
Submitted:
30 January 2025
Posted:
31 January 2025
You are already at the latest version
Abstract
Background: Growing evidence emphasizes the beneficial effect of flavonoids such as anthocyanins as key constituents in lowering cardiovascular risk, and incorporating daily anthocyanin-rich fruits into the diet significantly impacts public health. Methods: The effect of raspberry fruit bioactive polyphenols on the molecular pathways in inflammation was analyzed in activated RAW 264.7 macrophages, and protective potential against endothelial dysfunction was evaluated using TNF-α-induced human umbilical vein endothelial cells (HUVECs). Results: The results have shown that raspberry fruit (RBF) extract, as well as anthocyanin and polyphenol fractions, exhibited a potent anti-inflammatory effect in macrophage cell culture by inhibition of LPS-elicited induction of pro-inflammatory gene expressions such as IL-6, IL-1β, TNF-α, and NF-κB. Moreover, RBF and both fractions have shown protective effect on endothelial functions by decreasing the expression of several inflammation-related genes and adhesion molecules such as IL-6, IL-1b, VCAM-1, ICAM-1, and SELE in TNF-α-induced HUVECs. Conclusions: Consumption of raspberry fruit and/or polyphenol-rich extract can help prevent the development of early atherosclerosis. The underlying mechanisms are related to improved inflammation status and vascular endothelial function. Due to raspberries' strong anti-inflammatory properties, incorporating them into the daily diet may significantly lower the risk of inflammation-related non-communicable diseases.

Keywords:
raspberry
; anthocyanins
; polyphenols
; HUVECs
; RAW 264.7
; inflammation
; anti-inflammatory
; cytokine
; chemokine
1. Introduction
Cardiovascular disease (CVD) is responsible for nearly half of all non-communicable disease deaths worldwide. The causes of CVD are well-known and include hypertension, high cholesterol, and age. However, CVD is now recognized as chronic inflammation [1]. Atherosclerosis, the primary cause of CVD, is a chronic inflammatory condition with immune-competent cells producing pro-inflammatory cytokines. Macrophages, which may be tissue-resident or derived from monocytes, play a key role in the progression and regression of atherosclerosis [1,2]. Vascular macrophages impact and direct atherosclerosis by interacting with all the cell types in the vessel wall. Pro-inflammatory macrophages are commonly found in progressing plaques with active inflammation [1,2]. The healthy endothelial monolayer lining the vascular lumen regulates cell adhesion and vascular tone and maintains vascular homeostasis through anticoagulant, antithrombotic, and anti-inflammatory activity [3]. During atherosclerosis, the homeostatic properties of the endothelium are lost due to the release of inflammatory cytokines by tissue-resident macrophages such as TNF-α and IL-1β, which induce the rapid expression of extracellular adhesion molecules such as E- and P-selectin, an intercellular adhesion molecule 1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1) [4]. Although the accumulation of macrophages and their interactions with other cells in the vessel wall directly cause atherosclerosis, they are also necessary for the stabilization and regression of plaques [5].
Standard therapy for treating atherosclerosis is based on lipid-lowering agents reducing the high circulating cholesterol level and artery inflammation. Due to the side effects of these agents and the disease significance even when cholesterol is low, there is a need for widely available, safe, and low-cost anti-inflammatory therapies that directly target inflammation [6]. Clinical trials proved the efficacy of targeting vascular macrophages and inflammation as a therapy for treating CVD [6,7,8]. Data showed that an improved diet could prevent over half of all cardiovascular events. Diets rich in fruits are the third most important factor for reducing global rates of non-communicable diseases [9]. Growing evidence emphasizes the beneficial effect of flavonoids such as anthocyanins as key constituents in lowering CVD risk, and incorporating daily anthocyanin-rich fruit into the diet significantly impacts public health [10]. The best dietary sources of anthocyanin’s are berries, including blueberries, blackberries, raspberries, and strawberries. For instance, regular consumption of red raspberries decreases the risk of chronic diseases [11]. Their daily intake prevents the development of early atherosclerosis by improving antioxidant status and serum lipid profiles [12], eliminating vascular endothelial dysfunction [13], and showing anti-inflammatory effects [14,15].
This study aimed to evaluate the ability of red raspberry fruit (RBF) extract and RBF anthocyanin (RBF-ACN) and non-anthocyanin polyphenol (RBF-PP) fractions to prevent and treat endothelial dysfunction imitated in the in vitro models. The effect of raspberry fruit bioactive polyphenols on the molecular pathways in inflammation was analyzed in activated RAW 264.7 macrophages. Protective potential against endothelial dysfunction was evaluated using TNF-α-induced human umbilical vein endothelial cells (HUVECs). Several crucial pro-inflammatory and anti-inflammatory cytokines were analyzed in inflamed cell culture models upon raspberry fruit polyphenols treatment.
2. Results
2.1. Polyphenol Composition of the Raspberry Fruit Extract and Anthocyanin and Non-Anthocyanin Fractions
The polyphenolic compounds present in freeze-dried RBF extract, and in RBF antho-cyanin (RBF-ACN) and RBF non-anthocyanin (RBF-PP) fractions are shown in Table 1. The compounds identified in RBF extract were classified as anthocyanins (peaks 1–4), flavanols (peaks 5–7), hydroxycinnamic acid derivatives (peaks 8–11), and flavonols (peaks 12–19). The HPLC-DAD chromatograms acquired from the extract and both frac-tions at the four wavelengths are shown in Figures S1–S3. The anthocyanin derivatives were assessed at 5.7 ± 0.39 mg/g, and the most abundant anthocyanins were cya-nidin-3-O-sophoroside, cyanidin-3-O-glucoside, cyanidin-3-O-glucosyl-rutinoside, and cyanidin-3-O-rutinoside, contributing an average of 53.9 %, 24.7 %, 12.5%, and 8.9 % to the total anthocyanin content, respectively. Procyanidin B, catechin, and epicatechin have been identified in the group of flavan-3-ols. The level of flavanols was calculated at 1.00 ± 0.09 mg/g. In the RBF extract, free ellagic acid, its sugar conjugates, three quercetin, and one kaempferol glycoside were found only in low concentrations. The most abundant among quercetin glycosides was quercetin-3-O-galactoside, with an average content of 0.14 ± 0.01 mg/g (Table 1). RBF-ACN fraction consisted of four main anthocyanin com-pounds, which including cyanidin-3-O-sophoroside (53.1 %), cya-nidin-3-O-glucosyl-rutinoside (13.0%), cyanidin-3-O-glucoside (24.0%) and cya-nidin-3-O-rutinoside (9.9 %) (Table 1).
The RBF-PP fraction included polyphenolic compounds belonging to three groups: flavan-3-ols, hydroxycinnamic acid derivatives, and flavonols, which accounted for 75.7 %, 17.3 %, and 7.0 %, respectively. Moreover, in the RBF-PP fraction, anthocyanin compounds’ residue (2.9 %) such as cyanidin-3-O-sophoroside were detected. In the RBF-PP fraction, polyphenols quantified in a significant amount (>5%) were: procyanidin B, catechin, epicatechin, caffeoyl hexoside, p-coumaryl hexoside isomer 1, and quercetin-3-O-galactoside (Table 1).
2.2. Anti-Inflammatory Effects of Raspberry Fruit Extract
The cytotoxic effects of RBF extract on RAW 264.7 macrophages were evaluated using the MTT assay to identify non-cytotoxic doses for subsequent anti-inflammatory experiments. As shown in Figure 1A, RBF extract concentrations ranging from 0.01 to 1 mg/ml did not affect the viability of LPS-activated macrophages, indicating that the observed anti-inflammatory effects were not due to cytotoxicity of the extract. In these experiments, macrophages were treated with LPS to stimulate an immune response and promote the release of inflammatory cytokines, as depicted in Figure 1B.
The increase in cytokine production induced by LPS was reduced when macrophages were treated with RBF extract and budesonide. At all tested non-cytotoxic doses, the RBF extract significantly inhibited the expression of the genes IL-6, IL-1β, COX-2, TNF-α, and NF-κB (Figure 1B). At the highest concentration of 1 mg/ml, the extract decreased the expression of these genes by 86%, 93%, 78%, 74%, and 99%, respectively (p < 0.001). The inhibitory effects of the 1 mg/ml RBF extract were more potent than those of budesonide (Figure 1B). Additionally, the RBF extract reduced the enhanced secretion of IL-6 protein in LPS-activated macrophages by 59% and 81% after treatment with 0.1 mg/ml and 1 mg/ml, respectively (p < 0.001) (Figure 1C). In contrast, there was no statistically significant effect of the RBF extract on MCP-1 gene expression (Figure 1B) or MCP-1 protein secretion (Figure 1C). The extract also had a moderate impact on the mRNA level of IL-10, with an increase in IL-10 expression of 25% (p < 0.01) observed only when LPS-induced macrophages were treated with the extract at the maximum dose of 1 mg/ml (Figure 1B).
2.3. Anti-Inflammatory Effects of Anthocyanin and Non-Anthocyanin Polyphenol Fractions of Raspberry Fruit Extract
In this study, we investigated the anti-inflammatory potential of two fractions derived from raspberry fruit extract: the anthocyanin (RBF-ACN) and non-anthocyanin polyphenol (RBF-PP) fractions, using LPS-activated macrophages. We first identified non-cytotoxic doses of the analyzed fractions suitable for anti-inflammatory experiments using an MTT test. As shown in Figure 2A, neither fraction affected macrophage viability at concentrations ranging from 0.1 µg/ml to 10 µg/ml. Real-time PCR analysis demonstrated the significant suppressive effects of both the RBF-PP and RBF-ACN fractions, revealed by a dose-dependent decrease in the expression of pro-inflammatory markers: IL-6, IL-1β, TNF-α, and NF-κB (Figure 2B). Notably, the RBF-PP fraction exhibited a slightly stronger effect. Depending on the dosage, the RBF-PP fraction reduced the expression of inflammatory mediators as follows: IL-6 (↓ 42%-69%), IL-1β (↓ 41%-72%), TNF-α (↓ 20%-43%), and NF-κB (↓ 38%-56%). The impact of the RBF-ACN fraction on IL-6 (↓18%-35%), IL-1β (↓ 16%-61%), and NF-κB (↓ 28%-56%) expression was comparable to that of the RBF-PP fraction. A statistically significant effect on COX-2 expression was observed only when macrophages were treated with the RBF-PP fraction at a concentration of 10 µg/ml, resulting in an 18% decrease (p < 0.05). This effect was similar to that obtained with budesonide. Both fractions, at the highest dose of 10 µg/ml, upregulated IL-10 expression significantly: the RBF-PP fraction increased IL-10 expression by 138% (p < 0.001), while the RBF-ACN fraction resulted in a 60% increase (p < 0.01) (Figure 2B). The secretion of the IL-6 cytokine changed in the LPS-activated macrophage cultures in parallel with the altered expression levels induced by RBF-ACN and RBF-PP fractions (Figure 2C). Similar to the whole RBF extract, neither the RBF-ACN nor the RBF-PP fractions had a statistically significant effect on MCP-1 gene expression and protein secretion (Figure 2B,C).
2.4. Potential of Raspberry Fruit Extract, Anthocyanin Fraction, and Non-Anthocyanin Polyphenol Fraction in Counteracting Vascular Endothelial Dysfunction
The inflammatory cytokine TNF-α is pivotal in disrupting macrovascular and microvascular circulation. Intra-arterial TNF-α infusion in healthy volunteers showed direct evidence about TNF-α stimulated vascular dysfunction. It has been found that blocking or decreasing TNF-α function may reduce the risk of vascular complications in various diseases [18]. The markers of endothelial dysfunction are elevated plasma levels of soluble vascular cell adhesion molecule (sVCAM), soluble intercellular adhesion molecule (sICAM), endothelin 1, E-selectin, and markers of low-grade inflammation such as C-reactive protein (CRP), IL-1β, IL-6, and TNF-α [18].
In this study, we investigated the ability of RBF extract, as well as the RBF-ACN and RBF-PP fractions, to counteract endothelial inflammation induced in HUVECs by TNF-α. TNF-α significantly stimulated the expression of several inflammation-related cytokines and adhesion molecules in HUVECs, including IL-6, IL-1β, VCAM-1, ICAM-1, and SELE (Figure 3 and Figure 4). In comparison to control cells, the RBF extract, along with the RBF-PP and RBF-ACN fractions, reduced the TNF-α-induced increase in the expression of IL-6, IL-1β, VCAM-1, ICAM-1, and SELE in a dose-dependent manner (Figure 3 and Figure 4).
After incubating HUVECs with RBF extract at concentrations of 0.1 µg/ml and 1 µg/ml, we observed a statistically significant reduction in the mRNA expression levels of IL-6 and IL-1β. Specifically, IL-6 mRNA expression decreased by 67% and 77% (p < 0.001), while IL-1β expression decreased by 22% (p < 0.05) and 48% (p < 0.001). Additionally, the effects of RBF extract on the expression of VCAM-1, ICAM-1, and SELE were significant at the 1 mg/ml concentration. VCAM-1 expression decreased by 58% (p < 0.001), ICAM-1 by 39% (p < 0.001), and SELE by 37% (p < 0.001) (Figure 3).
The RBF-ACN and RBF-PP fractions exhibited the most potent inhibitory effects on the mRNA expression of IL-6, IL-1β, VCAM-1, ICAM-1, and SELE at doses of 1 µg/ml and 10 µg/ml. Notably, the RBF-ACN fraction demonstrated a slightly stronger impact than the RBF-PP fraction. Specifically, the RBF-ACN fraction reduced the expression of IL-6 by 28% and 54%, IL-1β by 32% and 60%, and VCAM-1 by 40% and 60%. The RBF-PP fraction decreased the number of IL-6 transcripts by 33% and 39%, IL-1β by 23% and 32%, and VCAM-1 by 21% and 39% (Figure 4). Significant effects on ICAM-1 and SELE expression were observed only at the higher dose of 10 µg/ml for both fractions. The RBF-ACN and RBF-PP fractions similarly reduced the mRNA expression of ICAM-1 and SELE, decreasing ICAM-1 expression by 25% and 28% and SELE expression by 24% and 25%, respectively (Figure 4).
3. Discussion
Chronic inflammation has an extensive role in the development and progression of several disorders including type 2 diabetes mellitus, CVD, as well as atherosclerosis and neurodegenerative diseases. Therefore, the potential to target inflammation is an appealing strategy to combat inflammatory-related diseases [19]. Atherosclerosis is caused by lipid metabolism, endothelial dysfunction, and inflammatory cell invasion. Macrophages are a key to atherosclerotic disease progression and regression [1]. They release pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α in response to various inflammatory stimuli. However, macrophages are also necessary for the stabilization and regression of plaques [1]. Plaque macrophages promote tissue repair and angiogenesis by releasing anti-inflammatory cytokines such as IL-10 and TGF-β [1,5]. Reducing macrophage inflammatory response has been associated with anti-atherogenic properties. Many potential treatments for CVD target pro-inflammatory cytokines released by macrophages. In vivo studies have found that inhibiting of macrophage IL-1β expression significantly reduced cardiovascular events [20].
Evidence from prospective studies and randomized controlled trials suggests that more than half of all cardiovascular events could be prevented by improving diet [10]. Research, both in vivo and in vitro, indicates that polyphenols found in berry fruits may help reduce inflammation, and consuming foods rich in anthocyanins is associated with a lower risk for chronic, non-communicable diseases [19]. Dietary anthocyanins and other phenolic compounds have the potential to influence inflammatory status by modulating the expression of pro-inflammatory and anti-inflammatory cytokines [21]. Common sources of anthocyanins include blueberries, blackberries, elderberries, aronia fruit, and raspberries [19].
This study evaluated the anti-inflammatory effects of whole RBF extract and its fractions, RBF-ACN and RBF-PP, by measuring their impact on pro-inflammatory and anti-inflammatory gene expression in LPS-activated RAW 264.7 macrophages. The results showed that the mRNA expression levels of IL-6, IL-1β, COX-2, TNF-α, and NF-κB were significantly reduced by the whole RBF extract, demonstrating a more potent effect than budesonide. Both fractions, RBF-ACN and RBF-PP, also inhibited the expression of pro-inflammatory cytokines, but their effects were not as strong as those observed with the whole extract. These findings suggest synergistic interactions among multiple components that may significantly enhance the anti-inflammatory potential. Additionally, the RBF extract, RBF-ACN, and RBF-PP fractions at the highest doses increased IL-10 mRNA expression in LPS-activated macrophages.
Several previous studies have reported that berry anthocyanins can modulate inflammatory responses in cell cultures and animal models. Li et al. investigated the anti-inflammatory activities of crude extracts, anthocyanin-rich fractions, and des-anthocyanin fractions from seven berries in LPS-activated RAW 264.7 macrophages. They discovered that the anthocyanin fractions from red raspberries were the most effective in inhibiting the expression of iNOS and COX-2. Additionally raspberry anthocyanin fractions significantly reduced the mRNA levels of IL-1β and IL-6. These fractions also suppressed the NF-κB and AP-1 signaling pathways in RAW 264.7 cells in a concentration-dependent manner [22].
Previous research has shown that the anthocyanin fraction from blueberries, blackberries, and blackcurrants at 10 µg/ml or 20 µg/ml demonstrates significant anti-inflammatory effects in LPS-activated macrophages. These effects were achieved by downregulating the expression of IL-1β and TNF-α, primarily through inhibiting the nuclear transcription factor-κB (NF-κB) pathway [23]. Reported studies have shown that the exposure of LPS-stimulated human THP-1 macrophages to cyanidin-3-O- β-glucoside significantly reduces the expression and secretion of pro-inflammatory cytokines like TNF-α and IL-6 [24]. Furthermore, a dietary intervention involving red raspberry fruit (5.3% freeze-dried raspberry) in obese diabetic mice over 8 weeks decreased IL-6 plasma concentration [25]. In a study conducted by Mykkanen et al., it was demonstrated that incorporating Nordic wild blueberries (10% by weight) into a high-fat diet (comprising 45% fat) for 12 weeks effectively reduced serum concentrations of key pro-inflammatory markers, including IL-1, IL-2, and TNF-α, in C57BL/6 mice [26].
In a randomized controlled trial, daily red raspberry supplementation for 4 weeks was associated with significantly lower biomarkers of inflammation, particularly IL-6 and TNF-α in in adults with established type 2 diabetes mellitus [27]. In metabolic syndrome subjects, 4 weeks of anthocyanin-rich berry consumption significantly down-regulated the expression of NF-κB dependent genes, including TNF-α, IL-6, IL-1A, PCAM-1, and COX-2. The results suggested that berry supplementation can mitigate features of metabolic syndrome and related cardiovascular risk factors, and these benefits may be due to the inhibition of NF-κB dependent gene expression [28].
NF-κB is a crucial regulator of many genes that play a role in inflammation. Activation of NF-κB by kinases stimulates the transcription of genes encoding pro-inflammatory chemokines and cytokines. Therefore, diminishing NF-κB activation is being explored as a potential strategy for preventing chronic inflammatory diseases [29]. Many studies have indicated that downregulating NF-κB signaling is the primary mechanism through which berries and their bioactive compounds exert their anti-inflammatory effects [28,29,30]. Our research showed that the RBF extract and the RBF-ACN and RBF-PP fractions significantly suppressed the expression of LPS-induced NF-κB. This suggests that the anti-inflammatory effects of raspberry phytochemicals are achieved by inhibiting NF-κB activation.
Inflammation is crucial in causing vascular endothelial dysfunction by activating transcription factors, such as NF-κB. Activation of the NF-κB up-regulates the expression of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6), chemokines, and adhesion molecules (ICAMs and VCAM-1) [24]. Vascular macrophage-related pro-inflammatory cytokines induce inflammation-mediated increases in vascular permeability [31]. Our data demonstrated that RBF extract, as well as RBF-ACN and RBF-PP fractions, had a protective effect on endothelial functions by reducing the expression of several inflammation-related genes and adhesion molecules, including IL-6, IL-1β, VCAM-1, ICAM-1, and SELE in TNF-α-induced HUVECs. In vitro studies and in vivo animal models testing the effects of raspberry polyphenolic components or raspberry fruit extracts have shown their efficacy in improving endothelial function [11].
In cell culture, purified anthocyanin mixture inhibited IL-6 and IL-1β-induced CRP production in the HepG2 cell line and LPS-induced VCAM-1 secretion in the porcine iliac artery endothelial cell line. Moreover, the reduction of inflammatory cytokines associated with the anthocyanin mixture was more potent when compared with the effects of delphinidin-3-O-β-glucoside and cyanidin-3-O-β-glucoside separately, indicating additive or synergistic effects of the anthocyanin mixture in mediating anti-inflammatory responses [32]. Pretreatment of human endothelial cells with cyanidin-3-O-glucoside significantly attenuated TNF-α-induced expression of adhesion molecules ICAM-1, VCAM-1, and E-selectin, and inhibited NF-κB activation [33]. Polyphenol sea buckthorn berry extract significantly decreased the level of serum TNF-α and IL-6, as well as alleviated vascular impairment by decreasing eNOS and ICAM-1 mRNA expression and protein secretion in the aortas of rats with hyperlipidemia [34]. In female patients with metabolic syndrome, consuming 4 cups of freeze-dried strawberries daily for 8 weeks resulted in lower levels of LDL-cholesterol and VCAM-1 [35]. In adults with hypercholesterolemia, a purified anthocyanin mixture significantly decreased serum CRP, sVCAM-1, and plasma IL-1β compared to the placebo [32]. In patients with metabolic syndrome, black raspberry consumption over 12 weeks significantly decreased serum total cholesterol levels and inflammatory cytokines such as IL-6, TNF-α, CRP, sICAM-1, and sVCAM-1, thereby improving vascular endothelial function [36].
In summary, the results have shown that RBF extract, as well as both fractions analyzed, exhibited a potent anti-inflammatory effect in macrophage cell culture by inhibition of LPS-elicited induction of pro-inflammatory gene expressions such as IL-6, IL-1β, TNF-α, and NF-κB indicating a protective effect of raspberries to prevent chronic inflammation. Moreover, RBF extract and RBF-ACN and RBF-PP fractions have shown a protective effect on endothelial functions by decreasing the expression of several inflammation-related genes and adhesion molecules such as IL-6, IL-1b, VCAM-1, ICAM-1, and SELE in TNF-α-induced HUVECs. These results suggest that consumption of raspberry fruit can help to prevent the development of early atherosclerosis, with the underlying mechanisms related to improved inflammation status and vascular endothelial function. These data indicate that raspberry fruit supplementation may have a role in preventing or treating chronic inflammatory diseases by inhibiting pro-inflammatory chemokines, cytokines, and inflammatory mediators. Thus, raspberry may be an effective dietary agent for treating inflammatory and cardiovascular complications.
4. Materials and Methods
4.1. Reagents
Dulbecco’s Modified Eagle’s Medium (DMEM), lipopolysaccharides (LPS) from E. coli O127, and TNF-α were purchased from Merck (Merck Life Science, Poland). Fetal bovine serum (FBS) was obtained from Gibco BRL (Grand Island, NY, USA). The total RNA was isolated using the Tri Reagent (Merck Life Science, Poland). Template cDNA synthesis was performed using the Transcriptor High Fidelity cDNA Synthesis Kit (Roche Diagnostics, Poland). PCR was performed using SYBR® Select Master Mix (Life Technologies, Carlsbad, CA, USA). IL-6 and MCP-1 cytokines were determined using ELISA kits from R & D Systems, Inc (Minneapolis, MN, USA). All other reagents were purchased from Merck Life Science unless otherwise noted.
4.2. Preparation of Raspberry Fruit Extract and Anthocyanin and Non-Anthocyanin Polyphenol Fractions
The mature raspberry fruit (Rubus idaeus L.) Polka cv were obtained from Bio Berry Poland (Warsaw Poland) and subjected to freeze-drying as described previously [16]. The freeze-dried raspberry fruit were dissolved in a complete cell culture medium, and the pH was adjusted to 7.4. The suspension was centrifuged (3000 g, 10 min.), filtered through a 0.22 μm membrane, and added to the cell culture at different concentrations, ranging from 0.01 mg to 1 mg of freeze-dried powder/ml of culture medium. RBF-ACN and RBF-PP fractions were prepared according to the previously described procedure [17]. Freeze-dried fractions were dissolved in a complete medium and added to the cell cultures at 0.1, 1, and 10 µg/ml concentrations.
4.3. Polyphenol Identification and Quantification
RBF extract and RBF-ACN and RBF-PP fractions were analyzed by the HPLC-DAD-ESI-MS method on an Agilent 1200 series HPLC system (Agilent Technologies, Inc., Santa Clara, CA, USA) with separation conditions described previously [17].
4.4. Cell Cultures and Anti-Inflammatory Experiment Procedure
Human umbilical vein endothelial cells (HUVEC, ATCC CRL-1730™) were obtained from the American Type Culture Collection (ATCC). HUVECs were cultured in F-12K medium (ATCC) supplemented with 10% FBS, endothelial cell growth supplement from bovine neural tissue (30 µg/mL), and heparin (100 µg/mL). HUVECs were seeded at 6 × 103 cells/cm2 density onto 24-well plates coated with rat tail collagen. After 24 h, HUVECs were exposed for 3 h to RBF extract and RBF-ACN and RBF-PP fractions, and subsequently treated with TNF-α (10 ng/mL) for an additional 3 h to induce inflammation.
RAW 264.7 macrophages were obtained from the European Collection of Cell Cultures (ECACC, 91062702). They were cultured in DMEM supplemented with 10% FBS. For experiments, RAW 264.7 cells were seeded onto 24-well plates at inoculum of 5×105 cells/cm2. After 24 h the cells were treated with RBF extract and RBF-ACN and RBF-PP fractions for 1 h before stimulation with LPS at 1 μg/ml for an additional 3 h.
For every experiment, one positive control (cells treated with budesonide at 1 μM, a glucocorticoid steroid with strong anti-inflammatory potential) and one negative control (cells treated with vehicle) were included.
HUVECs and RAW 264.7 cells were used for total RNA isolation and gene expression analysis, and culture medium was collected for the determination of IL-6 and MCP-1 protein secretion.
4.5. Cell Viability Assay
The viability of LPS-activated RAW 264.7 macrophages, treated and non-treated with RBF extract and RBF-ACN and RBF-PP fractions, were analyzed using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay following the procedure described previously [16].
4.6. RNA Extraction and Real-Time PCR Analysis
RAW 264.7 macrophages and HUVECs were treated with TRI-Reagent for total RNA isolation. First-strand cDNA synthesis was performed with 1 µg of total RNA using a Transcriptor High Fidelity cDNA Synthesis Kit based on the manufacturer’s instruction. The quantification of gene expression was conducted using a real-time PCR system (CFX96 Touch Real-Time PCR Detection System, Bio-Rad Laboratories, Inc.). PCR mixture in a final volume of 25 µL included a cDNA sample (1 µL), specific forward and reverse primers (5 µM/1 µL), and SYBR® Select Master Mix (12.5 µL). Primer sequences used in this experiment are shown in Table S1. The PCR cycling conditions included an initial denaturation at 94 OC for 10 min, followed by 40 PCR cycles: 40 s at 95 ℃, 30 s at 59 ℃, and 30 s at 72 ℃. The relative gene expression was calculated using the 2-ΔΔCT method. Transcript levels were normalized to β-actin for RAW 264.7 macrophages and to GAPDH for HUVECs. Relative mRNA expression was expressed as fold change compared with control (untreated) cells. All reactions were performer in triplicate.
4.7. Determination of IL-6 and MCP-1 Production
IL-6 and MCP-1 protein concentrations were measured with ELISA kits following the manufacturer’s protocols. Quantitation was performer using the calibration of standards. Each standard and sample was assayed in triplicate.
4.8. Statistical Analysis
Statistical analysis was performed using the STATISTICA version 13.3 software (Statsoft, Inc., Tulsa, OK, USA). One-way analysis of variance (ANOVA) and Tukey’s post hoc test were applied to estimate the differences between multiple groups’ mean values. Levene’s test verified the equality of variances assumption. Statistical significance was set at p < 0.05.
5. Conclusions
Chronic inflammation plays a central role in the development and progression of several disorders, amongst others, atherosclerosis, autoimmune diseases, neurodegenerative diseases, and cardiovascular complications; therefore, the development of anti-inflammatory drugs is a significant challenge. Diet is an essential factor that can decrease the risk of inflammation-related diseases by the ability of certain foods and their bioactive compounds to reverse or prevent the progression of the pathogenic inflammatory processes that underlie these diseases. Our studies provide evidence that incorporating raspberry fruit for their significant anti-inflammatory effect into the daily diet could considerably reduce the risk of inflammation-related non-communicable diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: HPLC-DAD chromatograms of raspberry fruit (RBF) extract recorded at 280 (A), 520 (B), 325 (C), and 355 nm (D) for optimal detection of flavan-3-ols, anthocyanins, hydroxycinnamic acid derivatives, and flavonols, respectively (peak numbers correspond to those in Table 1); Figure S2: HPLC-DAD chromatograms of anthocyanin raspberry fruit (RBF-ACN) extract recorded at 280 (A), 520 (B), 325 (C), and 355 nm (D) for optimal detection of flavan-3-ols, anthocyanins, hydroxycinnamic acid derivatives, and flavonols, respectively (peak numbers correspond to those in Table 1); Figure S3: HPLC-DAD chromatograms of raspberry fruit polyphenol (RBF-PP) extract recorded at 280 (A), 520 (B), 325 (C), and 355 nm (D) for optimal detection of flavan-3-ols, anthocyanins, hydroxycinnamic acid derivatives, and flavonols, respectively (peak numbers correspond to those in Table 1); Table S1: The primers sequence used for real-time PCR.
Author Contributions
Conceptualization, K.K. and A.O.; methodology, K.K., A.O. and R.D.; validation, K.K and A.O..; formal analysis, A.O.; investigation, K.K., R.D., and A.O; writing—original draft preparation, K.K.; writing—review and editing, A.O.; visualization, K.K. and A.O.; project administration, A.O.; funding acquisition, A.O.
Funding
This research was funded by NATIONAL SCIENCE CENTRE, POLAND, grant number 2015/19/B/NZ9/01054.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Susser, L.I.; Rayner, K.J. Through the layers: how macrophages drive atherosclerosis across the vessel wall. J. Clin. Invest. 2022, 132, e157011. [CrossRef]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: a dynamic balance. Nat. Rev. Immunol. 2013, 13, 709-721. [CrossRef]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O., Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science. 2007, 317, 666-670. [CrossRef]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774.
- Rahman, K.; Vengrenyuk, Y.; Ramsey, S.A.; Vila, N.R.; Girgis, M.; Liu, J.; Gusarova, V.; Gromada, J.; Weinstock, A.; Moore, K.J.; Loke, P.; Fisher, E.A. Inflammatory Ly6Chi monocytes and their conversion to M2 macrophages drive atherosclerosis regression. J. Clin. Invest. 2017, 127, 2904-2915.
- Libby, P.; Everett, B.M. Novel anti-atherosclerotic therapies. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 538–545.
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; An-ker, S.D.; Kastelein, J.J.P.; Cornel, J.H.; Pais, P.; Pella, D.; Genest, J.; Cifkova, R.; Lorenzatti, A.; Forster, T.; Kobalava, Z.; Vida-Simiti, L.; Flather, M.; Shimokawa, H.; Ogawa, H.; Dellborg, M.; Rossi, P.R.F.; Troquay, R.P.T.; Libby, P.; Glynn, R.J.; CAN-TOS Trial Group. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119-1131.
- Martínez, G.J.; Celermajer, D.S.; Patel, S. The NLRP3 inflammasome and the emerging role of colchicine to inhibit atherosclerosis-associated inflammation. Atherosclerosis. 2017, 269, 262–271.
- Ezzati, M.; Riboli, E. Behavioral and dietary risk factors for non-communicable diseases. N. Engl. J. Med. 2013, 369, 954-964.
- Cassidy, A. Berry anthocyanin intake and cardiovascular health. Mol. Aspects. Med. 2018, 61, 76-82. [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red Raspberries and Their Bioactive Polyphenols: Cardiometabolic and Neuronal Health Links. Adv. Nutr. 2016, 7, 44-65. [CrossRef]
- Suh, J. H.; Romain, C.; González-Barrio, R.; Cristol, J. P.; Teissèdre, P. L.; Crozier, A.; Rouanet, J. M. Raspberry juice consumption, oxidative stress and reduction of atherosclerosis risk factors in hypercholesterolemic golden Syrian hamsters. Food Funct. 2011, 2, 400–405.
- Jia, H.; Liu, J.W.; Ufur, H.; He, G.S.; Liqian, H.; Chen, P. (2011). The antihypertensive effect of ethyl acetate extract from red raspberry fruit in hypertensive rats. Pharmacogn. Mag. 2011, 7, 19–24.
- Kowalska, K.; Olejnik, A.; Zielińska-Wasielica, J.; Olkowicz, M. Raspberry (Rubus idaeus L.) fruit extract decreases oxidation markers, improves lipid metabolism and reduces adipose tissue inflammation in hypertrophied 3T3-L1 adipocytes. J. Func. Food. 2019, 62, 103568.
- Jean-Gilles, D.; Li, L.; Ma, H.; Yuan, T.; Chichester, C.O. 3rd; Seeram, N.P. Anti-inflammatory effects of polyphenolic-enriched red raspberry extract in an antigen-induced arthritis rat model. J. Agric. Food. Chem. 2012, 60, 5755-5762.
- Kowalska, K.; Olejnik, A.; Rychlik, J.; Grajek, W. Cranberries (Oxycoccus quadripetalus) inhibit adipogenesis and lipogenesis in 3T3-L1 cells. Food Chem. 2014, 148, 246–252. [CrossRef]
- Kowalska, K.; Dembczyński, R.; Gołąbek, A.; Olkowicz, M.; Olejnik, A. ROS Modulating Effects of Lingonberry (Vaccinium vitis idaea L.) Polyphenols on Obese Adipocyte Hypertrophy and Vascular Endothelial Dysfunction. Nutrients. 2021, 13, 885. [CrossRef]
- Zhang, H.; Park, Y.; Wu, J.; Chen, X.; Lee, S.; Yang, J.; Dellsperger, K.C.; Zhang, C. Role of TNF-alpha in vascular dysfunction. Clin. Sci. 2009, 116, 219–230.
- Speer, H.; D'Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health-A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants. 2020, 9, 366. [CrossRef]
- Stevens, T.W;, Khalaf, F.K.; Soehnlen, S.; Hegde, P.; Storm, K.; Meenakshisundaram, C.; Dworkin, L.D.; Malhotra, D.; Haller, S.T.; Kennedy, D.J.; Dube, P. Dirty Jobs: Macrophages at the Heart of Cardiovascular Disease. Biomedicines. 2022, 10, 1579. [CrossRef]
- Vendrame, S.; Klimis-Zacas, D. Anti-inflammatory effect of anthocyanins via modulation of nuclear factor-κB and mitogen-activated protein kinase signaling cascades. Nutr. Rev. 2015, 73, 348-358. [CrossRef]
- Li, L.; Wang, L.; Wu, Z.; Yao, L.; Wu, Y.; Huang, L.; Liu, K.; Zhou, X.; Gou, D. Anthocyanin-rich fractions from red raspberries attenuate inflammation in both RAW264.7 macrophages and a mouse model of colitis. Sci. Rep. 2014, 4, 6234. [CrossRef]
- Lee, S.G.; Kim, B.; Yang, Y.; Pham, T.X.; Park, Y.K.; Manatou, J.; Koo, S.I.; Chun, O.K.; Lee, J.Y. Berry anthocyanins suppress the expression and secretion of pro-inflammatory mediators in macrophages by inhibiting nuclear translocation of NF-κB independent of NRF2-mediated mechanism. J. Nutr. Biochem. 2014, 25, 404-411.
- Zhang, Y.; Lian, F.; Zhu, Y.; Xia, M.; Wang, Q.; Ling, W.; Wang, X.D. Cyanidin-3-O-beta-glucoside inhibits LPS-induced expression of inflammatory mediators through decreasing IkappaBalpha phosphorylation in THP-1 cells. Inflamm. Res. 2010, 59, 723-730.
- Noratto, G.D.; Chew, B.P.; Atienza, L.M. Red raspberry (Rubus idaeus L.) intake decreases oxidative stress in obese diabetic (db/db) mice. Food Chem. 2017, 227, 305-314. [CrossRef]
- Mykkanen, O.T.; Huotari, A.; Herzig, K.H.; Dunlop, T.W.; Mykkanen, H.; Kirjavainen, P.V. Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS ONE 2014, 9, e114790.
- Schell, J.; Betts, N.M.; Lyons, T.J.; Basu, A. Raspberries Improve Postprandial Glucose and Acute and Chronic Inflammation in Adults with Type 2 Diabetes. Ann. Nutr. Metab. 2019, 74, 165-174.
- Aboonabi, A.; Aboonabi A. Anthocyanins reduce inflammation and improve glucose and lipid metabolism associated with inhibiting nuclear factor-kappaB activation and increasing PPAR-γ gene expression in metabolic syndrome subjects. Free Radic. Biol. Med. 2020, 150, 30-39. [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjørnsson, B. The Role of Mitogen-Activated Protein Kinase-Activated Protein Kinases (MAP-KAPKs) in Inflammation. Genes (Basel). 2013, 4, 101-133. [CrossRef]
- Land Lail, H.; Feresin, R.G.; Hicks, D.; Stone, B.; Price, E.; Wanders, D. Berries as a Treatment for Obesity-Induced Inflammation: Evidence from Preclinical Models. Nutrients. 2021, 13, 334. [CrossRef]
- Farahi, L.; Sinha, S.K.; Lusis, A.J. Roles of Macrophages in Atherogenesis. Front. Pharmacol. 2021, 12, 785220. [CrossRef]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y; Li, G.; Xiao, Y.; Liu, F.; Li, Z.; Shi, Z.; Yang, Y. Anti-inflammatory effect of purified dietary anthocyanin in adults with hypercholesterolemia: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [CrossRef]
- Speciale, A.; Canali, R.; Chirafisi, J.; Saija, A.; Virgili, F.; Cimino, F. Cyanidin-3-O-glucoside protection against TNF-a-induced endothelial dysfunction: involvement of nuclear factor-ĸB signaling. J. Agric. Food Chem. 2010, 58, 12048–12054.
- Yang, F.; Suo, Y.; Chen, D.; Tong, L. Protection against vascular endothelial dysfunction by polyphenols in sea buckthorn berries in rats with hyperlipidemia. Biosci. Trends 2016, 10, 188–196.
- Basu, A.; Fu, D.X.; Wilkinson, M.; Simmons, B.; Wu, M.; Betts, N.M.; Du, M.; Lyons, T.J. Strawberries decrease atherosclerotic markers in subjects with metabolic syndrome. Nutr. Res. 2010, 30, 462–469.
- Jeong, H.S.; Hong, S.J.; Lee, T.B.; Kwon, J.W.; Jeong, J.T.; Joo, H.J.; Park, J.H.; Ahn, C.M.; Yu, C.W.; Lim, D.S. Effects of black raspberry on lipid profiles and vascular endothelial function in patients with metabolic syndrome. Phytother. Res. 2014, 28, 1492-1498. [CrossRef]
Figure 1.
Effect of raspberry fruit extract (RBF) on the cell viability (A), inflammatory-related gene expression (B) and protein secretion (C) in LPS-stimulated RAW 264.7 macrophages. Budesonide (BD) was used as a positive control to verify anti-inflammatory effects. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences between exposed cells and negative control K(+) / 0 with significant differences ap < 0.05, bp < 0.01, cp < 0.001.
Figure 1.
Effect of raspberry fruit extract (RBF) on the cell viability (A), inflammatory-related gene expression (B) and protein secretion (C) in LPS-stimulated RAW 264.7 macrophages. Budesonide (BD) was used as a positive control to verify anti-inflammatory effects. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences between exposed cells and negative control K(+) / 0 with significant differences ap < 0.05, bp < 0.01, cp < 0.001.
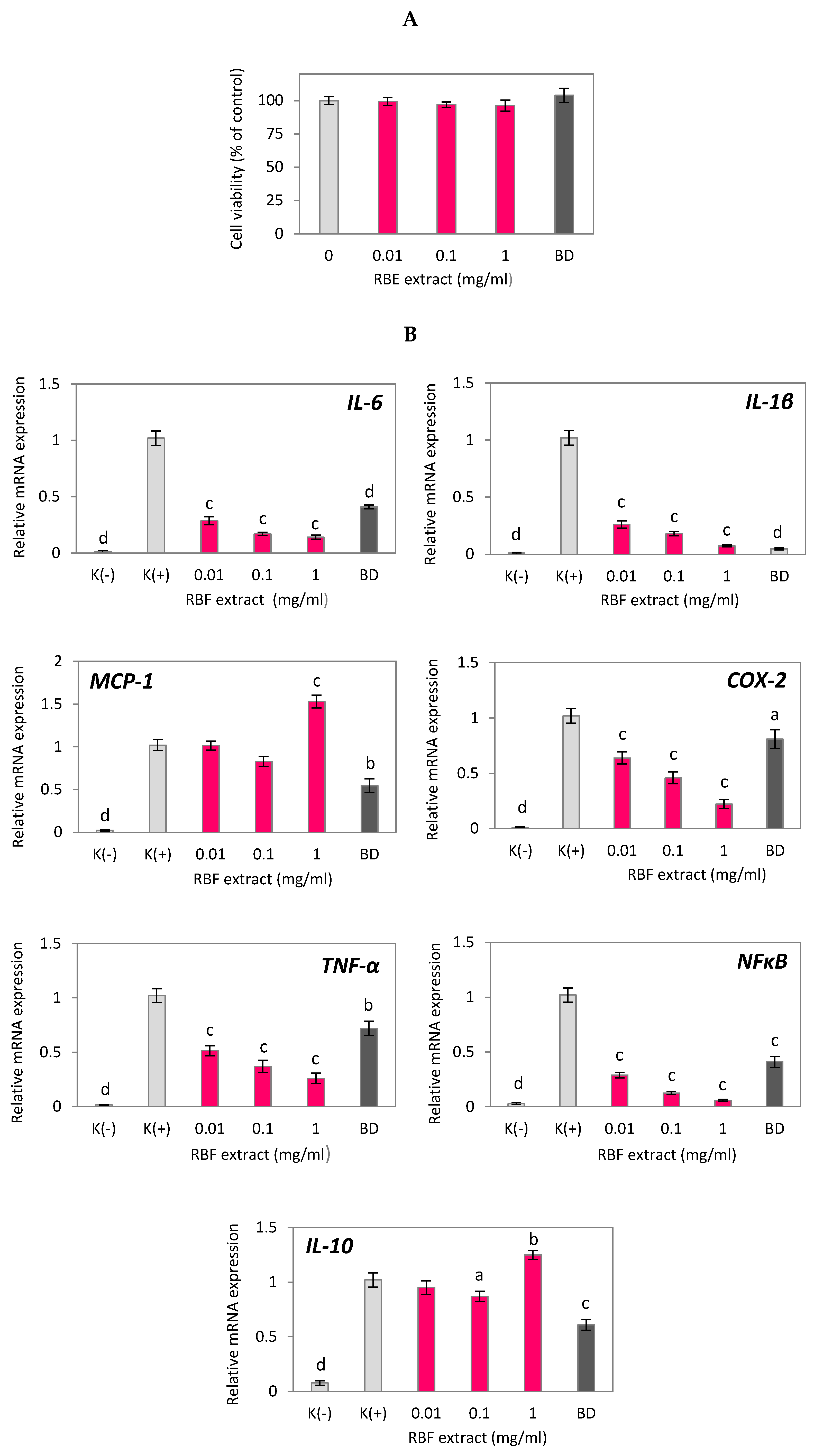
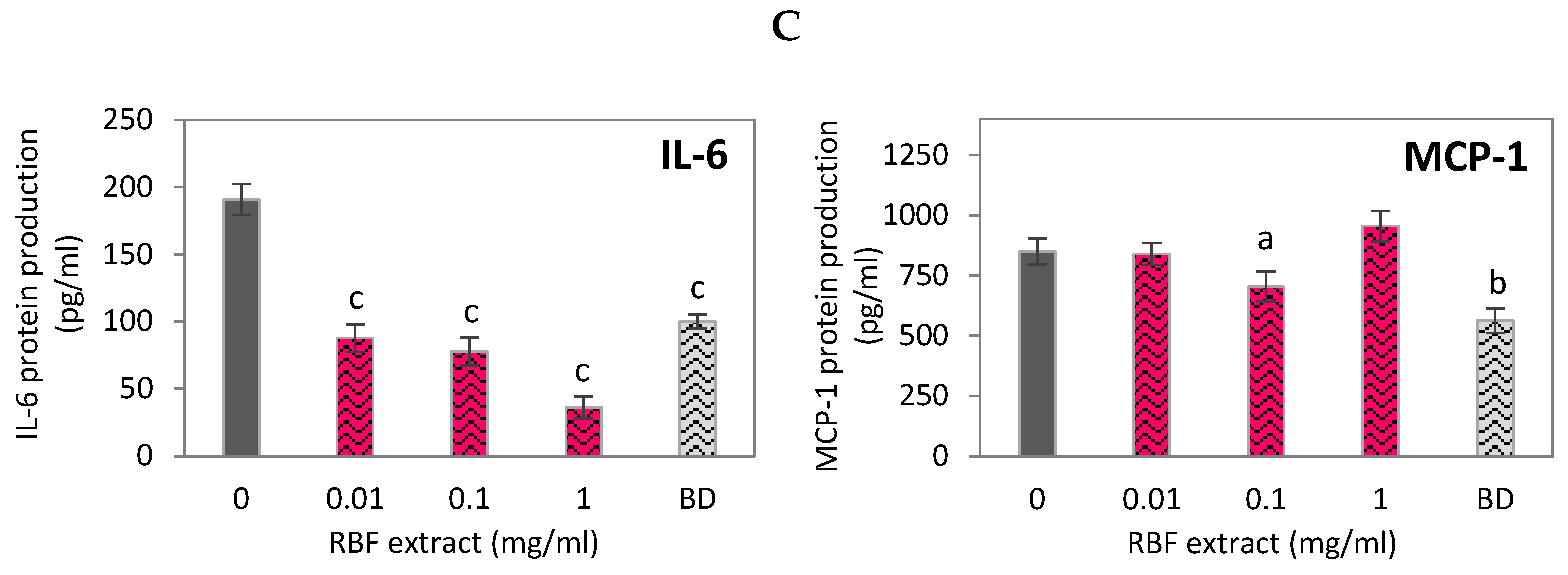
Figure 2.
Effect of raspberry fruit anthocyanin fraction (RBF-ACN) and non-anthocyanin polyphenol fraction (RBF-PP) on the cell viability (A), inflammatory-related gene expression (B) and protein secretion (C) in LPS-stimulated RAW 264.7 macrophages. Budesonide (BD) was used as a positive control to verify anti-inflammatory effects. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences between exposed cells and negative control K(+) / 0 with significant differences ap < 0.05, bp < 0.01, cp < 0.001.
Figure 2.
Effect of raspberry fruit anthocyanin fraction (RBF-ACN) and non-anthocyanin polyphenol fraction (RBF-PP) on the cell viability (A), inflammatory-related gene expression (B) and protein secretion (C) in LPS-stimulated RAW 264.7 macrophages. Budesonide (BD) was used as a positive control to verify anti-inflammatory effects. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences between exposed cells and negative control K(+) / 0 with significant differences ap < 0.05, bp < 0.01, cp < 0.001.
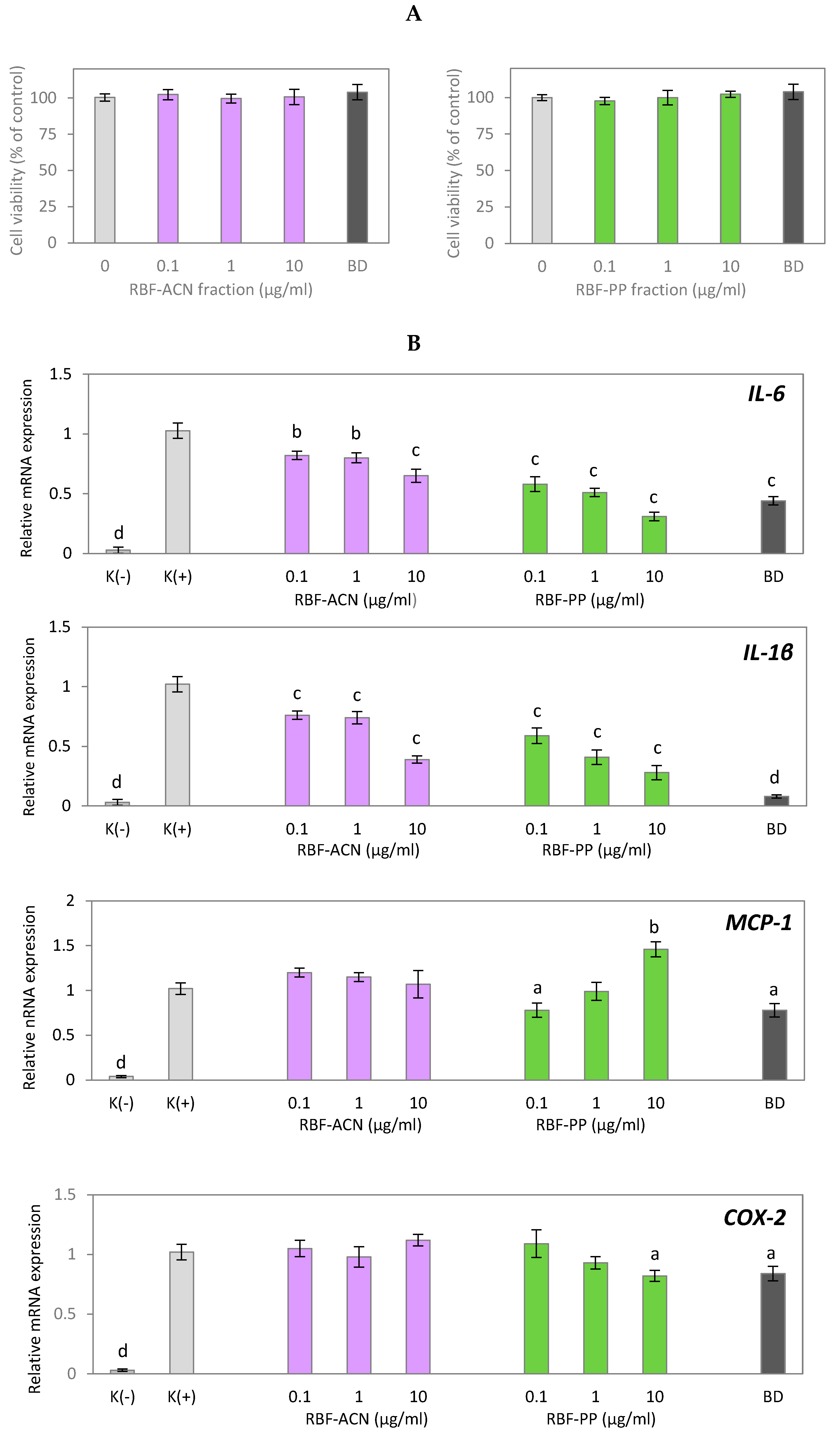
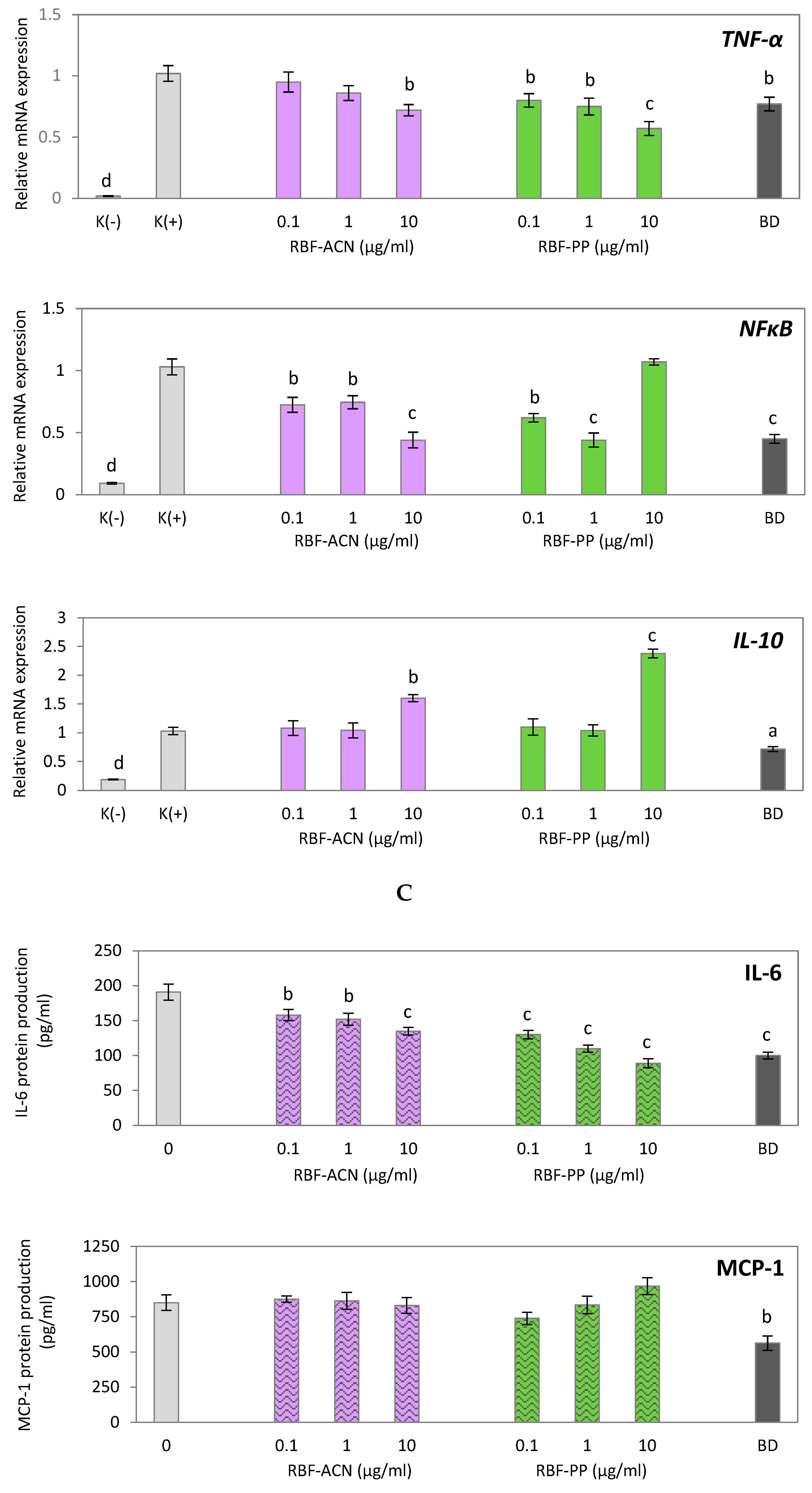
Figure 3.
Effect of raspberry fruit (RBF) extract on inflammatory-related gene expression in HUVECs treated with TNF-α. Negative control K(-) was not induced by TNF-α. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences compared to the positive control K(+) with significant differences ap < 0.05, bp < 0.01, cp < 0.001, dp < 0.0001.
Figure 3.
Effect of raspberry fruit (RBF) extract on inflammatory-related gene expression in HUVECs treated with TNF-α. Negative control K(-) was not induced by TNF-α. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences compared to the positive control K(+) with significant differences ap < 0.05, bp < 0.01, cp < 0.001, dp < 0.0001.
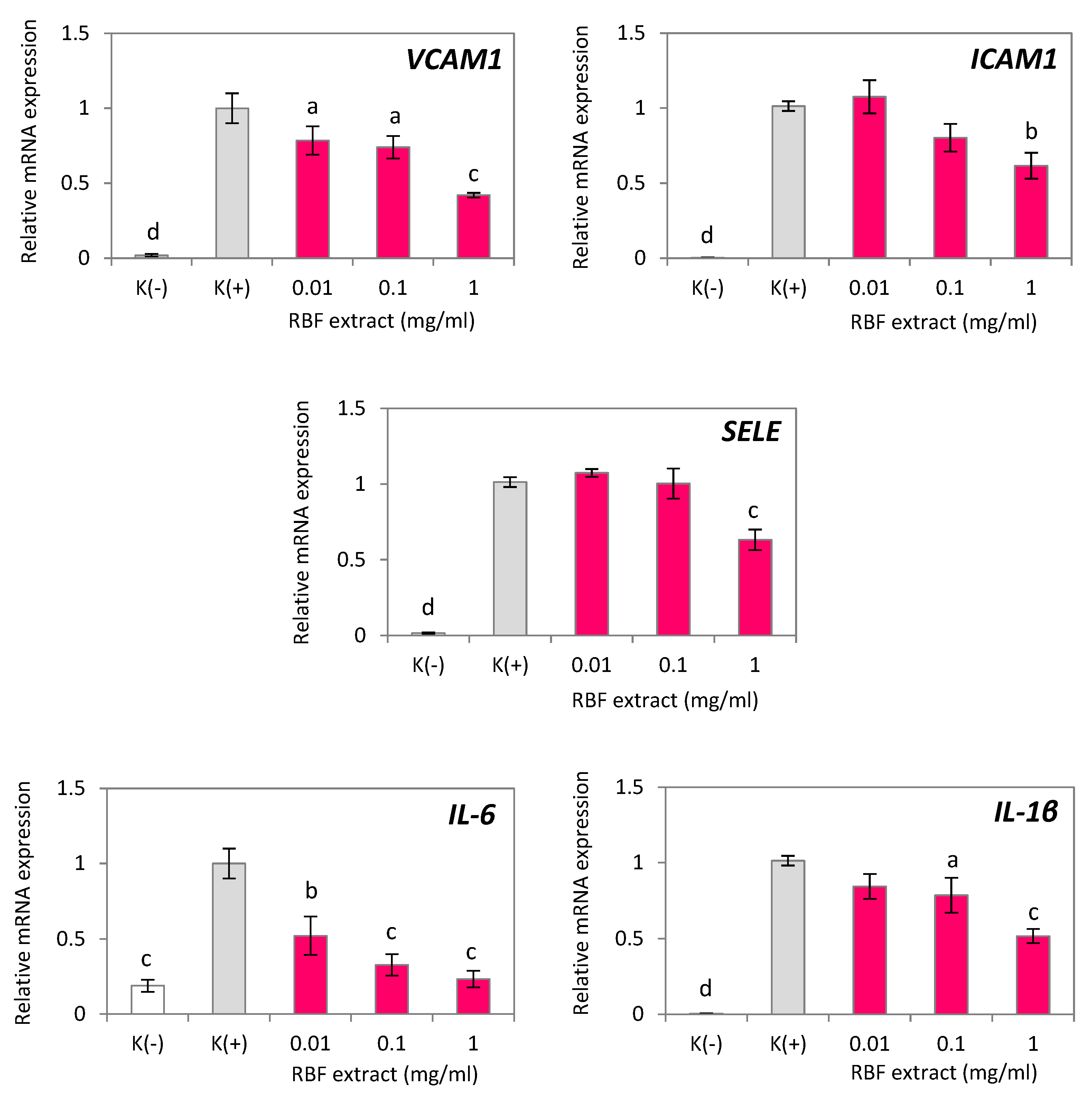
Figure 4.
Effect of anthocyanin (RBF-ACN) and non-anthocyanin polyphenol (RBF-PP) fractions on inflammatory-related gene expression in HUVECs treated with TNF-α. Negative control K(-) was not induced by TNF-α. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences compared to the positive control K(+) with significant differences ap < 0.05, bp < 0.01, cp < 0.001, dp < 0.0001.
Figure 4.
Effect of anthocyanin (RBF-ACN) and non-anthocyanin polyphenol (RBF-PP) fractions on inflammatory-related gene expression in HUVECs treated with TNF-α. Negative control K(-) was not induced by TNF-α. Data are the mean values ± SD (n = 3). Statistical analysis assessed differences compared to the positive control K(+) with significant differences ap < 0.05, bp < 0.01, cp < 0.001, dp < 0.0001.
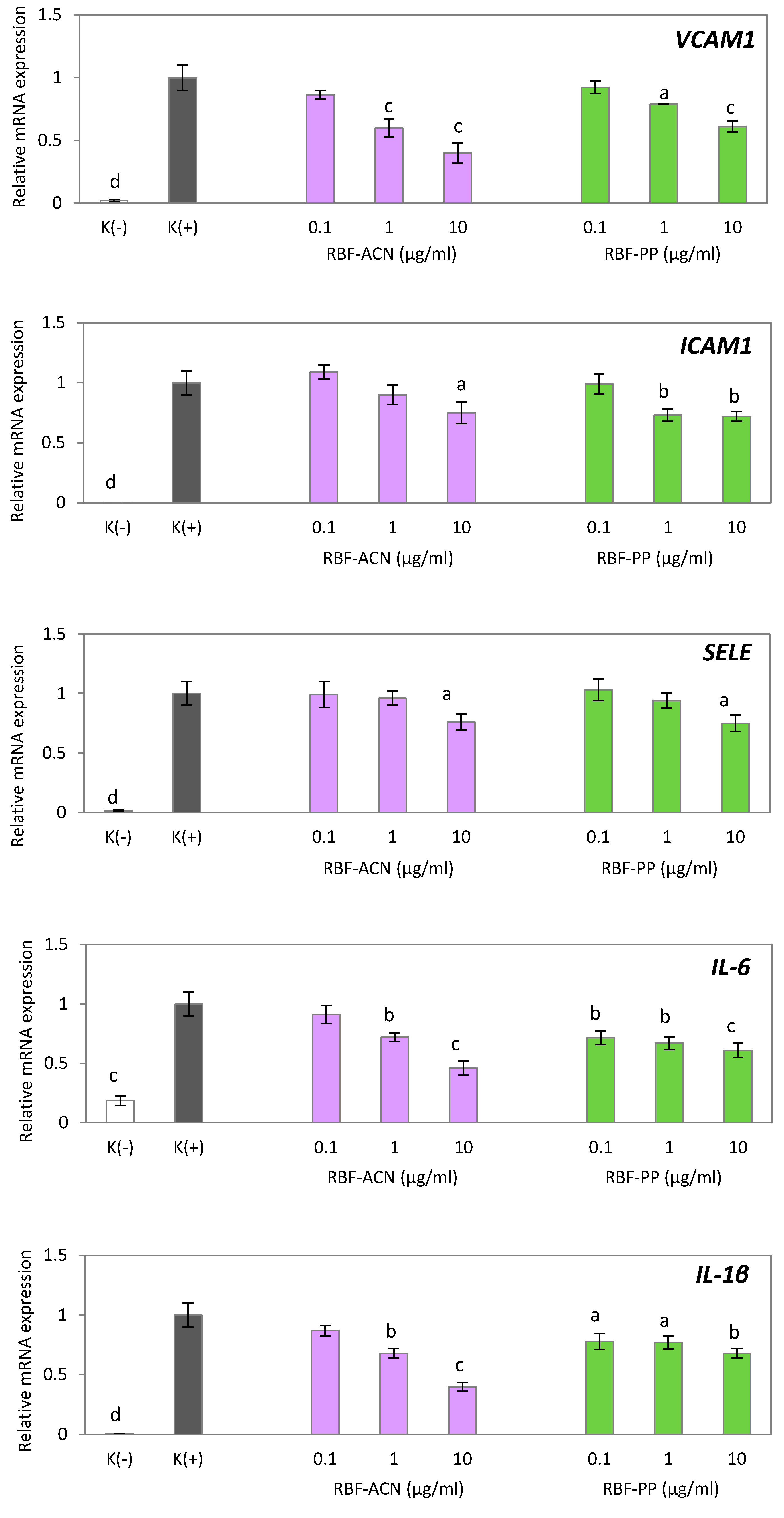

Table 1.
Identification and quantification of phenolic compounds contained in raspberry fruit (RBF), RBF anthocyanin (RBF-ACN), and RBF non-anthocyanin (RBF-PP) extracts. Quantitative data are expressed as mg/g lyophilized powder.
Table 1.
Identification and quantification of phenolic compounds contained in raspberry fruit (RBF), RBF anthocyanin (RBF-ACN), and RBF non-anthocyanin (RBF-PP) extracts. Quantitative data are expressed as mg/g lyophilized powder.
| Peak No. |
RT (min) |
UV λ max (nm) | [M − H]- / / [M + H]+ (m/z) |
MS/MS (m/z) |
Tentative identification |
Concentration (mg/g)* | ||
|---|---|---|---|---|---|---|---|---|
| RBF | RBF-ACN | RBF-PP | ||||||
| RBF ANTHOCYANINS | ||||||||
| 1 2 3 4 |
15.77 16.44 17.18 18.20 |
280, 520 280, 520 280, 520 280, 520 |
611.1642 (+) 757.2220 (+) 449.1123 (+) 595.1734 (+) |
449.1078 287.0615 595.1663 287.0572 287.0589 287.0578 |
Cyanidin-3-O-sophoroside Cyanidin-3-O-glucosyl-rutinoside Cyanidin-3-O-glucoside Cyanidin-3-O-rutinoside |
3.07 ± 0.18 0.71 ± 0.05 1.41 ± 0.12 0.51 ± 0.04 |
520.02 ± 12.54 127.25 ± 6.23 235.26 ± 5.22 97.25 ± 3.41 |
28.29 ± 1.95 Trace amounts Trace amounts Trace amounts |
| RBF FLAVANOLS | ||||||||
| 5 6 7 |
9.84 10.74 12.61 |
280 280 280 |
577.1355 (-) 289.1139 (-) 289.1142 (-) |
289.0712 245.1203 245.1210 |
Procyanidin B Catechin Epicatechin |
0.30 ± 0.03 0.12 ± 0.02 0.58 ± 0.04 |
- - - |
42.43 ± 2.05 26.06 ± 1.13 140.39 ± 5.97 |
| RBF HYDROXYCINNAMIC ACID DERIVATIVES | ||||||||
| 8 9 10 11 |
9.61 12.60 14.30 14.93 |
278, 320 234, 316 234, 316 235, 325 |
341.0880 (-) 325.0933 (-) 325.0933 (-) 163.0402 (-) |
161.0238 145.0294 145.0293 119.0496 |
Caffeoyl hexoside p-Coumaryl hexoside isomer 1 p-Coumaryl hexoside isomer 2 p-Coumaric acid |
0.10 ± 0.01 0.08 ± 0.01 0.07 ± 0.01 0.09 ± 0.02 |
- - Trace amounts Trace amounts |
19.98 ± 0.96 27.72 ± 1.08 Trace amounts Trace amounts |
| RBF FLAVONOLS | ||||||||
| 12 13 14 15 16 17 18 19 |
22.19 23.35 23.69 24.41 24.62 25.88 28.45 29.52 |
254, 360 262, 363 266, 354 268, 354 272, 355 266, 348 256, 360 256, 360 |
433.0417 (-) 300.9992 (-) 463.0874 (-) 463.0882 (-) 477.0679 (-) 461.0733 (-) 475.0525 (-) 475.0525 (-) |
300.9982 229.0131 301.0334 301.0354 301.0345 285.0391 300.9978 299.9904 |
Ellagic acid-O-pentoside Ellagic acid Quercetin-3-O-galactoside Quercetin-3-O-glucoside Quercetin 3-O-glucuronide Kaempferol 3-O-glucuronide Ellagic acid acetyl pentoside isomer 1 Ellagic acid acetyl pentoside isomer 2 |
Trace amounts 0.21 ± 0.03 0.14 ± 0.01 Trace amounts Trace amounts Trace amounts Trace amounts Trace amounts |
Trace amounts Trace amounts 19.45 ± 1.37 Trace amounts Trace amounts Trace amounts Trace amounts Trace amounts |
|
* Values were expressed as mean ± SEM for three independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



