Submitted:
09 February 2025
Posted:
10 February 2025
You are already at the latest version
Abstract
Drought is one of the most significant agrometeorological hazards which limit wheat production and yield. Wheat exhibits a variety of morphological, physiological, biochemical and molecular responses to drought stress. In this study, we simulated the effects of drought stress in wheat by using the CiteSpace visualization tool, sumarized climate impact and adaptation to drought stress of wheat pruduction, drought tolerance in wheat, progress and technologies on breeding wheat for drought tolerance, and drought stress impact on vigour, plant traits and yield. Under drought stress, plant height, water content, leaf area index, chlorophyll content and chlorophyll fluorescence parameters of wheat showed different degrees of decline, and water stress significantly increased the content of osmoregulatory substances (proline, free amino acid, total soluble sugar) in wheat. Drought tolerance is a complex trait controlled by multiple genes, and its expression is influenced by various environmental factors. Research on the evolutionary trend of dry matter accumulation in different ploidies of wheat during the process of hybridization and domestication is necessary, and molecular-level studies on the drought tolerance mechanism in wheat should be conducted to provide a basis for wheat breeding.Wheat breeding with new traits is a climate change adaptation option, however its effect will vary among different regions and limited under soil conditions and local environmental conditions. This review paper focused on water limitation and its effects on morphological, physiological, biochemical and molecular responses of wheat leaves and possible losses due to drought stress.
Keywords:
wheat
; growth period
; leaf water potential
; chlorophyll fluorescence
; molecular mechanisms
; photosynthesis
1. Introduction
Since the 1970s, global climatic droughts have been intensifying due to increasing warming and changes in atmospheric circulation patterns [1]. Droughts are characterized by high frequencies, long durations, and wide coverage, and they continue to pose a major threat to food security in today’s environment, characterized by rising temperatures and decreasing rainfall [2]. The Sixth Assessment Report of the IPCC (IPCC, 2023) states that the negative impacts of highly regional and complex drought hazards on grain yields have continued to intensify, and the contradiction between the increasing demand for wheat and decreasing yields is becoming increasingly prominent. Wheat (Triticum aestivum L., 2n = 6x = 42, AABBDD) is a major food crop globally, and individuals who consume wheat as a staple food account for approximately 35-40% of the world’s population [3]. As one of the three major food crops in China, wheat is grown and consumed in huge quantities throughout the country. Global wheat production in the major production regions is being threatened by recurrent grought that is predicted to increase with climate change [4]. Drought is one of the most common environmental stresses that affect growth and development of crops. In the context of the expanding arid and semi-arid areas, confined and regional drought disasters occur frequently [5,6]. South Africa has a semi-arid climate and faces significant drought challenges, with prolonged drought leading to water scarcity, which affects both rain-fed and irrigated agriculture [7]. In India and Pkistan, temperature increases and erratic monsoon patterns have been reported to be collectively linked to decreased wheat and rice yields, highlighting the vulnerability of these staple crops to climate variability [8]. Australia is one of the largest world grains exporting countries, where agriculture is dominated by rainfed cropping in arid and semi-arid regions, experiencing frequent severe droughts in recent decades [9,10,11,12]. The droughts caused significant reductions and failures in crop yields across the primary grain-producing areas of south-eastern Australia, particularly in 2019 [13]. They pose a serious threat to China’s food security and ecological safety and are important factors affecting China’s sustainable socioeconomic development [4,5].
Models based on the Coupled Model Intercomparison Project Phase 6 (CMIP6) project increased drought frequecy and severity in Australia under greenhouse warming scenarios [16,17], indicated that how the rainfed crop yield responds to different drought conditions across regions is essential to develop effective adaptation options to manage the risk of future droughts. Tolerance to water stress is a complicated parameter in which crops’ performance can be influenced by several characteristics [18]. Standardized Precipitation Evapotranspiration Index (SPEI) and the Palmer Drought Severity Index (PDSI) were developed to assess different drought metrics such as duration, frequency, and severity [19], whose advantages are easily calculated using readily available meteorological and hydrolodical data. SPEI set up to above and /or below certain thresholds [20]. In recent years, statistical analysis and crop simulation modeling have emerged as prevalent methodologies for evaluating the impact of climate change on crop production [21,22,23,24]. Moreover, Lobell et al. (2015) used a modified APSIM (Agricultural Production Systems Simulator) model to quantify the influence of drought and heat stresses on crop production in northeast Australia. By leveraging such approached, researchers can enhance our understanding and prediction of agro-climatic effects on agricultural systems, thereby aiding in strategies. Additionally, the deterministic nature of empirical and biophysical models [25] makes them accessible for direct use and easy to understand. However, these methods need to be equipped with sufficient input data. In some cases, input data may not be available with detailed measurements at a large scale.
Tolerance can be divided into two parts including drought avoidance and dehydration tolerance [26]. Drought avoidance includes root depth, appropriate use of available water by plants, and changes in plants’ lifestyle to use rainfall [27]. Adaption of plants to drought stress is a important points to develop new improve approachs for increasing stress tolerant plants [28]. Many factors can affect plants’ responses to drought stress such as plant genotype, growth stage, severity and duration of stress, physiological process of growth [29], different patterns of genes expression [30], different patterns of the activity of respiration [31], activity of photosynthesis machinery [32], and environmental factors [28,33]. Wheat has a certain degree of drought tolerance and can implement complex physiological and biochemical responses under drought in order to ensure optimal growth and development under such adverse conditions [34]. The leaf is one of the most sensitive organs to drought stress, which inhibits wheat’s growth and development, reduces the leaf water potential, affects the photosynthetic mechanism and organic matter synthesis, and, in severe cases, affects the leaf morphology and causes irreversible damage to the leaf. As the main site for photosynthesis, the physiological state of the leaf has a critical impact on the yield of wheat.
Many factors can affect plants’ responses to drought stress such as plant genotype, growth stage, severity and duration of stress, physiological process of growth [35], different patterns of genes expression [36], different patterns of the activity of respiration [37], activity of photosynthesis machinery [38], and environmental factors [39]. Drought stress can also influence plants in terms of protein changes, antioxidant production, osmotic adjustment, hormone composition, root depth and extension, opening and closing of stomata, cuticle thickness, inhibition of photosynthesis, decrease in chlorophyll content, reduction in transpiration, and growth inhibition [40,41,42,43] to stand with some osmotic changes in their organs. The role of genes can be distinguished by expression of a gene to high resistance levels among varieties [44]. In many drought studies, the role of reactive oxygen species (ROS) has been identified. Increase in ROS can be caused by drought stress in which oxidative balance of the cell is changed [45,46]. A rise in the generation of ROS prompts to the generation of ABA (abscisic acid) which is a general signal under drought [47,48,49,50] and can consequently reglate the antioxidant genes expressions by produing superoxide dismutase (SOD) and catalase (CAT) [51]. Drought can laso cause pollen sterility, grain loss, accumulation of abscisic acid in spikes of drought-susceptible wheat genotypes, and abscisic acid synthesis genes in the anthers [45].
Physiological responses include closure of stomata, decrease in the activity of photosynthesis,development of oxidative stress, alteration in the integrity of cell wall, production of metabolites which are toxic and cause plants’ death [52], signal recognition of roots, turgor loss and adjustment of osmosis, reduction in water potential of leaf, decrease in stomata conductance to CO2, reduction of internal CO2 concentration, and reduction of growth rates. For measuring drought tolerance, various scientists considered maintenance of membrane integrity and its role under water stress [33,34]. K. Wiśniewski and B. Zagdańska [53] also observed that the role of cysteine was improved, but its role was negatively related to the degree of drought tolerance of ten lines of spring wheat. Transpiration efficiency (TE) can be influenced by cultivar and drought [54,55].
Studying the effects of different water stresses on the photosynthesis, growth, yield, water use efficiency (WUE) and irrigation water productivity (IWP) of water wheat will provide data for the development of scientific irrigation strategies for water-saving agricultural methods. The seasonal changes in photosynthetic parameters, such as net photosynthetic rate (), intercellular carbon concentration (), stomatal conductance () and transpiration (E), significantly decreased under moderate and severe stress. As a result, the height, biomass and grain size of winter wheat decreased significantly, which led to low WUE and IWP. The of the mild stress group only slightly decreased compared to that of the well-watered group, and was actually higher during the flowering and grain-filling stages, resulting in increases in dry biomass and 1000 grain weight of 2.07% and 1.95%, respectively. Higher WUE and IWP were attributed to higher yields anf less water use. Thus, mild stress (60-80% field water capacity) resulted in the optimal use of water resources without a significant reduction in yield in the North China Plain (NCP). Therefore, mild stress can be considered a suitable environment for winter wheat growth in arid areas [56]. The stress of water deficit prompted a marked reduction in agronomic parameters while biochemical attributes were increased under conditions of water stress expect for total soluble proteins. Si application improves plant biochemical attributes, absorbs nutrients and shows higher growth rate which ultimately helps the plant mitigate the drought-stressing effects. At the anthesis stage, foliar spray was stronger while at the tillering stage, fertigation produced good results [57]. Salicylic acid (SA) is a promising compound to increase plant tolerance to drought stress, and it can affect many aspects of physiological and biochemical processes. Membrane electrolyte leakage, and lipid peroxidation of all ecotypes, were obviously increased under drought stress conditions. Drought stress decreased leaf chlorophyll content, photosynthetic rate, stomatal conductance, carboxylation effeciency,and transpiration rate [58].
The study aims to visualize the intellectual background, current research status and state-of-the-art knowledge structure of drought stress in wheat related literature using CiteSpace based scientometric investigation. The drought stress in wheat related original articles, published from 2013 to 2023, were retrieved from the Web of Science core collection (WOS) and China National Knowledge Infrastructure (CNKI). The most prolific publications, contries and journals involved in the flourishment of wheat drought stress research were identified. Moreover, visualization methods were employed to determine the highly priductive articles, keywords,hotspots, and research frontiers in the wheat drought stress domain. Furthermore, the classification of wheat drought stress kowledge was performed in the form of clusters and knowledge structure to achieve five distinct sub-domains [59,60,61]. The study can help the researchers to spot the new research frontiers and distinguish among the most drought in wheat based knowdge. The research papers were retrieved from the China National Knowledge Infrastructure (CNKI) and Web of Science core collection (WOSCC) databases and the bibliometric investigation was conducted using CiteSpace software to answer the following questions:
- (a)
- What is the aspects of growth of published articles on drought stress in wheat research?
- (b)
- What are the recent and emerging research frontiers in drought stress in wheat research?
- (c)
- What is the knowledge structure of drought stress in wheat research based the different key words co-occurrence network?
In this study, we analyze the literature related to wheat and drought through CiteSpace visualization on China National Knowledge Infrastructure (CNKI) and Web of Science (WOS) databases, covering a 10-year range, and we explore and summarize the vast research content. The main work can be summarized as follows:
- (1)
- ComprehensiveAnalysis: This study provides a comprehensive analysis of research of drought stress in wheat by testing various topics, such as publication trends, journal distributions, author networks, institutional networks, national networks, and keyword co-occurrences, as well as undertaking timeline analysis and emergent word analysis. This comprehensive approach offers a holistic understanding of the research landscape in research of drought stress in wheat;
- (2)
- Identification of research themes: Through the analysis of keywords and clustering, this study identifies key research themes in research of drought stress in wheat. It highlights the dominant areas of research, including spring wheat fertility in response to drought stress, leaf water potential in response to drought stress, chlorophyll content in response to drought stress, photosynthesis in response to drought stress, chlorophyll fluorescence parameters in response to drought stress, drought stress in wheat at the molecular level. This identification of research themes helps to delineate the major focus areas in the field;
- (3)
- Visualization of research patterns: The use of visualization techniques, such as network diagrams and timeline analysis, helps to visualize and understand the evolution of research hotspots over time. It allows researchers to observe the dynamics of research themes, emerging trends, and the interconnections between different topics [62,63,64].
In summary, this review explores the changes in the physiological properties of wheat leaves under drought, so as to provide a theoretical basis for the cultivation and breeding of drought-resistant wheat.
2. Materials and Methods
2.1. Data Sources
IIn this study, we conduct a statistical analysis through bibliometrics, analyze and summarize the keywords in the literature, and explore their structural characteristics and internal laws in order to obtain a comprehensive and objective understanding of the field. The literature data were obtained from the China National Knowledge Infrastructure (CNKI) databases (https://www.cnki.net) and Web of Science core collection (WOS) core collection databases (https://webofscience.clarivate.cn/wos/author/search). The Chinese database was searched and screened, spanning the period from 2013 to 2023, with the themes of “spring wheat”, “drought stress”, “leaf”, and “photosynthesis physiology”. The screened literature was exported in the RefWorks format as a sample for data analysis. The English database was searched with “spring wheat drought”, “ spring wheat leaf”, and “crop yield” as the themes to achieve an advanced search, and the documents were exported as plain text files in the form of “Full Record with Cited References” as a sample for data analysis.
2.2. Data Processing
2.2.1. Literature Selection Criteria
Advanced searches were conducted on CNKI and WOS using keywords, and the results were organized to exclude documents such as dissertations, conferences, newspapers, book reviews, patents, and almanacs [65,66]. The main proceeds are as follows:
- (1)
- Inclusion Criteria:The literature chosed for this study focused on research of drought stress in wheat, encompassing topics such as spring wheat fertility in response to drought stress, leaf water potential in response to drought stress, chlorophyll content in response to drought stress, photosynthesis in response to drought stress, chlorophyll fluorescence parameters in response to drought stress, drought stress in wheat at the molecular level.
- (2)
- Exclusion Criteria: Articles unrelated to the topic, such as achievements, conference papers, patents, advertisements, popular science articles, etc.; Non-original research, such as systematic reviews, meta-analyses, and reviews of wildfire prediction research; Articles with incomplete information, such as author, year, keywords, etc.; Duplicate or withdrawn publications [64,67].
After applying the screening criteria, a total of 11557 literatures were included as the sample data for this study.
2.2.2. Data Processing Software
CiteSpace, a citation visualization analysis software package, was utilized in this study to analyze the potential knowledge included in the scientific literature. CiteSpace is specifically designed for scientific metrics and data visualization, aiming to uncover dynamic development patterns within disciplines and identify research frontiers. The version used in this study was CiteSpace 6.2.R2 (64-bit), Advanced, developed by Dr. Chaomei Chen and their team [68,69]. CiteSpace 6.2.R2 software was used to generate intuitive knowledge graphs from the analyzed samples of data obtained from CNKI and WOS. In the software, according to the node type, the screened literature was visualized and analyzed by “keyword”, the time frame was set as 2013-2023, and the time slice was 1. A keyword network diagram was constructed so that we could determine the current research hotspots and development trends.
3. Results
3.1. Spring Wheat Fertility in Response to Drought Stress
The fertility period of spring wheat is an important indicator reflecting the growth and development process of this crop, which is affected not only by its genetic characteristics but also by the climatic conditions of the planting area. The alignment between the fertility period and water and heat resources is an important factor affecting the physiological and biochemical characteristics of the crop.
Based on the growth and development characteristics of spring wheat at each stage, the fertility period can be divided into seedling, triticale, tillering, nodulation, gestation, tasseling, flowering, milky, and maturity. Among these, the seedling to tasseling stage constitutes the nutritive growth period, and the flowering to maturity stage constitutes the reproductive growth period [70]. Figure 1 shows the keyword visualization knowledge map for research on spring wheat’s fertility in response to drought stress. From the figure, it can be seen that the research on this topic is extensive. In addition to the most common keywords, the current yield, semi-arid zone, post-flowering drought, growth and development, fertility, and photosynthetic characteristics are the main focal points of the research in this field. It can be seen that the semi-arid zone and post-flowering drought represent hotspots in the research on drought stress, and the yield, growth and development, fertility, and photosynthetic characteristics are considered in relation to the effects of drought stress on spring wheat during the fertility period.
The effects of drought stress on wheat plants during different fertility periods are directly related to the wheat yield. From the perspective of the whole fertility period, delayed sowing under high temperatures and drought causes an increase in the number of days constituting the nutritional growth stage, leading to a gradual decrease in the duration of postponement. There is also a decrease in the number of days constituting the reproductive growth stage, which shows an advancing trend on average. Moreover, the postponement of sowing and the advancement of maturity are the main factors associated with the shortening of the fertility period in spring wheat [71]. It has also been found that, with a delay in the sowing period, the wheat yield fluctuates, exhibiting a unimodal curve. Thus, coordinating the allocation of water and heat resources with the wheat sowing period is important to realize high wheat yields [72,73]. Er Y. H. et al. found that the length of spring wheat’s fertility period is negatively correlated with the early and late sowing period, and the effective cumulative temperature decreases accordingly due to drought and the late sowing in the early fertility period [74]. Mei L. et al. found that, with a delayed sowing period, the wheat fertility period is shortened, while the seedling emergence period is extended [75]. Zhang K. et al. found that the later the sowing date, the slower the nutrient growth, the higher the peak leaf area index, and the slower the leaf senescence [76]. Cui Y. Z. et al. found that the wheat yield increased and then decreased with delayed sowing [77]. Zhang Z. Z. et al. found that delayed sowing reduced the number of days constituting the entire reproductive period in wheat, resulting in the insufficient accumulation of nutrients and a decrease in the wheat yield [78].
When plants were subjected to drought stress, dry matter was redistributed between the above- and belowground parts of the plant. This was typically characterized by an increase in the biomass of the belowground part and a decrease in the leaf area index (LAI) and stem partitioning ratio (SPR). Leaf photosynthesis was inhibited, the light compensation point and light saturation point decreased significantly, and the apparent quantum rate decreased continuously with the increase in the stress intensity. Nutrient deficiencies promoted protein catabolism, and plant senescence was significantly accelerated, causing a reduction in the yield and grain quality in spring wheat [79,80]. During different fertility stages, it was found that the physiological and biochemical characteristics of wheat leaves under drought stress decreased to a certain extent. Drought stress during seed germination affects the wheat germination rate and germination potential; it also significantly reduces the wheat seedling biomass, dry matter conversion, and soluble sugar content. Moreover, biomass is preferentially allocated to the root system, disrupting the coordination of the root–crown ratio and reducing the efficiency of resource utilization [81]. Drought stress at the tiller stage causes leaf tiller degeneration and death, resulting in a lower leaf area index and reduced water consumption in wheat [82]. The destruction of the chlorophyll structure in flag leaf cells and the reduction in protective enzyme activity allow excess reactive oxygen radicals to damage the membrane system, consequently accelerating leaf senescence [83,84]. The period from nodulation to the tassel stage exhibits vigorous growth, and the growth status of spring wheat shifts from nutrient growth to reproductive growth. The plant is able to resist drought stress by reducing the biomass of the aboveground part and increasing that of the belowground part, as well as increasing the density of the root length to promote the absorption of water and accelerating the metabolism of the chloroplasts to replenish the nutrients required for reproductive fruiting. Moreover, the leaf area is reduced to the minimum in the tasseling stage [85,86]. Under the same degree of drought stress, the magnitude of the decrease in each photosynthetic index and the chlorophyll fluorescence parameter was found to be greater at the filling stage than at the flowering stage. Post-flowering drought caused a decrease in the soluble protein content of each functional leaf in spring wheat, and the trend consisted of a gradual decrease from the physiological upper level to the physiological lower level [87]. In this crop, under warming or water deficit conditions, the yield and fertile grain size decreased with increasing stress, posing a serious threat to the yield and grain quality [88,89].

Table 1.
Yield losses at vegetative growth stages under drought in wheat.
| Vegetative stage | Yield loss (%) | Reference |
|---|---|---|
| Early season stress | 22 | [90] |
| Midseason stress | 58 | [90] |
| Booting stage | 20.74 | [91] |
| Tillering stage | 46.85 | [91] |
| 1000-grain weight (vegetative stage) | 38.67 | [92] |
| Earlier stages | 79.70 | [93,94] |
| Spike length (vegetative stage) | 16.90 | [92] |
| Number of spikelets per spike (vegetative stage) | 28.63 | [92] |
| Grains number (vegetative stage) | 72.51 | [92] |
| Grains number (vegetative stage) | 61.38 | [92] |
Table 2.
Yield losses at reproductive growth stages under drought in wheat.
| High grain protein content, fewer days to physiological maturity, smaller kernel weight and diameter, less grain yield | Not applicable | [95] |
| Less grain yield (drought-tolerant variety) | 43 | [96] |
| Less grain yield (drought-sensitive variety) | 26 | [96] |
| 1000-grain weight | 18.29 | [18] |
| 5 | [27] | |
| 1000-grain weight (anthesis stage) | 38.67 | [92] |
| Biological yield | 10 | [18] |
| Maximum grain yield | 22 | [18] |
| Decreased seed number | 64 | [27] |
| Grain formation stage | 101.23 | [91] |
| Grain formation stage | 65.5 | [93,94] |
| Number of spikes | 19.85 | [92] |
| Number of spikes (anthesis stage) | 15.79 | [92] |
| Spike length (anthesis stage) | 16.90 | [92] |
| Number of spikelets per spike (anthesis stage) | 26.20 | [92] |
| Grain number (anthesis stage) | 72.51 | [92] |
| Grain yield (anthesis stage) | 64.46 | [92] |
Growth is sensitive to drought and can be affected by reduction in turgor pressure, whose reason is low turgor pressure,water stress quenches cell expansion and growth. However, when turgor pressure bigger than the cell wall yield, cell expansion can occur [97,98]. Osmotic adjustment is a remarkable part of plants’ physiology by which they respond to water deficits [29,99,100,101,102,103]. Yield losses at vegetative growth and reproductive stages under drought in wheat are provided in Table 1 and Table 2.
3.2. Leaf Water Potential in Response to Drought Stress in Spring Wheat
WThe water potential is an important indicator of the plant’s water status or the degree of water deficit. In particular, the leaf water potential is the most sensitive physiological indicator, reflecting the plant’s water surplus or deficit; thus, it directly reflects the water status of the plant body. Studies have shown that maintaining the stability of the leaf water potential is important to enhance the drought resistance of rice [104,105]. It has also been pointed out that the daily variation in the leaf water potential in spring wheat shows that it tends to increase and then decrease, reaching its highest in the early morning and decreasing to its lowest level in the late afternoon [106]. The early morning leaf water potential reflects the recovery of the plant from a water deficit, and the midday leaf water potential reflects the maximum water deficit in the plant [107]. With decreased soil moisture content and increased evapotranspiration under mild drought stress, the plant establishes a larger water potential difference in the soil–plant–atmosphere continuum (SPAC) by lowering the leaf water potential. This causes water to be transported upward through the water conductance pathway to reach the leaves [108]. In the soil–plant pathway, the soil water potential is positively correlated with the leaf water potential. The leaf water conductance pathway consists of the xylem pathway and extra-xylem pathway. Under severe drought stress, when the leaf water potential decreases beyond a certain threshold, the low-resistance xylem pathway forms an embolism. The resistance to the water supply in the aboveground part increases, and the lack of water reduces the number of chloroplasts, causing the senescence and death of the leaves [109]. Drought tolerance in wheat drives it to attenuate this suppression through different water supply or self-protection mechanisms. In a study, it was found that, with increasing drought stress, the transpiration water consumption of the plant was negative at night, which suggests the existence of water backsorption from the atmosphere to the plant in the plant–atmosphere pathway. This serves to maintain the leaf water potential and ensure a normal xylem water supply to the plant when seedlings are subjected to severe drought stress [110].
An analysis of the covariate mapping shown in Figure 2 indicates that most of the previous studies on the leaf water potential in wheat have focused on the stomatal conductance and photosynthetic characteristics. It can be seen from the inter-nodal connecting lines that, in wheat, the photosynthetic characteristics are closely linked to the yield and leaf senescence, and stomatal conductance is mainly limited by water stress. The leaf net photosynthetic rate is closely related to the leaf water potential. It was found that when the stress was mild, the leaf net photosynthetic rate appeared to be low near noon, which might indicate that the plant undergoes a period of respite. Moreover, the net photosynthetic rate was lower than that in the blank control group, even reaching half of this value throughout the day when the stress was severe. Mild drought stress rehydration has a promotional effect on plant root and leaf growth, and plants show compensatory effects, where the leaf water potential and photosynthetic capacity are improved. However, severe drought stress can seriously impair water transport in the roots, stems, and leaves and irreversibly obstruct the physiological and biochemical activities of the plant, causing leaf shriveling, premature senescence, and even plant death [111,112,113]. In a study, it was found that a reduction in the leaf water potential under severe drought stress caused the destruction of the chloroplast structure, the blockage of chlorophyll synthesis, a reduction in protective enzyme activity, damage to the membrane system caused by the accumulation of reactive oxygen species in the organism, and a sharp decrease in photosynthesis, which was manifested in yellowing and senescence of the leaves [114]. Nitrogen application has a compensatory effect on plant water deficits, facilitates the recovery of the nocturnal water potential, enhances the antioxidant properties, and protects the photosynthetic mechanisms [115]. Drought increases the soluble sugar content within leaf cells, altering the osmotic potential and significantly increasing the vein density of the plant in order to increase the water transport channels and the contact area for water exchange with the leaf pulp; thus, the safety of water transport within the leaf is ensured [116,117,118]. Plant transpiration is linearly related to stomatal conductance, and the attenuation of transpiration under drought stress is mainly regulated by the stomatal length and width. Meanwhile, increasing the stomatal density of the lower epidermal leaves to ensure the CO2 supply improves wheat’s photosynthesis and increases the plant’s stress tolerance [119,120].
Some researchers studied crop water stress index (CWSI) and midday flag leaf water potential () on wheat (Triticum aestivum L. Adana 99) under the three different supplemental and conventional irrigation strategies using spinkler line-source system during 2014 and 2015 in Adana, Turkey. They found that is recommended when there is no water shortage; however, under water scarcity conditions , and , with higher WUE and relatively higher yields, are recommended [121]. A technique for studying variation in the accumulation of abscisic acid (ABA) in response to drought stress is decreased. The results was that there was considerable genotypic variation in the rate at which water potential decreased, partly explained by variation in plant size [122]. Identification and understanding of the mechanism of drought tolerance in wheat (Triticum aestivum L.) cultivars has long been a major goal of plant breeders. A test was conducted to illustrate Whether leaf water potential, osmotic potential, yield, and yield components could be used to differentiate apparent drought tolerance among the cultivars or not. It found that stress applied at either tillering or heading gave similar results but effects were more pronounced when stress was applied at heading. It was not possible to determine if osmotic adjustment contributed to the differences between cultivars in response to water stress [123].
Currently, the leaf water potential is the best indicator of a plant’s water status and it has been extensively investigated, but few studies have comprehensively examined the changes in water c irculation and storage in various tissue types under drought. Changes in the leaf water potential, as affected by the soil moisture status and atmospheric water and air pressure deficits, have been addressed in several studies, but most have investigated one of these aspects individually. There is a lack of research that links the two through the SPAC system to explore the similarities and differences in the patterns of response and bias regarding the changes in the leaf water potential.
3.3. Chlorophyll Content in Response to Drought Stress
Chloroplasts are the main sites for photosynthesis in plants, and photosynthetic pigments increase the energy available for photosynthetic carbon assimilation through the absorption, transfer, and conversion of light energy. This indirectly affects wheat’s growth and development and yield through effects on photosynthesis, responding to the rate of leaf photosynthesis and the leaf senescence process. This can be used, to a certain extent, to measure the plant’s resilience and strength [124]. A reduction in efficiency of photochemical, reduced Rubisco efficiency, gathering of stress metabolites (glutathione, MDHA, glybet, and polyamines), antioxidative enzymes (superoxide dismutase (SOD)), peroxidase(POD), catalase (CAT), ascorbate peroxidase(APX), glutathione reductase (GR), glutathione-S-transferase (GST), glutathine peroxidase (GP)), monodehydroascorbate reductase (MDHAR), and reduced ROS accumulation are biochemical responses of plants to drought stress. Tolerance to drought correlates with a positive response of plants’ antioxidant system [125].
As shown in Figure 3, the top ten keywords identified were drought stress, physiological characteristics, photosynthetic characteristics, antioxidant enzymes, chlorophyll, yield, photosynthesis, physiological indexes, drought resistance, seed germination, microstructure, and nitrogen metabolism. Overall, the chlorophyll content in spring wheat under drought stress can be studied from two perspectives: one involves the study of the chlorophyll content and chloroplast structure, and the other involves the study of the plant’s photosynthetic characteristics.
PChanges in activity of lipid peroxidation, chlorophyll, protein oxidation,and nucleic acids are crucial for the resistance of various plants to drought stress [126]. In one study, the photosynthetic pigment content increased slightly under the influence of mild drought and showed some drought tolerance, while the chlorophyll content decreased, and the decrease was greater with increasing drought stress [127]. Carotenoids are important antioxidant substances in plants and can participate in the dissipation of excess light energy in the plant photosynthetic system, mitigate and eliminate the damage caused by excited-state chlorophyll molecules, and maintain metabolic homeostasis [128,129]. A moisture deficit beyond a given threshold causes reactive oxygen radicals to accumulate beyond the scavenging capacity of antioxidants, and reactive oxygen species accumulate in large quantities [130]. During the observation of the ultrastructure of the chloroplast, the accumulation of reactive oxygen species caused damage to the chloroplasts’ membranes, the arrangement of cyst-like lamellae was disorganized and loose, the number of osmophilic granule particles increased significantly, and the structures and functions of the photosynthetic organs were destroyed [131,132,133]. It was also found that the degree of damage to the chloroplast structure was positively correlated with the degree of membrane lipid peroxidation. Moreover, drought stress induced the excessive production of reactive oxygen radicals, reduced the peroxidase activity and membrane lipid peroxidation, damaged the integrity of the membrane system, and disrupted the chloroplast structure, as well as causing the inhibition of chlorophyll synthesis, reduced light energy capture, and weakened photosynthesis [134]. Chlorophyll a was found to be more sensitive to drought, with structural damage leading to a significant decrease in its content, while the chlorophyll b content was essentially unchanged and the total chlorophyll content was significantly reduced. The ratio of chlorophyll a/b reflects the drought tolerance of a plant—the higher the ratio, the more drought-tolerant the plant is [135,136,137].
Drought stress at different growth stages significantly alters growth, yield, and quality traits of wheat, however, great variability exists among genotypes regarding their reponse to drought stress. There fore, determining the immpacts of drought stress on yield and quality traits would help to select the superior genotypes. Bin Wang et al. Investigated the impact of seasonal droughts on yield variation in different subregions of the wheat belt. They identified significant positive correlations between wheat yields and SPEI (Standardized Precipition Evaportranspiration Index). They observed that lower yield corresponded with lower SPEI values, vice versa. This indicates that there was a close correlation between wheat yields and SPEI, confirming that SPEI is an effective grought index. The results are consisitent with those from prior studies [138]. These correlation could play a important role in assessing the impacts of drought on wheat productivity.
Carbon and nitrogen substances are the main components associated with the wheat yield and nutritional quality. The carbon and nitrogen substances in the kernel originate from two sources: the re-transportation of substances stored in the nutrient organs prior to flowering and the direct input of photosynthesis products synthesized by wheat leaves during the plumping period. Different varieties rely on both sources to different extents [139]. Terminal drought reduces the leaf area index of wheat and its ability to synthesize photosynthesis products, which severely reduces the accumulation of photosynthesis products in the grain and the amount and rate of translocation of pre-flowering storage material in the plant’s organs, resulting in poor yield traits and ultimately leading to yield reductions [140].
3.4. Photosynthesis in Response to Drought Stress
Photosynthesis is the most important physiological and biochemical process in spring wheat, and balanced photosynthesis is essential for improved plant survival and agricultural benefits in terms of biomass and yield. However,drought stress (DS) strongly perturbs photosynthetic efficiency due to biochemical and diffusive limitations that reduce key photosynthetic components and close stomata. While stomatal closure, disrupted photosynthetica systems, over-reduced electron transport rates (ETR), partial hindrance of the Calvin cycle, and reduced pigment contents strongly affect the repertoire of photosynthetic processes under DS, chloroplast retrograde signalling also has a plausible role in preserving photosyntheic capacity. Progress in agronomic, genetic engineering approaches and isoprene regulation would help to rescue photosynthetic apparatus under DS [141,142,143] The stabilization of the photosynthetic efficiency under drought stress is the goal during the response to this type of stress. According to Figure 4, the most important keyword is “drought stress”, which encompasses the three aspects of photosynthetic characteristics, photosynthesis, and photosynthetic parameters. In a study of the related literature, it was found that the net photosynthetic rate, light saturation point, and dark respiration rate of spring wheat under drought stress showed a decreasing trend with the increase in the drought duration, and the magnitude of this decrease was proportional to the intensity of the stress [144,145,146]. Research on photosynthesis under drought stress has focused on stomatal limitations, chlorophyll, and the photosynthetic systems. Drought affects photosynthesis through a combination of stomatal and non-stomatal factors [147]. The mentioned study concluded that the reduction in the photosynthetic rate was mainly due to stomatal factors. Moisture stress decreases the stomatal conductance in spring wheat to reduce water loss; stomatal closure hinders CO2 uptake by the leaves, while a reduction in the dark respiration rate, light compensation point, and light saturation point reduces the photosynthetic carbon loss, enabling the plant to adapt to the lower net photosynthetic rate. Moreover, normal photosynthetic carbon cycling is inhibited, and the photosynthetic pigment content decreases, as well as the net photosynthesis [148]. However, it has also been suggested that the leaves respond to mild drought by closing the stomata to reduce transpiration, and the reduction in the intracellular CO2 content decreases the net photosynthetic rate. At this time, stomatal factors dominate; when the water stress exceeds a certain threshold, the water deficit leads to the destruction of the leaf’s photosynthetic apparatus, and the changes in the net photosynthetic rate and stomatal conductance are no longer linearly related, at which time non-stomatal factors dominate [149,150]. The decrease in the photosynthetic rate caused by stomatal factors can be quickly recovered via rehydration, and the plant shows compensatory effects, enhancing its photosynthetic capacity and improving its stress tolerance. Meanwhile, the decrease in the net photosynthetic rate caused by non-stomatal factors is irreversible, as the photosynthetic system is impaired and the leaves are curled and deformed, which can lead to the death of the plant in severe cases [151,152,153]. The leaf photosynthetic rate was found to be positively correlated with the soil moisture, which affected the photosynthetic rate by influencing the photosynthetically active area. Moreover, the effect of lower soil drought on the photosynthetic rate was greater than that of upper soil drought [154,155]. At the same time, a reduction in the water use efficiency reduces the basic efficiency of organic matter accumulation, and, in spring wheat, the upper leaves are the first to be affected; this is manifested by the curling and deformation of the flag leaf and the yellowing of the tip [156,157].
3.5. Chlorophyll Fluorescence Parameters in Response to Drought Stress
Leaf photosynthesis involves adaptive and protective mechanisms, and the photosynthetic system stabilizes the photosynthetic performance by increasing the rate of electron transfer in the non-Calvin cycle, converting excess light energy into heat, eliminating excess light energy, and avoiding damage to photosynthetic structures such as PSII and the electron transport chain, as evidenced by an increase in the non-photochemical quenching coefficient (NPQ) [160]. In a study, the qP and NPQ showed contrasting patterns of change, and the degree of change increased with the degree of drought stress. It was found that, under the action of antioxidant enzymes, PSII was able to mitigate the damage caused by photo-oxidative stress and stabilize the photosynthetic efficiency under photo-oxidative conditions by increasing the rate of electron transfer, enhancing the conversion of light energy, ensuring smooth electron transfer, and dissipating excess light energy. However, excessive drought stress leads to the extreme photoinhibition of PSII receptors, where electrons are unable to function properly, causing severe damage to the photosystem, from which it is difficult to recover. This will attenuate or disrupt the plant’s photochemistry, thereby causing premature plant failure [161,162]. At the same time, the leaf stomata are closed, and the insufficiency of the intracellular CO2 content limits the fixation of carbon via the Calvin cycle. The excessive electron transfer to O2 and the reduced activity of leaf superoxide dismutase (SOD) and catalase (CAT) lead to the accumulation of excess reactive oxygen species in the cell, and the cellular metabolism becomes dysfunctional, causing photo-oxidative damage [163].
LLeaf photosynthesis involves adaptive and protective mechanisms, and the photosynthetic system stabilizes the photosynthetic performance by increasing the rate of electron transfer in the non-Calvin cycle, converting excess light energy into heat, eliminating excess light energy, and avoiding damage to photosynthetic structures such as PSII and the electron transport chain, as evidenced by an increase in the non-photochemical quenching coefficient (NPQ) [160]. In a study, the qP and NPQ showed contrasting patterns of change, and the degree of change increased with the degree of drought stress. It was found that, under the action of antioxidant enzymes, PSII was able to mitigate the damage caused by photo-oxidative stress and stabilize the photosynthetic efficiency under photo-oxidative conditions by increasing the rate of electron transfer, enhancing the conversion of light energy, ensuring smooth electron transfer, and dissipating excess light energy. However, excessive drought stress leads to the extreme photoinhibition of PSII receptors, where electrons are unable to function properly, causing severe damage to the photosystem, from which it is difficult to recover. This will attenuate or disrupt the plant’s photochemistry, thereby causing premature plant failure [161,162]. At the same time, the leaf stomata are closed, and the insufficiency of the intracellular CO2 content limits the fixation of carbon via the Calvin cycle. The excessive electron transfer to O2 and the reduced activity of leaf superoxide dismutase (SOD) and catalase (CAT) lead to the accumulation of excess reactive oxygen species in the cell, and the cellular metabolism becomes dysfunctional, causing photo-oxidative damage [163].
As the raw material for photosynthesis, water supplies primary electrons and plays an important role in photochemical reactions. The inhibitory effects of water deficits on the photosynthetic electron transfer rate (ETR) and reaction center complex hinder photosynthesis, and the photosynthetic fluorescence parameters will show different degrees of fluctuation. In a study, it was found that the initial fluorescence () increased under drought stress, the maximum fluorescence () and photochemical quenching coefficient (qP) values decreased significantly, and the maximum photochemical rate () values changed from maintaining dynamic stabilization to a gradual decrease, where the decrease was positively correlated with the stress level [164,165,166]. denotes the intensity of chlorophyll fluorescence emission when the photosystem II (PSII) reaction center is fully open, and the degree of change is negatively correlated with the plant’s stress tolerance. An increase in under drought stress indicates that the chlorophyll structure is damaged, which affects the efficiency of PSII in capturing excitation energy and the primary response of photosynthesis [167]. The primary photochemical reaction is the initiation of light energy conversion, which is operated by both PSII and PSI. The light energy absorbed by the plant undergoes the PSII reaction, and the released electrons are directed along the electron carrier in the cystoid membrane to PSI, driving the formation of the reducing substance NADPH [168]. The reduction in qP under the combined effect of drought and high temperatures indicates that both the light energy transfer efficiency and photosynthetic phosphorylation in PSII are impeded, severely curtailing the process of electron flow from the oxidizing side of PSII to the reaction centers in PSII, and the capacity for photosynthesis is weakened [169].
3.6. Drought Stress and Wheat at the Molecular Level
The selection, breeding, and identification of drought-tolerant varieties have, to a great extent, relied on the observation of physiological traits. Based on previous research, it has been concluded that drought-tolerant wheat has a higher root–crown ratio and chlorophyll content and lower stomatal conductance and leaf water potential [170,171]. Some genes are known to be influenced and produced different types of drought stress related proteins and enzymes including dehydrins [172], vacuolar acid invertase [173], glutathione S-transferase (GST) [174], and late embryo abundant (LEA) protein [175]; expression of ABA genes and production of proteins like RAB, rubisco, helicase, proline, and carbohydrates are molecular basis of drought tolerance. In wheat seedling stage, a lot of research are conducted in gene expression, but the most sensitive stage is the junction stage which is susceptible to drought. Junction stage is an important linkage point between the vegetative and flowering growth stage, which is vital for development and reproduction [176]. For example, Sivamani et al. [94] indicated that HVA1 gene assists to increase wheat growth under drought stress. Proline is a important protein in water stress tolerance, which can be produced from pyrroline-5-carboxylate synthetase or P5CR, and the responsible gene for this enzyme has been distinguished in some crops [177,178,179]. In photosystem II (PS II) reaction center, psbr has an indispensable task in oxidation of water [180], and in Calvin cycle, Rubisco is the key enzyme under drought stress [181]. Some plant proteins can be over-expressed including late embryogenesis abundant (LEA) that are saved in vegetative tissues during desiccation of seeds under drought stress. LEA proteins are shaped by drought stress and their size in wheat studies reach 200 kDa (Wcs 200) [182,183,184]. Researchers cost a lot of time to engineer LEA producing genes for promoting crop drought stress resistance. PMA1959 and PMA80 improved water deficit resistance in rice [185]. In wheat, the Em gene of wheat which encodes LEA protein first groud has been vastly researched [186,187,188]. In durum wheat, Td27e, Td29b, and Td16, gene transcripts were saved late in embryogenesis and throughout seed development [189]. In seeds,transcripts of Td11 gene were presented whereas no transcripts of Td25a gene were detected. Wheat V-PPase genes, TaVP3, TaVP2, and TaVP1 were investigated by Wang et al. [190]. TaRZF70 as a RING-H2 zinc finger gene presented various responsible genes in wheat for drought stress upregulated in leaf and downregulated in root [191]. Another genes expressed in wheat root were TaRZF38 and TaRZF70. However, TaRZF74 and TaRZF59 were mainly expressed in embryo and endosperm. TACCGACAT, the 9-bp consensus sequence, was first distinguished in the promoter of Arabidopsis rd29A/lti78 and presented to be vital for drought induction in abscisic acid absence [192]. The function and express about DRE-binding (DREB) proteins have detected by some researchers [193,194]. Lucas et al. Used a sequence of putative DREB labelled DREB3A from wheat (TaDREB3A, Gen bank ID: AY781349) to seclude a DREBfrom wild wheat ( T. turgidum ssp. dicoccoides) and to detect its function in higher drought resistance. The results found that DREB proteins are numerous and vastly upreguleted in reaction to drought in root tissue rather than leaf [194]. Additionally RD gene (responsive to desiccation) are also involved in drought stress [195,196]. RD gene comprise two parts: one is expression of regulatory gene and signal direction during the crops’ reaction to stress, the other is proteins which directly protect cells from stresses [197]. There are about 265 genes detected at the junction stage and 146 genes distinguished at the seedling stage in response to drought stress in wheat.
The limited success of the physiological and molecular breeding approaches now suggests that a careful rethink is needed of our strategies in order to understand better and breed for drought tolerance. Genomics technologies were seen as a path to understand the genetic and environmental complexity of drought stress. To be relevant to breeding programs, genomic studies must consider the nature of drought stress in the target environment and use plant material and phenotyping techniques that relate to field conditions [198,199]. Breeding for drought and heat stress tolerance was initiated by the International Wheat Improvement Network (IWIN) led by the International Maize and Wheat Improvement Centre (CIMMYT) established in 1966 in Mexico and the International Center for Agricultural Research in the Dry Areas (ICARDA)/Lebanon established in 1977 and partnered with Egypt in 1979. Both centres adopted complementary breeding technologies involving conventional and molecular breeding approaches to develop high-yielding germplasm with tolerance to heat and drought stress, diseases and insect pests with acceptable end-use qualities [200,201]. The major limiting factor for breeding progress and genetic gains is the reduced genetic diversity [205]. Some assessments revealed inconsistency contributed by different plant breeding methods perceived to reduce genetic gains [203]. The use of genetic resources such as landraces, obsolete lines and modern pure or breeding lines have potential to contribute to increased genetic gains [204]. A research programme for increasing drought tolerance of wheat should tackle the problem in a multi-disciplinary approach, considering interaction between multiple stressses and plant phenology, and integrating the physiological dissection of drought-tolerance traits and the genetic and genomics tools, such as quantitative trait loci (QTL), microarrays, and transgenic crops [205].
The rapid improvements in modern biomolecular technology have facilitated research on the relationship between abiotic stress and wheat at the molecular level, providing a basis for improvements in the drought tolerance of wheat and for the selection and breeding of resistant wheat varieties. The current research is aimed at addressing the changes in wheat’s physiological traits by identifying and utilizing drought-tolerance-related genes. Such genes protect plants against drought by encoding functional proteins that act both directly and indirectly as transcription factors. Proteins are the direct functional molecules of organisms, and functional proteins mainly include antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT); enzymes involved in the synthesis of osmoregulatory substances, such as glutamate synthetase; and proteins that protect the cellular structure, such as thermokinins and proteases [206,207]. In a study, it was found that heat stress reduced the total protein content and caused the most severe damage to the photosynthetic proteins and enzymes involved in metabolism. In contrast, heat-stimulated proteins were rapidly synthesized to repair the damaged cytoskeleton through phosphorylation reactions, and the level of peroxidase was increased to combat oxidative senescence in wheat cells [208,209]. The amino acid sequence of HSP70 depolymerizes proteins and attenuates organismal damage. TaAQP genes are involved in the stress response under the high expression of water channel proteins, which selectively control the flow of water and ionic molecules inside and outside of the cell membrane system; this is closely related to the leaf water potential [210]. Transcription factors regulate gene-specific transcription through binding to promoters. Changbiao Wang et al. found that, as the primary transcription factors, B-type ARR transcription factors regulate plants’ hormone responses, as well as regulating cell growth and development in response to drought [211]. Du P. P. et al. found that the TaNF-YB4 gene, which regulates downstream gene transcription in the nuclear factor Y (NF-Y) transcription factor, resists adverse conditions by maintaining the dynamic balance of sugar and nitrogen accumulation [212]. Genes can also improve plants’ drought tolerance by directly or indirectly inducing hormone synthesis [213,214]. With the continued, in-depth exploration of wheat, new genes are constantly being discovered and characterized.
4. Conclusions
Climate change is projected to decrease global wheat production by - 1.9% by mid-century. Top wheat-producing countries such as India, Russia, Australia and Pakistan are projected to have declining wheat yields. Wheat breeding with new traits is a climate change adaptation option, however its effect will vary among different regions and limited under soil conditions (i. e., intercropping, mulching, and crop rotation) and local environmental conditions (e. g., rainfall, temperature and soil nutrients) where water and N stress limit benefits of traits for high-temperature tolerance, early vigor, and delayed flowering adaptations. Therefore, future research should target improving prevailing stresses concurrently, to achieve improved grain yield and quality of wheat under water limited conditions.
Strategies such as breeding high-yielding cultivars, high-throughput phenotyping, and improving irrigation and water use efficiency are extremely important. Understanding the environmental variables and agronomic factors that determine wheat’s response to drought and yield is critical. The exploitation of genetic resources such as landraces, synthetic lines and wild relatives will help to identify the wealth of important alleles that can be used in breeding and improvement programs. Progress in molecular markers and marker technologies such as QTL analysis and detection could speed up genetic selections for prominent breeding traits thus reducing the breeding cycle. Therefore, elucidation of gene loci for important breeding traits that promote adaptation and drought tolerance in wheat and their expression patterns in response to drought stress will provide a solid foundation for gene-based breeding approaches and strategies to improve germplasm for adaptation to multiple and specific environments. Although plant breeding provides strategies to develop drought-resistant cereal species reducing yield losses under unpredictable climate change, integration into the local climatic context should also be a key consideration in the application of technological innovations.
Author Contributions
Conceptualization, Chunying Li and Chunlong Zhang; methodology, Yuting Feng; software, Chuying Li and Yuting Feng; validation, Yuting Feng, Chunlong Zhang and Fengyin Zhang; formal analysis, Chunying Li; investigation, Yuting Feng; resources, Chunying Li; data curation, Yuting Feng and Fengyin Zhang; writing—original draft preparation, Yuting Feng and Chunying Li; writing—review and editing, Chunying Li, Chunlong Zhang and Yuting Feng; visualization, Yuting Feng; supervision, Chunying Li; project administration, Chunying Li; funding acquisition, Chunying Li. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Key Laboratory Open Funding Project of the High-level Talent Scientific Research Start-up Project of West Anhui University(AUCIEERC-2022-03), West Anhui University of Science and Technology, School-level Natural Key Programs (WXZR202212) and 2022 Key Projects on Educational and Teaching Research at the School Level of West Anhui University (wxxy2022108).
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgments
We thank the anonymous reviewers for their constructive comments and suggestions regarding the revision of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, Q.; Li, Y.; Huang, G.; Wang, H.; Li, Y.; Shen, Z. Multivariate time series convolutional neural networks for long-term agricultural drought prediction under global warming. Agric. Water Manag. 2024, 292, 12. [Google Scholar] [CrossRef]
- Colombo, S.M.; Rodgers, T.F.M.; Diamond, M.L.; Bazinet, R.P.; Arts, M.T. Projected declines in global DHA availability for human consumption as a result of global warming. AMBIO 2020, 49, 865–880. [Google Scholar] [CrossRef]
- Zhao, G.C.; CHang, X.H.; Wang, D.M.; Tao, Z.; Wang, Y.; Yang, Y.; Zhu, Y. Overview of wheat production and its development. Crops 2018, 4, 7. [Google Scholar] [CrossRef]
- Li, Y.; Ye, W.; Wang, M.; Yan, X. Climate change and drought: a risk assessment of crop-yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Lesk, C.; Anderson, W.; Rigden, A.; Coast, O.; Jägermeyr, J.; McDermid, S.; Davis, K.F.; Konar, M. Compound heat and moisture extreme impacts on global crop yields under climate change. Nat. Rev. Earth Environ. 2022, 3, 872–889. [Google Scholar] [CrossRef]
- Archer, E.R.M.; Landman, W.A.; Tadross, M.A.; Malherbe, J.; Weepener, H.; Maluleke, P.; Marumbwa, F.M. Understanding the evolution of the 2014–2016 summer rainfall seasons in southern Africa: Key lessons. Clim. Risk Manag. 2017, 16, 22–28. [Google Scholar] [CrossRef]
- Saleem, F.; Arshad, A.; Mirchi, A.; Khaliq, T.; Zeng, X.; Rahman, M.; Dilawar, A.; Pham, Q.B.; Mahmood, K. Observed Changes in Crop Yield Associated with Droughts Propagation via Natural and Human-Disturbed Agro-Ecological Zones of Pakistan. Remote. Sens. 2022, 14, 2152. [Google Scholar] [CrossRef]
- Yuan, C.; Yamagata, T. Impacts of IOD, ENSO and ENSO Modoki on the Australian Winter Wheat Yields in Recent Decades. Sci. Rep. 2015, 5, 17252–17252. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Kansara, P.; Dandridge, C.; Lakshmi, V. Drought monitoring using high spatial resolution soil moisture data over Australia in 2015–2019. J. Hydrol. 2021, 594. [Google Scholar] [CrossRef]
- van Dijk, A.I.J.M.; et al. The Millennium Drought in southeast Australia (2001–2009): natural and human causes and implications for water resources, ecosystems, economy, and society. Water Resour. Res. 2013, 49, 1040–1057. [Google Scholar] [CrossRef]
- Nguyen, H.; Wheeler, M.C.; Hendon, H.H.; Lim, E.-P.; Otkin, J.A. The 2019 flash droughts in subtropical eastern Australia and their association with large-scale climate drivers. Weather. Clim. Extremes 2021, 32. [Google Scholar] [CrossRef]
- BOM. Special climate statement 70 update-drought conditions in Australia and impact on water resources in the murray–darling basin. 2020. Available online: http://www.bom.gov.au/ climate/current/statements/scs70.pdf.
- Zhang, Q.; Yao, Y.; Li, Y.; Huang, J.; Ma, Z.; Wang, Z.; Wang, S.; Wang, Y.; Zhang, Y. Causes and Changes of Drought in China: Research Progress and Prospects. J. Meteorol. Res. 2020, 34, 460–481. [Google Scholar] [CrossRef]
- Wang, F.; Lai, H.; Li, Y.; Feng, K.; Zhang, Z.; Tian, Q.; Zhu, X.; Yang, H. Dynamic variation of meteorological drought and its relationships with agricultural drought across China. Agric. Water Manag. 2022, 261, 107301. [Google Scholar] [CrossRef]
- A Araujo, D.S.; Marra, F.; Merow, C.; I Nikolopoulos, E. Today’s 100 year droughts in Australia may become the norm by the end of the century. Environ. Res. Lett. 2022, 17, 044034. [Google Scholar] [CrossRef]
- Kirono, D.G.; Round, V.; Heady, C.; Chiew, F.H.; Osbrough, S. Drought projections for Australia: Updated results and analysis of model simulations. Weather. Clim. Extremes 2020, 30, 100280. [Google Scholar] [CrossRef]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef] [PubMed]
- Askarimarnani, S.S.; Kiem, A.S.; Twomey, C.R. Comparing the performance of drought indicators in Australia from 1900 to 2018. Int. J. Clim. 2021, 41, E912–E934. [Google Scholar] [CrossRef]
- Ribeiro, A.F.; Russo, A.; Gouveia, C.M.; Páscoa, P. Copula-based agricultural drought risk of rainfed cropping systems. Agric. Water Manag. 2019, 223. [Google Scholar] [CrossRef]
- Bönecke, E.; Breitsameter, L.; Brüggemann, N.; Chen, T.; Feike, T.; Kage, H.; Kersebaum, K.; Piepho, H.; Stützel, H. Decoupling of impact factors reveals the response of German winter wheat yields to climatic changes. Glob. Chang. Biol. 2020, 26, 3601–3626. [Google Scholar] [CrossRef]
- Mäkinen, H.; Kaseva, J.; Trnka, M.; Balek, J.; Kersebaum, K.; Nendel, C.; Gobin, A.; Olesen, J.; Bindi, M.; Ferrise, R.; et al. Sensitivity of European wheat to extreme weather. Field Crop. Res. 2018, 222, 209–217. [Google Scholar] [CrossRef]
- Riedesel, L.; Möller, M.; Horney, P.; Golla, B.; Piepho, H.-P.; Kautz, T.; Feike, T. Timing and intensity of heat and drought stress determine wheat yield losses in Germany. PLOS ONE 2023, 18, e0288202. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, J.; Offermann, F.; Soder, M.; Frühauf, C.; Finger, R. Extreme weather events cause significant crop yield losses at the farm level in German agriculture. Food Pol. 2022, 112, 102359. [Google Scholar] [CrossRef]
- Leng, G. Keeping global warming within 1.5 °C reduces future risk of yield loss in the United States: a probabilistic modeling approach. Sci. Total Environ. 2018, 644, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kramer, P.J.; Boyer, J.S. Water Relations of Plants and Soils; Academic Press: New York, NY, USA, 1995. [Google Scholar]
- Salekdeh, G.; Siopongco, J.; Wade, L.; Ghareyazie, B.; Bennett, J. A proteomic approach to analyzing drought- and salt-responsiveness in rice. Field Crop. Res. 2002, 76, 199–219. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The Combined Effect of Drought Stress and Heat Shock on Gene Expression in Tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Marôco, J.P.; Pereira, J.S. Understanding plant responses to drought — from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Denby, K.; Gehring, C. Engineering drought and salinity tolerance in plants: lessons from genome-wide expression profiling in Arabidopsis. Trends Biotechnol. 2005, 23, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Carbo, M.; Taylor, N.L.; Giles, L.; Busquets, S.; Finnegan, P.M.; Day, D.A.; Lambers, H.; Medrano, H.; Berry, J.A.; Flexas, J. Effects of Water Stress on Respiration in Soybean Leaves. Plant Physiol. 2005, 139, 466–473. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.J.S.; Davies, W.J. Keeping in touch: responses of the whole plant to deficits in water and nitrogen supply. Adv. Bot. Res. 1996, 22, 229–300. [Google Scholar]
- Aliyeva, D.R.; Gurbanova, U.A.; Rzayev, F.H.; Gasimov, E.K.; Huseynova, I.M. Biochemical and Ultrastructural Changes in Wheat Plants during Drought Stress. Biochem. (Moscow) 2023, 88, 1944–1955. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Marôco, J.P.; Pereira, J.S. Understanding plant responses to drought — from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Denby, K.; Gehring, C. Engineering drought and salinity tolerance in plants: lessons from genome-wide expression profiling in Arabidopsis. Trends Biotechnol. 2005, 23, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Carbo, M.; Taylor, N.L.; Giles, L.; Busquets, S.; Finnegan, P.M.; Day, D.A.; Lambers, H.; Medrano, H.; Berry, J.A.; Flexas, J. Effects of Water Stress on Respiration in Soybean Leaves. Plant Physiol. 2005, 139, 466–473. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.J.S.; Davies, W.J. Keeping in touch: responses of the whole plant to deficits in water and nitrogen supply. Adv. Bot. Res. 1996, 22, 229–300. [Google Scholar]
- Szegletes, Z.; Erdei, L.; Tari, I.; Cseuz, L. Accumulation of osmoprotectants in wheat cultivars of different drought tolerance. Cereal Res. Commun. 2000, 28, 403–410. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant, Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant responses to drought, acclimation, and stress tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Ouvrard, O.; Cellier, F.; Ferrare, K.; et al. Differential expression of water stress-regulated genes in drought tolerant or sensitive sunflower genotypes. In Proceedings of the International Conference of Integrated Studies on Drought Tolerance of Higher Plants, Inter Drought; 1995; vol. 95. [Google Scholar]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.D.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant, Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. CMLS 2000, 57, 779–795. [Google Scholar] [CrossRef]
- Davies, W.J.; Zhang, J. Root Signals and the Regulation of Growth and Development of Plants in Drying Soil. Annu. Rev. Plant Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene Expression and Signal Transduction in Water-Stress Response. Plant Physiol. 1997, 115, 327–334. [Google Scholar] [CrossRef]
- Pei, Z.-M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.M.; Zhao, J.; Scandalios, J.G. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J. 2000, 22, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.A. Classification of Genes Differentially Expressed during Water-deficit Stress in Arabidopsis thaliana: an Analysis using Microarray and Differential Expression Data. Ann. Bot. 2002, 89, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, K.; Zagdańska, B. Genotype-dependent prote- ’olytic response of spring wheat to water deficiency. J. Exp. Bot. 2001, 52, 1455–1463. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Patanè, C.; Sanzone, E.; Copani, V.; Foti, S. Effects of soil water content and nitrogen supply on the productivity of Miscanthus × giganteus Greef et Deu. in a Mediterranean environment. Ind. Crops Prod. 2007, 25, 75–88. [Google Scholar] [CrossRef]
- Sankar, B.; Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Somasundaram, R.; Panneerselvam, R. Relative efficacy of water use in five varieties of Abelmoschus esculentus(L.) Moench under water-limited conditions. Colloids Surf. B 2008, 62, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of Water Stress on Photosynthesis, Yield, and Water Use Efficiency in Winter Wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Bukhari; Adnan, M.; Ahmad, Z.; Ashraf, M.Y.; Afzal, M.; Nawaz, F.; Nafees, M.; Jatoi, W.N.; Malghani, N.A.; Shah, A.N.; et al. Silicon mitigates drought stress in wheat (Triticum aestivum L.) through improving photosynthetic pigments, biochemical and yield characters. Silicon 2021, 13, 4757–4772. [Google Scholar] [CrossRef]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon 2021, 7, e05908. [Google Scholar] [CrossRef] [PubMed]
- Azam, A.; Ahmed, A.; Wang, H.; Wang, Y.; Zhang, Z. Knowledge structure and research progress in wind power generation (WPG) from 2005 to 2020 using CiteSpace based scientometric analysis. J. Clean. Prod. 2021, 295. [Google Scholar] [CrossRef]
- Ahsan, M.M.; Cheng, W.; Hussain, A.B.; Chen, X.; Wajid, B.A. Knowledge mapping of research progress in vertical greenery systems (VGS) from 2000 to 2021 using CiteSpace based scientometric analysis. Energy Build. 2022, 256. [Google Scholar] [CrossRef]
- WAN, M.; SONG, Y.; SUN, X.; WANG, F.; TAN, Y. Citespace-based visual analysis of overseas research frontiers and hot spots about home-based care. Chinese General Practice 2020, 23, 1465. [Google Scholar]
- Junjia, Y.; Alias, A.H.; Haron, N.A.; Abu Bakar, N. A Bibliometric Review on Safety Risk Assessment of Construction Based on CiteSpace Software and WoS Database. Sustainability 2023, 15, 11803. [Google Scholar] [CrossRef]
- Peng, W.; Haron, N.A.; Alias, A.H.; Law, T.H. Knowledge Map of Climate Change and Transportation: A Bibliometric Analysis Based on CiteSpace. Atmosphere 2023, 14, 434. [Google Scholar] [CrossRef]
- Pan, M.; Zhang, S. Visualization of Prediction Methods for Wildfire Modeling Using CiteSpace: A Bibliometric Analysis. Atmosphere 2023, 14, 1009. [Google Scholar] [CrossRef]
- Alviz-Meza, A.; Orozco-Agamez, J.; Quinayá, D.C.P.; Alviz-Amador, A. Bibliometric Analysis of Fourth Industrial Revolution Applied to Material Sciences Based on Web of Science and Scopus Databases from 2017 to 2021. Chemengineering 2023, 7, 2. [Google Scholar] [CrossRef]
- Eriksson, C.P.; Johansson, N.; McNamee, M. The performance of wildfire danger indices: A Swedish case study. Saf. Sci. 2022, 159. [Google Scholar] [CrossRef]
- Dong, Y.; Ding, S.; Zhang, T.; Zhou, W.; Si, H.; Yang, C.; Liu, X. Visual analysis of hot spots and trends in research of meteorology and hemorrhagic fever with renal syndrome: a bibliometric analysis based on CiteSpace and VOSviewer. Front. Environ. Sci. 2024, 12, 1395135. [Google Scholar] [CrossRef]
- Chen, C.; Hu, Z.; Liu, S.; Tseng, H. Emerging trends in regenerative medicine: A scientometric analysis in CiteSpace. Expert Opin. Biol. Ther. 2012, 12, 593–608. [Google Scholar] [CrossRef]
- Zhao, X.; Nan, D.; Chen, C.; Zhang, S.; Che, S.; Kim, J.H. Bibliometric study on environmental, social, and governance research using CiteSpace. Front. Environ. Sci. 2023, 10. [Google Scholar] [CrossRef]
- Du, J.; Gao, J.J.; Suo, L.T.J.; Ci, W.D.Z. Response of spring wheat fertility to climate change in Tibet, 1991-2020. J. Triticeae Crops. 2021, 41, 1044–1054. [Google Scholar]
- Liu, Y.J.; Chen, Q.M.; Ge, Q.S.; Dai, J.H.; Qin, Y.; Dai, L.; Zou, X.T.; Chen, J. Modelling the impacts of climate change and crop management on phenological trends of spring and winter wheat in China. Agric. For. Meteorol. 2018, 248, 518–526. [Google Scholar] [CrossRef]
- Ruan, X.M.; Chen, X.; Yue, W.; Zhan, X.C.; Cong, X.H.; Du, H.Y.; Shi, F.Z.; Wu, D.X.; Luo, Z.X. Influence of sowing period on annual yield, fertility and utilization of temperature and light resources of rice and wheat in the coastal plain of Anhui Province, China. China Rich. 2021, 27, 38–46. [Google Scholar]
- Flohr, B.; Hunt, J.; Kirkegaard, J.; Evans, J.; Trevaskis, B.; Zwart, A.; Swan, A.; Fletcher, A.; Rheinheimer, B. Fast winter wheat phenology can stabilise flowering date and maximise grain yield in semi-arid Mediterranean and temperate environments. Field Crop. Res. 2018, 223, 12–25. [Google Scholar] [CrossRef]
- E, Y.H.; Huo, Z.G.; MA, Y.P.; XV, Y.J. Regional variability and climate adaptation of spring wheat fertility changes in northern China. Chin. J. Appl. Ecol. 2013, 33, 6295–6302. [Google Scholar]
- Mei, L.; Lian, Y.J.; Meng, F.Y.; Zhao, J.H.; LI, R.Z.; Wang, J.Y. The effect of late and over-late sowing on the growth, development and yield of winter wheat. Tianjin Agric. Sci. 2023, 29, 27–33. [Google Scholar]
- Zhang, K.; Li, Q.Z.; Wang, R.Y.; Guo, L.; Lei, J.; Wang, Y.B.; Wang, H.L. Effect of sowing period on growth, development and yield of spring wheat. Chin. J. Ecol. 2012, 31, 324–331. [Google Scholar] [CrossRef]
- Cui, Y.Z.; Lv, L.H.; Li, Q.; Meng, J.; Li, M.Y.; Jia, X.L. Effect of sowing date on grain quality and yield traits of wheat kernels. J. Hebei Agric. Sci. 2023, 27, 28–35. [Google Scholar]
- Zhen-zhen, Z.; Shuang, C.; Peng, F.; Nian-bing, Z.; Zhi-peng, X.; Ya-jie, H.; Fang-fu, X.; Bao-wei, G.; Hai-yan, W.; Hong-cheng, Z. Effects of sowing date and ecological points on yield and the temperature and radiation resources of semi-winter wheat. J. Integr. Agric. 2023, 22, 1366–1380. [Google Scholar]
- Liu, C.M.; Zhao, Y. Effects of drought stress on leaf microstructure and chlorophyll in medicinal chrysanthemums. Science Mosaic. 2017, 26–29. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Tabassum, T.; Nazir, U. Growth and developmental responses of crop plants under drought stress: a review. Zemdirbyste-Agriculture 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Shi, C.X.; Chen, T.; Wang, C.J.; Qin, X.L.; Liao, Y.C. Effect of drought stress on seed germination and seedling root-shoot biomass partitioning in wheat with different drought resistances. J. Triticeae Crops 2016, 36, 483–490. [Google Scholar]
- Zhao, H.; Dai, Y.B.; Jiang, D.; Jing, Q.; Cao, W.X. Effects of drought and waterlogging at high temperatures on photosynthetic characteristics and substance transport in flag leaves of winter wheat after anthesis. Chin. J. Appl. Ecol. 2007, 333–338. [Google Scholar]
- Wang, C.; Wang, L.X.; Ma, G.F.; Zhang, X.Y.; Li, Q. Response of morphological structure and dry matter allocation of spring wheat to different periods of drought stress in Ningxia irrigation area. Chin. J. Ecol. 2019, 38, 2049–2056. [Google Scholar] [CrossRef]
- Xiong, D.; Zhen, G.; Xiaonuo, S.; Dahong, B.; Jianhong, R.; Peng, Y.; Yanhong, C. Increasing temperature during early spring increases winter wheat grain yield by advancing phenology and mitigating leaf senescence. Sci. Total Environ. 2021, 812, 152557–152557. [Google Scholar]
- LI, Y.B.; ZHU, Y.N.; LI, D.X.; GAO, Y. Effects of stage drought and rehydration on growth and development, photosynthesis and yield of wheat. J. Irrig. Drain. 2018, 37, 76–82. [Google Scholar] [CrossRef]
- Science - Soil Science; Study Findings from University of Bonn Broaden Understanding of Soil Science (Estimation of the Impact of Precrops and Climate Variability On Soil Depth Differentiated Spring Wheat Growth and Water, Nitrogen and Phosphorus Uptake). Food Farm Week 2019, 195.
- Bheemanahalli, R.; Sunoj, V.S.J.; Saripalli, G.; Prasad, P.V.V.; Balyan, H.S.; Gupta, P.K.; Grant, N.; Gill, K.S.; Jagadish, S.V.K. Quantifying the Impact of Heat Stress on Pollen Germination, Seed Set, and Grain Filling in Spring Wheat. Crop. Sci. 2019, 59, 684–696. [Google Scholar] [CrossRef]
- WANG, H.L.; ZHANG, Q.; WANG, R.Y.; GAN, Y.T.; NIU, J.Y.; ZHANG, K.; ZHAO, F.N.; ZHAO, H. Effects of warming and precipitation changes on yield and quality of spring wheat in the semi-arid zone of Northwest China. Chin. J. Appl. Ecol. 2015, 26, 67–75. [Google Scholar] [CrossRef]
- Vignjevic, M.; Wang, X.; Olesen, J.E.; Wollenweber, B. Traits in Spring Wheat Cultivars Associated with Yield Loss Caused by a Heat Stress Episode after Anthesis. J. Agron. Crop. Sci. 2014, 201, 32–48. [Google Scholar] [CrossRef]
- Milad, I.; Wahba, L.E.; Barakat, M.N. Identification of RAPD and ISSR markers associated with flag leaf senescence under water-stressed conditions in wheat (Triticum aestivum L.). Aust. J. Crop Sci. 2011, 5, 337–343. [Google Scholar]
- Schneekloth, J.; Bauder, T.; Hansen, N. Limited irrigation management: principles and practices. 2012. Available online: http://www.ext.colostate.edu/pubs/crops/04720.html.
- Rizza, F.; Badeck, F.W.; Cattivelli, L.; Lidestri, O.; Di Fonzo, N.; Stanca, A.M. Use of a Water Stress Index to Identify Barley Genotypes Adapted to Rainfed and Irrigated Conditions. Crop. Sci. 2004, 44, 2127–2137. [Google Scholar] [CrossRef]
- Tuberosa, R.; Salvi, S. Genomics-based approaches to improve drought tolerance of crops. Trends Plant Sci. 2006, 11, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Sivamani, E.; Bahieldin, A.; Wraith, J.M.; Al-Niemi, T.; E Dyer, W.; Ho, T.-H.D.; Qu, R. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 2000, 155, 1–9. [Google Scholar] [CrossRef]
- Singh, T.; Paleg, I.; Aspinall, D. Stress Metabolism I. Nitrogen Metabolism and Growth in the Barley Plant During Water Stress. Aust. J. Biol. Sci. 1973, 26, 45–56. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Karthikeyan, B.; Jaleel, C.A.; Gopi, R.; Deiveekasundaram, M. Alterations in seedling vigour and antioxidant enzyme activities inCatharanthus roseus under seed priming with native diazotrophs. J. Zhejiang Univ. Sci. B 2007, 8, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Manivannan, P.; Kishorekumar, A.; Sridharan, R.; Panneerselvam, R. Studies on germination, seedling vigour, lipid peroxidation and proline metabolism in Catharanthus roseus seedlings under salt stress. South Afr. J. Bot. 2007, 73, 190–195. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant Responses to Water Stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Erdei, L.; Tari, I.; Csiszar, J.I.; et al. smotic stress responses of wheat species and cultivars differing in drought tolerance: some interesting genes (advices for gene hunting). Acta Biol. Szeged. 2002, 46, 63–65. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Plaut, Z. Crop plants: critical development stages of water. Encyclopedia of Water Science 2003, 95–100. [Google Scholar]
- Maathuis, F.J.M.; Filatov, V.; Herzyk, P.; Krijger, G.C.; Axelsen, K.B.; Chen, S.; Green, B.J.; Li, Y.; Madagan, K.L.; Sánchez-Fernández, R.; et al. Transcriptome analysis of root transporters reveals participation of multiple gene families in the response to cation stress. Plant J. 2003, 35, 675–692. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, P.G.; Cutforth, H.W. Comparative forage yield, water use, and water use efficiency of alfalfa, crested wheatgrass and spring wheat in a semiarid climate in southern Saskatchewan. Can. J. Plant Sci. 2005, 85, 877–888. [Google Scholar] [CrossRef]
- Papastefanou, P.; Zang, C.S.; Pugh, T.A.M.; Liu, D.; Grams, T.E.E.; Hickler, T.; Rammig, A. A Dynamic Model for Strategies and Dynamics of Plant Water-Potential Regulation Under Drought Conditions. Front. Plant Sci. 2020, 11, 373. [Google Scholar] [CrossRef]
- Du, H.L.; Wang, H.L.; Zhang, K.; Chen, P.; Wang, R.Y.; Sun, X.Y.; Huang, P.C. Changes in leaf water potential of spring wheat during the critical period of water demand under drought stress and its influencing factors. Agric. Res. Arid. Areas 2021, 39, 230–237. [Google Scholar]
- Sofo, A.; Dichio, B.; Montanaro, G.; Xiloyannis, C. Shade effect on photosynthesis and photoinhibition in olive during drought and rewatering. Agric. Water Manag. 2009, 96, 1201–1206. [Google Scholar] [CrossRef]
- Deng, X.; Ran, H.; Yv, T.G.; Peng, X.L.; Ji, S.S.; Hu, X.T. Response of stomatal conductance and leaf water potential to water and nitrogen stress during the filling period of seed-producing maize in the dry zone of Northwest China. Agric. Res. Arid. Areas 2020, 38, 253–258. [Google Scholar]
- Xv, J.X.; Wang, Q.; Gao, Y.; Sun, J.S.; Hou, Z.L. Effects of water and salt stress on the hydraulic properties of maize stem xylem. J. Irrig. Drain. 2020, 39, 45–51. [Google Scholar] [CrossRef]
- Li, J.Y.; Zhao, P.; Zhao, L.J. Effect of drought stress on transpiration water consumption of seedlings. Acta Ecol. Sin. 2002, 1380–1386. [Google Scholar]
- Fan, Y.; Lv, Z.; Ge, T.; Li, Y.; Yang, W.; Zhang, W.; Ma, S.; Dai, T.; Huang, Z. Night-Warming Priming at the Vegetative Stage Alleviates Damage to the Flag Leaf Caused by Post-anthesis Warming in Winter Wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 13. [Google Scholar] [CrossRef]
- Nan, S.R.; Luo, Y.Z.; Yv, S.M.; Tong, H.X.; HE, Y. Effects of water stress on water potential and biomass allocation in alfalfa leaves. Grassland and Turf. 2022, 42, 63–70. [Google Scholar] [CrossRef]
- Guo, J.S.; Cao, L.R.; Zhang, X.; Zhang, Q.J.; Wei, L.M.; Wang, Z.H.; Lu, X.M. Effects of exogenous ABA on photosynthetic capacity and stomatal opening of maize leaves under drought stress. Chinese Agricultural Science Bulletin 2019, 35, 31–35. [Google Scholar]
- Shokat, S.; Großkinsky, D.K.; Roitsch, T.; Liu, F. Activities of leaf and spike carbohydrate-metabolic and antioxidant enzymes are linked with yield performance in three spring wheat genotypes grown under well-watered and drought conditions. BMC Plant Biol. 2020, 20, 1–19. [Google Scholar] [CrossRef]
- Rafiq, M.; Jawad, H.; Javed, T.; Hussain, S.; Arif, M.; Ali, B.; Sultan, M.; Abbas, G.; Aziz, M.; Al-Sadoon, M.; et al. Improving Quantitative and Qualitative Characteristics of Wheat (Triticum aestivum L.) through Nitrogen Application under Semiarid Conditions. Phyton 2023, 92, 1001–1017. [Google Scholar] [CrossRef]
- Nardini, A.; Pedà, G.; La Rocca, N. Trade-offs between leaf hydraulic capacity and drought vulnerability: morpho-anatomical bases, carbon costs and ecological consequences. New Phytol. 2012, 196, 788–798. [Google Scholar] [CrossRef]
- Zhu, X.; Gong, H.; Chen, G.; Wang, S.; Zhang, C. Different solute levels in two spring wheat cultivars induced by progressive field water stress at different developmental stages. J. Arid. Environ. 2005, 62, 1–14. [Google Scholar] [CrossRef]
- Xv, H.M.; Shao, J.X.; Li, Y.Y. Response of leaf vein characteristics to water and nitrogen supply in wheat flag leaves and their relation to leaf hydrophysiological functions. Plant Nutrition and Fertilizer Science 2020, 26, 1636–1646. [Google Scholar]
- Liu, Y.Q.; Zhang, X.Y.; Bai, B. Analysis of differences in leaf stomatal density and morphology of wheat varieties of different ecotypes. Acta Agriculturae Boreali-Occidentalis Sinica 2023, 32, 677–684. [Google Scholar]
- Baloch, M.J.; Dunwell, J.; Khan, N.U.; Jatoi, W.A.; Khakhwani, A.A.; Vessar, N.F.; Gul, S. Morpho-physiological Characterization of Spring Wheat Genotypes under Drought Stress. Int. J. Agric. Biol. 2013, 15, 945–950. [Google Scholar]
- Alghory, A.; Yazar, A. Evaluation of crop water stress index and leaf water potential for deficit irrigation management of sprinkler-irrigated wheat. Irrig. Sci. 2018, 37, 61–77. [Google Scholar] [CrossRef]
- Quarrie, S.A.; Jones, H.G. Genotypic Variation in Leaf Water Potential, Stomatal Conductance and Abscisic Acid Concentration in Spring Wheat Subjected to Artificial Drought Stress. Ann. Bot. 1979, 44, 323–332. [Google Scholar] [CrossRef]
- Moustafa, M.A.; Boersma, L.; Kronstad, W.E. Response of Four Spring Wheat Cultivars to Drought Stress. Crop. Sci. 1996, 36, 982–986. [Google Scholar] [CrossRef]
- Hou, M.; Yao, N.; Li, Y.; Liu, F.; Biswas, A.; Pulatov, A.; Hassan, I. Better Drought Index between SPEI and SMDI and the Key Parameters in Denoting Drought Impacts on Spring Wheat Yields in Qinghai, China. Agronomy 2022, 12, 1552. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef]
- van Rensburg, L.; Krüger, G. Evaluation of Components of Oxidative Stress Metabolism for Use in Selection of Drought Tolerant Cultivars of Nicotiana tabacum L. J. Plant Physiol. 1994, 143, 730–737. [Google Scholar] [CrossRef]
- Xv, Y.P.; Pan, Y.D.; Ren, C.; Yao, Y.H.; Jia, Y.C.; Chen, W.Q.; Huo, K.C.; Bao, Q.J.; Zhao, F.; Zhang, H.Y. Effects of drought stress and rehydration on yield quality and chlorophyll content of beer barley. Gansu Agric. Sci. Technol. 2019, 19–24. [Google Scholar]
- Yang, X.Z.; Yang, L.; Qin, Y.; Liu, L.; Yang, H.; Xie, W. Effect of PEG-8000 simulated drought stress on chlorophyll and carotenoid content of potato seedlings grown in histoculture. Chin. Potato J. 2019, 33, 193–202. [Google Scholar]
- Khodabin, G.; Tahmasebi-Sarvestani, Z.; Rad, A.H.S.; Modarres-Sanavy, S.A.M. Effect of Drought Stress on Certain Morphological and Physiological Characteristics of a Resistant and a Sensitive Canola Cultivar. Chem. Biodivers. 2020, 17, e1900399. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Lee, J.G. Effect of Drought Stress on Chlorophyll Fluorescence Parameters, Phytochemical Contents, and Antioxidant Activities in Lettuce Seedlings. Horticulturae 2021, 7, 238. [Google Scholar] [CrossRef]
- Saha, S.; Begum, H.H.; Nasrin, S.; Samad, R. EFFECTS OF DROUGHT STRESS ON PIGMENT AND PROTEIN CONTENTS AND ANTIOXIDANT ENZYME ACTIVITIES IN FIVE VARIETIES OF RICE (ORYZA SATIVA L.). Bangladesh J. Bot. 2020, 49, 997–1002. [Google Scholar] [CrossRef]
- Song, Y.H.; Zhang, M.H.; Zhou, R.X.; Chen, X.Y.; Hua, X.; Qi, X.L.; Wang, Y.X.; Li, Y. Analysis of photosynthetic rate and chloroplast ultrastructure of Zheng mai 1860 flag leaves during filling period. J. Triticeae Crops 2023, 43, 744–752. [Google Scholar]
- Zhang, F.-J.; Zhang, K.-K.; Du, C.-Z.; Li, J.; Xing, Y.-X.; Yang, L.-T.; Li, Y.-R. Effect of Drought Stress on Anatomical Structure and Chloroplast Ultrastructure in Leaves of Sugarcane. Sugar Tech 2014, 17, 41–48. [Google Scholar] [CrossRef]
- Chen, B.W.; Qin, Z.H.; Zhang, Y.; Xiao, Y.F.; Zhang, X.N.; Liu, H.L. Physiological activity and endogenous hormone dynamics of Australian tea tree under drought stress. Shandong Agric. Sci. 2019, 51, 55–59. [Google Scholar] [CrossRef]
- Gilani, M.; Danish, S.; Ahmed, N.; Rahi, A.A.; Akrem, A.; Younis, U.; Irshad, I.; Iqbal, R.K. Mitigation of drought stress in spinach using individual and combined applications of salicylic acid and potassium. Pak. J. Bot. 2020, 52, 1505–1513. [Google Scholar] [CrossRef]
- Gujjar, R.S.; Banyen, P.; Chuekong, W.; Worakan, P.; Roytrakul, S.; Supaibulwatana, K. A Synthetic Cytokinin Improves Photosynthesis in Rice under Drought Stress by Modulating the Abundance of Proteins Related to Stomatal Conductance, Chlorophyll Contents, and Rubisco Activity. Plants 2020, 9, 1106. [Google Scholar] [CrossRef]
- Zhu, Z.C.; Wu, Y.F.; Ma, J.C.; Yi, L.; Liu, B.H.; Jin, H.L. Chlorophyll changes and canopy spectral response of winter wheat under water stress based on unmanned airborne remote sensing. Spectrosc. Spectr. Anal. 2023, 43, 3524–3534. [Google Scholar]
- Labudová, L.; Labuda, M.; Takáč, J. Comparison of SPI and SPEI applicability for drought impact assessment on crop production in the Danubian Lowland and the East Slovakian Lowland. Theor. Appl. Clim. 2016, 128, 491–506. [Google Scholar] [CrossRef]
- Touzy, G.; Lafarge, S.; Redondo, E.; Lievin, V.; Decoopman, X.; Le Gouis, J.; Praud, S. Identification of QTLs affecting post-anthesis heat stress responses in European bread wheat. Theor. Appl. Genet. 2022, 135, 947–964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, Y.; Li, M.; Ma, J.; Wang, C.; Su, J.; Yang, D. Abscisic acid associated with key enzymes and genes involving in dynamic flux of water soluble carbohydrates in wheat peduncle under terminal drought stress. Plant Physiol. Biochem. 2020, 151, 719–728. [Google Scholar] [CrossRef]
- Zahra, N.; Hafeez, M.B.; Kausar, A.; Al Zeidi, M.; Asekova, S.; Siddique, K.H.M.; Farooq, M. Plant photosynthetic responses under drought stress: Effects and management. J. Agron. Crop. Sci. 2023, 209, 651–672. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of Water Stress on Photosynthesis, Yield, and Water Use Efficiency in Winter Wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon 2021, 7, e05908. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.W.; Zhang, Y.Z.; Shi, Y. Effects of a new compound water-holding agent on the growth and physiological characteristics of wheat seedlings under drought stress. Soils Fertil. Sci. China 2021, 255–261. [Google Scholar]
- Molero, G.; Reynolds, M.P. Spike photosynthesis measured at high throughput indicates genetic variation independent of flag leaf photosynthesis. Field Crop. Res. 2020, 255, 107866. [Google Scholar] [CrossRef]
- Silva-Pérez, V.; De Faveri, J.; Molero, G.; Deery, D.M.; Condon, A.G.; Reynolds, M.P.; Evans, J.R.; Furbank, R.T. Genetic variation for photosynthetic capacity and efficiency in spring wheat. J. Exp. Bot. 2019, 71, 2299–2311. [Google Scholar] [CrossRef]
- Luo, D.D.; Wang, C.K.; Jin, Y. Plant water regulation responses: iso-hydric and non-iso-hydric behavior. Chin. J. Plant Ecol. 2017, 41, 1020–1032. [Google Scholar]
- Wang, H.Z.; Xv, Y.L.; Dai, Y.Q.; Han, L. Photosynthetic physiological properties of heteromorphic leaves of Populus tremula and their relationship with leaf morphology. J. Tarim Univ. 2020, 32, 35–43. [Google Scholar]
- Wang, Y.; Wang, L.N.; Zhang, J.P.; Li, J.; Liu, Q.; Wang, S.P.; Gen, Z.K.; Zhou, J.; ZHANG, Y.Q. Response mechanism of photosynthesis to drought stress in Ginseng leaves. J. Shandong Univ. Tradit. Chin. Med. 2022, 46, 252–259. [Google Scholar] [CrossRef]
- Zhou, X.L.; Wen, L.L.; Jiang, D.C.; Xiao, C.P. Photosynthetic characteristics and physiological responses to drought stress in Northern Cangzhu (Aesculus nigra). J. Chin. Med. Mater. 2020, 43, 15–19. [Google Scholar] [CrossRef]
- Zhao, F.-N.; Zhou, S.-X.; Wang, R.-Y.; Zhang, K.; Wang, H.-L.; Yu, Q. Quantifying key model parameters for wheat leaf gas exchange under different environmental conditions. J. Integr. Agric. 2020, 19, 2188–2205. [Google Scholar] [CrossRef]
- Li, Q.; Gao, Y.; Hamani, A.K.M.; Fu, Y.; Liu, J.; Wang, H.; Wang, X. Effects of Warming and Drought Stress on the Coupling of Photosynthesis and Transpiration in Winter Wheat (Triticum aestivum L.). Appl. Sci. 2023, 13, 2759. [Google Scholar] [CrossRef]
- Sciarresi, C.; Patrignani, A.; Soltani, A.; Sinclair, T.; Lollato, R.P. Plant Traits to Increase Winter Wheat Yield in Semiarid and Subhumid Environments. Agron. J. 2019, 111, 1728–1740. [Google Scholar] [CrossRef]
- He, L.; Li, H.; He, J.; Han, D.; Li, R.; Wang, H. Irrigation at appearance of top 2nd or flag leaf could improve canopy photosynthesis by regulating light distribution and LAI at each leaf layer. Field Crop. Res. 2023, 295, 10. [Google Scholar] [CrossRef]
- Ferroni, L.; Živčak, M.; Kovar, M.; Colpo, A.; Pancaldi, S.; Allakhverdiev, S.I.; Brestič, M. Fast chlorophyll a fluorescence induction (OJIP) phenotyping of chlorophyll-deficient wheat suggests that an enlarged acceptor pool size of Photosystem I helps compensate for a deregulated photosynthetic electron flow. J. Photochem. Photobiol. B: Biol. 2022, 234, 112549. [Google Scholar] [CrossRef] [PubMed]
- Rane, J.; Sharma, D.; Ekatpure, S.; Aher, L.; Kumar, M.; Prasad, S.V.; Nankar, A.N.; Singh, N.P. Relative tolerance of photosystem II in spike, leaf, and stem of breadand durum wheat under desiccation. Photosynthetica 2019, 57, 1100–1108. [Google Scholar] [CrossRef]
- Zhu, L.; Wen, W.; Thorpe, M.R.; Hocart, C.H.; Song, X. Combining Heat Stress with Pre-Existing Drought Exacerbated the Effects on Chlorophyll Fluorescence Rise Kinetics in Four Contrasting Plant Species. Int. J. Mol. Sci. 2021, 22, 10682. [Google Scholar] [CrossRef] [PubMed]
- Spanic, V.; Mlinaric, S.; Zdunic, Z.; Katanic, Z. Field Study of the Effects of Two Different Environmental Conditions on Wheat Productivity and Chlorophyll Fluorescence Induction (OJIP) Parameters. Agriculture 2021, 11, 1154. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, B.; Zhao, F. Quantifying Agricultural Drought Severity for Spring Wheat Based on Response of Leaf Photosynthetic Features to Progressive Soil Drying. Atmosphere 2022, 13, 531. [Google Scholar] [CrossRef]
- Sales, C.R.G.; Molero, G.; Evans, J.R.; Taylor, S.H.; Joynson, R.; Furbank, R.T.; Hall, A.; Carmo-Silva, E. Phenotypic variation in photosynthetic traits in wheat grown under field versus glasshouse conditions. J. Exp. Bot. 2022, 73, 3221–3237. [Google Scholar] [CrossRef] [PubMed]
- Nehe, A.S.; Misra, S.; Murchie, E.H.; Chinnathambi, K.; Tyagi, B.S.; Foulkes, M.J. Nitrogen partitioning and remobilization in relation to leaf senescence, grain yield and protein concentration in Indian wheat cultivars. Field Crop. Res. 2020, 251, 107778. [Google Scholar] [CrossRef]
- Botyanszka, L.; Zivcak, M.; Chovancek, E.; Sytar, O.; Barek, V.; Hauptvogel, P.; Halabuk, A.; Brestic, M. Chlorophyll Fluorescence Kinetics May Be Useful to Identify Early Drought and Irrigation Effects on Photosynthetic Apparatus in Field-Grown Wheat. Agronomy 2020, 10, 1275. [Google Scholar] [CrossRef]
- Tan, C.-W.; Wang, D.-L.; Zhou, J.; Du, Y.; Luo, M.; Zhang, Y.-J.; Guo, W.-S. Assessment of Fv/Fm absorbed by wheat canopies employing in-situ hyperspectral vegetation indexes. Sci. Rep. 2018, 8, 9525. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Li, Y.T.; Hu, F.F.; Li, Y.; Wi, H.L.; Kang, J.H. Effect of post-flowering drought on fluorescence kinetic parameters in spring wheat. Jiangsu Agric. Sci. 2016, 44, 139–142. [Google Scholar] [CrossRef]
- Jia, Q.; Liu, Z.; Guo, C.; Wang, Y.; Yang, J.; Yu, Q.; Wang, J.; Zheng, F.; Lu, X. Relationship between Photosynthetic CO2 Assimilation and Chlorophyll Fluorescence for Winter Wheat under Water Stress. Plants 2023, 12, 3365. [Google Scholar] [CrossRef]
- Liu, G.C.; Yue, L.H.; Liu, C.H.; Zhao, P.Z.; Deng, Z.Y.; Li, Y.L. Effects of drought stress on photosynthetic fluorescence parameters of Russian sea buckthorn seedlings with large fruits. Northern Horticulture 2022, 16–23. [Google Scholar]
- Posch, B.C.; Hammer, J.; Atkin, O.K.; Bramley, H.; Ruan, Y.-L.; Trethowan, R.; Coast, O. Wheat photosystem II heat tolerance responds dynamically to short- and long-term warming. J. Exp. Bot. 2022, 73, 3268–3282. [Google Scholar] [CrossRef] [PubMed]
- ebela, D.; Bergkamp, B.; Somayanda, I.M.; Fritz, A.K.; Jagadish, S.V.K. Impact of post-flowering heat stress on spike and flag leaf senescence in wheat tracked through optical signals. Agron. J. 2020, 112, 3993–4006. [Google Scholar]
- Ren, H.L.; Zhu, X.; Zhang, F.Y.; Zhang, R.P.; Hong, H.L.; Zhnag, B.X.; Qui, L.J. Localization and exploitation of drought resistance genes in crops. Molecular Plant Breeding, 1–10.
- Li, L.; Wang, J.; Li, C.; Mao, X.; Zhang, X.; Sun, J.; Zhang, K.; Liu, Y.; Reynolds, M.P.; Yang, Z.; et al. Insights into progress of wheat breeding in arid and infertile areas of China in the last 14 years. Field Crop. Res. 2023, 306. [Google Scholar] [CrossRef]
- Shokat, S.; Großkinsky, D.K.; Singh, S.; Liu, F. The role of genetic diversity and pre-breeding traits to improve drought and heat tolerance of bread wheat at the reproductive stage. Food Energy Secur. 2023, 12. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: emergence of a biochemical role of a family of plant dehydration proteins. Physiologia Plantarum 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Trouverie, J.; Thévenot, C.; Rocher, J.; Sotta, B.; Prioul, J. The role of abscisic acid in the response of a specific vacuolar invertase to water stress in the adult maize leaf. J. Exp. Bot. 2003, 54, 2177–2186. [Google Scholar] [CrossRef]
- JAnderson, V.; Davis, D.G. Abiotic stress alters transcript profiles and activity of glutathione S-transferase, glutathione peroxidase, and glutathione reductase in Euphorbia esula. Plant Physiology 2004, 120, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Pnueli, L.; Hallak-Herr, E.; Rozenberg, M.; Cohen, M.; Goloubinoff, P.; Kaplan, A.; Mittler, R. Molecular and biochemical mechanisms associated with dormancy and drought tolerance in the desert legume Retama raetam. Plant J. 2002, 31, 319–330. [Google Scholar] [CrossRef]
- Lv, L.; Zhang, W.; Sun, L.; Zhao, A.; Zhang, Y.; Wang, L.; Liu, Y.; Li, Z.; Li, H.; Chen, X. Gene co-expression network analysis to identify critical modules and candidate genes of drought-resistance in wheat. PLOS ONE 2020, 15, e0236186. [Google Scholar] [CrossRef] [PubMed]
- de Ronde, J.A.; Cress, W.A.; Krüger, G.H.J.; Strasser, R.J.; van Staden, J. Photosynthetic response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat and drought stress. J. Plant Physiol. 2004, 161, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Morishita, H.; Urano, K.; Shiozaki, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Yoshiba, Y. Effects of free proline accumulation in petunias under drought stress. J. Exp. Bot. 2005, 56, 1975–1981. [Google Scholar] [CrossRef]
- Gubiš, J.; Vaňková, R.; Červená, V.; Dragúňová, M.; Hudcovicová, M.; Lichtnerová, H.; Dokupil, T.; Jureková, Z. Transformed tobacco plants with increased tolerance to drought. South Afr. J. Bot. 2007, 73, 505–511. [Google Scholar] [CrossRef]
- Suorsa, M.; Sirpiö, S.; Allahverdiyeva, Y.; Paakkarinen, V.; Mamedov, F.; Styring, S.; Aro, E.-M. PsbR, a Missing Link in the Assembly of the Oxygen-evolving Complex of Plant Photosystem II. J. Biol. Chem. 2006, 281, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Esquível, M.G.; Pinto, T.S.; Marín-Navarro, J.; Moreno, J. Substitution of tyrosine residues at the aromatic cluster around the βA-βB loop of rubisco small subunit affects the structural stability of the enzyme and the in vivo degradation under stress conditions. Biochemistry 2006, 45, 5745–5753. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, F.; Houde, M.; Sarhan, F. Purification, Characterization and cDNA Cloning of the 200 kDa Protein Induced by Cold Acclimation in Wheat. Plant Cell Physiol. 1993, 34, 59–65. [Google Scholar] [CrossRef]
- Dure, L. The LEA proteins of higher plants. In Control of Plant Gene Expression; Verma, D.P.S., Ed.; CRC Press: Boca Raton, Fla, USA, 1983; pp. 325–335. [Google Scholar]
- Campbell, S.A.; Close, T.J. Dehydrins: genes, proteins, and associations with phenotypic traits. New Phytol. 1997, 137, 61–74. [Google Scholar] [CrossRef]
- Cheng, Z.; Targolli, J.; Huang, X.; Wu, R. Wheat LEA genes, PMA80 and PMA1959, enhance dehydration tolerance of transgenic rice (Oryza sativa L.). Mol. Breed. 2002, 10, 71–82. [Google Scholar] [CrossRef]
- Litts, J.C.; Colwell, G.W.; Chakerian, R.L.; Quatrano, R.S. The nucleotide sequence of a cDNA clone encoding the wheat Em protein. Nucleic Acids Res. 1987, 15, 3607–3618. [Google Scholar] [CrossRef]
- Marcotte, W.R.; Bayley, C.C.; Quatrano, R.S. Regulation of a wheat promoter by abscisic acid in rice protoplasts. Nature 1988, 335, 454–457. [Google Scholar] [CrossRef]
- Rock, C.D.; Quatrano, R.S. Lanthanide ions are agonists of transient gene expression in rice protoplasts and act in synergy with ABA to increase Em gene expression. Plant Cell Reports 1996, 15, 371–376. [Google Scholar] [CrossRef]
- Ali-Benali, M.A.; Alary, R.; Joudrier, P.; Gautier, M.-F. Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. et Biophys. Acta (BBA) - Gene Struct. Expr. 2005, 1730, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, H.; Zhang, G.; Zhu, H.; Zhang, L.; Zhang, Z.; Zhang, C.; Ma, Z. Expression and responses to dehydration and salinity stresses of V-PPase gene members in wheat. J. Genet. Genom. 2009, 36, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Kamei, A.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Molecular responses to drought, salinity and frost: common and different paths for plant protection. Curr. Opin. Biotechnol. 2003, 14, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [PubMed]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; et al. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in droughtand low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Durmaz, E.; Akpınar, B.A.; Budak, H. The drought response displayed by a DRE-binding protein from Triticum dicoccoides. Plant Physiol. Biochem. 2011, 49, 346–351. [Google Scholar] [CrossRef]
- Yan, M.; Xue, C.; Xiong, Y.; Meng, X.; Li, B.; Shen, R.; Lan, P. Proteomic dissection of the similar and different responses of wheat to drought, salinity and submergence during seed germination. J. Proteom. 2020, 220, 103756. [Google Scholar] [CrossRef]
- Zhu, J.; Dong, C.H.; Zhu, J.K. Interplay between coldresponsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Abe, H.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Carninci, P.; Hayashizaki, Y.; Shinozaki, K. Monitoring the Expression Pattern of 1300 Arabidopsis Genes under Drought and Cold Stresses by Using a Full-Length cDNA Microarray. Plant Cell 2001, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Langridge, P.; Reynolds, M.P. Genomic tools to assist breeding for drought tolerance. Curr. Opin. Biotechnol. 2015, 32, 130–135. [Google Scholar] [CrossRef]
- Bapela, T.; Shimelis, H.; Tsilo, T.J.; Mathew, I. Genetic Improvement of Wheat for Drought Tolerance: Progress, Challenges and Opportunities. Plants 2022, 11, 1331. [Google Scholar] [CrossRef]
- Braun, H.J.; Rajaram, S.; Ginkel, M. CIMMYT’s approach to breeding for wide adaptation. In Adaptation in Plant Breeding; Springer: Dordrecht, The Netherlands, 1997; pp. 197–205. [Google Scholar]
- Tadesse, W.; Sanchez-Garcia, M.; Assefa, S.G.; Amri, A.; Bishaw, Z.; Ogbonnaya, F.C.; Baum, M. Genetic gains in wheat breeding and its role in feeding the world. Crop Breed. Genet. Genome 2019, 1, 1–28. [Google Scholar]
- Bonnin, I.; Bonneuil, C.; Goffaux, R.; Montalent, P.; Goldringer, I. Explaining the decrease in the genetic diversity of wheat in France over the 20th century. Agric. Ecosyst. Environ. 2014, 195, 183–192. [Google Scholar] [CrossRef]
- Van de Wouw, M.; Kik, C.; van Hintum, T.; van Treuren, R.; Visser, B. Genetic erosion in crops: Concept, research results and challenges. Plant Genet. Resour. 2010, 8, 1–15. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.A.; Crossa, J.; Huerta-Espino, J.; Vargas, M.; Mondal, S.; Velu, G.; Payne, T.S.; Braun, H.; Singh, R.P. Genetic Gains for Grain Yield in CIMMYT's Semi-Arid Wheat Yield Trials Grown in Suboptimal Environments. Crop. Sci. 2018, 58, 1890–1898. [Google Scholar] [CrossRef]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Ru, C.; Hu, X.; Chen, D.; Wang, W.; Zhen, J. Photosynthetic, antioxidant activities, and osmoregulatory responses in winter wheat differ during the stress and recovery periods under heat, drought, and combined stress. Plant Sci. 2022, 327, 111557. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Gupta, A.K.; Bains, N.S.; Kaur, K. Variability in enzymatic and non enzymatic antioxidants in wheat (Triticum aestivum L.) genotypes. Plant Physiol. Rep. 2021, 26, 1–15. [Google Scholar] [CrossRef]
- Michaletti, A.; Naghavi, M.R.; Toorchi, M.; Zolla, L.; Rinalducci, S. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Zhao, W.; Liao, A.M.; Ji, X.G.; Lv, X.; Bian, K.; Lin, J.T.; Zhang, X.S.; Hou, Y.C.; Lv, T. Progress in the study of heat shock proteins in wheat. J. Henan Univ. Technol. (Nat. Sci. Ed.) 2018, 39, 119–127. [Google Scholar] [CrossRef]
- Ma, D.F.; Hu, Y.F.; Niu, H.L.; Zeng, F.S. Genome-wide identification of the wheat water channel protein gene family and its role in salt and drought stress response. J. Yangtze Univ. (Nat. Sci. Ed.) 2021, 18, 84–103. [Google Scholar] [CrossRef]
- Wang, C.B.; Yi, Y.R.; Cheng, Z.; Ren, Y.K.; Niu, Y.Q.; Liu, J.; Han, B.; Yang, S.; Tang, Z.H. Identification and expression pattern analysis of wheat B-ARR transcription factor family genes. J. Plant Genet. Resour. 1–16. [CrossRef]
- Du, P.P.; Zhao, Y.J.; Shi, G.Q.; Xiao, K. Functional studies on the wheat NF-YB-type transcriptional gene TaNF-YB4 mediating plant resistance to drought. J. Hebei Agric. Univ. 2021, 44, 1–8. [Google Scholar] [CrossRef]
- Li, X.; Wei, W.; Li, F.; Zhang, L.; Deng, X.; Liu, Y.; Yang, S. The Plastidial Glyceraldehyde-3-Phosphate Dehydrogenase Is Critical for Abiotic Stress Response in Wheat. Int. J. Mol. Sci. 2019, 20, 1104. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Xue, C.; Xiong, Y.; Meng, X.; Li, B.; Shen, R.; Lan, P. Proteomic dissection of the similar and different responses of wheat to drought, salinity and submergence during seed germination. J. Proteom. 2020, 220, 103756. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Keyword co-occurrence network mapping for the study of spring wheat fertility response to drought stress.
Figure 1.
Keyword co-occurrence network mapping for the study of spring wheat fertility response to drought stress.
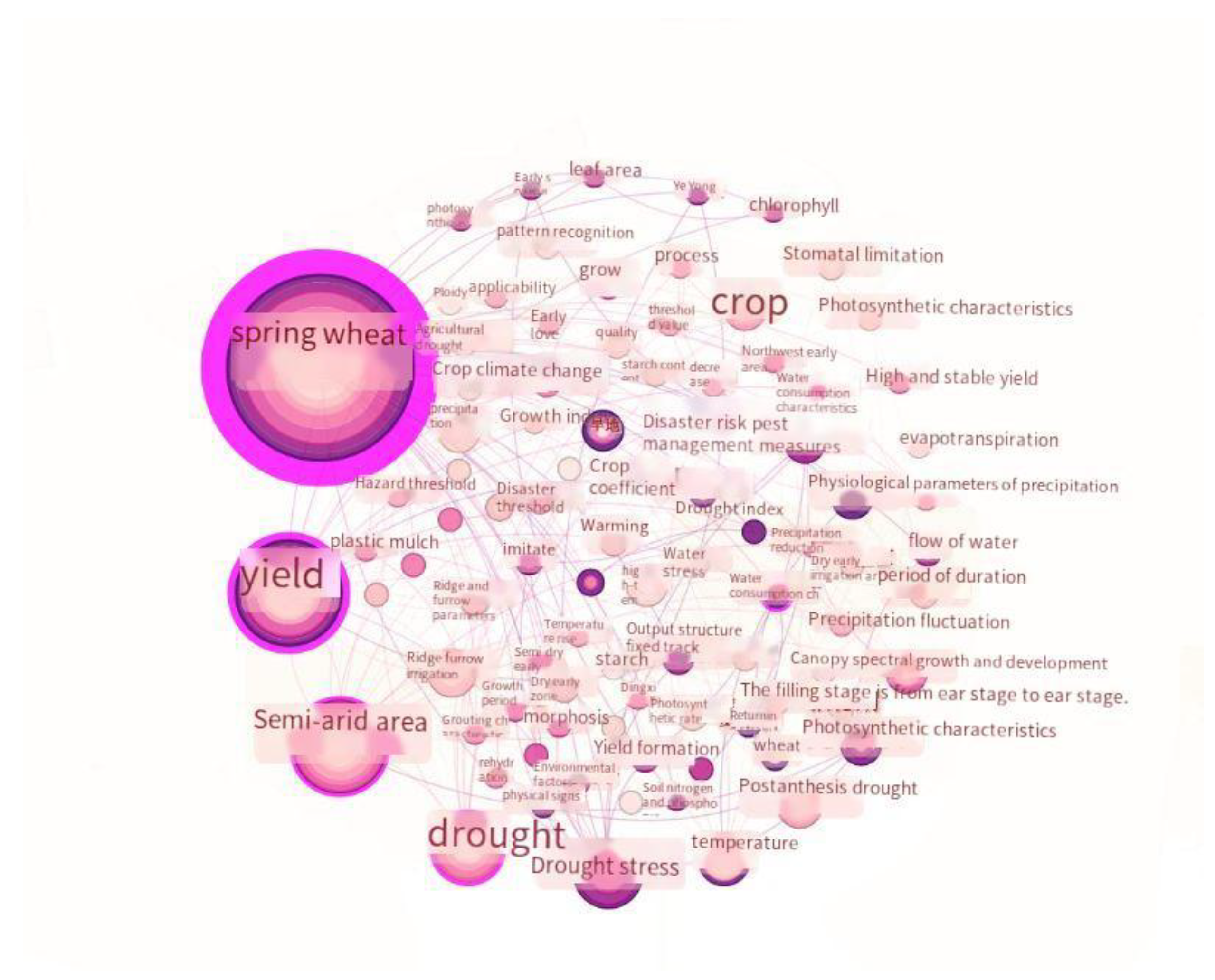
Figure 2.
Keyword covariance network mapping for the study of leaf water potential in response to drought stress in spring wheat.
Figure 2.
Keyword covariance network mapping for the study of leaf water potential in response to drought stress in spring wheat.

Figure 3.
Keyword covariance network mapping for the study of chlorophyll response to drought slopes in spring wheat.
Figure 3.
Keyword covariance network mapping for the study of chlorophyll response to drought slopes in spring wheat.
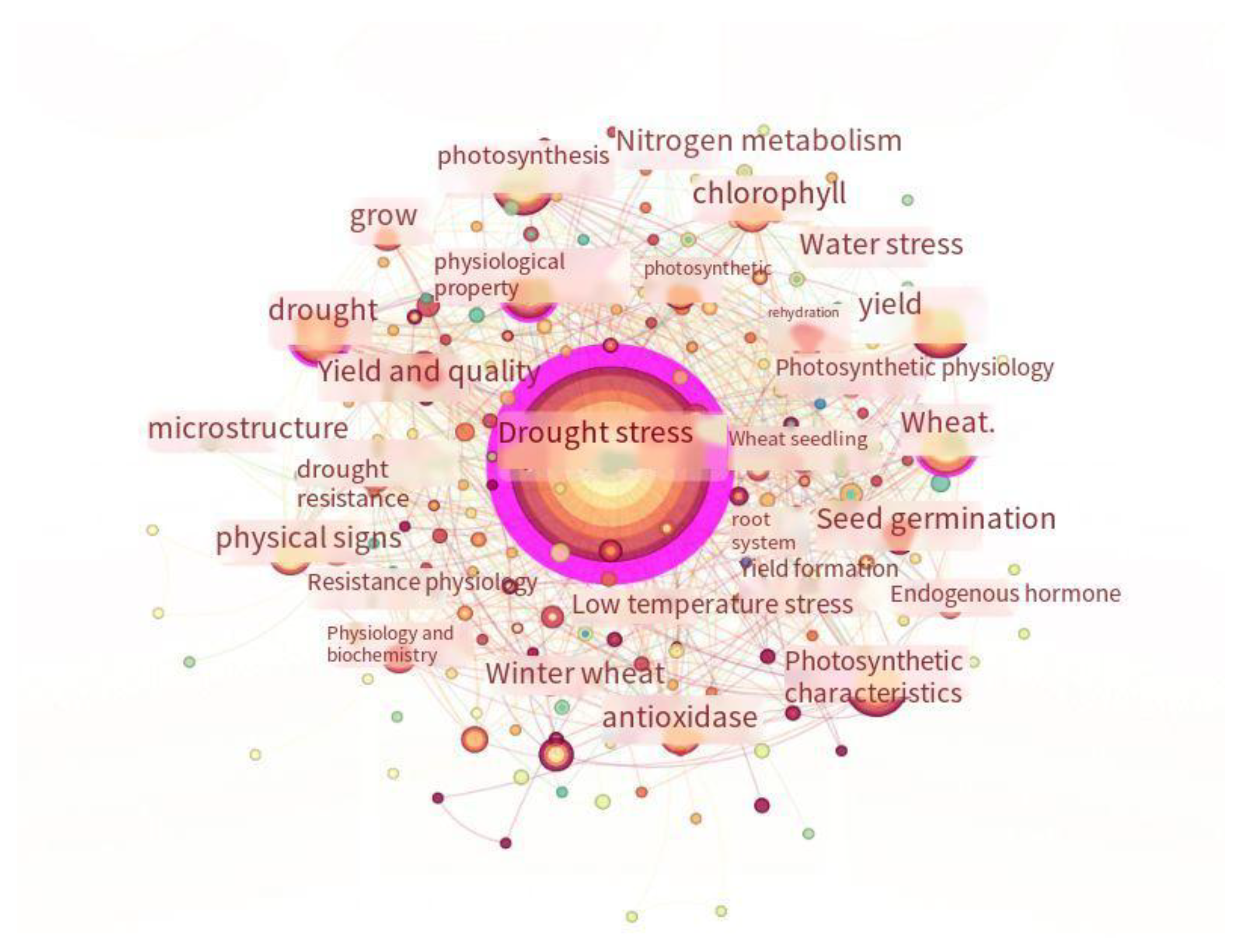
Figure 4.
Research keyword covariance network mapping of the effect of drought stress on photosynthesis in spring wheat leaves.
Figure 4.
Research keyword covariance network mapping of the effect of drought stress on photosynthesis in spring wheat leaves.
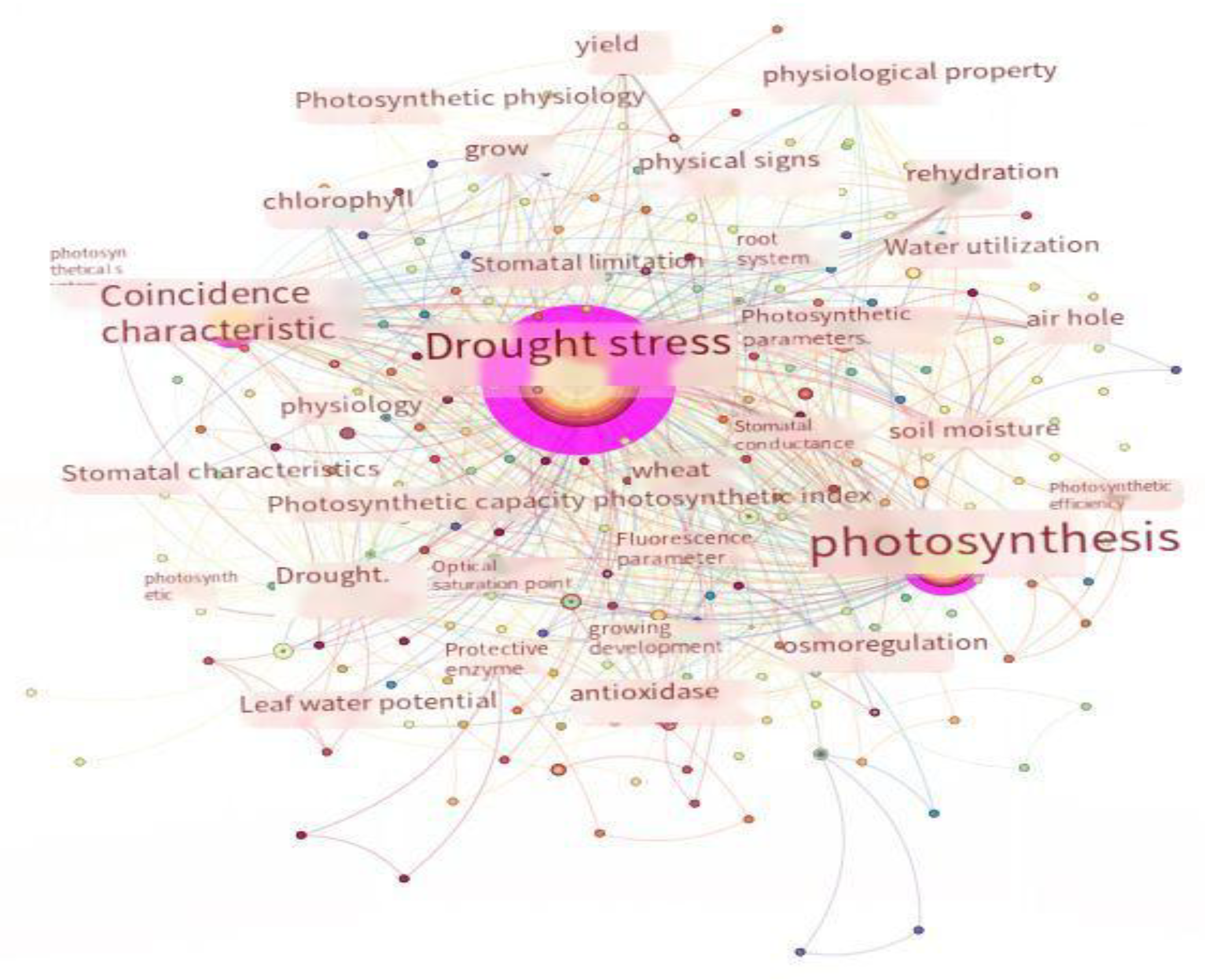
Figure 5.
Study of changes in chlorophyll fluorescence parameters of leaves under drought stress Keywords covariance network mapping.
Figure 5.
Study of changes in chlorophyll fluorescence parameters of leaves under drought stress Keywords covariance network mapping.
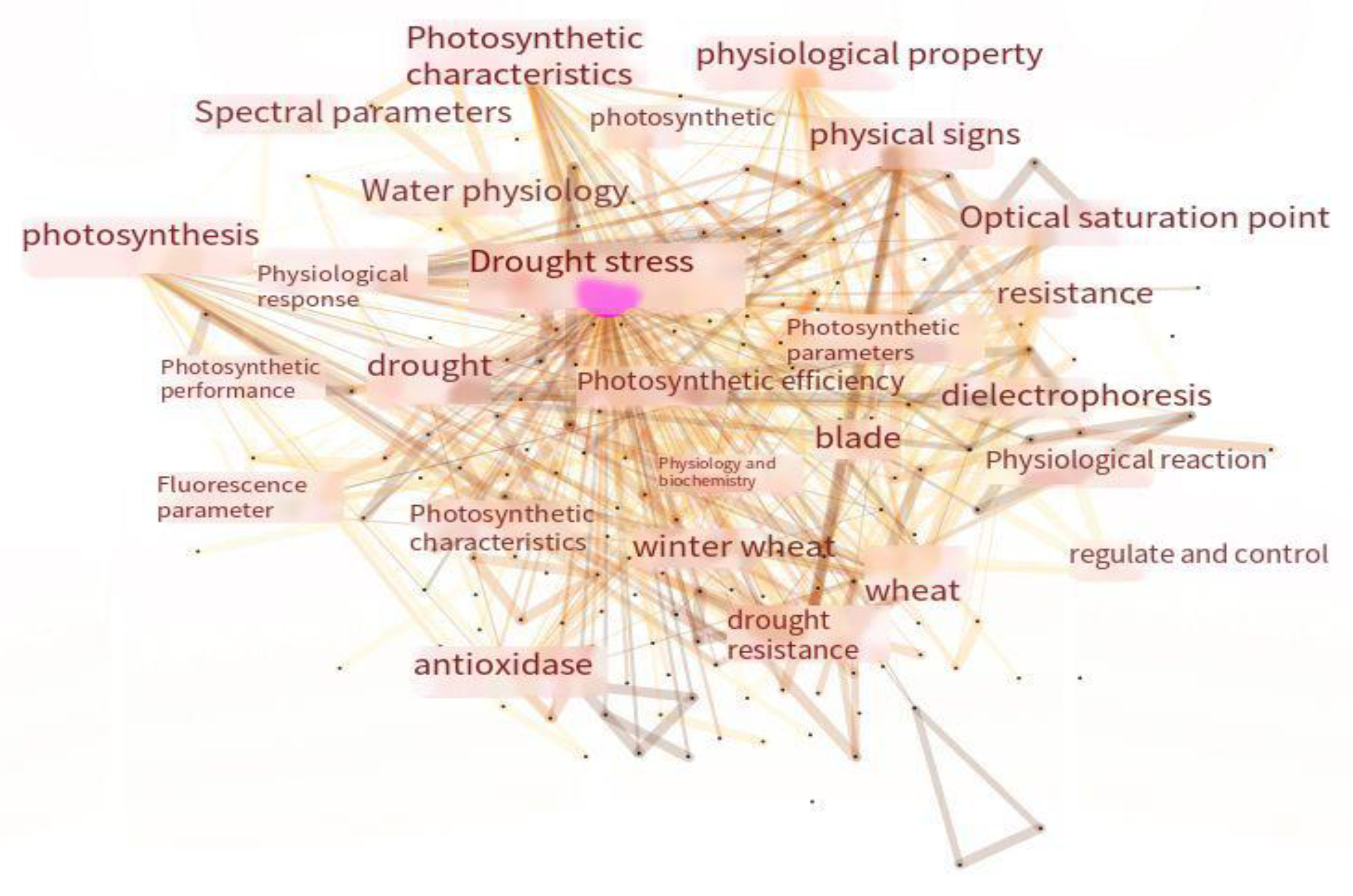
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



