Submitted:
29 January 2025
Posted:
31 January 2025
You are already at the latest version
Abstract
Gonadotropin-inhibitory hormone (GnIH) has a well-established role in regulating reproductive functions, but recent data has also demonstrated its involvement of in the control of metabolic processes. GnIH neurons and fibers have been identified in hypothalamic regions associated with feeding behavior and energy homeostasis with GnIH receptors being expressed throughout the hypothalamus. GnIH does not act alone in the hypothalamus, but rather interacts with the melanotropin system as well as with other neuropeptides. GnIH and its receptors are also expressed in peripheral tissues involved in important metabolic functions. Therefore, the local action of GnIH in peripheral organs, including the pancreas, gastrointestinal tract, gonad, and adipose tissue, is also suggested. This review aims to provide a comprehensive summary of the emerging role of GnIH in metabolic regulation at both central and peripheral levels.
Keywords:
gonadotropin-inhibitory hormone (GnIH)
; RFamide-Related Peptide-3 (RFRP-3)
; metabolism
; food intake
; hypothalamus
; adipose tissue
1. Introduction
Since gonadotropin-inhibitory hormone (GnIH) was first discovered in 2000 as the hypothalamic neuropeptide that actively inhibits gonadotropin release, much evidence now supports GnIH as having a role as a key neurohormone to inhibit reproduction by regulating the hypothalamic-pituitary function. Given that a complex reciprocal mechanism exits between the reproductive axis and other endocrine systems, accumulating evidence has also suggested the involvement of GnIH in the adrenal and thyroid axes. Moreover, recent evidence suggests that GnIH also influences feeding behavior and energy homeostasis. GnIH-mediated metabolic control may occur at multiple levels. Although most interactions between metabolic factors and GnIH occur in the hypothalamus, the direct influence of GnIH on peripheral metabolic tissues has recently been recognized. In this review, we aim to highlight the endocrine role of central and peripheral GnIH in the regulation of metabolic process.
2. Overview of GnIH and Its Impact on the Endocrine System
2.1. GnIH and Its Receptors
GnIH was initially isolated from the Japanese quail (Coturnix japonica) hypothalamus and it was shown that GnIH inhibited gonadotropin release from the Japanese quail cultured anterior pituitary gland [1]. The GnIH precursor gene encodes one GnIH and two GnIH-related peptides, GnIH-RP-1 and GnIH-RP-2, in all avian species studied [2,3,4,5,6]. Subsequently, GnIH peptides have been identified in all classes of vertebrates that share an LPXRFamide (X = L or Q) motif at their C-termini [7,8,9], thus also known as RFamide-related peptides (RFRPs). Mammalian GnIH precursor gene is translated and cleaved into at least two GnIH peptides, RFRP-1 (also known as neuropeptide SF, NPSF) and RFRP-3 (also known as neuropeptide VF, NPVF) [7,8,9]; RFRP-2 is not an LPXRFamide peptide.
Two G protein-coupled receptors, GPR147 (also known as neuropeptide-FF receptor 1, NPFFR1; NPFF1; NPFF1R1; OT7T022) and GPR74 (also known as NPFFR2; NPFF2; NPGPR; HLWAR77) have been identified as GnIH receptors (GnIH-Rs) [3,10,11,12]. Given the higher GnIH binding affinity for GPR147 than GPR74, it is postulated that GPR147 is the principal receptor for GnIH [3,11]. Both GnIH-Rs couple to Gαi, which inhibits the activity of adenylate cyclase (AC), thus reducing intracellular cAMP levels and protein kinase A (PKA) activity [10,13,14,15]. The molecular mechanism of GnIH-mediated cAMP inhibition was investigated using cellular model systems derived from the hypothalamus, anterior pituitary gland, and gonadal tissue [16,17,18]. The results indicate that inhibition by GnIH is through specific targeting of the AC/cAMP/PKA pathway.
2.2. Regulation of the Endocrine System by GnIH
GnIH is localized in various regions of the brain in many species, particularly in the diencephalon and mesencephalon, with a specific focus on the hypothalamic region [19,20,21,22]. GnIH neuronal cell bodies are located in the paraventricular nucleus (PVN) in birds [1,23,24] and the dorsomedial hypothalamic area (DMH) in most mammals [10,20,25,26,27]. GnIH neuronal projections are also identified throughout the brain, including the preoptic area (POA), lateral septum, arcuate nucleus (ARC), and anterior hypothalamus in mammals [22,25,28,29]. Similarly, GnIH-Rs are also expressed throughout the hypothalamus, specifically in the POA, rostral periventricular area of the third ventricle, and ARC. The widespread localization of GnIH neurons and GnIH-Rs within the hypothalamic region strongly suggests multiple neuroendocrine functions of the GnIH system.
2.2.1. Role of GnIH on the Reproductive Axis
The control of reproduction depends on the intact hypothalamic-pituitary-gonadal (HPG) axis. Reproductive neuropeptides synthesized in hypothalamic neurons, including the well-known gonadotropin-releasing hormone (GnRH), kisspeptin, and GnIH, regulate the release of pituitary gonadotropin, follicle stimulating hormone (FSH) and luteinizing hormone (LH) that, in turn, control gonadal functions and the production of sex hormones. Extensive studies have established that GnIH peptides reduce the synthesis and/or release of gonadotropin (reviewed in [7,8,30,31,32,33]). At the hypothalamic level, GnIH inhibits the activity of GnRH and/or kisspeptin neurons. Based on the morphological evidence, GnIH has been considered to act at the most upstream level of the HPG axis, as described below.
GnRH stimulates the release of gonadotropin and plays a fundamental role in controlling physiological aspects of reproductive process. The projection of GnIH neurons to GnRH neurons is the most conserved characteristic of GnIH neurons across species. GnIH neuronal axon terminals form both axo-somatic and axo-dendritic contacts with GnRH neurons, which express GnIH-Rs in the POA [20,25,34,35,36,37]. Direct application of GnIH to hypothalamic brain slices decreased the firing rate of a subpopulation of GnRH neurons [38]. Electrophysiological recordings also support this observation that a direct postsynaptic inhibition of GnRH neuronal firing may occur via GnIH [39]. Additionally, intracerebroventricular (ICV) administration of GnIH suppressed cFOS activity in GnRH neurons, indicating the direct suppressive effects of GnIH on the regulation of GnRH neurons.
Kisspeptin serves as a stimulatory regulator for GnRH release across various vertebrate species, excluding birds. In mammals, kisspeptin neurons are found in the anteroventral periventricular nucleus (AVPV) and ARC. AVPV kisspeptin neurons are regarded as a target of estrogen-positive feedback to induce preovulatory GnRH/LH surge, whereas ARC kisspeptin neurons are negatively regulated by sex steroids on GnRH/LH secretion. In mice, only a small portion (5-15%) of kisspeptin neurons in the AVPV expresses GPR147 or GPR74, whereas higher co-expression with either of the GnIH-Rs is observed in the ARC kisspeptin neurons. Approximately 35% of kisspeptin neurons in the ARC are contacted by GnIH neuronal fibers [40], suggesting that GnIH may directly modulate the kisspeptin neuronal activity associated with the generation of pulsatile GnRH/LH secretions.
GnIH neuronal fibers are also observed in the median eminence (ME) and control anterior pituitary function via GnIH-Rs expressed in the gonadotropes [1,19,23,37,41,42,43]. Although there is some debate regarding whether GnIH can act directly on the anterior pituitary gland in some species, evidence from numerous studies indicates that GnIH can decrease pituitary gonadotropin synthesis and/or release in many species [22,35,42,44,45,46,47].
While the inhibitory effect of GnIH on reproduction is mainly accomplished at the hypothalamic-pituitary levels, GnIH and GnIH-Rs are also expressed in steroidogenic cells and germ cells in the gonads, testis and ovary, indicating that autocrine or paracrine mechanisms of GnIH system exist to control gonadal functions by inhibiting sex steroid production and gametogenesis [16,48,49,50]. Therefore, GnIH is a key regulator that inhibits each level of the HPG axis by controlling the activities of hypothalamic GnRH and kisspeptin neurons, pituitary gonadotropin secretion, and gonadal function.
2.2.2. Involvement of GnIH in the Stress Axis
The stress system, known as the hypothalamic-pituitary-adrenal (HPA) axis is closely linked to the HPG axis. Prolonged exposure to stress can disrupt the equilibrium between the HPA and HPG axes, resulting in reduced reproductive function and disorders associated with infertility. Stress response is characterized by hypothalamic release of corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP), which in turn induce the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH causes the adrenal cortex to secrete glucocorticoids (GC) such as corticosterone in birds and rodents. When GC levels reach a certain concentration, GC exerts negative feedback via CRH, AVP, and ACTH to end the stress response, thereby returning to the systemic homeostasis. GnIH neurons in the PVN direct contact with CRH [51]. Furthermore, the CRH receptor-1 is expressed in GnIH neurons [52], and GnIH neurons express GC receptor (GR) [52,53], indicating that adrenal GC can mediate the stress effect through direct action on GnIH neurons.
Both the HPA and HPG axes exhibit striking sex differences, and there are controversial effects depending on the species and stress paradigm. Nonetheless, most experimental stress protocols, such as restraint/immobilization [52,54,55,56,57,58,59], nutritional/metabolic [60,61,62,63], and thermal stress [64,65], have been shown to activate GnIH and/or GnIH-Rs in the hypothalamus and/or gonad with showing negatively related gonadotropin or sex hormone levels (Reviewed in [66]). Importantly, the suppressive effects of stress on the HPA activity are prevented by ablation of GnIH neurons [58], knockdown of GnIH [54], knockout (KO) of GPR147 [60], or administration of GPR147 antagonist [62]. These findings suggest that GnIH plays a crucial role in mediating stress-induced reproductive dysfunction. In addition, pharmacologically induced stress status by administration of ACTH or corticosterone, increases the expression of GnIH or GnIH-Rs [53,67,68]. Importantly, GR is recruited to the GnIH promoter region in response to corticosterone [52,53], further supporting the direct activation of GnIH in response to stress.
GnIH also exerts a stimulatory effect on the HPA axis. ICV administration of GnIH significantly elevated serum corticosterone levels in conscious male rats [69]. Mice that received chronic infusions of GnIH exhibited anxiogenic effects and demonstrated elevated basal circulating corticosterone levels [70]. It is noteworthy that a highly selective GPR147 antagonist, GJ14, effectively impeded GnIH-induced corticosterone release and CRH neuronal activation, while also reversing the anxiogenic impact of GnIH [70]. Furthermore, acute stimulation of GnIH neurons resulted in a significant, dose-dependent release of corticosterone [58]. These results indicate the existence of a positive feedback loop whereby stressful stimuli activate the GnIH system, which in turn further activates the HPA axis. The expression of GPR147 in the PVN, where the CRH/AVP neurons are located, may provide support for the potential modulatory action of GnIH in this positive feedback loop.
2.2.3. Involvement of GnIH in the thyroid axis
Thyroid hormones (THs; thyroxine, T4 and triiodothyronine, T3), are well known as regulators of metabolism, development, and growth. They play an vital role in the normal development and function of the reproductive system, suggesting close interactions between the hypothalamic-pituitary-thyroid (HPT) and HPG axes. Therefore, thyroid disorders, such as hypothyroidism and hyperthyroidism, cause abnormal reproductive function. As elevated TH levels are known to activate GnRH neurons and indirectly suppress the activity of GnIH [71,72,73], T4-induced hyperthyroidism leads to a decrease in hypothalamic GnIH expression, and T3 treatment suppresses GnIH mRNA expression in hypothalamic explants [74]. In contrast, hypothyroidism induced by long-term administration of propylthiouracil (PTU), which inhibits the production of new TH, in juvenile female mice results in delayed pubertal onset with increased hypothalamic GnIH expression and decreased pituitary-gonadal activity, and the effect of hypothyroidism in delaying pubertal onset is prevented by GnIH KO [74]. Although to date limited information is available for the TH-mediated GnIH regulation, the expression of TH receptors in GnIH neurons [74] suggests a direct involvement of GnIH in mediating HPT-HPG interactions.
3. Metabolic Regulation by Hypothalamic GnIH
The hypothalamus is the primary target of metabolic programming and the principal regulatory center of energy metabolism, acting through both neuronal and hormonal mechanisms, and each hypothalamic area has a distinct role in the metabolic regulation. The wide distribution of GnIH fibers and GnIH-Rs throughout regions of the hypothalamus involved in appetite and energy homeostasis, including the ARC, PVN, lateral hypothalamus, and ventromedial nucleus, supports the significance of GnIH beyond the reproductive axis. Therefore, there have been attempts to determine whether GnIH administration would affect metabolism.
3.1. Effect of Central GnIH Administration on Feeding Behavior
It has been confirmed that central GnIH administration affects the appetite in many species. The stimulatory effect of GnIH on food intake was first demonstrated in domestic chicken chicks [75]. Tachibana et al. showed that ICV injection of GnIH stimulates food intake, whereas anti-GnIH antiserum injection significantly inhibits food intake in food deprivation-induced feeding [75]. Subsequent studies have demonstrated that GnIH exerts stimulatory effects on food intake in other bird species, and mammals including mice, hamsters, rats, sheep, and cynomolgus monkeys, as evidenced by an increase in food intake following ICV injection of GnIH [22,76,77,78,79,80,81]. However, other factors also play as role such as sex or duration of GnIH administration. For example, chronic injections of GnIH did not stimulate food intake in female hamsters [80] or following acute injection in male hamsters [81] or rats [82].
A single study has reported the inhibitory effect of GnIH (specifically RFRP-1) on feeding behavior through direct brain microinjection of GnIH into amygdala [83]. The amygdala plays an important role in the regulation of food intake and body weight. GnIH-immunoreactive fibers and GPR147 are found in the amygdala [84]. Kovács et al. showed that intra-amygdaloid microinjection of GnIH decreased liquid food intake in rats, and this effect was prevented by the pretreatment with the GPR147 antagonist RF9 in amygdala [83]. The metabolic effects of central GnIH administration described above are summarized in Table 1.
3.2. Interaction with the Melanocortin System
The hypothalamus achieves the energy homeostasis mainly through the melanocortin system, which is located in the ARC and DMH. The melanocortin system has direct connections to the PVN and other brain regions to regulate feeding behavior as well as energy homeostasis. The melanocortin system involves the orexigenic agouti-related peptide (AgRP)/neuropeptide Y (NPY) neurons, and the anorexigenic proopiomelanocortin (POMC)/cocaine-and-amphetamine regulated transcript (CART) neurons. GnIH fibers form close apposition with the NPY and POMC neurons in the ARC [43,51,85]. Although it is considered that GnIH has a stimulatory effect on feeding behavior, the direct role of GnIH on the melanocortin system has yielded conflicting results.
Consistent with the orexigenic function of GnIH, GnIH has been found to reduce the electrophysiological function of POMC neurons in hypothalamic slices from mice [85,86]. Similarly, hypothalamic POMC expression was shown to be increased in GPR147 KO male mice, which exhibited a decline in spontaneous food intake, further suggesting the orexigenic action of endogenous GnIH/GnIH-R signaling [87]. The orexigenic effect was also observed in male mice [79] and hamsters [80] after chronic GnIH ICV injection ; however, the expression of hypothalamic metabolic genes, POMC, NPY, or AgRP were not altered.
Despite its well-documented ability to stimulate food intake in various species, including mice, GnIH showed a predominantly inhibitory effect on the electrophysiological and functional activity of NPY neurons (80% of those tested) in mice [85]. Furthermore, GnIH also inhibited the secretion of NPY in incubated hypothalamic slices [85], but GnIH excited less than 20% of NPY neurons, and a similar percentage of NPY neurons received close input from GnIH-immunoreactive fibers. These findings suggest therefore, the control of NPY neurons in the mouse ARC appears to be complex.
3.3. Hypothlamic GnIH Regulation by Peripheral Hormones, Leptin and Ghrelin
The adipose-derived hormone leptin has strong effects on hypothalamic regulation of satiety, energy expenditure, and body weight. Leptin acts centrally through its specific receptor, LepR. Earlier studies have shown that leptin has little or no effect on GnIH neurons. Poling et al. showed that LepR mRNA is found to be co-expressed in ∼15% of GnIH neurons [88], but other research concluded that LepR mRNA is undetectable in semi-purified GnIH neuronal preparations [89]. Regardless of whether LepR is expressed in GnIH neurons or not, both groups indicated that leptin-deficient mice exhibit either a minor reduction in GnIH mRNA levels or no detectable difference compared to wild-type mice. Furthermore, the postnatal development of GnIH neurons appears to be unaffected by leptin deficiency [88]. Together, these results indicate that GnIH is unlikely to be an important neuronal pathway for the direct regulation of metabolism by leptin.
Ghrelin, which is secreted from the gastrointestinal (GI) tract, exerts orexigenic effects by acting on the hypothalamic ARC. Although ghrelin is primarily linked to the GI tract, some studies have reported ghrelin-immunoreactive cells in the hypothalamus [90,91] and the existence of ghrelin-producing neurons was reported in the ARC [92]. To date, the direct interaction between GnIH and ghrelin has not been extensively studied, and the potential impact of ghrelin on the activity of GnIH neurons is not well understood. However, given the widespread distribution of GnIH and GnIH-Rs in the brain, it is hypothesized that GnIH neurons may project to ghrelin neurons in the ARC and contribute to the regulation of its central effects (reviewed in [93]).
Despite the absence of direct evidence for their interactions in in hypothalamus, GPR147 KO male mice exhibited an altered food intake response induced by ICV administration of leptin and ghrelin [87]. Anorectic response to leptin were exaggerated in GPR147 KO mice compared to wild-type during low-fat diet conditions. Conversely, orexigenic responses to ghrelin were blunted in GPR147 KO mice during high-fat diet (HFD) conditions. Thus, this study suggests the possible involvement of the GnIH system in the feeding response by these hormones, in which the effects of leptin and ghrelin might partially depend on the integrity of GnIH/GPR147 signaling pathway.
4. Metabolic Regulation by Peripheral GnIH
Although GnIH is primarily produced in the hypothalamus, similar to other neuropeptides, it can potentially impact peripheral tissues expressing GnIH-Rs through endocrine signaling via the bloodstream, paracrine signaling for local regulation, or autocrine signaling for self-regulation. Recent evidence suggests a broader role for GnIH in peripheral tissues, as indicated by the expression of GnIH and/or GnIH-Rs in peripheral endocrine organs, not just the brain. Among these organs, the intra-gonadal role of GnIH has been well investigated in relation to the integration of reproduction and metabolism (reviewed in [94]). Thus, in this review we focus on the potential role of GnIH in metabolic tissues, such as adipose tissue, pancreas, GI tract, and liver, where expression of the GnIH system is confirmed (Table 2). As this is a relatively new field of GnIH research, in this section we discuss the latest progress in peripheral metabolic control by the GnIH system.
4.1. Effect of Peripheral GnIH Administration
The peripheral administration of GnIH has been demonstrated to induce orexigenic effects that resemble those observed following central ICV injection. Chronic intraperitoneal (IP) injection of GnIH has been shown to increase food intake in male mice [102,103], as well as in both male and female rats [96], and female piglets [104]. These chronic IP injection models have shown the hyperphagia-induced weight gain, adiposity, and impaired glucose homeostasis, suggesting the potential involvement of GnIH in obesity-induced metabolic disorders and related reproductive dysfunction. However, it remains unclear whether peripheral GnIH can enter the brain via its receptor-mediated transport crossing the blood-brain barrier. Alternatively, IP-injected GnIH may act on the peripheral organs expressing GnIH-Rs. We have summarized the effect of IP injection of GnIH on metabolism in Table 3, and the changes in metabolic tissues induced by GnIH injection are described in the following section.
4.2. Adipose Tissue
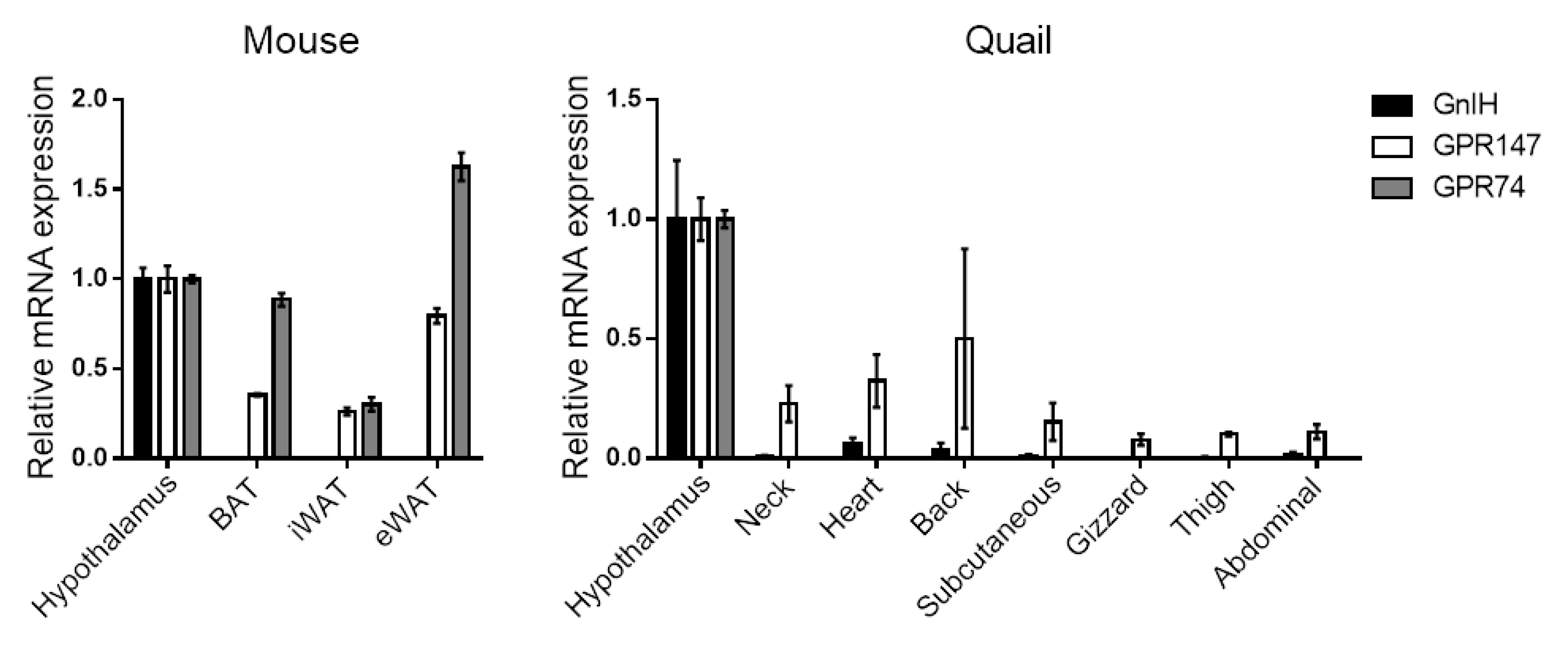
It appears that GnIH itself is not produced in adipose tissues, and we have also confirmed that GnIH mRNA is not detected in various adipose depots in both mice and Japanese quail (Figure 1). However, GnIH-Rs have been observed in human [95] and rat [96] adipose tissues, suggesting the direct effect of GnIH on adipose tissues via its receptors. Our expression profiling also indicates that GPR147 and GPR74 are abundantly expressed in mouse brown and white adipose tissues (BAT and WATs), with higher levels of GPR74 than GPR147 (Figure 1, left). In Japanese quail adipose tissues, GPR147 is predominantly expressed in all depots tested, compared to GPR74 (Figure 1, right). In human samples, both GPR147 and GPR74 were expressed at the protein level in freshly isolated mature adipocytes as well as in vitro differentiated adipocytes from the omental and subcutaneous region [95]. Particularly, GPR74 mRNA expression levels were higher in adipose tissue from obese as compared with non-obese subjects [95]. In mouse 3T3-L1 differentiated adipocytes, both GnIH-Rs were also expressed [97]. Given the characteristics of GnIH-Rs, which are coupled with Gαi to inhibit cAMP pathway, it can be hypothesized that higher gene expression of either receptors may be associated with lower lipolysis activity by inhibiting the noradrenaline/cAMP/PKA pathway [106]. Indeed, in vitro differentiated adipocytes isolated from human subjects, RFRP-1 was indicated to be important for the regulation of lipolysis probably acting through GPR74 and GPR147 [95]. Although the direct effect of RFRP-3 was not investigated in this study, its binding affinity to these receptors suggest that RFRP-3 may have a similar effect to RFRP-1 on lipolysis.
Chronic GnIH-treatment causes an increase in the mass of adipose tissue [79,96,102,103,104] in many species. Chronic ICV infusion of GnIH for 13 days increased BAT mass, resulting in WAT-like appearance. BAT is the thermoregulatory organ to enhance energy expenditure, thus the observed decreases in energy expenditure and core body temperature could be explained by the impaired BAT function in GnIH-injected mice [79]. Although, centrally injected GnIH did not change WAT mass, peripheral administration of GnIH significantly increased WAT mass and altered its function related to glucose and lipid metabolism. Anjum et al. showed that in vivo (IP injection for 8 days) and in vitro treatment of GnIH resulted in upregulation of glucose transporter 4 (GLUT4), a major insulin-stimulated glucose transporter, and increased triglyceride uptake in abdominal visceral WAT, contributing to obesity-related metabolic abnormalities in male mice [102]. Similarly, chronic IP injection of GnIH for 14 days in male mice led to hyperlipidemia, hyperglycemia, glucose intolerance, and insulin resistance through changes in the expression of glucose and lipid metabolism-related genes in perigonadal visceral WAT [103]. A similar set of effects was observed in rats [96] and female piglets [104] after IP administration of GnIH, including an increase in WAT mass and impaired glucose transport and insulin signaling in WAT. These effects were associated with a disruption in whole-body glucose homeostasis. The molecular mechanism of GnIH-induced impaired glucose metabolism via inhibition of AKT-GSK3-β signaling is suggested [96]; however, it remains unclear whether this effect is directly mediated by GnIH-Rs in WAT. Further studies, using GnIH-Rs antagonists or the overexpression/knockdown of GnIH-Rs are required to further elucidate the direct involvement of GnIH via its receptors.
Figure 1.
Expression of GnIH and GnIH-Rs in various adipose tissues isolated from mouse (left) and Japanese quail (right). Mouse adipose tissues include interscapular BAT, subcutaneous iWAT, and visceral eWAT. Japanese quail adipose tissues are collected close to the indicated depot and organ. RT-PCR was performed according to our previous report [107]. Expression levels are shown by setting “Hypothalamus” to 1.
Figure 1.
Expression of GnIH and GnIH-Rs in various adipose tissues isolated from mouse (left) and Japanese quail (right). Mouse adipose tissues include interscapular BAT, subcutaneous iWAT, and visceral eWAT. Japanese quail adipose tissues are collected close to the indicated depot and organ. RT-PCR was performed according to our previous report [107]. Expression levels are shown by setting “Hypothalamus” to 1.
4.3. Pancreas
Given the effect of GnIH on food intake and glucose homeostasis, it has been hypothesized that GnIH/GnIH-Rs may play a role in the pancreas, a vital organ in regulating blood glucose levels through the secretion of insulin and glucagon. In addition, other RFRP members, 43RFa and 26RFa, have been shown to promote the survival of INS-1E rat pancreatic β-cells and human pancreatic islets [108], suggesting the possible involvement of GnIH in the pancreatic regulation. Indeed, abundant GnIH-immunoreactive cells were found to be concentrated in the pancreatic islets of rats [96] and male piglets [98] . In addition, GPR147 expression was also confirmed in the islets of rats [96] and mice [99]. Specifically, intense GnIH-immunoreactive cells were observed in some α-cells, but not in β-cells, indicating that GnIH colocalized primarily with glucagon [96]. In contrast, moderate GPR147 immunoreactivity was observed around the islet and exocrine β-cells of the pancreas and weak immunoreactivity with α-cells, indicating that GPR147 colocalized primarily with insulin in the pancreatic islets [96].
Using αTC1, a mouse islet α-cell line which was also validated to express GPR147, it was reported that GnIH promoted survival of α-cells under conditions of hyperglycemia and serum starvation [99]. Mechanistically, GnIH activated PI3K/AKT and ERK1/2 signaling cascades, and treatment with RF9 (GPR147 antagonist) blocked the activation of both pathways, thereby indicating the possible role of GnIH in promoting α-cell survival, probably via GPR147. In vivo studies have shown that IP-injected GnIH induces changes in islet histomorphology, thereby altering insulin and glucagon secretion from pancreas, which contributes to hyperglycemia and insulin resistance in GnIH-treated models. In rat models, chronic GnIH treatment for 14 days resulted in a dramatic induction of pancreatic islet hyperplasia and significantly inhibited insulin secretion, accompanied by elevated glucagon secretion [96]. Similarly, female piglets treated with GnIH for 14 days showed increased pancreatic mass and islet hypertrophy, along with hyperinsulinism and hyperglucagon [104]. Taken together, these in vitro and in vivo findings suggest a direct action of GnIH on pancreatic islets, potentially via GPR147, to regulate glucose homeostasis.
4.4. GI Tract
Despite the limitations of a few studies, GnIH-immunoreactive cells were found to be widely distributed throughout the GI tract including the esophagus, stomach, small intestine, and large intestine, in male piglets [98]. Additionally, low levels of GPR147 mRNA were detected in female pigs [100]. Both GnIH and GPR147 mRNA were also observed at low levels in the stomach, ileum, and colon of mice [101]. Xu et al. recently reported the potential role of GnIH in stress-induced intestinal dysfunction in domestic chickens. [105]. In this study, chronic IP injection of GnIH for 14 days not only induced intestinal and systemic stress but also led to the disruption of the physical, chemical, and microbial barriers of the intestine, as well as an increase in intestinal inflammation. The in vivo findings were further validated by treatment with GnIH in jejunal explants in vitro, revealing that GnIH directly damaged the intestinal barriers.
4.5. Liver and Skeletal Muscle
GnIH was not detected in liver and muscle of rats [96] and male piglets [98], whereas GPR147 was expressed in liver and skeletal muscle of rats at both mRNA and protein levels, to a similar extent as in WAT [96]. Hou et al. conducted a comparative analysis of GnIH-mediated effects in liver, skeletal muscle, and WAT [96]. Chronic GnIH injection (IP for 14 days) resulted in similar effects in liver and WAT, including decreased insulin receptor and GLUT4 mRNA expression and increased pro-inflammatory gene expression. Importantly, these changes were more evident in liver than in WAT, implying that liver might also serve as the target for GnIH action. Altered signaling pathways following GnIH administration showed similar patterns, marked by increased AKT phosphorylation and inhibited GSK3-β phosphorylation in both liver and WAT. In contrast, skeletal muscle exhibited a minimal response in GLUT4 and proinflammatory gene expression, along with inhibited AKT phosphorylation after GnIH administration. The changes observed in each tissue following GnIH administration are believed to contribute to the increased systemic insulin resistance and inflammatory response.
5. Nutritional Status and GnIH: Obesity and Fasting
In genetically obese (ob/ob) and HFD-induced obese mouse models, the expression of GnIH itself remained unchanged [89]. However, GPR147 KO female mice exhibited increased body weight gain during long-term HFD feeding compared to WT, associated with increased fat mass and decreased total energy expenditure [87]. In the same study, HFD-fed GPR147 KO male mice showed impaired glucose tolerance and insulin sensitivity without changes in body weight and body composition. These data suggest that GnIH/GPR147 signaling regulates metabolic homeostasis in a sexually dimorphic manner.
Increased expression of GnIH has been observed in response to fasting. In female hamsters, there was a gradual increase in the activation of GnIH neurons (cFOS activity) in the DMH, concomitant with an increase in the duration of food restriction for 12 days. These gradual changes in activation of GnIH neurons were remarkably similar to the changes in appetitive ingestive behavior (food hoarding) and were the exact opposite of changes in appetitive sexual behavior (male preference) [63,109,110]. In male zebra finches, 10 hours of fasting increased gonadal GnIH mRNA expression, but did not affect the hypothalamic GnIH expression at both immunoreactivity and mRNA levels [111]. It should be noted that fasting/food deprivation is a widely used stress protocol to induce the endocrine and metabolic changes in experimental animals, accompanied by elevated circulating corticosterone levels [112,113,114]. Thus, increased GnIH expression induced by fasting condition may be attributable to the direct effect of corticosterone on the GnIH system, as described in section 2.2.2.
6. Conclusions
Since its discovery in 2000, an increasing number of functions have been identified for GnIH neuropeptide. The widespread expression of GnIH and its receptors, in both the brain and peripheral tissues, suggests that it plays an expanding range of roles in general physiology, beyond its originally reported role in reproduction. For example, the orexigenic function of GnIH is now well accepted, as evidenced by many studies showing increased food intake following ICV administration. However, the close contact of GnIH fibers with NPY and POMC neurons does not fully explain the orexigenic effect, since their neuronal activities or gene expression patterns were not associated with the orexigenic changes caused by GnIH treatment in vivo and in vitro. Furthermore, there is no evidence for GnIH-R expression in these neurons. Therefore, the detailed mechanism linking the central GnIH system and feeding behavior requires further investigation. As described above, chronic IP administration of GnIH has been found to induce metabolic abnormalities, accompanied by significant alteration in peripheral tissue metabolism. The increased food intake observed following IP GnIH injection may be the primary contributor to weight gain and adiposity, but it is unclear how IP-injected GnIH stimulates food intake and which organ is most responsible for these GnIH-induced metabolic disorders. Currently there is insufficient evidence for the direct action of GnIH via its receptors other than the fact that GnIH/GnIH-Rs are expressed in the peripheral metabolic tissues. Further cellular, animal, and human studies are required to fully elucidate GnIH’s effects across various physiological contexts. In parallel, the detailed molecular mechanism of the GnIH action should be investigated to explore its potential as a therapeutic target.
Author Contributions
Y.L.S wrote the first draft of the manuscript. S.L.M and Y.T provided revisions that were incorporated by Y.L.S. All authors have read and approved the final manuscript.
Funding
This work was partly supported by JSPS KAKENHI 21K06240 and 24K0951104 to Y.L.S. and by UKRI Biotechnology and Biological Sciences Research Council (BBSRC) BB/5015760/1, and Roslin Institute Strategic Grant funding, BB/P013759/1 and BBS/E/RL/230001C, to S.L.M.
Institutional Review Bord Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank the students of the Birdsong Research Unit at Azabu University for assistant maintaining the Japanese quail.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tsutsui, K.; Saigoh, E.; Ukena, K.; Teranishi, H.; Fujisawa, Y.; Kikuchi, M.; Ishii, S.; Sharp, P.J. A novel avian hypothalamic peptide inhibiting gonadotropin release. Biochem Biophys Res Commun 2000, 275, 661-667. [CrossRef]
- Satake, H.; Hisada, M.; Kawada, T.; Minakata, H.; Ukena, K.; Tsutsui, K. Characterization of a cDNA encoding a novel avian hypothalamic neuropeptide exerting an inhibitory effect on gonadotropin release. Biochem J 2001, 354, 379-385. [CrossRef]
- Ikemoto, T.; Park, M.K. Chicken RFamide-related peptide (GnIH) and two distinct receptor subtypes: identification, molecular characterization, and evolutionary considerations. J Reprod Dev 2005, 51, 359-377. [CrossRef]
- Osugi, T.; Ukena, K.; Bentley, G.E.; O'Brien, S.; Moore, I.T.; Wingfield, J.C.; Tsutsui, K. Gonadotropin-inhibitory hormone in Gambel's white-crowned sparrow (Zonotrichia leucophrys gambelii): cDNA identification, transcript localization and functional effects in laboratory and field experiments. J Endocrinol 2004, 182, 33-42.
- Ubuka, T.; Kim, S.; Huang, Y.C.; Reid, J.; Jiang, J.; Osugi, T.; Chowdhury, V.S.; Tsutsui, K.; Bentley, G.E. Gonadotropin-inhibitory hormone neurons interact directly with gonadotropin-releasing hormone-I and -II neurons in European starling brain. Endocrinology 2008, 149, 268-278.
- Tobari, Y.; Iijima, N.; Tsunekawa, K.; Osugi, T.; Okanoya, K.; Tsutsui, K.; Ozawa, H. Identification of gonadotropin-inhibitory hormone in the zebra finch (Taeniopygia guttata): Peptide isolation, cDNA cloning and brain distribution. Peptides 2010, 31, 816-826. [CrossRef]
- Tsutsui, K. A new key neurohormone controlling reproduction, gonadotropin-inhibitory hormone (GnIH): Biosynthesis, mode of action and functional significance. Prog Neurobiol 2009, 88, 76-88. [CrossRef]
- Tsutsui, K.; Bentley, G.E.; Bedecarrats, G.; Osugi, T.; Ubuka, T.; Kriegsfeld, L.J. Gonadotropin-inhibitory hormone (GnIH) and its control of central and peripheral reproductive function. Front Neuroendocrinol 2010, 31, 284-295. [CrossRef]
- Tsutsui, K.; Ubuka, T.; Bentley, G.E.; Kriegsfeld, L.J. Review: regulatory mechanisms of gonadotropin-inhibitory hormone (GnIH) synthesis and release in photoperiodic animals. Front Neurosci 2013, 7, 60. [CrossRef]
- Hinuma, S.; Shintani, Y.; Fukusumi, S.; Iijima, N.; Matsumoto, Y.; Hosoya, M.; Fujii, R.; Watanabe, T.; Kikuchi, K.; Terao, Y.; et al. New neuropeptides containing carboxy-terminal RFamide and their receptor in mammals. Nat Cell Biol 2000, 2, 703-708.
- Bonini, J.A.; Jones, K.A.; Adham, N.; Forray, C.; Artymyshyn, R.; Durkin, M.M.; Smith, K.E.; Tamm, J.A.; Boteju, L.W.; Lakhlani, P.P.; et al. Identification and characterization of two G protein-coupled receptors for neuropeptide FF. J Biol Chem 2000, 275, 39324-39331. [CrossRef]
- Yin, H.; Ukena, K.; Ubuka, T.; Tsutsui, K. A novel G protein-coupled receptor for gonadotropin-inhibitory hormone in the Japanese quail (Coturnix japonica): identification, expression and binding activity. J Endocrinol 2005, 184, 257-266.
- Mollereau, C.; Mazarguil, H.; Marcus, D.; Quelven, I.; Kotani, M.; Lannoy, V.; Dumont, Y.; Quirion, R.; Detheux, M.; Parmentier, M.; et al. Pharmacological characterization of human NPFF(1) and NPFF(2) receptors expressed in CHO cells by using NPY Y(1) receptor antagonists. Eur J Pharmacol 2002, 451, 245-256. [CrossRef]
- Gouarderes, C.; Mazarguil, H.; Mollereau, C.; Chartrel, N.; Leprince, J.; Vaudry, H.; Zajac, J.M. Functional differences between NPFF1 and NPFF2 receptor coupling: high intrinsic activities of RFamide-related peptides on stimulation of [35S]GTPgammaS binding. Neuropharmacology 2007, 52, 376-386.
- Ubuka, T.; Son, Y.L.; Bentley, G.E.; Millar, R.P.; Tsutsui, K. Gonadotropin-inhibitory hormone (GnIH), GnIH receptor and cell signaling. Gen Comp Endocrinol 2013, 190, 10-17.
- Oishi, H.; Klausen, C.; Bentley, G.E.; Osugi, T.; Tsutsui, K.; Gilks, C.B.; Yano, T.; Leung, P.C. The human gonadotropin-inhibitory hormone ortholog RFamide-related peptide-3 suppresses gonadotropin-induced progesterone production in human granulosa cells. Endocrinology 2012, 153, 3435-3445. [CrossRef]
- Son, Y.L.; Ubuka, T.; Millar, R.P.; Kanasaki, H.; Tsutsui, K. Gonadotropin-inhibitory hormone inhibits GnRH-induced gonadotropin subunit gene transcriptions by inhibiting AC/cAMP/PKA-dependent ERK pathway in LbetaT2 cells. Endocrinology 2012, 153, 2332-2343.
- Son, Y.L.; Ubuka, T.; Soga, T.; Yamamoto, K.; Bentley, G.E.; Tsutsui, K. Inhibitory action of gonadotropin-inhibitory hormone on the signaling pathways induced by kisspeptin and vasoactive intestinal polypeptide in GnRH neuronal cell line, GT1-7. Faseb J 2016, 30, 2198-2210.
- Ubuka, T.; Morgan, K.; Pawson, A.J.; Osugi, T.; Chowdhury, V.S.; Minakata, H.; Tsutsui, K.; Millar, R.P.; Bentley, G.E. Identification of human GnIH homologs, RFRP-1 and RFRP-3, and the cognate receptor, GPR147 in the human hypothalamic pituitary axis. PLoS One 2009, 4, e8400. [CrossRef]
- Ubuka, T.; Inoue, K.; Fukuda, Y.; Mizuno, T.; Ukena, K.; Kriegsfeld, L.J.; Tsutsui, K. Identification, expression, and physiological functions of Siberian hamster gonadotropin-inhibitory hormone. Endocrinology 2012, 153, 373-385.
- Yano, T.; Iijima, N.; Kakihara, K.; Hinuma, S.; Tanaka, M.; Ibata, Y. Localization and neuronal response of RFamide related peptides in the rat central nervous system. Brain Res 2003, 982, 156-167. [CrossRef]
- Johnson, M.A.; Tsutsui, K.; Fraley, G.S. Rat RFamide-related peptide-3 stimulates GH secretion, inhibits LH secretion, and has variable effects on sex behavior in the adult male rat. Horm Behav 2007, 51, 171-180.
- Ukena, K.; Ubuka, T.; Tsutsui, K. Distribution of a novel avian gonadotropin-inhibitory hormone in the quail brain. Cell Tissue Res 2003, 312, 73-79.
- Ubuka, T.; Ueno, M.; Ukena, K.; Tsutsui, K. Developmental changes in gonadotropin-inhibitory hormone in the Japanese quail (Coturnix japonica) hypothalamo-hypophysial system. J Endocrinol 2003, 178, 311-318. [CrossRef]
- Kriegsfeld, L.J.; Mei, D.F.; Bentley, G.E.; Ubuka, T.; Mason, A.O.; Inoue, K.; Ukena, K.; Tsutsui, K.; Silver, R. Identification and characterization of a gonadotropin-inhibitory system in the brains of mammals. Proc Natl Acad Sci U S A 2006, 103, 2410-2415.
- Legagneux, K.; Bernard-Franchi, G.; Poncet, F.; La Roche, A.; Colard, C.; Fellmann, D.; Pralong, F.; Risold, P.Y. Distribution and genesis of the RFRP-producing neurons in the rat brain: comparison with melanin-concentrating hormone- and hypocretin-containing neurons. Neuropeptides 2009, 43, 13-19.
- Revel, F.G.; Saboureau, M.; Pevet, P.; Simonneaux, V.; Mikkelsen, J.D. RFamide-related peptide gene is a melatonin-driven photoperiodic gene. Endocrinology 2008, 149, 902-912. [CrossRef]
- Ukena, K.; Tsutsui, K. Distribution of novel RFamide-related peptide-like immunoreactivity in the mouse central nervous system. Neurosci Lett 2001, 300, 153-156.
- Mason, A.O.; Duffy, S.; Zhao, S.; Ubuka, T.; Bentley, G.E.; Tsutsui, K.; Silver, R.; Kriegsfeld, L.J. Photoperiod and reproductive condition are associated with changes in RFamide-related peptide (RFRP) expression in Syrian hamsters (Mesocricetus auratus). J Biol Rhythms 2010, 25, 176-185. [CrossRef]
- Tsutsui, K.; Bentley, G.E.; Kriegsfeld, L.J.; Osugi, T.; Seong, J.Y.; Vaudry, H. Discovery and evolutionary history of gonadotrophin-inhibitory hormone and kisspeptin: new key neuropeptides controlling reproduction. J Neuroendocrinol 2010, 22, 716-727.
- Tsutsui, K.; Ubuka, T.; Bentley, G.E.; Kriegsfeld, L.J. Gonadotropin-inhibitory hormone (GnIH): discovery, progress and prospect. Gen Comp Endocrinol 2012, 177, 305-314. [CrossRef]
- Tsutsui, K.; Ubuka, T.; Son, Y.L.; Bentley, G.E.; Kriegsfeld, L.J. Contribution of GnIH Research to the Progress of Reproductive Neuroendocrinology. Front Endocrinol (Lausanne) 2015, 6, 179. [CrossRef]
- Kriegsfeld, L.J.; Ubuka, T.; Bentley, G.E.; Tsutsui, K. Seasonal control of gonadotropin-inhibitory hormone (GnIH) in birds and mammals. Front Neuroendocrinol 2015, 37, 65-75.
- Smith, J.T.; Coolen, L.M.; Kriegsfeld, L.J.; Sari, I.P.; Jaafarzadehshirazi, M.R.; Maltby, M.; Bateman, K.; Goodman, R.L.; Tilbrook, A.J.; Ubuka, T.; et al. Variation in kisspeptin and RFamide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain: a novel medium for seasonal breeding in the sheep. Endocrinology 2008, 149, 5770-5782.
- Kriegsfeld, L.J.; Gibson, E.M.; Williams, W.P., 3rd; Zhao, S.; Mason, A.O.; Bentley, G.E.; Tsutsui, K. The roles of RFamide-related peptide-3 in mammalian reproductive function and behaviour. J Neuroendocrinol 2010, 22, 692-700.
- Rizwan, M.Z.; Poling, M.C.; Corr, M.; Cornes, P.A.; Augustine, R.A.; Quennell, J.H.; Kauffman, A.S.; Anderson, G.M. RFamide-related peptide-3 receptor gene expression in GnRH and kisspeptin neurons and GnRH-dependent mechanism of action. Endocrinology 2012, 153, 3770-3779. [CrossRef]
- Bentley, G.E.; Perfito, N.; Ukena, K.; Tsutsui, K.; Wingfield, J.C. Gonadotropin-inhibitory peptide in song sparrows (Melospiza melodia) in different reproductive conditions, and in house sparrows (Passer domesticus) relative to chicken-gonadotropin-releasing hormone. J Neuroendocrinol 2003, 15, 794-802.
- Ducret, E.; Anderson, G.M.; Herbison, A.E. RFamide-related peptide-3, a mammalian gonadotropin-inhibitory hormone ortholog, regulates gonadotropin-releasing hormone neuron firing in the mouse. Endocrinology 2009, 150, 2799-2804.
- Wu, M.; Dumalska, I.; Morozova, E.; van den Pol, A.N.; Alreja, M. Gonadotropin inhibitory hormone inhibits basal forebrain vGluT2-gonadotropin-releasing hormone neurons via a direct postsynaptic mechanism. J Physiol 2009, 587, 1401-1411. [CrossRef]
- Poling, M.C.; Quennell, J.H.; Anderson, G.M.; Kauffman, A.S. Kisspeptin neurones do not directly signal to RFRP-3 neurones but RFRP-3 may directly modulate a subset of hypothalamic kisspeptin cells in mice. J Neuroendocrinol 2013, 25, 876-886.
- Maddineni, S.; Ocon-Grove, O.M.; Krzysik-Walker, S.M.; Hendricks, G.L., 3rd; Proudman, J.A.; Ramachandran, R. Gonadotrophin-inhibitory hormone receptor expression in the chicken pituitary gland: potential influence of sexual maturation and ovarian steroids. J Neuroendocrinol 2008, 20, 1078-1088.
- Clarke, I.J.; Sari, I.P.; Qi, Y.; Smith, J.T.; Parkington, H.C.; Ubuka, T.; Iqbal, J.; Li, Q.; Tilbrook, A.; Morgan, K.; et al. Potent action of RFamide-related peptide-3 on pituitary gonadotropes indicative of a hypophysiotropic role in the negative regulation of gonadotropin secretion. Endocrinology 2008, 149, 5811-5821. [CrossRef]
- Ubuka, T.; Lai, H.; Kitani, M.; Suzuuchi, A.; Pham, V.; Cadigan, P.A.; Wang, A.; Chowdhury, V.S.; Tsutsui, K.; Bentley, G.E. Gonadotropin-inhibitory hormone identification, cDNA cloning, and distribution in rhesus macaque brain. J Comp Neurol 2009, 517, 841-855.
- Murakami, M.; Matsuzaki, T.; Iwasa, T.; Yasui, T.; Irahara, M.; Osugi, T.; Tsutsui, K. Hypophysiotropic role of RFamide-related peptide-3 in the inhibition of LH secretion in female rats. J Endocrinol 2008, 199, 105-112.
- Gibson, E.M.; Humber, S.A.; Jain, S.; Williams, W.P., 3rd; Zhao, S.; Bentley, G.E.; Tsutsui, K.; Kriegsfeld, L.J. Alterations in RFamide-related peptide expression are coordinated with the preovulatory luteinizing hormone surge. Endocrinology 2008, 149, 4958-4969.
- Kadokawa, H.; Shibata, M.; Tanaka, Y.; Kojima, T.; Matsumoto, K.; Oshima, K.; Yamamoto, N. Bovine C-terminal octapeptide of RFamide-related peptide-3 suppresses luteinizing hormone (LH) secretion from the pituitary as well as pulsatile LH secretion in bovines. Domest Anim Endocrinol 2009, 36, 219-224.
- Sari, I.P.; Rao, A.; Smith, J.T.; Tilbrook, A.J.; Clarke, I.J. Effect of RF-amide-related peptide-3 on luteinizing hormone and follicle-stimulating hormone synthesis and secretion in ovine pituitary gonadotropes. Endocrinology 2009, 150, 5549-5556.
- Bentley, G.E.; Ubuka, T.; McGuire, N.L.; Chowdhury, V.S.; Morita, Y.; Yano, T.; Hasunuma, I.; Binns, M.; Wingfield, J.C.; Tsutsui, K. Gonadotropin-inhibitory hormone and its receptor in the avian reproductive system. Gen Comp Endocrinol 2008, 156, 34-43. [CrossRef]
- McGuire, N.L.; Bentley, G.E. A functional neuropeptide system in vertebrate gonads: Gonadotropin-inhibitory hormone and its receptor in testes of field-caught house sparrow (Passer domesticus). Gen Comp Endocrinol 2010, 166, 565-572. [CrossRef]
- Zhao, S.; Zhu, E.; Yang, C.; Bentley, G.E.; Tsutsui, K.; Kriegsfeld, L.J. RFamide-related peptide and messenger ribonucleic acid expression in mammalian testis: association with the spermatogenic cycle. Endocrinology 2010, 151, 617-627. [CrossRef]
- Qi, Y.; Oldfield, B.J.; Clarke, I.J. Projections of RFamide-related peptide-3 neurones in the ovine hypothalamus, with special reference to regions regulating energy balance and reproduction. J Neuroendocrinol 2009, 21, 690-697.
- Kirby, E.D.; Geraghty, A.C.; Ubuka, T.; Bentley, G.E.; Kaufer, D. Stress increases putative gonadotropin inhibitory hormone and decreases luteinizing hormone in male rats. Proc Natl Acad Sci U S A 2009, 106, 11324-11329. [CrossRef]
- Son, Y.L.; Ubuka, T.; Narihiro, M.; Fukuda, Y.; Hasunuma, I.; Yamamoto, K.; Belsham, D.D.; Tsutsui, K. Molecular basis for the activation of gonadotropin-inhibitory hormone gene transcription by corticosterone. Endocrinology 2014, 155, 1817-1826. [CrossRef]
- Geraghty, A.C.; Muroy, S.E.; Zhao, S.; Bentley, G.E.; Kriegsfeld, L.J.; Kaufer, D. Knockdown of hypothalamic RFRP3 prevents chronic stress-induced infertility and embryo resorption. Elife 2015, 4. [CrossRef]
- Moustafa, A. Changes in nitric oxide, carbon monoxide, hydrogen sulfide and male reproductive hormones in response to chronic restraint stress in rats. Free radical biology & medicine 2021, 162, 353-366. [CrossRef]
- Yang, J.A.; Song, C.I.; Hughes, J.K.; Kreisman, M.J.; Parra, R.A.; Haisenleder, D.J.; Kauffman, A.S.; Breen, K.M. Acute Psychosocial Stress Inhibits LH Pulsatility and Kiss1 Neuronal Activation in Female Mice. Endocrinology 2017, 158, 3716-3723. [CrossRef]
- Yang, J.A.; Hughes, J.K.; Parra, R.A.; Volk, K.M.; Kauffman, A.S. Stress rapidly suppresses in vivo LH pulses and increases activation of RFRP-3 neurons in male mice. J Endocrinol 2018, 239, 339-350. [CrossRef]
- Mamgain, A.; Sawyer, I.L.; Timajo, D.A.M.; Rizwan, M.Z.; Evans, M.C.; Ancel, C.M.; Inglis, M.A.; Anderson, G.M. RFamide-Related Peptide Neurons Modulate Reproductive Function and Stress Responses. J Neurosci 2021, 41, 474-488. [CrossRef]
- Calisi, R.M.; Rizzo, N.O.; Bentley, G.E. Seasonal differences in hypothalamic EGR-1 and GnIH expression following capture-handling stress in house sparrows (Passer domesticus). Gen Comp Endocrinol 2008, 157, 283-287.
- León, S.; García-Galiano, D.; Ruiz-Pino, F.; Barroso, A.; Manfredi-Lozano, M.; Romero-Ruiz, A.; Roa, J.; Vázquez, M.J.; Gaytan, F.; Blomenrohr, M.; et al. Physiological roles of gonadotropin-inhibitory hormone signaling in the control of mammalian reproductive axis: studies in the NPFF1 receptor null mouse. Endocrinology 2014, 155, 2953-2965. [CrossRef]
- Simonin, F.; Schmitt, M.; Laulin, J.-P.; Laboureyras, E.; Jhamandas, J.H.; MacTavish, D.; Matifas, A.; Mollereau, C.; Laurent, P.; Parmentier, M.; et al. RF9, a potent and selective neuropeptide FF receptor antagonist, prevents opioid-induced tolerance associated with hyperalgesia. Proceedings of the National Academy of Sciences of the United States of America 2006, 103, 466-471. [CrossRef]
- Batool, A.; Naz, R.; Wazir, M.; Azam, A.; Ullah, R.; Wahab, F.; Shahab, M. Acute fasting-induced repression of the hypothalamic-pituitary-gonadal axis is reversed by RF-9 administration in the adult male macaque. Horm Metab Res 2014, 46, 927-832. [CrossRef]
- Schneider, J.E.; Benton, N.A.; Russo, K.A.; Klingerman, C.M.; Williams, W.P., 3rd; Simberlund, J.; Abdulhay, A.; Brozek, J.M.; Kriegsfeld, L.J. RFamide-related Peptide-3 and the Trade-off between Reproductive and Ingestive Behavior. Integrative and comparative biology 2017, 57, 1225-1239. [CrossRef]
- Chowdhury, V.S.; Tomonaga, S.; Nishimura, S.; Tabata, S.; Cockrem, J.F.; Tsutsui, K.; Furuse, M. Hypothalamic gonadotropin-inhibitory hormone precursor mRNA is increased during depressed food intake in heat-exposed chicks. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2012, 162, 227-233. [CrossRef]
- Bahry, M.A.; Yang, H.; Tran, P.V.; Do, P.H.; Han, G.; Eltahan, H.M.; Chowdhury, V.S.; Furuse, M. Reduction in voluntary food intake, but not fasting, stimulates hypothalamic gonadotropin-inhibitory hormone precursor mRNA expression in chicks under heat stress. Neuropeptides 2018, 71, 90-96. [CrossRef]
- Son, Y.L.; Ubuka, T.; Tsutsui, K. Regulation of stress response on the hypothalamic-pituitary-gonadal axis via gonadotropin-inhibitory hormone. Front Neuroendocrinol 2022, 64, 100953. [CrossRef]
- McGuire, N.L.; Koh, A.; Bentley, G.E. The direct response of the gonads to cues of stress in a temperate songbird species is season-dependent. PeerJ 2013, 1, e139. [CrossRef]
- Clarke, I.J.; Bartolini, D.; Conductier, G.; Henry, B.A. Stress Increases Gonadotropin Inhibitory Hormone Cell Activity and Input to GnRH Cells in Ewes. Endocrinology 2016, 157, 4339-4350. [CrossRef]
- Samson, W.K.; Keown, C.; Samson, C.K.; Samson, H.W.; Lane, B.; Baker, J.R.; Taylor, M.M. Prolactin-releasing peptide and its homolog RFRP-1 act in hypothalamus but not in anterior pituitary gland to stimulate stress hormone secretion. Endocrine 2003, 20, 59-66. [CrossRef]
- Kim, J.S.; Brownjohn, P.W.; Dyer, B.S.; Beltramo, M.; Walker, C.S.; Hay, D.L.; Painter, G.F.; Tyndall, J.D.; Anderson, G.M. Anxiogenic and Stressor Effects of the Hypothalamic Neuropeptide RFRP-3 Are Overcome by the NPFFR Antagonist GJ14. Endocrinology 2015, 156, 4152-4162. [CrossRef]
- Prevot, V.; Croix, D.; Bouret, S.; Dutoit, S.; Tramu, G.; Stefano, G.B.; Beauvillain, J.C. Definitive evidence for the existence of morphological plasticity in the external zone of the median eminence during the rat estrous cycle: implication of neuro-glio-endothelial interactions in gonadotropin-releasing hormone release. Neuroscience 1999, 94, 809-819. [CrossRef]
- Yamamura, T.; Hirunagi, K.; Ebihara, S.; Yoshimura, T. Seasonal morphological changes in the neuro-glial interaction between gonadotropin-releasing hormone nerve terminals and glial endfeet in Japanese quail. Endocrinology 2004, 145, 4264-4267. [CrossRef]
- Yamamura, T.; Yasuo, S.; Hirunagi, K.; Ebihara, S.; Yoshimura, T. T(3) implantation mimics photoperiodically reduced encasement of nerve terminals by glial processes in the median eminence of Japanese quail. Cell Tissue Res 2006, 324, 175-179. [CrossRef]
- Kiyohara, M.; Son, Y.L.; Tsutsui, K. Involvement of gonadotropin-inhibitory hormone in pubertal disorders induced by thyroid status. Sci Rep 2017, 7, 1042. [CrossRef]
- Tachibana, T.; Sato, M.; Takahashi, H.; Ukena, K.; Tsutsui, K.; Furuse, M. Gonadotropin-inhibiting hormone stimulates feeding behavior in chicks. Brain Res 2005, 1050, 94-100. [CrossRef]
- Clarke, I.J.; Smith, J.T.; Henry, B.A.; Oldfield, B.J.; Stefanidis, A.; Millar, R.P.; Sari, I.P.; Chng, K.; Fabre-Nys, C.; Caraty, A.; et al. Gonadotropin-inhibitory hormone is a hypothalamic peptide that provides a molecular switch between reproduction and feeding. Neuroendocrinology 2012, 95, 305-316.
- Fraley, G.S.; Coombs, E.; Gerometta, E.; Colton, S.; Sharp, P.J.; Li, Q.; Clarke, I.J. Distribution and sequence of gonadotropin-inhibitory hormone and its potential role as a molecular link between feeding and reproductive systems in the Pekin duck (Anas platyrhynchos domestica). Gen Comp Endocrinol 2013, 184, 103-110. [CrossRef]
- McConn, B.; Wang, G.; Yi, J.; Gilbert, E.R.; Osugi, T.; Ubuka, T.; Tsutsui, K.; Chowdhury, V.S.; Furuse, M.; Cline, M.A. Gonadotropin-inhibitory hormone-stimulation of food intake is mediated by hypothalamic effects in chicks. Neuropeptides 2014, 48, 327-334. [CrossRef]
- Moriwaki, S.; Narimatsu, Y.; Fukumura, K.; Iwakoshi-Ukena, E.; Furumitsu, M.; Ukena, K. Effects of Chronic Intracerebroventricular Infusion of RFamide-Related Peptide-3 on Energy Metabolism in Male Mice. Int J Mol Sci 2020, 21. [CrossRef]
- Cazarez-Marquez, F.; Milesi, S.; Laran-Chich, M.P.; Klosen, P.; Kalsbeek, A.; Simonneaux, V. Kisspeptin and RFRP3 modulate body mass in Phodopus sungorus via two different neuroendocrine pathways. J Neuroendocrinol 2019, 31, e12710. [CrossRef]
- Cázarez-Márquez, F.; Laran-Chich, M.P.; Klosen, P.; Kalsbeek, A.; Simonneaux, V. RFRP3 increases food intake in a sex-dependent manner in the seasonal hamster Phodopus sungorus. J Neuroendocrinol 2020, 32, e12845. [CrossRef]
- Cázarez-Márquez, F.; Eliveld, J.; Ritsema, W.; Foppen, E.; Bossenbroek, Y.; Pelizzari, S.; Simonneaux, V.; Kalsbeek, A. Role of central kisspeptin and RFRP-3 in energy metabolism in the male Wistar rat. J Neuroendocrinol 2021, 33, e12973. [CrossRef]
- Kovacs, A.; Laszlo, K.; Galosi, R.; Toth, K.; Ollmann, T.; Peczely, L.; Lenard, L. Microinjection of RFRP-1 in the central nucleus of amygdala decreases food intake in the rat. Brain Res Bull 2012, 88, 589-595. [CrossRef]
- Liu, Q.; Guan, X.M.; Martin, W.J.; McDonald, T.P.; Clements, M.K.; Jiang, Q.; Zeng, Z.; Jacobson, M.; Williams, D.L., Jr.; Yu, H.; et al. Identification and characterization of novel mammalian neuropeptide FF-like peptides that attenuate morphine-induced antinociception. J Biol Chem 2001, 276, 36961-36969. [CrossRef]
- Jacobi, J.S.; Coleman, H.A.; Enriori, P.J.; Parkington, H.C.; Li, Q.; Pereira, A.; Cowley, M.A.; Clarke, I.J. Paradoxical effect of gonadotrophin-inhibiting hormone to negatively regulate neuropeptide Y neurones in mouse arcuate nucleus. J Neuroendocrinol 2013, 25, 1308-1317. [CrossRef]
- Fu, L.Y.; van den Pol, A.N. Kisspeptin directly excites anorexigenic proopiomelanocortin neurons but inhibits orexigenic neuropeptide Y cells by an indirect synaptic mechanism. J Neurosci 2010, 30, 10205-10219. [CrossRef]
- Leon, S.; Velasco, I.; Vázquez, M.J.; Barroso, A.; Beiroa, D.; Heras, V.; Ruiz-Pino, F.; Manfredi-Lozano, M.; Romero-Ruiz, A.; Sanchez-Garrido, M.A.; et al. Sex-Biased Physiological Roles of NPFF1R, the Canonical Receptor of RFRP-3, in Food Intake and Metabolic Homeostasis Revealed by its Congenital Ablation in mice. Metabolism: clinical and experimental 2018, 87, 87-97. [CrossRef]
- Poling, M.C.; Shieh, M.P.; Munaganuru, N.; Luo, E.; Kauffman, A.S. Examination of the influence of leptin and acute metabolic challenge on RFRP-3 neurons of mice in development and adulthood. Neuroendocrinology 2014, 100, 317-333. [CrossRef]
- Rizwan, M.Z.; Harbid, A.A.; Inglis, M.A.; Quennell, J.H.; Anderson, G.M. Evidence that hypothalamic RFamide related peptide-3 neurones are not leptin-responsive in mice and rats. J Neuroendocrinol 2014, 26, 247-257. [CrossRef]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschop, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003, 37, 649-661. [CrossRef]
- Lu, S.; Guan, J.L.; Wang, Q.P.; Uehara, K.; Yamada, S.; Goto, N.; Date, Y.; Nakazato, M.; Kojima, M.; Kangawa, K.; et al. Immunocytochemical observation of ghrelin-containing neurons in the rat arcuate nucleus. Neurosci Lett 2002, 321, 157-160. [CrossRef]
- Kageyama, H.; Kitamura, Y.; Hosono, T.; Kintaka, Y.; Seki, M.; Takenoya, F.; Hori, Y.; Nonaka, N.; Arata, S.; Shioda, S. Visualization of ghrelin-producing neurons in the hypothalamic arcuate nucleus using ghrelin-EGFP transgenic mice. Regulatory peptides 2008, 145, 116-121. [CrossRef]
- Celik, O.; Celik, N.; Aydin, S.; Aygun, B.K.; Haberal, E.T.; Kuloglu, T.; Ulas, M.; Aktun, L.H.; Acet, M.; Celik, S. Ghrelin action on GnRH neurons and pituitary gonadotropes might be mediated by GnIH-GPR147 system. Hormone molecular biology and clinical investigation 2016, 25, 121-128. [CrossRef]
- Singh, P.; Anjum, S.; Srivastava, R.K.; Tsutsui, K.; Krishna, A. Central and peripheral neuropeptide RFRP-3: A bridge linking reproduction, nutrition, and stress response. Front Neuroendocrinol 2022, 65, 100979. [CrossRef]
- van Harmelen, V.; Dicker, A.; Sjolin, E.; Blomqvist, L.; Wiren, M.; Hoffstedt, J.; Ryden, M.; Arner, P. Effects of pain controlling neuropeptides on human fat cell lipolysis. Int J Obes (Lond) 2010, 34, 1333-1340. [CrossRef]
- Huo, K.; Li, X.; Hu, W.; Song, X.; Zhang, D.; Zhang, X.; Chen, X.; Yuan, J.; Zuo, J.; Wang, X. RFRP-3, the Mammalian Ortholog of GnIH, Is a Novel Modulator Involved in Food Intake and Glucose Homeostasis. Front Endocrinol (Lausanne) 2020, 11, 194. [CrossRef]
- Lefrere, I.; De Coppet, P.; Camelin, J.C.; Le Lay, S.; Mercier, N.; Elshourbagy, N.; Bril, A.; Berrebi-Bertrand, I.; Feve, B.; Krief, S. Neuropeptide AF and FF modulation of adipocyte metabolism. Primary insights from functional genomics and effects on beta-adrenergic responsiveness. J Biol Chem 2002, 277, 39169-39178. [CrossRef]
- Wang, X.; Li, X.; Hu, C. Distribution of gonadotropin-inhibitory hormone (GnIH) in male Luchuan piglets. Gene expression patterns : GEP 2018, 28, 42-53. [CrossRef]
- Zhang, W.; Wang, L.; Yu, X.; Jia, A.; Ming, J.; Ji, Q. RFamide-related peptide-3 promotes alpha TC1 clone 6 cell survival likely via GPR147. Peptides 2018, 107, 39-44. [CrossRef]
- Li, X.; Su, J.; Lei, Z.; Zhao, Y.; Jin, M.; Fang, R.; Zheng, L.; Jiao, Y. Gonadotropin-inhibitory hormone (GnIH) and its receptor in the female pig: cDNA cloning, expression in tissues and expression pattern in the reproductive axis during the estrous cycle. Peptides 2012, 36, 176-185. [CrossRef]
- Gospodarska, E.; Kozak, L.P.; Jaroslawska, J. Isolation and identification of endogenous RFamide-related peptides 1 and 3 in the mouse hypothalamus. J Neuroendocrinol 2019, 31, e12668. [CrossRef]
- Anjum, S.; Krishna, A.; Tsutsui, K. Possible Role of GnIH as a Mediator between Adiposity and Impaired Testicular Function. Front Endocrinol (Lausanne) 2016, 7, 6. [CrossRef]
- Luo, R.; Chen, L.; Song, X.; Zhang, X.; Xu, W.; Han, D.; Zuo, J.; Hu, W.; Shi, Y.; Cao, Y.; et al. Possible Role of GnIH as a Novel Link between Hyperphagia-Induced Obesity-Related Metabolic Derangements and Hypogonadism in Male Mice. Int J Mol Sci 2022, 23. [CrossRef]
- Chen, L.; Zhang, X.; Song, X.; Han, D.; Han, K.; Xu, W.; Luo, R.; Cao, Y.; Shi, Y.; Liu, C.; et al. Peripheral Gonadotropin-Inhibitory Hormone (GnIH) Acting as a Novel Modulator Involved in Hyperphagia-Induced Obesity and Associated Disorders of Metabolism in an In Vivo Female Piglet Model. Int J Mol Sci 2022, 23. [CrossRef]
- Xu, C.; Han, D.; Song, X.; Zhang, X.; Liu, C.; Zhang, J.; Shen, B.; Li, Z.; Ma, R.; Li, Y.; et al. The possibly role of GnIH in stress and gut dysfunction in chicken. Poultry science 2024, 103, 103757. [CrossRef]
- De Jong, K.A.; Siddig, S.; Pfeifer, A.; Nikolaev, V.O. The role of compartmentalized beta-AR/cAMP signaling in the regulation of lipolysis in white and brown adipocytes. The FEBS journal 2025, 292, 261-271. [CrossRef]
- Son, Y.L.; Ubuka, T.; Millar, R.P.; Kanasaki, H.; Tsutsui, K. Gonadotropin-inhibitory hormone inhibits GnRH-induced gonadotropin subunit gene transcriptions by inhibiting AC/cAMP/PKA-dependent ERK pathway in LβT2 cells. Endocrinology 2012, 153, 2332-2343. [CrossRef]
- Granata, R.; Settanni, F.; Trovato, L.; Gallo, D.; Gesmundo, I.; Nano, R.; Gallo, M.P.; Bergandi, L.; Volante, M.; Alloatti, G.; et al. RFamide peptides 43RFa and 26RFa both promote survival of pancreatic beta-cells and human pancreatic islets but exert opposite effects on insulin secretion. Diabetes 2014, 63, 2380-2393. [CrossRef]
- Benton, N.A.; Russo, K.A.; Brozek, J.M.; Andrews, R.J.; Kim, V.J.; Kriegsfeld, L.J.; Schneider, J.E. Food restriction-induced changes in motivation differ with stages of the estrous cycle and are closely linked to RFamide-related peptide-3 but not kisspeptin in Syrian hamsters. Physiol Behav 2018, 190, 43-60. [CrossRef]
- Klingerman, C.M.; Williams, W.P., 3rd; Simberlund, J.; Brahme, N.; Prasad, A.; Schneider, J.E.; Kriegsfeld, L.J. Food Restriction-Induced Changes in Gonadotropin-Inhibiting Hormone Cells are Associated with Changes in Sexual Motivation and Food Hoarding, but not Sexual Performance and Food Intake. Front Endocrinol (Lausanne) 2011, 2, 101. [CrossRef]
- Lynn, S.E.; Perfito, N.; Guardado, D.; Bentley, G.E. Food, stress, and circulating testosterone: Cue integration by the testes, not the brain, in male zebra finches (Taeniopygia guttata). Gen Comp Endocrinol 2015, 215, 1-9. [CrossRef]
- Tang, F.; Hsieh, A.C.; Lee, C.P.; Baconshone, J. Interaction of cold and starvation in the regulation of plasma corticosterone levels in the male rat. Horm Metab Res 1984, 16, 445-448. [CrossRef]
- Pirke, K.M.; Spyra, B. Catecholamine turnover in the brain and the regulation of luteinizing hormone and corticosterone in starved male rats. Acta Endocrinol (Copenh) 1982, 100, 168-176. [CrossRef]
- Palmblad, J.; Levi, L.; Burger, A.; Melander, A.; Westgren, U.; von Schenck, H.; Skude, G. Effects of total energy withdrawal (fasting) on thelevels of growth hormone, thyrotropin, cortisol, adrenaline, noradrenaline, T4, T3, and rT3 in healthy males. Acta Med Scand 1977, 201, 15-22. [CrossRef]

Table 1.
Effect of central GnIH administration on metabolism. Avian GnIH peptide or mammalian RFRP-3 peptide is used for ICV injection, except for intra-amygdaloid microinjection of RFRP-1 peptide.
Table 1.
Effect of central GnIH administration on metabolism. Avian GnIH peptide or mammalian RFRP-3 peptide is used for ICV injection, except for intra-amygdaloid microinjection of RFRP-1 peptide.
| Animal models | Injection | Metabolic effects | Ref |
|---|---|---|---|
| Male domestic chicken chick | Single ICV (0.3, 0.9, or 2.6 nmol) | ・Increased food intake at 1 and 2 h | [75] 2005 |
| Single ICV of anti-GnIH antiserum | ・Reduction in food deprivation-induced feeding ・No effect on ad libitum feeding |
||
| Male domestic chicken chick | Single ICV (0.9, 2.6, or 7.8 nmol) | ・Increased food intake at 0.5 to 2 h ・No effect on water intake ・Increased feeding pecks at 5 min to 30 min |
[78] 2014 |
| Adult male Pekin duck | Single ICV (100 ng) | ・Increased food intake at 2 h | [77] 2013 |
| Adult male mouse | Single ICV (25, 50, or 100 ng) | ・Increased food intake at 0.5, 1, 2 h | [76] 2012 |
| Adult male mouse | ICV (6 nmol/day) for 13 days | ・Increased food intake, BW, BAT and liver mass ・No effects on WATs and muscle mass ・Decreased O2/CO2 metabolism, energy expenditure, and core body temperature1 ・No effects on locomotor activity |
[79] 2020 |
| SD-adapted male hamster | ICV (8.25 pmol/h) for 5 weeks | ・Increased BW and food intake ・Increased circulating insulin and leptin ・No effects on hypothalamic metabolic genes |
[80] 2019 |
| SD-adapted female hamster | ・No effects on BW, food intake, circulating insulin/leptin, and hypothalamic metabolic genes | ||
| Male hamster | Single ICV (0.5 or 1.5 µg) | ・No effect on food intake regardless of LD or SD | [81] 2020 |
| Female hamster | ・Increased food intake in both LD (0.5 µg) and SD (1.5 µg), and NPY expression at 3 h | ||
| Adult male rat | Single ICV (100 or 500 ng) | ・Increased food intake at 2 h ・No effect on BW at 24 h |
[22] 2007 |
| Adult male rat | ICV (1 µg/h) for 5 days | ・Increased food and water intake ・No effects on whole-body energy expenditure and BAT thermogenesis |
[76] 2012 |
| Adult male rat | Single ICV (50 or 250 pmol) | ・No effect on food intake | [82] 2021 |
| Adult male rat | Intraamygdaloid microinjection of RFRP-1 (37.8 pmol) | ・Decreased liquid food intake over 1 h ・No effect on locomotor activity |
[83] 2012 |
| RF9 (41.4 pmol) + RFRP-1 | ・Prevents RFRP-1 effect on food intake | ||
| Ovariectomized adult female sheep | ICV (40 µg/h) for 4 h | ・Increased food intake at 2, 4 h ・No effects on thermogenesis of muscle and visceral WAT |
[76] 2012 |
| Adult male cynomolgus macaque monkey | ICV (3 µg/kg/h) for 9 days | ・Increased food intake | [76] 2012 |
1 Measured during a short time-period in the dark phase. 2. NPY, POMC, and somatostatin expression in the hypothalamus. BW, body weight; BAT, brown adipose tissue; WAT, white adipose tissue; SD or LD, short-day or long-day photoperiod.
Table 2.
Expression of GnIH or GnIH-Rs in peripheral metabolic tissue.
| Tissue | Subject | Expression of GnIH or GnIH-Rs | Method | Ref | |
|---|---|---|---|---|---|
| Adipose tissue | Human | Tissue | GPR147/GPR74 | RT-PCR | [95] |
| Mature adipocyte | GPR147/GPR74 | Western | |||
| Rat | Tissue | GPR147 | RT-PCR, Western | [96] | |
| Mouse | Tissue | GPR147/GPR74 | RT-PCR | Figure 1 | |
| Japanese quail | Tissue | GPR147 | RT-PCR | Figure 1 | |
| Mouse 3T3-L11 | Mature adipocyte2 | GPR147/GPR74 | RT-PCR | [97] | |
| Pancreas | Piglet | Islet | GnIH | Immunostaining | [98] |
| Rat | Tissue | GnIH and GPR147 | RT-PCR, Western, Immunostaining |
[96] | |
| Mouse | Islet | GPR147 | Immunostaining | [99] | |
| Mouse αTC11 | α-cells of islet | GPR147 | RT-PCR, Immunostaining | [99] | |
| GI tract | Piglet | Esophagus, stomach, small and large intestine | GnIH | Immunostaining | [98] |
| Female pig | Intestine | GPR147 | RT-PCR | [100] | |
| Mouse | Stomach, ileum, and colon | GnIH and GPR147 | RT-PCR | [101] | |
| Liver | Rat | Tissue | GPR147 | RT-PCR, Western | [96] |
| Skeletal muscle | Rat | Tissue | GPR147 | RT-PCR, Western | [96] |
1 In vitro cell models. 2 In vitro differentiated 3T3-L1 cells.
Table 3.
Effect of peripheral GnIH administration on metabolism. Chicken GnIH peptide or mammalian RFRP-3 peptide is used for IP injection.
Table 3.
Effect of peripheral GnIH administration on metabolism. Chicken GnIH peptide or mammalian RFRP-3 peptide is used for IP injection.
| Animal models | IP injection of GnIH | Metabolic effects | Ref. |
|---|---|---|---|
| Female domestic chicken | 30 nmol × twice/day for 14 days | ・Disrupts the physical and chemical barriers of the intestine ・Increased intestinal inflammation |
[105] 2024 |
| Male mouse | 20 ng, 200 ng, or 2 µg/day for 8 days | ・Increased food intake ・Increased BW ・Increased WAT mass |
[102] 2016 |
| Male mouse | 20 µg × twice/day for 21 days | ・Increased food intake and BW ・Increased liver and eWAT mass ・Decreased testis mass ・Glucose intolerance and insulin resistance |
[103] 2022 |
| Rat (male and female mixed population) | 1 or 10 µg × twice/day for 14 days | ・Increased food intake during photophase ・Increased meal frequency ・Increased BW ・Glucose intolerance and insulin resistance ・Increased inflammation in liver, skeletal muscle, or WAT |
[96] 2020 |
| Female piglet | 0.1 or 1 mg × twice/day for 14 days | ・Increased food intake and BW ・Increased organ mass in pancreas, pgWAT, iWAT and liver ・Glucose intolerance ・Altered gene expression in liver, pgWAT, and iWAT related to lipid and glucose-metabolism |
[104] 2022 |
eWAT, epididymal white adipose tissue; pgWAT, perigonadal white adipose tissue; iWAT, inguinal white adipose tissue.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



