% homologous to Aspergillus niger isolate SUMS0061 FJ011541.1,ZGCL49 was 99% homologous to Aspergillus flavus strain 176.1B2 KP784374.1, ZGCL18 showed 100% similarity with Fusarium oxysporum isolate FoxySIN8 KJ466113.1,ZGCL44 was 100 % homology with Pencilliumdigitatum, strain FRr 1313 AY373910.1, ZGCL51 showed 100% homology with Trichoderma harzonium strain HEND AF055216.1 and ZGCL12 showed 100% homology withAspergillus oryzae NRRL 506. AF459735.1.The results of this research go in similar with identification of fungal strain BCC7928 by their morphological characteristics and comparison of ITS (internal transcribed spacer) sequences of rRNA gene. ITS-F and ITS-R primers were used to amplify the partial fragment of DNA of the It was amplified by PCR. The PCR product, was examined to about 600 bp length. Phylogenetic analysis and phenotypic characteristics of ITS data revealed that BCC7928 strain was recognized as Marasmius sp. Aspergillus niger wild xyn B gene size was found to be about 800 bp. xyn B gene from the genomic library of DNA of Aspergillus niger was isolated ZGCL22 (KT964482) [27,28].
5. Conclusion
These local fungal isolates were good producers of xylanase and should be explored for enhanced biological production of xylanases under optimized conditions. Fungal production of xylanase will be cheaper and easier.
References
- Palaniappan, A.; Mapengo, C.R.; Emmambux, M.N. Properties of water-soluble xylan from different agricultural by-products and their production of xylooligosaccharides. Starch-Stärke. 2023, 75, 2200122. [Google Scholar] [CrossRef]
- Santibáñez, L.; Henríquez, C.; Corro-Tejeda, R.; Bernal, S.; Armijo, B.; Salazar, O. Xylooligosaccharides from lignocellulosic biomass: A comprehensive review. Carbohydrate Polymers. 2021, 251, 117118. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Thakur, M.; Tondan, D.; Bamrah, G.K.; Misra, S.; Kumar, P.; Kulshrestha, S. Nanomaterial conjugated lignocellulosic waste: cost-effective production of sustainable bioenergy using enzymes. Biotech. 2021, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.K.; Dixit, M.; Kapoor, R.K.; Shukla, P. Xylanolytic enzymes in pulp and paper industry: new technologies and perspectives. Molecular biotechnology. 2022, 64, 130–143. [Google Scholar] [CrossRef]
- Kumar, V.; Verma, P. A critical review on environmental risk and toxic hazards of refractory pollutants discharged in chlorolignin waste of pulp and paper mills and their remediation approaches for environmental safety. Environmental Research. 2023, 116728. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Kim, S.W. Nutritional and functional roles of phytase and xylanase enhancing the intestinal health and growth of nursery pigs and broiler chickens. Animals. 2022, 12, 3322. [Google Scholar] [CrossRef]
- Güler, F.; Özçelik, F. Screening of xylanase producing Bacillus species and optimization of xylanase process parameters in submerged fermentation. Biocatalysis and Agricultural Biotechnology. 2023, 51, 102801. [Google Scholar] [CrossRef]
- Al Mousa, A.A.; Abo-Dahab, N.F.; Hassane, A.M.; Gomaa, A.E.R.F.; Aljuriss, J.A.; Dahmash, N.D. Harnessing Mucor spp. for Xylanase Production: Statistical Optimization in Submerged Fermentation Using Agro-Industrial Wastes. BioMed Research International. 2022, 2022, 3816010. [Google Scholar] [CrossRef]
- Šuchová, K.; Fehér, C.; Ravn, J.L.; Bedő, S.; Biely, P.; Geijer, C. Cellulose-and xylan-degrading yeasts: Enzymes, applications and biotechnological potential. Biotechnology Advances. 2022, 59, 107981. [Google Scholar] [CrossRef]
- Kumar, A.; Kumari, S.; Dindhoria, K.; Manyapu, V.; Kumar, R. Efficient utilization and bioprocessing of agro-industrial waste. In Sustainable agriculture reviews 56: Bioconversion of food and agricultural waste into value-added materials. 2022, (pp. 1–37). Cham: Springer International Publishing.
- Dhaver, P.; Pletschke, B.; Sithole, B.; Govinden, R. Isolation, screening, preliminary optimisation and characterisation of thermostable xylanase production under submerged fermentation by fungi in Durban, South Africa. Mycology. 2022, 13, 271–292. [Google Scholar] [CrossRef]
- Dar, F.M.; Dar, P.M. Fungal xylanases for different industrial applications. Industrially Important Fungi for Sustainable Development: Bioprospecting for Biomolecules 2021, 2, 515–539. [Google Scholar]
- Ellatif, S.A.; Abdel Razik, E.S.; Al-Surhanee, A.A.; Al-Sarraj, F.; Daigham, G.E.; Mahfouz, A.Y. Enhanced production, cloning, and expression of a xylanase gene from endophytic fungal strain Trichoderma harzianum kj831197. 1: unveiling the in vitro anti-fungal activity against phytopathogenic fungi. Journal of Fungi. 2022, 8, 447. [Google Scholar] [CrossRef] [PubMed]
- Saroj, P.P.M.; Narasimhulu, K. Biochemical characterization of thermostable carboxymethyl cellulase and β-glucosidase from Aspergillus fumigatus JCM 10253. Applied Biochemistry and Biotechnology. 2022, 194, 2503–2527. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.; Barzkar, N.; Michaud, P.; Tamadoni Jahromi, S.; Babich, O.; Sukhikh, S.; Nahavandi, R. Cellulolytic and xylanolytic enzymes from yeasts: Properties and industrial applications. Molecules. 2022, 27, 3783. [Google Scholar] [CrossRef]
- Corbu, V.M.; Gheorghe-Barbu, I.; Dumbravă, A.Ș.; Vrâncianu, C.O.; Șesan, T.E. Current insights in fungal importance—a comprehensive review. Microorganisms. 2023, 11, 1384. [Google Scholar] [CrossRef]
- Trovão, M.; Schüler, L.M.; Machado, A.; Bombo, G.; Navalho, S.; Barros, A.; Varela, J. Random mutagenesis as a promising tool for microalgal strain improvement towards industrial production. Marine drugs. 2022, 20, 440. [Google Scholar] [CrossRef]
- Yadav, H.; Singh, S.; Sinha, R. Fermentation Technology for Microbial Products and Their Process Optimization. In Industrial Microbiology and Biotechnology: A New Horizon of the Microbial World. 2024, 35-64. Singapore: Springer Nature Singapore.
- Banoon, S.R.; Salih, T.S.; Ghasemian, A. Genetic mutations and major human disorders: A review. Egyptian Journal of Chemistry. 2022, 65, 571–589. [Google Scholar] [CrossRef]
- Akin, C.; Arock, M.; Valent, P. Tyrosine kinase inhibitors for the treatment of indolent systemic mastocytosis: are we there yet? Journal of Allergy and Clinical Immunology. 2022, 149, 1912–1918. [Google Scholar] [CrossRef]
- Liguori, R.; Pennacchio, A.; Vandenberghe, L.P.D.S.; De Chiaro, A.; Birolo, L.; Soccol, C.R.; Faraco, V. Screening of fungal strains for cellulolytic and xylanolytic activities production and evaluation of brewers’ spent grain as substrate for enzyme production by selected fungi. Energies. 2021, 14, 4443. [Google Scholar] [CrossRef]
- Chmelová, D.; Legerská, B.; Kunstová, J.; Ondrejovič, M.; Miertuš, S. The production of laccases by white-rot fungi under solid-state fermentation conditions. World Journal of Microbiology and Biotechnology. 2022, 38, 21. [Google Scholar] [CrossRef]
- Bellaouchi, R.; Abouloifa, H.; Rokni, Y.; Hasnaoui, A.; Ghabbour, N.; Hakkou, A.; Asehraou, A. Characterization and optimization of extracellular enzymes production by Aspergillus niger strains isolated from date by-products. Journal of Genetic Engineering and Biotechnology. 2021, 19, 50. [Google Scholar] [CrossRef] [PubMed]
- Siwach, R.; Sharma, S.; Khan, A.A.; Kumar, A.; Agrawal, S. Optimization of xylanase production by Bacillus sp. MCC2212 under solid-state fermentation using response surface methodology. Biocatalysis and Agricultural Biotechnology 2024, 103085. [Google Scholar] [CrossRef]
- Hassan, Z.; Khan, G. . PCR-Based Techniques. In Molecular Techniques for Studying Viruses: Practical Notes. 2024, 25-31. Singapore: Springer Nature Singapore.
- Abena, T.; Simachew, A. A review on xylanase sources, classification, mode of action, fermentation processes, and applications as a promising biocatalyst. BioTechnologia. 2024, 105, 273. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Kaushik, R.; Sirohi, R. Insights into recent advances in agro-waste derived xylooligosaccharides: production, purification, market, and health benefits. Biomass Conversion and Biorefinery. 2024, 1–21. [Google Scholar] [CrossRef]
- Chen, J.; Huang, B.; Liu, Y.; Sun, X.; Xiong, L.; Zhu, T.; Liu, H. Characterization of a novel cold-active β-Xylosidase from Parabacteroides distasonis and its synergistic hydrolysis of beechwood xylan. International Journal of Biological Macromolecules. 2025, 284, 137895. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



