
Submitted:
22 April 2025
Posted:
22 April 2025
You are already at the latest version
Abstract
Wheat Black Point, a seed-borne disease caused by Alternaria alternata, significantly impacts wheat production, reducing yield and quality. This study combined experimental and in silico approaches to evaluate the efficacy of fungicides against the esterase domain of A. alternata. Field trials assessed fungicide performance and optimal dosages, while laboratory analyses tested fungal susceptibility using the agar well diffusion method. Computational modeling, including AlphaFold 2 and molecular docking, investigated fungicide binding mechanisms. Leveraging machine learning and Explainable AI (XAI) techniques, this study integrates SHAP (SHapley Additive exPlanations) interaction values with XGBoost, a robust gradient boosting algorithm, to analyze the interaction of temperature, humidity, and precipitation with various pesticides in controlling Black Point Disease in wheat. By employing 1000 Monte Carlo simulations, the study provides a comprehensive evaluation of both individual and combined effects of these weather parameters on pesticide efficacy, offering a transparent and interpretable framework for optimizing pest control strategies. The findings revealed that Propiconazole (Tilt) demonstrated superior efficacy, with Dose 4 providing the highest disease suppression and yield improvement in field trials. Structural analysis of six fungal proteins revealed three important proteins, AFE1, AFE2, and AFE3. These proteins contain the Esterase_PHD domain, which is critical for the survival of fungi in host cells. Docking simulations identified robust interactions between Propiconazole and the C1 binding pocket of these proteins, yielding favorable Vina scores (-5.1 for AFAE1 and AFAE2; -4.7 for AFAE3). Results of SHAP indicated that the moisture-dependent pesticides (Propiconazole and Cymoxanil + Mancozeb), worked better under wet and humid climates, whereas region-specific pesticides (Metribuzine and Thiophanate-methyl), were effective in semi-arid and moderate climates. Adaptable pesticides, like Chlorothalonil + Metalaxyl-M and Sulfosulfuron, demonstrated consistent efficacy across variable climatic conditions. The integration of machine learning and AI tools with traditional agricultural practices is recommended to optimize fungicide application strategies. These findings highlight the critical role of tailoring pesticide application to regional environmental parameters to improve disease management strategies.
Keywords:
A. alternata
; Black Point
; Esterase_PHD
; Propiconazole (Tilt)
; Triticum
; Explainable AI (XAI)
; SHAP (SHapley Additive exPlanations)
; XGBoost
Introduction
Wheat (Triticum spp.) is an important crop that provides key nutrients and energy in the human diet (Shukla et al., 2020). Pakistan is the second most rapidly growing population at the rate of 3.6% in South Asia and the fourth largest population in general. The most significant and second largest food crop is wheat after rice. Both the biotic and abiotic factors are involved in causing black point disease which causes an estimated 22% yield losses (Figueroa et al., 2018). Annually about 20% of global crop yield is reduced by pathogenic fungi. The most common fungal diseases in wheat include Spot Blotch (Cochliobolus sativus), Black Point (A. alternata), and leaf Rust (Puccinia triticina) (Savary et al., 2019). In 2021 and 2022, the huge impact of fungal diseases was reported, 3.9% reduction in wheat production and 2.1% in wheat acreage resulting in approximately 15 to 20% yield losses in all parts of the country (Saeed & Hammad, 2022-23).
Black point is a fungal seed-born disease that has economic importance and negatively impacts the wheat crop globally. It is differentiated by dark brown or black discoloration at the embryo's end and surrounding areas of the wheat kernel. This disease generally reduces grain quality, germination, and crop production (Shukla et al., 2020). The primary pathogen of this disease is A. alternata which accounts for up to 24% loss of wheat crop (El-Gremi et al., 2017). The A. alternata fungus affects plants and humans and causes diseases in them, also its pathogenicity affects agricultural products. This fungus belongs to Ascomycota which is known to cause diseases in about 400 plant species and damage the postharvest outputs (Masiello et al., 2020). It is necessary to effectively manage diseases from the danger caused by fungal pathogens (Fisher et al., 2012; Strange & Scott, 2005).
To prevent fungal growth and the development of disease, fungicide application is important (Achilonu et al., 2023). Propiconazole is a systematic fungicide that shows a response against black point, powdery mildew, and leaf spot (Kassaw et al., 2021). A. alternata contains an esterase enzyme which is involved in the pathogenicity of this fungus so the esterase domain interaction with fungicide is important to understand (Hou et al., 2022). Furthermore, feruloyl esterases contribute to the pathogenicity of A. alternata by breaking down the cell wall of plants specifically the lignocellulose-containing part (Crepin et al., 2004). However, there is little information is available on how the A. alternata’s esterases are affected by the fungicides.
Crop diseases pose a danger to food security but due to the unavailability of infrastructure, it is difficult to detect and identify them in their initial stages which is significant for the control of disease (Long et al., 2023; Mohanty et al., 2016). AI-based computer vision methods must be investigated to quickly and economically identify disease incidents on each plot of land under cropping (Kamilaris & Prenafeta-Boldú, 2018b; Abade et al., 2021; Arnal Barbedo, 2013; Haque et al., 2022). The use of AI-based techniques reduces the accidental use of chemicals, carries out regular health checks of plants, and gives information about the minimum number or amount of pesticides required to cure plant diseases (Ferentinos, 2018). Recent studies reported that deep machine models give 98% accurate results of disease identification on the dataset (Naik et al., 2022; Sutaji & Yıldız, 2022).
The study examined the efficacy of commercially available fungicides against A. alternata by integrating experimental trials and in silico studies. The experimental analysis includes field trials and agar well diffusion technique while in silico analysis includes protein structure modeling with AlphaFold3 and docking simulations to investigate the interactions between fungicides and the fungal esterase-PHD domain. Machine learning models were also used to evaluate how well the fungicides would work in different environmental conditions and the impact of climate factors such as precipitation, temperature, and humidity on fungicides' performance. This research attempts to improve fungicide application strategies and adapt them to challenges posed by climate change to ensure sustainable wheat production and efficient management of disease. It is important to know how the climate affects the fungicides' efficacy since black point seriously lowers crop yields. This study seeks to the individual and combined effects of climatic variables on pesticide efficacy to understand the interconnections and the selection of appropriate pesticides for certain
Material and Methods
This study's methodology consists of four primary components: fieldwork, laboratory work, computational analysis, and machine learning employing XGBoost (Extreme Gradient Boosting) in conjunction with Explainable AI (XAI) techniques, particularly utilizing SHAP (SHapley Additive exPlanations) to deliver interpretable and transparent insights regarding feature interactions and their influence on pesticide efficacy (Prendin et al., 2023).
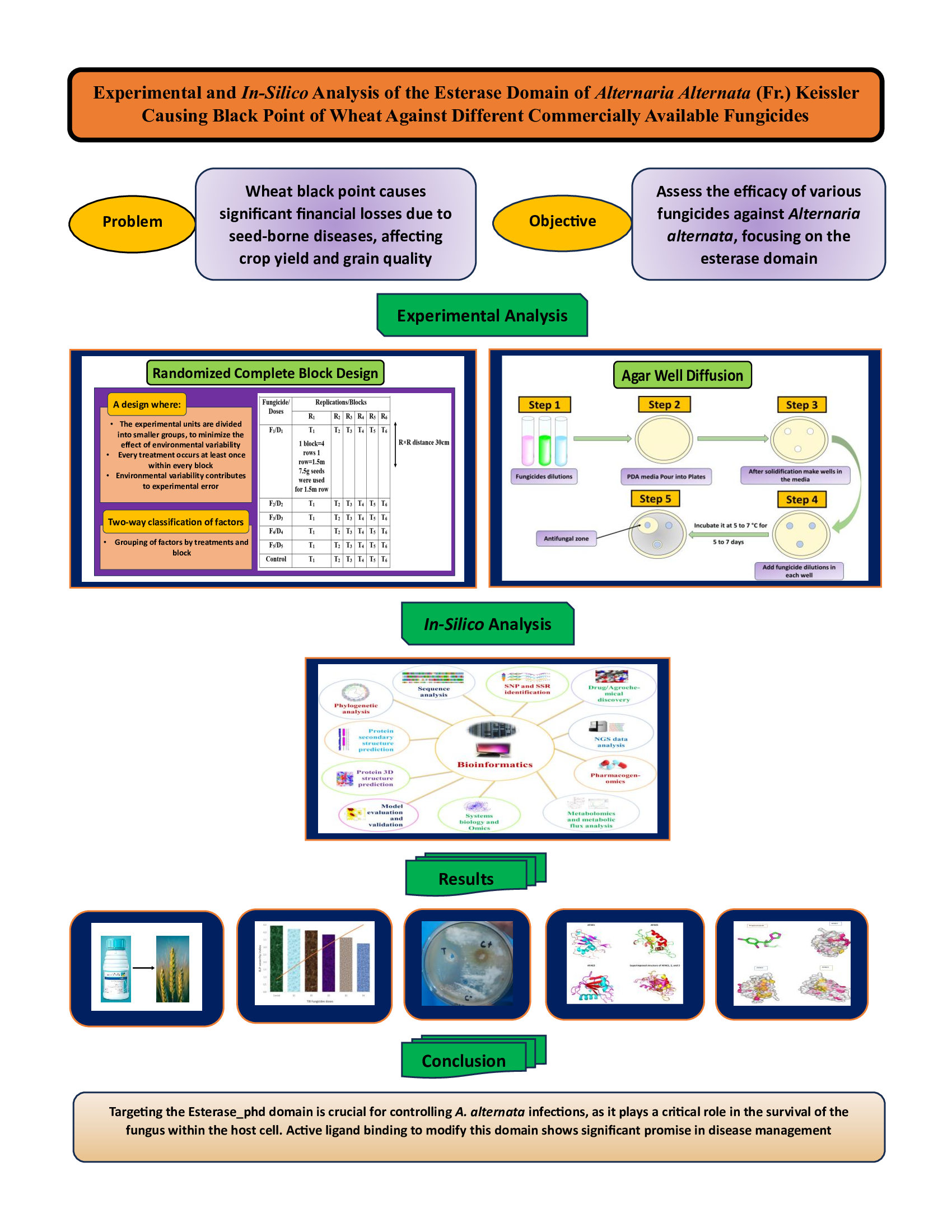
Figure 1.
Depicts the systematic steps of this study such as field work, laboratory work, computational work, and deep machine learning.
Figure 1.
Depicts the systematic steps of this study such as field work, laboratory work, computational work, and deep machine learning.
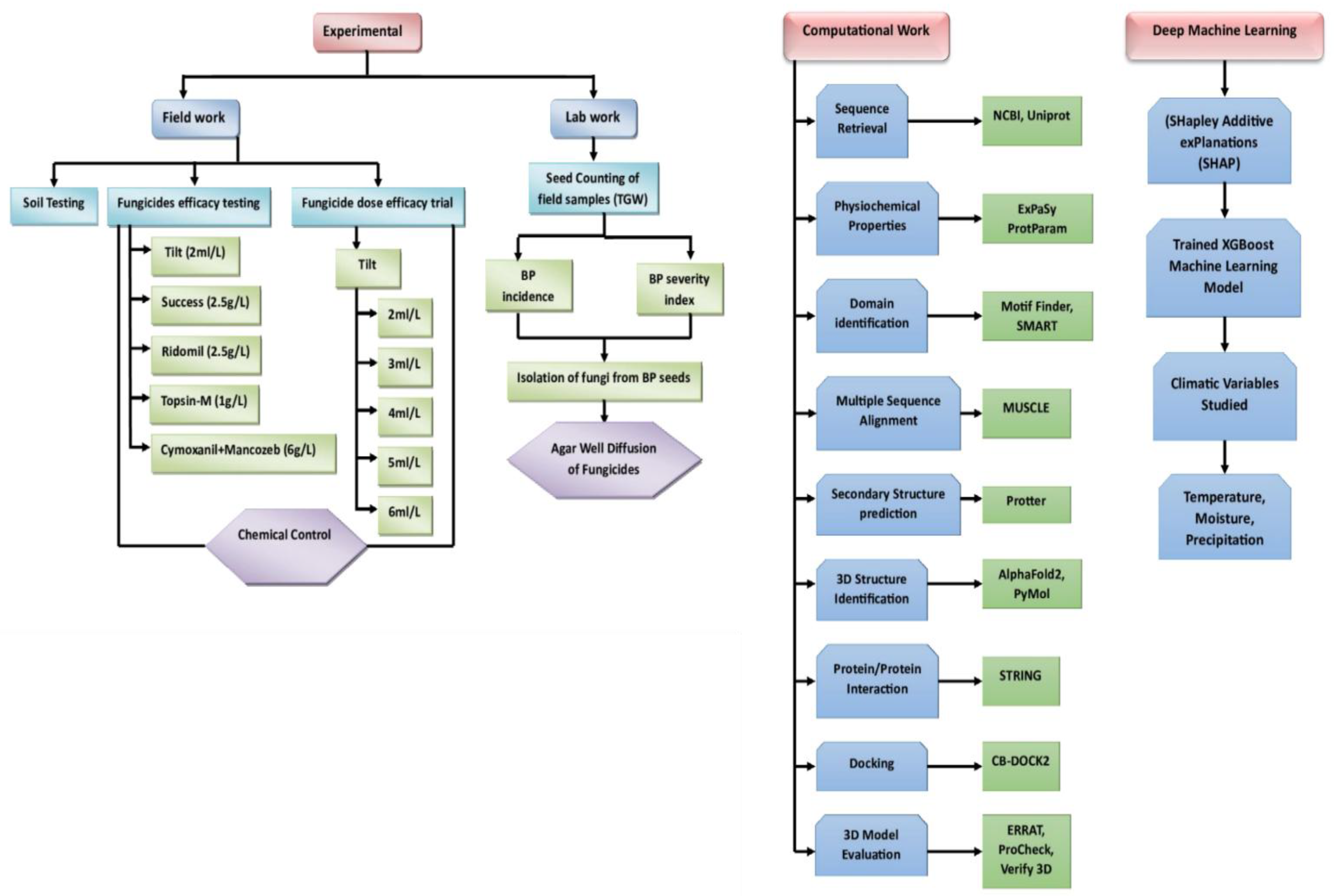
Study Design
The study was designed by the Department of Microbiology, Shaheed Benazir Bhutto Women University Peshawar, in collaboration with the Plant Pathology Group at the Nuclear Institute for Food and Agriculture (NIFA) in Peshawar. The fieldwork was conducted at the NIFA farm, while the in-vitro experiments were carried out in the NIFA Plant Pathology Laboratory. The study was carried out over a year, from November 2022 to December 2023. Figure S1 shows the location of the field and experimental work.
Ethical Approval
This study was approved by the ethical committee of Shaheed Benazir Bhutto Women University Peshawar.
Field Work
During the 2022-23 cropping season, two trials were conducted at the NIFA farm to assess the efficacy of five fungicides and different doses of one fungicide against black point on the susceptible variety "Morocco." The trials were raised in a Randomized Complete Block (RCB) design with six replications. In the first trial, five fungicides (Tilt, Success, Ridomil, Topsin-M, Cymoxanil+Mancozeb) were evaluated, while in the second trial, different doses of the fungicide "Tilt" (2ml/l, 3ml/l, 4ml/l, 5ml/l, 6ml/l) were tested as shown in Table S1 to S5 (Asif et al., 2021). Morocco was sown on 0.125 acres, with appropriate fertilizers applied (Degewione & Alamerew, 2013). Seeds were sown manually at specific densities, and fungicide applications were carried out as per company recommendations in the first trial. Harvesting took place in mid-May 2023, followed by threshing and collection of seeds for further analysis, specifically for the black point study (Nithya et al., 2011) (Figure S3). The physicochemical properties of the soil were also tested by the Soil and Environmental Sciences Division of NIFA, Peshawar. The soil was collected from a depth of 0-15 cm in the wheat farm (Peila et al., 2016).
Laboratory Work
An automatic seed counter and a Sartorius balance were utilized to generate treatment-wise/replication-wise working samples of 1,000 seeds and recorded their weight in grams (Moshatati & Gharineh, 2012) (Figure S4). Black point infection was identified by manually counting black-pointed seeds, observed through careful examination using a stereo microscope at 20X magnification. The percent disease incidence was calculated using a specific formula (Shukla et al., 2020). (Figure S5)
The black point severity index was also calculated as described by (Draz et al., 2016) using the severity score from 0 to 3 (Figure S6)
The black point severity index was calculated by using the mentioned formula:
Pathogens associated with black-point-infected seeds were isolated using the agar plating technique on Potato Dextrose Agar (PDA) as shown in Figure S8. Surface-sterilized seeds were placed on PDA plates under a laminar flow unit and incubated for 7 days (Xu et al., 2018). Pure cultures of the isolated fungi were maintained on PDA and identified based on morphological features observed under a compound microscope (Manamgoda et al., 2015). Fresh cultures of the identified fungi were sub-cultured for further analysis. The efficacy of chemical fungicides, including Tilt, Success, Ridomil, Topsin M, and Cymoxanil+Mancozeb, against black point pathogens, was assessed using the agar well diffusion technique. Inhibition zone diameters were measured after 5 days of incubation at 25-27°C (Balouiri et al., 2016). (Figure S9)
The soil fungal flora of both trials was also assessed using the process of serial dilution (Alastruey et al., 2015). Fungal colonies were identified based on their morphological and diagnostic features using a compound microscope (Manamgoda et al., 2015). The soil was collected from three depths, such as 0-15 cm, 15-30 cm, and 30-45 cm, respectively as shown in Figure S10.
In-Silico Analysis
Six A. alternata proteins were retrieved from UniProt and NCBI databases for analysis, focusing on those without solved experimental protein structures (Griffin & Griffin, 1995; “UniProt: A Hub for Protein Information,” 2015). Physicochemical properties were assessed using the ProtParam tool, including molecular weight, theoretical pI, amino acid composition, and other parameters (Gasteiger et al., 2005). Protein domain analysis was conducted using SMART (Schultz et al., 1998), and Motif finder (Brown et al., 2013). These tools were used to identify domains, motifs, and families. Sequence alignment was done using MUSCLE (Edgar & Batzoglou, 2006). Secondary structures were predicted using Protter-interactive protein feature visualization https://wlab.ethz.ch/protter/start/. Tertiary protein structures were predicted using AlphaFold 3 (Abramson et al., 2024) software and visualized with PyMol (DeLano & Bromberg, 2004). Stereochemical quality was assessed using ProCheck (Laskowski et al., 1993), ERRAT (Dym & Eisenberg, 2012), ProSA-Web (Wiederstein & Sippl, 2007), and verify3D (Eisenberg et al., 1997). Active sites were predicted using the Computed Atlas of Surface Topography of Proteins (CASTp) (Binkowski, 2003). To examine protein to protein interactions, STRING software was utilized (Szklarczyk et al., 2015). For finding reputed ligands to bind with active sites, ChEMBL (https://www.ebi.ac.uk/chembl/) database was used (Gaulton et al., 2012). CB-Dock2 (https://cadd.labshare.cn/cb-dock2/php/index.php) was used for blind docking purposes which is a web-based application (Liu et al., 2022).
Molecular Docking
CB-Dock2, a cavity-detection-based docking server, was used to blind dock the three fungal esterase homologs (AFAE1, AFAE2, and AFAE3) in order to find possible ligand-binding pockets. CB-Dock2 predicted several binding cavities for every ligand-protein set, and AutoDock Vina [1] was used to score the cavities according to their volume and binding energy. The pocket with the highest Vina score, or the top-ranked pocket, was chosen for further targeted docking.
Ligand structures were created using Open Babel's [2] 3D geometry generating feature from .mol files in PDBQT format. To ensure compatibility with rigid docking processes, protein receptors were processed using MGLTools' [3] prepare_receptor4.py and then cleaned to eliminate unnecessary torsional annotations.
AutoDock Vina (v1.2) was used for the final docking, with grid box parameters based on the chosen pocket coordinates that CB-Dock2 had supplied. A custom Python pipeline automated ligand/receptor preparation, execution of Vina docking, and parsing of results, including binding affinity values and predicted interacting residues. A structured summary table containing all of the docking data was created in order to choose ligand-protein complexes for molecular dynamics simulation.
Molecular Dynamics Simulations
GROMACS (v2021.6) [4]was used to conduct molecular dynamics (MD) simulations in order to assess the dynamic stability of the docked ligand–protein complexes. Top-ranked docking poses (based on Vina scores) were extracted, and ligand topologies were generated using ACPYPE [5] with AMBER force field parameters. Ligand coordinate and topology files were patched and standardized across all systems for consistency.
The AMBER99SB-ILDN force field and SPC/E water model were used to process protein receptors (AFAE1, AFAE2, and AFAE3) using pdb2gmx. After being solvated with explicit water and neutralised by the addition of Na+ and Cl+ ions, each ligand–protein complex was centred in a cubic box with a spacing of 0.8 nm.
Using the steepest descent algorithm, energy was minimised. Following this, 100 ps of NVT and 100 ps of NPT equilibration were performed employing position restrictions on the ligand's heavy atoms and protein. Using a Parrinello-Rahman barostat and a velocity-rescaling thermostat, final production runs were conducted for 5 ns under periodic boundary conditions.
To process all systems in a sequential manner and guarantee uniform ligand preparation, topology merging, restraint creation, and simulation execution, a completely automated Python pipeline was created. GPU acceleration was used for all simulations, and the results were post-processed to extract solvent-accessible surface area (SASA), radius of gyration (Rg), RMSD, RMSF, and hydrogen bonds.
Machine Learning and Explainable AI Analysis of Climatic Impacts on Fungicide Efficacy
Dataset Collection and Overview
In this study, the dataset used was taken from the Department of Agriculture, Peshawar which gives a detailed compilation of agricultural and environmental data collected from six key districts of wheat production in Khyber Pakhtunkhwa, Pakistan including Peshawar, Nowshera, Mardan, Dera Ismail Khan, Charsadda and Swabi. These districts constitute diverse agro-climatic zones, to ensure a thorough understanding of the agronomic and environmental factors affecting the dynamics of black point disease and wheat yield. The data collected compass 2020–2024, employing systematic field surveys and monitoring systems to capture inter-annual climate variability and cropping patterns. Rigorous validation processes were employed to ensure data accuracy, incorporating checks for consistency and completeness. The dataset consists of 2,000 rows, offering a rich and representative sample of real-world agricultural conditions.
The key climatic parameters were included in the dataset such as Humidity (%), measuring daily average relative humidity which is critical for the growth of fungus; Total Precipitation (mm), reflecting cumulative rainfall contributing to soil and atmospheric moisture; and Average Temperature (°C), capturing daily mean temperature that impacts the crop physiology and activity of pathogens. These variables provide a multidimensional view of weather conditions driving black point disease incidence. The target variable, black point disease incidence, quantifies the percentage of crops affected, serving as the outcome for prediction and analysis.
The dataset also includes detailed pesticide application records, encompassing fungicides such as Propiconazole, Metalaxyl-M + Mancozeb, Propoxycarbazone, Sulfosulfuron, Thiophanate-methyl, Metribuzine, Cymoxanil + Mancozeb, Chlorothalonil + Metalaxyl-M, and Dichlorodiphenyltrichloroethane (DDT). These records facilitated a nuanced evaluation of pesticide efficacy under varying climatic conditions.
Preprocessing
A comprehensive preprocessing pipeline was applied to the dataset to ensure data quality, enhance feature interpretability, and optimize model performance. This pipeline addressed missing values, duplicate entries, categorical encoding, feature extraction, numerical scaling, and outlier detection.
Missing Value Imputation: Addressed missing entries in climatic variables (e.g., temperature, humidity, precipitation) and pesticide efficacy data to ensure completeness.
Outlier Detection and Removal: Identified and removed anomalous values in key numeric columns, such as temperature and Black Point disease incidence, to improve the quality of the data.
Normalization: Applied Z-score scaling to normalize climatic parameters (temperature, humidity, and precipitation) for better model performance and feature comparability.
Encoding Categorical Variables: Encoded pesticide names into numeric representations (e.g., using label encoding) to prepare the data for machine learning models while maintaining interpretability.
Data Integrity Checks: Verified that the dataset was free of duplicate rows and logical inconsistencies, ensuring a clean and reliable data foundation.
SHAP Analysis to Quantify and Interpret Climate and Pesticide Interactions The Explainable AI techniques, specifically SHAP (SHapley Additive exPlanations), were utilized to quantify interaction effects and feature importance. An XGBoost model was used to facilitate SHAP analysis for its accuracy, efficiency, and ability to handle datasets with mixed numerical and categorical features, while effectively capturing interactions between predictors. The predictors included climate variables (humidity, precipitation, and temperature) and pesticide types. Black point disease incidence, expressed as the percentage of affected crops was the target variable. The preprocessed dataset, comprising normalized climatic variables and encoded pesticide types, was leveraged to train the XGBoost model, allowing it to learn complex relationships between climatic conditions, pesticide applications, and disease incidence. To ensure reliability and minimize variability, SHAP interaction values were averaged over 1,000 Monte Carlo simulations, providing robust insights into the individual and combined effects of climatic variables on fungicide performance. This comprehensive methodology enabled the identification of temperature-, precipitation-, and humidity-dependent pesticide behaviors, offering actionable data to optimize fungicide application under diverse environmental conditions (Lee et al., 2021; Prendin et al., 2023; Taylor & Cunniffe, 2023).
Data Analysis
Social Sciences software (IBM SPSS Statistics 20) was used to analyze the ANOVA, LSD, and correlation on the experimental dataset.
ResultsField Results
The soil’s physicochemical properties of the wheat field were tested which include soil texture (Slit loam), pH (7.9), nitrogen (0.06%), electrical conductivity (0.50 ds/m), phosphorous (9.25ppm), potash (110 ppm) and organic matter (1%) as shown in Table S6 respectively. The field samples were evaluated in the laboratory. For the fungicides trial, tilt (Propiconazole) showed the highest efficacy, with an incidence of 5.7%, followed by Cymoxanil+Mancozeb (6.0%), Success (6.3%), Topsin-M (6.5%), and Ridomil (7.0%). The control group had the highest disease incidence i.e., 8.6%. Propiconazole also resulted in the highest TKW i.e., 35.5g, followed by Cymoxanil+Mancozeb (34g), Topsin-M (30.9g), Success (30.6g), and Ridomil (29.7g), while the control had the lowest TKW i.e., 25.9g as shown in Table S7 and Figure S11. In case of severity index, Cymoxanil+Mencozeb had the lowest severity index (2.7%), followed by Tilt (3%), Success (3.1%), and Ridomil (3.3%). The highest severity index was observed for Topsin-M (4.7%). The control group exhibited a severity index of 4.1%. as shown in Table S8 and Figure S12. The ANOVA results indicated that there were no significant differences among treatments for Severity (F = 1.28, p = 0.3046) and Incidence (F = 1.05, p = 0.4109), while TKW showed significant differences (F = 14.92, p < 0.0001). The LSD pairwise comparisons revealed significant differences among treatments for Severity and TKW, while no significant pairwise differences were observed for Incidence. Notably, TKW means varied significantly among treatments, highlighting distinct treatment effects. Correlation analysis showed a strong positive correlation between Severity and Incidence (r = 0.7107), moderate negative correlations between Incidence and TKW (r = -0.2901), and a moderate positive correlation between TKW and Treatment (r = 0.3475). These findings suggest significant treatment effects on TKW but not on Severity and Incidence, with notable correlations among the response variables.
For the dose trail, the most effective dose was dose 4 (5.7%), followed by dose 1 (6%), dose 2 (6.3%), dose 5 (6.6%), and dose 3 (7.4%) respectively. The control group had the highest disease incidence i.e., 9.1%. The maximum TKW was recorded for dose 5 (37.0g), followed by dose 4 (36.7g), dose 3 (36.6g), dose 2 (36.3g), and dose 1 (35.8g). The control had the lowest TKW of 30.3g as shown in Table S9 and Figure S13. The severity index for kernel black point disease in the tilt fungicide trial varied across different doses of the fungicide. Dose 4 had the lowest severity index of 3.3%, followed by dose 2 (3.6%), dose 3 (3.8%), dose 5 (4.1%), and dose 1 (4.2%). The control group had the highest severity index of 4.4% as shown in Table S10 and Figure S14. The ANOVA results indicated no significant differences among treatments for Sev (F=1.75, p=0.1599), while significant differences were found for both Inc (F=4.30, p=0.0059) and TKW (F=20.12, p<0.0001). The LSD All-Pairwise Comparisons Test reveals significant differences among treatments for severity, incidence, and TKW, with specific treatments showing distinct separation from others within each variable. Correlation analysis showed a moderate positive correlation between Sev and Inc (r=0.5170) and Treatments and TKW (r=0.6424). Negative correlations were observed between Inc and TKW (r=-0.5131) and Sev and TKW (r=-0.2482), indicating potential trade-offs among these traits. These findings provide insights into the treatment effects and relationships between key agricultural variables.
Laboratory Results
Fungus detected and isolated from black-pointed seeds was identified as Alternaria alternata (Figure S15), exhibiting characteristic morphological features (Figure S16) (Woudenberg et al., 2013). The efficacy of fungicides against A. alternata was assessed using the agar well diffusion technique. Tilt (Propiconazole) displayed the highest efficacy with a 28 mm inhibition zone, followed by Cymoxanil+Mancozeb with an 8 mm inhibition zone. Success, Ridomil, and Topsin M showed no significant activity against A. alternata under laboratory conditions. These results indicate varying effectiveness among the tested fungicides, with Tilt and Cymoxanil+Mancozeb demonstrating a positive lab-based inhibition of A. alternata growth as shown in Figures S17 and S18. From the serial dilution process, Aspergillus niger, Aspergillus fumigatus, and A. alternata were consistently isolated across all dilutions as shown in Figure S19. These three fungi have been identified as the causative agents of Black Point disease in wheat.
In-Silico Analysis Results
Sequence Retrieval
To identify proteins involved in the virulence of A. alternaria, a total of six proteins were searched and selected based on data from the Universal Protein Resource (UniProt) and the National Center for Biotechnology Information (NCBI) databases. To determine whether the structures of these proteins had been previously elucidated, a search was conducted in the Protein Data Bank (PDB). Only those proteins whose structures had not been experimentally determined were selected for further analysis, relying solely on the AlphaFold-predicted structures or not even predicted by AlphaFold. After validation, the FASTA sequences for each protein were obtained using UniProt. Among the six proteins involved in the virulence of A. alternata, AFAE1 (Accession no: A0A4Q4NLE1), AFAE2 (Accession no: A0A177DJG1), and AFAE3 (Accession no: A0A177DMH0) were selected for further investigation due to their functional relevance, as they feature the esterase-PHD domain. By hydrolyzing ester linkages between hemicelluloses and lignin, feruloyl esterases in particular are linked to the breakdown of plant cell walls, which is a crucial step in impairing plant defenses and promoting infection. These proteins are important targets for structural and functional research since it is thought that they contribute significantly to A. alternata's pathogenicity. (Table S11).
These three proteins were chosen based on a number of factors, such as their possible functional significance in A. alternata, their high correlation with virulence, and the presence of the esterase-PHD domain. Existing research and bioinformatic predictions that emphasized these particular proteins as important contributors to pathogenicity served as the impetus for the focus on them. A more thorough structural and functional examination is possible when the study is restricted to three proteins, guaranteeing correctness and viability. Other proteins with esterase domains may be the subject of future research, but this preliminary analysis lays the groundwork for examining the most promising possibilities first. In addition to offering important insights into the function of esterases in fungal virulence, this phased strategy guarantees that the study stays focused and manageable.
AFAE1 likely stands for Alternaria feruloyl esterase 1. AFAE1 can be broken down as follows: A stands for Alternaria (the genus of the fungus), F stands for Feruloyl (referring to feruloyl esterase activity), A stands for Acid (commonly linked to enzymatic function in biological nomenclature), E stands for Esterase (the enzyme class), and 1 indicates the number of protein or enzyme.
Analysis of Physiochemical Properties
Physiochemical properties of UniProt sequences were analyzed, focusing on molecular weight (kDa), amino acid composition, theoretical pI, and instability index. The instability index assesses protein stability; values <40 denote stability, while >40 suggest potential challenges in handling and expression, aiding researchers in anticipating stability issues. AFAE1 contained 324 amino acids, weighed approximately 35.20395 kDa, and had a theoretical pI of 4.67 and an instability index of 37.63, indicating potential instability. AFAE2 also comprised 324 amino acids, weighed about 35.27207 kDa, and had a theoretical pI of 4.82 and an instability index of 38.18, suggesting moderate instability. AFAE3 consisted of 330 amino acids, weighed approximately 35.59258 kDa, and had a theoretical pI of 6.70, indicating acidity. Its instability index of 43.40 suggested a higher propensity for instability under laboratory conditions. These findings provided insights for researchers to anticipate and address potential stability issues in experimental work. (Table S12)
Protein Domain Analysis
Functional domain analysis identified conserved protein domains crucial for understanding protein function and evolutionarily conserved sites. Protein domains contribute to stability and function as distinct functional or structural units within a protein. The Esterase_PHD domain, shared by AFAE1 and AFAE3, spanned residues 55–206, while for AFAE2, it spanned residues 55–209 (Figure 2). This domain in A. alternata facilitated the degradation of plant cell walls by targeting feruloyl-arabinose ester bonds in arabinoxylans, crucial polysaccharides found in wheat cells. The proteins' signal peptides aided in efficient trafficking within the cell, ensuring targeted deployment at specific cellular locations. A signal peptide is not considered a common domain. It is a short sequence that directs the protein to a specific cellular location, unlike domains, which are functional or structural units within a protein.
Protein Sequence Alignment
Protein sequence alignment via MUSCLE [version 3.8] was performed on Feruloyl esterase proteins to identify shared domains (Figure S21). However, the alignment results did not reveal common motifs, underscoring the diversity and complexity of Feruloyl esterase proteins. Despite this, the findings offer valuable insights for future research, particularly in understanding the role of Feruloyl esterase proteins from A. alternata in causing the black point of wheat. While shared domains were not observed, functional similarities or conserved regions may still exist. Further investigation using complementary approaches is warranted to explore potential functional relationships among these proteins.
Secondary Structure Prediction
The Protter was utilized to predict the secondary structures of the Esterase_PHD domains from three selected feruloyl esterase proteins. Protein secondary structure prediction (PSSP) seeks to develop a method that translates an amino acid sequence into its corresponding secondary structure, enabling the determination of the protein's secondary structure from its amino acid sequence as shown in Figure S22.
Visualization of Alphafold3 predicted Structures
The AlphaFold3 server was utilized to predict the structures of the Esterase_phd domains from three selected feruloyl esterase proteins. The predicted protein structures exhibited almost identical configurations and demonstrated generally high degrees of success. These structures were superimposed, resulting in a model with a high level of confidence and reliability. (Figure S23).
Verify 3D, Errat and ProCheck
The evaluation metrics for each protein based on its Protein ID, presented in Table S15, including the proportion of residues in the core region ("ProCheck"), the percentage of acceptable residues in the 3D structure ("Verify 3D"), and the percentage of prohibited residues ("ERRAT"). "AFAE1" achieved a quality factor of 99.3056%, with 88.0% of residues in the core region and only 0.8% disallowed residues. "AFAE2" scored 96.5753%, with 89.1% core residues and 0.8% disallowed residues. Despite generally high scores for all AlphaFold3 Esterase_phd models, "AFAE3" demonstrated lower scores of 67.11% and 90.9722%, indicating variability in model quality. This assessment offered insights into the precision and dependability of protein models across VERIFY 3D, ERRAT, and PROCHECK evaluations.
Ramachandran Plot
Ramachandran Plot analysis, as displayed in Table S16, showed the distribution of residues across different regions for each protein. For protein AFAE1, 88.0% of residues resided in the most favored region, followed by 11.2% in the additional allowed region, and 0.8% in the disallowed region. AFAE2 showed 89.1% of residues in the most preferred region, 10.2% in the additional allowed region, and 0.8% in the disallowed region. AFAE3 exhibited even higher percentages, with 88.9% in the most favored region, 9.5% in the additional allowed region, and 0.8% in both generously allowed and disallowed regions. These investigations offered standards for assessing the Ramachandran distributions of AlphaFold 3D models of A. alternata proteins.
Protein to Protein Interactions
Significant connections involving the esterase domain of A. alternata's Feruloyl esterase proteins were discovered through the identification of protein interaction partners using the STRING webserver. The significance of protein-protein interactions in cellular processes was highlighted by the Esterase_PHD domain's notable strong connections with both AA0117_g1633 and with itself. This implied a functional link with AA0117_g1633 and possible oligomerization or dimerization events that are essential for function (Figure S24 and S25). Deciphering intricate biological processes and disease mechanisms required an understanding of protein-protein interactions. These discoveries clarify the functions of proteins in networks and provide information for the creation of treatments that target these relationships.
Ligand Binding
The ChEMBL database, which provides bioactive compounds with drug-like qualities, was used to identify a potential molecule for binding to certain proteins. The Yang Cao Laboratory's web-based program CB-Dock2, which offers a sophisticated tool for protein-ligand blind docking, was utilized to help with ligand binding. Various fungicides were tested as ligands for docking with the Esterase_PHD of A. alternata. Propiconazole (Tilt) was found to disrupt ergosterol biosynthesis, potentially affecting ester bond hydrolysis. Chlorothalonil (Cymoxanil) interfered with multiple fungal processes, potentially hindering esterase function. Additionally, Cymoxanil inhibited enzyme activity by targeting cytochrome C oxidase. These interactions were visually represented, offering insights into their mechanisms within the Esterase_phd domain (Table S17).
Docking and MD Simulations
Molecular Docking Identifies Metalaxyl and Pyraclostrobin as Top Candidates
Pyraclostrobin–AFAE3 (–6.6 kcal/mol) , thiophanate-methyl_AFAE2 (–5.7 kcal/mol) and propiconazole_ AFAE1 (-5.6 kcal/mol) exhibited the most favorable binding characteristics among the simulated complexes, with AutoDock Vina-based docking scores ranging from –6.6 to –4.4 kcal/mol (Table. 1). These values suggest strong potential for stable, energetically favorable binding across the conserved catalytic sites.
Molecular Dynamics Confirms Binding Stability for Metalaxyl and Cymoxanil Complexes
For every chosen ligand–protein complex, 5 ns all-atom MD simulations were run to confirm the stability of docked postures in a dynamic environment. The findings showed significant variations in complicated behaviour after docking.
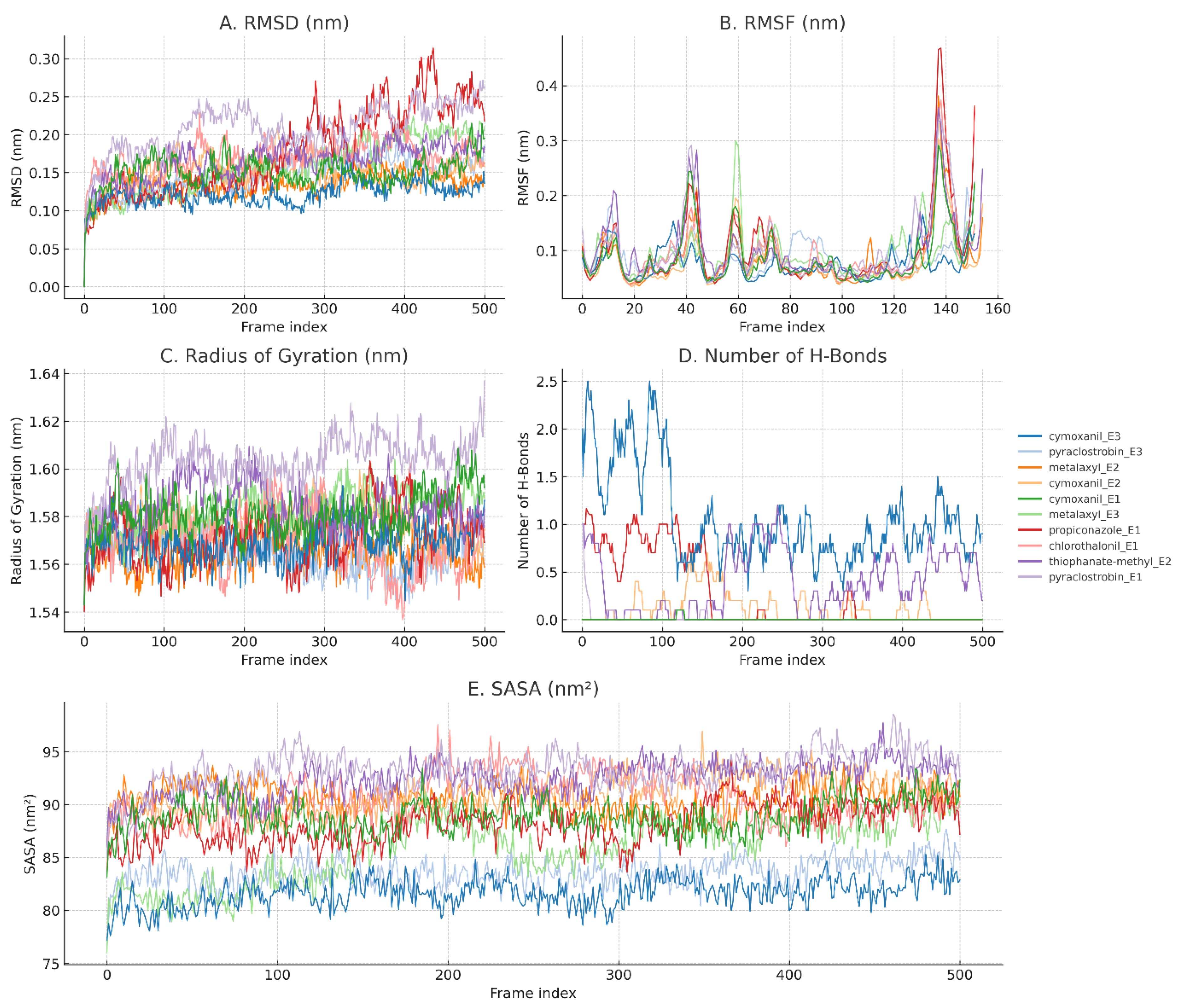
RMSD (Root Mean Square Deviation)
The best positional stability of the ligand at the binding site over time was indicated by the lowest RMSD values for Metalaxyl–AFAE2 (0.13 nm), Pyraclostrobin–AFAE3 (0.14 nm), and Cymoxanil–AFAE2 (0.14 nm) (shown in Figure 6A). Although not one of the absolute lowest, propiconazole–AFAE1 maintained an acceptable RMSD (~0.17 nm), indicating moderate positional stability.
Radius of Gyration (Rg)
The Rg values of all complexes remained between 1.56 and 1.60 nm, suggesting that the protein was generally stable and not unfolding. Comparable Rg to other ligands was displayed by propiconazole-containing complexes, indicating the receptor's structural integrity throughout simulation (Figure 6C).
RMSF (Root Mean Square Fluctuation)
All systems displayed minimal residue flexibility (average RMSF < 0.1 nm). Interestingly, the binding site residues in Metalaxyl–AFAE2 and Cymoxanil–AFAE2 were particularly rigid, indicating that the ligands had stabilising effects. In catalytic regions, propiconazole–AFAE2 also showed minimal RMSF, which is consistent with its capacity to sustain close connections (Figure 6B)).
SASA (Solvent Accessible Surface Area)
Among the SASA values, which varied from 81.62 to 93.23 nm² , cymoxanil–AFAE3 had the lowest value, signifying profound burial. The moderate SASA of propiconazole–AFAE2 (88.16 nm²) indicates partial but consistent pocket accommodation (Figure 6E).
Hydrogen Bonding
The only complex that consistently formed hydrogen bonds during simulation was cycloxanil–AFAE2 (mean = 0.12). Despite having a high in vitro efficacy, propiconazole–AFAE2 did not show substantial hydrogen bonding, suggesting that hydrophobic or shape-complementary binding rather than polar interactions may be the primary governing factors in their interaction (Figure 6D).
While Pyraclostrobin and Metalaxyl depend primarily on nonpolar and steric interactions for stable complex formation, Cymoxanil seems to use particular polar connections. Notably, propiconazole has a hybrid mode that is consistent with its potent experimental antifungal action, producing some hydrogen bonds while preserving favourable burial and RMSD.

Table 1.
Docking Affinities, Structural Stability (RMSD, Rg), Hydrogen Bonding, and Solvent Accessibility (SASA) of Selected Ligand–AFAE1–3 Complexes Based on 5ns MD Simulations.
Table 1.
Docking Affinities, Structural Stability (RMSD, Rg), Hydrogen Bonding, and Solvent Accessibility (SASA) of Selected Ligand–AFAE1–3 Complexes Based on 5ns MD Simulations.
| Ligand_Protein | Docking_Affinity (kcal/mol) | RMSD_Mean (nm) | Rg_Mean (nm) | Hbond_Avg | SASA_Avg (nm²) |
|---|---|---|---|---|---|
| pyraclostrobin_e3 | -6.6 | 0.147 | 1.566 | 0 | 83.58 |
| pyraclostrobin_e1 | -6.4 | 0.209 | 1.603 | 0.01 | 93.23 |
| thiophanate-methyl_e2 | -5.7 | 0.165 | 1.584 | 0.39 | 92.4 |
| propiconazole_e1 | -5.6 | 0.178 | 1.572 | 0.26 | 88.16 |
| metalaxyl_e2 | -5.4 | 0.135 | 1.564 | 0 | 90.54 |
| cymoxanil_e2 | -5.1 | 0.148 | 1.576 | 0.12 | 90.9 |
| metalaxyl_e3 | -5 | 0.157 | 1.583 | 0 | 85.78 |
| cymoxanil_e1 | -4.6 | 0.151 | 1.581 | 0 | 88.95 |
| cymoxanil_e3 | -4.5 | 0.122 | 1.569 | 1.05 | 81.62 |
| chlorothalonil_e1 | -4.4 | 0.17 | 1.569 | 0 | 90.54 |
Figure 3.
Composite Stability Ranking of Ligand–AFAE Complexes.
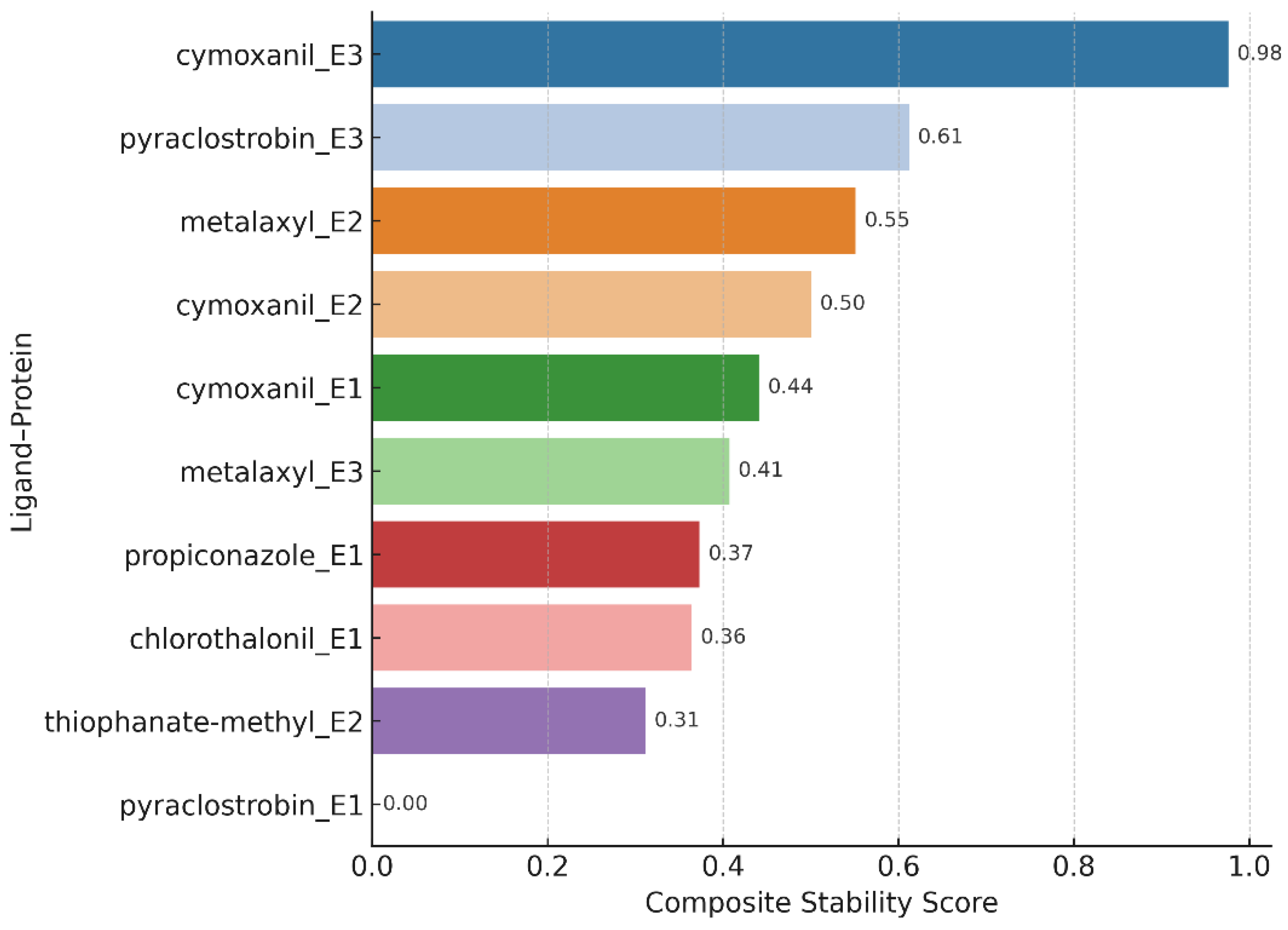
Figure 6.
Molecular Dynamics Simulation Metrics for Ligand–AFAE Complexes. (A) Root Mean Square Deviation (RMSD) trajectories of ligand–protein complexes over 5 ns show overall positional stability, particularly for metalaxyl_E2, cymoxanil_E2, and pyraclostrobin_E3. (B) Root Mean Square Fluctuation (RMSF) profiles demonstrate local flexibility of protein residues, with most complexes showing limited fluctuations (<0.15 nm). (C) Radius of Gyration (Rg) indicates compactness of the protein-ligand complex, with consistent values (~1.56–1.60 nm) across all systems. (D) Number of Hydrogen Bonds between ligand and protein throughout the trajectory. Notably, cymoxanil_E3 and thiophanate-methyl_E2 formed more stable hydrogen bonds compared to other ligands. (E) Solvent Accessible Surface Area (SASA) shows burial trends of ligands in the binding pocket; lower SASA values suggest deeper binding and reduced solvent exposure.
Figure 6.
Molecular Dynamics Simulation Metrics for Ligand–AFAE Complexes. (A) Root Mean Square Deviation (RMSD) trajectories of ligand–protein complexes over 5 ns show overall positional stability, particularly for metalaxyl_E2, cymoxanil_E2, and pyraclostrobin_E3. (B) Root Mean Square Fluctuation (RMSF) profiles demonstrate local flexibility of protein residues, with most complexes showing limited fluctuations (<0.15 nm). (C) Radius of Gyration (Rg) indicates compactness of the protein-ligand complex, with consistent values (~1.56–1.60 nm) across all systems. (D) Number of Hydrogen Bonds between ligand and protein throughout the trajectory. Notably, cymoxanil_E3 and thiophanate-methyl_E2 formed more stable hydrogen bonds compared to other ligands. (E) Solvent Accessible Surface Area (SASA) shows burial trends of ligands in the binding pocket; lower SASA values suggest deeper binding and reduced solvent exposure.
Machine Learning & xAI: Performance of Pesticides Under Various Climate Conditions
Effect of Individual Weather Parameters on Pesticide Performance
This study highlighted the effects of individual parameters and their combined impact on pesticide performance. The analysis examined the interactions between individual weather parameters (temperature, humidity, and precipitation) and various pesticides in controlling Black Point disease. Using averaged SHAP interaction values from 1,000 Monte Carlo simulations, the examination offered solid insights into the separate effects of each climate variable on pesticide efficacy. The various impacts of meteorological conditions on pesticide performance are reported in Table S19.
Temperature and Fungicides Combined Efficacy
The findings shed important light on how temperature affects the effectiveness of pesticides. Propiconazole, Metribuzine, and Propoxycarbazone are examples of extremely temperature-dependent pesticides that were found to significantly improve their potency under warm settings, making them ideal for usage in hotter climates or throughout the summer. Thiophanate-methyl, on the other hand, worked best in colder temperatures, indicating that it is appropriate for early-season use or temperate regions. Pesticides that are widely adjustable, such as Metalaxyl-M + Mancozeb, Sulfosulfuron, and Chlorothalonil + Metalaxyl-M, showed steady performance throughout a wide range of temperatures, giving them flexible choices for areas with erratic or fluctuating climates. Indicating broad performance without particular optimization for extreme temperature circumstances, moderate performers like Dichlorodiphenyltrichloroethane and Cymoxanil + Mancozeb showed consistent yet moderate efficacy over the temperature spectrum (Figure S26). Black Point disease is more effectively managed by coordinating pesticide use with climate, improving crop rotation, reduce resource waste. To maintain the sustainability of agriculture in the face of shifting environmental conditions, these results highlighted that climate-smart pest management techniques are crucial.
Humidity and Fungicides Combined Efficacy
Because high moisture levels foster the growth of fungal pathogens, targeted treatments of fungicides are essential in high-humidity environments, which boosts their efficacy. On the other hand, fungicide efficacy decreases under low humidity, which is indicative of decreased disease pressure and decreased pathogen activity.
Based on SHAP interaction values averaged over 1,000 Monte Carlo simulations, Figure S27 shows how different pesticides and humidity conditions interact to suppress black point disease. Propiconazole and Cymoxanil + Mancozeb demonstrated robust positive responses under high humidity circumstances, according to the analysis, which makes them very useful in tropical areas or during rainy seasons. Metribuzine, Propoxycarbazone, and Thiophanate-methyl, on the other hand, worked best at moderate humidity levels, which makes them appropriate for temperate climates with consistent moisture levels. Broadly versatile pesticides, such as Chlorothalonil + Metalaxyl-M, Metalaxyl-M + Mancozeb, and Sulfosulfuron, showed consistent efficacy at all humidity levels, making them dependable choices for areas with erratic or changing moisture conditions. Dichlorodiphenyltrichloroethane performed steadily and moderately in a variety of settings; it was not significantly optimized for extremely dry or wet climates. Overall, the results highlighted how crucial it is to adjust pesticide applications to local humidity levels in order to maximize crop yields, minimize resource waste, and control pests. The need of climate-smart pest management techniques for sustainable agricultural production was emphasized by this study.
Precipitation and Fungicide Efficacy
Higher fungicide utility is correlated with increased precipitation. In order to reduce the severity of the disease, timely fungicide applications are necessary because wet circumstances create an ideal setting for fungal growth. However, the disease's decreased frequency in arid environments reduces the requirement for fungicides.
Based on SHAP interaction values averaged over 1,000 Monte Carlo simulations, Figure S28 shows how different pesticides and precipitation levels interact to affect the incidence of Black Point Disease. Propiconazole and Cymoxanil + Mancozeb showed the most effectiveness under high rainfall, according to the analysis, which makes them perfect for tropical or subtropical areas with a lot of precipitation. Propoxycarbazone, Metribuzine, and Thiophanate-methyl, on the other hand, worked well in areas with moderate rainfall, which makes them ideal for temperate or semi-arid climates with consistent precipitation. Pesticides that are widely adjustable, such as Metalaxyl-M + Mancozeb, Sulfosulfuron, and Chlorothalonil + Metalaxyl-M, showed dependable performance at all precipitation levels, guaranteeing dependability in regions with erratic or changeable rainfall. Despite its lack of optimization for harsh conditions, Dichlorodiphenyltrichloroethane demonstrated stable but moderate performance throughout a range of precipitation levels. Climate-smart pest control tactics can be implemented by coordinating pesticide treatments with regional rainfall patterns, which can enhance crop yields, improve disease management, and encourage sustainable farming practices.
Pairwise Interactions Between Climate Variables and Pesticides
In controlling black point disease, important information about the effectiveness of pesticides was uncovered by examining the pairwise interactions between humidity, temperature, and precipitation. With their great dependence on high humidity and precipitation, Propiconazole, Metalaxyl-M + Mancozeb, and Cymoxanil + Mancozeb were the most effective in wet and humid conditions, making them perfect for tropical or subtropical regions. Sulfosulfuron and Chlorothalonil + Metalaxyl-M showed steady performance across a range of climates, demonstrating their flexibility in response to changing weather patterns. Propoxycarbazone and Metribuzine showed their most effectiveness in balanced conditions with moderate temperature, humidity, and precipitation, whereas Thiophanate-methyl and Sulfosulfuron functioned consistently in semi-arid areas with moderate precipitation and reduced humidity. These results underlined how crucial it is to match pesticide application methods to local climate circumstances to maximize crop protection, minimize waste, and improve sustainability. It was advised to prioritize moisture-dependent pesticides like Propiconazole or Cymoxanil + Mancozeb in wet conditions, while Thiophanate-methyl or Sulfosulfuron produced more dependable outcomes in dry areas. To increase agricultural productivity, this study laid the groundwork for climate-smart pest management techniques (Figure 4).
Triple Interaction Analysis of Climate Variables on Pesticides Efficacy
Different patterns in the effectiveness of pesticides in treating black point disease were found by analyzing the triple interaction of temperature, humidity, and precipitation. Metalaxyl-M + Mancozeb is perfect for rainy areas because it showed maximum effectiveness in conditions of moderate temperature, high humidity, and regular precipitation. Propoxycarbazone was less effective in harsh environments but did well in areas with steady temperatures and moderate rainfall. Sulfosulfuron and Chlorothalonil + Metalaxyl-M showed extensive adaptability, delivering constant performance in a range of conditions, making them dependable for erratic weather patterns. Propiconazole and Cymoxanil + Mancozeb flourished in wet and humid settings but were less effective in dry ones, whereas metribuzine worked best in semi-arid areas with mild temperatures and balanced moisture levels. In semi-arid regions with moderate precipitation and reduced humidity, thiophanate-methyl proved to be dependable. Dichlorodiphenyltrichloroethane, on the other hand, had an uneven performance in extreme weather conditions and performed better in stable, moderate temperatures, necessitating cautious administration. These results highlight how crucial it is to match pesticide use to local climate conditions in order to improve sustainability and efficacy.
Figure 5.
Pesticides combined impact on precipitation, temperature, and humidity.
Discussion
Black point is an economically significant disease of wheat prevalent in wheat-growing regions worldwide. It causes both qualitative and quantitative losses. In sensitive cultivars, disease-related grain losses range from 24 to 27% (El-Gremi et al., 2017). This study tested the efficacy of various systemic fungicides in two trials (a fungicides trial and a Tilt fungicide dose trial) conducted at NIFA, Peshawar, using the common variety “Morocco.” Parameters such as disease incidence, disease severity index, thousand kernel weight (TKW), and the pathogens responsible for causing the disease were examined. The efficacy of field-used chemical fungicides, including Tilt (Propiconazole, foliar), Success, Ridomil, Topsin M, and Cymoxanil+Mancozeb, against A. alternata was assessed using the agar well diffusion technique. Additionally, the study aimed to identify and model the virulent proteins of A. alternata, for which the 3D structures have not yet been solved or predicted. The study also analyzed the individual and combined influence of temperature, humidity, and precipitation on pesticide efficacy against black point disease using SHAP interaction values and Monte Carlo simulations.
In this study, the efficacy of five selected fungicides was evaluated against black point disease. Tilt (5.7%) was the most effective fungicide, resulting in the lowest disease incidence compared to the control (8.6%), followed by Cymoxanil+Mancozeb (6.0%), Success (6.3%), Topsin-M (6.5%), and Ridomil (7.0%). Different doses of Tilt fungicide also reduced disease incidence compared to the control. In the dose trial, the most effective dose was dose 4 (5.7%), followed by dose 1 (6%), dose 2 (6.3%), dose 5 (6.6%), and dose 3 (7.4%). At 9.1%, the illness incidence was highest in the control group. Tilt successfully manages black point infection, according to other researchers. According to Malaker et al., (2009), black point incidents were successfully reduced by foliar spraying Tilt-250EC (Propiconazole) on different schedules (Malaker et al., 2009). According to Iqbal et al., (2014), Tilt (propiconazole @ 500 ml/ha) was the most effective against black point disease and yielded an outstanding economic return of Rs. 13200/ha (Iqbal et al., 2014). Other authors also examined the effectiveness of six fungicides in 3 concentrations (100, 300, and 500 ppm) against A. alternata on PDA in vitro. The mycelial growth of A. alternata was reduced with propiconazole (94, 100, and 100%), followed by trifloxystrobin+tebuconazole (75, 90.11, and 95.88%). The mycelial growth was less affected by fungicides such as hexaconazole (70, 76.33, and 85.44%), carbendazim + mancozeb (64.25, 69.15, and 80.10%), azoxystrobin (57.00, 62.33, and 70.00%), and mancozeb (55.70, 59, and 65.66%). Then the most effective fungicides such as propiconazole and trifloxystrobin + tebuconazole in vitro were also studied as foliar sprays against A. alternata in microplots. Both fungicides considerably outperformed controls in terms of lowering disease intensity at 60 and 90 days of transplantation (DAT). At 60 and 90 DAT, trifloxystrobin + tebuconazole (7.80 and 27.90%) and propiconazole (5.00 and 23.00%) exhibited the lowest disease intensity (Yadav et al., 2020). Propiconazole fungicide is essential for resistance control measures. Farmers can mix or rotate fungicides with different modes of action to prevent the development of resistance. Because of its unique mode of action, propiconazole is essential for integrated pest management (IPM) programs. Environmentally, Propiconazole has a favorable profile when used correctly. Adhering to recommended rates and intervals minimizes environmental impact and health risks. However, efficacy may vary based on timing, conditions, and targeted pathogens. Following local regulations and label instructions is imperative for all agricultural chemicals (Box, 2015).
In this study, the severity index for both trials was also evaluated. Cymoxanil+Mancozeb had the lowest severity index (2.7%), followed by Tilt (3%), Success (3.1%), and Ridomil (3.3%). The highest severity index was observed for Topsin-M (4.7%). The control group exhibited a severity index of 4.1%. The severity index for kernel black point disease in the Tilt fungicide trial varied across different doses of the fungicide. Dose 4 had the lowest severity index (3.3%), followed by dose 2 (3.6%), dose 3 (3.8%), dose 5 (4.1%), and dose 1 (4.2%). The control group had the highest severity index of 4.4%. Cromey and Mulholland (1988) described the black point severity index, noting significant variation in black point infection levels in grain samples (Cromey and Mulholland, 1988). In another study, Draz et al., (2016) examined the black point disease severity index over two consecutive years, finding that the illness index ranged from 0.29 percent to 19.48 percent on average across both seasons (Draz et al., 2016). Black point disease severity varied widely between seed samples. Wheat cultivars with the lowest black point disease incidence and severity index may be valuable as sources of black point resistance germplasm. According to Srivastava et al., (2015), relatively higher temperatures, high relative humidity, and intermittent rains during anthesis promote the manifestation of black point symptoms. It was also noted that delayed watering (irrigation) combined with high wind velocity increased the severity of black point illness (Srivastava et al., 2015). The infection of winter wheat seeds by black point varies depending on the year of research and the variety.
For both fungicide experiments in this investigation, the weight of a thousand kernels was measured. TKW affects germination, seed vigor, seedling establishment, and yield, making it a crucial indication of seed quality. At 35.5 g, tilt (Propiconazole) had the greatest TKW, followed by Ridomil (29.7 g), Cymoxanil + Mancozeb (34 g), Topsin-M (30.9 g), and Succes (30.6 g). Compared to the control group, which had the lowest TKW (25.9 g), samples with the highest TKW showed a decreased incidence of black point illness. According to Shahwani et al., (2014), the most promising agronomic performance was shown by wheat with big seeds (Shahwani et al., 2014). Muhsin et al., (2021) found that bold seeds yielded the highest TKW (36.97 g) and the most grains per spike (45.17), whereas medium seeds produced the lowest TKW (32.03 g) and the fewest grains per spike (34). Bold seeds produce the greatest growth and increase yield as compared to other seed size classes. Therefore, wheat yield can be increased even under late-sown conditions because of their bold seed size, which enhances yield-related features and encourages vigorous germination (Muhsin et al., 2021). Another study found no significant difference between TKW and germination percent, germination rate, or mean germination time using a standard germination test. Nonetheless, germination percent and germination rate grew in conjunction with TGW, with the highest TGW being linked to the highest germination rate and percent. According to Moshatati and Gharineh (2012), the treatments with the highest and lowest TKW had the largest and lowest mean germination times, respectively (Moshatati & Gharineh, 2012). In conclusion, bold seeds significantly enhanced growth and yield as compared to other seed size classes. In late-planted situations, bold seed size can increase wheat production because of its strong and quick germination and improved yield-related characteristics. Before offering suggestions to the farming community, more research is necessary on different soil types and agroecosystems.
From the black-pointed diseased seeds, the fungus was isolated in the study. The frequently isolated fungus from all the disease seeds was A. alternata. Draz et al., (2016) and El-Gremi et al., (2017) conducted a study in which A. alternata, Cochliobolus sativus (Helminthosporium sativum), and Fusarium graminearum were commonly isolated from diseased seeds (El-Gremi et al., 2017; Draz et al., 2016). Cotuna et al., (2020) described that A. alternata is a common causal agent of wheat kernels infected by black point disease (Cotuna et al., 2020). In a study conducted by Draz et al., (2016), A. alternata isolated from black-pointed wheat grains was responsible for the occurrence of leaf spots, reduced grain germination, limited seedling growth, and finally losses in yield quantity and quality (Draz et al., 2016). Numerous bacteria are linked with the discoloration of wheat grains which destroys the appearance of the final product (Fernandez & Conner, 2011). However, the prevalence and incidence of these pathogens differ. The disease's prevalence also affects how much rain falls during seed development or early leaf senescence, which encourages black point. The pathogen's capacity to infect wheat types and raise the incidence and prevalence of black point disease is facilitated by increased precipitation and the frequency of wet days. The growth of black point or kernel smudge is highly weather-dependent. There is limited control over this issue, particularly in regions that receive significant rainfall (Ijaz et al., 2018). SESİZ (2023) also stated that current temperature, precipitation, and humidity, particularly during grain-filling periods, create an ideal environment for the development of black point components, which could be caused by fungal pathogens or oxidation reactions (SESİZ, 2023).
The five fungicides—Tilt (Propiconazole), Success, Ridomil, Topsin M, and Cymoxanil+Mancozeb—used in the field were also utilized in the laboratory to assess their efficacy against A. alternata using the agar well diffusion technique. Among these fungicides, tilt (Propiconazole) demonstrated the highest efficacy in inhibiting the growth of A. alternata, displaying a zone of inhibition measuring 28 mm. Cymoxanil+Mancozeb showed an 8 mm inhibitory zone after tilt. However, in lab settings, Success, Ridomil, and Topsin M—the other fungicides—did not show any discernible efficacy against A. alternata. This means that the fungicides had different degrees of efficacy, with propiconazole showing the highest reduction in the growth of A. alternata in the laboratory. Achilonu et al., (2023), studied the effect of six fungicides on the mycelial development of isolates of A. alternata derived from Alternaria Black Spot symptoms in vitro. Three different concentrations (0.2, 1, and 5 μg mL−1) of fungicides were tested, including Boscalid + Pyraclostrobin, Fentin Hydroxide, Azoxystrobin, and Propiconazole. Notably, Tilt and Bumper 250 EC, which has propiconazole as an active ingredient and acts as a demethylation inhibitor, showed the best performance, completely stopping mycelial growth for as long as six days after incubation (Achilonu et al., 2023). Dasharathbhai Ajayabhai et al., (2018) assessed nine fungicides and five bioagents against A. alternata (Fr.) Keissler causing leaf blight disease, at three concentrations (250, 500, and 1000 ppm). These included Carbendazim 50 WP, Hexaconazole 5 EC, , Chlorothalonil 75 WP, Propiconazole 25 EC, Benomil 50 WP, Difenconazole 20 + Azoxystrobulin 12.5 EC, Mancozeb 75 WP, Tebuconazole 25.9 EC, and Carbendazim 12% + Mancozeb 63% 75 WP. The fungal growth was completely inhibited by Propiconazole and Hexaconazole (250 ppm) at all three concentrations (Dasharathbhai Ajayabhai et al., 2018). Based on these investigations, A. alternata was best controlled by propiconazole as compared to other fungicides. Propiconazole had a high capacity to prevent the growth of A. alternata. Propiconazole is more efficient in the treatment of A. alternata infections which emphasizes its applicability in agricultural settings.
Ipyrobin (Iprodione + Pyraclostrobin), is a combination fungicide, which is used to manage a variety of fungal diseases in crops in agriculture. In lab work studies, this fungicide was used as a positive control to compare its effectiveness with field-applied fungicides. It showed the ability to limit the A. alternata’s growth. However, we suggested propiconazole because it is less harmful to human health. Further in silico analysis was performed for Propiconazole, due to its lower toxicity to humans. Propiconazole often shows a reduced toxicity when compared to ipyrobin. Propiconazole is categorized as moderately dangerous when taken orally and slightly toxic when used topically but ipyrobin may have higher toxicity levels depending on the precise formulation and concentration (Wang et al., 2018). Propiconazole generally reduces environmental persistence by controlling the risk of environmental buildup and exposure to fungal disease which may result in a quicker breakdown in soil and water.
This study combines Explainable AI (XAI) techniques, notably SHAP (SHapley Additive exPlanations), with an XGBoost (Extreme Gradient Boosting) machine learning model to provide interpretable insights into the effect of climatic variables on the efficacy of fungicides. The lab-based inhibitory zone data and simulated climate conditions were used and the model evaluated the individual and combined effects of precipitation, temperature, and humidity on fungicide performance. The impact of these factors and their combinations on pesticide effectiveness was brought to light by the SHAP analysis (Prendin et al., 2023). The results of this study showed that different fungicides performed differently in a range of meteorological circumstances, such as temperature, humidity, and precipitation. Propiconazole and Cymoxanil + Mancozeb, two moisture-dependent fungicides, showed their best results in areas with high precipitation and humidity. While Thiophanate-methyl proved more effective in semi-arid settings with moderate rainfall and lower humidity, region-specific fungicides such as Metribuzine worked best at slightly warmer temperatures and moderate humidity. Adaptable fungicides, such as Sulfosulfuron and Chlorothalonil + Metalaxyl-M, on the other hand, demonstrated steady performance over a broad range of temperatures, humidity levels, and precipitation circumstances, making them appropriate for regions with erratic weather patterns. These results emphasize that while designing fungicide treatment techniques for efficient black point disease control, particular meteorological conditions, such as temperature, humidity, and precipitation, must be taken into account. Extreme weather patterns such as increasing temperatures, fluctuating precipitation, and changes in humidity provide serious obstacles to sustainable agriculture as climate variability increases (Kharin et al., 2013). According to research, a 1°C rise in temperature can result in a 10–20% drop in agricultural yield worldwide (Barlow et al., 2015). These temperature changes have a major effect on yield, grain weight, and grain size, especially during crucial phases of wheat growth including blooming, anthesis, and the milking stage (Talukder et al., 2014). For example, it has been demonstrated that dry climates in northern Europe can interfere with flowering timing, resulting in significant output losses (Bodner et al., 2015). The necessity for dynamic, climate-responsive solutions to sustain fungicide efficacy and agricultural output is highlighted by the rising frequency of extreme climate events. Globally more robust and sustainable wheat production systems can be ensured by incorporating climate data into disease management frameworks to help reduce yield losses brought on by climate-induced variability.
Conclusions
It is concluded from this study that propiconazole was the most effective fungicide in both field trials and lab work and showed the highest efficacy in the field resulting in lower disease incidence and increased yield. It also proved effective in the laboratory against A. alternata, showing significant growth inhibition. Numerous A. alternata proteins implicated in virulence against the host were identified by computational research. To better understand the methods used by A. alternata to tolerate even advanced-generation fungicides, it is still necessary to conduct experiments to clarify the structure of these proteins. Additionally, the study found that three targeted proteins' Esterase_PHD domain is essential to A. alternata's ability to survive inside the host cell. By changing the esterase_PHD domain via active ligand binding, A. alternata infections may be possibly controlled. Propiconazole resulted in the highest vina scores by interacting with specific amino acid residues of targeted proteins at the C1 pocket.
Climate variables like as temperature, humidity, and precipitation all have a substantial impact on fungicide effectiveness in the management of Black Point Disease. Moisture-dependent fungicides, such as Propiconazole and Cymoxanil + Mancozeb, performed best in areas with high humidity and frequent rainfall. Region-specific fungicides, such as Metribuzine, worked best in moderate temperature and humidity environments, but Thiophanate-methyl proved more effective in semi-arid regions with moderate rainfall and lower humidity. Adaptable fungicides like Chlorothalonil + Metalaxyl-M and Sulfosulfuron have shown consistent effectiveness over a wide range of climatic circumstances, making them ideal for places with changing weather. These findings emphasize the need of taking into account local meteorological parameters when selecting fungicides to maximize disease control and agricultural operations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Figure S1: Location of the study area, NIFA. Table S1: Mode of action, active ingredients, and doses of five fungicides against black point. Table S2: Fungicide concentrations used in the field. Table S3: Five Tilt doses used against black point disease. Table S4: Replications/Treatments of five fungicides against the black point in Randomized Complete Block Design. Table S5: Treatments of five doses of fungicides “Tilt” against the black point disease in Randomized Complete Block Design. Figure S2: Fungicides and Sprayer used in this field to control black point disease of wheat. Figure S3: Collection of samples from the field. (A, B) The wheat treatment-wise seeds were harvested at the NIFA farm; (C) Threshing of the wheat samples; (D) The seeds were collected from the thresher machine; (E) The wheat samples were put in plastic bags containing their variety number. Figure S4: Counting of the wheat seeds samples. (A) Seed Buro 801, an automatic seed counter machine; (B) Process of seed counting; (C) Thousand-grain weight of wheat seed sample was recorded by using sartorius balance. Figure S5: Incidence of black point wheat disease. (A) Black pointed seeds separated from healthy seeds; (B) Magnifying glass lens (10X) used for the separation of black pointed seeds; (C) Wheat seeds containing dark black color at embryo end. Figure S6: wheat kernels severity index of Black Point disease. (A) Infected wheat seeds by black point disease-causing pathogens; (B) Seeds according to severity scores 1, 2, and 3. (Score 0 = no symptoms of black point, Score 1 = mild smudging, particularly on every side of the embryo with ventral crease, Score 2 = a distant darkening near the embryo's end, as well as a ventral crease, Score 3 = the grain one-third part has been extensively blackened). Figure S7: Slide preparation and microscopy of fungus isolated from black pointed seeds. Figure S8: The figure shows the before, after, front, and back pictures of some black pointed seed samples that were inoculated on PDA media. Figure S9: Agar well diffusion of 5 fungicides against A. alternata. Figure S10: Serial dilution of wheat farm soil used for two trials investigated in this study. This technique was employed to isolate fungi from the soil using PDA. First, 1 gram of soil was added to 9 ml of distilled water, thoroughly stirred for 15 minutes, and then vortexed. Serial dilutions from 10-1 to 10-5 were prepared. To isolate fungi from the diluted samples, the spread plate method was used. Specifically, 1 ml from each dilution was added onto the PDA and evenly distributed using an L-shaped glass rod (spreader). The plates were incubated for 5 to 7 days at a temperature of 25 to 27°C and then observed for growth. Table S6: The physiochemical properties of wheat field soil. Table S7 Thousand kernel weight, number of black point infected seeds, and black point disease incidence percentage of five fungicides used in this study. (The data presented in the table represents the average values obtained from six replications). Figure S11: Thousand kernel weight, number of black point infected seeds, and black point disease incidence percentage of five fungicides used in this study. Table S8: The infection severity index for kernel black point disease in five test fungicides used in this study. (The data presented in the table represents the average values obtained from six replications). Figure S12: Different doses efficacy used against black point wheat disease. Table S9 Thousand kernel weight, number of black point infected seeds, and black point disease incidence percentage of five doses of fungicide Tilt used in this study. (The data presented in the table represents the average values obtained from six replications). Figure S13: Thousand kernel weight, number of black point infected seeds, and black point disease incidence percentage of five doses of fungicide Tilt used in this study. Table 10: The infection severity index for kernel black point disease in five doses of Tilt fungicide used in this study. (The data presented in the table represents the average values obtained from six replications). Figure S14: The infection severity index for kernel black point disease in five doses of Tilt fungicide used in this study. Figure S15: Pure culture Alternaira alternata isolated from black pointed seeds. Figure S16: Microscopy of Alternaria alternata under a compound microscope. (A, B) Microscopy of Alternaria alternata under 40x magnification lens; (C, D) Microscopy of Alternaria alternata under 100x magnifying lens. The colonies of A. alternata were often black, though they might also be white or grey sometimes. Conidiophores were golden brown and found in small clusters, they could be straight or flexuous, branched or simple. Long chains of obclavate, ellipsoidal, or obpyriform conidia were observed, which had a small conical or cylindrical beak, up to eight transverse, and numerous longitudinal septa. The spores of A. alternata are 10–20 um thick and 20–70 um in diameter. Figure S17: Tilt (Propiconazole) fungicide demonstrates activity against the A. alternata fungus, effectively inhibiting its growth. The C+ well serves as a positive control, containing ipyrobin fungicide, a commercially available fungicide used for comparison. The C- well contains distilled water as the negative control, as all the fungicides were dissolved in it. Figure S18: The figure displays the results of all five fungicides, with Tilt demonstrating the highest activity in inhibiting the growth of A. alternata, followed by Cymoxanil+Mancozeb. None of the other fungicides were effective against A. alternata under laboratory conditions. Figure S19: Pure cultures of fungi i.e.: Aspergillus Niger, Aspergillus Fumigatus and Alternaria alternata isolated through a process of serial dilution. Figure S20: The Ipyrobin fungicide used as a positive control in the agar well diffusion method. Table S11: The table shows the Protein ID, Accession number, protein name, and FASTA sequence of 3 selected A. alternata proteins. Table S12: Physiochemical properties of three selected proteins of A. alternata. Table S13: Domain analysis of A. alternata proteins by using motif finder. Table S14: Domain analysis of Alternaria alternata protein by using SMART. Figure S21: Depicts the multiple sequence alignment of three Esterase_PHD domain-containing proteins from A. alternata, conducted using MUSCLE. In the alignment, amino acids marked in red represent the conserved domain, while those in blue denote semi-conserved domains. Figure S22: Secondary structure prediction of A. alternata by using Protter. Figure S23: AFAE1, AFAE2, AFAE3 AlphaFold 2 Colab structures and superimposition of Esterase phd of all three proteins. Table S15: ERRAT, Verified 3D, and ProCheck of all the selected proteins of A. alternata. Table S16: Ramachandran Plot of all three selected proteins. Figure S24: AFAE2 protein of A. alternata interaction with other proteins. Figure S25: AFAE3 protein of A. alternata interaction with other proteins. Table S17: Curpockets of five selected fungicides including positive control. Table S18: Bind docking of five selected fungicides including positive control with three selected proteins.
Funding
This work was supported by the Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2025R411), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia and by the Nuclear Institute for Food and Agriculture (NIFA) Peshawar.
Declaration of Interest Statement
The authors declare no conflicts of interest regarding the publication of this manuscript. The research was conducted without any involvement of organizations or entities that could have influenced the outcomes. The study received institutional support and was performed in compliance with ethical research standards.
Disclosure Statement
The authors reported no potential conflicts of interest.
References
- Abade, A., Ferreira, P. A., & de Barros Vidal, F. (2021). Plant diseases recognition on images using convolutional neural networks: A systematic review. Computers and Electronics in Agriculture, 185, 106125.
- Achilonu, C. C., Gryzenhout, M., Ghosh, S., & Marais, G. J. (2023). In Vitro Evaluation of Azoxystrobin, Boscalid, Fentin-Hydroxide, Propiconazole, Pyraclostrobin Fungicides against Alternaria alternata Pathogen Isolated from Carya illinoinensis in South Africa. Microorganisms, 11(7).
- Alastruey-Izquierdo, A., Melhem, M. S., Bonfietti, L. X., & Rodriguez-Tudela, J. L. (2015). Susceptibility test for fungi: clinical and laboratorial correlations in medical mycology. Revista do Instituto de Medicina Tropical de São Paulo, 57, 57-64.
- Ali, Y., Abbas, T., Aatif, H. M., Ahmad, S., Khan, A. A., & Hanif, C. M. S. (2022). Impact of Foliar Applications of Different Fungicides on Wheat Stripe Rust Epidemics and Grain Yield. Pakistan Journal of Phytopathology, 34(1), 135–141.
- Arnal Barbedo, J. G. (2013). Digital image processing techniques for detecting, quantifying and classifying plant diseases. SpringerPlus, 2(1), 660.
- Asif, M., Strydhorst, S., Strelkov, S. E., Terry, A., Harding, M. W., Feng, J., & Yang, R. C. (2021). Evaluation of disease, yield and economics associated with fungicide timing in Canadian Western Red Spring wheat. Canadian Journal of Plant Science, 101(5), 680–697.
- Balouiri, M., Sadiki, M., & Ibnsouda, S. K. (2016). Methods for in vitro evaluating antimicrobial activity: A review. Journal of Pharmaceutical Analysis, 6(2), 71–79.
- Binkowski, T. A. (2003). CASTp: Computed Atlas of Surface Topography of proteins. Nucleic Acids Research, 31(13), 3352–3355.
- Barlow, K. M., Christy, B. P., O’Leary, G. J., Riffkin, P. A., & Nuttall, J. G. (2015). Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field crops research, 171, 109-119.
- Box, P. O. (2015). States Environmental Protection Agency. Toxics Release Inventory (TRI) National Analysis, 703.
- Brown, P., Baxter, L., Hickman, R., Beynon, J., Moore, J. D., & Ott, S. (2013). MEME-LaB: motif analysis in clusters. Bioinformatics, 29(13), 1696–1697.
- Chi, D. H., Giap, V. D., Anh, L. P. H., & Nghi, D. H. (2017). Feruloyl esterase from Alternaria tenuissima that hydrolyses lignocellulosic material to release hydroxycinnamic acids. Applied Biochemistry and Microbiology, 53(6), 654–660.
- COTUNA, O., PARASCHIVU, M., SĂRĂȚEANU, V., DURĂU, C., & RECHIȚEAN, I. (2020). Research Regarding the Identification of the Fungus“Black Point” in Several Wheat Varieties Cultivated in Western Romania (Case Study). Life Science and Sustainable Development, 1(2), 25–31.
- Crepin, V. F., Faulds, C. B., & Connerton, I. F. (2004). Functional classification of the microbial feruloyl esterases. Applied Microbiology and Biotechnology, 63(6), 647–652.
- Cromey, M. G., & Mulholland, R. I. (1988). Blackpoint of wheat: Fungal associations, cultivar susceptibility, and effects on grain weight and germination. New Zealand Journal of Agricultural Research, 31(1), 51–56.
- Dasharathbhai Ajayabhai, C., Kedar Nath, Bekriwala, T., & Mabhu Bala. (2018). Management of Alternaria leaf blight of groundnut caused by Alternaria alternata. Indian Phytopathology, 71(4), 543–548.
- Degewione, A., & Alamerew, S. (2013). Genetic Diversity in Bread Wheat (Triticum aestivum L.) Genotypes. Pakistan Journal of Biological Sciences, 16(21), 1330–1335.
- DeLano, W., & Bromberg, S. (2004). PyMOL User’s Guide (Original). DeLano Scientific LLC, 1–66.
- Draz, I. S., El-Gremi, S. M., & Wassief Abd-Elsamad Youssef. (2016). Pathogens associated with wheat black-point disease and responsibility in pathogenesis. Journal of Environmental and Agricultural Sciences, 8(September 2016), 71–78.
- Draz, I. S., El-Gremi, S. M., & Youssef, W. A. (2016). Response of Egyptian wheat cultivars to kernel black point disease alongside grain yield. Pakistan Journal of Phytopathology, 28(1 PG-15–17), 15–17.
- Edgar, R. C., & Batzoglou, S. (2006). Multiple sequence alignment. Current Opinion in Structural Biology, 16(3), 368–373.
- Eisenberg, D., Lüthy, R., & Bowie, J. U. (1997). [20] VERIFY3D: Assessment of protein models with three-dimensional profiles (pp. 396–404).
- El-Gremi, S. M., Draz, I. S., & Youssef, W. A.-E. (2017). Biological control of pathogens associated with kernel black point disease of wheat. Crop Protection, 91, 13–19.
- Ferentinos, K. P. (2018). Deep learning models for plant disease detection and diagnosis. Computers and Electronics in Agriculture, 145, 311–318.
- Fernandes, C., Casadevall, A., & Gonçalves, T. (2023). Mechanisms of Alternaria pathogenesis in animals and plants. FEMS Microbiology Reviews, 47(6).
- Fernandez, M. R., & Conner, R. L. (2011). Black Point and Smudge in Wheat Kernel Discolouration in Common and Durum Wheat. 4, 158–164.
- Figueroa, M., Hammond-Kosack, K. E., & Solomon, P. S. (2018). A review of wheat diseases-a field perspective. Molecular Plant Pathology, 19(6), 1523–1536.
- Fisher, M. C., Henk, D. A., Briggs, C. J., Brownstein, J. S., Madoff, L. C., McCraw, S. L., & Gurr, S. J. (2012). Emerging fungal threats to animal, plant and ecosystem health. Nature, 484(7393), 186–194. h.
- Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M. R., Appel, R. D., & Bairoch, A. (2005). Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook (pp. 571–607). Humana Press.
- Gaulton, A., Bellis, L. J., Bento, A. P., Chambers, J., Davies, M., Hersey, A., Light, Y., McGlinchey, S., Michalovich, D., Al-Lazikani, B., & Overington, J. P. (2012). ChEMBL: a large-scale bioactivity database for drug discovery. Nucleic Acids Research, 40(D1), D1100–D1107.
- Griffin, H. G., & Griffin, A. M. (1995). National centre for biotechnology information (NCBI): Services provided for the molecular biotechnologist. Molecular Biotechnology, 4(2), 206–206.
- Guillermo Fuentes-Dávila, Ivón Alejandra Rosas-Jáuregui, Carlos Antonio Ayón-Ibarra, José Luis Félix-Fuentes, Pedro Félix-Valencia, & María Monserrat Torres-Cruz. (2023). Incidence of black point (Alternaria sp.) in elite advanced bread wheat (Triticum aestivum) lines. GSC Biological and Pharmaceutical Sciences, 25(2), 222–229.
- Haque, M. A., Marwaha, S., Deb, C. K., Nigam, S., Arora, A., Hooda, K. S., ... & Agrawal, R. C. (2022). Deep learning-based approach for identification of diseases of maize crop. Scientific reports, 12(1), 6334.
- Hou, Y.-H., Yang, Z.-H., Wang, J.-Z., & Yang, Q.-Z. (2022). Characterization of a thermostable alkaline feruloyl esterase from Alternaria alternata and its synergism in dissolving pulp production. Biochemical Engineering Journal, 187, 108657.
- Ijaz, M., Afzal, A., Ashraf, M. M., Ali, S. R., & ul Haq, M. I. (2018). Integrated Disease Management of Black point in Wheat in Punjab (Pakistan). Plant Protection, 2(2), 63-68.
- Iqbal, M. F., Hussain, M., Ali, M. A., Nawaz, R., & Iqbal, Z. (2014). Efficacy of fungicides used for controlling black point disease in wheat crop. Int. J. Adv. Res. Bio. Sci, 1(6), 59-64.
- Kassaw, A., Mihretie, A., & Ayalew, A. (2021). Rate and Spraying Frequency Determination of Propiconazole Fungicide for the Management of Garlic Rust at Woreilu District, Northeastern Ethiopia. Advances in Agriculture, 2021, 1–9.
- Kamilaris, A., & Prenafeta-Boldú, F. X. (2018b). Deep learning in agriculture: A survey. Computers and Electronics in Agriculture, 147, 70–90.
- Kharin, V. V., Zwiers, F. W., Zhang, X., & Wehner, M. (2013). Changes in temperature and precipitation extremes in the CMIP5 ensemble. Climatic change, 119, 345-357.
- Kustrzeba-Wójcicka, I., Siwak, E., Terlecki, G., Wolańczyk-Mędrala, A., & Mędrala, W. (2014). Alternaria alternata and Its Allergens: a Comprehensive Review. Clinical Reviews in Allergy & Immunology, 47(3), 354–365.
- Laskowski, R. A., MacArthur, M. W., Moss, D. S., & Thornton, J. M. (1993). PROCHECK: a program to check the stereochemical quality of protein structures. Journal of Applied Crystallography, 26(2), 283–291.
- Lee, G. Y., Alzamil, L., Doskenov, B., & Termehchy, A. (2021). A survey on data cleaning methods for improved machine learning model performance. arXiv preprint arXiv:2109.07127.
- Liu, Y., Yang, X., Gan, J., Chen, S., Xiao, Z.-X., & Cao, Y. (2022). CB-Dock2: improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Research, 50(W1), W159–W164.
- Long, M., Hartley, M., Morris, R. J., & Brown, J. K. M. (2023). Classification of wheat diseases using deep learning networks with field and glasshouse images. Plant Pathology, 72(3), 536–547.
- Lore, J. S., & Thind, T. S. (2012). Performance of different fungicides against multiple diseases of rice. Indian Phytopathology.
- Malaker, P. K., & Mian, I. H. (2009). Effect of seed treatment and foliar spray with fungicides in controlling black point disease of wheat. Bangladesh Journal of Agricultural Research, 34(3), 425-434.
- Manamgoda, D. S., Rossman, A. Y., Castlebury, L. A., Chukeatirote, E., & Hyde, K. D. (2015). A taxonomic and phylogenetic re-appraisal of the genus Curvularia (Pleosporaceae): Human and plant pathogens. Phytotaxa, 212(3), 175–198.
- Masiello, M., Somma, S., Susca, A., Ghionna, V., Logrieco, A. F., Franzoni, M., Ravaglia, S., Meca, G., & Moretti, A. (2020). Molecular identification and mycotoxin production by alternaria species occurring on durum wheat, showing black point symptoms. Toxins, 12(4).
- Merck. (2006). Potato Dextrose Agar. Merck Microbiology Manual, 12, 1–2.
- Mohanty, S. P., Hughes, D. P., & Salathé, M. (2016). Using Deep Learning for Image-Based Plant Disease Detection. Frontiers in Plant Science, 7.
- Moshatati, A., & Gharineh, M. H. (2012). Effect of grain weight on germination and seed vigor of wheat. International Journal of Agriculture and Crop Sciences, 4(8), 458–460.
- Muhsin, M., Nawaz, M., Khan, I., Chattha, M. B., Khan, S., Aslam, M. T., Iqbal, M. M., Amin, M. Z., Ayub, M. A., Anwar, U., Hassan, M. U., & Chattha, M. U. (2021). Efficacy of Seed Size to Improve Field Performance of Wheat under Late Sowing Conditions. Pakistan Journal of Agricultural Research, 34(1).
- Naik, B. N., Malmathanraj, R., & Palanisamy, P. (2022). Detection and classification of chilli leaf disease using a squeeze-and-excitation-based CNN model. Ecological Informatics, 69, 101663.
- Nithya, U., Chelladurai, V., Jayas, D. S., & White, N. D. G. (2011). Safe storage guidelines for durum wheat. Journal of Stored Products Research, 47(4), 328–333.
- Peila, D., Picchio, A., Martinelli, D., & Negro, E. D. (2016). Laboratory tests on soil conditioning of clayey soil. Acta Geotechnica, 11(5), 1061–1074.
- Rani, P., & Singh, A. (2018). Effect of Black Point Infection on Germination of different varieties of Wheat Seed. I.
- Prendin, F., Pavan, J., Cappon, G., Del Favero, S., Sparacino, G., & Facchinetti, A. (2023). The importance of interpreting machine learning models for blood glucose prediction in diabetes: an analysis using SHAP. Scientific reports, 13(1), 16865.
- Savary, S., Willocquet, L., Pethybridge, S. J., Esker, P., McRoberts, N., & Nelson, A. (2019). The global burden of pathogens and pests on major food crops. Nature Ecology & Evolution, 3(3), 430–439.
- Schultz, J., Milpetz, F., Bork, P., & Ponting, C. P. (1998). SMART, a simple modular architecture research tool: Identification of signaling domains. Proceedings of the National Academy of Sciences, 95(11), 5857–5864.
- SESİZ, U. (2023). The Screening of Black Point in Commercial Bread Wheat Cultivars Grown in Turkey, and The Effect of Black Point on Thousand Grain Weight. Yüzüncü Yıl Üniversitesi Fen Bilimleri Enstitüsü Dergisi, 28(1), 230–238.
- Shahwani, A. R., Sana, U. B., Shahbaz, K. B., Baber, M., Waseem, B., Hafeez, N. B., Rameez, A. B., Sial, A. H., Sabiel, S. A. I., Kamran, R., Ayaz, A. S., & Ashraf, M. (2014). INFLUENCE OF SEED SIZE ON GERMINABILITY AND GRAIN YIELD OF WHEAT ( Triticum Aestivum L . ) VARIETIES. Journal of Natural Sciences Research, 4(23), 1–11.
- Shukla, D. N., Shrivastava, J. P., & Yadav, M. K. (2020). Occurrence of Black Point Disease Complex of Wheat in Eastern Uttar Pradesh. Www. Groupexcelindia. Com, November.
- Srivastava, J. P., Kushwaha, G. D., & Shukla, D. N. (2015). Black point disease of wheat and its implications on seed quality. Crop Research, 47(1to3), 21-23.
- Abramson, J., Evans, R., Bryant, P., Ingale, A., Johansson-Åkhe, I., Pan, J. et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 629, 289–298 (2024).
- Strange, R. N., & Scott, P. R. (2005). Plant Disease: A Threat to Global Food Security. Annual Review of Phytopathology, 43(1), 83–116.
- Sutaji, D., & Yıldız, O. (2022). LEMOXINET: Lite ensemble MobileNetV2 and Xception models to predict plant disease. Ecological Informatics, 70, 101698.
- Szklarczyk, D., Franceschini, A., Wyder, S., Forslund, K., Heller, D., Huerta-Cepas, J., Simonovic, M., Roth, A., Santos, A., Tsafou, K. P., Kuhn, M., Bork, P., Jensen, L. J., & von Mering, C. (2015). STRING v10: protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Research, 43(D1), D447–D452.
- Talukder, A. S. M. H. M., McDonald, G. K., & Gill, G. S. (2014). Effect of short-term heat stress prior to flowering and early grain set on the grain yield of wheat. Field Crops Research, 160, 54-63.
- Taylor, N. P., & Cunniffe, N. J. (2023). Coupling machine learning and epidemiological modelling to characterise optimal fungicide doses when fungicide resistance is partial or quantitative. Journal of the Royal Society Interface, 20(201), 20220685.
- UniProt: a hub for protein information. (2015). Nucleic Acids Research, 43(D1), D204–D212.
- Wang, Y., Wu, S., Chen, J., Zhang, C., Xu, Z., Li, G., Cai, L., Shen, W., & Wang, Q. (2018). Single and joint toxicity assessment of four currently used pesticides to zebrafish (Danio rerio) using traditional and molecular endpoints. Chemosphere, 192, 14–23.
- Wiederstein, M., & Sippl, M. J. (2007). ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Research, 35(Web Server), W407–W410.
- Xu, K. G., Jiang, Y. M., Li, Y. K., Xu, Q. Q., Niu, J. S., Zhu, X. X., & Li, Q. Y. (2018). Identification and Pathogenicity of Fungal Pathogens Causing Black Point in Wheat on the North China Plain. Indian Journal of Microbiology, 58(2), 159–164.
- Yadav, R. K., Ghasolia, R. P., & Yadav, R. K. (2020). Management of Alternaria alternata of Tomato (Lycopersicon esculentum Mill.) through Plant Extract and Fungicides in vitro and Natural Condition. International Journal of Current Microbiology and Applied Sciences, 9(5), 514–523.
- Zubrod, J. P., Bundschuh, M., Arts, G., Brühl, C. A., Imfeld, G., Knäbel, A., Payraudeau, S., Rasmussen, J. J., Rohr, J., Scharmüller, A., Smalling, K., Stehle, S., Schulz, R., & Schäfer, R. B. (2019). Fungicides: An Overlooked Pesticide Class? Environmental Science & Technology, 53(7), 3347.
Figure 2.
Diagram showing positions of Esterase PHD domain and signal peptide of three feruloyl esterase (AFAE1, AFAE2, AFAE3) proteins.
Figure 2.
Diagram showing positions of Esterase PHD domain and signal peptide of three feruloyl esterase (AFAE1, AFAE2, AFAE3) proteins.
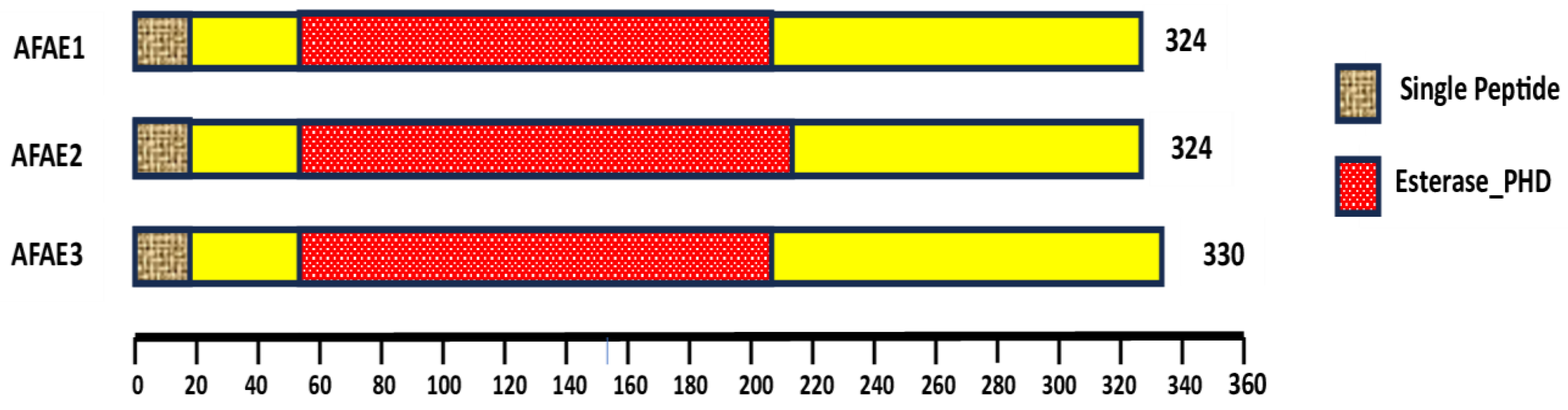
Figure 4.
The pairwise interaction of Temperature, humidity, and precipitation with pesticides.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



