
Submitted:
21 January 2025
Posted:
22 January 2025
You are already at the latest version
Abstract
Maternally inherited endosymbionts are widespread in arthropods, with multiple symbionts commonly co-existing within a single host, potentially competing for or sharing limited host resources and space. Wolbachia and Rickettsia, two maternally-inherited symbionts in arthropods, can co-infect hosts, yet research on their combined impacts on host reproduction and interaction remains scarce. This study used diverse parthenogenetic backcross and antibiotic screening to ex-plore the reproductive effects of these two symbionts on Tetranychus turkestani. Results showed that single Rickettsia infection induced male-killing in the amphigenesis of T. turkestani, leading to ar-rhenotokous embryo death and fewer offsprings. Single Wolbachia infection induced strong cyto-plasmic incompatibility (CI). During dual infection, CI intensity decreased, as Rickettsia's male-killing antagonized Wolbachia - induced CI. Dual-infected mites had increased oviposition, lower mortality, a higher female-to-male ratio, and more offsprings, thus enhancing T. turkestani's fitness. These findings will be helpful to understand the nature of host-endosymbiont interaction and the potential for evolutionary conflicts, offering insights into their co-evolutionary relationship.
Keywords:
Rickettsia
; Wolbachia
; male-killing
; CI
; Tetranychus turkestani
1. Introduction
Endosymbiotic bacteria are common in arthropods, with over 50% of species infected, primarily via maternal inheritance [1,2]. Endosymbiotic bacteria have co-evolved with their hosts, influencing host's nutrition, digestion, resistance, and defense against predators, thereby play critical role in host colonization and ecological evolution in specific habitats [3,4]. At present, the most studied secondary endosymbionts including Wolbachia, Cardinium, Rickettsia, Spiroplasma, are known to manipulate the host reproductive and developmental processes by inducing cytoplasmic incompatibility, male-killing, parthenogenesis, heat resistance and drug resistance in their hosts [5,6,7,8].
Wolbachia is a maternally transmitted gram-negative bacteria found in arthropods. The host range of Wolbachia is extremely wide, and approximately 65% of insect species naturally carry this endosymbiont. Wolbachia is abundantly present in insect ovaries and testes, and is also distributed in non-reproductive tissues such as head, muscles, midgut, salivary gland, Malpighian tubules, hemolymph and fat body of insects [9,10]. The regulatory effects of Wolbachia on its host has always been a hot topic in Wolbachia-related research. Currently, the documented Wolbachia's reproductive regulation of the host include cytoplasmic incompatibility (CI), male-killing, feminization and parthenogenesis etc. CI is the most common reproductive regulation induced by Wolbachia, which refers to the phenomenon where mating between Wolbachia-infected male and uninfected female insect results in either no or few offspring. In addition, some strains of Wolbachia can also affect the host's sense of smell, lifespan, immunity, nutrition, fertility and developmental processes etc [9,10,11,12,13,14,15,16].
Rickettsia is an intracellular symbiotic bacteria that spreads and cause diseases in humans and animals, and is also a secondary endosymbionts existing in insects. Rickettsia, gram-negative bacteria belongs to the family Rickettsiaceae in the α subgroup of Proteobacteria. It is widely distributed in nature, and its hosts include vertebrates, arthropods, annelids, amoebas, ciliates, hydrozoans and plants [7]. Research has found that Rickettsia and its host insects have a mutualistic symbiosis and are co-evolved. Rickettsia can affect the reproductive behavior of their host by inducing male-killing, parthenogenesis, and also has an impact on the fitness of the host insects [8,15,16,17,18].
In nature, co-infection of arthropod hosts by different symbiotic bacteria is quite common. The impacts of multiple infection on the host may be cumulative [19]. The interactions between co-infecting symbiotic bacteria may lead to reproductive phenotypes that are completely different from those seen in singly infected hosts. If co-infection confers a higher fitness than single infection, it can be stably maintained within the host population [20]. Rickettsia and Wolbachia sometimes co-infect arthropods. However, little research has been conducted on the interactions between these two bacteria [2,21,22,23,24], and studies on co-infections have only focused on the expression of cytoplasmic incompatibility, while the impact of co-infection of these two bacteria on host reproduction has not been reported.
Tetranychus turkestani (Ugarov et Nikolski) is an important agricultural pest, which is distributed in Russia, Kazakhstan, United States, Middle East, and in Xinjiang, China [25,26]. This spider mite reproduces rapidly, has a short generation cycle, and is the dominant pest in the cotton fields in northern Xinjiang. Various endosymbiotic bacteria are present in this mite, including Wolbachia, Cardinium, Rickettsia, etc [27]. In this study, we compared the different hybridization types of four different infected strains of Tetranychus turkestani (double-infected Rickettsia and Wolbachia strain IWR, single-infected Rickettsia strain IR, single-infected Wolbachia strain IW and double-uninfected strain IU), and investigated the effects of Wolbachia and Rickettsia on the host and their interaction. These results will further enhance our understanding of the reproductive manipulation induced by the co-infection of symbiotic bacteria in arthropods.
2. Materials and Methods
2.1. Collection and Rearing of Spider Mites
The Tetranychus turkestani were collected in 2019 from the experimental field of the College of Agriculture, Shihezi University in 2019. Since then, they have been reared within a light incubator at the Insect Physiology Laboratory of the College of Agriculture, Shihezi University, under controlled conditions (25 ℃, a photoperiod of 16 hours of light and 8 hours of darkness, and a relative humidity of 60%). The mites were fed on Phaseolus vulgaris L. throughout the rearing process, with no exposure to any pesticides.
2.2. Detection of Infections by Different Symbiotic Bacteria
Extraction of total DNA: 25 μL of STE buffer (100 mmol/L NaCl, 10 mmol/L Tris-HCl, 1 mmol/L EDTA, pH = 8.0) was added to a 1.5 mL centrifuge tube. A single spider mite was picked with an insect needle and placed in the tube, then thoroughly crushed with a plastic pestle. Subsequently, 2 μL of proteinase K (10 mg/mL) was added. The mixture was centrifuged at 3000 r/min for 2 min, incubated at 37 °C for 30 min, then heated at 95 °C for 5 min, and centrifuged again at 3000 rpm for 2 min. 2 μL of the supernatant was used as the template for PCR amplification.
Primers were designed using the Beacon Designer 7 software to detect whether Tetranychus turkestani was infected with Wolbachia and Rickettsia (see Appendix Table A2 and Table A5).
2.3. Establishment of Strains Infected with Different Endosymbiotic Bacteria
Establishment of a strain co-infected with Wolbachia and Rickettsia: A complete and fresh kidney bean leaf was put in a Petri dish with sponge (9 cm diameter), and was divided into four approximately equal chambers using moistened cotton strips according to the leaf size. Unmated female mites were selected in the static III state from the laboratory strain and were placed individually into each chamber for parthenogenesis. When the offspring developed into adult male mites, the mother was backcrossed with her male offspring. After two days of backcrossing, the mother was transferred to a new chamber for oviposition. After seven days, perform PCR was performed to detect the mother. These above steps were repeated for five generations with the offspring of the female mites with co-infection of Wolbachia and Rickettsia, and then 30 of them were selected for PCR detection of the infection rates of Wolbachia and Rickettsia. Once all were infected, a strain co-infected with Wolbachia and Rickettsia was obtained.
The experimental strains with single infection of Rickettsia and single infection of Wolbachia were obtained using the same method.
Establishment of a completely uninfected strain of Tetranychus turkestani: A complete and fresh kidney bean leaf were soaked in a 0.2% tetracycline solution for 24 hours and then placed into a 9 cm diameter Petri dish with sponge. Moist cotton strips were placed around the bean leaf to prevent the spider mites from escaping. Newly hatched Tetranychus turkestani larvae (unfed, nearly white) were selected and placed on the leaf, where they were allowed to grow and reproduce naturally. Distilled water was added daily to the Petri dish to maintain the moisture of the sponge, and the leaf were replaced with a fresh one in a timely manner. Once the larvae matured, about 30 individuals were selected for PCR detection of Wolbachia and Rickettsia infections. If no infections was detected, the offspring of this strain were continuously cultured, to obtaining an experimental strain uninfected with Wolbachia and Rickettsia.
Nomenclature of spider mite strains: IW represented the strain singly infected with Wolbachia, IR represented the strain singly infected with Rickettsia, IWR represented the strain co-infected with Wolbachia and Rickettsia, and IU represented the uninfected strain. F stands for female, and M stands for male. Tetranychus turkestani can be abbreviated as T. turkestani.
2.4. Wolbachia and Rickettsia Phylogenetic Tree Construction
The wsp sequence of Wolbachia and the gltA sequence of Rickettsia (see Appendix Table A2 , Table A3 and Table A4) were used in the PCR. PCR amplification products were detected using 1% agarose gel electrophoresis, and positive results were further purified using gel recovery, and then the purified products were sent to Youkang Biotechnology Co., Ltd. for bidirectional sequencing. Sequences of Wolbachia wsp and Rickettsia gltA from different species were searched and downloaded from the NCBI database. ClustaIW sequence alignment was performed using MEGA11, and an NJ (Neighbor - joining) phylogenetic tree was constructed. Bootstrap analysis with 1000 replicates was conducted.
2.5. Detection of the Maternal Inheritance Efficiency of Wolbachia and Rickettsia
The maternal inheritance efficiency of the symbiotic bacteria Wolbachia and Rickettsia was determined by measuring the infection rates of the two bacteria in the male offspring from parthenogenesis of single female mites or the female offspring of sexual reproduction of single pairs of Tetranychus turkestani. The parthenogenetic offspring of IW, IR, and IWR female mites, the bisexual reproductive offspring of IW female mites and IU male mites, the bisexual reproductive offspring of IR female mites and IU male mites, and the bisexual reproductive offspring of IWR female mites and IU male mites were selected respectively. Using the primers for Rickettsia gltA and Wolbachia wsp, the infection status of Rickettsia and Wolbachia was detected by PCR. A total of 10/50 female mites were randomly selected (since the number of parthenogenetic offspring of IR female mites is relatively small, 50 IR female mites were selected) to determine whether they undergo arrhenotokous parthenogenesis or bisexual reproduction. Subsequently, 10/2 male or female offspring from each female mite were tested, with a total of 100 offspring individuals in each group. Based on the PCR amplification results, the infection rates of Wolbachia and Rickettsia were calculated.
2.6. Detection of the Titers of Wolbachia and Rickettsia in Tetranychus turkestani
Based on the gltA gene sequence of Rickettsia and the wsp gene sequence of Wolbachia, specific quantitative primers were designed to detect the titers of Rickettsia and Wolbachia in Tetranychus turkestani. The RPSI8 reference gene was selected as an internal control for data standardization and quantification [28](see Appendix Table A5, A6, A7). Adult male and female mites from different infected strains were quantified, with 200-300 individuals per group constituting one replicate, and the experiment was repeated three times. The quantitative PCR (qPCR) reactions were performed on ABI Prism 7500 qPCR instrument. The PCR cycling conditions were as follows: 95°C for 30 seconds; 95°C for 5 seconds, 60°C for 30 seconds, 40 cycles. To verify the specificity of the qPCR products, a melting curve (95°C for 15 seconds, 60°C for 1 minute, 95°C for 15 seconds) was conducted at the end of the reaction. Three technical replicates were performed for each sample. A negative control was set for each reaction. The titer data of Wolbachia and Rickettsia in Tetranychus turkestani were analyzed using SPSS software, and the expression levels were calculated by the 2-ΔΔCt method. The statistical significance analysis was performed using Student's t test.
2.7. Effects of Different Symbiotic Bacteria Infections on the Fecundity of Tetranychus turkestani
Parthenogenesis: Fresh kidney bean leaves were taken and each leaf was divided into four circular sections with an area of approximately 4 cm² each. Single female mites in the static III stage with different infection statuses were selected and placed onto each section of the leaf. Number of eggs laid were counted daily and the counting was started from the first day the female mite begins to lay eggs. After laying eggs for five consecutive days, the female mite was removed. The daily egg-laying count and the total number of eggs laid were recorded. Once the eggs hatch into larvae, the hatching rate was recorded and when they develop into adult mites, the sex ratio (female/male) was noted.
Sexual reproduction: Four different strains of Tetranychus turkestani were selected using different crossbreeding combinations to conduct hybridization experiments. Fresh leaves were taken and divided each leaf into four circular sections, each approximately 4 cm². A single female and male mite in the static III stage with different infection statuses were placed together in each section of the leaf, with one pair per section. Two days after the female molted into a mature adult, the male was removed. Starting from the first day of egg-laying, the female mites were removed after laying eggs for five days. Daily egg-laying count and total eggs laid were recorded. After the eggs hatched into larvae, the hatching rate was recorded, and the sex ratio (female/male) was noted once the mites reached adulthood. If the parental male adult mite died before the female mite starts laying eggs, it was promptly replaced with another male adult mite. If the parental female adult mite died before completing five days of egg laying, the data for that pair was discarded. The CI level (CI%) was calculated using the formula: CI% = (1−F /FC) × 100, where F represnts the number of female offspring from incompatible crosses (♀Iu×♂IW, ♀Iu×♂IWR), and FC is the average number of female offspring from the control cross (♀Iu×♂IU) [29]. The embryonic mortality (EMs) of different Tetranychus turkestani strains (parthenogenetic individuals or sexually reproducing individuals) was calculated using the formula EM = TE-HE, where TE is the total number of eggs in a single cross, and HE is the number of hatched eggs. The post embryonic mortality (PEM) of each crossbreeding combination (♀IU×♂IWR, ♀IU×♂IW) was calculated using the following formula: PEM% = (1–AO/HE )×EM, where AO is the number of adult offspring in a single cross [30].
The above experiments were repeated 30 times. Under a microscope, the number of eggs laid by a single female mite or each pair of parents was counted, and the number of embryonic deaths and nymph deaths was recorded. Adult mites were collected for gender identification.
2.8. Data Processing
A one-way analysis of variance (SPSS 26.0) was used to compare the outputs of arrhenotokous parthenogenesis and bisexual reproduction in the IWR, IW, IR, and IU strains, and to analyze the cytoplasmic incompatibility (CI) function of Wolbachia in the IWR and IW strains. Pairwise comparisons of all variables were performed using Duncan's multiple range test. Independent samples t-tests (SPSS 26.0, P<0.05) were employed to analyze the infection titers of Wolbachia and Rickettsia in Tetranychus turkestani of different genders in the IWR, IW, and IR strains, to compare the outputs of arrhenotokous parthenogenesis and bisexual reproduction in the IWR, IR, and IW strains, and to analyze the impact of male killing induced by Rickettsia on the CI function induced by Wolbachia. Graph Pad software was used for graphing.
3. Results and Analysis
3.1. Phylogenetic Analysis of Wolbachia and Rickettsia
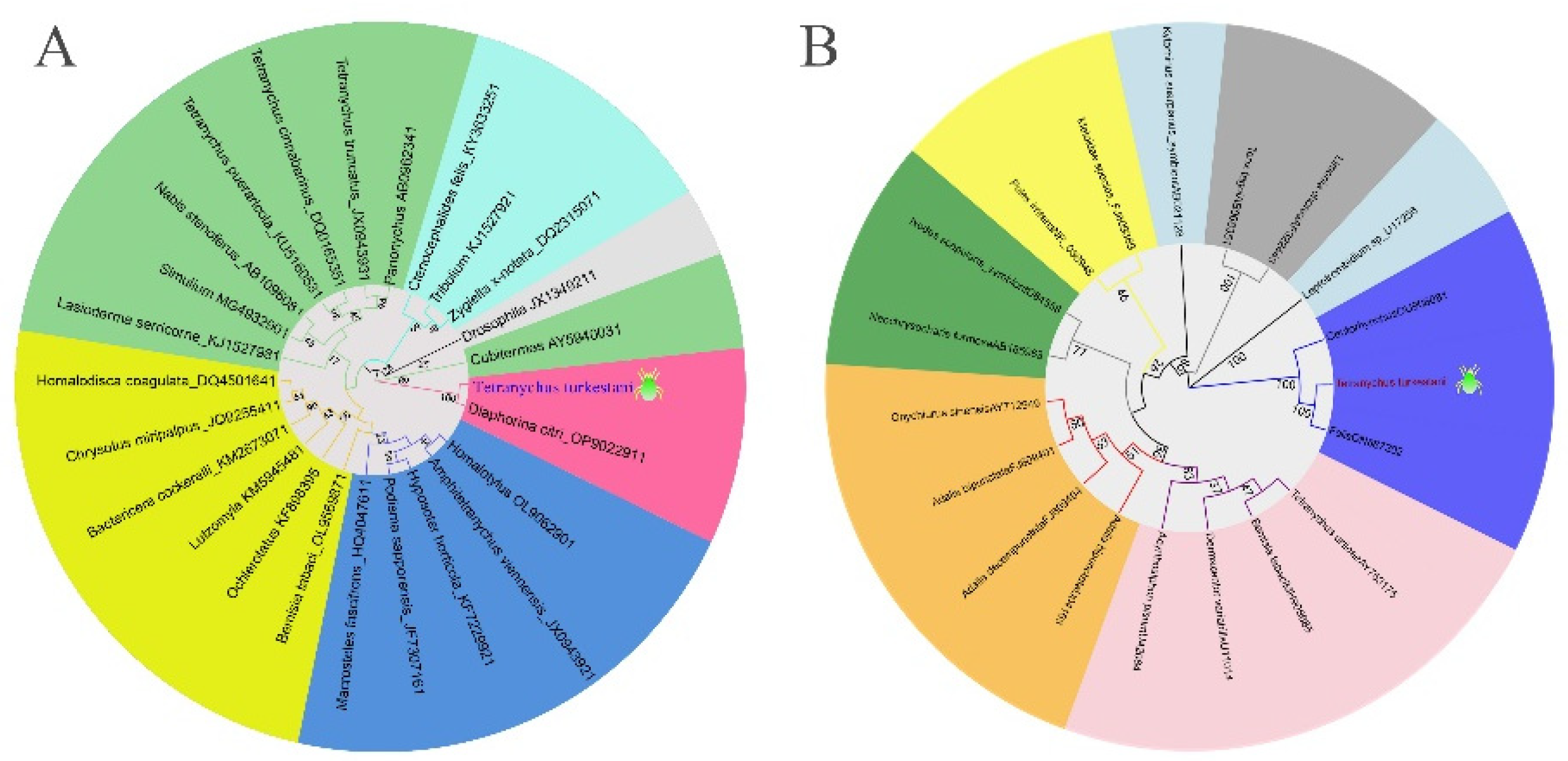
A phylogenetic tree was constructed based on the Wolbachia wsp sequence and 25 Wolbachia strains from different species in the database, and the Wolbachia infecting Tetranychus turkestani was classified into group B. The Wolbachia infecting Tetranychus turkestani was found to have a relatively close evolutionary relationship with the Wolbachia infecting Diaphorina citri. A phylogenetic tree was also constructed based on the Rickettsia gltA sequence and 19 Rickettsia strains from different species in the database. The Rickettsia infecting Tetranychus turkestani was found to have a relatively close evolutionary relationship with the Rickettsia infecting Leptotrombidium and Ceutorhynchus.
Figure 1.
Figure 1. Phylogenetic Trees. (A) Wolbachia. (B) Rickettsia. The numbers at the nodes are bootstrap values, which were used to evaluate the reliability of the branching structure.
Figure 1.
Figure 1. Phylogenetic Trees. (A) Wolbachia. (B) Rickettsia. The numbers at the nodes are bootstrap values, which were used to evaluate the reliability of the branching structure.
3.2. Analysis of the Maternal Inheritance Efficiency of Wolbachia and Rickettsia
The maternal inheritance efficiency of the two symbiotic bacteria was determined through the infection rates of Wolbachia and Rickettsia in the offspring of Tetranychus turkestani. The results showed that in all male and female offspring from the IWR, IW, and IR lines, regardless of whether they originated from parthenogenesis or sexual reproduction, Rickettsia and Wolbachia were transmitted from the mother with complete infection (100%).

Table 1.
The maternal inheritance efficiency of Rickettsia and Wolbachia.
|
T. turkestani Strains |
Number of adult female mites |
Number of offspring |
Total number of specimens tested |
Rickettsia | Wolbachia | ||||
| n➕ | n➖ | % | n➕ | n➖ | % | ||||
| ♀IW | 10 | 10 | 100 | 0 | 100 | 0 | 100 | 0 | 100 |
| ♀IR | 50 | 2 | 100 | 100 | 0 | 100 | 0 | 100 | 0 |
| ♀IWR | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
| ♀IW×♂IU | 10 | 10 | 100 | 0 | 100 | 0 | 100 | 0 | 100 |
| ♀IR×♂IU | 10 | 10 | 100 | 100 | 0 | 100 | 0 | 100 | 0 |
| ♀IWR×♂IU | 10 | 10 | 100 | 100 | 0 | 100 | 100 | 0 | 100 |
Note: n, number; n +, number of positive individuals; n➖ , number of negative individuals.
3.3. Detection of the Titers of Wolbachia and Rickettsia in Tetranychus turkestani
The target bands were obtained by PCR amplification using Wolbachia-specific primers WSP-236F/44R and Rickettsia-specific primers RICTG - F/R (see Appendix Figure A1 ). The IWR strain of Tetranychus turkestani was co-infected with both endosymbiotic bacteria, the Iw strain was infected only with Wolbachia, the IR strain was infected only with Rickettsia, and the Iu strain was uninfected with either of these two symbiotic bacteria. Real time quantitative PCR was used to measure the titers of Wolbachia and Rickettsia in male and female adult mites of different strains. The results showed that there were significant differences in the contents of Wolbachia and Rickettsia between male and female adult mites, with female adult mites having significantly higher content than male adult mites. The content of Wolbachia in the IW strain was significantly higher than that in the IWR strain (Figure 2A). In contrast, the content of Rickettsia was the opposite, with the content in female mites of the IWR strain being higher than that in the IR strain (Figure 2B).
3.4. Effects of Different Endosymbiont Infections on the Parthenogenesis of Tetranychus turkestani
Four strains of Tetranychus turkestani reproduced parthenogenetically, with all offspring being male. The average number of eggs laid (per female) of the IWR strain was significantly higher than that of the Iu, IW, and IR strains (number of eggs laid: 43.80 ± 6.04 vs 36.20 ± 5.47, 36.20 ± 4.16, 35.10 ± 3.56, P < 0.001, Figure 3A). The IWR strain had the lowest embryonic mortality rate, while the IR strain had the highest (0.92 ± 0.05, P<0.001, Figure 3B). There were no significant differences in the nymph survival rates among the four strains (Figure 3C). The number of male offspring in the IWR strain was the highest, and that in the IR strain was the lowest. The numbers in the IU and IW strains were at an intermediate level and significantly different from those in the IWR and IR strains (35.47 ± 4.89 vs 2.50 ± 1.57, 28.97 ± 5.04, 28.67 ± 4.20, P<0.001, Figure 3D). Eggs produced by parthenogenesis from Tetranychus turkestani strain singly infected with Rickettsia failed to hatch normally, with a large number of male embryos died, which might lead to the least number of male offspring in the IR strain.
3.5. Effects of Different Endosymbiotic Bacterial Infections on the Sexual Reproduction of Tetranychus turkestani
In all four strains, the mated females produced both female and male offspring. The fecundity of mated female mites of the IWR strain was the highest (45.03 ± 7.78 vs 38.97 ± 6.13, 34.93 ± 4.32, 38.90 ± 5.63, P < 0.013, Figure 4A). Both the embryonic and nymphal mortality rates of the IWR strain were significantly lower than those of the IW and IU strains (embryonic mortality rate: 0.11 ± 0.04 vs 0.18 ± 0.07, 0.25 ± 0.08, P < 0.001, Figure 4B; nymphal mortality rate: 0.12 ± 0.05 vs 0.16 ± 0.04, 0.17 ± 0.04, P < 0.001, Figure 4C). The number of female offspring and the female to male sex ratio in the IWR strain were significantly higher than those in the other three strains (number of female offspring: 29.80 ± 5.01 vs 16.70 ± 3.74, 15.30 ± 2.89, 24.43 ± 5.39, P < 0.001, Figure 4E; female to male sex ratio: 0.85 ± 0.04 vs 0.62 ± 0.07, 0.70 ± 0.04, 0.80 ± 0.06, P < 0.001, Figure 4F). However, at the same time, the number of male offspring in the IWR strain was significantly lower than that in the IU strain (5.20 ± 1.88 vs 10.17 ± 2.10, P < 0.001, Figure 4D). This indicates that in Tetranychus turkestani co-infection with Wolbachia and Rickettsia, male offspring die, resulting in an increase in the female to male sex ratio. In other words, co-infection with Wolbachia and Rickettsia can induce male death in Tetranychus turkestani.
3.6. Verification of the Male - Killing Effect of Rickettsia in Tetranychus turkestani
Parthenogenesis and sexual reproduction of female mites in the IWR and IW strains were further investigated to verify the male-killing effect of Rickettsia. Both IWR and IW strains produced male offspring parthenogenetically, and there was no significant difference in the nymphal mortality rate of the offspring (Figure 5A). Intraspecific sexual reproduction of female mites in the IWR and IW strains simultaneously produced both female and male offspring. The results showed that compared with the IW strain without Rickettsia, the IWR strain with Rickettsia had a significantly lower nymphal mortality rate (0.12 ± 0.05 vs 0.17 ± 0.04, P < 0.001, Figure 5A). Compared with the IW strain, the number of male offspring in the IWR strain was significantly reduced (5.20 ± 1.88 vs 6.40 ± 1.25, P < 0.01, Figure 5B), while the number of female offspring was significantly increased (29.80 ± 5.01 vs 15.30 ± 2.89, P < 0.001, Figure 5C), and the female to male sex ratio in the IWR strain was significantly higher than that in the IW strain (0.85 ± 0.04 vs 0.70 ± 0.04, P < 0.001, Figure 5D). It can be inferred that the infection of Rickettsia led to the death of more male offspring, resulting in an increase in the female to male sex ratio. This verifies that Rickettsia induced male killing in Tetranychus turkestani.
3.7. Verification of the Cytoplasmic Incompatibility (CI) Induced by Wolbachia in Singly-infected Tetranychus turkestani
A hybridization experiment using IU and Iw strains was carried out to verify that Wolbachia inducd CI in singly-infected Tetranychus turkestani. Among the four different mating combinations, the fecundity of the ♀IU×♂IW combination was significantly lower than that of the other three combinations (Figure 6A). The embryonic and nymph mortality rates of the ♀IU×♂IW combination were significantly higher than those of the other three combinations (embryonic mortality rate: 0.37 ± 0.05, P < 0.001, Figure 6B; nymph mortality rate: 0.22 ± 0.03, P < 0.001, Figure 6C), and the female to male sex ratio was the lowest (0.52 ± 0.07, P < 0.002, Figure 6D). These results all indicated that Wolbachia induces strong CI in Tetranychus turkestani.
3.8. Verification of the Cytoplasmic Incompatibility (CI) Induced by Wolbachia and Rickettsia in co-infected Tetranychus turkestani
A hybridization experiment using Iu and IWR strains was conducted to verify that Wolbachia and Rickettsia induced CI in co-infected Tetranychus turkestani. The fecundity of the ♀Iu×♂IWR combination was lower than that of the ♀IWR×♂IWR combination (40.57 ± 4.53 vs 45.03 ± 7.78, P < 0.004, Figure 7A), but there was no significant difference compared with the other two hybridization combinations. The embryonic and nymph mortality rates of the ♀Iu×♂IWR combination were the highest, significantly higher than those of the ♀IWR×♂IWR combination (embryonic mortality rate: 0.20 ± 0.03 vs 0.11 ± 0.04, P < 0.001, Figure 7B; nymph mortality rate: 0.18 ± 0.03 vs 0.12 ± 0.05, P < 0.001, Figure 7C). Meanwhile, the female - to - male sex ratio of the ♀IU×♂IWR combination was significantly lower than that of the other three groups (0.54 ± 0.08 vs 0.62 ± 0.07, 0.71 ± 0.08, 0.85 ± 0.04, P < 0.001, Figure 7D). These results indicate that Wolbachia and Rickettsia also induce CI in co-infected Tetranychus turkestani.
3.9. Antagonistic Effect of Rickettsia-induced Male-killing on the Strength of Wolbachia-induced CI
After verifying that Rickettsia infection can induce male-killing in Tetranychus turkestani, we further investigated the impact of Rickettsia-induced male-killing on Wolbachia-induced CI. Compared with the singly-infected ♀IU×♂IW combination, the fecundity of the co-infected ♀IU×♂IWR combination was significantly increased (40.57 ± 4.53 vs 32.50 ± 2.61, P < 0.001, Figure 8A), and the embryonic and nymphal mortality rates were significantly decreased (embryonic mortality rate: 0.37 ± 0.05 vs 0.20 ± 0.03, P < 0.001, Figure 8B; nymph mortality rate: 0.22 ± 0.03 vs 0.18 ± 0.03, P < 0.001, Figure 8C). Compared with the ♀IU×♂IW combination without Rickettsia infection, the co-infected ♀IU×♂IWR combination had more female and male offspring (female offspring: 14.47 ± 2.80 vs 8.33 ± 1.40, P < 0.001, Figure 8D; male offspring: 12.20 ± 2.54 vs 7.63 ± 1.59, P < 0.001, Figure 8E). Moreover, the CI level of the co-infected ♀IU×♂IWR combination was significantly lower than that of the single-infected ♀IU×♂IW combination (9.90 ± 22.77 vs 48.51 ± 10.13, P < 0.001; Figure 8F). This indicated that the IWR strain induced a weaker CI, which might be due to partially antagonism of Rickettsia-induced male - killing against CI induced by Wolbachia.
3.10. Wolbachia Does Not Have a Male-Killing Effect on Tetranychus turkestani
Female mites of the IW and IU strains produced male offspring through parthenogenesis, with no significant differences in the fecundity and hatching rate (fecundity: IW vs. IU = 36.00 ± 3.01 vs. 36.08 ± 4.21, p > 0.05, Figure 9A; hatching rate: IW vs. IU = 0.87 ± 0.05 vs 0.89% ± 0.05, p > 0.05, Figure 9B). This indicated that Wolbachia did not exhibit the male-killing effect on Tetranychus turkestani.
4. Discussion
In this study, different strains of Tetranychus turkestani infected with different endosymbiotic bacteria (Wolbachia and Rickettsia) were examined, and phylogenetic analysis was carried out. The results showed that the Wolbachia infected Tetranychus turkestani belonged to supergroup B and can induce cytoplasmic incompatibility (CI) in the host. The transmission efficiency of Wolbachia and Rickettsia were statistically analyzed. The results revealed that, regardless of parthenogenesis or sexual reproduction, both symbiotic bacteria completely follow maternal transmission. Real- time quantitative PCR was used to determine the titers of the endosymbiotic bacteria. The results showed significant differences in the abundance of Wolbachia and Rickettsia between male and female adult mites.
Rickettsia is a maternally inherited symbiotic bacterium. In some hosts, it acts as a nutritional symbiont, while in others, it influences the host reproduction through reproductive regulations such as parthenogenesis induction and male killing. It can also enhance the host resistance to pesticides and improve the host ability to resist to predators, high temperatures, or other lethal factors [3,4,16,17,31,32,33,34]. To date, no experimental studies have investigated the reproductive regulation of this bacterium in mites. Our research demonstrated that Rickettsia infected spider mite resulted in parthenogenesis producing only male offspring, but the hatching rate of male embryos was extremely low. Sexual reproduction in Rickettsia singly infected mites produced both female and male offspring, with increased number of female offspring than male, resulting in a high female to male sex ratio. This indicated that Rickettsia infection leads to a male - killing phenotype.
Cytoplasmic incompatibility (CI) is the most common reproductive regulation induced by Wolbachia, typically occurs in two forms. The first form is characterized by high embryonic mortality rate, resulting in a decrease number of female offspring, which is called female lethality. The second form does not decreased the total number of offspring, but results in an increase in the number of male offspring, known as male development. Both female lethality and male development induced by Wolbachia may occur simultaneously in a single insect host, such as the Wolbachia wLhetl strain in parasitic wasps [35,36]. In this study, sexual reproduction of Tetranychus turkestani (♀IU×♂IW) significantly increased the mortality rates of embryos and nymphs, and significantly decreased the female to male sex ratio, which belongs to the female lethality type. Wolbachia singly-infected in Tetranychus turkestani induced strong CI.
Currently, research on the interaction between Rickettsia and Wolbachia is limited, especially regarding the impact of Rickettsia and Wolbachia co-infection on the reproductive regulation of host insects, which has not yet been reported. In this study, we found that the Rickettsia and Wolbachia co-infection induced cytoplasmic incompatibility (CI) in Tetranychus turkestani, but the intensity was much weaker than that induced by single-infection of Wolbachia. This may be because the male killing effect of the Rickettsia in co-infection reduced the CI level induced by Wolbachia. Subsequent hybridization experiments clearly demonstrated an antagonistic interaction between the male killing effect of Rickettsia and the CI induced by Wolbachia. Previous studies have shown that there are two types of Wolbachia in nature: one that maintains a high prevalence in insect host and weakly induces CI, such as Wolbachia in Drosophila melanogaster [37]; the another that induces strong CI but maintains a low prevalence and titer, like Wolbachia in Drosophila melanogaster [38]. The Wolbachia in Tetranychus turkestani studied here was most similar to the former. Furthermore, compared with the control group (IU×IU) without symbiotic bacteria infection, the Rickettsia-Wolbachia co-infection had a higher fecundity, lower mortality, a higher female to male ratio, and more offspring. In conclusion, the synergistic effect of the two symbiotic bacteria significantly improved the fitness of Tetranychus turkestani.
In conclusion, our study revealed five findings (Figure 10): Rickettsia infection induced male-killing effect in the sexual reproduction of Tetranychus turkestani; Rickettsia infection caused the death of parthenogenetically produced male embryos in Tetranychus turkestani, leading to a reduction in the number of offspring; Wolbachia single infection induced strong cytoplasmic incompatibility (CI) in Tetranychus turkestani, while Rickettsia-Wolbachia co-infection induced weaker CI; the male killing effect induced by Rickettsia antagonized the strong CI induced by Wolbachia; Rickettsia-Wolbachia co-infection promoted the fitness of Tetranychus turkestani, suggesting synergistic mutualistic relationship between the two symbiotic bacteria within the same host Tetranychus turkestani. We gained a better understanding of the complex interactions between symbiotic bacteria and host, as well as between different symbionts, providing strategies and insights for future applications of symbionts in biological control.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, S.W and Y.Z.; methodology, S.W and Y.Z.; software, S.W.; validation, S.W., X.W. and A.B.; formal analysis, X.W.; investigation, S.W.; resources, S.W and X.W.; data curation, S.W.; writing—original draft preparation, S.W.; writing—review and editing, S.W., Y.Z., Q.W., K.Z. and A.B.; visualization, S.W.; supervision, X.W.; project administration, Y.Z.; funding acquisition, Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. 32260676 and No. 31860508), the Program for Young Leading Scientists in Science and Technology of XPCC (No. 2022CB002 - 06), and the Natural Science Foundation of XPCC(No. 2024DA018).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How Many Species Are Infected with Wolbachia? – A Statistical Analysis of Current Data: Wolbachia Infection Rates. FEMS Microbiology Letters 2008, 281, 215–220. [CrossRef]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The Incidence of Bacterial Endosymbionts in Terrestrial Arthropods. Proc. R. Soc. B. 2015, 282, 20150249. [CrossRef]
- Brumin, M.; Levy, M.; Ghanim, M. Transovarial Transmission of Rickettsia Spp. and Organ-Specific Infection of the Whitefly Bemisia Tabaci. Appl Environ Microbiol 2012, 78, 5565–5574. [CrossRef]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative Bacterial Symbionts in Aphids Confer Resistance to Parasitic Wasps. Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 1803–1807. [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat Rev Microbiol 2008, 6, 741–751. [CrossRef]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and Virus Protection in Insects. Science 2008, 322, 702. [CrossRef]
- Perlman, S.J.; Hunter, M.S.; Zchori-Fein, E. The Emerging Diversity of Rickettsia. Proc Biol Sci 2006, 273, 2097–2106. [CrossRef]
- Liu, B.-Q.; Bao, X.-Y.; Yan, J.-Y.; Zhang, D.; Sun, X.; Li, C.-Q.; Chen, Z.-B.; Luan, J.-B. Rickettsia Symbionts Spread via Mixed Mode Transmission, Increasing Female Fecundity and Sex Ratio Shift by Host Hormone Modulating. Proc Natl Acad Sci U S A 2024, 121, e2406788121. [CrossRef]
- Dobson, S.L.; Bourtzis, K.; Braig, H.R.; Jones, B.F.; Zhou, W.; Rousset, F.; O’Neill, S.L. Wolbachia Infections Are Distributed throughout Insect Somatic and Germ Line Tissues. Insect Biochemistry and Molecular Biology 1999, 29, 153–160. [CrossRef]
- Ilbeigi Khamseh Nejad, M.; Cappelli, A.; Damiani, C.; Falcinelli, M.; Catapano, P.L.; Nanfack-Minkeu, F.; Mayi, M.P.A.; Currà, C.; Ricci, I.; Favia, G. Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species. Microorganisms 2024, 12, 545. [CrossRef]
- Fast, E.M.; Toomey, M.E.; Panaram, K.; Desjardins, D.; Kolaczyk, E.D.; Frydman, H.M. Wolbachia Enhance Drosophila Stem Cell Proliferation and Target the Germline Stem Cell Niche. Science 2011, 334, 990–992. [CrossRef]
- Dobson, S.L.; Rattanadechakul, W.; Marsland, E.J. Fitness Advantage and Cytoplasmic Incompatibility in Wolbachia Single- and Superinfected Aedes Albopictus. Heredity (Edinb) 2004, 93, 135–142. [CrossRef]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The Endosymbiotic Bacterium Wolbachia Induces Resistance to Dengue Virus in Aedes Aegypti. PLoS Pathog 2010, 6, e1000833. [CrossRef]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia Induces Reactive Oxygen Species (ROS)-Dependent Activation of the Toll Pathway to Control Dengue Virus in the Mosquito Aedes Aegypti. Proc Natl Acad Sci U S A 2012, 109, E23–E31. [CrossRef]
- Hagimori, T.; Abe, Y.; Date, S.; Miura, K. The First Finding of a Rickettsia Bacterium Associated with Parthenogenesis Induction among Insects. Curr Microbiol 2006, 52, 97–101. [CrossRef]
- Himler, A.G.; Adachi-Hagimori, T.; Bergen, J.E.; Kozuch, A.; Kelly, S.E.; Tabashnik, B.E.; Chiel, E.; Duckworth, V.E.; Dennehy, T.J.; Zchori-Fein, E.; et al. Rapid Spread of a Bacterial Symbiont in an Invasive Whitefly Is Driven by Fitness Benefits and Female Bias. Science 2011, 332, 254–256. [CrossRef]
- Chen, X.; Li, S.; Aksoy, S. Concordant Evolution of a Symbiont with Its Host Insect Species: Molecular Phylogeny of Genus Glossina and Its Bacteriome-Associated Endosymbiont, Wigglesworthia Glossinidia. J Mol Evol 1999, 48, 49–58. [CrossRef]
- Fan, Z.-Y.; Liu, Y.; He, Z.-Q.; Wen, Q.; Chen, X.-Y.; Khan, M.M.; Osman, M.; Mandour, N.S.; Qiu, B.-L. Rickettsia Infection Benefits Its Whitefly Hosts by Manipulating Their Nutrition and Defense. Insects 2022, 13, 1161. [CrossRef]
- Zélé, F.; Weill, M.; Magalhães, S. Identification of Spider-Mite Species and Their Endosymbionts Using Multiplex PCR. Experimental and Applied Acarology 2018, 74, 123–138. [CrossRef]
- Frank, S.A. Dynamics of Cytoplasmic Incompatability with MultipleWolbachiaInfections. Journal of Theoretical Biology 1998, 192, 213–218. [CrossRef]
- Zchori-Fein, E.; Perlman, S.J. Distribution of the Bacterial Symbiont Cardinium in Arthropods. Mol Ecol 2004, 13, 2009–2016. [CrossRef]
- Dobson, S.L. Evolution of Wolbachia Cytoplasmic Incompatibility Types. Evolution 2004, 58, 2156–2166. [CrossRef]
- Ros, V.I.D.; Breeuwer, J. a. J. The Effects of, and Interactions between, Cardinium and Wolbachia in the Doubly Infected Spider Mite Bryobia Sarothamni. Heredity (Edinb) 2009, 102, 413–422. [CrossRef]
- Zhao, D.-X.; Zhang, X.-F.; Hong, X.-Y. Host-Symbiont Interactions in Spider Mite Tetranychus Truncates Doubly Infected with Wolbachia and Cardinium. Environ Entomol 2013, 42, 445–452. [CrossRef]
- Ben-David, T.; Melamed, S.; Gerson, U.; Morin, S. ITS2 Sequences as Barcodes for Identifying and Analyzing Spider Mites (Acari: Tetranychidae). Exp Appl Acarol 2007, 41, 169–181. [CrossRef]
- Navajas, M.; Boursot, P. Nuclear Ribosomal DNA Monophyly versus Mitochondrial DNA Polyphyly in Two Closely Related Mite Species: The Influence of Life History and Molecular Drive. Proc Biol Sci 2003, 270 Suppl 1, S124-127. [CrossRef]
- Chu, D.; Gao, C.S.; De Barro, P.; Zhang, Y.J.; Wan, F.H.; Khan, I.A. Further Insights into the Strange Role of Bacterial Endosymbionts in Whitefly, Bemisia Tabaci: Comparison of Secondary Symbionts from Biotypes B and Q in China. Bull Entomol Res 2011, 101, 477–486. [CrossRef]
- Sun, W.; Jin, Y.; He, L.; Lu, W.-C.; Li, M. Suitable Reference Gene Selection for Different Strains and Developmental Stages of the Carmine Spider Mite, Tetranychus Cinnabarinus, Using Quantitative Real-Time PCR. J Insect Sci 2010, 10, 208. [CrossRef]
- Mouton, L.; Dedeine, F.; Henri, H.; Boulétreau, M.; Profizi, N.; Vavre, F. Virulence, Multiple Infections and Regulation of Symbiotic Population in the Wolbachia-Asobara Tabida Symbiosis. Genetics 2004, 168, 181–189. [CrossRef]
- Nguyen, D.T.; Morrow, J.L.; Spooner-Hart, R.N.; Riegler, M. Independent Cytoplasmic Incompatibility Induced by Cardinium and Wolbachia Maintains Endosymbiont Coinfections in Haplodiploid Thrips Populations. Evolution 2017, 71, 995–1008. [CrossRef]
- Sakurai, M.; Koga, R.; Tsuchida, T.; Meng, X.-Y.; Fukatsu, T. Rickettsia Symbiont in the Pea Aphid Acyrthosiphon Pisum: Novel Cellular Tropism, Effect on Host Fitness, and Interaction with the Essential Symbiont Buchnera. Appl Environ Microbiol 2005, 71, 4069–4075. [CrossRef]
- Ferrari, J.; Scarborough, C.L.; Godfray, H.C.J. Genetic Variation in the Effect of a Facultative Symbiont on Host-Plant Use by Pea Aphids. Oecologia 2007, 153, 323–329. [CrossRef]
- Kontsedalov, S.; Zchori-Fein, E.; Chiel, E.; Gottlieb, Y.; Inbar, M.; Ghanim, M. The Presence of Rickettsia Is Associated with Increased Susceptibility of Bemisia Tabaci (Homoptera: Aleyrodidae) to Insecticides. Pest Manag Sci 2008, 64, 789–792. [CrossRef]
- Shan, H.-W.; Lu, Y.-H.; Bing, X.-L.; Liu, S.-S.; Liu, Y.-Q. Differential Responses of the Whitefly Bemisia Tabaci Symbionts to Unfavorable Low and High Temperatures. Microb Ecol 2014, 68, 472–482. [CrossRef]
- Vavre, F.; Dedeine, F.; Quillon, M.; Fouillet, P.; Fleury, F.; Bouletreau, M. Within-Species Diversity of Wolbachia-Induced Cytoplasmic Incompatibility in Haplodiploid Insects. Evolution 2001, 55, 1710–1714. [CrossRef]
- Vala, F.; Breeuwer, J. a. J.; Sabelis, M.W. Sorting out the Effects of Wolbachia, Genotype and Inbreeding on Life-History Traits of a Spider Mite. Exp Appl Acarol 2003, 29, 253–264. [CrossRef]
- Kriesner, P.; Conner, W.R.; Weeks, A.R.; Turelli, M.; Hoffmann, A.A. Persistence of a Wolbachia Infection Frequency Cline in Drosophila Melanogaster and the Possible Role of Reproductive Dormancy. Evolution 2016, 70, 979–997. [CrossRef]
- Richardson, K.M.; Schiffer, M.; Griffin, P.C.; Lee, S.F.; Hoffmann, A.A. Tropical Drosophila Pandora Carry Wolbachia Infections Causing Cytoplasmic Incompatibility or Male Killing. Evolution 2016, 70, 1791–1802. [CrossRef]
Figure 2.
Density of Wolbachia and Rickettsia in female and male T. turkestani of different infection strains. (A) Relative quantity of Wolbachia. (B) Relative quantity of Rickettsia. IUF: Female of the IU population, IuM: Male of the Iu population, IRF: Female of the IR population, IRM: Male of the IR population, IwF: Female of the Iw population, IwM: Male of the Iw population, IWRF: Female of the IWR population, IWRM: Male of the IWR population. The symbol “*” indicates a statistically significant difference between the two groups (P < 0.05), while “ns” represents no significant difference. All error bars represent the standard error of the mean.
Figure 2.
Density of Wolbachia and Rickettsia in female and male T. turkestani of different infection strains. (A) Relative quantity of Wolbachia. (B) Relative quantity of Rickettsia. IUF: Female of the IU population, IuM: Male of the Iu population, IRF: Female of the IR population, IRM: Male of the IR population, IwF: Female of the Iw population, IwM: Male of the Iw population, IWRF: Female of the IWR population, IWRM: Male of the IWR population. The symbol “*” indicates a statistically significant difference between the two groups (P < 0.05), while “ns” represents no significant difference. All error bars represent the standard error of the mean.
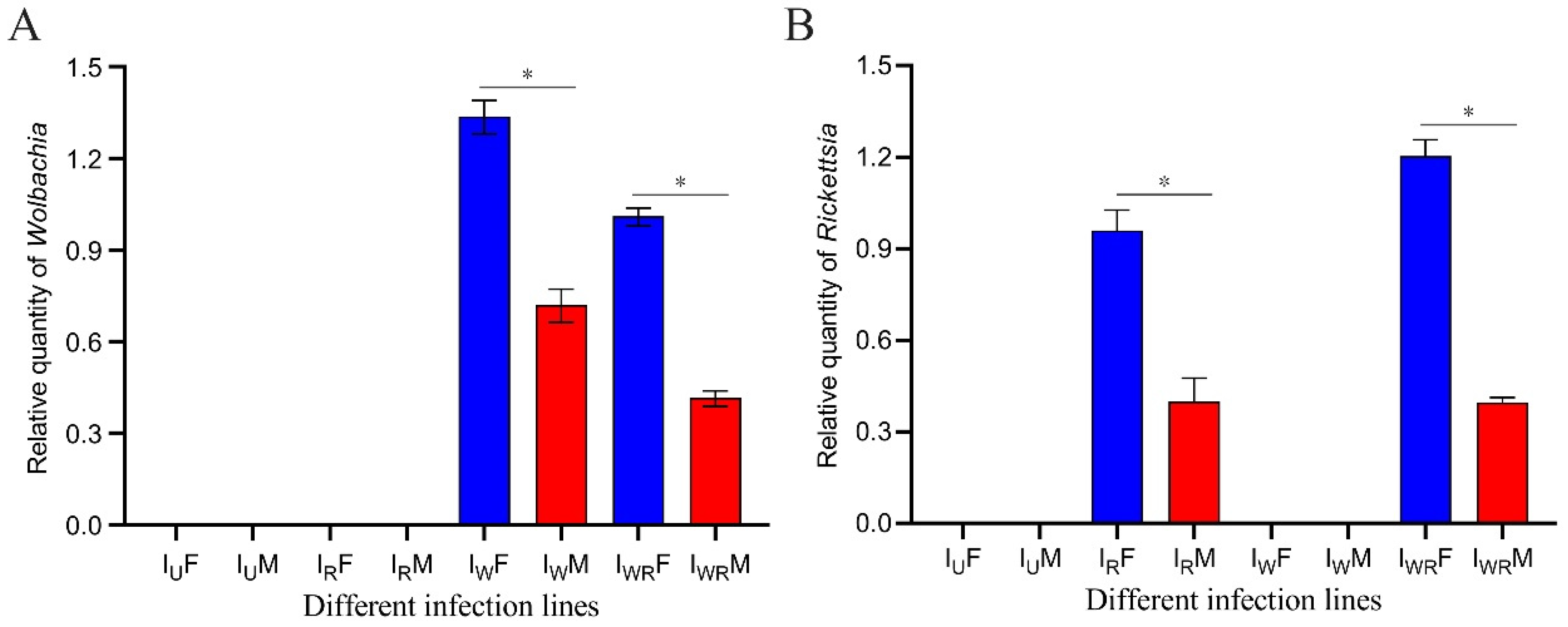
Figure 3.
Parthenogenetic parameters of IU、IW、IR、IWR in T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) The survival rate of nymphs. (D) Male offspring. The data in the figure are average ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 3.
Parthenogenetic parameters of IU、IW、IR、IWR in T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) The survival rate of nymphs. (D) Male offspring. The data in the figure are average ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
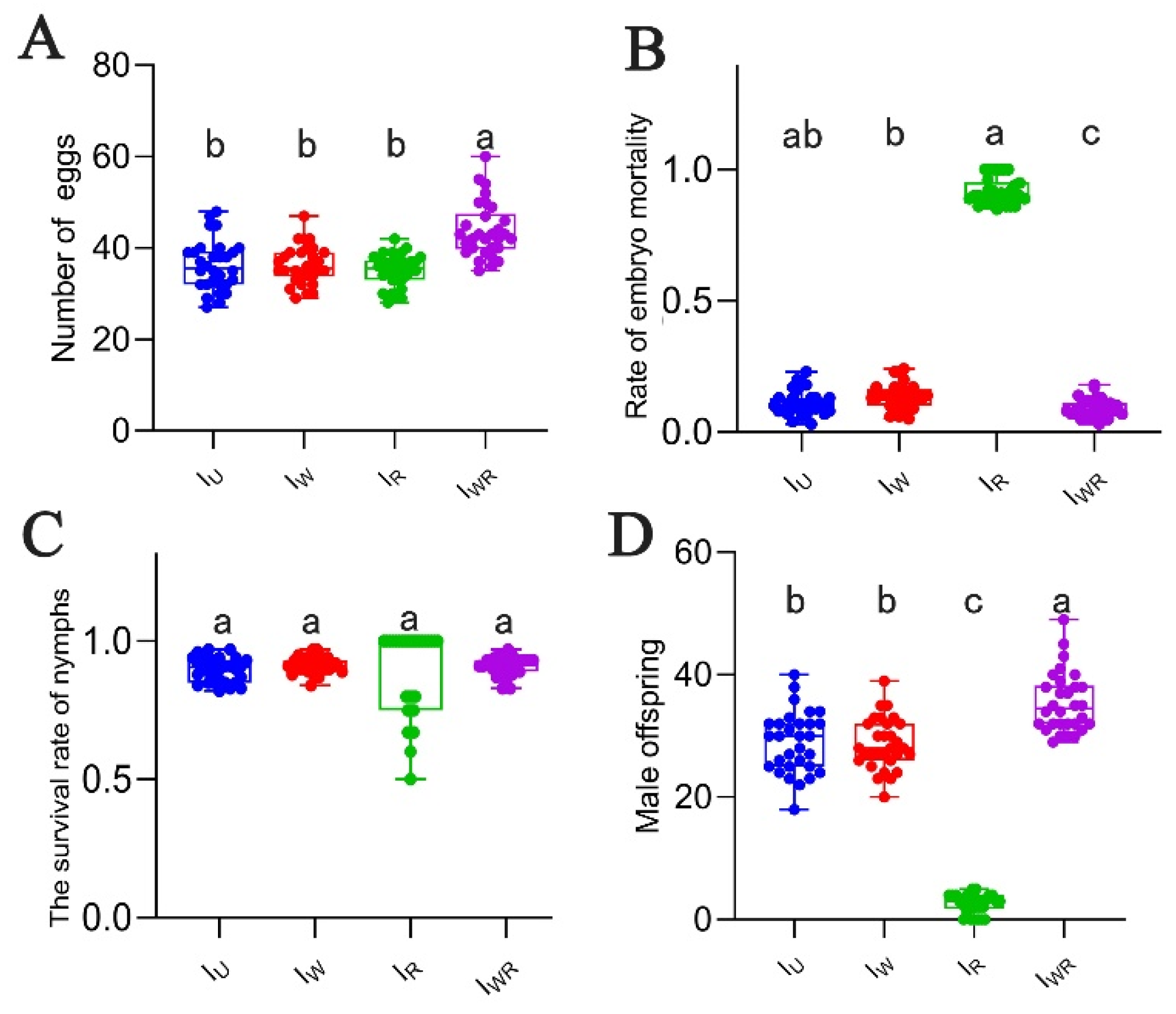
Figure 4.
Sexual reproductive parameters of IW、IR 、IWR and IU mating in T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Male offspring. (E) Female offspring. (F) Female sex ratio. The data in the figure are mean ±standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 4.
Sexual reproductive parameters of IW、IR 、IWR and IU mating in T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Male offspring. (E) Female offspring. (F) Female sex ratio. The data in the figure are mean ±standard error; The mean values of different letter markers were statistically significant (P < 0.05).
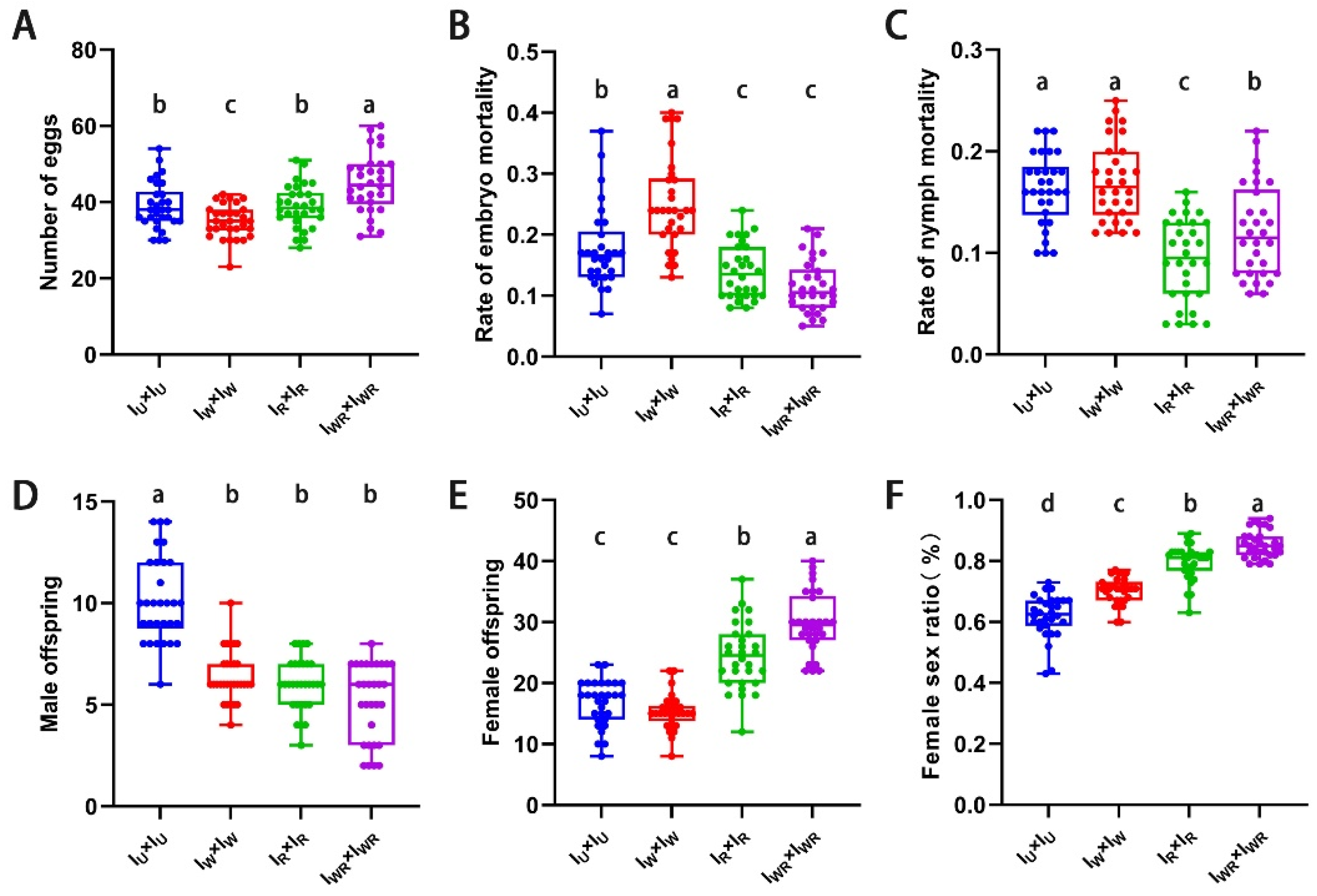
Figure 5.
Parthenogenesis and intraspecific sexual reproduction of IWR and IW in T. turkestani within 5 days of oviposition. (A) Rate of nymph mortality. (B) Male offspring. (C) Female offspring. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 5.
Parthenogenesis and intraspecific sexual reproduction of IWR and IW in T. turkestani within 5 days of oviposition. (A) Rate of nymph mortality. (B) Male offspring. (C) Female offspring. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
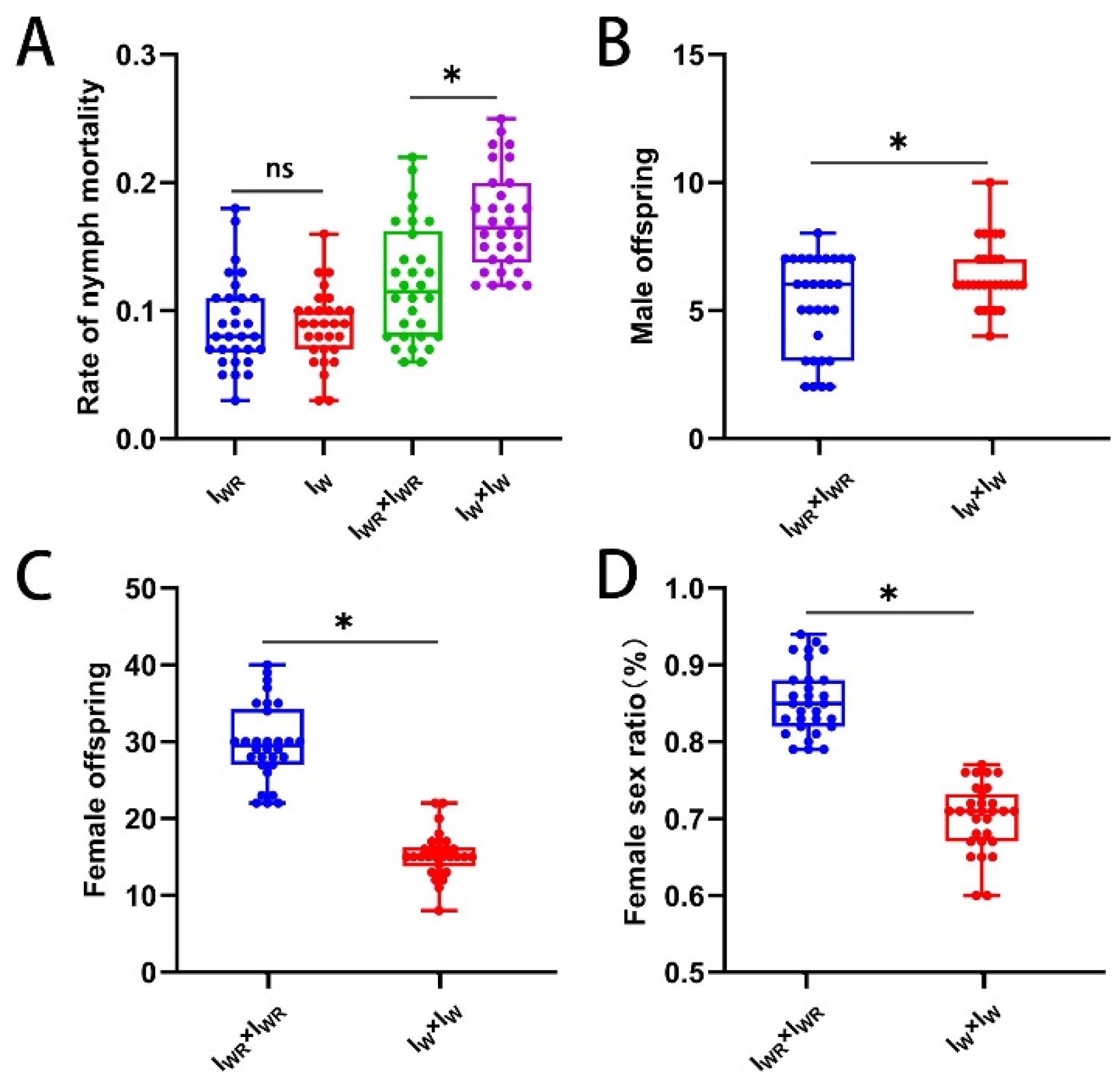
Figure 6.
CI identification of Wolbachia in Iw of T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 6.
CI identification of Wolbachia in Iw of T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
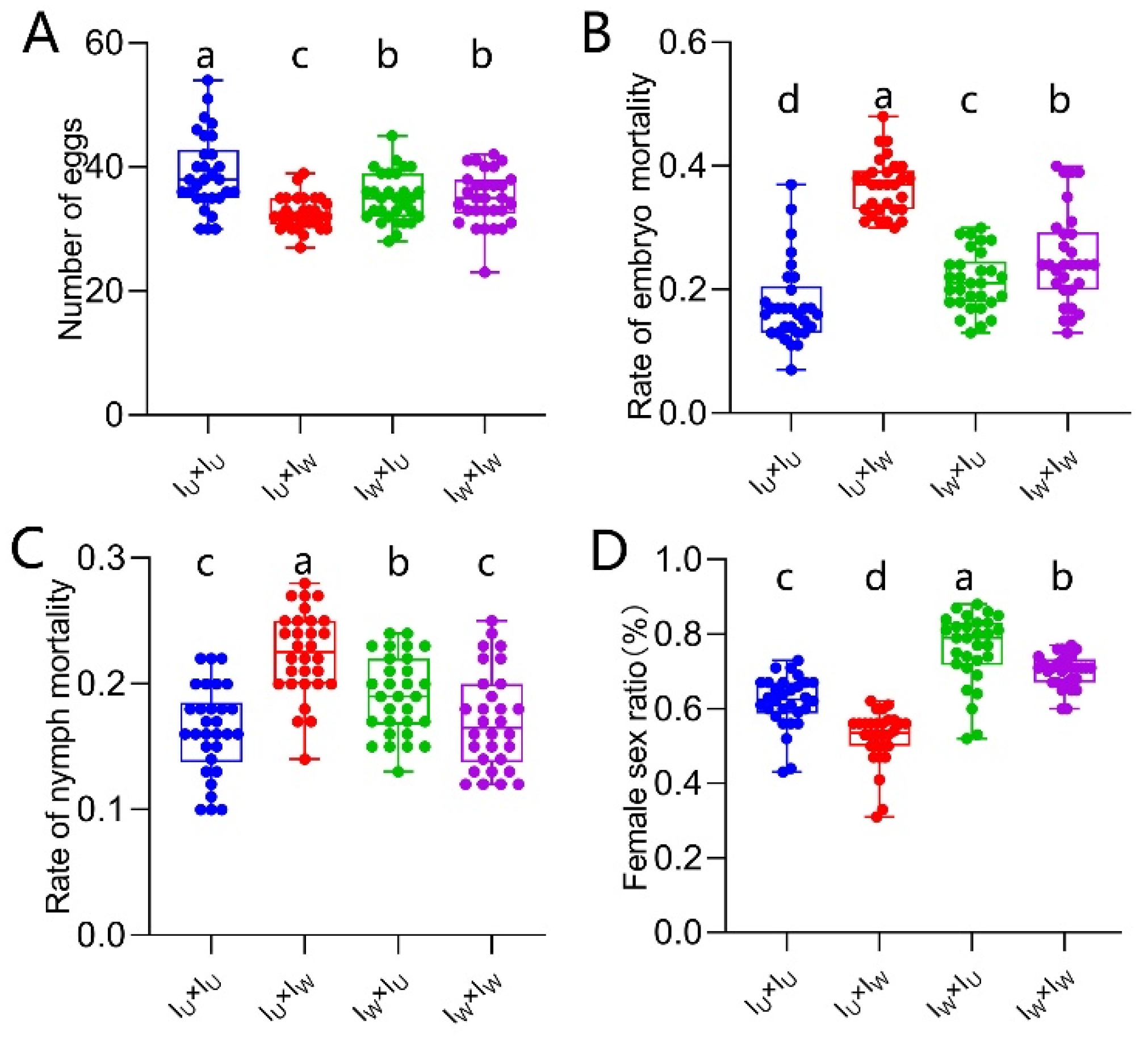
Figure 7.
CI identification of Wolbachia and Rickettsia in IWR of T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 7.
CI identification of Wolbachia and Rickettsia in IWR of T. turkestani. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female sex ratio. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
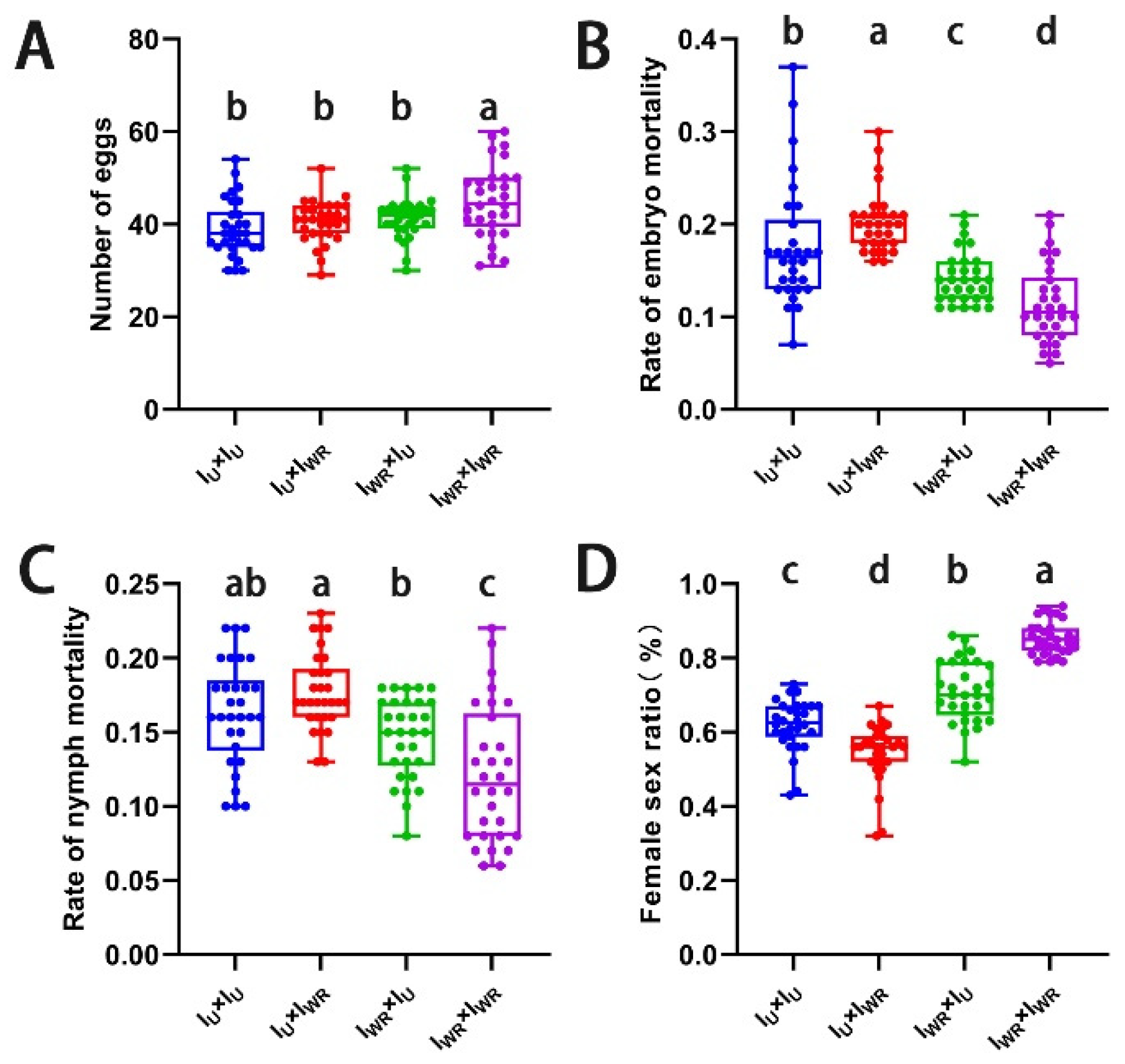
Figure 8.
Antagonistic effect of Rickettsia co-infection on CI in Wolbachia. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female offspring. (E) Male offspring. (F) CI%. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
Figure 8.
Antagonistic effect of Rickettsia co-infection on CI in Wolbachia. (A) Number of eggs. (B) Rate of embryo mortality. (C) Rate of nymph mortality. (D) Female offspring. (E) Male offspring. (F) CI%. The data in the figure are mean ± standard error; The mean values of different letter markers were statistically significant (P < 0.05).
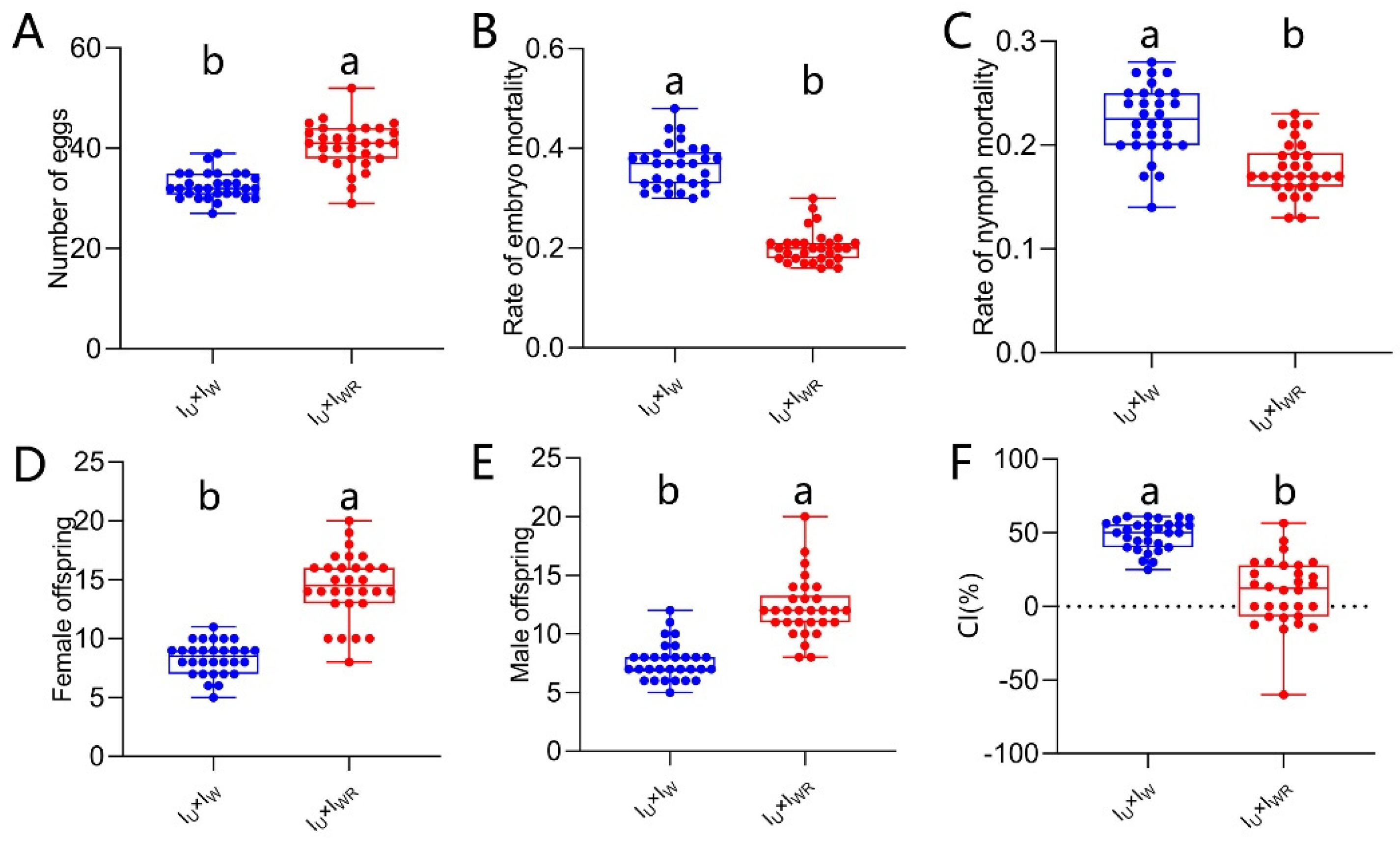
Figure 9.
The reproductive parameters of different combinations of T. turkestani. (A) Egg production of IW and IU in parthenogenesis. (B) Hatching rate of IW and IU in parthenogenesis. IU,Wolbachia-uninfected;IW,Wolbachia-infected. Crossing combinations of strains are shown as‘Female×Male’. The data in the figure are presented as mean ± standard error; means marked with different letters indicate a statistically significant difference (P < 0.05).
Figure 9.
The reproductive parameters of different combinations of T. turkestani. (A) Egg production of IW and IU in parthenogenesis. (B) Hatching rate of IW and IU in parthenogenesis. IU,Wolbachia-uninfected;IW,Wolbachia-infected. Crossing combinations of strains are shown as‘Female×Male’. The data in the figure are presented as mean ± standard error; means marked with different letters indicate a statistically significant difference (P < 0.05).
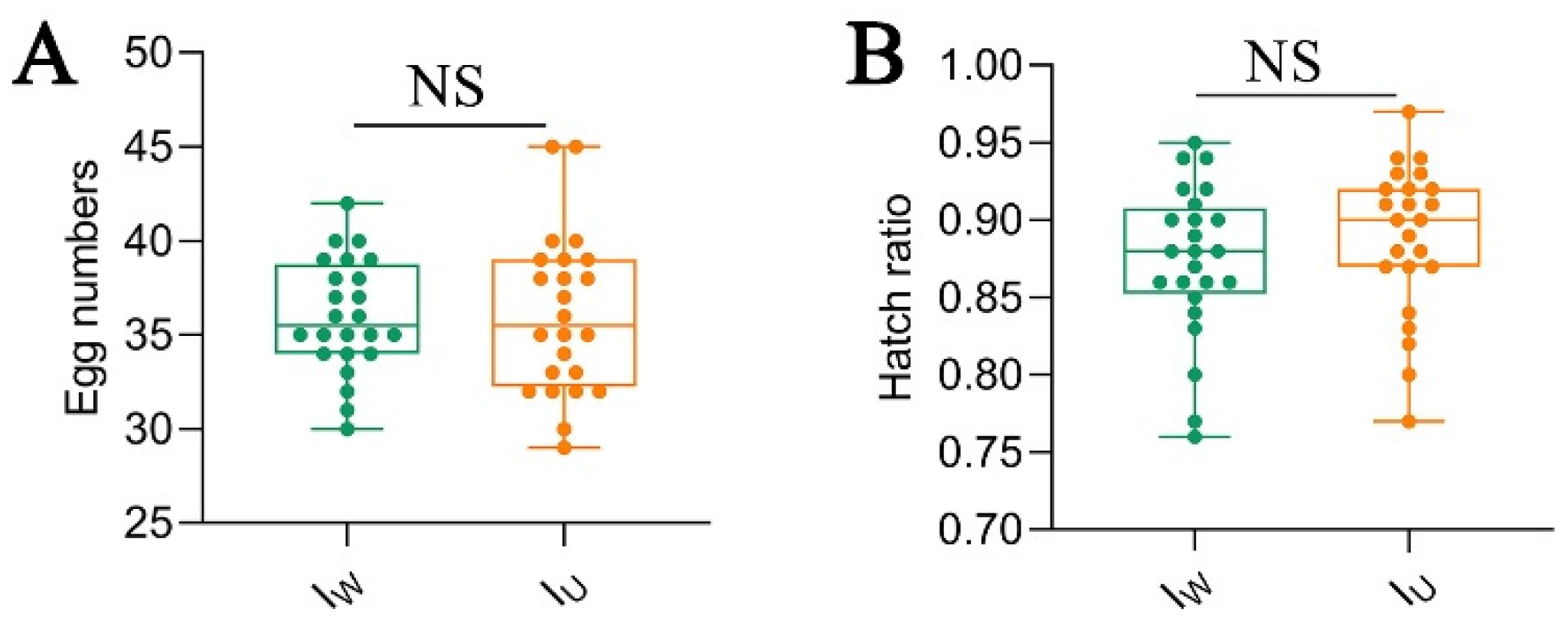
Figure 10.
The action model of Rickettsia and Wolbachia on the reproductive regulation of T. turkestani. RI: T. turkestani with single Rickettsia infection; R: Parthenogenesis with single Rickettsia infection; WCI: Cytoplasmic incompatibility (CI) induced by single Wolbachia infection; WRCI: Cytoplasmic incompatibility (CI) induced by co-infection of Wolbachia-Rickettsia; WR: T. turkestani with co-infection of Wolbachia-Rickettsia.
Figure 10.
The action model of Rickettsia and Wolbachia on the reproductive regulation of T. turkestani. RI: T. turkestani with single Rickettsia infection; R: Parthenogenesis with single Rickettsia infection; WCI: Cytoplasmic incompatibility (CI) induced by single Wolbachia infection; WRCI: Cytoplasmic incompatibility (CI) induced by co-infection of Wolbachia-Rickettsia; WR: T. turkestani with co-infection of Wolbachia-Rickettsia.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



