
Submitted:
24 January 2025
Posted:
24 January 2025
You are already at the latest version
Abstract
Chestnut trees are an essential source of both food and timber. However, the severe threats from invasive pests and diseases compromise their existence and productivity. In Europe, chestnut hybridization programs have been initiated to produce resilient rootstocks in response to ink disease. However, there remains a gap in the identification of these hybrid plants. Our study presents a methodology for distinguishing between chestnut hybrids and varieties using microsatellite (SSR) markers and bioinformatics tools. This tool is essential for identifying CA90 (Castanea sativa × Castanea crenata), a hybrid with demonstrated resistance to Phytophthora cinnamomi. We used 35 chestnut samples, including three CA90 controls, hybrids sampled within Portugal. Nine modified distinct SSR primers from the already developed 43 simple sequence repeat (SSR) markers from expressed sequence tags (ESTs). PCR amplification and agarose gel electrophoresis were used to visualize the DNA bands, and the PCR products were sequenced to confirm genetic variations. We aimed to define the SSR motifs in the chestnut varieties used in this study to improve the use of chestnut hybrids. Twenty-seven amplified bands were sequenced by Sanger sequencing. This analysis identified 31 SSRs across 22 SSR-containing sequences, with trinucleotide (67.74%) repeats being the most common, (22.58%) dinucleotide, (6.45%) mononucleotide, and hexanucleotide (3.23%). A total of 18 alleles were observed for the nine loci. The alleles ranged from 1 to 3 per locus for the 35 samples. It was also observed that primer locus CP4 could only be found in CA90 hybrids. This tool can aid in identifying and selecting disease-resistant hybrids, thereby contributing to chestnut production and management strategies.
Keywords:
Chestnut hybrids
; Castanea sativa
; Microsatellites Markers (SSR)
; CA90
1. Introduction
The chestnut tree (Castanea: Fagaceae) has been valued for its economic, cultural, and ecological significance in Europe, Asia, and North America. Their family includes other important tree species, such as oaks (Quercus spp.) [1]. In Portugal, chestnut trees (mainly Castanea sativa) have significant cultural and economic importance. The regions of Trás-os-Montes and Beiras are known for their extensive chestnut orchards and high-quality production, accounting for more than 80% of production (34,000 to 38,000 tons) [2], particularly varieties like “Martainha,” “Longal,” and “Judia.” Chestnuts are celebrated annually at the Magusto festival in early November, where locals gather to roast and celebrate the harvest [3,4].
Chestnut trees (Castanea spp.) are susceptible to invasive pests and diseases, which can seriously jeopardize their existence and productivity due to their genetic adaptation to different environmental conditions. Chestnut ecosystems are currently threatened by different stress factors (natural or anthropogenic), such as climate change, abandonment of traditional orchards, wildfire, and an increased incidence of pests and diseases [5]. The pathogens of Ink disease, the oomycetes Phytophthora cinnamomi and Phytophthora cambivora, and the chestnut light fungi Cryphonectria parasitica are the most severe chestnut disease pathogens [6,7]. At the end of the nineteenth century, ink disease drastically threatened European chestnuts. The first record of this disease in Portugal was in 1838, although it is believed to have been present in Spain since 1726 [8]. The repercussions of biotic and abiotic stress factors, predictions on implications of climate change on pathogens shift distribution, coupled with the consequent manifestation of distinct disease phenotypes, led to the development of novel chestnut hybrids that are better suited to prevailing climatic conditions and biotic stress [9].
To counter these severe diseases, mitigating research methodologies have been diligently employed, ultimately culminating in identifying a promising approach: developing hybridized plant varieties fortified with resistance mechanisms [7,10]. Hybrid chestnuts derived from Asian and European sources have proven notably resistant to ink diseases [11]. Since the 1950s, breeding programs with European and Japanese chestnuts (C. sativa × C. crenata) have been established in Portugal, France, and Spain to obtain hybrids that are tolerant to ink disease while maintaining fruit production and quality traits to satisfy commercial demands [12,13]. More common control approaches are correctly managing nurseries/orchards, using resistant rootstocks for propagation, or planting resistant hybrids for production [14]. However, clear strategies for distinguishing and maintaining established hybrid plants are yet to emerge, resulting in farmers inadvertently cultivating non-hybrid varieties. The selection of cultivars is an essential step in establishing and maintaining orchards. Helping nurseries and growers maintain good genetic records will help answer questions about specific cultivars’ performance. In the Mediterranean countries, Chestnut cultivar identification and genetic resources assessment are significant issues, such as Italy [15], Spain [16], and Portugal [4,17].
In Portugal, chestnut genetic diversity was analyzed through a preliminary study using single sequence repeat (SSR), and it was successfully used to type traditional Portuguese chestnut varieties [4]. They suggested low genetic variability among the Portuguese chestnut varieties. [18] revealed that C. crenata and C. mollissima have been the foundation of the Portuguese chestnut breeding programs. Another breeding program, more recently by [19], focused on the development of C. sativa × C. crenata and C. sativa × C. mollissima hybrids, where Four F1 hybrids were chosen for extensive propagation according to their potential to proliferate and root in vitro, ink disease resistance levels and field development [20,21]. A clonal collection of Japanese chestnut hybrids was crossed with European chestnut (C. sativa) to produce nuts, timber, and rootstocks for local cultivars [11]. Moreover, a group of hybrid genotypes selected as the most resistant to P. cinnamomi is being propagated as improved genetic materials for new rootstocks released to the market [12].
Chestnut varietal characterization has been carried out in recent decades using different approaches, and more recently, the development of DNA markers has provided a direct study of genotypes that enable the identification of chestnut cultivars [22]. In the last 20 years, microsatellite markers (SSRs) consisting of tandem repeats of polymorphic short-sequence motifs (2-8 bp), Co-dominant, inherited, highly polymorphic, and reproducible are the most suitable to detect genetic variability [11,23].
In Portugal, non-hybrid chestnut plants are cheap, whereas hybrids such as “ferosacre” (CA90) command a higher price. However, in certain instances, rootstock vendors may offer Castanea sativa plants while asserting that they are hybrid cultivars. To date, some farmers still depend on traditional methods of hybrid identification, such as morphological characteristics and field observations. However, these methods have limitations in distinguishing closely related individuals from genetically similar hybrids [24]. Molecular techniques, particularly microsatellite analysis, have revolutionized the field of hybrid identification by providing a precise means of differentiating between individuals based on their genetic profiles. Meanwhile, more attention has been paid to genetic diversity and not to hybrid identification tools that enable farmers to plant resistant varieties. Our study was designed to fight plant diseases and find answers to cultivar identification challenges. We aimed to develop a molecular diagnostic tool through SSR to differentiate between chestnut hybrids and non-hybrid plants. This study is essential because the chestnut industry lacks certification and validation programs for cultivars. Genetic marker technology can play a significant role in helping the industry maintain genetic standards.
2. Material And Methods
2.1. Plant Materials and DNA Extraction
A total of 35 samples consisting of hybrids and traditional Portuguese varieties were collected from chestnut trees within Portugal: 12 from Sta Comba, 10 from Vila-Boa de Serapicos, and 13 from Deifil Green Biotechnology, a research institution specializing in in vitro plant propagation for species with inefficient natural reproduction systems. Error! Reference source not found. provides a detailed description of each accession. Three (3) samples were positive controls, CA90 hybrids (C. sativa × C. crenata), with well-defined genetic profiles among Sta Comba samples. After sample collection, the leaves were cleaned with 70% ethanol and stored at -20°C until DNA extraction.

Table 1.
Details of the 35 Chestnut Samples, Including Laboratory Designated Codes.
| Lab code | Samples | GPS / Field Tag | Sampling location |
|---|---|---|---|
| A | CA90 positive control | M111111,2 P222973,9 | Sta Comba |
| B | CA90 positive control | M110916,0 P2225509 | Sta Comba |
| C | CA90 positive control | M110932,6 P222509,7 | Sta Comba |
| D | C. Sativa | TD | Vila-Boa de Serapicos |
| 1 | CA90 | T1 | Vila-Boa de Serapicos |
| 2 | CA90 | T2 | Vila-Boa de Serapicos |
| 3 | CA90 | T3 | Vila-Boa de Serapicos |
| 4 | CA90 | T4 | Vila-Boa de Serapicos |
| 5 | CA90 | T5 | Vila-Boa de Serapicos |
| 6 | CA90 | T6 | Vila-Boa de Serapicos |
| 7 | CA90 | T7 | Vila-Boa de Serapicos |
| 8 | CA90 | T8 | Vila-Boa de Serapicos |
| 9 | CA90 | T9 | Vila-Boa de Serapicos |
| 10 | C. mollissima 60907 | T10 | Deifil Green Biotechnology |
| 11 | C. mollissima E2604 | T11 | Deifil Green Biotechnology |
| 12 | C. mollissima Y0204 | T12 | Deifil Green Biotechnology |
| 13 | C. mollissima Z1408 | T13 | Deifil Green Biotechnology |
| 14 | CA90 | T14 | Deifil Green Biotechnology |
| 15 | Martaínha 2 | T15 | Deifil Green Biotechnology |
| 16 | Bouche de Betizac 1 | T16 | Deifil Green Biotechnology |
| 17 | Bouche de Betizac 2 | T17 | Deifil Green Biotechnology |
| 18 | Martaínha 1 | T18 | Deifil Green Biotechnology |
| 19 | Cota | T19 | Deifil Green Biotechnology |
| 20 | Judia | T20 | Deifil Green Biotechnology |
| 21 | Marsol | T21 | Deifil Green Biotechnology |
| 22 | Précoce Migoule | T22 | Deifil Green Biotechnology |
| 23 | Putative CA90 | M111478,8 P223081,2 | Sta Comba |
| 24 | Putative CA90 | M11609,9 P222967,4 | Sta Comba |
| 25 | Putative CA90 | M111649,4 P222888,7 | Sta Comba |
| 26 | Putative CA90 | M111494,7 P222815,0 | Sta Comba |
| 27 | Putative CA90 | M111497,6 P223077,5 | Sta Comba |
| 28 | Putative CA90 | M111191,6 P222934,5 | Sta Comba |
| 29 | Putative CA90 | M111512,1 P222808,6 | Sta Comba |
| 30 | Putative CA90 | M111486,4 P222934,5 | Sta Comba |
| 31 | Putative CA90 | M111477,6 P222821,3 | Sta Comba |
In the laboratory, all harvested leaves were deprived of veins and reduced to a fine powder in liquid nitrogen using a mortar and pestle. Genomic DNAs were extracted from 0.05 g of leaves using a Thermo Scientific GeneJET Plant Genomic DNA Purification Mini Kit following the manufacturer’s protocol.
2.2. SSR Amplification
In order to define a set of microsatellite sequences useful for the fingerprinting and mapping of chestnut, nine (9) SSR primer pairs (Error! Reference source not found.) obtained from the National Institute for Agricultural and Veterinary Research (INIAV) Portugal were used to define the fingerprints of the Castanea hybrids and varieties in this study. However, we noticed post-experimentation that the primer sets were modified from the developed 43 simple sequence repeat (SSR) markers from expressed sequence tags (ESTs) by [21]. Ten (10) primers were selected and modified from that study. The primers loci CcPT 0002 and CcPT 0003 were maintained but renamed CP2 and CP3 according to this study (Error! Reference source not found.). The modifications affected CP4 to CP11, including adding one or two nucleotides to the 3’ end of the forward and reverse primers. This may have influenced the binding specificity or efficiency of the primers. However, we had some exciting outcomes.
Target loci for this study were amplified by performing a PCR reaction in a final volume of 50μl containing 0.2 mM dNTPs, 1X reaction buffer, 0.8X Kcl, 1.5 mM Mgcl2, 2.5 units DFS-DNA polymerase, 2.5 μM each of Forward and Reverse primers, 4 μg genomic DNA (20 ng/μl), 0.6 mg/ml BSA, and 29.25μl deionized water. The following thermocycling conditions were used: initial denaturation, final denaturation, annealing, extension, and final extension at 94 ◦ for C-2 min; 94 ◦ for C-10 s, 57 ◦ for C-20 s, and 72 ◦C-30 s for 36 cycles and 72 °C for 5 min. The internal temperature of the thermocycler was adjusted to 4°C for the optimal maintenance of amplified DNA fragments. A summary of all the primers used is shown (Table 2). The PCR programme was based on [25] with little modifications. After amplification, the fragments were separated using agarose gel electrophoresis. The PCR products were labeled with ethidium bromide, run at 80 Volts, and visualized using the Bio-Rad gel DocTM Ez Imager (Image Lab application 5.2.1).
2.3. Sequencing
Twenty-seven DNA amplicons were randomly selected from different sets of nine primers and chestnut samples, purified using a DNA Clean & Concentrator kit from ZYMO Research, quantified with a nanodrop to assess DNA concentration and purity following the Beer-Lambert Law, and sent to the University of Salamanca for Sanger sequencing.
2.4. Bioinformatic Analysis
Briefly, (i) BioEdit was used to manipulate, analyze, and visualize the raw obtained biological sequences. (ii) BLASTn NCBI for sequence comparison with sequences published in the GenBank, (iii) Molecular Evolutionary Genetics Analysis (MEGA11) and ClustalW combined feature for sequence alignments and phylogenetic analyses, and (iv) MISA-web for detecting perfect microsatellites.
3. Results and Discussion
3.1. Effects on Primer Locus Modification
The modifications of the primers affected the results of this study. Our primer set consistently amplified different regions and could be considered a new variant or loci. We named them locus (CP2, CP3, CP4, CP5, CP6, CP8, CP9, CP10, CP11). The results compared with [21], from which the primers were modified, showed differences regarding SSR motifs, Number of alleles, and allelic size, except for CP2, which was not modified. See (Error! Reference source not found.).
3.2. Locus Amplifications and Band Pattern Representation
In this study, all primer sets gave good amplifications at all loci, depicting monomorphic bands on gel and high polymorphisms when sequenced, except for CP8, which showed polymorphic bands. A heatmap (Error! Reference source not found.) was created based on the visual representation of band appearance patterns from the gel images to create an effective identification tool and select the most effective primer pairs that differentiate hybrids and varieties.
Figure 1.
A Heatmap of Band Patterns from Gel Images. (ABC)-CA90 controls, (D)-C. sativa, (1-9) - CA90, (10) - C. mollissima 60907, (11) -C. mollissima E2604, (12) - C. mollissima a Y0204, (13) - C. mollissima Z1408, (14) - CA90-deifil, (15) -Martainha 2, (16) -Bouche de Betizac 1, (17) -Bouche de Betizac 2, (18) - Martainha 1, (19) -Cota, (20) -Judia, (21) -Marsol, (22) -Precoce migoule, (23-31) -Putative CA90.
Figure 1.
A Heatmap of Band Patterns from Gel Images. (ABC)-CA90 controls, (D)-C. sativa, (1-9) - CA90, (10) - C. mollissima 60907, (11) -C. mollissima E2604, (12) - C. mollissima a Y0204, (13) - C. mollissima Z1408, (14) - CA90-deifil, (15) -Martainha 2, (16) -Bouche de Betizac 1, (17) -Bouche de Betizac 2, (18) - Martainha 1, (19) -Cota, (20) -Judia, (21) -Marsol, (22) -Precoce migoule, (23-31) -Putative CA90.
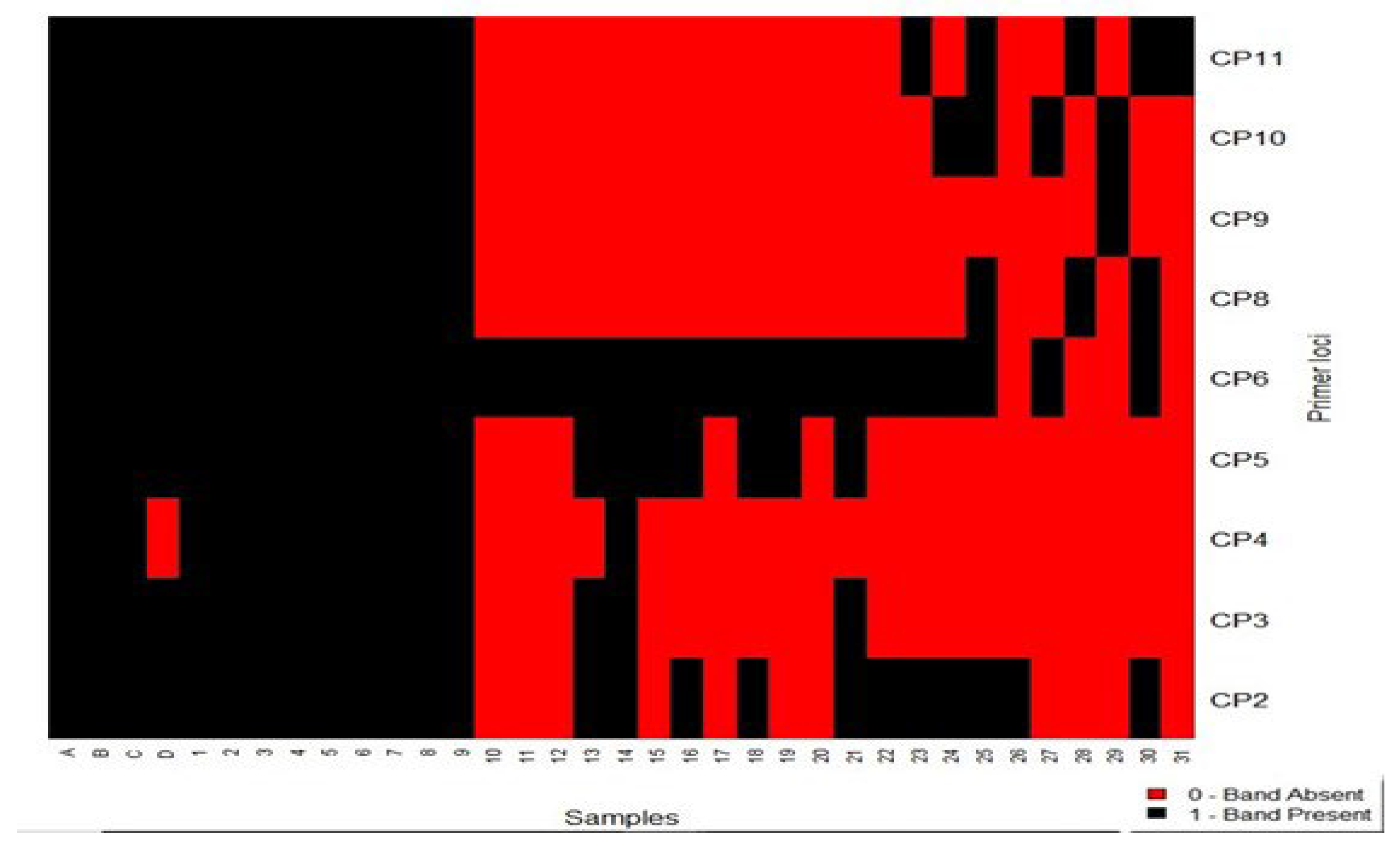
It was evident from the heatmap that samples (1–9) from Vila-Boa de Serapicos collectively exhibited a remarkable 100% genetic identity with the three controls in all nine primer loci, confirming them as CA90 hybrids. In stark contrast, Sample (14), identified as (CA90) from Deifil Green Biotechnology, displayed a questionable genetic match of 55.6% and could not be identified as CA90. The Sta Comba samples (23-31) could not be identified as CA90 since their genetic status did not match the controls. Additionally, sample (D), recognized as “C. sativa,” exhibited a striking 88.9% genetic resemblance to the controls in all primer loci. This high genetic match with the controls supports CA90 as a natural interspecific hybrid derived from (C. crenata × C. sativa), indicating gene inheritance from both parental varieties.
3.3. Sequencing Results
The obtained raw DNA sequence reads were subjected to BioEdit (version 7.2.5) for quality control assessment. The resulting sequences were then submitted to ClustalW for sequence alignment and finally to MISA-web following the procedures of [26] for microsatellite prediction (Table 3) to confirm the results of the gel images. The 27 examined sequences collectively comprised 8899 base pairs. Among these, 31 SSRs were identified and distributed across 22 sequences containing SSRs, nine sequences contained more than one SSR, and six SSRs were found to be involved in compound formation. The search parameters for SSRs were set, specifying a motif length ranging from 1 to 6, a repetition minimum of 5, and no base pairs between the two microsatellites for compound SSR detection. The analysis revealed that the majority (67.74%) of the identified microsatellites were of the trinucleotide type, with dinucleotide (22.58%), mononucleotide (6.45%), and hexanucleotide (3.23%) microsatellites also present, albeit in smaller proportions. A total of 18 alleles were obtained for the nine loci. The alleles ranged from 1 to 3 per locus for the 35 samples. Our results confirm the low genetic variability of Portuguese chestnut varieties, which aligns with the studies (Costa et al., 2005). The range of variability makes it possible to select loci for specific applications. For example, markers with only a few alleles are well-suited for population genetic studies [27]. We also observed some intra-varietal differences within the studied accessions; for example, CA90 has been characterized by the same SSR motifs but different numbers of repeats. This aligns with the results [28]. They used SSR markers to assess the heterogeneity of the genotype of the cultivar “Judia” in Trás-os-Montes.
While the gel images in this study showed monomorphic bands, the sequences from all primer loci were characterized by distinct motifs, as detailed in (Error! Reference source not found.), proving all nine primer pairs are highly polymorphic. This was attributed to the samples’ homoplasy, allelic variation, and inherent genetic diversity. Sequences from different samples at locus CP4, using forward primer (5’-GCTGCTTCACAACCTTCCTC-3’) and reverse primer (5’-GCAAGAGATTCCCTTTGCTG-3’) consistently exhibited identical SSR motifs [(GA)10 and (GA)11] and variations in alleles. This validation underscores the success of our objective: establishing a robust tool for differentiating chestnut hybrids. The sequences, when compared with sequences published in the GenBank database using the BLAST algorithm, showed nucleotide similarity greater than 95%, belonging to Quercus Lobata, Quercus variabilis, Quercus suber, and Quercus rober, confirming the lack of DNA sequence data for Castanea, which supports the view that most SSRs currently available are developed from repeat-enriched genomic DNA libraries from Quercus sequences being helpful in Castanea [29,30].
3.4. Phylogenetic Tree Analysis
An evolutionary history was inferred with a phylogenetic tree analysis (Figure 2) using the Maximum Likelihood method and the Tamura-Nei model [31] with a bootstrap consensus tree inferred from 100 and BioNJ algorithms to a matrix of estimated pairwise distances using the Tamura-Nei model. The analysis involved all the 27 nucleotide sequences. There were 613 positions in the final dataset. The evolutionary analyses were conducted using the MEGA11 software [31]. The phylogenetic tree analysis validated the results obtained by gel electrophoresis and sequencing. These results reaffirmed the close genetic relatedness among samples with bands for primers CP2, CP4, CP6, CP9, and CP10 loci while highlighting considerable genetic diversity among hybrids with bootstrap values above 95%. The sequences from these loci were considered, suggesting the robustness of the inferred relationship. However, there were differences in the leaf figures, except for CP4, implying that the taxa represented by the leaf nodes were distinct from each other in some way, either genetically, morphologically, or ecologically. The phylogenetic tree confirmed that C. mollissima Z1408 (Chinese hybrid) and Marsol shared genetic relationships with C. sativa and CA90.
4. Conclusions
From the gel image analysis on the heatmap, Locus CP4 emerged as the most significant locus in this investigation, showing bands exclusively in the CA90 samples. The absence of bands in C. sativa at locus CP4 and the presence in all loci ultimately confirm this loci’s significance in differentiating this cultivar. Therefore, for time and cost management, the primer loci CP2, CP4, CP6, CP9, and CP10 could serve as effective primer combinations for differentiating CA90 hybrids and C. Sativa since they gave good amplifications and clustering specificity according to the phylogenetic tree.
Molecular marker-based characterization is faster, cheaper, and more reliable than traditional morphological classification, although both methods have complementary values. To enhance the effectiveness of SSR markers for chestnut identification, it would be beneficial to evaluate markers designed for species closely related to Castanea and those within the Fagaceae family. Implementing these technologies in Portugal’s chestnut industry could bring several advantages: (1) certification of cultivars and germplasm; (2) precise identification of species, cultivars, and rootstocks; and (3) improved screening of selection in breeding programs. This information will aid in identifying pollinators that influence fruit set, yield, size, and quality. Further research is recommended to expand the SSR marker set for enhanced accuracy across various chestnut species because of the lack of certification and validation programs for cultivars.
Author Contributions
Conceptualization, A.C. and T.S.Y.; Funding Acquisition, E.G.; Investigation, T.S.Y. and A.C..; Methodology, T.S.Y., A.C., and N.E.; Visualization, S.T.Y.; Writing—Original Draft, S.T.Y.; Writing—Review and Editing, A.C., E.G., and V.C. All authors have read and agreed to the version of the manuscript for publication.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Compliance with Ethical Standards
This article does not contain any studies with human or animal subjects performed by any of the authors.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Clark, S.L.; Marcolin, E.; Patrício, M.S.; Loewe-Muñoz, V. A Silvicultural Synthesis of Sweet (Castanea sativa) and American (C. dentata) Chestnuts. For Ecol Manag. 2023, 539, 121041. [CrossRef]
- Ribeiro, M.; Freitas, M.; Domínguez-Perles, R.; Barros, A.I.R.N.A.; Ferreira-Cardoso, J.; Igrejas, G. Nutriproteomics Survey of Sweet Chestnut (Castanea sativa Miller) Genetic Resources in Portugal. Food Biosci. 2020, 36, 100622. [CrossRef]
- Braga, M.; Araújo, S. de S.; Sales, H.; Pontes, R.; Nunes, J. Portuguese Castanea sativa Genetic Resources: Characterization, Productive Challenges and Breeding Efforts. Agriculture (Switzerland). 2023, 13, 1629.
- Costa, Rita.; Valdiviesso, Teresa.; Marum, Liliana.; Borges, Olga.; Soeiro, José.; Assunção, Augusto.; Soares, F.Matos.; Sequeira, José. Characterisation of Traditional Portuguese Chestnut Cultivars by Nuclear SSRs. Acta Horticulturae. 2005, 693, 437–440. [CrossRef]
- Freitas, T.R.; Santos, J.A.; Silva, A.P.; Fraga, H. Influence of Climate Change on Chestnut Trees: A Review. Plants. 2021, 10, 1463. [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria Parasitica, the Causal Agent of Chestnut Blight: Invasion History, Population Biology and Disease Control. Mol Plant Pathol 2018, 19, 7–20. [CrossRef]
- Anagnostakis, S.L. Chestnut Breeding in the United States for Disease Insect Resistance. Plant Dis. 2012, 96, 1392–1403. [CrossRef]
- Vannini, A.; Vettraino, A.M. Ink Disease in Chestnuts: Impact on the European Chestnut. For Snow Landsc. 2001, 76, 345-350.
- Burgess, T.I.; Scott, J.K.; Mcdougall, K.L.; Stukely, M.J.C.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and Projected Global Distribution of Phytophthora cinnamomi, One of the World’s Worst Plant Pathogens. Glob Chang Biol. 2017, 23, 1661–1674.
- Anagnostakis, S.L. Chestnut Blight: The Classical Problem of an Introduced Pathogen1. Mycologia 1987, 79, 23.
- Pereira-Lorenzo, S.; Lourenço Costa, R.; Serdar Ondokuz, U.; Üniversitesi, M.; Ramos-Cabrer, A. Interspecific Hybridization of Chestnut. Polyploidy and Hybridization for Crop Improvement, 1st ed; Annaliese S. M, CRC press, 2016, pp.379-408.
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First Interspecific Genetic Linkage Map for Castanea sativa x Castanea crenata Revealed QTLs for Resistance to Phytophthora cinnamomi. PLoS One. 2017, 12, e0184381. [CrossRef]
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea Root Transcriptome in Response to Phytophthora cinnamomi Challenge. Tree Genet Genomes. 2015, 11. [CrossRef]
- Hardham, A.R. Phytophthora cinnamomi. Mol Plant Pathol 2005, 6,589-604.
- Boccacci, P.; Akkak, A.; Torello, D.; Bounous, G.; Botta, R. Typing European Chestnut (Castanea sativa Mill.) Cultivars Using Oak Simple Sequence Repeat Markers. HortSci. 2004, 39, 1212-1216. [CrossRef]
- Ramos-Cabrer, A.M.; Pereira-Lorenzo, S. Genetic Relationship between Castanea sativa Mill. Trees from North-Western to South Spain Based on Morphological Traits and Isoenzymes. Genet Resour Crop Evol. 2005, 52, 879–890. [CrossRef]
- Goulao, L.; Valdiviesso, T.; Santana, C.; Oliveira, C.M. Comparison between Phenetic Characterisation Using RAPD and ISSR Markers and Phenotypic Data of Cultivated Chestnut (Castanea Sativa Mill). Genetic Resources and Crop Evolution. 2001, 48, 329-338. [CrossRef]
- Santos, C.; Machado, H.; Serrazina, S.; Gomes, F.; Gomes-Laranjo, J.; Correia, I.; Zhebentyayeva, T.; Duarte, S.; Bragança, H.; Fevereiro, P.; et al. Comprehension of Resistance to Diseases in Chestnut. Revista de Ciências Agrárias. 2016, 39, 189–193. [CrossRef]
- Costa, R.; Santos, C.; Tavares, F.; Machado, H.; Gomes-Laranjo, J.; Kubisiak, T.; Nelson, C.D. Mapping and Transcriptomic Approches Implemented for Understanding Disease Resistance to Phytophthora cinammomi in Castanea Sp. BMC Proc. 2011, 5. [CrossRef]
- Fernandes, P.; Tedesco, S.; da Silva, I.V.; Santos, C.; Machado, H.; Costa, R.L. A New Clonal Propagation Protocol Develops Quality Root Systems in Chestnut. Forests. 2020, 11, 826. [CrossRef]
- Santos, C.; Zhebentyayeva, T.; Serrazina, S.; Nelson, C.D.; Costa, R. Development and Characterization of EST-SSR Markers for Mapping Reaction to Phytophthora cinnamomi in Castanea spp. Sci Hortic. 2015, 194. [CrossRef]
- Martin, M.A.; Alvarez, J.B.; Mattioni, C.; Cherubini, M.; Villani, F.; Martin, L.M. Identification and Characterisation of Traditional Chestnut Varieties of Southern Spain Using Morphological and Simple Sequence Repeat (SSRs) Markers. Annals of Applied Biology .2009, 154, 389–398. [CrossRef]
- Rasoarahona, R.; Wattanadilokchatkun, P.; Panthum, T.; Thong, T.; Singchat, W.; Ahmad, S.F.; Chaiyes, A.; Han, K.; Kraichak, E.; Muangmai, N.; et al. Optimizing Microsatellite Marker Panels for Genetic Diversity and Population Genetic Studies: An Ant Colony Algorithm Approach with Polymorphic Information Content. Biology (Basel). 2023, 12, 1280. [CrossRef]
- Marinoni, D.; Akkak, A.; Bounous, G.; Edwards, K.J.; Botta, R. Development and Characterization of Microsatellite Markers in Castanea sativa (Mill.). Molecular Breeding. 2003, 11, 127-136. [CrossRef]
- Botta, R.; Marinoni, D.; Beccaro, G.; Akkak, A.; Bounous, G.; Development of a DNA Typing Technique for the Genetic Certification of Chestnut Cultivars. For snow Landsc. 2001, 76, 425-428.
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-Web: A Web Server for Microsatellite Prediction. Bioinformatics. 2017, 33, 2583–2585. [CrossRef]
- Yang, H.C.; Lin, H.C.; Huang, M.C.; Li, L.H.; Pan, W.H.; Wu, J.Y.; Chen, Y.T. A New Analysis Tool for Individual-Level Allele Frequency for Genomic Studies. BMC Genomics. 2010, 11, 415. [CrossRef]
- Dinis, L.-T.J.; Peixoto, F.; Costa, R.; Gomes-Laranjo, J. Molecular Characterization of “Judia” (Castanea sativa Mill.) from Several Trás-Os-Montes Regions by Nuclear Microsatellite Markers. Acta Horticul. 2010, 866, 225-232.
- Bodénès, C.; Chancerel, E.; Gailing, O.; Vendramin, G. G.; Bagnoli, F.; Durand, J. Comparative mapping in the Fagaceae and beyond with EST-SSRs. BMC plant biology. 2012, 12, 1-18. [CrossRef]
- Barreneche, T.; Casasoli, M.; Russell, K.; Akkak, A.; Meddour, H.; Plomion, C.; Villani, F.; Kremer, A. Comparative Mapping between Quercus and Castanea Using Simple-Sequence Repeats (SSRs). Theoretical and Applied Genetics 2004, 108, 558–566. [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. 1993, Molecular Bio and Evol. [CrossRef]
Figure 2.
Bootstraps consensus Phylogenetic tree Showing Similarities Between Sequences from Primer Loci.
Figure 2.
Bootstraps consensus Phylogenetic tree Showing Similarities Between Sequences from Primer Loci.
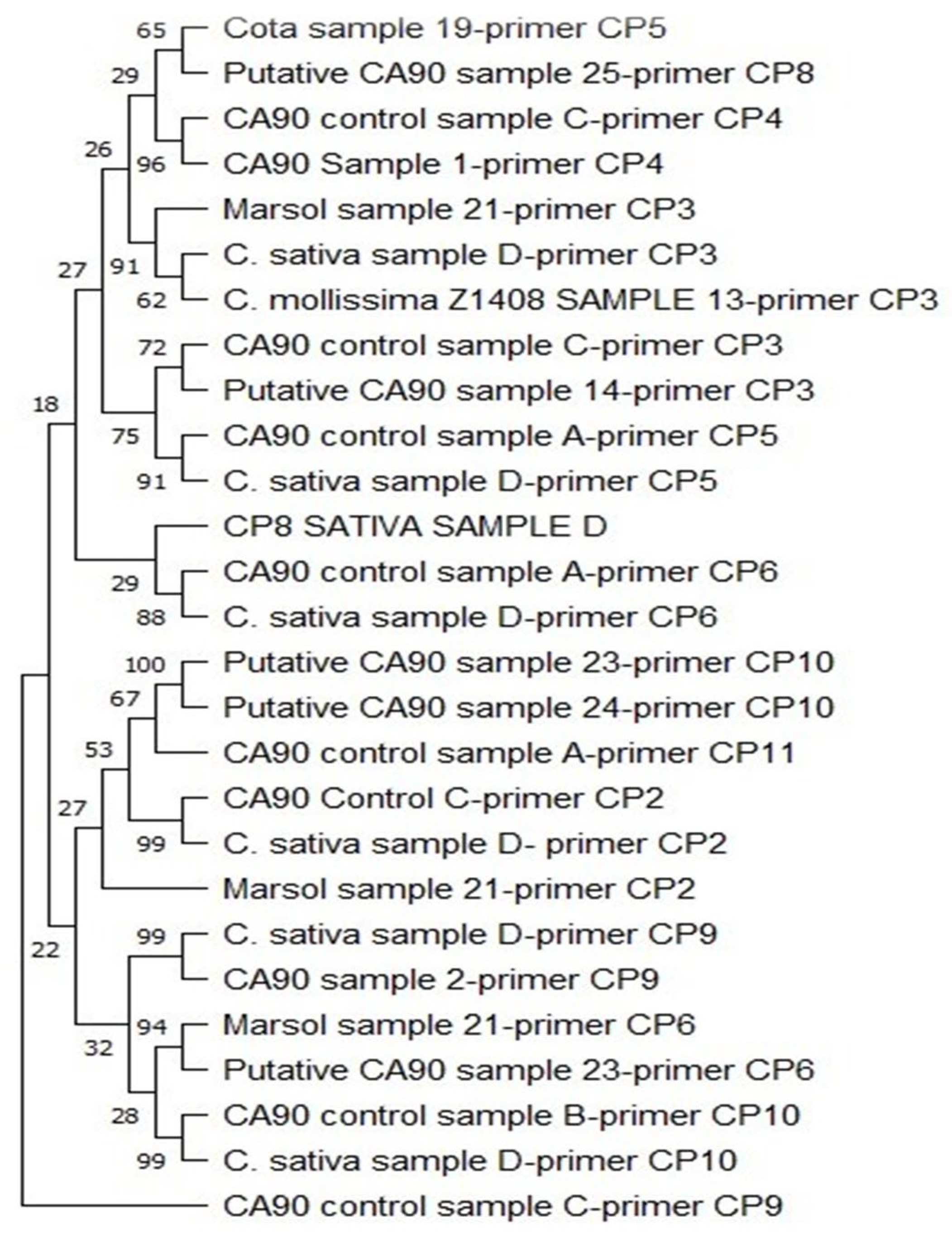
Table 2.
Primer Sequences Used for SSR Analysis and Their Characteristics.
| NO | loci | Forward (5’-3’) | Reverse (5’-3’) | Size (bp) | Annealing Tm (℃) |
|---|---|---|---|---|---|
| 1 | CP2 | AGTTCTCCACGAGGCTCAAA | TCCAAGCTGGAGAATCATCA | 220 | 55.3 |
| 2 | CP3 | GGTGCCCAGATTTACGAGAA | ATCGCTTGGAGTCACAGCTT | 240 | 57.3 |
| 3 | CP4 | GCTGCTTCACAACCTTCCTC | GCAAGAGATTCCCTTTGCTG | 220 | 57.3 |
| 4 | CP5 | ACACATGGGGGTGTGAACTT | TTATGGGAAACGGCATCTTC | 125 | 55.3 |
| 5 | CP6 | CCTGTGAGGCTAAGAGAGCG | ACCACGTCGGTGCTTCTAGT | 200 | 59.4 |
| 6 | CP8 | TCGTCCCCTTCTTCATCATC | ATATGGCCAAAAACCCATCA | 250 | 53.2 |
| 7 | CP9 | TTCCACCCAATTGTTACCAC | GATGAAGAAGGGGACGA | 200 | 55.3 |
| 8 | CP10 | ATCCATGAGTGAAAGCCACC | TGGAACAAGAAGCCTCGATT | 250 | 55.6 |
| 9 | CP11 | TCATCCAAGAAGCCCTCAAC | TTCTGCCTCTTTTGTTGCCT | 230 | 55.3 |
Table 3.
Comparison of results for the modified SSR primer loci: number of alleles (Na), *-No motifs found.
Table 3.
Comparison of results for the modified SSR primer loci: number of alleles (Na), *-No motifs found.
| Loci | Sample | Na | Motif | Reference | Loci | Na | Motif | Reference |
|---|---|---|---|---|---|---|---|---|
| Cp2 | Marsol | 1 | (GTG)9 | This study | CcPT 0002 | 1 | (GTG)9 | [21]. |
| Cp2 | CA90 | * | This study | |||||
| Cp2 | C. sativa | * | This study | |||||
| Cp3 | Marsol | 3 | (ATC)6 | This study | CcPT 0003 | 7 | (ATC)8 | [21]. |
| Cp3 | C. sativa | (ATC)6 | This study | |||||
| Cp3 | C. mollissima. Z1408 | (ATC)6 | This study | |||||
| Cp3 | CA90 | (TGA)5tcatgatgaccacaaggattgaagttagtcacagcatctcggccaccaacgcgttgggccgcgatgtcgtacgcttttgca (TCT)5 | This study | |||||
| Cp3 | Martainha | (TGA)8tcatcagtaccacccggcgggttgaagttcacaacatcttctccaccacaaaagcggggggccatgatgtcatattttgttg (CTT)5 | This study | |||||
| Cp4 | CA90 | 2 | (GA)10 | This study | CcPT 0004 | 6 | (CT)10 | [21]. |
| Cp4 | CA90 | (GA)11 | This study | |||||
| Cp5 | CA90 | 3 | (CT)8 | This study | CcPT 0005 | 7 | (CT)1 | [21]. |
| Cp5 | C. sativa | (CT)12 | This study | |||||
| Cp5 | COTA | (AG)7 | This study | |||||
| Cp6 | Marsol | 2 | (GAA)5 | This study | CcPT 0006 | 4 | (TTC) | [21]. |
| Cp6 | CA90 | (GAA)5 gaggaagaagaac(A)12 | This study | |||||
| Cp6 | CA90 | * | This study | |||||
| Cp6 | C. sativa | * | This study | |||||
| Cp8 | C. sativa | 2 | (CTCAGA)5 gtacaacaaccgacagca (AAG)11 | This study | CcPT 0008 | 5 | (TCT)1 | [21]. |
| Cp8 | CA90 | (A)12 (TCT)8 | This study | |||||
| Cp9 | CA90 | 2 | (AG)10 | This study | CcPT 0009 | 8 | (TC) | [21]. |
| Cp9 | C. sativa | (TC)8 | This study | |||||
| Cp9 | CA90 | * | This study | |||||
| Cp10 | CA90 | 2 | (CAC)6 (AAG)5 | This study | CcPT 0010 | 5 | (GGT) | [21]. |
| Cp10 | C. sativa | (CAC)6 (AAG)5 | This study | |||||
| Cp10 | CA90 | (GGT)5 gggggagccttc (TCT)6 | This study | |||||
| Cp10 | CA90 | (GGT)5 gggggagccttc (TCT)6 | This study | |||||
| Cp11 | CA90 | 1 | (GGT)7 | This study | CcPT 0011 | 5 | (CAC) | [21]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



