Submitted:
26 January 2025
Posted:
27 January 2025
You are already at the latest version
Abstract
Background/Objectives: The prefrontal cortex (PFC) plays a crucial role in cognitive functions such as attention, working memory and executive functions. Physical activity (PA), even at light-intensity, has been shown to increase PFC activity. However, most studies have focused on moderate- to vigorous-intensity exercises or on adults, while little data is available for children engaging in light-intensity PA (LPA). The aim of this study was to investigate the changes in the hemodynamics of the PFC during short-term, light-intensity exercises in children using functional near-infrared spectroscopy (fNIRS). The secondary aim was to compare the responses between two exercise patterns differing in duration (10 vs. 20 seconds).Methods: Thirty-eight healthy children aged 10 to 14 years from private schools in Barcelona participated in this study. Seven light-intensity exercises were performed in randomized order according to two patterns: Pattern 1 (10 seconds per exercise) and Pattern 2 (20 seconds per exercise). Changes in oxygenated hemoglobin (oxy-Hb) were measured by fNIRS in three PFC regions: right (R-PFC), middle (M-PFC) and left (L-PFC). Analysis of covariance (ANCOVA) was used to determine significant differences, with gender, age and resting oxy-Hb Z-scores as covariates.Results: Significant increases in oxy-Hb Z-scores were observed in the M-PFC during event A (Upward Stretch) and in all PFC regions during event D (Trunk Twist) in pattern 1. Events related to coordination and balance, such as balancing on one leg, also elicited significant activation of the PFC. However, simpler movements, such as hand washing and elbow circles, showed no significant changes. Comparisons between patterns 1 and 2 showed no significant differences, suggesting that duration alone has no effect on PFC activation.Conclusions: Light-intensity coordination and balance exercises stimulate PFC activity, even for short durations. These results emphasize the potential for integrating such exercises in school or rehabilitation settings to promote cognitive and motor development in children.
Keywords:
prefrontal cortex activation
; functional near-infrared spectroscopy
; child cognitive development
; motor coordination
; light-intensity physical activity
1. Introduction
Physical activity (PA) exerts a significant impact on neural development, especially in the formative years of childhood [1,2,3]. Engaging in PA activates complex biochemical processes that improve cerebral blood flow [4], stimulate the release of growth factors [5] such as Brain-Derived Neurotrophic Factor (BDNF) [6], [7] and promote the restructuring of neural circuits [8]. These physiological responses are crucial for the development of cognitive functions such as memory [9], [10], attention and executive processing [11], which are largely mediated by the prefrontal cortex (PFC) [12].
The PFC is significantly involved in higher-order cognitive tasks and behavioral regulation, making its development critical for academic performance and adaptive functioning [13,14]. Physical activities, especially low-intensity ones, contribute uniquely to the activation of this brain region [15], [16]. In contrast to vigorous physical activity (VPA), light-intensity physical activity (LPA) does not significantly stress the cardiovascular system [17,18], but is thought to stimulate sufficient neurophysiological changes that can improve cognitive function [19]. This form of exercise is particularly valuable in educational settings, as it can be seamlessly integrated into daily routines and provides cognitive benefits without the logistical challenges of more intense exercise programs [20,21].
Research has shown that even minimal increases in PA can improve cerebral blood flow and metabolic function in the PFC [22,23,24], leading to better cognitive performance. However, detailed insights into the specific hemodynamic changes induced by PLA in children's brains is still lacking. Given the potential of these activities to be regularly integrated into the classroom, there is an urgent need to understand their direct impact on children's neurodevelopment.
This study aims to address this gap by using functional near-infrared spectroscopy (fNIRS) to measure the hemodynamic responses of the PFC to short-duration, LPA in a pediatric population. fNIRS is particularly suited to this task because it is non-invasive and provides real-time insights into the changes in cerebral blood flow and oxygenation associated with brain activity. By elucidating how these exercises affect PFC activity, the study aims to underpin the role of LPA in improving cognitive function and potentially shape future educational and developmental strategies to effectively incorporate such exercises. This could not only improve academic performance, but also contribute to children's overall neurological health.
2. Materials and Methods
2.1. Participants
38 children aged 10 to 14 from private schools in Barcelona took part in this study. Of these, 16 participants were selected with the consent of the principals of their respective schools, while the remaining 22 were recruited through an extended snowball sampling method. Written informed consent was obtained from both the children and their parents or guardians, which also covered the use of the participants’ images in freely accessible online publications. The study was ethically approved by the Ethics Committee of the University of Zagreb (protocol code: 29; 4 2023). All research procedures were in strict compliance with international ethical standards and guidelines for research involving human subjects, with participant safety and data integrity being paramount throughout the study.
2.1.1. Number of Participants and fNIRS Data Analyzed
Of the 38 participants recruited for the study, data from 33 participants were included in the final analysis after excluding those with excessive motion artifacts based on predefined assessment criteria. For Pattern 1 (10 s), 30 participants (81.6% males) provided valid data, while for Pattern 2 (20 s), 31 participants (84.2% males) were eligible for analysis. Table 1 summarizes the demographic characteristics of the participants included in the study.
2.2. Procedure
The experimental sessions were conducted in a multipurpose room in a private facility and in a specific area of the collaborating private school in Barcelona. Before starting the experiment, the research team verified the name, age, gender and dominant hand of each participant. Detailed instructions were given for the LPA, emphasizing the importance of moving the head as little as possible, avoiding rapid movements and not holding the breath or straining excessively during the exercises.
To demonstrate the exercises and provide clarity, the researchers performed each movement as it would be performed during the actual experiment. Participants were then asked to rate their perceived exertion for each exercise using the modified Borg scale.
An fNIRS system (model and manufacturer's specifications, similar to the OEG-16H; Spectratech Inc. but localized for the study) was then carefully attached to each participant’s head while they were seated. The device was calibrated to ensure accurate measurement of cerebral blood flow. After verifying the accuracy of the device setting, participants performed the prescribed LPA as planned.
2.3. LPA
The study included seven different light intensity exercises, labeled A through G (see Table 2). The selection of these exercises was based on two criteria: (1) the exercises had to be of light intensity and simple enough that they could be comfortably performed by all children, including those with limited motor skills, in everyday situations, such as during short breaks in class or as part of regular class activities; and (2) the exercises required minimal head movement to reduce movement artifacts in the fNIRS data and thus avoid interference related to changes in cerebral blood flow due to head tilt. Consequently, the selected exercises excluded any form of running, jumping or extensive dynamic movements involving significant trunk and head movement in the sagittal or frontal plane.
The exercises selected were primarily gentle static and dynamic stretching exercises that are known to be feasible in typical school settings. As there is evidence that exercises involving fine motor skills and balance can activate the PFC, such movements were also integrated into the training program. All exercises were performed in a seated position, with the exception of a standing balance task to further limit movement noise during fNIRS monitoring.
To investigate the effects of exercise duration on cerebral blood flow, two exercise patterns were designed: one with a movement duration of 10 seconds (pattern 1) and another with a movement duration of 20 seconds (pattern 2). While static stretching exercises typically last 15 to 60 seconds per movement, a shorter duration was chosen in our protocol to focus the study on educational application rather than flexibility improvement. To control for possible sequence effects on the response variables, the order of the exercises (A-G) and the two patterns (1 and 2) was randomized for each participant, as shown in Figure 1 and Figure 2.
Instructional videos were produced for each child showing the order of the exercises as they were to be performed. Participants watched these exercises on a monitor at eye level, with a 10-second pause between the first and second movement. After each event, there was a 30-second pause during which participants viewed a calming naturalistic image to facilitate mental recovery. After completing the first exercise pattern, participants rested in a seated position for five minutes before recalibrating the fNIRS device to perform the second pattern.
3. Measures
3.1. fNIRS
Functional Near-Infrared Spectroscopy (fNIRS) is a non-invasive optical imaging technique that tracks the dynamics of cerebral blood flow by emitting near-infrared light through the skull. This light penetrates skin, bone and muscle tissue to a depth of approximately 2 cm and is therefore suitable for monitoring cortical activity, but not deeper brain structures. fNIRS measures variations in the concentrations of oxygenated hemoglobin (oxy-Hb) and deoxygenated hemoglobin (deoxy-Hb), providing insight into hemodynamic responses to neural activity. Compared to technologies such as functional magnetic resonance imaging (fMRI), fNIRS offers lower spatial resolution, but has the advantage of being less sensitive to motion artifacts and can record brain activity under more natural conditions without strict postural constraints.
In our study, we used an fNIRS system to monitor hemodynamic changes in the PFC of children during light exercise and at rest. The system was equipped with six transmitters and six receivers distributed over 16 channels to ensure comprehensive coverage of the PFC area. We used near-infrared wavelengths of 770 nm and 840 nm, with a distance of 3 cm between the transmitters and receivers to optimize signal detection. The placement of the sensors was strategically aligned with the Fpz position, according to the international 10–20 system used for EEG recordings. The data acquisition rate was set to 6.10 Hz, which corresponds to 0.08102 seconds per sample. This setup allowed us to closely monitor changes in oxy-Hb, deoxy-Hb, and total hemoglobin levels. Consistent with established research, our study focused on oxy-Hb levels as the primary indicator of regional cerebral blood flow, which is a reliable measure of PFC activity both during exercise and at rest.
3.2. fNIRS Data Processing
The fNIRS data was pre-processed using proprietary software integrated into the fNIRS system. First, all acquired data underwent a hemodynamic separation process to filter out signals associated with increased blood flow in the skin that were not indicative of cerebral activity. This step was crucial to ensure the accuracy of our measurements, which relate exclusively to brain function.
To further refine the data quality and remove extraneous noise from physiological sources such as heart rhythms and respiratory movements, a dual filtering approach was applied. A high-pass filter with a frequency of 0.01 Hz was used to suppress slow signal shifts, and a low-pass filter with a frequency of 0.3 Hz was applied to exclude high-frequency noise. This band-pass filtering effectively isolated the cerebral blood flow signals of interest.
In the analysis, the waveforms of oxygenated and deoxygenated hemoglobin (oxy-Hb and deoxy-Hb) were closely examined. The classification of the data was based on the phase relationship and stability of these signals:
- Data where oxy-Hb and deoxy-Hb were in antiphase were considered for further analysis.
- Data were retained when deoxy-Hb levels were stable and approached zero, indicating minimal extracerebral interference.
- Simultaneous increases or decreases in oxy-Hb and deoxy-Hb levels were specifically analyzed to assess their coherence.
- Data were selected if the proportion of deoxy-Hb remained lower than that of oxy-Hb, unless there was significant variation in the deoxy-Hb signal, which led to data exclusion.
After preprocessing, the data was extracted in CSV format and then imported into Microsoft Excel 2019 for detailed analysis. For each channel, the average oxy-Hb values at rest (0-5 seconds before exercise) and during exercise (10 or 20 seconds after the start of exercise) were calculated for all events and patterns. As each exercise pattern was repeated, the average values of the first and second event were calculated to increase the reliability of the results.
Channels were categorized to reflect specific regions of the PFC:
- -
- Channels 1 to 4 were assigned to the right PFC (R-PFC).
- -
- Channels 7 to 10 corresponded to the middle PFC (M-PFC).
- -
- Channels 13 to 16 represented the left PFC (L-PFC).
Mean oxy-Hb concentrations for each region were calculated both at rest and during exercise, and these values were subsequently converted to Z-scores to standardize results across different sessions and participants. This allowed a comparative analysis of hemodynamic responses under different conditions within the study.
4. Statistical Analysis
The analysis included the examination of differences in oxy-Hb-Z-scores at rest and during exercise in the regions of the PFC (R-PFC, M-PFC and L-PFC) for both exercise patterns (pattern 1 and pattern 2). To assess these differences, an analysis of covariance (ANCOVA) was performed, with age and gender included as covariates to account for potential confounding effects.
To further investigate whether the rate of change in oxy-Hb levels varied between pattern 1 and pattern 2, an additional ANCOVA was performed. In this analysis, baseline oxy-Hb Z values (measured at rest) were included as covariates along with age and gender to ensure a robust comparison between conditions.
A significance threshold of P < 0.05 was applied to all statistical tests to determine meaningful differences. Statistical analyses were performed using IBM SPSS Statistics software (version 29.0; IBM Corp., Armonk, NY, USA) to ensure consistency and reliability of data analysis. This approach allowed for a comprehensive assessment of hemodynamic responses to LPA in all regions and patterns of the PFC.
5. Results
Ratings of Perceived Exertion (RPE)
Perceived exertion for each exercise category was rated using the Borg scale, ranging from 6 ("no exertion") to 20 ("maximum exertion"). All reported RPE scores fell below "11"," which categorised the exercises as light-intensity. The mean RPE score for the group was 7.5 ± 1.6, with most participants describing the exercises as "very light" (score 9) or "extremely light" (score 7) according to the Borg scale descriptors. These results confirm that all exercises performed in this study met the criteria for LPA.
Hemodynamic changes in the PFC
Comparison between rest and LPA (pattern 1)
In the 10-second movement pattern, certain regions of the PFC showed significant changes in oxygenated hemoglobin (oxy-Hb) Z-scores during exercise compared to rest. For event A, the M-PFC showed a significant increase (F [1, 64] = 4.60; P = 0.039). For event B, no significant differences were observed in any of the PFC regions.
For event C, significant increases were observed in both the M-PFC (F [1, 64] = 3.87; P = 0.042) and the L-PFC (F [1, 64] = 5.21; P = 0.019). Similarly, an increase in M-PFC (F [1, 64] = 4.90; P = 0.033) and L-PFC (F [1, 64] = 5.87; P = 0.017) was observed for event D.
In event E, a significant increase in oxy-Hb-Z-scores were identified only in the L-PFC (F [1, 64] = 3.87; P = 0.049). Events F and G showed significant increases in all PFC regions. For event F, significant changes were observed in the R-PFC (F [1, 64] = 4.98; P = 0.028), M-PFC (F [1, 64] = 6.73; P = 0.012) and L-PFC (F [1, 64] = 8.53; P = 0.005).
In event G, increases in oxy-Hb Z-scores were observed in R-PFC (F [1, 64] = 8.23; P = 0.008), M-PFC (F [1, 64] = 10.80; P = 0.002) and L-PFC (F [1, 64] = 14.55; P < 0.001) during exercise compared to rest (Table 3).
Comparison Between Rest and LPA (Pattern 2)
During the 20-second movement pattern (pattern 2), significant increases in oxygenated hemoglobin (oxy-Hb) Z-scores were observed in several regions of the PFC for certain events. For event A, the R-PFC showed a significant increase during exercise compared to rest (F [1, 64] = 7.12; P = 0.011). No significant changes were observed in the M-PFC and L-PFC during this event.
For event B, a significant increase in M-PFC was observed (F [1, 64] = 3.95; P = 0.049). However, no significant differences in R-PFC or L-PFC were observed.
Event C showed significant increases in both the M-PFC (F [1, 64] = 5.42; P = 0.026) and the R-PFC (F [1, 64] = 4.28; P = 0.041). No significant differences were observed in the L-PFC during this event.
For event D, all three PFC regions showed significant increases in oxy-Hb Z-scores. In particular, the R-PFC (F [1, 64] = 18.31; P < 0.001), the M-PFC (F [1, 64] = 22.10; P < 0.001) and the L-PFC (F [1, 64] = 26.87; P < 0.001) showed significant changes.
For event E, the R-PFC (F [1, 64] = 7.21; P = 0.012), the M-PFC (F [1, 64] = 13.47; P < 0.001) and the L-PFC (F [1, 64] = 5.19; P = 0.029) all showed a significant increase during exercise.
For event F, the R-PFC (F [1, 64] = 11.65; P = 0.001), the M-PFC (F [1, 64] = 4.71; P = 0.032) and the L-PFC (F [1, 64] = 9.88; P = 0.003) showed significant increases in oxy-HbZ-scores during exercise compared to rest.
Similarly, event G showed significant increases in all three PFC regions. Significant changes were observed in the R-PFC (F [1, 64] = 16.20; P < 0.001), M-PFC (F [1, 64] = 19.10; P < 0.001) and L-PFC (F [1, 64] = 22.71; P < 0.001) (Table 4).
Comparison Between Pattern 1 and Pattern 2
The comparison of the oxy-Hb-Z-scores between Pattern 1 and Pattern 2 showed no significant differences in any of the regions of the PFC for most events. Slight differences were observed in the mean oxy-Hb-Z-scores, but none of the changes reached statistical significance (P > 0.05). This consistency between patterns suggests a similar hemodynamic response in the R-PFC, M-PFC, and L-PFC during LPA (Table 5).
6. Discussion
This study investigated the hemodynamic responses in the PFC during short-term, light-intensity exercises using functional near-infrared spectroscopy (fNIRS). The results revealed several significant findings that shed light on how specific movements and exercise patterns affect oxygenated hemoglobin (oxy-Hb) levels in different regions of the PFC. At the same time, there were some unexpected findings that lead to further research into the relationship between light exercise and brain activity.
One of the most notable findings was the significant increase in oxy-Hb-Z-scores in the middle PFC (M-PFC) during event A (upward stretch) in pattern 1, which is consistent with previous studies [25,26,27,28] indicating that the M-PFC is particularly sensitive to even subtle motor demands, especially when associated with cognitive processes related to the maintenance of postural control. Similar activation of the M-PFC during simple aerobic exercises was reported [29,30], suggesting that even light-intensity tasks can activate this region, possibly due to its role in the integration of motor and executive functions. Although the upward stretch was simple, it may have required sufficient coordination to stimulate activity in the PFC. Another important finding was the significant increase in oxy-Hb-Z-scores across all three PFC regions — right (R-PFC), middle (M-PFC) and left (L-PFC) — during event D (Trunk Twist) in pattern 1. This finding supports the hypothesis that exercises requiring both motor coordination and bilateral movements activate broader PFC regions. Similar conclusions [31,32] emphasized that tasks involving cross-body movements activate broader cortical networks due to the integration of motor and cognitive functions. Trunk twist probably required not only physical coordination but also spatial awareness and balance, activating a broader range of PFC networks. In contrast, some events, such as event C (elbow circles) and event E (washing hands), showed no significant differences in oxy-Hb-Z-scores between rest and exercise conditions. This was unexpected as previous literature [33,34] that indicates that even light-intensity activities typically lead to measurable activation of the PFC. One plausible explanation is that these movements were too simple and repetitive to require substantial activation of the PFC. Exercises with minimal cognitive or motor complexity may not activate the PFC to the same extent as more demanding tasks. Furthermore, these non-significant results may reflect individual variability in neuronal responses to light-intensity activity, as shown in previous studies. The comparison between patterns 1 and 2 across multiple events also revealed no significant differences in oxy-Hb-Z scores, suggesting that doubling the exercise duration (from 10 to 20 seconds) did not induce significant changes in PFC activation. This result is consistent with previous studies [35,36,37], who observed that the intensity and complexity of a task plays a more important role in triggering PFC activation than its duration. The lack of significant differences suggests that the short duration used in this study may have limited the ability to detect robust effects, especially for movements that inherently require less cognitive involvement. The results of this study emphasize the nuanced relationship between exercise type, complexity and duration in influencing PFC activation. For example, the results show that tasks requiring coordination, balance or bilateral movements, such as trunk twists and single-leg balance exercises, consistently elicit higher activation in the PFC regions. This is consistent with the concept of dual-tasking, in which simultaneous physical and cognitive demands lead to increased cortical activation [38,39,40]. In contrast, simple and repetitive movements, even when performed for longer durations, appear to have limited effects on PFC activation, highlighting the importance of task complexity in exercise interventions to promote brain activity. Comparisons with previous studies show both similarities and discrepancies. The significant increases in oxy-Hb-Z-scores observed during certain events, particularly those related to coordination and balance, were confirmed with previous studies [29,38,41,42,43], who showed that PA, even at light intensity, can induce significant changes in PFC activity under certain conditions. However, the non-significant findings in some events are in contrast to some previous studies [44,45,46,47], who reported consistent PFC activation during light-intensity exercises. These differences could be due to variations in the types of exercises studied, participant characteristics or measurement methods. In addition, the limited duration of exercises in this study may have contributed to the lack of consistent results, that emphasise the importance of sufficient exercise duration for the demonstration of reliable neural effects [48,49,50,51]. Despite their contributions, this study has several limitations. The short duration of exercises may have limited the ability to observe robust changes in oxy-Hb-Z Z-scores, particularly for events that inherently require minimal cognitive or motor involvement. Future studies should extend the duration of the exercises or include more complex tasks to better understand the relationship between light activity and PFC activation. Furthermore, while the sample size is sufficient to detect significant trends, the generalizability of the results is limited. Expanding the participant pool to include individuals of different ages, fitness levels, and cognitive abilities could provide a more comprehensive understanding of how light-intensity exercise affects PFC activity. Finally, the lack of a control condition, such as a baseline without training, limits the ability to isolate the specific effects of training from other potentially confounding factors, such as participant engagement or external stimuli.
7. Conclusions
In conclusion, this study demonstrates that light-intensity exercises can induce significant hemodynamic changes in the PFC, especially during tasks requiring coordination, balance or bilateral movements. However, the lack of consistent activation across events emphasises the critical role of task complexity and duration in PFC activation. These findings contribute to the growing body of evidence for the role of PA in promoting brain health and provide valuable insights for the design of PA interventions in educational and clinical settings.
Author Contributions
Conceptualization, T.P.; methodology, T.P.; software, T.P.; validation, T.P.; formal analysis, T.P.; investigation, T.P.; resources, T.P.; data curation, T.P.; writing—original draft preparation, T.P.; writing—review and editing, T.P.; visualization, T.P.; supervision, T.P.; project administration, T.P.; funding acquisition, T.P.
Funding
This research received no external funding.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data supporting the findings of this study are not publicly available due to privacy/ethical restrictions. Data are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- H. İ. Ceylan, A. F. Silva, R. Ramirez-Campillo, and E. Murawska-Ciałowicz, “Exploring the Effect of Acute and Regular Physical Exercise on Circulating Brain-Derived Neurotrophic Factor Levels in Individuals with Obesity: A Comprehensive Systematic Review and Meta-Analysis,” May 01, 2024, Multidisciplinary Digital Publishing Institute (MDPI). [CrossRef]
- M. Kukla-Bartoszek and K. Głombik, “Train and Reprogram Your Brain: Effects of Physical Exercise at Different Stages of Life on Brain Functions Saved in Epigenetic Modifications,” Nov. 01, 2024, Multidisciplinary Digital Publishing Institute (MDPI). [CrossRef]
- F. Latino and F. Tafuri, “The role of physical activity in the physiological activation of the scholastic pre-requirements,” AIMS Neurosci, vol. 11, no. 3, pp. 244–259, 2024. [CrossRef]
- X. Li, X. Qu, K. Shi, Y. Yang, and J. Sun, “Physical exercise for brain plasticity promotion an overview of the underlying oscillatory mechanism,” 2024, Frontiers Media SA. [CrossRef]
- J. Q. Jing, S. J. Jia, and C. J. Yang, “Physical activity promotes brain development through serotonin during early childhood,” Aug. 30, 2024, Elsevier Ltd. [CrossRef]
- M. H. Khalil, “The BDNF-Interactive Model for Sustainable Hippocampal Neurogenesis in Humans: Synergistic Effects of Environmentally-Mediated Physical Activity, Cognitive Stimulation, and Mindfulness,” Dec. 01, 2024, Multidisciplinary Digital Publishing Institute (MDPI). [CrossRef]
- R. R. Kraemer and B. R. Kraemer, “The effects of peripheral hormone responses to exercise on adult hippocampal neurogenesis,” 2023, Frontiers Media SA. [CrossRef]
- M. H. Khalil, “Neurosustainability,” Front Hum Neurosci, vol. 18, 2024. [CrossRef]
- S. L. Plas et al., “Neural circuits for the adaptive regulation of fear and extinction memory,” 2024, Frontiers Media SA. [CrossRef]
- S. Battaglia, A. Avenanti, L. Vécsei, and M. Tanaka, “Neural Correlates and Molecular Mechanisms of Memory and Learning,” Mar. 01, 2024, Multidisciplinary Digital Publishing Institute (MDPI). [CrossRef]
- M. Ribeiro, Y. N. Yordanova, V. Noblet, G. Herbet, and D. Ricard, “White matter tracts and executive functions: A review of causal and correlation evidence,” Feb. 01, 2024, Oxford University Press. [CrossRef]
- F. Mao, F. Huang, S. Zhao, and Q. Fang, “Effects of cognitively engaging physical activity interventions on executive function in children and adolescents: a systematic review and meta-analysis,” 2024, Frontiers Media SA. [CrossRef]
- M. A. Salehinejad, E. Ghanavati, M. H. A. Rashid, and M. A. Nitsche, “Hot and cold executive functions in the brain: A prefrontal-cingular network,” Brain Neurosci Adv, vol. 5, p. 239821282110077, Jan. 2021. [CrossRef]
- N. Boyle, S. Betts, and H. Lu, “Monoaminergic Modulation of Learning and Cognitive Function in the Prefrontal Cortex,” Brain Sci, vol. 14, no. 9, p. 902, Sep. 2024. [CrossRef]
- M. E. Anderson, E. J. Wind, and L. S. Robison, “Exploring the neuroprotective role of physical activity in cerebral small vessel disease,” Jun. 15, 2024, Elsevier B.V. [CrossRef]
- A. Kaiser et al., “A Randomized Controlled Trial on the Effects of a 12-Week High- vs. Low-Intensity Exercise Intervention on Hippocampal Structure and Function in Healthy, Young Adults,” Front Psychiatry, vol. 12, Jan. 2022. [CrossRef]
- R. B. Batacan, M. J. Duncan, V. J. Dalbo, P. S. Tucker, and A. S. Fenning, “Effects of Light Intensity Activity on CVD Risk Factors: A Systematic Review of Intervention Studies,” 2015, Hindawi Limited. [CrossRef]
- D. Greenwalt, S. Phillips, C. Ozemek, R. Arena, and A. Sabbahi, “The Impact of Light Physical Activity, Sedentary Behavior and Cardiorespiratory Fitness in Extending Lifespan and Healthspan Outcomes: How Little is Still Significant? A Narrative Review,” Oct. 01, 2023, Elsevier Inc. [CrossRef]
- L. Youssef et al., “Neurophysiological effects of acute aerobic exercise in young adults: a systematic review and meta-analysis,” Sep. 01, 2024, Elsevier Ltd. [CrossRef]
- J. Siette et al., “A Pilot Study of BRAIN BOOTCAMP, a Low-Intensity Intervention on Diet, Exercise, Cognitive Activity, and Social Interaction to Improve Older Adults’ Dementia Risk Scores,” Journal of Prevention of Alzheimer’s Disease, Oct. 2024. [CrossRef]
- C. Martínez-Díaz and L. Carrasco Páez, “Little but Intense: Using a HIIT-Based Strategy to Improve Mood and Cognitive Functioning in College Students,” Healthcare (Switzerland), vol. 11, no. 13, Jul. 2023. [CrossRef]
- Xu, J. Yu, G. Li, and Y. Wang, “Exercise intervention on the brain structure and function of patients with mild cognitive impairment: systematic review based on magnetic resonance imaging studies,” 2024, Frontiers Media SA. [CrossRef]
- Huang et al., “Comparative effects of different types of exercise on endothelial function, arterial stiffness, and executive function in sedentary young individuals: a randomized controlled trial,” Mar. 11, 2024. [CrossRef]
- X. Chen, N. Hu, H. Han, G. Cai, and Y. Qin, “Effects of high-intensity interval training in a cold environment on arterial stiffness and cerebral hemodynamics in sedentary Chinese college female students post-COVID-19,” Front Neurol, vol. 15, 2024. [CrossRef]
- S. Keshmiri, H. Sumioka, R. Yamazaki, and H. Ishiguro, “Differential effect of the physical embodiment on the prefrontal cortex activity as quantified by its entropy,” Entropy, vol. 21, no. 9, Sep. 2019. [CrossRef]
- S. Zimmermann, R. Richardson, and K. D. Baker, “Maturational changes in prefrontal and amygdala circuits in adolescence: implications for understanding fear inhibition during a vulnerable period of development,” Mar. 01, 2019, MDPI AG. [CrossRef]
- Trofimova, “Functional Constructivism Approach to Multilevel Nature of Bio-Behavioral Diversity,” Oct. 27, 2021, Frontiers Media S.A. [CrossRef]
- A. Kampaite et al., “Brain connectivity changes underlying depression and fatigue in relapsing-remitting multiple sclerosis: A systematic review,” PLoS One, vol. 19, no. 3 March, Mar. 2024. [CrossRef]
- T. Naito, K. Oka, and K. Ishii, “Hemodynamics of short-duration light-intensity physical exercise in the prefrontal cortex of children: a functional near-infrared spectroscopy study,” Sci Rep, vol. 14, no. 1, Dec. 2024. [CrossRef]
- G. Cruz, Y. N. Leow, N. M. Le, E. Adam, R. Huda, and M. Sur, “CORTICAL-SUBCORTICAL INTERACTIONS IN GOAL-DIRECTED BEHAVIOR,” Physiol Rev, vol. 103, no. 1, pp. 347–389, Jan. 2023. [CrossRef]
- T. W. Cacciatore, D. I. Anderson, and R. G. Cohen, “Central mechanisms of muscle tone regulation: implications for pain and performance,” Front Neurosci, vol. 18, 2024. [CrossRef]
- H. B. Coslett, J. Medina, D. K. Goodman, Y. Wang, and A. Burkey, “Can they touch? A novel mental motor imagery task for the assessment of back pain,” Frontiers in Pain Research, vol. 4, 2023. [CrossRef]
- E. Sanchis-Navarro, F. G. Luna, J. Lupiáñez, and F. Huertas, “Benefits of a light- intensity bout of exercise on attentional networks functioning,” Sci Rep, vol. 14, no. 1, p. 25745, Dec. 2024. [CrossRef]
- Jung et al., “A mechanistic understanding of cognitive performance deficits concurrent with vigorous intensity exercise,” Brain Cogn, vol. 180, Oct. 2024. [CrossRef]
- Guan et al., “Effects and neural mechanisms of different physical activity on major depressive disorder based on cerebral multimodality monitoring: a narrative review,” 2024, Frontiers Media SA. [CrossRef]
- Jung, S. Ryu, M. Kang, A. H. Javadi, and P. D. Loprinzi, “Evaluation of the transient hypofrontality theory in the context of exercise: A systematic review with meta-analysis,” Quarterly Journal of Experimental Psychology, vol. 75, no. 7, pp. 1193–1214, Jul. 2022. [CrossRef]
- J. Crum et al., “Body fat predictive of acute effects of exercise on prefrontal hemodynamics and speed,” Neuropsychologia, vol. 196, Apr. 2024. [CrossRef]
- C. Kan et al., “Differences in cortical activation characteristics between younger and older adults during single/dual-tasks: A cross-sectional study based on fNIRS,” Biomed Signal Process Control, vol. 99, Jan. 2025. [CrossRef]
- Saraiva, M. A. Castro, and J. P. Vilas-Boas, “Muscular and Prefrontal Cortex Activity during Dual-Task Performing in Young Adults,” Eur J Investig Health Psychol Educ, vol. 13, no. 4, pp. 736–747, Apr. 2023. [CrossRef]
- P. H. M. Monteiro, A. J. Marcori, N. R. da Conceição, R. L. M. Monteiro, D. B. Coelho, and L. A. Teixeira, “Cortical activity in body balance tasks as a function of motor and cognitive demands: A systematic review,” Nov. 01, 2024, John Wiley and Sons Inc. [CrossRef]
- E. Zangen et al., “Prefrontal cortex neurons encode ambient light intensity differentially across regions and layers,” Nat Commun, vol. 15, no. 1, Dec. 2024. [CrossRef]
- D. Kimura, T. Hosokawa, T. Ujikawa, and T. Ito, “Effects of different exercise intensities on prefrontal activity during a dual task,” Sci Rep, vol. 12, no. 1, Dec. 2022. [CrossRef]
- Y. Hong, D. Bao, B. Manor, Y. Zhou, and J. Zhou, “Effects of endurance exercise on physiologic complexity of the hemodynamics in prefrontal cortex,” Neurophotonics, vol. 11, no. 01, Mar. 2024. [CrossRef]
- C. Damrongthai et al., “Slow running benefits: Boosts in mood and facilitation of prefrontal cognition even at very light intensity,” Jan. 30, 2024. [CrossRef]
- Jung, S. Ryu, M. Kang, A. H. Javadi, and P. D. Loprinzi, “Evaluation of the transient hypofrontality theory in the context of exercise: A systematic review with meta-analysis,” Quarterly Journal of Experimental Psychology, vol. 75, no. 7, pp. 1193–1214, Jul. 2022. [CrossRef]
- S. Cheng, L. Cong, D. Hui, C. Teng, W. Li, and J. Ma, “Enhancement of prefrontal functional connectivity under the influence of concurrent physical load during mental tasks,” Front Hum Neurosci, vol. 18, 2024. [CrossRef]
- J. Liu, Y. Liu, and L. Wu, “Exploring the dynamics of prefrontal cortex in the interaction between orienteering experience and cognitive performance by fNIRS,” Sci Rep, vol. 14, no. 1, Dec. 2024. [CrossRef]
- Sahnoune et al., “Exercise ameliorates neurocognitive impairments in a translational model of pediatric radiotherapy,” Neuro Oncol, vol. 20, no. 5, pp. 695–704, Apr. 2018. [CrossRef]
- L. Hosang, E. Mouchlianitis, S. M. R. Guérin, and C. I. Karageorghis, “Effects of exercise on electroencephalography-recorded neural oscillations: a systematic review,” Int Rev Sport Exerc Psychol, 2022. [CrossRef]
- F. Sanaeifar et al., “Beneficial effects of physical exercise on cognitive-behavioral impairments and brain-derived neurotrophic factor alteration in the limbic system induced by neurodegeneration,” Oct. 01, 2024, Elsevier Inc. [CrossRef]
- Zhang, J. Lu, Y. Sun, and J. Li, “Recreational gymnastics exercise of moderate intensity enhances executive function in Chinese preschoolers: A randomized controlled trial,” Psych J, Dec. 2024. [CrossRef]
Figure 1.
fNIRS Measurement Setup: Randomized Order of Patterns (1 and 2) and Events (A–G).
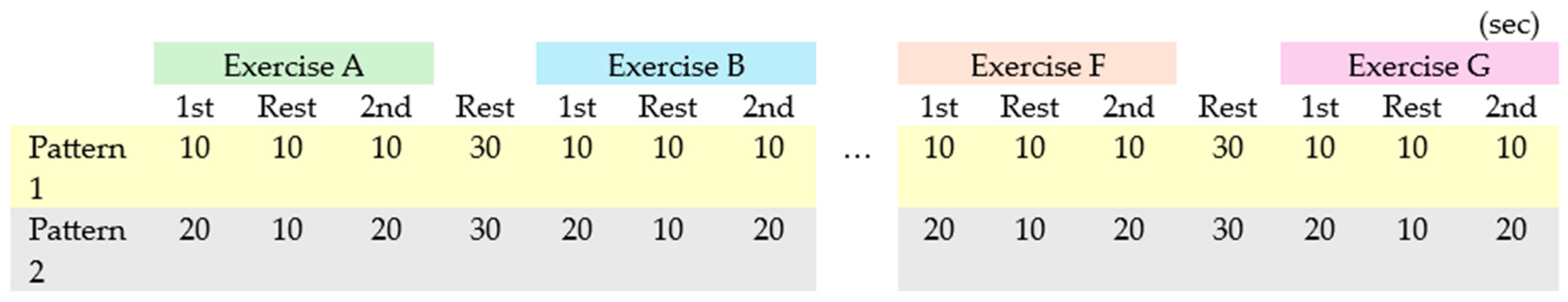
Figure 2.
Placement Points and Channel Coordinates of Functional Near-Infrared Spectroscopy Probes.
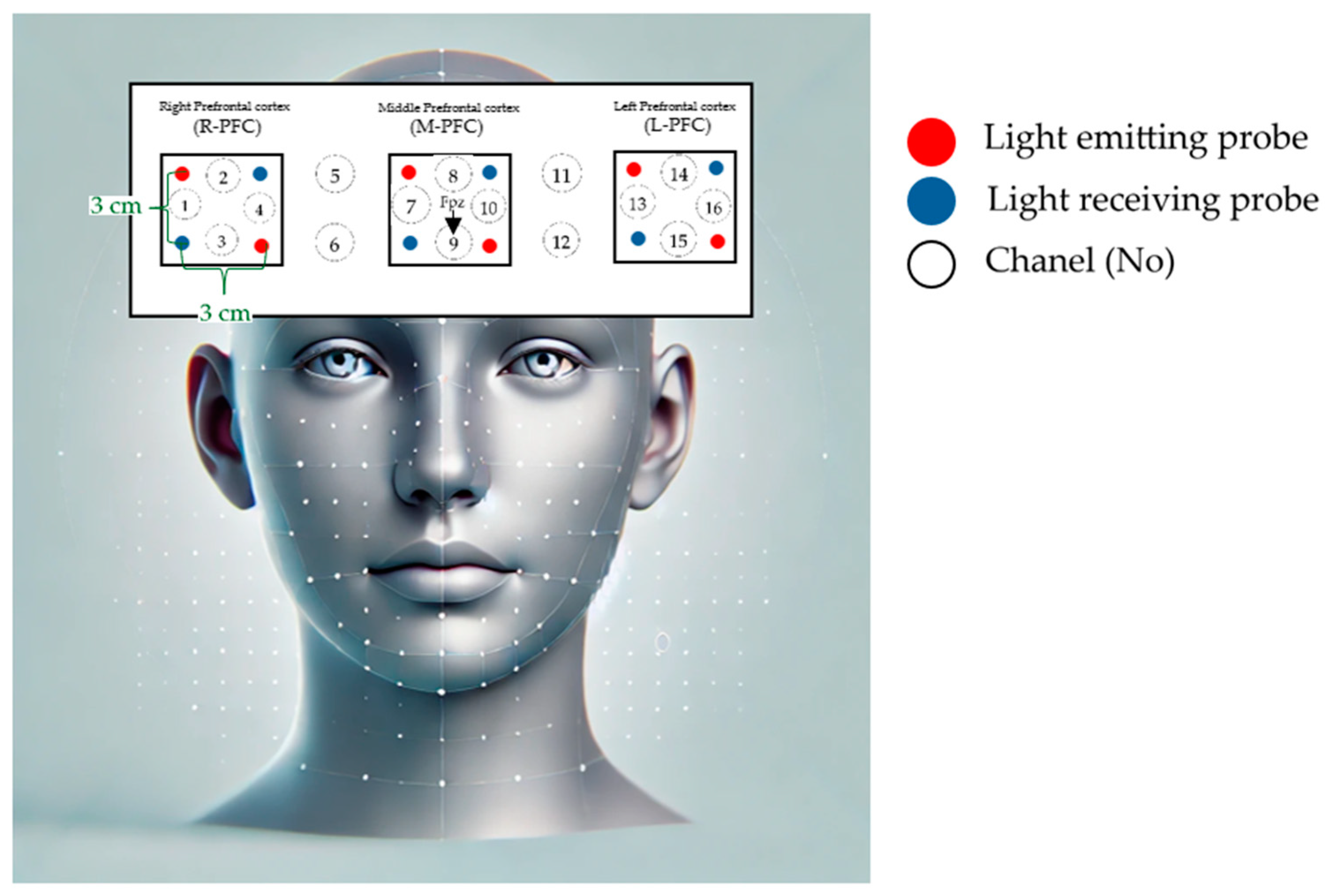

Table 1.
Participant characteristics and data analyzed in Patterns 1 and 2.
| All | Pattern 1 | Pattern 2 | ||||
|---|---|---|---|---|---|---|
| N | % | N | % | N | % | |
| Age, mean ± SD, years | 12.3 ± 1.4 | 12.3 ± 1.3 | 12.3 ± 1.3 | |||
| Gender | ||||||
| Male | 24 | 63 | 21 | 66 | 22 | 65 |
| Female | 14 | 37 | 11 | 34 | 12 | 35 |
| Age Groups | ||||||
| 10–11 years | 12 | 31 | 9 | 29 | 10 | 30 |
| 12 years | 15 | 39 | 12 | 38 | 13 | 38 |
| 13–14 years | 11 | 29 | 8 | 25 | 9 | 27 |
| Dominant Hand | ||||||
| Right | 35 | 92 | 30 | 91 | 31 | 91 |
| Left | 3 | 8 | 3 | 9 | 3 | 9 |
SD: Standard deviation. Pattern 1: Exercises performed with a 10-second duration. Pattern 2: Exercises performed with a 20-second duration.
Table 2.
Light-intensity exercise utilized in protocol.
| Event ID | Exercise | Description |
|---|---|---|
| A | Nick Tilts | Gently tilt your head toward one shoulder and hold for a few seconds, then repeat on the other side. |
| B | Arm Cross Stretch | Extend one arm straight across your chest. Use the other hand to pull the extended arm closer, stretching the shoulder. |
| C | Wrist Rolls | Extend your arms out in front of you and gently roll your wrists clockwise and then counterclockwise. |
| D | Side Bends | Stand with feet shoulder-width apart. Slowly bend to one side, sliding your hand down your leg, then switch to the other side. |
| E | Heel Digs | Stand upright and dig one heel into the ground, toe pointing upwards. Alternate legs. |
| F | Torso Twists | Stand with feet planted firmly, twist your torso to one side as far as is comfortable, then switch sides. |
| G | Marching in Place | Lift your knees high one at a time, as if marching on the spot. |
| Ankle Circles | Lift one foot off the ground and rotate the ankle in a circular motion. Rotate a few times in one direction, then switch directions and finally switch feet. |
Table 3.
Changes in oxy-Hb Z-scores observed during rest and LPA (Pattern 1).
|
Event (ID-Seconds) |
Region |
Oxy-Hb (mM・mm) | Oxy-Hb (Z-score) |
F |
P |
η² |
||||||
| Rest | Exercise | Rest | Exercise | |||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||||
| A-10 Upward Stretch | R-PFC | 0,05 | 0,11 | 0,09 | 0,12 | -0,10 | 0,08 | 0,65 | 0,10 | 1,35 | 0.430 | 0,01 |
| M-PFC | -0,03 | 0,10 | 0,06 | 0,14 | -0,27 | 0,09 | 0,68 | 0,11 | 4,60 | 0.039* | 0,07 | |
| L-PFC | 0,02 | 0,15 | 0,08 | 0,16 | -0,15 | 0,13 | 0,80 | 0,14 | 1,22 | 0.276 | 0,03 | |
| B-10 Shoulder Stretch | R-PFC | 0,01 | 0,09 | 0,02 | 0,10 | -0,07 | 0,07 | 0,50 | 0,08 | 1,08 | 0.362 | 0,00 |
| M-PFC | 0,00 | 0,08 | 0,12 | 0,15 | -0,12 | 0,10 | 0,85 | 0,12 | 2,03 | 0.092 | 0,04 | |
| L-PFC | 0,01 | 0,11 | 0,10 | 0,12 | -0,10 | 0,09 | 0,89 | 0,11 | 1,40 | 0.249 | 0,02 | |
| C-10 Elbow Circles | R-PFC | -0,02 | 0,12 | 0,08 | 0,14 | -0,16 | 0,11 | 0,85 | 0,12 | 1,88 | 0.085 | 0,05 |
| M-PFC | -0,05 | 0,14 | 0,10 | 0,16 | -0,50 | 0,13 | 0,69 | 0,14 | 3,87 | 0.042* | 0,08 | |
| L-PFC | -0,03 | 0,13 | 0,11 | 0,15 | -0,42 | 0,12 | 0,87 | 0,13 | 5,21 | 0.019* | 0,09 | |
| D-10 Trunk Twist | R-PFC | 0,00 | 0,10 | 0,07 | 0,12 | -0,10 | 0,08 | 0,62 | 0,10 | 1,35 | 0.440 | 0,01 |
| M-PFC | -0,03 | 0,11 | 0,08 | 0,14 | -0,27 | 0,09 | 0,80 | 0,11 | 4,90 | 0.033* | 0,07 | |
| L-PFC | -0,04 | 0,12 | 0,09 | 0,15 | -0,29 | 0,12 | 0,84 | 0,13 | 5,87 | 0.017* | 0,10 | |
| E-10 Washing Hands | R-PFC | 0,03 | 0,10 | 0,06 | 0,12 | -0,10 | 0,08 | 0,63 | 0,10 | 1,27 | 0.402 | 0,01 |
| M-PFC | 0,02 | 0,09 | 0,11 | 0,14 | -0,22 | 0,09 | 0,48 | 0,12 | 2,78 | 0.068 | 0,05 | |
| L-PFC | 0,00 | 0,08 | 0,09 | 0,11 | -0,34 | 0,07 | 0,89 | 0,10 | 3,87 | 0.049* | 0,06 | |
| F-10 Thumb & Pinky | R-PFC | -0,01 | 0,11 | 0,07 | 0,13 | -0,20 | 0,10 | 0,72 | 0,11 | 1,70 | 0.285 | 0,02 |
| M-PFC | -0,03 | 0,12 | 0,10 | 0,14 | -0,29 | 0,11 | 0,72 | 0,13 | 4,98 | 0.028* | 0,08 | |
| L-PFC | -0,02 | 0,13 | 0,12 | 0,15 | -0,34 | 0,12 | 0,89 | 0,13 | 6,73 | 0.012* | 0,12 | |
| G-10 Single Leg Balance | R-PFC | -0,04 | 0,10 | 0,09 | 0,12 | -0,28 | 0,08 | 0,90 | 0,10 | 6,10 | 0.017* | 0,11 |
| M-PFC | -0,08 | 0,09 | 0,15 | 0,12 | -0,38 | 0,07 | 0,96 | 0,09 | 8,23 | 0.008* | 0,14 | |
| L-PFC | -0,06 | 0,11 | 0,13 | 0,13 | -0,42 | 0,10 | 0,78 | 0,11 | 9,45 | 0.004* | 0,16 | |
Gender and age were included as covariates. R-PFC: right prefrontal cortex, M-PFC: middle prefrontal cortex, L-PFC: left prefrontal cortex, SD: standard deviation, oxy-Hb: oxygenated hemoglobin. *Indicates a statistically significant difference between rest and exercise Z-scores (P < .05).
Table 4.
Changes in oxy-Hb Z-scores observed during rest and LPA (Pattern 2).
|
Event (ID-Seconds) |
Region |
Oxy-Hb (mM mm) | Oxy-Hb (Z-score) |
F |
P |
η² |
||||||
| Rest | Exercise | Rest | Exercise | |||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||||
| A-20 Upward Stretch | R-PFC | -0,05 | 0,22 | 0,10 | 0,30 | -0,32 | 0,90 | 0,28 | 0,92 | 7,12 | 0.011* | 0,10 |
| M-PFC | 0,02 | 0,25 | 0,12 | 0,28 | -0,22 | 0,78 | 0,64 | 1,15 | 3,32 | 0.068 | 0,05 | |
| L-PFC | -0,03 | 0,23 | 0,14 | 0,27 | -0,14 | 0,80 | 0,71 | 1,10 | 2,10 | 0.125 | 0,03 | |
| B-20 Shoulder Stretch | R-PFC | 0,00 | 0,14 | 0,08 | 0,18 | -0,08 | 0,68 | 0,62 | 1,01 | 1,82 | 0.220 | 0,02 |
| M-PFC | 0,04 | 0,16 | 0,15 | 0,20 | -0,16 | 0,80 | 0,82 | 0,98 | 3,95 | 0.049* | 0,06 | |
| L-PFC | -0,01 | 0,15 | 0,09 | 0,19 | -0,12 | 0,73 | 0,68 | 1,03 | 2,37 | 0.099 | 0,04 | |
| C-20 Elbow Circles | R-PFC | 0,02 | 0,13 | 0,11 | 0,24 | -0,18 | 0,72 | 0,71 | 1,15 | 4,28 | 0.041* | 0,07 |
| M-PFC | -0,03 | 0,17 | 0,14 | 0,25 | -0,25 | 0,65 | 0,89 | 1,10 | 5,42 | 0.026* | 0,09 | |
| L-PFC | -0,02 | 0,14 | 0,12 | 0,23 | -0,15 | 0,68 | 0,74 | 1,07 | 3,10 | 0.073 | 0,05 | |
| D-20 Trunk Twist | R-PFC | -0,06 | 0,12 | 0,15 | 0,20 | -0,38 | 0,70 | 0,85 | 1,10 | 18,31 | < 0.001* | 0,21 |
| M-PFC | -0,08 | 0,18 | 0,19 | 0,27 | -0,40 | 0,74 | 0,92 | 1,12 | 22,10 | < 0.001* | 0,24 | |
| L-PFC | -0,09 | 0,16 | 0,20 | 0,26 | -0,45 | 0,78 | 1,01 | 1,15 | 26,87 | < 0.001* | 0,28 | |
| E-20 Washing Hands | R-PFC | -0,04 | 0,13 | 0,09 | 0,18 | -0,30 | 0,68 | 0,64 | 0,98 | 7,21 | 0.012* | 0,11 |
| M-PFC | -0,05 | 0,15 | 0,13 | 0,22 | -0,36 | 0,62 | 0,81 | 0,93 | 13,47 | < 0.001* | 0,15 | |
| L-PFC | -0,03 | 0,12 | 0,10 | 0,18 | -0,34 | 0,59 | 0,74 | 0,85 | 5,19 | 0.029* | 0,07 | |
| F-20 Thumb & Pinky | R-PFC | -0,02 | 0,13 | 0,07 | 0,16 | -0,20 | 0,64 | 0,72 | 0,90 | 11,65 | 0.001* | 0,14 |
| M-PFC | 0,01 | 0,14 | 0,09 | 0,18 | -0,15 | 0,70 | 0,81 | 0,92 | 4,71 | 0.032* | 0,08 | |
| L-PFC | -0,03 | 0,15 | 0,11 | 0,20 | -0,31 | 0,78 | 0,85 | 1,03 | 9,88 | 0.003* | 0,12 | |
| G-20 Single Leg Balance | R-PFC | -0,05 | 0,12 | 0,14 | 0,18 | -0,41 | 0,70 | 0,88 | 1,08 | 16,20 | < 0.001* | 0,19 |
| M-PFC | -0,07 | 0,16 | 0,18 | 0,22 | -0,37 | 0,72 | 0,95 | 1,10 | 19,10 | < 0.001* | 0,23 | |
| L-PFC | -0,08 | 0,17 | 0,15 | 0,21 | -0,45 | 0,80 | 0,87 | 1,02 | 22,71 | < 0.001* | 0,26 | |
Covariates included gender and age. *Indicates a significant difference between mean Z-scores at rest and during exercise (P < .05). R-PFC: right prefrontal cortex, M-PFC: middle prefrontal cortex, L-PFC: left prefrontal cortex, SD: standard deviation, oxy-Hb: oxygenated hemoglobin.
Table 5.
Differences in oxy-Hb Z-score changes between Pattern 1 and Pattern 2.
|
Event (ID, Name) |
Pattern 1 | Pattern 2 |
F |
P |
|||
| Region | Mean | SD | Mean | SD | |||
| A Upward Stretch | R-PFC | 0,20 | 1,32 | 0,62 | 1,30 | 0,78 | 0.381 |
| M-PFC | 0,48 | 1,29 | 0,41 | 1,48 | 0,12 | 0.731 | |
| L-PFC | 0,28 | 1,64 | 0,31 | 0,84 | 0,15 | 0.699 | |
| B Shoulder Stretch | R-PFC | 0,07 | 1,40 | 0,16 | 1,10 | 0,18 | 0.672 |
| M-PFC | 0,12 | 1,28 | 0,34 | 1,53 | 0,09 | 0.762 | |
| L-PFC | 0,22 | 1,30 | 0,36 | 1,48 | 0,27 | 0.601 | |
| C Elbow Circles | R-PFC | 0,31 | 0,97 | 0,12 | 1,40 | 0,30 | 0.587 |
| M-PFC | 0,98 | 1,02 | 1,07 | 1,36 | 0,42 | 0.519 | |
| L-PFC | 0,82 | 1,16 | 1,03 | 1,50 | 0,53 | 0.469 | |
| D Trunk Twist | R-PFC | 0,18 | 1,50 | 0,62 | 1,38 | 0,51 | 0.479 |
| M-PFC | 0,52 | 1,69 | 0,86 | 1,32 | 0,36 | 0.549 | |
| L-PFC | 0,55 | 1,28 | 0,94 | 1,18 | 0,66 | 0.422 | |
| E Washing Hands | R-PFC | 0,22 | 1,46 | 0,60 | 1,38 | 0,33 | 0.564 |
| M-PFC | 0,45 | 1,52 | 0,72 | 1,24 | 0,20 | 0.656 | |
| L-PFC | 0,66 | 1,32 | 0,78 | 1,40 | 0,18 | 0.678 | |
| F Thumb & Pinky | R-PFC | 0,56 | 1,65 | 0,73 | 1,25 | 0,08 | 0.778 |
| M-PFC | 0,92 | 1,14 | 0,88 | 1,06 | 0,14 | 0.707 | |
| L-PFC | 0,75 | 1,20 | 0,90 | 1,14 | 0,11 | 0.741 | |
| G Single Leg Balance | R-PFC | 0,72 | 1,29 | 0,88 | 1,19 | 0,19 | 0.662 |
| M-PFC | 0,82 | 1,42 | 1,10 | 1,32 | 0,24 | 0.626 | |
| L-PFC | 0,85 | 1,25 | 1,12 | 1,30 | 0,17 | 0.681 | |
Covariates included gender, age, and oxy-Hb Z-scores at rest. R-PFC: right prefrontal cortex, M-PFC: middle prefrontal cortex, L-PFC: left prefrontal cortex, SD: standard deviation, oxy-Hb: oxygenated hemoglobin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



