Submitted:
14 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
The Japanese eel endothelial cell-infecting virus (JEECV) and Anguilla herpesvirus (AnHV) are major pathogens in farmed eels. JEECV causes eel viral endothelial cell necrosis (VECNE), while AnHV leads to symptoms such as head erythema and gill necrosis, respectively. Both viruses can result in severe mortality, either independently or in combination, necessitating rapid and early detection of their presence. In this study, we developed a highly efficient duplex quantitative PCR method (R² = 0.999) using TaqMan probes for the rapid and simultaneous detection of AnHV and JEECV. This new diagnostic method demonstrated a 1.7-fold higher detection rate for AnHV and a 2.5-fold higher detection rate for JEECV than those of conventional PCR. Quantitative analysis of water and eel tissue samples from aquaculture facilities revealed that viruses could be detected in water 1–3 months prior to mortality, enabling early identification of infections through water testing alone. Moreover, the method reliably detected low viral loads (<1 copy) in both water and tissue samples, facilitating preclinical detection and proactive disease management. This approach reduces the risk of mass mortality and economic losses in eel farming. These findings underscore the critical role of advanced molecular diagnostic technologies in enhancing health management in aquaculture.
Keywords:
JEECV
; AnHV
; qPCR
; eels
; aquaculture diagnostics
1. Introduction
Eels are among the most prominent species in the inland aquaculture industry of Korea. Between 2012 and 2021, eel production increased by approximately 3.6-fold, reaching 15,764 tons in 2021, which accounted for approximately 47% of the total inland aquaculture output [1]. In Korea, most eels are reared in recirculating aquaculture systems (RAS), where water temperatures are maintained between 27 and 32°C year-round, even during winter. Although this stable environment optimizes eel growth, it poses significant risks. Specifically, in the event of a disease outbreak, the shared filtration systems in RAS can facilitate rapid disease transmission across tanks, potentially leading to mass mortality and substantial economic losses. Therefore, early detection and rapid response to disease outbreaks are critical to mitigating these risks.
Recent studies have reported mass mortality in key cultured eel species in Korea, including Anguilla japonica, A. bicolor, and A. marmorata, due to infections by two DNA viruses: Anguillid herpesvirus-1 (AnHV) and Japanese eel endothelial cell-infecting virus (JEECV) [2,3,4,5]. JEECV, the causative agent of viral endothelial cell necrosis of eel (VECNE), is characterized by symptoms such as head reddening, pectoral fin necrosis, and gill congestion [4,6,7]. AnHV, first isolated in A. japonica and A. anguilla in Japan in 1985 [8], has been associated with disease outbreaks in farmed eels in Japan, Taiwan, and Korea. Infected eels typically exhibit symptoms such as snout reddening, skin hemorrhages, and necrosis of the skin and gills [2,3,5,9,10,11,12]. Although these two viruses can infect eels independently, they often occur in conjunction with bacterial co-infections. They can also present as dual infections involving both AnHV and JEECV, resulting in more severe disease and elevated mortality rates [2]. Notably, a recent survey of eel farms detected JEECV in 49% of cases, AnHV in 27%, and co-infections in 17%, highlighting the critical prevalence of these viral pathogens in aquaculture [2]. Therefore, the rapid and early detection of AnHV and JEECV is essential for effective disease prevention and management in eel farming. Moreover, developing highly sensitive diagnostic tools is crucial to enabling timely identification and response to infections.
In the present study, we aimed to establish a duplex quantitative PCR (qPCR) method utilizing TaqMan probes for the simultaneous detection of AnHV and JEECV. We validated the efficacy of this diagnostic approach through quantitative analyses of viral presence in water and eel tissue samples from aquaculture farms. This study demonstrates the potential of this method for early detection and improved disease control in eel farming operations.
2. Results
2.1. Phylogenetic Relationships of Target Genes
Primers designed for AnHV (Table 1) were utilized for nested PCR amplification, confirming its presence in 13 out of 28 eel samples. Sequencing analysis of the amplified products revealed that all AnHV-positive samples were successfully sequenced, with total lengths ranging from 825–846 bp. Variations in the length of these sequences were attributed to the insertion of four codons and the deletion of three codons compared to the reference DNA polymerase catalytic subunit (Pol) gene of AnHV (AnHV-1 Pol; GenBank accession number: KX027736). A phylogenetic tree constructed using the neighbor-joining method (Figure 1) revealed two distinct genotypes, type 1 and type 2, within AnHV. Notably, all AnHV-positive samples with the type 2 genotype exhibited no codon deletions.
Similarly, primers designed for JEECV (Table 1) were used for nested PCR amplification, identifying four JEECV-positive samples. Sequencing analysis revealed uniform sequence lengths of 723 bp across all samples, with 100% identity and no observed genetic variation. These sequences were aligned with the reference polyomavirus large T-antigen-like protein of JEECV (JEECV Ltlg) from the NCBI GenBank database, and a phylogenetic tree was constructed using the neighbor-joining method (Figure 2). The analysis confirmed the absence of genetic diversity among the JEECV-positive samples in this study.
2.2. Optimization of Duplex qPCR
To optimize duplex qPCR conditions for detecting AnHV and JEECV, the newly designed primers and TaqMan probes (Table 2) were tested. Primer and probe concentrations were evaluated in the range of 5–10 pmol/reaction, and annealing/extension temperatures were tested at 62 and 63°C. At 62°C, weak nonspecific signals were observed in the negative control, whereas no nonspecific signals occurred at 63°C. These nonspecific signals were enhanced with increasing primer and probe concentrations; therefore, the optimal concentration was determined to be 5 pmol/reaction. Consequently, the optimal duplex qPCR conditions were set at an annealing/extension temperature of 63°C with primer and probe concentrations of 5 pmol/reaction.
2.3. Standard Curve
To establish internal standards for the target gene regions—Pol for AnHV and Ltlg for JEECV—PCR amplification products were cloned into plasmid DNA using TA cloning for large-scale replication. The plasmid DNA was standardized to 100,000 copies based on molecular weight and concentration. Serial 10-fold dilutions were then prepared to generate standard materials with concentrations ranging from 100,000 copies/reaction to 1 copy/reaction. Standard curves were constructed for each virus, demonstrating a strong inverse relationship between Ct values and standard material concentration, with an R² value of 0.999, indicating high reliability. Nonspecific signals were not detected in the negative control (Figure 3).
2.4. Sensitivity of the qPCR Marker
The sensitivity of the duplex qPCR method, developed to detect AnHV and JEECV, was compared using conventional PCR (cPCR) on 28 eel samples (Table 3). For AnHV, cPCR identified positive bands in 7 out of 28 samples. In comparison, the duplex qPCR method detected 12 positive samples, representing a 1.7-fold increase in sensitivity. All cPCR-positive samples were also identified as positive by qPCR, with viral loads ranging from 167.6 to 76,209.1 copies. Notably, samples that were identified as negative by cPCR but positive by qPCR had viral loads ranging from 0.2 to 218 copies. For JEECV, cPCR failed to detect any positive samples in the initial PCR step, and only 4 samples were confirmed positive using nested PCR. In contrast, the duplex qPCR method identified 10 positive samples, achieving a 2.5-fold higher detection rate than that of nested PCR. Viral loads in qPCR-positive samples ranged from 0.1 to 2,104 copies.
2.5. Spike Tests on Culture Water
JEECV was successfully detected in groundwater samples (1, 2, 4, and 10 L) spiked with 4 mg/L of JEECV-infected tissue homogenate. The viral load increased proportionally with the volume of concentrated groundwater (Table 4). Specifically, the viral load in 2 L of concentrated groundwater was approximately 3.6 times higher (44,250 copies) than that in 1 L (12,367 copies). It also increased by 14-fold (173,072 copies) and nearly 30-fold (375,871 copies) in samples with volumes of 4 and 10 L, respectively. In contrast, no virus was detected in the corresponding negative control groundwater samples, regardless of volume.
2.6. Validation of the QPCR Diagnostic Method for Monitoring Viral Outbreaks in Eel Farms
Quantitative analysis of viral DNA from environmental DNA (eDNA) and eel tissue samples (gills, fins, kidneys, and spleen) collected from two eel farms is summarized in Table 5. In Farm A, no viruses were detected in either the eDNA or eel tissues during the first survey. During the second survey, JEECV was detected in the eDNA at a concentration of 1.0 × 10¹ copies/mL. By the third survey, JEECV was detected in all eel tissues and eDNA, with the highest concentration observed in the gills at 4.0 × 10³ copies/µL. Mortalities were also reported during this survey. In Farm B, AnHV was detected in the eDNA during the first survey; however, its concentration was below the quantification limit (<1 copy/mL), preventing precise quantification. In the second survey, no virus was detected in the eDNA, whereas AnHV was identified in all eel tissues, particularly in the fins and gills of all three sampled eels, with additional detections in some kidney and spleen samples. During the third survey, no virus was detected, although the eels appeared weak. In the fourth survey, both eDNA and eel tissue samples tested positive for the virus, and mortality events were recorded.
3. Discussion
In a previous study, we investigated the infection status of AnHV and JEECV in eel farms across Korea [2], confirming that these viral diseases are widespread in cultured eels, with frequent cases of co-infection. Such mixed infections, particularly during the juvenile stage, often led to mass mortality, resulting in significant economic losses. These findings emphasize the need for monitoring both viruses to improve disease surveillance in Korean eel farms. However, the available PCR methods [7,13] require individual detection of each virus, making the process labor-intensive and time-consuming. To address these limitations, we developed a highly sensitive duplex qPCR method capable of detecting both viruses simultaneously in the present study.
We conducted DNA sequencing of PCR-positive samples to design primers specific to JEECV and AnHV. While the sequencing results revealed no genetic variation in JEECV, two types of AnHV (AnHV-1 and AnHV-2) were detected, independent of host species or geographic region. AnHV-1 has been documented in various eel species, including A. anguilla, A. japonica, A. bicolor, A. marmorata, and A. rostrata, across Europe, East Asia, and Southeast Asia [5,14,15,16,17,18], with full genome sequences reported for strains from A. anguilla and A. rostrata [19,20]. AnHV-2, first isolated from A. japonica, has been partially sequenced, with the DNA polymerase gene (GenBank accession: LC092221) serving as a reference. While Kim et al. recently developed a duplex PCR method for detecting AnHV-1 [21], in our study, we designed primers and probes targeting conserved regions of the Pol gene shared by both AnHV-1 and AnHV-2. This approach enabled us to simultaneously detect both AnHV types. Moreover, standard curve analysis using plasmid DNA demonstrated high reliability (R² = 0.999) in detecting both viruses down to a single copy, confirming the exceptional sensitivity of our method for detecting trace viral loads. In addition, compared to cPCR, the duplex qPCR method developed in the present study was 1.7 times more sensitive for AnHV and 2.5 times more sensitive for JEECV. The ability of our method to detect low viral loads undetectable by cPCR significantly enhances the accuracy of disease monitoring. This heightened sensitivity enables the detection of viral infections at preclinical stages, prior to the onset of symptoms, thereby facilitating early diagnosis and intervention for AnHV and JEECV in farmed eels.
Using this diagnostic tool, we conducted a six-month monthly monitoring program at two eel farms and analyzed samples from rearing water and fish. We observed no cases of co-infection during the monitoring period; however, we detected AnHV and JEECV individually. At Farm A, initial investigations revealed no detectable viral DNA in either eel tissues or eDNA, suggesting either pre-infection conditions or a viral load too low to detect. In subsequent investigations, JEECV was detected in eDNA at a concentration of 1.0 × 10¹ copies/mL, indicating initial viral introduction. By the third investigation, JEECV was detected in all eel tissues, coinciding with an increase in eel mortality. The highest viral concentration (4.0 × 10³ copies/μL) was observed in the gills, suggesting that this tissue is the primary entry point and reservoir for infection. At Farm B, AnHV was initially detected in eDNA from rearing water at concentrations below the limit of quantification (<1 copy/mL), indicative of latent or low-level infection. In the second survey, the virus was undetectable in eDNA but was present in all eel tissues, highlighting the importance of tissue sampling for identifying subclinical infections that may be missed by water analysis alone. During the third survey, the virus was not detected, despite deterioration in eel health. By the fourth survey, AnHV was detected in all samples, and mortality rates had increased, marking the onset of an outbreak. These findings emphasize the necessity of continuous monitoring and early intervention to prevent mass mortality from viral infections.
The present study suggests that an integrated sampling approach, combining rearing water eDNA analysis with fish tissue examination, may provide a more accurate prediction of viral disease outbreaks than an individual approach. However, repeated sampling of live fish can impose significant stress on the stock. Therefore, continuous monitoring of rearing water eDNA using the high-sensitivity detection technology developed in this study offers an efficient and non-invasive alternative for disease surveillance and control. Moreover, when the timing of virus detection in stock water was compared to the onset of eel mortality, viruses were consistently detected in eDNA 3 weeks to 3 months before mortalities were observed (Table 5). This finding demonstrates that monitoring eDNA in rearing water alone is sufficient to predict disease outbreaks. Similarly, recent studies have highlighted the potential of eDNA methods for detecting fish viruses [22,23,24,25]. Notably, DNA viruses are more stable than RNA viruses, making them particularly suitable for eDNA-based monitoring [26]. Given that both AnHV and JEECV are DNA viruses, they are ideal candidates for detection using this method in aquaculture settings.
In conclusion, we developed a highly sensitive diagnostic technology capable of simultaneously detecting AnHV and JEECV, which are major causes of mortality in eel farms. The method is specifically designed to detect both types 1 and 2 of AnHV. With a sensitivity of detecting as little as one plasmid DNA copy, this technology supports non-destructive testing using fin or gill samples, avoiding the need to dissect internal organs. Furthermore, we confirmed that virus monitoring is feasible using eDNA from rearing water. Notably, eDNA-based virus monitoring eliminates the need for complex and invasive fish sampling procedures, enabling early detection of infections and prompt intervention to prevent disease spread. Our results also indicate that virus detection can be achieved with only 1 L of culture water, although elevated concentrations of culture water enhance detection sensitivity. These findings highlight eDNA-based monitoring as a practical tool for early diagnosis and outbreak prediction. This diagnostic technology has broad applications in viral disease monitoring, vaccine development, and treatment research, offering a significant contribution to advancing health management practices in eel farming.
4. Materials and Methods
4.1. Fish Sampling
Twenty-eight diseased eels were collected from aquaculture farms. Viral DNA was extracted from pooled spleen and kidney samples using the DNA Isolation Kit for Cells and Tissues (Roche, Basel, Switzerland), following the manufacturer’s instructions. DNA concentration and purity were assessed with a NanoVue™ Plus Spectrophotometer (GE Healthcare, Chicago, IL, USA). The DNA was diluted to 20 ng/μL, stored at -20°C, and used for sequencing target genes and verifying the qPCR markers developed in this study.
4.2. Sequencing of Target Genes
Analysis of the GenBank database revealed that the Pol gene of AnHV and the Ltlg gene of JEECV had the most abundant sequence information. Using these regions from AnHV (GenBank accession number: KX027736) and JEECV (NC_015123) as reference sequences, new PCR primers were designed (Table 1). PCR amplification was performed using these primers on DNA from the 28 eel specimens. PCR-positive samples were sequenced, and their Pol and Ltlg sequences were analyzed. The newly sequenced genes were aligned with corresponding sequences registered in GenBank. Molecular phylogenetic trees were constructed using the neighbor-joining method to establish phylogenetic relationships.
4.3. Design and Verification of QPCR Markers
The DNA sequences of AnHV registered in GenBank and the newly sequenced 13 AnHV-positive samples were aligned to identify a conserved region present in both AnHV-1 and AnHV-2. This conserved sequence was used to design highly specific qPCR primers and hydrolysis probes. The AnHV probe employed Cy5 as the fluorescent reporter dye and BHQ2 as the quencher (Table 2). The optimal annealing/extension temperature for the AnHV qPCR reaction was determined by testing temperatures in 2°C increments between 60°C and 70°C to identify the condition that yielded the highest amplification efficiency.
Similarly, the DNA sequences of JEECV in GenBank and the newly sequenced 4 JEECV-positive samples were aligned to identify a conserved region. Specific qPCR primers and hydrolysis probes were designed for JEECV detection, using HEX as the fluorescent reporter dye and BHQ1 as the quencher (Table 2). The annealing/extension temperature for JEECV qPCR amplification was optimized using the same temperature range (60–70°C) to achieve maximum amplification efficiency.
4.4. Optimization of QPCR Conditions
To optimize the duplex qPCR conditions for detecting AnHV and JEECV, primer concentrations were tested in the range of 5–10 pmol/reaction, and hydrolysis probe concentrations were tested in the same range. The annealing/extension temperatures were set at 62 and 63°C for final verification.
4.5. Standard Curves
Internal standards were established for the target genes (Pol for AnHV and Ltlg for JEECV). DNA fragments containing the forward and reverse primers, as well as hydrolysis probe binding sites, were synthesized and inserted into plasmids for mass replication (Bioneer, Daejeon, Republic of Korea). The plasmid DNA was quantified, and copy numbers were standardized to 100,000. Serial 10-fold dilutions were performed to establish standard materials with concentrations ranging from 100,000 copies/reaction to 1 copy/reaction. Calibration curves were generated for each virus to enable precise quantification of the target genes.
4.6. Sensitivity Tests
To evaluate the detection sensitivity of the diagnostic method developed in this study compared to cPCR, both cPCR and qPCR analyses were performed on 28 eel tissue samples, along with two no-template controls. All DNA samples were diluted to a concentration of 20 μg/μl, and 2 μl (40 μg) of each sample was used for analysis. For cPCR, the primers newly designed in this study for detecting AnHV or JEECV were used (Table 1). For JEECV detection, as the virus was not identified in the first round of PCR, nested PCR was performed to enhance sensitivity.
4.7. Spike Tests Using Culture Water
To simulate virus detection in aquaculture water, 350 µLof eel tissue homogenate was mixed thoroughly with 20 L of groundwater used in eel farming. The spiked groundwater was divided into portions of 1, 2, 4, and 10 L and filtered through cellulose nitrate membranes (50 mm diameter, 0.45 μm pore size; Whatman®, Maidstone, UK). Viral DNA was then extracted using the DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany), following the complete destruction of the membranes. This destruction was achieved by adding three stainless steel beads (2.4 mm diameter; OMNI International, Kennesaw, GA, USA) and processing the sample with a Bead Ruptor Elite homogenizer (OMNI International).
4.8. Validation of the QPCR Diagnostic Method for Monitoring Viral Outbreaks in Eel Farms
The qPCR diagnostic method developed in this study was validated by monitoring virus occurrence in eel tissues and culture water from two eel farms monthly. One liter of rearing water was collected from aquaculture tanks and transported to the laboratory. Suspended solids were removed, and the water was filtered through a nitrocellulose membrane filter (0.45 μm pore size). Subsequently, eDNA was extracted from the membrane filter using the DNeasy Blood & Tissue Kit (QIAGEN). To monitor virus presence in eel tissues, three eels were sampled from the same tanks where water was collected. Tissues were collected from the pectoral fins, gills, kidneys, and spleen. Genomic DNA was extracted from each tissue using the DNA Isolation Kit for Cells and Tissues (Roche) and eluted in 50 μL of TE buffer (10 mM Tris-HCl, pH 8.0; 1 mM EDTA, pH 8.0). Subsequently, DNA concentration and purity were determined using a spectrophotometer. The extracted eDNA from water samples and genomic DNA from tissue samples were used as templates for quantifying viral gene copy numbers using the qPCR method.
Author Contributions
Conceptualization, J-Y. S. and A. K.; methodology K-Y. K. and J-Y. S.; writing-original draft preparation, J-Y. S. and K-Y. K.; supervision, A. K.; project administration, A. K.; funding acquisition, A. K. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| eDNA | Environmental DNA |
| qPCR | Quantitative PCR |
| JEECV | Japanese eel endothelial cell-infecting virus |
| AnHV | Anguilla herpesvirus |
| VECNE | Eel viral endothelial cell necrosis |
| RAS | Recirculating aquaculture systems |
| Pol | DNA polymerase catalytic subunit |
| Ltlg | Polyomavirus large T-antigen-like protein of JEECV |
References
- Ministry of Oceans and Fisheries Statistics System; https://www.mof.go.kr/statPortal/search/searchList.do, accessed May 2, 2023.
- Jang, M.H.; Lee, N.; Cho, M.; Song, J. Status and characteristics of JEECV (Japanese eel endothelial cell-infecting virus) and AnHV (anguillid herpesvirus 1) infections in domestic farmed eels Anguilla japonica, Anguilla bicolor and Anguilla marmorata. J. Fish. Aquat. Sci.; Kor 2021, 54, 668–675.
- Kim, H. J.; Yu, J. H.; Kim, D. W.; Kwon, S. R.; Park, S. W. Molecular evidence of anguillid herpesvirus-1 (AngHV-1) in the farmed japanese eel, Anguilla japonica Temminck & schlegel, in korea. J. Fish Dis., Kor 2012, 35(4).
- Kim, S.M.; Ko, S.M.; Jin, J.H.; Seo, J.S.; Lee, N.S.; Kim, Y.S.; Gu, J.H.; Bae, Y.R. Characteristics of viral endothelial cell necrosis of eel (VECNE) from culturing eel (Anguilla japonica, Anguilla bicolor) in Korea. J. Ichthyol.; Kor 2018, 30, 185–193.
- Park, S.; Jung, E.; Kim, D. Outbreak of anguillid herpesvirus-1 (AngHV-1) infection in cultured shortfin eel (Anguilla bicolor) in Korea. J. Fish Pathol. 2012, 25, 151–158. [Google Scholar] [CrossRef]
- Egusa, S.; Tanaka, M.; Ogami, H.; Oka, H. Histopathological observations on an intense congestion of the gills in cultured Japanese eel Anguilla japonica. Fish Pathol.; Kor 1989, 24, 51-56.
- Mizutani, T.; Sayama, Y.; Nakanishi, A.; Ochiai, H.; Sakai, K.; Wakabayashi, K.; Tanaka, N.; Miura, E.; Oba, M.; Kurane, I.; et al. Novel DNA virus isolated from samples showing endothelial cell necrosis in the Japanese eel, Anguilla japonica. Virology 2011, 412, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Fukuda, H.; Sano, T. Isolation and characterization of a new herpesvirus from eel. Pathol. Mar. Sci. 1990, 15, 31. [Google Scholar]
- Chang, P.H.; Pan, Y.H.; Wu, C.M.; Kuo, S.T.; Chung, H.Y. Isolation and molecular characterization of herpesvirus from cultured European eels Anguilla anguilla in Taiwan. Dis. Aquat. Organ. 2002, 50, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Miyazaki, T. Characterization and pathogenicity of a herpesvirus isolated from cutaneous lesion in Japanese eel, Anguilla japonica. Fish Pathol. 1997, 32, 89–95. [Google Scholar] [CrossRef]
- Lee, *!!! REPLACE !!!*; et al. , 1999 Lee N.S.; Kobayashi J.; Miyazaki T. Gill filament necrosis in farmed Japanese eels, Anguilla japonica (Temminck & Schlegel), infected with herpesvirus anguillae. J. Fish Dis. 1999, 22, 457–463. [Google Scholar]
- Ueno, Y.; Shi, J.W.; Yoshida, T.; Kitao, T.; Sakai, M.; Chen, S.N.; Kou, G.H. Biological and serological comparisons of eel herpesvirus in Formosa (EHVF) and herpesvirus anguillae (HVA). J. Appl. Ichthyol. 1996, 12, 49–51. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Pritz-Verschuren, S.; Kerkhoff, S.; Botter, A.; Willemsen, M.; van Nieuwstadt, T.; Haenen, O. Development of a polymerase chain reaction for the detection of anguillid herpesvirus DNA in eels based on the herpesvirus DNA polymerase gene. J. Virol. Method. 2005, 124, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Davidse, A.; Haenen, O.L.M.; Dijkstra, S.G.; van Nieuwstadt, A. P.; van der Vorst, T. J. K.; Wagenaar, F.; Wellenberg, G. J. First isolation of herpesvirus of eel (Herpesvirus anguillae) in diseased European eel (Anguilla anguilla L.) in Europe. In: Bulletin of the European Association of Fish Pathologists. 1999, 19(4), 137-141.
- Guo, R.; Zhang, Z.; He, T.; Li, M.; Zhuo, Y.; Yang, X.; Fan, H.; Chen, X. Isolation and identification of a new isolate of anguillid herpesvirus 1 from farmed American eels (Anguilla rostrata) in China. Viruses 2022, 14, 2722. [Google Scholar] [CrossRef] [PubMed]
- Panicz, R.; Eljasik, P.; Nguyen, T.T.; Vo Thi, K.T.; Hoang, D.V. First detection of herpesvirus anguillae (AngHV-1) associated with mortalities in farmed giant mottled eel (Anguilla marmorata) in Vietnam. J. Fish Dis. 2021, 44, 847–852. [Google Scholar] [CrossRef] [PubMed]
- van Beurden, S.J.; Engelsma, M.Y.; Roozenburg, I.; Voorbergen-Laarman, M.A.; van Tulden, P.W.; Kerkhoff, S.; van Nieuwstadt, A.P.; Davidse, A.; Haenen, O.L.M. Viral diseases of wild and farmed European eel Anguilla anguilla with particular reference to the Netherlands. Dis. Aquat. Organ. 2012, 101, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.M.; Liu, P.C.; Nan, F.H. Complete Genome Sequence of Herpesvirus anguillae Strain HVA980811 Isolated in Chiayi, Taiwan. Genome Announc. 2017, 5, 01677–01616. [Google Scholar] [CrossRef] [PubMed]
- Kole, S.; Kim, H.J.; Jung, S.J. Complete genome sequence of anguillid herpesvirus 1 isolated from imported Anguilla rostrata (American eel) from Canada. Microbiol. Resour. Announc. 2022, 11, e0082922. [Google Scholar] [CrossRef] [PubMed]
- van Beurden, S.J.; Bossers, A.; Voorbergen-Laarman, M.H.A.; Haenen, O.L.M.; Peters, S.; Abma-Henkens, M.H.C.; Peeters, B.P.H.; Rottier, P.J.M.; Engelsma, M.Y. Complete genome sequence and taxonomic position of anguillid herpesvirus 1. J. Gen. Virol. 2010, 91, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Bathige, S.D.N.K.; Jeon, H.B.; Kim, S.H.; Yu, C.; Kang, W.; Noh, K.; Lee, E.; Lim, B.S.; Shin, D.H. Development of a duplex qPCR assay for the detection of coinfection in eels by Japanese eel endothelial cell-infecting virus and anguillid herpesvirus-1. Aquaculture 2024, 578, 740091. [Google Scholar] [CrossRef]
- Haramoto, E.; Katayama, H.; Oguma, K.; Ohgaki, S. Recovery of naked viral genomes in water by virus concentration methods. J. Virol. Method. 2007, 142, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Haramoto, E.; Kitajima, M.; Katayama, H.; Ohgaki, S. Detection of koi herpesvirus DNA in river water in Japan. J. Fish Dis. 2007, 30, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Taengphu, S.; Kayansamruaj, P.; Kawato, Y.; Delamare-Deboutteville, J.; Mohan, C.V.; Dong, H.T.; Senapin, S. Concentration and quantification of tilapia tilapinevirus from water using a simple iron flocculation coupled with probe-based RT-qPCR. PeerJ 2022, 10, e13157. [Google Scholar] [CrossRef] [PubMed]
- Weli, S.C.; Bernhardt, L.V.; Qviller, L.; Myrmel, M.; Lillehaug, A. Development and evaluation of a method for concentration and detection of salmonid alphavirus from seawater. J. Virol. Method. 2021, 287, 113990. [Google Scholar] [CrossRef] [PubMed]
- Bohara, K.; Yadav, A.K.; Joshi, P. Detection of fish pathogens in freshwater aquaculture using eDNA methods. Diversity 2022, 14, 1015. [Google Scholar] [CrossRef]
Figure 1.
Neighbor-Joining phylogenetic tree constructed based on the DNA sequences of the AnHV Pol gene region from 13 AnHV-positive samples.
Figure 1.
Neighbor-Joining phylogenetic tree constructed based on the DNA sequences of the AnHV Pol gene region from 13 AnHV-positive samples.
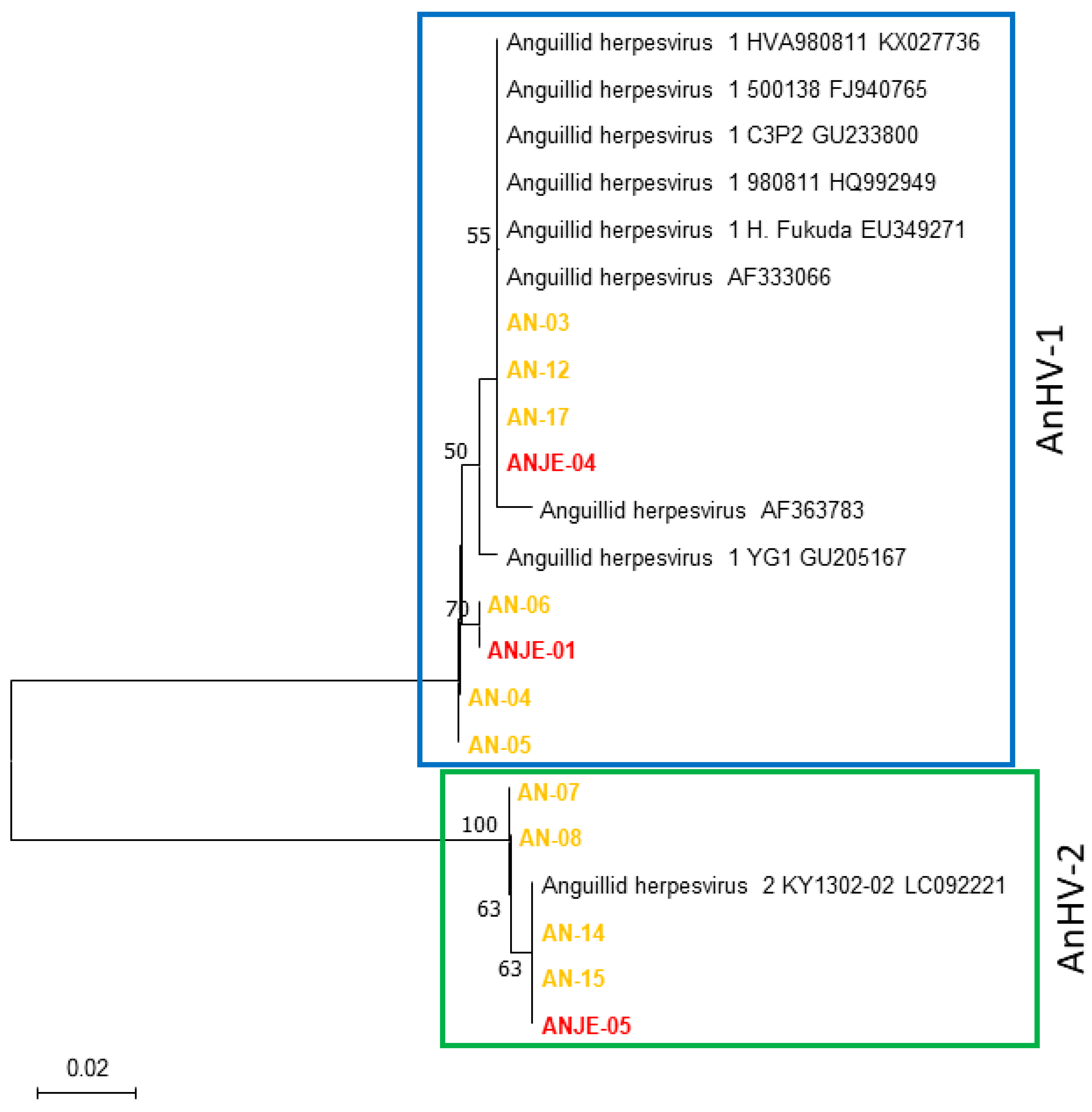
Figure 2.
Neighbor-Joining phylogenetic tree constructed based on the DNA sequences of the JEECV Ltlg gene region from four JEECV-positive samples.
Figure 2.
Neighbor-Joining phylogenetic tree constructed based on the DNA sequences of the JEECV Ltlg gene region from four JEECV-positive samples.
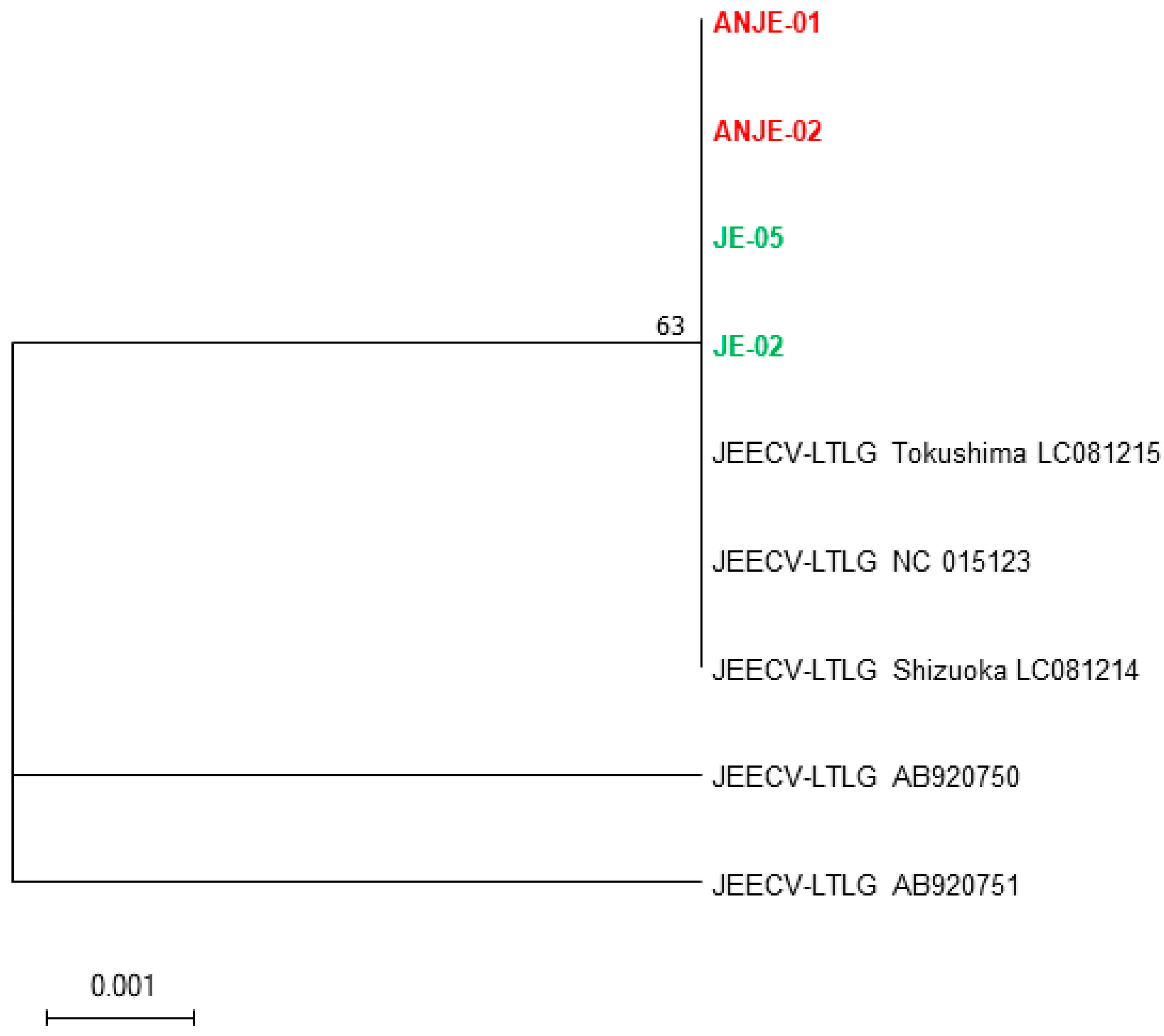
Figure 3.
Duplex qPCR amplification curves and standard curves for AnHV and JEECV at internal standard concentrations (100,000 copies/rxn, 10,000 copies/rxn, 1,000 copies/rxn, 100 copies/rxn, 10 copies/rxn, 1 copy/rxn), with an r2 value of 0.999.
Figure 3.
Duplex qPCR amplification curves and standard curves for AnHV and JEECV at internal standard concentrations (100,000 copies/rxn, 10,000 copies/rxn, 1,000 copies/rxn, 100 copies/rxn, 10 copies/rxn, 1 copy/rxn), with an r2 value of 0.999.
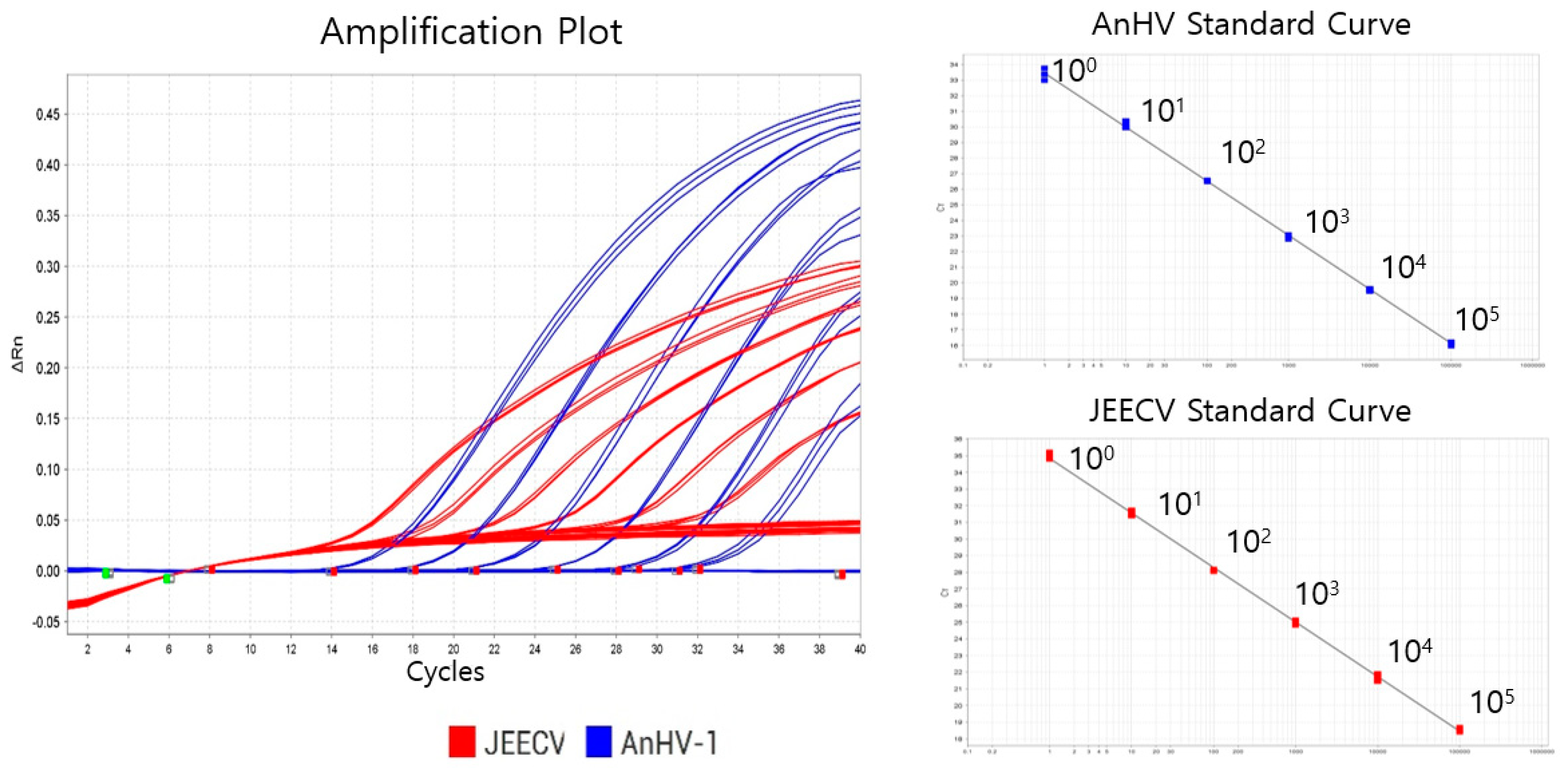

Table 1.
Primer sequence designed for AnHV and JEECV target gene sequences.
| Virus | Target gene | PCR | Primer name | Sequence (5'→3') | Reference |
|---|---|---|---|---|---|
| AnHV | Pol gene | 1st | 2061f | CAAGCTGAAGGAGAACAGAT | This study |
| 3086r | GTTCGTGATCAGAGAGTTGT | This study | |||
| Nested | 2108f | CCATGTGCTTGACTGAAAAG | This study | ||
| 3016r | TCCTCAAAGAACCACGCTTT | This study | |||
| JEECV | Ltlg gene | 1st | 0343f | TTTGTGCAGACTGTACTGGT | This study |
| 1455r | TACTCATTCATAGTGGCAATC | modified from Mizutani et al., 2011[7] | |||
| Nested | 0462f | AACAGGCATAGCAGTTGTGC | This study | ||
| 1219r | TCACCGTTCATGTTTAGGAC | modified from Mizutani et al., 2011[7] |
Table 2.
Duplex qPCR Markers for AnHV and JEECV detection designed in the present study.
| Target virus | Primers and Probe | Sequence (5'→3') |
|---|---|---|
| AnHV | ANHV-Pol-2173f | AAGGTGTTTCAGCCYACCAT |
| ANHV-Pol-2587r | ATGAAGATCT GCGCCAACTC | |
| ANHV-Pol-2260p | Cy5-AGCAACATGTGCGACGCCAA-BHQ2 | |
| JEECV | JEECV-LTLG-0546f | CAATGTGATGCAGGTAGCAA |
| JEECV-LTLG-0777r | TCTGTTGGTCGCTTCGACAT | |
| JEECV-LTLG-0662p | HEX-TGGGCTTTGACTACACGATGCT-BHQ1 |
Table 3.
Comparison of diagnostic performance between the newly developed qPCR method and conventional PCR using eel tissued.
Table 3.
Comparison of diagnostic performance between the newly developed qPCR method and conventional PCR using eel tissued.
| No. | Host | Fish size | AnHV | JEECV | ||||
|---|---|---|---|---|---|---|---|---|
| cPCR | qPCR (copies/ml) | cPCR1 | qPCR (copies/ml) | |||||
| 1 | A. japonica | 50.5 | ND | ND | ND | ND | ||
| 2 | A. japonica | 48.2 | ND | ND | ND | 1.1 | ||
| 3 | A. japonica | 42.3 | Positive | 2308.9 | ND | 2.9 | ||
| 4 | A. marmorata | 50.9 | ND | ND | ND | ND | ||
| 5 | A. marmorata | 11.6 | Positive | 76209.1 | ND | ND | ||
| 6 | A. bicolor | No data | Positive | 711.7 | ND | 0.1 | ||
| 7 | A. japonica | No data | ND | ND | ND | 2104.5 | ||
| 8 | A. japonica | 56.7 | ND | 11 | ND | 0.8 | ||
| 9 | A. australis | 14.5 | ND | ND | ND | ND | ||
| 10 | A. australis | 12.6 | ND | ND | ND | ND | ||
| 11 | A. australis | 55.8 | ND | ND | ND | ND | ||
| 12 | A. japonica | 53 | Positive | 1668.7 | ND | ND | ||
| 13 | A. japonica | 24.6 | ND | 0.6 | ND | 0.5 | ||
| 14 | A. japonica | 47 | ND | 28 | ND | ND | ||
| 15 | A. japonica | 43.5 | ND | 0.3 | ND | ND | ||
| 16 | A. bicolor | 39.5 | ND | ND | ND | ND | ||
| 17 | A. japonica | 46.5 | Positive | 167.6 | ND | ND | ||
| 18 | A. australis | 13.9 | ND | 0.2 | ND | ND | ||
| 19 | A. japonica | 59.5 | ND | ND | ND | ND | ||
| 20 | A. marmorata | 52.8 | ND | ND | Positive | ND | ||
| 21 | A. bicolor | 42.2 | ND | ND | ND | ND | ||
| 22 | A. australis | 48 | ND | ND | ND | ND | ||
| 23 | A. japonica | 53.4 | ND | ND | Positive | 2.5 | ||
| 24 | A. bicolor | No data | Positive | 464.9 | Positive | 0.3 | ||
| 25 | A. japonica | No data | ND | ND | Positive | 1182.9 | ||
| 26 | A. australis | 55.8 | ND | ND | ND | ND | ||
| 27 | A. japonica | 53 | Positive | 218.5 | ND | ND | ||
| 28 | A. japonica | 24.6 | ND | 0.2 | ND | 0.3 | ||
| 29 | NTC-1 | ND | ND | ND | ND | |||
| 30 | NTC-2 | ND | ND | ND | ND | |||
1 , nested PCR result of JEECV.
Table 4.
Results of the spike test using JEECV-positive eel tissue homogenate in groundwater.
| Sample Name | Sample volume (liters) | AnHV | JEECV | ||
|---|---|---|---|---|---|
| Cq value | Copy no. | Cq value | Copy no. | ||
| Ground water | 1 | ND | - | ND | - |
| Ground water | 2 | ND | - | ND | - |
| Ground water | 4 | ND | - | ND | - |
| Ground water | 10 | ND | - | ND | - |
| Spiked ground water | 1 | ND | - | 21.424 | 12,367 |
| Spiked ground water | 2 | ND | - | 19.606 | 44,250 |
| Spiked ground water | 4 | ND | - | 17.662 | 173,072 |
| Spiked ground water | 10 | ND | - | 16.556 | 375,871 |
Table 5.
Results of virus detection in eel farms (eel tissues and eDNA) by the diplex qPCR method developed in this study.
Table 5.
Results of virus detection in eel farms (eel tissues and eDNA) by the diplex qPCR method developed in this study.
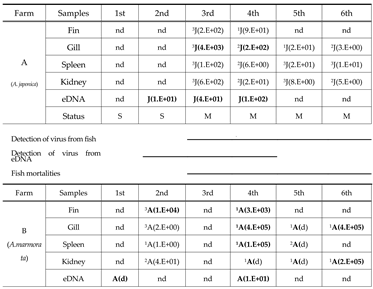
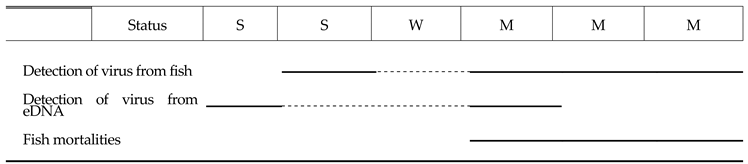
nd, not detected; d, detected (Cq value ≥ 35); J, JEECV; A, AnHV. Fish condition was stable (S), weakend (W) and mortality (M); 1-3, number of the fish detected with virus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



