Submitted:
17 January 2025
Posted:
17 January 2025
You are already at the latest version
Abstract
There is an increasing interest in yeasts isolated from natural sources to be used as probi-otics. Saccharomyces-based probiotics have been proposed as a valid alternative to the con-ventional drug therapy for the prevention and treatment of vulvovaginal candidiasis, also considering the resistance of some Candida strains to many antifungals. Here we isolated from an artisanal sourdough a new yeast strain which was identified as Kluyveromyces marxianus and assessed its probiotic and safety properties, which resulted comparable at all to those exhibited by the commercial probiotic Saccharomyces boulardii. Then, we checked the antagonistic activity of the new isolate against some clinical fluconazole re-sistant C. albicans strains, showing its ability to inhibit filamentation, biofilm formation, and adhesion of C. albicans to vaginal epithelial A-431 cells. Also, K. marxianus reduced the cell damage provoked by C. albicans and the expression of SAP2 and SAP6 genes. On the whole, our results enlarge the spectrum of the beneficial properties of the food-grade yeast K. marxianus showing for the first time its biotherapeutic potential as anticandidal agent.
Keywords:
probiotic yeasts
; Kluyveromyces marxianus
; vulvovaginal candidiasis
1. Introduction
The Yeasts are components of the microbiota of various fermented foods and beverages and are largely employed in industry. The discovery of the antagonistic effects of yeasts, especially against other fungal species, has paved the way to focused investigations on their use as probiotics, that is of beneficial use for animal and human health [1].
Most conventional probiotics are of bacterial origin, mainly belonging to the Lactobacillus and Bifidobacterium genera [2]. More recently, several yeast strains have been reported as promising probiotic candidates as shown by the increasing number of publications over the last decades indicating the relevance of this topic [3]. Some of the differences between yeasts and bacteria, such as the larger size, which can block the attachment of pathogen and spoilage species, the presence of a cell wall containing mannans and beta-glucans, which can positively influence the intestinal microbiome and stimulate innate and acquired immunity, as well as the intrinsic resistance to antibiotics, are considered advantageous for the use of yeasts as probiotics [3].
Saccharomyces cerevisiae var. boulardii, first isolated in 1920, has been established as a probiotic since 1950; and now it is commercially available throughout the world, mainly used for the treatment of gastrointestinal tract disorders [4]. Later, other Saccharomyces and non-Saccharomyces yeasts isolated from vegetables, fruits, grains, and traditional fermented foods and beverages have been proposed as probiotics [5,6].
More recently, it has shown how S. cerevisiae-based probiotics may also be used for the prevention and treatment of vulvovaginal candidiasis (VVC), a pathology which affect about 3-quarters of women during their reproductive age and subjected to frequent recurrencies [7]. Since the vaginal microbiome plays a relevant role in VVC, a probiotic-based approach could represent a valid alternative to the conventional drug therapy, also considering the resistance of some Candida strains to many antifungals and their toxicity for the patients [8].
The effects of Saccharomyces-based treatment against VVC has been investigated both in vitro, and in vivo [9-12], definitely proving its beneficial protective and therapeutic action.
In this work we isolated from a sourdough and then characterized a new yeast strain, which resulted to be Kluyveromyces marxianus, and assessed its potential probiotic and safety properties, compared to those exhibited by the commercial probiotic S. boulardii.
K. marxianus isolates from different sources have been reported to have beneficial effects on human health. They include cholesterol reducing capability [13,14], antiproliferative effects on cancer cells [15,16], immunomodulatory action on Caco-2 cells [17], and also antimicrobial activity against various pathogen species [16,18,19]. In particular, the antifungal activity of different K. marxianus strains has been investigated and demonstrated against plant pathogenic fungi [20,21] and food spoilage moulds present in dairy products, so that the yeast has been proposed as a potential biocontrol agent [22]. Instead, so far, the anticandidal activity of this species has not been investigated in detail, being reported only in the case of a K. marxianus strain isolated from a French cheese [18].
In a previous work we isolated several C. albicans strains from VVC patiens and characterized the clinical isolates in terms of antifungal resistance and biofilm forming ability [23], a characteristic associated to Candida virulence.
Therefore, here, aiming at enlarging the number and applications of available probiotic yeasts, we propose the new K. marxianus isolate as a novel anticandidal agent, testing its ability to antagonize fluconazole resistant Candida albicans vaginal strains, and presenting some insights into the mechanisms underlying its protective effect.
2. Materials and Methods
2.1 Strains and culture condition
For isolation of K. marxianus, 10 g of sourdough were suspended in 90 mL of PBS and homogenized with Fisherbrand 850 Homogenizer (Thermo Fisher Scientific, Waltham, MA, USA) for 20 min. The suspended sample was serially diluted in phosphate buffer saline solution (PBS) and plated onto Rosa Bengal agar (RB, Sigma-Aldrich, St. Louis, MO, USA), supplemented with chloramphenicol. The plates were incubated at 25 °C for 96 h until typical creamy yeast colonies appeared.
For most experiments, a strain of S. boulardii of commercial origin (Biocodex, Zambon, Italy) was used. It was obtained dissolving a capsule of the commercial product in PBS and then plating a proper diluted suspension on RB agar plates. After incubation of plates at 25 °C for 48 h, colonies appeared, and typical yeast morphology was checked by microscope observation.
Pre cultures and cultures of both K. marxianus and S. boulardii was performed in shake flasks containing YPD (yeast extract 2%, bactopeptone 1%, dextrose 2%) medium, starting from one single colony, and incubated at 25 °C for 24 h at 200 rpm. The strains of C. albicans used were: ATCC 90028, and four clinical C. albicans strains, namely C7, C14, C17, and C19 isolated from VVC patients [23]. All C. albicans strains were cultured in YPD or RPMI (Thermo Fisher Scientific Inc., Waltham, MA, USA) medium at 37 °C.
In some experiments, to distinguish C. albicans from K. marxianus colonies, the selective chromogenic medium Brillance Candida Agar Base (Oxoid Ltd, Wade Road, Basingstoke, Hants, RG24 8PW, UK) was used, which is properly formulated so that C. albicans colonies, after incubating agar plates at 37 °C, appear green, whereas K. marxianus colonies appear yellow/brown.
2.2. Molecular characterization of the yeast isolate
Isolated yeast colonies were, was sub-cultured ion Dichloran Rose Bengal Chloramphenicol agar (DRBC, Thermo Fisher Scientific Inc., Waltham, MA, USA). CTAB extraction protocol [24] was employed to extract fungal total DNA. Extracted DNA was amplified with PCR, targeting ITS-5.8S rDNA region of the fungal 18S rRNA gene (750 bp amplicon size) [25], employing a MiniAmp™ Thermal Cycler (Thermo Fisher Scientific Inc., Waltham, MA, USA)TECHNE Prime Thermal Cycler, and disposing of a ITS1_f (5’-GGA AGT AAA AGT CGT AAC AAG G-3’ 5’-TCC GTA GGT GAA CCTGCG G-3’) and ITS4_r (5’-TCC TCC GCT TAT TG A TAT GC-3’) primer set (Biofab Research, Rome, Italy). Templates were run on a 1.5% agarose gel, stained with GelRed (BIOTIUM), utilising a 100 bp DNA ladder as a reference. Sanger Sequencing reactions were performed by an external service (Biofab Research, Rome, Italy); the obtained FASTA sequences were interpreted using an editing tool, Chromas Lite v. 2.6.6 (Technelysium Pty Ltd., South Brisbane, Australia), and compared to NCBI Sequence Database sequences, using BLASTN ver. 2.2.29 (also referring to GenBank), selecting the highest percentage Identity, with a 98% cut-off and 0.0 e-value
2.3. Probiotic Evaluation of the K. marxianus isolate
2.3.1. Growth at 37 °C
To test the ability of the K. marxianus isolate and the commercial strain of S. boulardii to grow at the temperature of the human body, 37 °C, both strains were cultivated at this temperature in shake flasks in YPD for 24 h. Growth was assessed by OD590 determination.
2.3.2. Acid and Bile Salt Tolerance
Acid and bile salt tolerance of K. marxianus and S. boulardii were determined as already reported [26]. Yeast cells were collected from a 18 h preculture in YPD at 37 °C, washed with PBS and resuspended in 5 mL PBS plus HCl (pH = 3.0) or PBS plus 0.3% w/v bile salts (Sigma Aldrich St. Louis MO, USA), so as to have an initial cell density of 108 cells mL-1. After 4 h incubation at 37 °C, 150 rpm, the CFU mL-1 were determined using the standard plate count method, and the percentage of residual viable cells was calculated with respect to time 0 of incubation (100%)
2.3.3. Auto-Aggregation Capacity
The auto-aggregation capacity of both K. marxianus isolate and S. boulardii was determined after 24 h according to Maione et al. [26], by determining the OD590 of the yeast suspension before (OD0) and after (ODt) incubation. The auto-aggregation (A) percentage was calculated as:
2.3.4. Hydrophobicity Assay
Cell biomass was suspended in 5 mL of PBS and absorbance at 590 nm was measured
(OD0). Then, 3 mL xylene, were added to each sample. After incubation at 37 °C for 60 min without shaking, the absorbance of the interphase was measured (ODf) [27]. The hydrophobicity percentage was calculated as follows:
2.3.5. Antioxidant Activity
Radical scavenging activity of K. marxianus and S. boulardii was evaluated by 2,2- diphenyl-1-picrylhydrazyl (DPPH) free radical assay [28]. The degree of discoloration of DPPH indicates the scavenging potential of the strain. A 0.2 mM DPPH solution was freshly prepared in methanol 800 μL of each strain suspension was mixed with 1 mL of 0.2 mmol L₋1. DPPH solution in methanol. The samples were incubated for 30 min in the dark at room temperature with agitation and then centrifuged at 2000 rpm for 2 min. The supernatant absorbance was measured at 517 nm. In parallel, ascorbic acid was used as positive control. The scavenging ability (SA) of the strain was calculated using the formula:
2.3.6. Assessment of biofilm formation
2.3.7. A-431 cells
The human epithelial A-431 cell line derived from a vaginal epithelial squamous cell carcinoma was used. The cell line was purchased from DSZM (Germany). The cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Sigma Aldrich Co., St. Louis, MO, USA) supplemented with L-glutamine (2 mM) (Sigma Aldrich Co., St. Louis, MO, USA) , 1% w/v penicillin-streptomycin ((Sigma Aldrich Co., St. Louis, MO, USA), and heat-inactivated Fetal Bovine Serum (FBS), at 10% w/v (Sigma Aldrich Co., St. Louis, MO, USA); specifically, the cell line was kept in culture by-passages in fresh medium twice a week and incubated at 37 °C and 5% CO2.
2.3.8. Adhesion to A-431 cells
To allow adhesion to human cells, K. marxianus and S. boulardii cells at a density of 106 cells mL-1 were added to the cell monolayer in a 48 well multiwell plate and incubated for 2 h in DMEM at 37 °C. Then, wells were washed with PBS, and 100 μL di tripsin/EDTA solution (Sigma Aldrich Co., St. Louis, MO, USA), was added. After 5-6 min incubation, 900 μL di DMEM were added for trypsin inactivation. The content of the wells was serially diluted in PBS and plated onto chloramphenicol RB agar for the yeast cell count.
Also, invasiveness of K. marxianus and S. boulardii into A-431 cells was determined, following the above described protocol, but, before trypsin treatment, the infected A-431 cells were incubated with 0,5 mg mL-1 amphotericin B for 1 h, to eliminate the adhered cells on the surface and allow to count only the internalized yeast cells.
2.4. Safety Assessment of the K. marxianus isolate
2.4.1. Haemolytic activity
The haemolytic activity of the three isolates was carried out according to the method
described by Shen et al 2022 [31], by inoculating the strains on blood agar plates (Oxoid Ltd., Basingstoke, UK) containing 5% defibrinated sheep blood for 48 h of incubation at 37 °C in order to detect patterns of haemolysis. Next, cultured plates were observed to assess a haemolytic zone, and a clear zone of hydrolysis around the colonies was considered as a positive result (β-haemolysis).
2.4.2. Antibiotic and antifungal resistance
Antimicrobial resistance was screened by agar disk diffusion method as described by Turchi et al 2013 [32], in accordance with the Clinical and Laboratory Standards (M44, M45). The isolated yeast strain was tested for resistance against the most commonly used antibacterial and antifungal compounds, 12 belonging to the first category and 5 to the latter, each of which has a different mode of action. The molecules tested were gentamicin (10 μg GM), ampicillin (10 μg AMP), polymyxin-B (300 U PB), amoxycillin + clavulanic acid (30 μg AMC), norfloxacin (10 μg NOR), streptomycin (10 μg STR), cephotaxime (30 μg CTX),ciprofloxacin (10 μg CIP), azithromycin (15 μg AZ), ceftriaxone (30 μg CRO), vancomycin (20 μg VAN), rifampicin (20μg RIF), fluconazole (10μg FL) caspofungin (2 μg CSF), ketoconazole (30 μg KET), itraconazole (30 μg ITR), and amphotericin B (2 μg Amph B). After 24–48 h incubation at 30 °C, the diameter of the inhibition zone was measured and the strains were considered susceptible (S), intermediate (I), or resistant (R).
2.4.3. Galleria mellonella survival assay
To evaluate the safety of the K. marxianus isolate in vivo, larvae of G. mellonella were used. For each assay, 20 healthy larvae of a similar size, were selected and injected with 10 μL of the K. marxianus suspension in PBS through the last left pro-leg, at a density corresponding to 104, 105, 106, 107 yeast cells/larva. A similar procedure was performed for the commercial S. boulardii. Intact larvae or larvae injected with PBS alone were used as controls. All samples were incubated at 37 °C for 120 h Every 24 h the number of viable larvae was registered, and the survival percentage was calculated. Larvae were considered dead when they displayed no response to touch.
2.5. Anticandidal activity of the K. marxianus isolate
2.5.1. Co-aggregation of K. marxianus with C. albicans
The co-aggregation of the K. marxianus isolate with C. albicans, was determined as previously described [26]. Briefly, 4 mL of a K. marxianus overnight culture and 4 mL of C. albicans overnight culture were mixed, vortexed for 10 s, and incubated for 5 and 24 h at 37 °C. Each control tube contained 4 mL of each single suspension. The absorbance (OD) of the mixed suspension was then measured at 590 nm (ODmix) and compared with those of the control tubes containing K. marxianus (ODstrain) and the pathogen (ODpathogen) after incubation. The percentage of co-aggregation (C) was calculated as:
2.5.2. Exclusion test
The ability of pre-established K. marxianus biofilms to prevent the adhesion and biofilm formation of C. albicans on polystyrene microplates was evaluated performing an “exclusion test” according to Cisneros et al. 2021 [33], with some modifications. K. marxianus cells derived from overnight cultures in YPD were adjusted to 106 CFU mL-1. And seeded in individual wells of a 96 well multiplate. Microplates were first incubated at 37 °C for 24 to allow cells to attach to the surface of the wells. Thereafter, supernatants were discarded, and a suspension of each C. albicans strain in RPMI 106 CFU mL-1 was added to each well containing the K. marxianus pre-formed biofilm. Microplates were incubated at 37 °C for 24 h. Then, wells were carefully washed twice with PBS and then individually scraped to remove all the biofilm biomass. Cells recovered from each well were suspended, diluted in PBS, and then plated on Brillance Candida Agar Base plates to distinguish and determine the number of viable cells of C. albicans and K. marxianus, respectively. The plates were incubated at 37 °C for 48 h. Then, for each strain, the number of colonies on the plates was determined. Results were expressed as Log10 (CFU well-1). Biofilm formation of C. albicans strains was used as control.
2.5.3. Germ tube test in the presence of K. marxianus
The ability of C. albicans to form germ tubes was assessed using the germ tube test according to Pericolini et al. [9]. Briefly, an isolated colony of C. albicans of each strain (ATCC90028, C7, C14, C17, and C19) taken from a 24 h YPD agar plate, was inoculated into a total volume of 0,5 mL of RPMI plus 10% Fetal Bovine Serum (Fisher Thermo Scientific, Cleveland, OH, USA) in the well of a 24-well multi plate and incubated for 4 h at 37 °C, in the presence or not of 108 cells mL-1 of K. marxianus resuspended in RPMI medium. After incubation, the wells were examined at JuLI™ Stage inverted microscope (Nanoentek) at x20 magnification.
2.5.4. Adhesion of C- albicans to A-431 cells in the presence of K. marxianus
To investigate the probiotic effects of K. marxianus on Candida adhesion to A-431 cells, we follow the protocol by Kunyet et al. [11] with some modifications. The assays of adhesion on human cells were performed under pre-inoculation, co-inoculation, and post-inoculation conditions in 48 well microplates where A-431 cells in DMEM were at 80% confluency. In all cases the density of the probiotic yeast and of each C. albicans strain (ATCC 90028, C7, C14, C17, and C19) was 108 cells mL-1 and 107 cells mL-1, respectively.
In the pre-inoculation condition, K. marxianus was inoculated into wells for 2 h at 37 °C. Then, the Candida strain was introduced and incubated for an additional 2 h with mild shaking (90 rpm). In the co-inoculation condition, the probiotic strain was co-inoculated with the Candida strain and incubated for 2 h. In the post-inoculation condition, the Candida strain was inoculated for 2 h before inoculation of K. marxianus for further 2 h.
At the end of incubation, the wells were washed with PBS, and their content was serially diluted in PBS. Dilution were plated on Brillance Candida Agar Base to distinguish C. albicans from K. marxianus colonies, and count residual C. albicans colonies. Results were expressed as percentage inhibition of the adhesion, with respect to controls performed in the same condition without the inoculation of the probiotic.
2.5.5. Cell damage: LDH determination
A-431 Cells were then incubated in the presence or absence of K. marxianus (OD590 = 5) for 2 h at 37 °C plus 5% CO2, extensively washed with PBS and then incubated with C. albicans ATCC 90028 or C. albicans C19 (1×106 mL-1) for 18 h at 37 °C plus 5% CO2.
After incubation, the epithelial cell damage was determined by the release of lactate dehydrogenase (LDH) into the surrounding medium. Therefore, the medium was collected and LDH was measured spectrophotometrically at 492 nm using a Lactase Dehydrogenase Activity Assay kit (Sigma-Aldrich, St. Louis, MO, USA). The percent cell damage of A-431 infected with C. albicans was calculated as follows:
and compared to 100% C. albicans damage induced in each cell type [34]. The lysis buffer was used as positive control to determine the maximum LDH activity.
2.5.6. Quantitative analysis of SAP2 and SAP6 gene expression
To check the expression of SAP2 and SAP6 genes of C. albicans in the presence of K. marxianus , aliquots of C. albicans cells of ATCC 90028 and C19 strains (106 mL-1) were incubated in YPD medium or YPD medium plus 1% of bovine serum albumin (BSA), for 24 h at 37 °C under agitation (150 rpm) in the presence or absence of K. marxianus cells (OD.590=5.0) [9]. Briefly, total RNA was extracted using Direct-zolTM RNA Miniprep Plus Kit (ZYMO RESEARCH). Then, 1 μg of RNA was reverse transcribed to cDNA (Bio-Rad, Milan, Italy) and analyzed by quantitative PCR run in AriaMx Real-Time PCR instrument (Agilent Technologies, Inc., Milan, Italy) according to the manufacturer’s instructions. For real-time PCR reaction 100 ng of cDNA was used. PCR System used: 95 °C for 10 min, one cycle for cDNA denaturation; 95 °C for 15 s and 60 °C for 1 min, 40 cycles for amplification; 95 °C for 15 s, one cycle for final elongation; and one cycle for melting curve analysis (from 60 °C to 95 °C) to verify the presence of a single product. C. albicans SAP2 and SAP6 genes were detected by using primers previously reported [35]. The expression levels of genes were evaluated using the 2−∆∆CT method, where Ct was the average threshold cycle number from three independent experiments. Data were presented as the fold change in gene expression normalized to the ACT1 gene as internal control [36,37].
2.6. Statistical Analyses
GraphPad Prism Software (version 8.02 for Windows, GraphPad Software, La Jolla, CA, USA, www.graphpad.com, accessed on 14 February 2024) was used for data analysis. All data are shown as mean +/- standard deviation (SD) and were derived from two or three independent experiments. One-way or two-way analysis of variance (ANOVA) followed by Tukey’s test was used for the comparison test. The Kaplan–Meier method and log-rank (Mantel–Cox) test were used to plot G. mellonella survival curves.
3. Results
3.1. Isolation and molecular characterization of K. marxianus isolate
The yeast isolate from sourdough was subjected to molecular identification. Results of Sanger Sequencing of the yeast colonies is reported in Table 1. The isolate resulted to be Kluyveromyces. marxianus.
3.2. Probiotic properties of the K. marxianus isolate
Some characteristics of the K. marxianus isolate were determined to assess the potential probiotic properties of the strain. They were: ability to grow at 37 °C, tolerance to gastric and bile acids, auto-aggregation, hydrophobicity, antioxidant power, biofilm forming capacity, and adhesion to epithelial cell lines. In parallel, the same tests were conducted with the commercial S. boulardii (Biocodex) strain.
On the whole, these tests are indicative of the adaptation of the isolate to the human body (growth at 37 °C, resistance to acid and bile salts), with potential benefit effect (antioxidant power) and the ability to adhere and colonize the epithelial mucosal membranes (hydrophobicity, auto-aggregation, biofilm forming ability, adhesion).
The results of these tests were reported in Table 2, where a score from 0 (minimum) to 2 (maximum) was given to the result of each test response for both the K. marxianus isolate and S. boulardii.
For a better visualization of the overall probiotic characteristics of each strain, the indicators in Table 2, were reported in radar diagrams (Figure 1).
The K. marxianus isolate showed very good probiotic properties (total score 13,5), even better than the commercial S. boulardii strain (total score 11,4). Most characteristics result similar between the two yeasts, but the K. marxianus isolate showed a better adaptation to the body temperature (37 °C), a much better ability to form biofilm in vitro, and a higher adhesion to A-431 cells than S. boulardii (Table 2 and Figure 1). Though adhesion to A-431 cells was not so high, it is worth of note that invasiveness of both the strains into the A-431 cells was absent (data not shown).
3.3. Safety assessment
Safety of the K. marxianus isolate was assessed in vitro by testing both its haemolytic activity on blood agar plates and resistance to antibiotics and antifungals.
As shown in Figure 2, no haemolytic activity was shown by the K. marxianus isolate, as well as by the S. boulardii strain.
As regards resistance to antimicrobials, the yeast isolate was resistant to all the most common antibiotics, and sensitive to the tested antifungals, except for itraconazole (ITR). A similar response was obtained by S. boulardii (Table 3).
Due to their simplicity of use and low cost, the larvae of G. mellonella represent an optimal model to evaluate the virulence of fungi and other microbial species [38], so we used them to assess the safety in vivo of the K. marxianus isolate. Larvae survival after injection of the yeast strain at different final cell densities (from 104 to 107 yeast cells per larva) was determined. In parallel the same test was carried out with the S. boulardii strain. No effect of the yeast injection was observed in the case of the K. marxianus isolate, since larvae survival remained 100% up to 120 h incubation, likewise to what observed in the case of the commercial probiotic S. boulardii (Figure 3).
3.4. Anticandidal activity of K. marxianus
Co-aggregation of a probiotic with a pathogen is indicative of its capacity to prevent subsequent adhesion and colonization, therefore we first tested the ability of the K. marxianus isolate to co-aggregate with C. albicans. The percentages of co-aggregation were significant, being about 50% with both ATCC 90028 and C19 strain after 5 h incubation and increased up to 75% for both strains after 24 h.
Then, we wanted to assess the effect of the putative probiotic K. marxianus on filamentation of C. albicans strains, considering that filamentation is a key virulence factor for Candida species and positively correlated with adhesion and biofilm formation. In Figure 4 the results of a germ test of C. albicans ATCC and C. albicans C19 in the presence of K. marxianus are shown. A clear full inhibition of the germ tube formation was observed for both the strains. Similar results were obtained with all the other C. albicans vaginal strains (data not shown).
As previously reported [23], all the C. albicans strains used in this work were able to form biofilms in vitro on polystyrene microplates. Here, we wanted to assess the ability of a pre-established biofilm of the K. marxianus isolate to prevent such biofilm formation. So, the formation of biofilms of C. albicans ATTC 9028 and the clinical strains were evaluated in the so called “exclusion test”. The results clearly indicated that surface colonization and ability to form biofilms was significantly reduced in the presence of a pre-formed K. marxianus biofilm for all the C. albicans strains examined (Figure 5).
Also , we tested the effect of the putative probiotic on the adhesion of Candida strains on A-431 cells, investigating different conditions of treatment: pre-inoculation, where the probiotic yeast was added to the cell monolayer prior to application of the Candida strains; co-inoculation, where the live probiotic yeast and Candida strains were simultaneously applied; and post-inoculation, where probiotic yeast treatment was performed after the application of Candida strains. In the three conditions and for all the strains examined, the reduction of adhesion was clear (Figure 6), and resulted very high (90%) for the strains C7, C14 and C19 in the condition of co-inoculation.
The possible protective effect of K. marxianus against the damage provoked by C. albicans infection in A-431 cells was also ascertained, determining the LDH release from the cell line preincubated with the K. marxianus isolate and then infected with C. albicans ATCC 90028 or C. albicans C19 for 18 h. The results showed a clear prevention of the cell damage provoked by the C. albicans strains in the presence of the probiotic yeast (Figure 7). It is worth noting that the K. marxianus isolate did not exert any significative cytotoxic effect on A-431 cells, since the LDH released from cells treated with the probiotic alone was comparable with the spontaneous release of LDH (data not shown).
Since aspartyl proteases exert a main contribution to virulence and invasiveness of C. albicans, we checked the expression of two of the genes (SAP2 and SAP6) encoding for such proteins when C. albicans was co-incubated with the putative probiotic K. marxianus isolate. The results of the RTqPCR reported in Figure 8 clearly show that in the presence of the isolate the expression of both SAP2 and SAP6 resulted greatly reduced both in the ATCC and the clinical C19 strain.
4. Discussion
In the recent years the interest towards the probiotic potential of non-conventional yeasts has greatly increased. Non-Saccharomyces yeasts mainly belonging to the genera of Pichia, Debaryomyces, Wickerhamomyces, Zygosaccharomyces and Kluyveromyces have been isolated from different sources and tested as probiotics, in alternative to the well-established commercial species S. cerevisiae var. boulardii [6,39].
The food-grade yeast K. marxianus is characterized by rapid growth rate, easy cultivation, thermotolerance, versatile metabolism, and ability to secrete lytic enzymes, which make this species particularly suitable for many food and biotechnological applications [40,41].
As specifically regards the probiotic properties of K. marxianus, they were ascertained since 2011, when it was shown that the yeast strain isolated from a dairy product [17] was able to adhere to human enterocyte-like Caco-2 cells and modulate colon microbiota, increasing the bifidobacterial concentration, and inducing the formation of higher amounts of the short-chain carboxylic acids acetate and propionate. The strain was K. marxianus B0399, which was subsequently introduced in the market as a new generation of probiotic lactic yeast (Turval B0399®).
Later on, other authors have reported several other beneficial effects of K. marxianus mainly isolated from dairy products [42]. Very recently Nag et al. [43] observed in vitro that this yeast improved insulin sensitivity and reduced fat storage in fat cells, suggesting benefits for type 2 diabetes and obesity. Moreover, K. marxianus produce high value-added bioingredients (oligo-saccharides, -nucleotides, -peptides) that, used as prebiotics, have revealed to increase the intestinal microbiota or added to foods, enhance flavours, stabilize food emulsions, act as immunopotentiators, have hypocholesterolemic action, and promote protection against bacterial infections [44].
In this work, we isolated a K. marxianus strain from a sourdough used in an artisanal bakery as starter for the leaving of bread. This practice, which has recently undergone a revival also in industrial bakeries, exploits the spontaneous fermentation of yeasts and lactobacilli present in the sourdough, and confers the final product a distinctive taste and aroma.
Aiming at proposing the new K. marxianus isolate as a potential probiotic, we performed an initial in vitro screening of the properties necessary to adapt to the human body with beneficial effects. The tests were done in parallel with a reference probiotic strain, the commercial S. boulardii (Biocodex). To comprehensively evaluate and visualize the results, radar charts were used [31],which clearly indicate that the new strain has excellent probiotic properties, a result which is agreement with those reported for other K. marxianus isolates [22]. The comparison showed that K. marxianus better adapts to the body temperature and form thicker biofilm in vitro. A good biofilm ability on abiotic surfaces has been reported in the case of several K. marxianus strains from different dairy products, and it is considered an essential trait for the outcome of the final product [45]. As regards the adhesion to vaginal epithelial cells, it resulted higher than that of S. boulardii on the same cells, but not so high (10%). Many different values of adhesion to Caco-2 cells have been reported for K. marxianus, varying from 8 to 19 % depending on the strain [46].
Adhesion of probiotics to epithelial cells is generally viewed as a prerequisite for their colonization, which may constitute a physical barrier against the adhesion of pathogens to the mucosa. However, also low-adhering S. cerevisiae strains have shown to display protective effects towards bacterial infections, via reduction of the intestinal pro-inflammatory response [16]. For some authors [47,48] a good adhesion of a probiotic strain is even considered an undesiderable virulence factor, because preceding the possible invasion of the strain into the human cells. In our case, it is worth of note that invasion of K. marxianus into A-431 cells was totally absent.
Safety evaluation was conducted to confirm the non-pathogenic nature of the K. marxianus isolate: the yeast resulted non-haemolytic, resistant to all the most common antibiotics, and sensitive to the tested common antifungals. Then, to assess the safety of the K. marxianus isolate also in vivo, we used the G. mellonella model. Indeed, G. mellonella larvae have been established as an animal model alternative to mammals to test the pathogenity of fungal species since 2000 [49], and more recently used for the study of the safety of probiotic putative strains in vivo [26,50]. Our results clearly showed that the survival of the Galleria larvae was not affected at all by the injection with K. marxianus, even at the highest density tested, similarly to what observed with S. boulardii. So, the in vivo tests unequivocally assessed the safety of the new K. marxianus isolate.
The opportunistic pathogen yeast C. albicans is the agent of the vast majority of VVC, often associated to strains resistant to conventional antifungals. In the perspective of a possible use of K. marxianus as a novel probiotic for both prevention and therapy of VVC, we analysed the antagonistic activity of the yeast isolate against some vaginal strains of C. albicans.
Our results clearly showed the co-aggregation ability of K. marxianus with C. albicans and a full inhibition of the germ tube formation upon treatment with the live yeast, a result in agreement with other authors who found the same effects with Saccharomyces strains [9,10]. It is well known that the polymorphic C. albicans may shift from the yeast-like morphology to germ tube, pseudo-hyphae and hyphae. Such filamentation is pivotal for biofilm formation and epithelial invasion, representing a well-known virulence factor for Candida [51].
Further, and in accordance with this result, we demonstrated a reduction of biofilm formation in vitro for all the tested C. albicans strains in the presence of K marxianus preformed biofilm. Such “exclusion effect” [33] is indicative of a clear antagonism between the putative probiotic and Candida, due to a physical block for the pathogen colonization on the abiotic surface, without excluding the contribution of other mechanisms of action.
The adhesion of Candida to epithelial vaginal cells is certainly mandatory for invasiveness and pathogenesis, so we also investigated the effect of K. marxianus on Candida adhesion to A-431 cells. It resulted significantly impaired by the presence of the live putative probiotic, which showed its efficacy in both inhibition of adhesion and displacement of Candida cells, suggesting its potential use in both preventing and ameliorating Candida infections. These results are in agreement with those obtained by Kunyet et al. [11] with Saccharomyces strains able to hinder Candida adhesion both on polystyrene surfaces and A-431 cell lines.
Indeed, the presence of K. marxianus isolate protected the epithelial cells from the damage induced by C. albicans, as shown by the less amount of LDH released from A-431 cells pretreated with the probiotic strain and then infected with the pathogen.
The secretion of aspartyl proteases encoded by SAP genes plays an essential role in the development of pathogenesis of C. albicans, being involved in the shift of the yeast to hyphal morphology which is determinant for invasiveness [52]. Consistently with the observed inhibition of the germ tube formation, we also demonstrated the ability of the probiotic isolate to reduce the expression of SAP2 and SAP6 genes in Candida under conditions of serum-induced hyphal differentiation. This result is in agreement to what already observed for other yeasts proposed as anticandidal probiotic agents [9,10].
Apparently, K. marxianus is able not only to protect the epithelial cells from adhesion via a mechanical hindrance to Candida colonization, but also to affect some virulence factors such as hyphal differentiation and secretion of aspartyl proteases, presumably due to products of its metabolism and/or competition for those serum components inducing Candida hyphal morphogenesis. Further investigations are necessary to shed light in the details of Kluyveromyces/Candida interactions.
In conclusion, our results enlarge the spectrum of the beneficial properties of the food-grade yeast K. marxianus showing for the first time its biotherapeutic potential in the prevention and treatment of VVC.
Author Contributions
Conceptualization, E.d.A. and E.G.; methodology, E.d.A.; software, A.M.; validation, A.M. and A.B.; formal analysis, A.B. and F.C.; investigation, A. B., M.I., and F.C.; resources, M.G.; data curation, A.M. writing—original draft preparation, E.d.A.; writing—review and editing, E.d.A., E.G. and A.B.; visualization, E.G.; supervision, E.d.A.; project administration, M.G.; funding acquisition, E.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by funding from PRIN2022—Financed by European Union—Next Generation EU. Serum metabolomics in atopic dermatitis (MetabAD): the potential of a GC-MS based strategy to improve the clinical diagnosis, the understanding of the disease, and the treatment efficacy (No. 2022HC3FRM).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in thearticle, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: from fundamental to novel applications. Front Microbiol 2012, 3, 421. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Hsia, K.-C.; Kuo, Y.-W.; Chen, S.-H.; Huang, Y.-Y.; Li, C.-M.; Hsu, Y.-C.; Tsai, S.-Y.; Ho, H.-H. Safety Assessment and Probiotic Potential Comparison of Bifidobacterium longum subsp. infantis BLI-02, Lactobacillus plantarum LPL28, Lactobacillus acidophilus TYCA06, and Lactobacillus paracasei ET-66. Nutrients 2024, 16, 126. [Google Scholar] [CrossRef]
- Ryabtseva, S.A.; Khramtsov, A.G.; Sazanova, S.N.; Budkevich, R.O.; Fedortsov, N.M.; Veziryan, A.A. The Probiotic Properties of Saccharomycetes (Review). Applied Biochemistry and Microbiology 2023, 59, 111–121. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? Journal of Fungi 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Ebrahimi, M.; Shahryari, S.; Kharazmi, M.S.; Jafari, S.M. Food applications of probiotic yeasts; focusing on their techno-functional, postbiotic and protective capabilities. Trends in Food Science & Technology 2022, 128, 278–295. [Google Scholar] [CrossRef]
- Shruthi, B.; Deepa, N.; Somashekaraiah, R.; Adithi, G.; Divyashree, S.; Sreenivasa, M.Y. Exploring biotechnological and functional characteristics of probiotic yeasts: A review. Biotechnology Reports 2022, 34, e00716. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: a systematic review. Lancet Infect Dis 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Akinosoglou, K.; Schinas, G.; Polyzou, E.; Tsiakalos, A.; Donders, G.G.G. Probiotics in the Management of Vulvovaginal Candidosis. Journal of Clinical Medicine 2024, 13, 5163. [Google Scholar] [CrossRef]
- Pericolini, E.; Gabrielli, E.; Ballet, N.; Sabbatini, S.; Roselletti, E.; Cayzeele Decherf, A.; Pélerin, F.; Luciano, E.; Perito, S.; Jüsten, P.; et al. Therapeutic activity of a Saccharomyces cerevisiae-based probiotic and inactivated whole yeast on vaginal candidiasis. Virulence 2017, 8, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, E.; Pericolini, E.; Ballet, N.; Roselletti, E.; Sabbatini, S.; Mosci, P.; Decherf, A.C.; Pélerin, F.; Perito, S.; Jüsten, P.; et al. Saccharomyces cerevisiae-based probiotic as novel anti-fungal and anti-inflammatory agent for therapy of vaginal candidiasis. Beneficial Microbes 2018, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Kunyeit, L.; Kurrey, N.K.; Anu-Appaiah, K.A.; Rao, R.P. Probiotic Yeasts Inhibit Virulence of Non<i>-albicans Candida</i> Species. mBio 2019, 10, 10.1128–mbio.02307. [Google Scholar] [CrossRef]
- Tomičić, R.; Tomičić, Z.; Raspor, P. Influence of culture conditions on co-aggregation of probiotic yeast Saccharomyces boulardii with Candida spp. and their auto-aggregation. Folia Microbiologica 2022, 67, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-J.; Kim, D.-H.; Jeong, D.; Seo, K.-H.; Jeong, H.S.; Lee, H.G.; Kim, H. Characterization of yeasts isolated from kefir as a probiotic and its synergic interaction with the wine byproduct grape seed flour/extract. LWT 2018, 90, 535–539. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, H.; Liu, H.; Xiong, L.; Gao, X.; Jia, H.; Lian, Z.; Tong, N.; Han, T. Hypocholesterolemic effects of Kluyveromyces marxianus M3 isolated from Tibetan mushrooms on diet-induced hypercholesterolemia in rat. Brazilian Journal of Microbiology 2015, 46. [Google Scholar] [CrossRef] [PubMed]
- Saber, A.; Alipour, B.; Faghfoori, Z.; Yari Khosroushahi, A. Secretion metabolites of dairy Kluyveromyces marxianus AS41 isolated as probiotic, induces apoptosis in different human cancer cell lines and exhibit anti-pathogenic effects. Journal of Functional Foods 2017, 34, 408–421. [Google Scholar] [CrossRef]
- Fadda, M.E.; Mossa, V.; Deplano, M.; Pisano, M.B.; Cosentino, S. In vitro screening of Kluyveromyces strains isolated from Fiore Sardo cheese for potential use as probiotics. LWT 2017, 75, 100–106. [Google Scholar] [CrossRef]
- Maccaferri, S.; Klinder, A.; Brigidi, P.; Cavina, P.; Costabile, A. Potential probiotic Kluyveromyces marxianus B0399 modulates the immune response in Caco-2 cells and peripheral blood mononuclear cells and impacts the human gut microbiota in an in vitro colonic model system. Appl Environ Microbiol 2012, 78, 956–964. [Google Scholar] [CrossRef]
- Ceugniez, A.; Drider, D.; Jacques, P.; Coucheney, F. Yeast diversity in a traditional French cheese “Tomme d'orchies” reveals infrequent and frequent species with associated benefits. Food Microbiology 2015, 52, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Ceugniez, A.; Coucheney, F.; Jacques, P.; Daube, G.; Delcenserie, V.; Drider, D. Anti-Salmonella activity and probiotic trends of Kluyveromyces marxianus S-2-05 and Kluyveromyces lactis S-3-05 isolated from a French cheese, Tomme d'Orchies. Research in Microbiology 2017, 168, 575–582. [Google Scholar] [CrossRef]
- l-Qaysi SAS, A.N. , Jaffer MR, Abbas ZA. Biological Control of Phytopathogenic Fungi by Kluyveromyces marxianus and Torulaspora delbrueckii Isolated from Iraqi Date Vinegar. Journal of Pure and Applied Microbiology 2021, 15(1), 300–311. [Google Scholar] [CrossRef]
- Dos Santos, A.M.; Albuini, F.M.; Barros, G.C.; Pereira, O.L.; da Silveira, W.B.; Fietto, L.G. Identification of the main proteins secreted by Kluyveromyces marxianus and their possible roles in antagonistic activity against fungi. FEMS Yeast Res 2023, 23. [Google Scholar] [CrossRef]
- Goktas, H.; Dikmen, H.; Demirbas, F.; Sagdic, O.; Dertli, E. Characterisation of probiotic properties of yeast strains isolated from kefir samples. International Journal of Dairy Technology 2021, 74, 715–722. [Google Scholar] [CrossRef]
- Maione, A.; Imparato, M.; Buonanno, A.; Carraturo, F.; Schettino, A.; Schettino, M.T.; Galdiero, M.; de Alteriis, E.; Guida, M.; Galdiero, E. Anti-Biofilm Activity of Phenyllactic Acid against Clinical Isolates of Fluconazole-Resistant Candida albicans. Journal of Fungi 2023, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Salbitani, G.; Chianese, M.R.; Bossa, R.; Bencivenga, T.; Carraturo, F.; Nappo, A.; Guida, M.; Loreto, F.; Carfagna, S. Cultivation of barley seedlings in a coffee silverskin-enriched soil: effects in plants and in soil. Plant and Soil 2024, 498, 199–211. [Google Scholar] [CrossRef]
- Korabečná, M.; Liška, V.; Fajfrlík, K. PrimersITS1, ITS2 andITS4 detect the intraspecies variability in the internal transcribed spacers and 5.8S rRNA gene region in clinical isolates of fungi. Folia Microbiologica 2003, 48, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Maione, A.; Imparato, M.; Buonanno, A.; Salvatore, M.M.; Carraturo, F.; de Alteriis, E.; Guida, M.; Galdiero, E. Evaluation of Potential Probiotic Properties and In Vivo Safety of Lactic Acid Bacteria and Yeast Strains Isolated from Traditional Home-Made Kefir. Foods 2024, 13, 1013. [Google Scholar] [CrossRef] [PubMed]
- SYAL P., V. A. PROBIOTIC POTENTIAL OF YEASTS ISOLATED FROM TRADITIONAL INDIAN FERMENTED FOODS. International Journal of Microbiology Research 2013, 5, 390–398. [Google Scholar] [CrossRef]
- Merchán, A.V.; Benito, M.J.; Galván, A.I.; Ruiz-Moyano Seco de Herrera, S. Identification and selection of yeast with functional properties for future application in soft paste cheese. LWT 2020, 124, 109173. [Google Scholar] [CrossRef]
- Maione, A.; Galdiero, E.; Cirillo, L.; Gambino, E.; Gallo, M.A.; Sasso, F.P.; Petrillo, A.; Guida, M.; Galdiero, M. Prevalence, Resistance Patterns and Biofilm Production Ability of Bacterial Uropathogens from Cases of Community-Acquired Urinary Tract Infections in South Italy. Pathogens 2023, 12, 537. [Google Scholar] [CrossRef] [PubMed]
- STEPANOVIĆ, S.; VUKOVIĆ, D.; HOLA, V.; BONAVENTURA, G.D.; DJUKIĆ, S.; ĆIRKOVIĆ, I.; RUZICKA, F. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Bai, X.; Zhang, Y.; Gao, Q.; Bu, X.; Xu, Y.; Guo, N. Evaluation of the Potential Probiotic Yeast Characteristics with Anti-MRSA Abilities. Probiotics and Antimicrobial Proteins 2022, 14, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Turchi, B.; Mancini, S.; Fratini, F.; Pedonese, F.; Nuvoloni, R.; Bertelloni, F.; Ebani, V.V.; Cerri, D. Preliminary evaluation of probiotic potential of Lactobacillus plantarum strains isolated from Italian food products. World J Microbiol Biotechnol 2013, 29, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Cisneros, L.; Cattelan, N.; Villalba, M.I.; Rodriguez, C.; Serra, D.O.; Yantorno, O.; Fadda, S. Lactic acid bacteria biofilms and their ability to mitigate Escherichia coli O157:H7 surface colonization. Lett Appl Microbiol 2021, 73, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Wächtler, B.; Wilson, D.; Hube, B. Candida albicans Adhesion to and Invasion and Damage of Vaginal Epithelial Cells: Stage-Specific Inhibition by Clotrimazole and Bifonazole. Antimicrobial Agents and Chemotherapy 2011, 55, 4436–4439. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Moyes, D.; Makwana, J.; Kanzaria, P.; Tsichlaki, E.; Weindl, G.; Tappuni, A.R.; Rodgers, C.A.; Woodman, A.J.; Challacombe, S.J.; et al. Quantitative expression of the Candida albicans secreted aspartyl proteinase gene family in human oral and vaginal candidiasis. Microbiology 2008, 154, 3266–3280. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Research 2001, 29, e45–e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Research 2002, 30, e36–e36. [Google Scholar] [CrossRef] [PubMed]
- Giammarino, A.; Bellucci, N.; Angiolella, L. Galleria mellonella as a Model for the Study of Fungal Pathogens: Advantages and Disadvantages. Pathogens 2024, 13, 233. [Google Scholar] [CrossRef] [PubMed]
- Vergara, S.C.; Leiva, M.J.; Mestre, M.V.; Vazquez, F.; Nally, M.C.; Maturano, Y.P. Non-saccharomyces yeast probiotics: revealing relevance and potential. FEMS Yeast Res 2023, 23. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. International Journal of Food Microbiology 2020, 333, 108818. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Ji, L.; Xu, Y.; Xu, S.; Lin, Y.; Iqbal, H.M.N.; Cheng, H. Bioprospecting Kluyveromyces marxianus as a Robust Host for Industrial Biotechnology. Front Bioeng Biotechnol 2022, 10, 851768. [Google Scholar] [CrossRef] [PubMed]
- Aziz Homayouni-Rad, A.A. , Parvin Oroojzadeh and Hadi Pourjafar. Kluyveromyces marxianus as a Probiotic Yeast: A Mini-review. Current Nutriotion & Food Science 2020, 16, 1163–1169. [Google Scholar] [CrossRef]
- Nag, D.; Goel, A.; Padwad, Y.; Singh, D. In Vitro Characterisation Revealed Himalayan Dairy Kluyveromyces marxianus PCH397 as Potential Probiotic with Therapeutic Properties. Probiotics and Antimicrobial Proteins 2023, 15, 761–773. [Google Scholar] [CrossRef]
- Collins, S.; Reid, G. Distant Site Effects of Ingested Prebiotics. Nutrients 2016, 8, 523. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Tittarelli, F.; Suzzi, G.; Tofalo, R. Cell Wall Surface Properties of Kluyveromyces marxianus Strains From Dairy-Products. Front Microbiol 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Motey, G.A.; Johansen, P.G.; Owusu-Kwarteng, J.; Ofori, L.A.; Obiri-Danso, K.; Siegumfeldt, H.; Larsen, N.; Jespersen, L. Probiotic potential of Saccharomyces cerevisiae and Kluyveromyces marxianus isolated from West African spontaneously fermented cereal and milk products. Yeast 2020, 37, 403–412. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Llopis, S.; Jespersen, L.; Fernández-Espinar, T.; Querol, A. Clinical Saccharomyces cerevisiae isolates cannot cross the epithelial barrier in vitro. Int J Food Microbiol 2012, 157, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Moure, M.C.; Pérez Torrado, R.; Garmendia, G.; Vero, S.; Querol, A.; Alconada, T.; León Peláez, Á. Characterization of kefir yeasts with antifungal capacity against Aspergillus species. International Microbiology 2023, 26, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Fallon, J.P. Galleria mellonella larvae as models for studying fungal virulence. Fungal Biology Reviews 2010, 24, 79–83. [Google Scholar] [CrossRef]
- Tran, K.-D.; Le-Thi, L.; Vo, H.-H.; Dinh-Thi, T.-V.; Nguyen-Thi, T.; Phan, N.-H.; Nguyen, K.-U. Probiotic Properties and Safety Evaluation in the Invertebrate Model Host Galleria mellonella of the Pichia kudriavzevii YGM091 Strain Isolated from Fermented Goat Milk. Probiotics and Antimicrobial Proteins 2024, 16, 1288–1303. [Google Scholar] [CrossRef]
- Desai, J.V. Candida albicans Hyphae: From Growth Initiation to Invasion. Journal of Fungi 2018, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.; Albrecht, A.; Bader, O.; Hube, B. Candida albicans proteinases and host/pathogen interactions. Cell Microbiol 2004, 6, 915–926. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Radar diagrams of the probiotic properties (see Table 1) of K. marxianus (A) and S. boulardii (B). The total score of the strain is reported in the centre of the diagram.
Figure 1.
Radar diagrams of the probiotic properties (see Table 1) of K. marxianus (A) and S. boulardii (B). The total score of the strain is reported in the centre of the diagram.
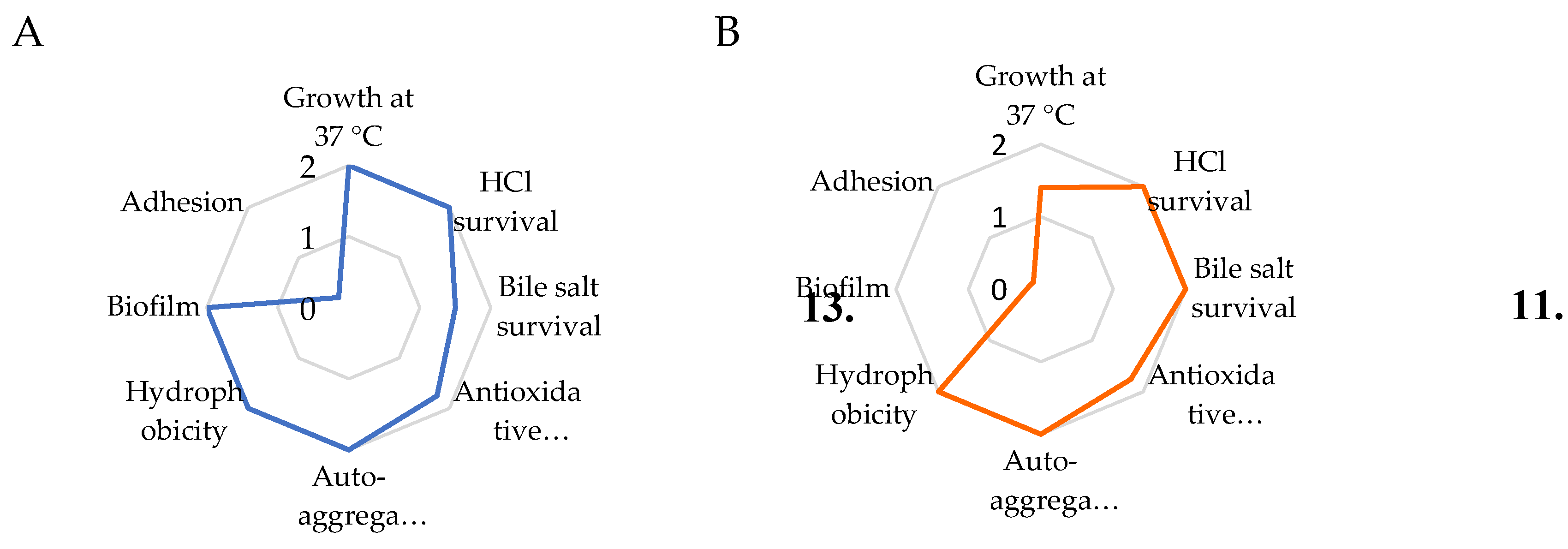
Figure 2.
Blood agar plates showing the absence of haemolytic activity for K. marxianus (A) and S. boulardii (B).
Figure 2.
Blood agar plates showing the absence of haemolytic activity for K. marxianus (A) and S. boulardii (B).
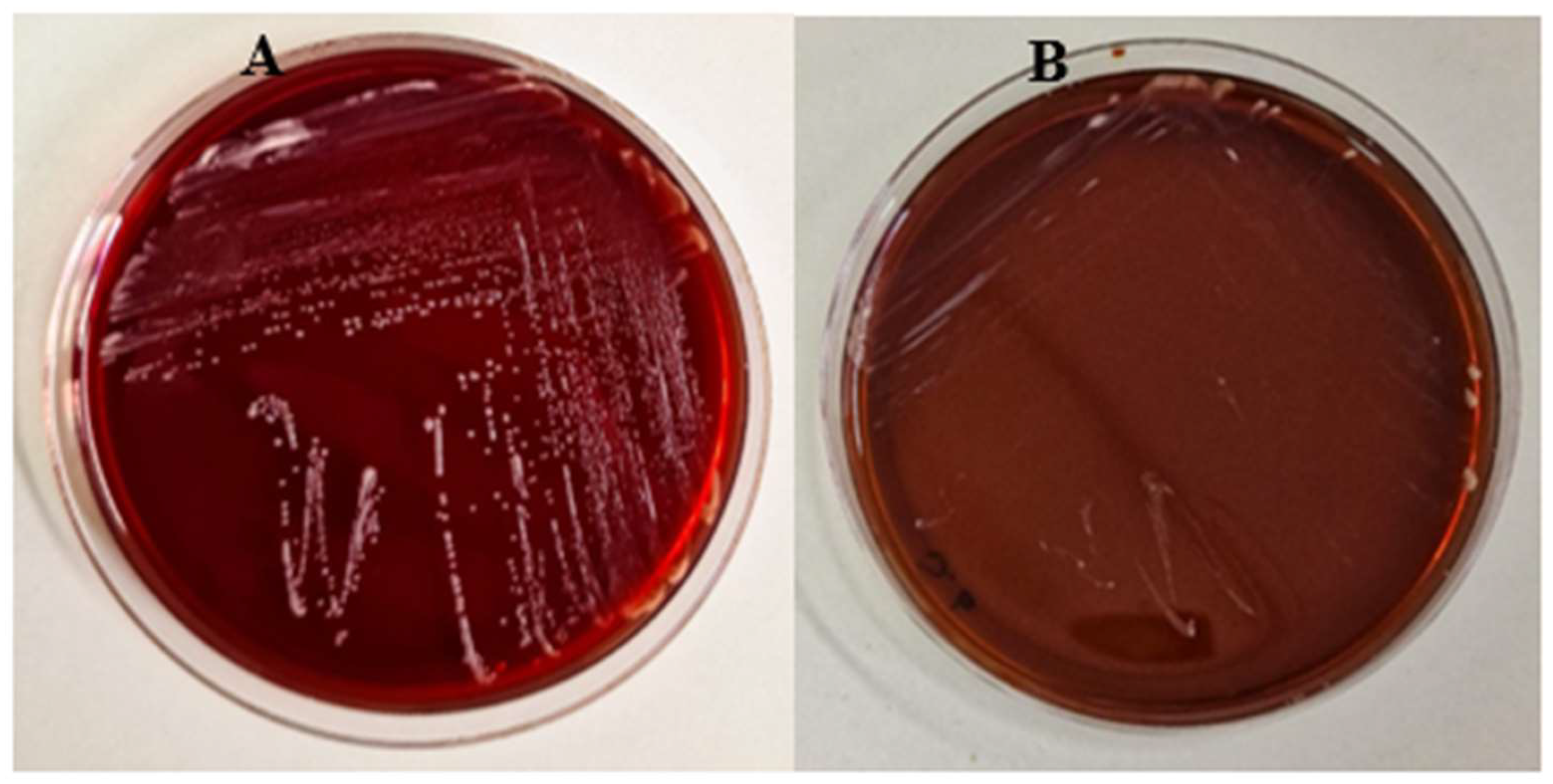
Figure 3.
Survival curves of G. mellonella larvae injected with K. marxianus (A) and S. boulardii (B) at density ranging from 104 to 10 7 CFU per larva.
Figure 3.
Survival curves of G. mellonella larvae injected with K. marxianus (A) and S. boulardii (B) at density ranging from 104 to 10 7 CFU per larva.
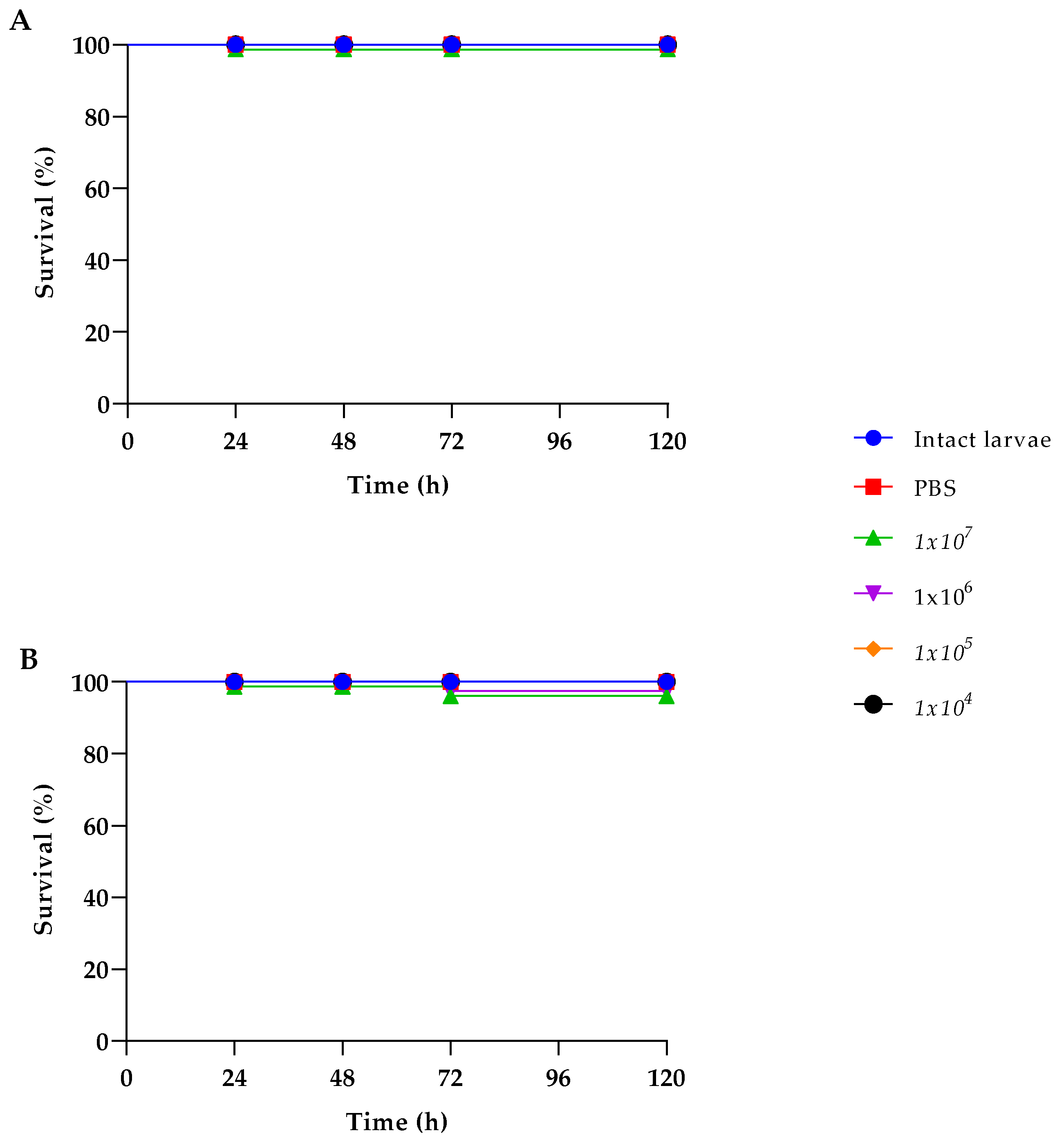
Figure 4.
Inhibition of filamentation of C. albicans ATTC 90028 (A, B) and C. albicans C19 (C, D) in the presence of K. marxianus (B, D). In panels A and C controls are shown. Bar corresponds to 125 μm.
Figure 4.
Inhibition of filamentation of C. albicans ATTC 90028 (A, B) and C. albicans C19 (C, D) in the presence of K. marxianus (B, D). In panels A and C controls are shown. Bar corresponds to 125 μm.
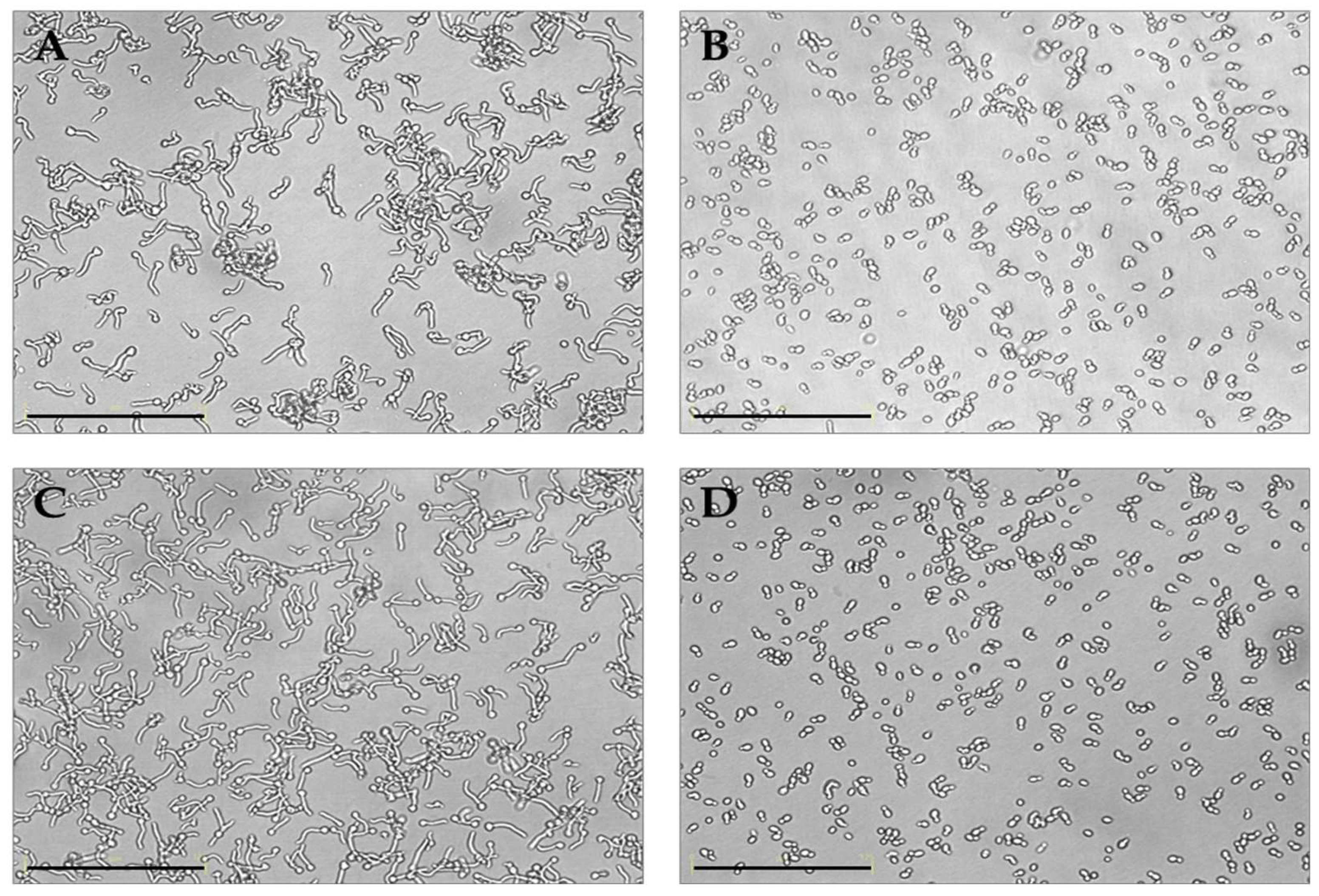
Figure 5.
Reduction of biofilm formation (Log10 CFU well-1) in polystyrene microwells of C. albicans ATCC 90028, C7, C14, C17 and C19 strains in the presence of a preformed K. marxianus biofilm. Data reported are the means of three independent experiments ± SDs; statistical significant is indicated by ** p < 0.01; *** p < 0.001 (Tukey’s test).
Figure 5.
Reduction of biofilm formation (Log10 CFU well-1) in polystyrene microwells of C. albicans ATCC 90028, C7, C14, C17 and C19 strains in the presence of a preformed K. marxianus biofilm. Data reported are the means of three independent experiments ± SDs; statistical significant is indicated by ** p < 0.01; *** p < 0.001 (Tukey’s test).
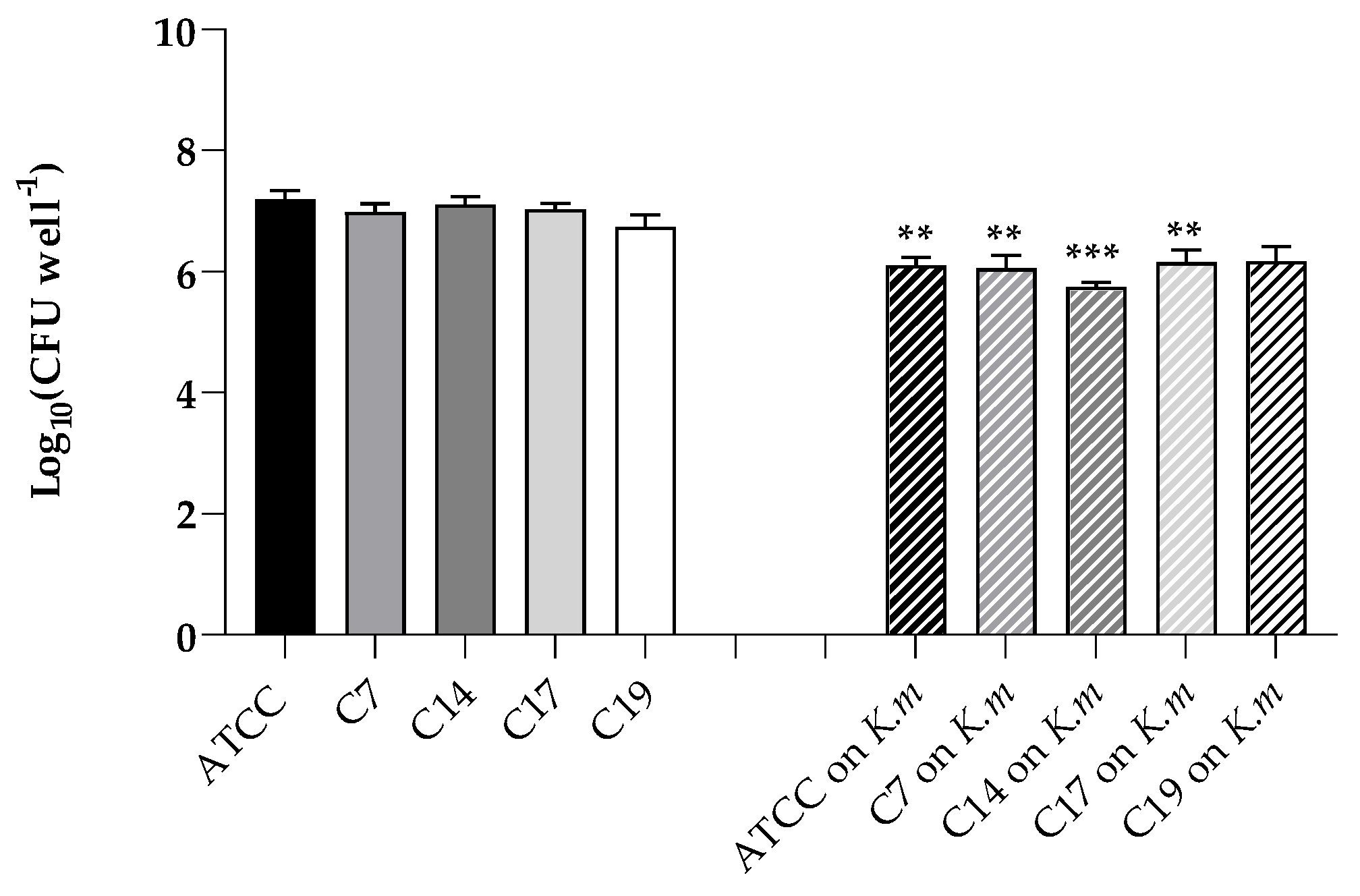
Figure 6.
Inhibition of adhesion of C. albicans ATCC 90028, C7, C14, C17 and C19 strains on A- 431 cells in conditions of pre-inoculation, co-inoculation, and post-inoculation with the K. marxianus isolate. Data reported are the means of three independent experiments ± SDs.
Figure 6.
Inhibition of adhesion of C. albicans ATCC 90028, C7, C14, C17 and C19 strains on A- 431 cells in conditions of pre-inoculation, co-inoculation, and post-inoculation with the K. marxianus isolate. Data reported are the means of three independent experiments ± SDs.
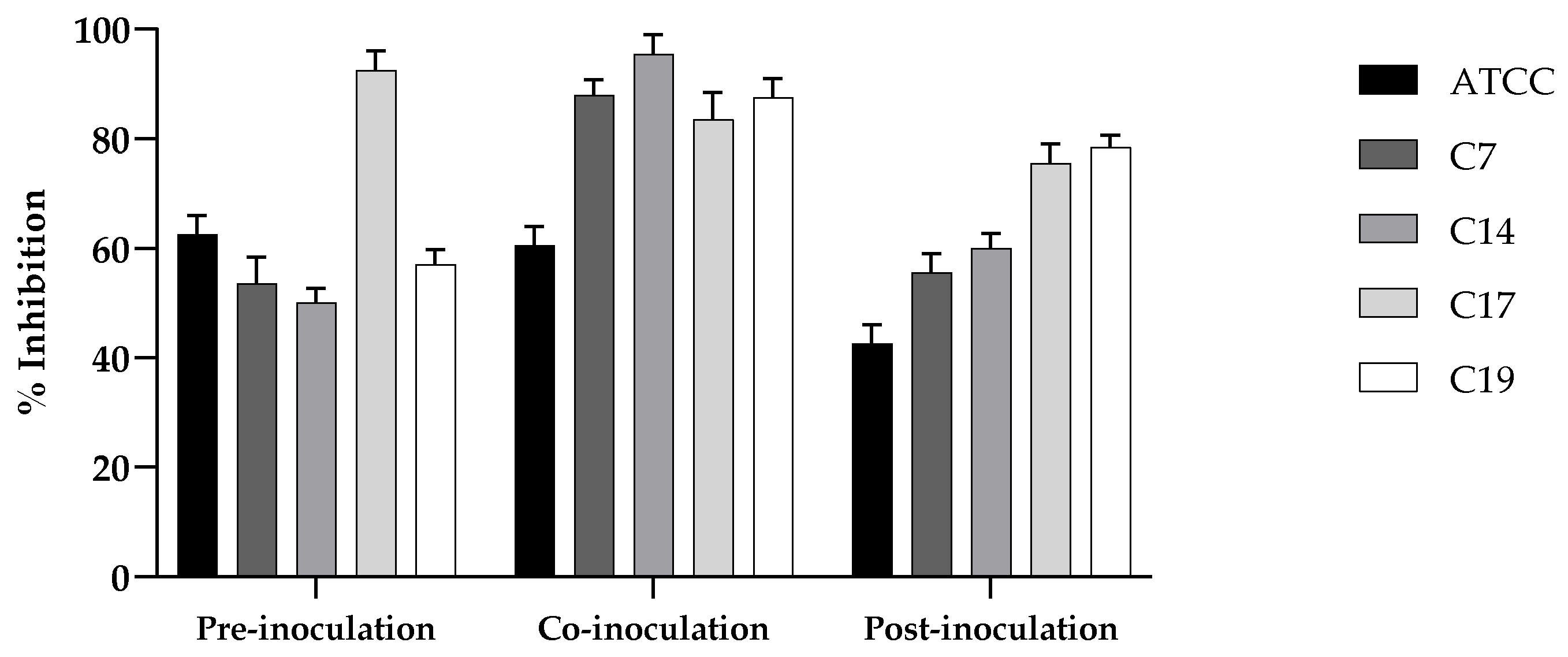
Figure 7.
Cell damage evaluated by LDH release from A- 431 cells infected with C. albicans ATCC 90028 and C19 strain and effect of the probiotic K. marxianus. Data reported are the means of three independent experiments ± SDs; statistical significant is indicated by ** p < 0.01 (Sidak’s test).
Figure 7.
Cell damage evaluated by LDH release from A- 431 cells infected with C. albicans ATCC 90028 and C19 strain and effect of the probiotic K. marxianus. Data reported are the means of three independent experiments ± SDs; statistical significant is indicated by ** p < 0.01 (Sidak’s test).
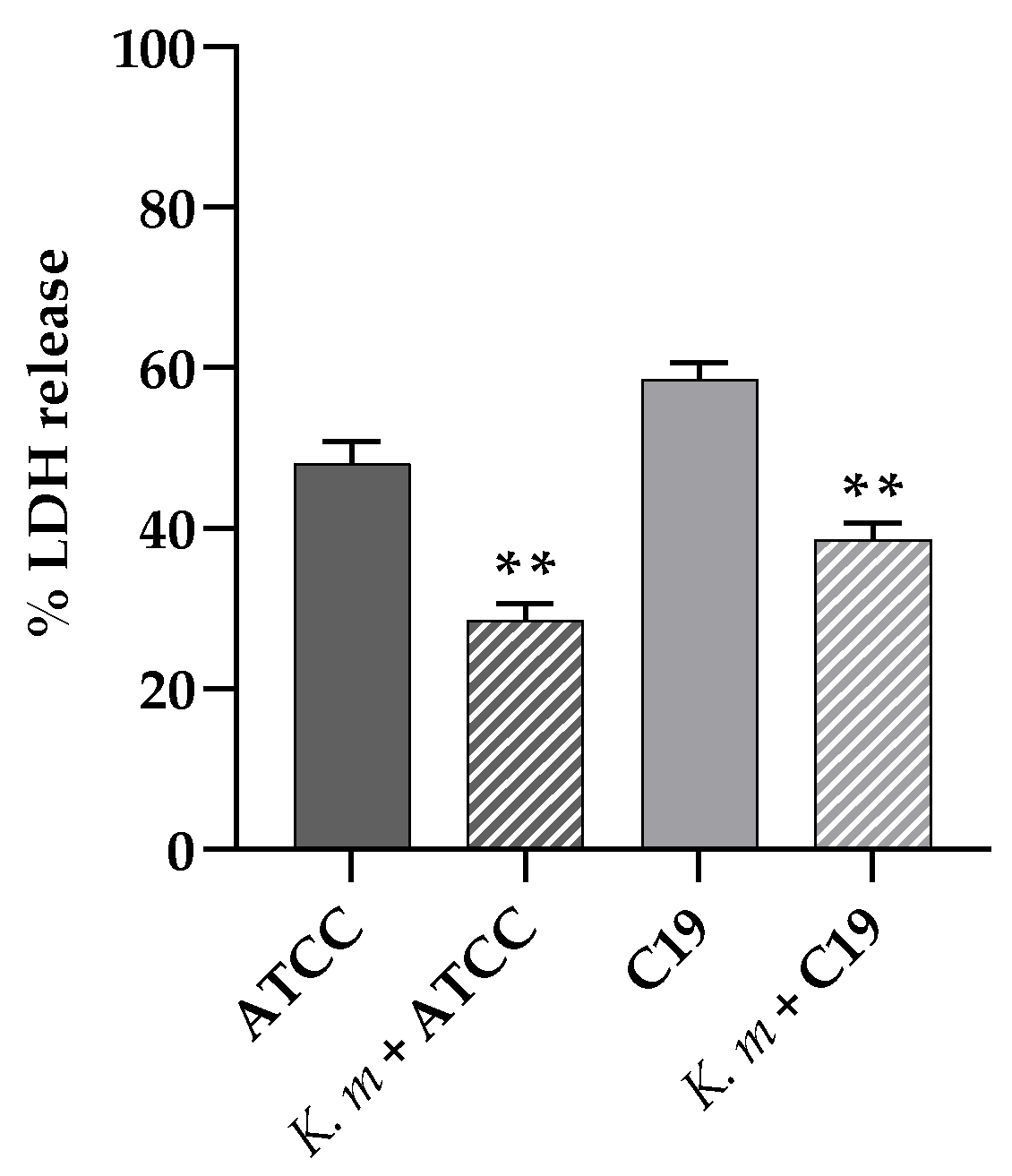
Figure 8.
Quantitative analysis of SAP2 and SAP6 genes expression in C. albicans ATTC 90028 (A) and C. albicans C19 (B) strains, incubated in YPD medium plus 1% BSA with or without K. marxianus. cDNA quantities were reported as fold changes relative to the Candida strain alone. Data reported are the means of three independent experiments ± SDs. Statistical significant is indicated by * p < 0.1 (Sidak’s test).
Figure 8.
Quantitative analysis of SAP2 and SAP6 genes expression in C. albicans ATTC 90028 (A) and C. albicans C19 (B) strains, incubated in YPD medium plus 1% BSA with or without K. marxianus. cDNA quantities were reported as fold changes relative to the Candida strain alone. Data reported are the means of three independent experiments ± SDs. Statistical significant is indicated by * p < 0.1 (Sidak’s test).
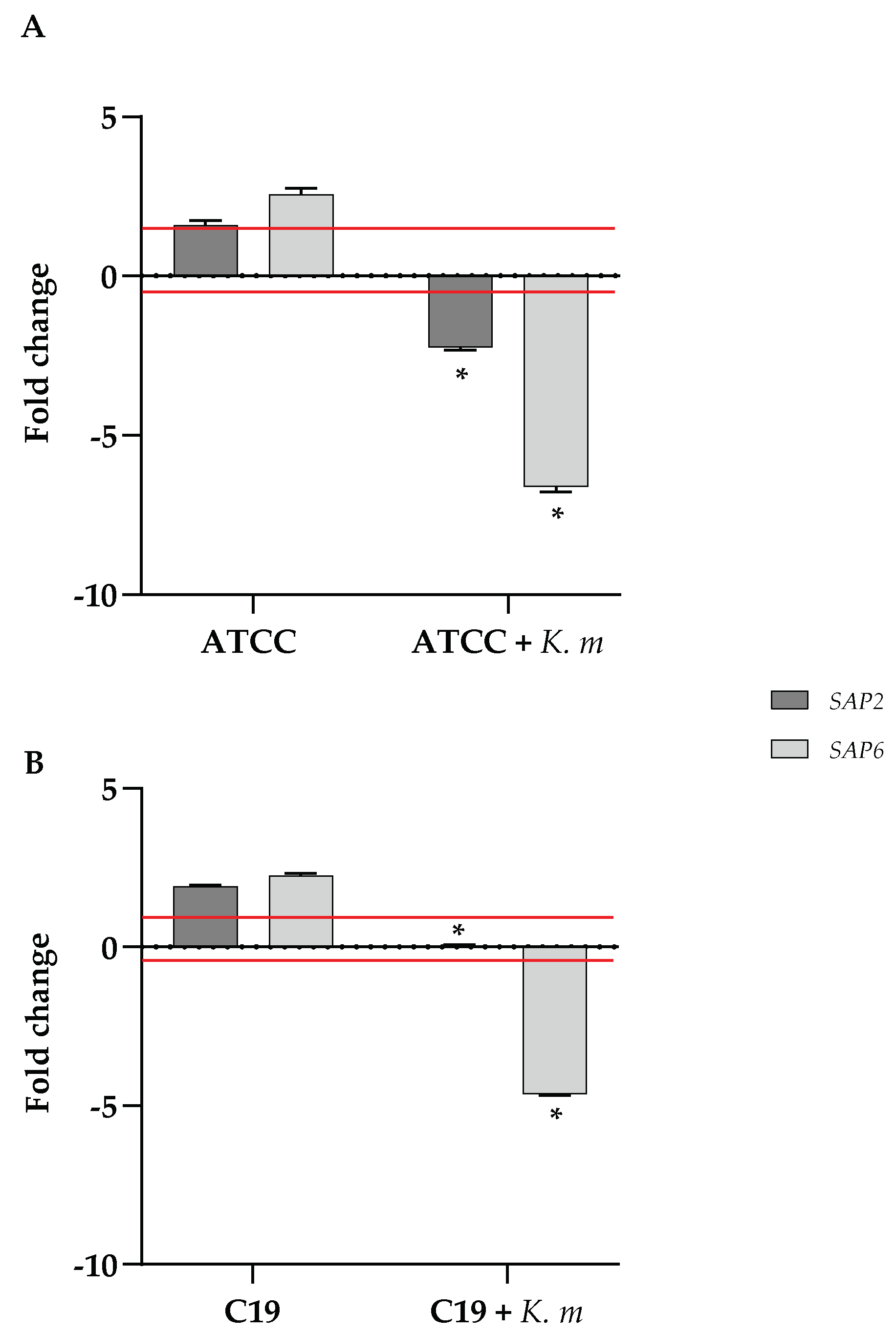

Table 1.
Results of Sanger Sequencing performed on yeast colonies isolated from sourdough. Obtained FASTA Sequences were compared to NCBI Sequence Database sequences, using BLASTN ver. 2.2.29.
Table 1.
Results of Sanger Sequencing performed on yeast colonies isolated from sourdough. Obtained FASTA Sequences were compared to NCBI Sequence Database sequences, using BLASTN ver. 2.2.29.
| Identified microrganism | Max score | Total score | Query Cover | e-value | % Identity | Accession N. |
| Kluyveromyces marxianus strain JYC2557 | 1223 | 1223 | 99% | 0.0 | 99.55% | MK044038.1 |
Table 2.
Results of the tests on the probiotic properties of K. marxianus (Km) and S. boulardii (Sb), and scores attributed to each test (in italics).
Table 2.
Results of the tests on the probiotic properties of K. marxianus (Km) and S. boulardii (Sb), and scores attributed to each test (in italics).
|
Growth at 37 °C |
HCl survival |
Bile salt survival | Antioxidative power |
Auto- aggregation |
Hydrophobicity | Biofilm | Adhesion | Total score | |
| (OD590) | (%) | (%) | (%) | (%) | (%) | (OD570) | (%) | ||
| Km | 6.86 | 100 | 73 | 78.5 | 100 | 90 | 4.5 | 10 | |
| 2 | 2 | 1.5 | 1.75 | 2 | 2 | 2 | 0.2 | 13.5 | |
| Sb | 4.93 | 100 | 98 | 77 | 100 | 81 | 0.3 | 2.35 | |
| 1.4 | 2 | 2 | 1.75 | 2 | 2 | 0.2 | 0.05 | 11.4 |
Table 3.
Table 3. Antimicrobial resistance of K. marxianus isolate (Km) and the commercial S. boulardii (Sb). The strains were considered susceptible (S), intermediate (I) or resistant (R).
Table 3.
Table 3. Antimicrobial resistance of K. marxianus isolate (Km) and the commercial S. boulardii (Sb). The strains were considered susceptible (S), intermediate (I) or resistant (R).
| GM | AMP | PB | AMC | NOR | STR | CTX | CIP | AZ | CRO | VAN | RIF | FLC | CSF | KET | ITR | AmphB | |
| Km | R | R | R | R | R | R | R | R | R | R | R | R | S | S | S | R | S |
| Sb | R | R | R | R | R | R | R | R | R | R | R | R | S | S | S | R | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



