
Submitted:
13 January 2025
Posted:
14 January 2025
You are already at the latest version
Abstract
Human pharmaceutical residues are increasingly recognized as environmental contaminants that pose significant risks to aquatic ecosystems. This review focuses on the oxidative stress induced in fish by these pharmaceutical residues. Pharmaceuticals, including antidepressants, anti-inflammatories, and other widely used medications, enter aquatic ecosystems through multiple pathways, most notably through their use in human and veterinary medicine, the subsequent metabolic excretion via urine and feces, and wastewater treatment plant effluents. These substances, often only partially removed during water treatment, persist in the environment and pose potential risks to aquatic organisms, impacting water quality and ecosystem health. Upon exposure, fish exhibit oxidative stress responses characterized by the overproduction of reactive oxygen species (ROS) and subsequent damage to cellular components such as lipids, proteins, DNA, and RNA. This review summarizes current research on the oxidative stress (OS) responses of various fish species to environmental concentrations of pharmaceuticals. It explores the mechanisms underlying OS, identifies the specific pharmaceuticals involved, and discusses these responses' physiological and ecological implications.
Keywords:
fish
; pharmaceutical pollutants
; antioxidant defense
; biomarkers of oxidative stress
; toxicological concerns
1. Introduction
1.1. Sources and Types of Human Pharmaceutical Active Compounds in the Aquatic Environment
Human activities, encompassing industrial production and agriculture, profoundly exacerbate environmental pollution through organic and inorganic contaminants. These pollutants include a wide spectrum, encompassing plastics, pharmaceutically active compounds, pesticides, heavy metals, etc.
Pharmaceutically active compounds (PhACs) are utilized by humans for their everyday health. Still, large quantities of pharmaceuticals are also used as veterinary medicine on farms worldwide [1] to prevent and treat animal diseases and increase economic benefits in intensive livestock.
In recent years, the consumption of human pharmaceutical products, which contain one or more pharmaceutically active compounds, has increased substantially. The European Union (EU) reported the use of over 3000 medicinal substances in human medicine [2], such as analgesics, anti-inflammatories, contraceptives, antibiotics, anticonvulsants, beta-blockers, antihistamines, anesthetics, vitamins, hormones, etc [3]. A report published by the IQVIA Institute for Human Data Science [4] projects that consumption per capita medicine use will increase across most regions, except in Africa and the Middle East, where overall volume growth is solely attributed to population growth.
The predominant pathways for these compounds to enter the aquatic environment are involuntary and purposeful human actions [5]. Involuntary excretion of medications through the body and their release into wastewater and intentional disposal of expired medicines into sewage or waste are the primary pathways through which these substances enter the surrounding environment. Pharmaceuticals and their metabolites reach soils and groundwater through the use of animal manure as fertilizer, as well as through sludge from wastewater treatment plants [6]. Pharmaceutical pollution can also originate from multiple sources, including industrial discharges, agricultural runoff, agricultural waste, improper discharge of effluents from the pharmaceutical industry or hospitals, or wastewater discharge from intensive animal farming [7,8,9].
Recent research conducted worldwide has revealed the presence of pharmaceuticals in surface water, groundwater, drinking water, soil, and aquatic plants, signaling environmental contamination and serious endangerment of aquatic ecosystems [10,11,12,13]. Pharmaceuticals and personal care products (PhACs) are present in water at low concentrations, typically ranging from ngL-1 to µgL-1 [14,15,16]. However, because some of these substances are long-lasting, they can bioaccumulate in organisms, leading to serious health problems [17,18,19].
The contamination of the aquatic environment with pharmaceuticals has emerged as a pressing global concern, drawing widespread public attention and prompting intensified efforts by scientists and governments to control further environmental deterioration. In this sense, the European Commission urges to classify them as emerging water pollutants under the Water Framework Directive.
In response to significant levels of pharmaceutical pollutants found in European waters, the European Commission acted in 2013 by placing three pharmaceutical substances—17-alpha-ethinylestradiol, 17-beta-estradiol, and diclofenac—on an initial watch list for monitoring water quality across its member states [20]. The objective of this directive was to gather data on these highly toxic pollutants, which are not currently part of broader European monitoring initiatives, to classify them as priority substances requiring targeted regulatory measures. However, in 2018, the European Commission determined that diclofenac should be removed from the watch list, citing the availability of “sufficient high-quality monitoring data” [21]. In 2020, this list was completed by the European Commission with five other PhACs such as amoxicillin, ciprofloxacin, sulfamethoxazole, trimethoprim, and venlafaxine [22]. In 2022, ofloxacin was added to the list of substances for Union-wide monitoring [23].
The most common PhACs found in aquatic environments are antibiotics (like ciprofloxacin, sulfamethoxazole, tetracycline, and azithromycin) [24,25,26], analgesics and anti-inflammatory drugs such as ibuprofen, diclofenac, aspirin, and naproxen [27,28,29,30], antidepressants including fluoxetine [31,32], anticonvulsants and mood stabilizers like carbamazepine [27,33,34], beta-blockers such as atenolol, propranolol, and metoprolol [35,36], hormones including estrogens (estradiol, ethinylestradiol) and progesterone [37,38], antihistamines [39], etc.
1.2. Oxidative Stress and Antioxidant Defense at Fish
Fish primarily encounter PhACs through two main routes: direct absorption from the water (bioconcentration) and ingestion through their diet. When both exposure pathways co-occur, bioaccumulation happens and is characterized by the uptake rate surpassing the elimination rate [40]. As these pharmaceutical pollutants accumulate in aquatic organisms, they can significantly disrupt homeostasis and essential organism functions by interfering with the endocrine system and hormonal secretions [41]. Additionally, the presence of these pollutants can induce oxidative stress, leading to further detrimental effects on the health and functioning of aquatic life.
1.2.1. Impacts of Oxidative Damage
Oxidative stress (OS) occurs when the production and accumulation of reactive oxygen species (ROS) within cells and tissues surpass the biological system’s ability to detoxify these reactive products, resulting in an imbalance [42]. The concept of OS was first introduced by Sies in 1985 [43], who described it as a disruption in the balance between pro-oxidants and antioxidants. This imbalance arises when the production of free radicals, generated through normal cellular processes or environmental exposures to oxidizing agents exceeds the cellular antioxidant defense capacity, which includes both enzymatic and non-enzymatic antioxidant molecules [44]. In a subsequent definition, Jones (2006) [45] further characterized OS as disrupting redox signaling control.
Aquatic organisms, like all aerobic organisms, require oxygen for metabolic processes. However, oxygen also has a negative effect, known as oxygen toxicity, due to the formation of ROS. Among the ROS, the most frequently produced are superoxide anion (O2-), hydroxyl radical (OH), and a non-radical compound, hydrogen peroxide (H2O2) [46]. Hydroxyl radicals have a short lifespan and are considered the most reactive and harmful ROS. Hydrogen peroxide (H2O2) is produced in large quantities in the mitochondrial matrix during oxygen reduction and has a longer lifespan than most ROS. [47]. Under normal conditions, electrons are transferred through the mitochondrial electron transport chain during the reduction of oxygen to water. However, approximately 1- 3 % of these electrons can leak from the system, forming superoxide radicals [48]. The superoxide radical (O2-) produced in cells initiates a series of chemical reactions, affecting multiple biological functions.
ROS are generated by all aerobic organisms through their normal metabolism, primarily in mitochondria and plasma membranes [49]. In moderate amounts, ROS play a beneficial role in the body; for instance, they help the immune system in attacking and eliminating pathogens [50]. However, excessive ROS production can overwhelm cellular defenses, disrupting immune homeostasis and resulting in OS. This imbalance adversely affects health and is implicated in the development of various chronic diseases. High levels of ROS can damage key biomolecules, leading to lipid peroxidation, protein oxidation, and the degradation of DNA, RNA, and carbohydrates. Such damage has severe cellular and physiological consequences, ultimately causing cells to self-destruct before completing their natural life cycle [51,52,53,54].
Lipid peroxidation is one primary effect, where ROS attacks cell membrane lipids, resulting in compromised membrane integrity and cell viability. This process produces reactive aldehydes, such as malondialdehyde (MDA), which can further propagate cellular damage [55]. This process frequently occurs during OS, and the lipid peroxidation index (MDA) is often used as a biomarker of oxidative degradation in fish.
Protein oxidation occurs when ROS modify amino acids, altering protein structure and function, potentially leading to enzyme inactivation and impaired cellular signaling. DNA oxidation is another critical outcome, where ROS causes strand breaks and base modifications that can lead to mutations, genomic instability, and even carcinogenesis in some organisms [46].
DNA and RNA damage. ROS can compromise the integrity of genetic material by causing oxidative damage to DNA and RNA. ROS can induce DNA strand breaks and base modifications, which may result in mutations and disrupt cellular functions. Similarly, in RNA, ROS production disrupts RNA stability, hinders protein synthesis, and impairs cellular signaling, contributing to cellular dysfunction and potentially leading to disease. Prolonged ROS exposure can increase mutation rates and inhibit protein synthesis, likely due to impaired ribosome function and enhanced mRNA degradation [56,57,58].
Carbohydrate oxidation, though less common, can disrupt cellular energy storage and metabolism.
These processes collectively drive the onset and progression of various diseases, including inflammatory conditions and neurodegenerative disorders. In aquatic organisms exposed to environmental stressors, such as pharmaceuticals, excessive ROS production, and the resulting biomolecular damage can adversely affect their health, growth, reproduction, and survival [59]. This poses significant risks to individual organisms, population sustainability, and overall ecosystem stability.
1.2.2. Antioxidant Defenses as Biomarkers of Oxidative Stress
To counteract the effects of OS, organisms have developed multiple defense mechanisms, involving the activity of specific enzymatic antioxidants that can be used as biomarkers of OS: superoxide dismutase (SOD), catalase (CAT), glutathione S-transferase (GST), glutathione peroxidase (GPx), reduced glutathione (GRed) [60]. In the antioxidant defense system of fish, in addition to the enzymatic antioxidants, low molecular weight antioxidants (non-enzymatic antioxidants) are also found, such as thioredoxin, carotenoids, vitamins E, K, and C, amino acids, and peptides [61]. Under typical physiological conditions, antioxidant defenses effectively manage to neutralize reactive oxygen species (ROS). However, disturbances in normal physiology can overwhelm these defenses, resulting in OS. This imbalance damages essential macromolecules, heightening vulnerability to disease, impairing immune responses, reducing survival chances, and hastening the aging process in animals.
Antioxidants serve as protective agents against the harmful effects of these reactive species, safeguarding cells and tissues from oxidative damage. They achieve this by converting ROS into non-reactive or less reactive compounds and repairing oxidative modifications that have already occurred. Non-enzymatic antioxidants, for instance, oxidize themselves to reduce free radicals. Research has established a clear relationship between physiological states and the efficacy of the body’s antioxidant system, emphasizing that enhancing the efficiency and levels of antioxidant defenses is vital for promoting overall health and resilience in organisms [62,63].
Among specific enzymatic antioxidants, superoxide dismutase (SOD) plays a crucial role as a primary line of defense against OS. It catalyzes the reduction of the superoxide anion radical (O2-) to hydrogen peroxide (H2O2) and O2, thereby protecting cells against OS and damage caused by free radicals. In fish, SOD is found in tissues such as the liver, kidneys, heart, and gills [64].
CAT degrades hydrogen peroxide (H2O2) produced from the breakdown of the superoxide anion by SOD [65]. The enzyme is present in both mitochondria and peroxisomes and catalyzes the decomposition of H2O2 into H2O and O2, thus maintaining a balance between ROS formation and elimination [66]. Additionally, GPx plays a detoxifying role by breaking down hydrogen peroxide (H2O2) into water (H2O) and oxygen (O2), thereby reducing the level of ROS in cells. Thus, catalase protects cells against OS caused by excessive ROS production.
GPx can convert GRed to oxidized glutathione (GSSG) via hydrogen peroxide and other organic peroxides, aiding in their elimination from cells and tissues. This enzyme maintains an optimal level of GRed in cells and prevents OS by removing hydroxyl radicals and hydrogen peroxide from the body [62].
Glutathione-S-transferase (GST) is a critical enzyme in the biotransformation process, particularly in phase II conjugation, where it plays a central role in cellular defense against OS. By catalyzing the conjugation of glutathione to reactive intermediates, GST helps neutralize ROS and maintain cellular redox balance, protecting cells from oxidative damage. This enzyme supports detoxification by binding glutathione to various endogenous and exogenous compounds, increasing their water solubility and facilitating bodily excretion. Through its -SH (sulfhydryl) group, GST also defenses cells from a range of hydrophobic and electrophilic substances, such as peroxidized lipids and xenobiotics. Conjugation of glutathione with these compounds forms mercapturic acids, which are subsequently excreted, completing the detoxification process [67].
GRed is present in nearly all body tissues, with higher concentrations in the liver. GRed is a tripeptide composed of the amino acid’s glutamate, cysteine, and glycine. It acts as a significant antioxidant in the body, protecting cells against the harmful effects of free radicals and OS. GRed also plays a crucial role in detoxifying toxic substances from the body and regulating metabolic processes, which are essential for normal immune system function and other physiological systems [68].
Exposure to pharmaceutical compounds can induce OS in organisms, particularly in aquatic environments where these substances accumulate and persist. These compounds, designed to exert specific biological effects can inadvertently generate reactive oxygen species (ROS) in non-target organisms such as fish and aquatic invertebrates. The OS can disrupt cellular redox balance, damaging lipids, proteins, and DNA. Gradual exposure to these substances can contribute to fish adaptation to new conditions, but sudden and prolonged exposure can cause significant biochemical and physiological changes. These changes weaken fish organisms and can intensify stress levels, potentially leading to death.
Understanding the OS responses elicited by pharmaceutical exposure is crucial for assessing environmental risks and developing mitigation strategies to safeguard aquatic ecosystems and the organisms dependent on them. Thus, by identifying and quantifying these OS biomarkers, we can better understand the molecular mechanisms underlying the development of various diseases and conditions associated with OS. Changes in the activity of these enzymes during the antioxidant response are used as biomarkers of OS [46,52,62,69]. Therefore, these markers, which provide crucial information about organism responses to OS, can be used to monitor the health of aquatic environments.
This review aims to comprehensively analyze the responses of OS biomarkers in various fish species exposed to environmentally relevant concentrations of pharmaceuticals (PhACs). Additionally, it aims to evaluate the utility of these biomarkers in advancing ecotoxicological risk assessments. This knowledge is important for shaping regulatory frameworks and enhancing environmental protection measures to safeguard aquatic ecosystems and mitigate potential risks to human populations posed by pharmaceutical pollutants.
2. Methodology
2.1. Data sources and Bibliometric Tools
In this study, the Web of Science core collection database was used to retrieve the publications related to pharmaceutical wastewater. VOSviewer and Publish of Perish 8 were used for the retrieval results for bibliometrics analysis.
After conducting a meta-analysis of studies on the Web of Science platform and applying the criteria of “pharmaceutical” and “fish oxidative stress,” 358 articles published between 2004-2024 were identified. For the definition of OS, we referred to some articles published between 1985-2006. The co-occurrence map of research items was generated using the VOSviewer program. The software automatically defined clusters based on a distance metric that reflects co-occurrence frequencies between terms. This analysis yielded five distinct clusters (Figure 1), each representing a group of terms closely related to pharmaceutical impacts on OS in aquatic organisms.
The bibliometric analysis’s first cluster (red color) focuses on various aspects of the aquatic environment. Key topics within this cluster include chronic exposure, diclofenac, exposure effects, fish, impact, paracetamol, relevant concentrations, toxic effects, and development. This cluster emphasizes the study of pharmaceuticals, particularly diclofenac and paracetamol, in aquatic ecosystems, especially concerning their chronic exposure effects on fish. The research highlights the importance of understanding these substances’ impacts and toxic effects at relevant environmental concentrations, which is essential for assessing their developmental implications in aquatic organisms.
Cluster 2 (green color) focuses on various aspects of fish response to contaminants. This cluster emphasizes the study of how different fish species, such as crucian carp (Carassius carassius) and rainbow trout (Oncorhynchus mykiss), respond to contaminants through antioxidant responses and bioconcentration. It also highlights the use of biomarkers to assess the effects and toxicity of these contaminants, providing insights into the health and environmental impact on these fish species.
Cluster 3 (blue color) is focused on the physiological and biochemical responses of Nile tilapia (Oreochromis niloticus) to pharmaceuticals. Key areas of study include gill and liver effects, OS biomarkers, and response pathways, highlighting the impact of pharmaceutical exposure on this species.
Cluster 4 (light green color) investigates the impact of pharmaceuticals like carbamazepine and ibuprofen on common carp (Cyprinus carpio). The research within this cluster emphasizes early life stages, OS, and the effects of short-term exposure to these substances.
Finally, cluster 5 (purple color), focuses on the biochemical and chronic effects of fluoxetine on zebrafish (Danio rerio). This cluster highlights studies that explore long-term impacts and stress responses in these fish, providing insights into the persistent effects of fluoxetine exposure.
3. Discussions
This review evaluates the impact of human pharmaceuticals on fish, focusing on the role of OOS biomarkers as indicators of sublethal toxicity.
Worldwide, the most frequently detected pharmaceuticals in aquatic environments include non-steroidal anti-inflammatory drugs (NSAIDs), antibiotics, antidepressants, and antiepileptics [70]. Understanding the prevalence of these compounds is essential for formulating effective strategies to mitigate their impact on aquatic ecosystems.
3.1. The Influence of Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) on Oxidative Stress in Fish
Non-steroidal anti-inflammatory drugs (NSAIDs) are one of the main groups detected in aquatic ecosystems. Between these, diclofenac (DCF), ibuprofen (IBF), naproxen (NPX), ketoprofen (KETO), and aspirin (ASA) are frequently reported [30,71].
In fish, exposure to NSAIDs disrupts the activity of antioxidant enzymes, increasing lipid peroxidation and DNA damage (Table 1). These changes indicate heightened oxidative stress and cellular damage caused by NSAIDs in aquatic organisms. NSAIDs’ primary mode of action is the inhibition of cyclooxygenase (COX) enzymes, specifically COX-1 and COX-2. These enzymes are essential for producing eicosanoids, signaling molecules that play a vital role in various physiological processes [72]. Importantly, the metabolites of NSAIDs are often more toxic than the compounds themselves, further amplifying their harmful effects in aquatic environments. However, the intricate mechanisms underlying NSAID toxicity in fish remain poorly understood [73].
Among the nonsteroidal anti-inflammatory drugs (NSAIDs) examined, diclofenac (DCF) emerges as a significant contaminant in aquatic ecosystems. Recognized as a pseudo-persistent micro-pollutant, DCF has been detected in surface waters at concentrations ranging from 21 to 722 ng/L [74]. In Romania, Chițescu et al. [75] investigated the presence of diclofenac in the Argeș River, finding concentrations up to 166 ng/L. Hallare et al. [76] reported that in Central European surface waters, DCF concentrations reach up to 0.54 μg/L. These findings confirm that the primary contamination sources are domestic and urban wastewater, hospital effluents, and uncontrolled discharges from pharmaceutical manufacturing processes.
The potential for DCF to undergo biotransformation in aquatic organisms after absorption has not yet been studied. In mammals, however, DCF is metabolized into two main metabolites: 4′-hydroxy diclofenac and 5′-hydroxy diclofenac. These metabolites are further oxidized into benzoquinone imine, a compound known to be toxic to fish. This process triggers the formation of reactive oxygen species (ROS) and leads to binding with sulfhydryl groups in the cytosol, as well as in enzymes and membrane proteins containing these groups [77,78].
Consequently, DCF and its environmental degradation products may impose significant ecological risks. Their presence can enhance ROS production, impairing fish’s antioxidant defense mechanisms.
For example, exposure of Danio rerio embryos and larvae to diclofenac (DCF) concentrations of 0, 0.5, 5, 50, and 500 μg/L for 96 hours resulted in elevated catalase (CAT) and glutathione peroxidase (GPx) activities, alongside a decrease in glutathione S-transferase (GST) activity [79]. These findings suggest the occurrence of pro-oxidative alterations associated with DCF metabolism. Elevated CAT levels indicate a defensive cellular response to OS caused by increased hydrogen peroxide production (H₂O₂). Meanwhile, heightened GPx activity likely reflects an adaptive mechanism to counteract excessive ROS. Conversely, the observed inhibition of GST activity may stem from its denaturation due to direct interactions with ROS, as Diniz et al. [80] hypothesized. The simultaneous increases in CAT and GPx activities support the notion of ROS overproduction following DCF exposure. Under conditions of increased ROS, GST isoenzymes may become inactivated due to oxidative damage. Additionally, the increased GPx activity suggests enhanced GSH oxidation, underscoring its critical role in mitigating OS and aiding the interpretation of these biochemical alterations.
Exposure of male tiger fish (Hoplies malabaricus) by intraperitoneal inoculation with DCF at several doses (0.2; 2.0 or 20.0 μg/kg) has significantly increased antioxidant responses in fish liver, suggesting the generation of free radicals. For instance, SOD activity increased at doses of 2 and 20 μg/kg, GSH levels at 20 μg/kg, and GPx activity was elevated across all tested doses. Also, DCF caused hepatic LPO in all exposed groups, indicating OS responses. Furthermore, DCF reduced the liver’s GST activity, demonstrating that biotransformation was inhibited (Table 1). These findings highlight the role of SOD as the first line of antioxidant defense, converting superoxide radicals (O₂⁻) into hydrogen peroxide (H₂O₂), which is then further neutralized by GPx and CAT to mitigate oxidative damage [81].
Similarly, a study by Eze et al. (2021) [82] investigated the effects of DCF exposure on Nile tilapia (Oreochromis niloticus). The experiment involved three sub-lethal concentrations of DCF (250, 320, and 480 µg/L), with fish exposed for 28 days followed by a 7-day recovery period. The findings demonstrated that DCF exposure induced OS in a dose- and time-dependent manner. The significant increase in lipid peroxidation (LPO) with DCF exposure underscores the drug’s capacity to enhance reactive oxygen species (ROS) production, which can lead to DNA damage, protein oxidation, and physiological disturbances. The observed reduction in the activities of superoxide dismutase (SOD) and catalase (CAT) aligns with the heightened LPO levels, suggesting that the inhibition of these critical antioxidant enzymes compromises the fish’s ability to mitigate OS. Additionally, the study reported significant increases in glutathione reductase (GRed) activities and glutathione peroxidase (GPx), which may represent a compensatory response to OS. Glutathione peroxidase (GPx) plays a crucial role in reducing hydrogen peroxide and lipid peroxides, thereby preventing the formation of radical intermediates through oxygen reduction mechanisms [83]. An observed increase in GPx activity likely reflects the enzyme’s effort to mitigate the effects of heightened ROS production [83]. In this context, elevated GPx levels further support the notion of an adaptive response to OS induced by diclofenac exposure. This adaptation, however, appears insufficient to fully counteract the oxidative damage, as evidenced by the persistent elevation of LPO. Interestingly, the increase in glutathione (GSH) activity indicates an upregulation of the antioxidant defense system, although not enough to prevent the adverse effects of prolonged diclofenac exposure. Generally, GSH plays a critical role in maintaining immune system function and exhibits antioxidative, integrative, and detoxifying effects. The slight recovery observed after a 7-day withdrawal period suggests that Oreochromis niloticus has some capacity to recover from OS. However, the authors reported un uncompleted recovery, particularly for SOD and CAT activities. This partial recovery highlights the potential for long-term environmental impacts of DCF, especially with continuous exposure (Table 1).
Contrary to the earlier information, some authors suggest that DCF may also have a protective effect on fish-specific physiological or biochemical processes. For instance, Guiloski et al. [84] found that a 21-day exposure to various environmental concentrations of DCF (0, 2, 20 μg/L) in the fish species Rhamdia quelen led to decreased activities of antioxidant enzymes such as SOD and CAT, alongside increased GSH concentrations and GST activity (Table 1).
Critical to the antioxidant defense system, these alterations likely contributed to the observed reduction in lipid membrane damage. The study highlights that DCF protects against lipid peroxidation (LPO), which may explain the absence of protein carbonylation and DNA damage. A particularly significant finding was the reduction in LPO at concentrations of 0.2 and 20 µg/L of DCF, where the authors reported no evidence of DNA damage. Furthermore, CAT activity decreased only in the 2 µg/L exposure group, and this effect was not concentration-dependent due to the low doses tested. The observed increase in GSH levels and GST activity likely played a critical role in preventing DNA damage in the blood and liver of Rhamdia quelen (Table 1).
Similarly, juveniles of Rhamdia quelen exposed to diclofenac (DCF) at concentrations of 0.2, 2, and 20 μg/L for 96 hours exhibited significant physiological changes. In the kidney, SOD activity increased at all tested concentrations, suggesting disruptions in H2O2 metabolism, although no DNA damage or lipid peroxidation was observed. These results underscore the potential for acute DCF exposure, even at low levels. However, the authors reported no evidence of oxidative stress in the liver [85].
These findings support the assertion of Petersen et al. [86], who states that low concentrations of DCF exposure can protect cells from OS. Moreover, Feito et al. [87] noted a decreased level of LPO after exposure to 0.03 µg/L DCF for 90 min in zebrafish embryos. Similarly, Praskova et al. [88] observed a reduction in LPO levels in 20-day-old zebrafish exposed to an environmental concentration of 0.02 µg/L DCF, typically found in Czech rivers, over 28 days. On the other hand, the authors declare no alteration of GST and GRed activity.
The reduction in LPO observed in these studies may be explained by the ability of increased GSH and GST activity to neutralize ROS and mitigate OS, thereby preventing damage to lipids and other cellular components. By enhancing the antioxidant defense mechanisms, these concentrations of DCF may induce a protective effect that counters the oxidative damage typically associated with higher concentrations. However, at higher concentrations, the authors suggest that LPO rises, eventually entering what is termed the “zone of compensation. In this zone, the „beneficial low-dose response” fails to reduce damage, and biomarker levels become comparable to those observed in the control group [87].
However, the ecotoxicological impacts of realistic DCF concentrations remain largely unexplored. Most of the studies used DCF concentrations in the range of mg/L, which is why these findings should be interpreted with caution, considering the complexity of interactions between chemical substances and aquatic organisms and the potential long-term effects on ecosystem health. Nonetheless, prolonged or high-dose exposure often overwhelms these defense mechanisms, leading to persistent oxidative damage and physiological disturbances. The variability in tissue-specific responses and the incomplete recovery of antioxidant systems following exposure underscore the long-term ecological risks associated with DCF contamination in aquatic environments, highlighting the need for further investigation into its sub-lethal and chronic effects.

Table 1.
The influence of pharmaceuticals on oxidative stress in fish.
| Category |
Pharm. prod. |
Species | Concentration | Effects on the physiological level | Ref. |
| NSAIDs | DCF |
Danio rerio (embryos and larvae) |
0; 0.5; 5; 50 and 500 μg/L for 96 hours | (+) CAT activity and GPx at 500 μg/L; (-) GSTs in all concentrations. |
[79] |
| Hoplias malabaricus | 0; 0.2; 2.0; 20 μg/kg, after intraperitoneal inoculation with 12 doses | (+) SOD, GPx, LPO, and GSH in the liver; (-) GST; No modifications of CAT activity. |
[81] | ||
| Oreochromis niloticus | 0; 250; 320; 480 μg/L for 28 days | (+) LPO; (+) GRed, GPx and GSH; (-) SOD and CAT. |
[82] | ||
| Rhamdia quelen | 0; 0.2; 2 and 20 μg/L for 21 days | (-) SOD; (+) GSH; (+) GST in all concentrations- in the liver: (-) LPO at 2 and 20 μg/L; (-) CAT at 2 μg/L; No modification of GPx activity. |
[84] | ||
| Rhamdia quelen | 0; 0.2; 2 and 20 μg/L for 96 hours | (+) SOD in the kidney at all concentrations; No alteration of CAT and GPx; (-) LPO; significant decrease of DNA damage in the kidney at 20 μg/L. |
[85] | ||
| IBF | Rhamdia quelen | 0.1, 1, and 10 μg/L for 14 days | (+) GST in all groups; (+) GPx and GSH activity at concentrations of 10 μg/L in the kidney; No modifications of SOD, CAT, and LPO activities. |
[94] | |
| Tinca tinca | 0.02 -60 μg/L for 35 days | (-) GST in 60 μg/L; (-) GST No modification of CAT activity and LPO |
[93] | ||
| Oncorhynchus mykiss | 2 and 200 μg/kg feed | In gills (-) GPX at all concentrations; In liver (+) LPO and GRed at 200 μg/kg feed; No changes in the activity of antioxidant enzymes in the kidney. |
[91] | ||
| ASA | Labeo rohita | 1, 10, 100 μg/L for 7, 14, 21 and 28 days | (-) SOD, CAT, GPx, GRed, GSH in liver at al conc.; (+) GST and LPO. | [104] | |
| Mugilogobius abei | 0.5, 5, and 50 μg/ L for 24, 72, 168 hours | (+) SOD, CAT, GPx, and GST activity; (-) GSH after 24 and 72 h; (+) GSH 168 hours; (+) MDA; (-) MDA after 168 hours |
[105] | ||
| APAP | Rhamdia quelen | 0, 0.25, 2.5 µg/ L, for 21 days | (+) SOD activity at a concentration of 2.5 µg/L; (-) GST at all concentrations; No modification of GPx, GSH, and LPO |
[109] | |
| Danio rerio embryos | 150, 300, 450, 600, 750, 900, 1050, and 1200 μg/L | (+) MDA and CAT; (+) SOD from conc. of 300 -1200 μg/L | [110] | ||
| Anguilla anguilla | 5, 25, 125, 625, and 3125 μg/L |
In liver: (+) GST at 625 and 3125 mg/L; LPO remained unaltered In gills: (-) GST; (+) LPO |
[111] | ||
| Antibiotics | OTC |
Danio rerio |
0, 0.1, 10, and 10,000 mg/L for 2 months | (-) GST and CAT | [117] |
| SMZ | Oreochromis niloticus | 0, 1, 10 and 100 μg/L SMZ for 7 and 30 days | In liver (+) SOD, CAT, GPx, GSH at 1 and 10 μg/L CAT and GSH, (-) LPO; At 100 μg/L SMZ (-) SOD and GSH; (+) LPO at both 7 and 30 days. |
[119] | |
| CIP | Danio rerio | 0,7 µg/L, 100, 650, 1100 and 3000 µg/L for 28 days |
(+) GST at 0,7 and 100 µg/L; (-) GST at 650, 1100, and 3 000 μg/L; (-) GRed at conc. of 1100 and 3000 µg/L; (-) GPX, at all tested concentrations, except for the 100 µg/L; (-) LPO at 100 μg/L |
[120] | |
| Antiepileptics | CBZ | Cyprinus carpio | 0, 1, 5, 50, or 100 µg/L of CBZ for 28 days | (+) CAT and GRed at 5 and 50 µg/ L) at 100 µg/L (-) GRed.; (-) SOD. | [126] |
| CBZ | Cyprinus carpio | 2000 µg/L exposure from 12-96 hours | (-) LPO in the brain after 24,48, 72 hours; (-) SOD in liver, gills and brain; (-) CAT; (+) GPx in liver, after 48 hours; (-)GPx at 96 hours; (-) GPx in the brain and gills. |
[128] | |
| Antidepressants | FLX | Danio rerio | 5, 16, 40 ng/L for 96 hours | (+) SOD, CAT, GPx in the liver, intestine, brain, and gills. (+) MDA in the brain and tissues after 96 hours at conc. of 5-40 ng/L |
[131] |
| FLX | Danio rerio | 0.0015, 0.05, 0.1, 0.5 and 0.8μM for 80 hours | (+) CAT (0.0015 and 0.5μM) (-) SOD (0.0015 and 0.5μM) |
[138] | |
| Pharm. mixtures | DCF+IBF | Oncorhynchus mykiss | DCF - 2 and 200 μg/kg); IBF- 2 and 200 μg/kg. Combination of DCF and IBF – (2 μg/kg DCF+ 2 μg/kg; 200 μg/kg IBF). |
In gills: (-) GPx activity at IBP 2 and 200 μg kg/kg and the combination of DCF and IBF; In liver: (+) LPO in DCF and IBP, and DCF conc. of 200 μg/kg (+) GR activity at IBF 200 μg/kg; In the posterior kidney: (+) CAT at DCF 200 μg/kg. |
[91] |
| DCF+ APAP | Cyprinus carpio | 50 μg of each/L, 1:1) | (+) SOD in the brain; (-) SOD in liver and gills; (+) CAT in the brain and gills; (+) GPx in brain and liver; (+) LPO in liver and gills. | [146] | |
| CBZ, irbesartan, APAP, NPX, DCF | Oncorhynchus mykiss | concentrations of 1x, 10x, and 100x the median levels found in the Meuse River, Belgium, over 42 days | No change of GST; (-) GSH after 24 hours; (-) GPx and CAT |
[147] |
Abbreviations: MDA – malondialdehyde; LPO – lipid peroxidation; TAC- Total antioxidant capacity; GSH – Glutathione; SOD - superoxide dismutase; CAT - catalase; GST - glutathione S-transferase; GpX - glutathione peroxidase; GRed - reduced glutathione; DCF- Diclofenac; DXM-Dexametasone; IBF- Ibuprofen; ASA-Acetylsalicylic acid; APAP- Paracetamol; SMZ- Sulfamethoxazole; OTC- oxytetracycline; CIP- Ciprofloxacin; FLOL- Florfenicol; CBZ- Carbamazepine; FLX- Fluoxetine, NPX-naproxen.
Another commonly detected non-steroidal anti-inflammatory medicine in aquatic environments is ibuprofen (IBF), widely used by humans for its analgesic (pain-relieving), antipyretic (fever-reducing), and anti-inflammatory properties.
Studies have reported concentrations of IBF in aquatic environmental samples typically in the μg/L range. For example, in the USA surface waters, IBF was found at values of 0.2 µg/L [89], while in Brazil higher concentrations were detected (0.326–2.094 µg/L) [90].
Ibuprofen has been shown to induce genotoxic effects in fish, cause histopathological damage to tissues such as gills and kidneys, and disrupt the activity of OS enzymes [91,92,93].
For instance, adult male Rhamdia quelen exposed to environmentally relevant concentrations of IBF (0.1, 1, and 10 μg/L) for 14 days showed increased GST, GPx, and GRed activity in the kidney (Table 1). However, no changes in DNA integrity were detected in the posterior kidney or liver. Additionally, IBF did not affect the biochemical biomarkers in the liver or gills. These enzymes play a crucial role in the antioxidant defense system by eliminating ROS, and the observed increase in GST and GPx activity, along with higher GSH levels, likely contributed to preventing DNA damage in the posterior kidney [94].
In another study, Stancová et al. [93] investigated the effects of IBF exposure on Tinca tinca over 35 days, using environmental concentrations of 0.02, 0.2, and 2 μg/L (Table 1). The study found significant biochemical responses, including reduced GPx and CAT activity. These changes could elevate oxidative stress and lipid peroxidation, alongside a decrease in the activity of GST, an enzyme critical for detoxification processes. Despite biochemical changes, authors reported no lipid oxidative damage. This absence of damage suggests a potential adaptive response to low-level OS during prolonged exposure. However, the lack of temporal data on antioxidant enzyme activity and LPO levels limits understanding of how these parameters evolved, indicating possible mitigation mechanisms in the fish. In conclusion, the findings suggest that the fish may have developed mechanisms to mitigate oxidative stress throughout the experiment.
For grass carp, IBF exposure (0, 4.8, 48.0, or 480.0 ng/L) for 14 days led to increased LPO and disrupted enzymatic activities [95]. Also, exposure of Danio rerio for a long term to environmentally relevant concentrations of IBF (0.1–11 μg/L) promotes the production of radical species and induces oxidative stress on the brain, gills, liver, and gut tissues [96].
In Oncorhynchus mykiss, IBF exposure through feed at 2 and 200 μg/kg caused reduced GPX activity in the gills, while at 200 μg/kg, it increased LPO and GRed levels in the liver. However, no significant effects were observed in kidney antioxidant enzymes, indicating organ-specific responses to IBF exposure [91].
In addition to NSAIDs like IBF and DCF, other medications frequently found in the aquatic environment are acetylsalicylic acid (ASA), commonly known as aspirin, and paracetamol (APAP) or acetaminophen [97,98].
Acetylsalicylic acid (ASA), is widely recognized for its anti-inflammatory, analgesic, and antipyretic properties. At the same time, Paracetamol (APAP) or acetaminophen is widely employed worldwide for pain relief and fever reduction, and it is commonly found in sewage treatment plant effluents, surface water, and drinking water.
Compared to DCF and IBF, acetylsalicylic acid (ASA) and paracetamol (APAP) are considered to have a relatively lower impact on water quality. For instance, ASA has been detected at concentrations ranging from 0.011 to 0.855 μg/L in the North Sea and the Scheldt Estuary [99], and from 0.025 to 0.29 μg/L in the Lis River, Portugal [100]. Similarly, APAP has been detected at concentrations of 9.6–183 ng/L in Romanian waters [101], 420–610 ng/L in Angke and Ancol, Jakarta Bay, Indonesia [102], and 65 μg/L in the Tyne River, UK [3], respectively in higher concentrations 246 μg/L in Spanish waters [103]. Although these substances are found in aquatic systems at low concentrations, studies have shown that exposure to ASA can lead to OS and decrease an organism’s ability to detoxify these harmful substances.
According to Gayen et al. [104], aspirin exposure to Labeo rohita at 1, 10, and 100 μg/L for 7, 14, 21, and 28 days, induces OS, evidenced by significant, dose- and duration-dependent decreases in antioxidant enzyme activities (CAT, GPx, GR) and GSH content. GST activity increased with higher doses but was not influenced by the duration of exposure. Additionally, the authors observed that lipid peroxidation (LPO) registered an increasing trend with dose and time of exposure. Statistical analysis revealed that concentration and exposure duration significantly impacted these biomarkers, with concentration having the most substantial effect on most parameters. The decrease in SOD activity and GSH content in the liver of Labeo rohita suggests disruption of the oxidative balance and impairment of the antioxidant defense system due to ASA exposure. This reduction in key antioxidants likely contributes to the loss of adaptive mechanisms, leaving the fish more vulnerable to severe OS.
Exposure of Mugilogobius abei at ASA concentrations of 0.5, 5, and 50 μg/ L for 24 h, 72 h, and 168 h induced significant changes in antioxidant and OS markers. SOD, CAT, GPx, and GST activities were generally increased, while GSH content decreased initially after 24 and 72 h after exposure, but showed a significant increase after 168 hours. MDA content exhibited a high growth throughout the exposure period, indicating OS, although it decreased after 168 hours. These results suggest that ASA exposure leads to OS in organisms, with varying responses over time [105].
Regarding the impact of the environmental concentrations of APAP on fish, the literature findings revealed teratogenic, neurotoxic, and cardiotoxic effects in embryos/larvae of Clarias gariepinus [106]. Additionally, other studies have demonstrated that chronic exposure of rainbow trout can lead to significant alterations in the histology and function of organs responsible for ion and nutrient homeostasis [107].
Generally, APAP undergoes a metabolic transformation in the body, resulting in the formation of ROS, primarily due to its conversion to N-acetyl-p-benzoquinone imine (NAPQI), an electrophilic metabolite. NAPQI is known to increase ROS levels, including superoxide anion, hydroxyl radical, and hydrogen peroxide, as documented by Yen et al., 2007 [108].
According to Guilkoski et al. [109], the elevation of ROS levels induced by APAP metabolism is corroborated by the observed induction of SOD activity and protein carbonylation in male fish of Rhamdia quelen exposed to environmental concentrations of APAP (0, 0.25, 2.5 μg/L) for 21 days. Notably, GPx activity remained unchanged, suggesting that the H2O2 generated by SOD may accumulate within cells rather than being efficiently converted to H2O. This accumulation likely contributes to protein carbonylation observed in fish exposed to both low and high concentrations of APAP, along with DNA damage in hepatocytes exposed to the lower concentration of the drug.
A study conducted by Rosas-Ramírez et al. [110] examined the impact of acetaminophen on zebrafish embryos at varying concentrations (0150, 300, 450, 600, 750, 900, 1050, and 1200 μg/L) (Table 1). The findings revealed that exposure to acetaminophen (150–1200 μg/L) led to an increase in the production of malondialdehyde (MDA) and catalase (CAT) activity. This effect is likely due to elevated H2O2 levels generated during acetaminophen exposure. Additionally, the study observed that exposure to acetaminophen at concentrations ranging from 300 to 1200 μg/L resulted in increased SOD activity in D. rerio embryos.
Nunes et al. [111] investigated the effects of acetaminophen (APAP) exposure on European eel (Anguilla anguilla) at concentrations of 5, 25 (typical for freshwater systems [3]), 125, 625, and 3125 μg/L. The study found no OS alterations at environmentally relevant concentrations (Table 1). However, at higher dosages, a paradoxical effect was observed, characterized by impaired CAT activity of GST and LPO. These findings suggest that eels possess efficient detoxification mechanisms, which help prevent metabolic disruptions and maintain their primary energy pathways.
3.2. The Influence of Antibiotics on Fish Oxidative Stress
The widespread use of antibiotics results in considerable contamination of aquatic environments. Excessive use of antibiotics poses a significant risk to the health of fish and aquatic ecosystems [112]. Even at low concentrations, antibiotics persist in water and can bioaccumulate in aquatic environments progressively increasing their levels, and being toxic to aquatic organisms [113]. Another concern is that antibiotics can facilitate the development of antibiotic-resistant strains, leading to unintended toxicity to fish [37].
Research generally suggests that antibiotic exposure in fish disrupts their oxidoreductive balance, leading to increased levels of ROS, and the inactivation of enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), resulting in damage to cellular components like lipids, proteins, and DNA [114]. Also, prolonged exposure can overwhelm these protective mechanisms, impairing physiological functions [115], inducing OS, and increasing fish susceptibility to diseases [116]. Generally, the specific nature and extent of these changes are likely influenced by several factors, including the type and dosage of the antibiotic, as well as the species and developmental stage of the fish.
For instance, Almeida et al. [117] conducted a study investigating the long-term (2 months) effects of oxytetracycline (OTC) on zebrafish (Danio rerio) oxidative stress. They exposed zebrafish to varying concentrations of OTC (0, 0.1, 10, and 10,000 mg/L), with the lower concentrations (0.1 and 10 mg/L) reported in environmental waters. Their findings showed a significant decrease in the activity of GST and CAT enzymes (Table 1). This reduction suggests that prolonged exposure to antibiotics can diminish the activity of antioxidant defenses, leading to oxidative damage likely caused by OTC accumulation. This damage may result in tissue damage and subsequently lower enzyme activity. However, as noted by Massarsky et al. [118], the activity of antioxidant enzymes can vary depending on the severity of OS.
Exposure to sulfamethoxazole (SMZ) has been shown to exert dose-dependent effects on juvenile Nile tilapia (Oreochromis niloticus), at concentrations up to 100 μg/L over 7 and 30 days. The biochemical and molecular markers revealed oxidative stress responses. At low concentrations (1 and 10 μg/L), SMZ enhanced antioxidant enzyme activities (SOD, CAT, GPx), increased glutathione (GSH) levels, and reduced lipid peroxidation (LPO), suggesting a compensatory activation of defense mechanisms. In contrast, higher concentrations (100 μg/L) suppressed SOD activity, depleted GSH, and exacerbated LPO, indicating oxidative damage. These findings, supported by transcriptional changes in antioxidant enzyme genes, highlight the dual role of SMZ, where low doses elicit adaptive responses, while high doses overwhelm the oxidative balance. Such dose-dependent effects underline the importance of considering sub-lethal endpoints in environmental risk assessments of pharmaceutical pollutants [119].
Ciprofloxacin (CIP) exposure of zebrafish (Danio rerio) at an environmental concentration of 0.7 μg/L, and higher concentrations of 100, 650, 1100, and 3 000 μg/L, induced variable responses in antioxidant enzymes and OS markers, even at 0.7 μg/L (Table 1). For example, GST activity was significantly elevated suggesting an adaptive enzymatic response to oxidative challenge. However, a marked reduction in GRed activity was observed at higher concentrations (1100 and 3000 μg/L), indicating potential disruption of the antioxidant regeneration system under more severe exposure. Similarly, GPx activity decreased significantly across all tested concentrations except 100 μg/L, highlighting a dose-specific impact on enzymatic detoxification pathways. Interestingly, the concentration of thiobarbituric acid reactive substances (TBARS), indicative of lipid peroxidation, was reduced only at 100 μg/L, suggesting a potential protective effect at this intermediate level [120]. These findings underscore the complexity of CIP impact, where both adaptive and adverse responses are concentration-dependent.
3.3. The Influence of Antiepileptics Drugs on Fish Oxidative Stress
Of the common antiepileptic drugs met in the aquatic environment carbamazepine (CBZ) is among the most widely detected [121]. CBZ is an antiepileptic medication, derived from dibenzoazepine, and is used as an antiepileptic agent in the treatment of certain types of epilepsy and also for neuropathic pain. Conventional methods in wastewater treatment plants do not biodegrade CBZ and may enter the aquatic environment by different pathways [122], posing a significant risk to aquatic organisms due to its potential for absorption and bioconcentration.
CBZ concentrations in aquatic ecosystems have been detected at varying levels: ranging from 20 to 49 ng/L according to Chițescu et al. 2020 [123], while other studies reported higher concentrations varying from 0.1 to 1.3 µg/L [124]. Exposure of fish to CBZ leads to the modulation of liver GST activity and a decrease in CAT activity after 63 days of continuous exposure [125].
In a study conducted by Liang et al. [126], common carp (Cyprinus carpio) were exposed to various environmentally relevant subacute concentrations of CBZ (0, 1, 5, 50, and 100 μg/L) for 28 days. The results revealed significant changes in the activities of antioxidant defence system enzymes in the liver, indicating a pronounced oxidative stress effect caused by CBZ. Specifically, during the first seven days of exposure, there was an initial surge in the activities of SOD and GRed. However, after the 14th day of exposure, the activities of these enzymes had decreased to levels below those observed in the control group and remained suppressed in fish exposed for 28 days. In contrast, the activity of CAT showed a marked increase throughout the 28-day exposure period compared to the control group. Moreover, a notable rise in GST activity was also observed. According to some authors [127], the decline of SOD and GRed enzymes may be due to factors such as lipid peroxidation and direct OS from reactive oxygen species, reduced protein levels for ROS scavenging, or an energy deficiency (NADPH) following prolonged exposure to CBZ. The rise of the antioxidant enzyme activity may be attributed to the increased levels of ROS, which activate the antioxidant defense system after exposure to CBZ. This activation, including elevated superoxide dismutase (SOD) activity in the fish tissues, helps to convert the ROS generated by CBZ into less harmful molecules.
Even short exposure of fish to CBZ can induce OS. A study by Gasca Perez et al. [128], investigated the impact of short-term exposure to carbamazepine (CBZ) on oxidative stress in common carp (Cyprinus carpio). Fish were exposed to CBZ at a concentration of 2000 µg/L for durations ranging from 12 to 96 hours. The researchers observed significant responses in oxidative stress markers across different organs. Specifically, lipid peroxidation (LPO) levels in the brain showed a notable decrease after 24, 48, and 72 hours of exposure, indicating a potential adaptive response to mitigate oxidative damage. Conversely, SOD activity in the liver, gills, and brain decreased significantly within 12 hours of exposure, suggesting an early onset of oxidative stress induced by CBZ. CAT activity exhibited a pronounced reduction across all studied organs throughout the exposure period, underscoring the vulnerability of antioxidant defenses to CBZ-induced OS. GPx activity displayed varying trends among organs, with an initial increase in the liver at 48 hours followed by a decline, while gills and brain tissues consistently showed reduced GPx activity over time. These findings highlight organ-specific responses to CBZ exposure and emphasize the potential ecological implications of pharmaceutical contaminants in aquatic environments.
3.4. The Influence of Antidepressant Drugs on Fish Oxidative Stress
Antidepressant drugs, commonly prescribed for treating mental health disorders in humans, are increasingly detected in aquatic environments due to their persistence and incomplete removal during wastewater treatment processes [129,130]. These pharmaceuticals can significantly impact non-target organisms, such as fish, by altering their biochemical and physiological processes.
Specifically, antidepressants affect OS biomarkers and the activity of antioxidant defense mechanisms in fish. It is well established that OS can lead to protein oxidation, resulting in altered functionality and the formation of new low molecular weight aggregates [131].
Among these drugs, fluoxetine (FLX) is the most frequently detected in aquatic environments, with concentrations ranging from 0.33 to 32.1 ng/L [132] and 0.012 to 1.4 μg/L [133,134]. The findings revealed that FLX prompted the induction of OS in various organs of the fish, encompassing the liver, gut, brain, and gills.
Orozco-Hernández et al. [131], assessed the potential toxicological effects of fluoxetine (FLX) at environmentally relevant concentrations (5, 16, and 40 ng/L) during a 96-hour acute exposure of Danio rerio. Their findings indicated an increase in the activity levels of SOD, CAT, and GPx in the liver, intestine, brain, and gills. Particularly, the study reported a significant increase in MDA levels in Danio rerio embryos and brain tissue after 96-hour exposure to FLX at concentrations between 5 and 40 ng/L. This finding suggests heightened oxidative stress in these sensitive developmental and neurological tissues, even at low environmental concentrations of FLX.
Also, other authors reported elevated values of CAT and SOD enzymatic activity in juvenile Argyrosomus regius after 15 days of exposure to FLX concentrations of 300 and 3000 ng/L [135]. In another study, acute exposure for 3 and 6 days to 0.1 μg/L of FLX has been reported to increase the CAT and SOD activities and the total antioxidant capacity (TAC) in adult Carassius auratus liver [136].
In contrast, other studies have reported decreased antioxidant activities following exposure to moderate and high concentrations of fluoxetine. Ding et al. [137], found that SOD activity was inhibited in adult Carassius auratus after a 7-day exposure to 4 and 100 μg/L of FLX. Similarly, Cunha et al. [138], reported SOD inhibition in Danio rerio embryos exposed for 80 hours to 0.0015 and 0.5μM FLX, increasing CAT activity at the same concentrations (Table 1). A possible explanation for these fluctuations is the transient nature of antioxidant enzymes, meaning that the duration of exposure can significantly influence their responses.
Additionally, exposure to fluoxetine increased LPO in the liver of Carassius auratus at concentrations of 4, 20, and 100 μg/L for 7 days, indicating oxidative damage. This suggests that FLX can oxidize proteins, altering their functionality and forming new low molecular weight aggregates. However, decreased LPO levels have also been observed, such as in juvenile Argyrosomus regius exposed for 15 days to 0.3 and 3 μg/L of FLX [139], highlighting the variability in responses depending on the species and conditions of exposure.
Nevertheless, the exact mechanism by which FLX induces oxidative stress is not fully understood, this pharmaceutical agent is believed to trigger oxidative stress through multiple pathways. Several researchers have suggested that FLX may affect mitochondrial function [140]. Consequently, the disruption of mitochondrial function by FLX can lead to increased ROS production, ultimately resulting in the induction of oxidative stress.
3.5. The Influence of Pharmaceutical Mixture on Fish Oxidative Stress
Another significant concern among scientists is a mixture of pharmaceutical drugs in aquatic environments. For example, the presence of a mixture of NSAIDs at concentrations ranging from a few ng/L to a few μg/L has been frequently reported in marine and estuarine waters [71], surface waters, groundwater [141,142], and even in drinking water [143].
Beyond pharmaceuticals, contaminants such as pesticides, industrial chemicals, and heavy metals frequently coexist in aquatic systems, creating a mixture of stressors [144,145]. These complex mixtures pose a significant challenge to aquatic ecosystems, as their combined effects often challenge predictions based on single-compound toxicity, underscoring the need for integrated approaches to assess their influence on fish health and OS biomarkers [145].
For example, when fish are exposed to multiple NSAIDs simultaneously, the cumulative concentration of these drugs can overwhelm the body’s detoxification and repair mechanisms. A study by Hodkovicova et al. [91] investigated the toxic effects of oral administration of environmental doses of DCF and IBP and their mixture on rainbow trout (Oncorhynchus mykiss). After exposing the fish to these NSAIDs for six weeks, the study revealed visible signs of inflammation and OS, along with impaired homeostasis and innate immunity, particularly in fish exposed to the combination of DCF and IBP (2 μg/kg DCF and 2 μg/kg IBP). Reduction of the GPx activity in the gills of fish exposed to the mixture of DCF and IBP indicates that these organs might be experiencing oxidative stress, potentially leading to damage at the cellular or tissue level.
Exposure of Common carp (Cyprinus carpio) to a mixture of DCF and APAP (50 μg of each/L, 1:1), revealed that OS responses vary across tissues based on physiological roles and exposure routes (Table 1) [146]. In the brain and gills, increased SOD and CAT activities suggest an adaptive response to elevated reactive oxygen species (ROS), while higher GPx in the brain highlights its vulnerability to oxidative damage. The liver shows a contrasting pattern, with decreased SOD but increased GPx and LPO, indicating significant oxidative stress that may overwhelm its antioxidant defenses during detoxification processes. Elevated LPO in the liver and gills signifies membrane damage caused by excessive free radicals, reflecting their roles in metabolizing pollutants and direct exposure to contaminants, respectively. While antioxidant enzyme activity increases in some tissues to counteract ROS, others show signs of damage, particularly in the liver, which may be a primary target of pharmaceutical toxicity.
While certain pharmaceutical mixtures have been shown to cause significant oxidative stress in aquatic organisms, Beghin et al. [147] proved that not all combinations lead to detrimental effects. In their study, juvenile rainbow trout (Oncorhynchus mykiss) were exposed to a mixture of five pharmaceuticals (carbamazepine, irbesartan, paracetamol, naproxen, and diclofenac), from the categories of neuroleptic, antihypertensive, analgesic and nonsteroidal anti-inflammatory drugs, at concentrations of 1x, 10x, and 100x the median levels found in the Meuse River, Belgium, over 42 days (Table 1). The results showed no change in GST activity, while GSH levels decreased significantly after 24 hours, with a concentration-dependent decline observed by day 42, particularly in the 10× group. This GSH depletion was linked to reduced GPx activity, and CAT activity was lower in the 100× group on day 1. However, despite these changes, the fish’s detoxification mechanisms effectively cleared the pharmaceuticals, preventing significant oxidative damage.
4. Conclusions
This review presents the impact of the most common pharmaceuticals found in the aquatic environment (non-steroidal anti-inflammatory drugs (NSAIDs), antibiotics, antiepileptics, and antidepressant drugs) on fish oxidative stress response.
As a main conclusion, we observed that research on the effects of pharmaceuticals on aquatic life at ecologically relevant concentrations is limited. Most of the studies that investigate the effects of these pharmaceuticals on aquatic organisms focus on high concentrations that are not representative of the low, environmentally relevant levels, typically found in natural water systems. While these high-concentration studies provide valuable insights into acute toxicity and potential mechanisms of action, they may overestimate the risks and fail to capture the subtle, chronic effects that occur under environmental conditions.
Despite their presence at low concentrations in aquatic environments, pharmaceuticals pose significant toxicological and ecotoxicological risks, particularly when found as components of complex mixtures. Interactions within these mixtures can result in unpredictable and synergistic effects, heightening the risks to aquatic ecosystems. Evaluating their impact on aquatic organisms and the environment remains challenging due to these substances’ constantly changing concentrations and interactions.
Assessing the impact of these substances is challenging due to the dynamic variability in their concentrations and ratios, as well as the number of pharmaceuticals involved. Also, a big gap is that few studies employ multiple biomarkers to evaluate contaminant effects across tissues and systems in fish, an approach essential for understanding species-specific toxicity, which seldom varies widely.
In conclusion, the exposure of fish to environmental concentrations of pharmaceuticals predominantly induces chronic long-term toxic effects rather than acute responses. Prolonged exposure increases ROS production, increases lipid peroxidation production, can impair enzymatic activity, and weakens antioxidant defenses, such as glutathione and catalase. These biochemical disruptions highlight how the duration and intensity of exposure critically influence the responses of organisms.
To address these risks, comprehensive studies are needed to explore the long-term ecotoxicological effects of pharmaceuticals, at lower concentrations (ng/L). Additionally, careful monitoring and evaluation are essential to mitigate potential harm to aquatic ecosystems and safeguard public health against environmental contamination.
In addition to the biochemical disruptions caused by pharmaceuticals, their impact extends to critical biological processes such as fish growth and reproduction. Altered OS responses can disrupt endocrine signaling pathways, impair gametogenesis, and hinder larval development. Further, prolonged oxidative damage may lead to reduced fish reproductive success, developmental abnormalities, and diminished growth rates, ultimately threatening fish populations and biodiversity. These findings emphasize the need to investigate the links between OS, physiological performance, and population-level effects in fish exposed to environmentally relevant concentrations of pharmaceuticals. Such studies are critical for understanding and mitigating the broader ecological consequences of pharmaceutical pollution.
Author Contributions
Conceptualization, L.G., L.D. M.C.; methodology, M.C., L.G., validation C.V., L.D., M.C.; formal analysis, L.G., and M.C.; investigation, M.C., L,D., L.G., writing—original draft preparation, L.D., M.C..; writing—review and editing, L.G.., L.D., M.C.; visualization, L.G.; C.V., L.D., supervision, L.D., M.C and C.V.; All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data are available from the first author, and can be delivered if required.
Acknowledgments
The authors would like to acknowledge to the project POCU/993/6/13 (Program to increase performance and innovation in excellent doctoral and postdoctoral research PROINVENT). The project was co-financed by the European Social Fund through the Human Capital Operational Program.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cunningham, V.L.; Constable, D.J.C.; Hannah, R.E. Environmental Risk Assessment of Paroxetine. Environ. Sci. Technol. 2004, 38, 3351–3359. [CrossRef]
- Fent, K.; Weston, A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [CrossRef]
- Roberts, P.; Thomas, K. The occurrence of selected pharmaceuticals in wastewater effluent and surface waters of the lower Tyne catchment. Sci. Total. Environ. 2006, 356, 143–153. [CrossRef]
- IQVIA Institute for Human Data Science. https://www.iqvia.com/insights/the-iqvia-institute/reports-and-publications/reports/the-global-use-of-medicines-2023, accessed at 29 May 2024.
- Daughton, C.G. Pharmaceuticals in the environment: sources and their management. Wilson Wilsons Compr. Anal. Chem., 2007, 50, 1-58.
- Chonova, T.; Keck, F.; Labanowski, J.; Montuelle, B.; Rimet, F.; Bouchez, A. Separate treatment of hospital and urban wastewaters: A real scale comparison of effluents and their effect on microbial communities. Sci. Total. Environ. 2016, 542, 965–975. [CrossRef]
- Wollenberger, L.; Halling-Sørensen, B.; Kusk, K. Acute and chronic toxicity of veterinary antibiotics to Daphnia magna. Chemosphere 2000, 40, 723–730. [CrossRef]
- Brausch, J.M.; Connors, K.A.; Brooks, B.W.; Rand, G.M. Human Pharmaceuticals in the Aquatic Environment: A Review of Recent Toxicological Studies and Considerations for Toxicity Testing. In Reviews of Environmental Contamination and Toxicology Volume 218; Whitacre, D.M., Ed.; Springer: Boston, MA, USA, 2012; Volume 218, pp. 1–99, ISBN 978-1-4614-3137-4.
- Koubová, A.; Van Nguyen, T.; Grabicová, K.; Burkina, V.; Aydin, F.G.; Grabic, R.; Nováková, P.; Švecová, H.; Lepič, P.; Fedorova, G.; et al. Metabolome adaptation and oxidative stress response of common carp (Cyprinus carpio) to altered water pollution levels. Environ. Pollut. 2022, 303, 119117. [CrossRef]
- Santos, L.H.M.L.M.; Araújo, A.N.; Fachini, A.; Pena, A.; Delerue-Matos, C.; Montenegro, M.C.B.S.M. Ecotoxicological aspects related to the presence of pharmaceuticals in the aquatic environment. J. Hazard. Mater. 2010, 175, 45–95. [CrossRef]
- Taylor, D.; Senac, T. Human pharmaceutical products in the environment–the “problem” in perspective. Chemosphere, 2014, 115, 95-99.
- Burkina, V.; Zlabek, V.; Zamaratskaia, G. Effects of pharmaceuticals present in aquatic environment on Phase I metabolism in fish. Environ. Toxicol. Pharmacol. 2015, 40, 430–444. [CrossRef]
- Sharma, J.; Joshi, M.; Bhatnagar, A.; Chaurasia, A.K.; Nigam, S. Pharmaceutical residues: One of the significant problems in achieving ‘clean water for all’ and its solution. Environ. Res. 2022, 215, 114219. [CrossRef]
- García-Morales, R.; García-García, A.; Orona-Navar, C.; Osma, J.F.; Nigam, K.; Ornelas-Soto, N. Biotransformation of emerging pollutants in groundwater by laccase from P. sanguineus CS43 immobilized onto titania nanoparticles. J. Environ. Chem. Eng. 2018, 6, 710–717. [CrossRef]
- Kondor, A.C.; Molnár, É.; Vancsik, A.; Filep, T.; Szeberényi, J.; Szabó, L.; Maász, G.; Pirger, Z.; Weiperth, A.; Ferincz, Á.; et al. Occurrence and health risk assessment of pharmaceutically active compounds in riverbank filtrated drinking water. J. Water Process. Eng. 2021, 41. [CrossRef]
- Forsberg, M. Occurrence of organic micropollutants and hormones in Swedish surface water. Uppsala University, Disciplinary Domain of Science and Technology, Earth Sciences, Department of Earth Sciences, 2022, 47p.
- Grabicova, K.; Grabic, R.; Fedorova, G.; Fick, J.; Cerveny, D.; Kolarova, J.; Turek, J.; Zlabek, V.; Randak, T. Bioaccumulation of psychoactive pharmaceuticals in fish in an effluent dominated stream. Water Res. 2017, 124, 654–662. [CrossRef]
- Corcoran, J.; Winter, M.J.; Tyler, C.R. Pharmaceuticals in the aquatic environment: A critical review of the evidence for health effects in fish. Crit. Rev. Toxicol. 2010, 40, 287–304. [CrossRef]
- Nakano, T.; Hayashi, S.; Nagamine, N. Effect of excessive doses of oxytetracycline on stress-related biomarker expression in coho salmon. Environ. Sci. Pollut. Res. 2015, 25, 7121–7128. [CrossRef]
- Directive 2013/39/eu https://eur-lex.europa.eu/legal-content/RO/TXT/PDF/?uri=CELEX:32013L0039&from=IT [Last accessed on 1 June 2024].
- Commission Implementing Decision (EU) 2018/840 of 5 June 2018 establishing a watch list of substances for Union-wide monitoring in the field of water policy pursuant to Directive 2008/105/EC of the European Parliament and of the Council and repealing Commission Implementing Decision (EU) 2015/495 (notified under document C(2018) 3362). Available from: https://eurlex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018D0840. [Last accessed on 1 June 2024].
- European Commission, 2020. EUR-Lex – Regulation (EU) 2020/741 of the European Parliament and of the Council of 25 May 2020 on minimum requirements for water reuse. Off. J. Eur. Union L177, 32–54. [Last accessed on 1 June 2024].
- Eu, 2022 https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32022D1307, [Last accessed on 1 June 2024].
- Kümmerer, K. Antibiotics in the aquatic environment–a review–part I. Chemosphere, 2009, 75(4), 417-434.
- Mukhtar, A.; Manzoor, M.; Gul, I.; Zafar, R.; Jamil, H.I.; Niazi, A.K.; Ali, M.A.; Park, T.J.; Arshad, M. Phytotoxicity of different antibiotics to rice and stress alleviation upon application of organic amendments. Chemosphere 2020, 258, 127353. [CrossRef]
- Sosa-Hernández, J.E.; Rodas-Zuluaga, L.I.; López-Pacheco, I.Y.; Melchor-Martínez, E.M.; Aghalari, Z.; Limón, D.S.; Hafiz M.N.; Parra-Saldívar, R. Sources of antibiotics pollutants in the aquatic environment under SARS-CoV-2 pandemic situation. Case stud. Chem. Environ. Eng., 2021, 4, 100127.
- Chițescu, C.L.; Kaklamanos, G.; Nicolau, A.I.; Stolker, A.A.M.L. High sensitive multi-residue analysis of pharmaceuticals and antifungals in surface water using U-HPLC-Q-Exactive Orbitrap HRMS. Application to the Danube River basin on the Romanian territory. Sci. Total Environ., 2015, 532, 501-511.
- Das, S.A.; Karmakar, S.; Chhaba, B.; Rout, S.K. Ibuprofen: its toxic effect on aquatic organisms. J. Exp. Zoology India, 2019, 22(2), 1125-1131.
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman Jr., C.U.; Mohan, D. Pharmaceuticals of emerging concern in aquatic systems: chemistry, occurrence, effects, and removal methods. Chem. Rev., 2019, 119(6), 3510-3673.
- Placova, K.; Halfar, J.; Brozova, K.; Heviankova, S. Issues of Non-Steroidal Anti-Inflammatory Drugs in Aquatic Environments: A Review Study. The 4th International Conference on Advances in Environmental Engineering. p. 13.
- Schultz, M.M.; Furlong, E.T. Trace Analysis of Antidepressant Pharmaceuticals and Their Select Degradates in Aquatic Matrixes by LC/ESI/MS/MS. Anal. Chem. 2008, 80, 1756–1762. [CrossRef]
- Correia, D.; Domingues, I.; Faria, M.; Oliveira, M. Effects of fluoxetine on fish: What do we know and where should we focus our efforts in the future Sci. Total Environ, 2023, 857, 159486.
- Bahlmann, A.; Weller, M.G.; Panne, U.; Schneider, R.J. Monitoring carbamazepine in surface and wastewaters by an immunoassay based on a monoclonal antibody. Anal. Bioanal. Chem. 2009, 395, 1809–1820. [CrossRef]
- Baali, H.; Cosio, C; Effects of carbamazepine in aquatic biota. Environ. Sci. Process. Impacts, 2022, 24(2), 209-220.
- Cleuvers, M. Initial risk assessment for three β-blockers found in the aquatic environment. Chemosphere 2005, 59, 199–205. [CrossRef]
- Sumpter, J.P.; Runnalls, T.J.; Donnachie, R.L.; Owen, S.F. A comprehensive aquatic risk assessment of the beta-blocker propranolol, based on the results of over 600 research papers. Sci. Total. Environ. 2021, 793, 148617. [CrossRef]
- Young, R.B.; Borch, T. Sources, Presence, Analysis, and Fate of Steroid Sex Hormones in Freshwater Ecosystems–A review. Reprod., 2009, 1(3).
- Laurenson, J.P.; Bloom, R.A.; Page, S.; Sadrieh, N. Ethinyl Estradiol and Other Human Pharmaceutical Estrogens in the Aquatic Environment: A Review of Recent Risk Assessment Data. AAPS J. 2014, 16, 299–310. [CrossRef]
- Berninger, J.P.; Du, B.; Connors, K.A.; Eytcheson, S.A.; Kolkmeier, M.A.; Prosser, K.N.; Theodore W.’ Valenti Jr. C.; Chambliss C.K.; Brooks B.W. Effects of the antihistamine diphenhydramine on selected aquatic organisms. Environ. Toxicol. Chem., 2011, 30(9), 2065-2072.
- A Arnot, J.; Gobas, F.A. A review of bioconcentration factor (BCF) and bioaccumulation factor (BAF) assessments for organic chemicals in aquatic organisms. Environ. Rev. 2006, 14, 257–297. [CrossRef]
- Eapen, J.V.; Thomas, S.; Antony, S.; George, P.; Antony, J. A review of the effects of pharmaceutical pollutants on humans and aquatic ecosystem. 2024, 2, 484–507. [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [CrossRef]
- Sies, H. Oxidative stress: damage to intact cells and organs. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 1985, 311(1152), 617-631.
- Rock C.L; Jacob R.A; Bowen P.E; Update on biological characteristics of the antioxidant micronutrients - Vitamin C, Vitamin E and the carotenoids. J. Am. Diet. Assoc. 1996, 96, 693– 702.
- Jones, D.P. Redefining Oxidative Stress. Antioxidants Redox Signal. 2006, 8, 1865–1879. [CrossRef]
- Bolaric, S; Vokurka, A; Batelja Lodeta, K; Bencic, Ð. Genotyping of Croatian Olive Germplasm with Consensus SSR Markers. Horticulturae. 2024, 10, 417. [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [CrossRef]
- Fridovich, I. Mitochondria: are they the seat of senescence?. Aging Cell 2003, 3, 13–16. [CrossRef]
- Hong, Y.; Boiti, A.; Vallone, D.; Foulkes, N.S. Reactive Oxygen Species Signaling and Oxidative Stress: Transcriptional Regulation and Evolution. Antioxidants 2024, 13, 312. [CrossRef]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxidants & redox signaling, 2014, 20(6), 1000-1037.
- Ghosh, N.; Das, A.; Chaffee, S.; Roy, S.; Sen, C. K. Reactive oxygen species, oxidative damage and cell death. In Immunity and inflammation in health and disease, 2018, (pp. 45-55). Academic Press.
- Ji, L.L.; Yeo, D. Oxidative stress: an evolving definition. Fac. Rev. 2021, 10. [CrossRef]
- Juan, C.A.; de la Lastra, J.M.P.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [CrossRef]
- Bresciani, G.; da Cruz, I.B.M.; González-Gallego, J. Manganese superoxide dismutase and oxidative stress modulation. Adv. Clin. Chem, 2015, 68, 87-130.
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [CrossRef]
- Kong, Q.; Lin, C.-L.G. Oxidative damage to RNA: mechanisms, consequences, and diseases. Cell. Mol. Life Sci. 2010, 67, 1817–1829. [CrossRef]
- Martins, S.G.; Zilhão, R.; Thorsteinsdóttir, S.; Carlos, A.R. Linking Oxidative Stress and DNA Damage to Changes in the Expression of Extracellular Matrix Components. Front. Genet. 2021, 12, 673002. [CrossRef]
- Khan, F.; Garg, V.K.; Singh, A.K.; Tinku, T. Role of free radicals and certain antioxidants in the management of huntington’s disease: a review. J. Anal. Pharm. Res. 2018, 7, 1. [CrossRef]
- Srivastava, B.; Reddy, P. B. Impacts of human pharmaceuticals on fish health. Int. J. Pharm. Sci. Res, 2021, 12(5185), 5185-94.
- Rodriguez, Y.E.; Laitano, M.V.; Pereira, N.A.; López-Zavala, A.A.; Haran, N.S.; Fernández-Gimenez, A.V. Exogenous enzymes in aquaculture: Alginate and alginate-bentonite microcapsules for the intestinal delivery of shrimp proteases to Nile tilapia. Aquaculture 2018, 490, 35–43. [CrossRef]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as Potential Antioxidants: A Systematic Review. J. Agric. Food Chem. 2015, 63, 3615–3626. [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [CrossRef]
- Olufunmilayo, E.O.; Gerke-Duncan, M.B.; Holsinger, R.M.D. Oxidative Stress and Antioxidants in Neurodegenerative Disorders. Antioxidants 2023, 12, 517. [CrossRef]
- SanJuan-Reyes, N.; Gómez-Oliván, L.M.; Galar-Martínez, M.; Vieyra-Reyes, P.; García-Medina, S.; Islas-Flores, H.; Neri-Cruz, N. Effluent from an NSAID-Manufacturing Plant in Mexico Induces Oxidative Stress on Cyprinus carpio. Water, Air, Soil Pollut. 2013, 224, 1–14. [CrossRef]
- Ramos, A.; Correia, A.; Antunes, S.; Gonçalves, F.; Nunes, B. Effect of acetaminophen exposure in Oncorhynchus mykiss gills and liver: Detoxification mechanisms, oxidative defence system and peroxidative damage. Environ. Toxicol. Pharmacol. 2014, 37, 1221–1228. [CrossRef]
- Devi, G.; Harikrishnan, R.; Paray, B.A.; Al-Sadoon, M.K.; Hoseinifar, S.H.; Balasundaram, C. Effect of symbiotic supplemented diet on innate-adaptive immune response, cytokine gene regulation and antioxidant property in Labeo rohita against Aeromonas hydrophila. Fish Shellfish. Immunol. 2019, 89, 687–700. [CrossRef]
- Sherratt, P.J.; Hayes, J.D. Glutathione S-transferases. Enzyme systems that metabolize drugs and other xenobiotics, 2019, 319-352.
- Waggiallah, H.; Alzohairy, M. The effect of oxidative stress on human red cells glutathione peroxidase, glutathione reductase level, and prevalence of anemia among diabetics. North Am. J. Med Sci. 2011, 3, 344–347. [CrossRef]
- Kumari, K.; Khare, A.; Dange, S. The Applicability of Oxidative Stress Biomarkers in Assessing Chromium Induced Toxicity in the FishLabeo rohita. BioMed Res. Int. 2014, 2014, 1–11. [CrossRef]
- wiacka, K.; Maculewicz, J.; Kowalska, D.; Caban, M.; Smolarz, K.; Świeżak, J. Presence of pharmaceuticals and their metabolites in wild-living aquatic organisms–current state of knowledge J. Hazard. Mater., 2022, 424, 127350.
- Blasco, J.; Trombini, C. Ibuprofen and diclofenac in the marine environment - a critical review of their occurrence and potential risk for invertebrate species. Water Emerg. Contam. Nanoplastics 2023, 2, 14. [CrossRef]
- Hayashi, Y.; Heckmann, L.-H.; Callaghan, A.; Sibly, R.M. Reproduction recovery of the crustacean Daphnia magna after chronic exposure to ibuprofen. Ecotoxicology 2008, 17, 246–251. [CrossRef]
- Świacka, K.; Michnowska, A.; Maculewicz, J.; Caban, M.; Smolarz, K. Toxic effects of NSAIDs in non-target species: A review from the perspective of the aquatic environment. 2020, 273, 115891. [CrossRef]
- Acuña, V.; Ginebreda, A.; Mor, J.; Petrovic, M.; Sabater, S.; Sumpter, J.; Barceló, D. Balancing the health benefits and environmental risks of pharmaceuticals: Diclofenac as an example. Environ. Int. 2015, 85, 327–333. [CrossRef]
- Chitescu, C.L.; Kaklamanos, G.; Nicolau, A.I.; Stolker, A.A. High sensitive multiresidue analysis of pharmaceuticals and antifungals in surface water using U-HPLC-Q-Exactive Orbitrap HRMS. Application to the Danube river basin on the Romanian territory. Sci. Total Environ. 2015, 532, 501–511. [CrossRef]
- Hallare, A.V.; Köhler, H.-R.; Triebskorn, R. Developmental toxicity and stress protein responses in zebrafish embryos after exposure to diclofenac and its solvent, DMSO. Chemosphere 2004, 56, 659–666. [CrossRef]
- Oviedo-Gómez, D.G.C.; Galar-Martínez, M.; García-Medina, S.; Razo-Estrada, C.; Gómez-Oliván, L.M. Diclofenac-enriched artificial sediment induces oxidative stress in Hyalella azteca. Environ. Toxicol. Pharmacol. 2010, 29, 39–43. [CrossRef]
- Grillo, M.P.; Hua, F.; Knutson, G.C.; Ware, A.J.; Li, C.Mechanistic studies on the bioactivation of diclofenac: Identification of diclofenac-S-acyl-glutathione in vitro in incubations with rat and human hepatocytes. Chem. Res. Toxicol. 2003, 16, 1410–1417.
- Bio, S.; Nunes, B. Acute effects of diclofenac on zebrafish: Indications of oxidative effects and damages at environmentally realistic levels of exposure. Environ. Toxicol. Pharmacol. 2020, 78, 103394. [CrossRef]
- Diniz, M.S.; de Matos, A.P.A.; Lourenço, J.; Castro, L.; Peres, I.; Mendonça, E.; Picado, A. Liver Alterations in Two Freshwater Fish Species (Carassius auratusandDanio rerio) Following Exposure to Different TiO2Nanoparticle Concentrations. Microsc. Microanal. 2013, 19, 1131–1140. [CrossRef]
- Guiloski I.C.; Ribas J.L.C.; da Pereira L.S.; Neves A.P.P.; Silva de Assis H.C. Effects of Trophic Exposure to Dexamethasone and Diclofenac in Freshwater Fish. Ecotoxicol. Environ. Saf. 2015, 114, 204–211.
- Eze, C.C.; Nwamba, H.O.; Omeje, F.U.; Anukwu, J.U.; Okpe, M.N.; Nwani, C.D. Effects of Diclofenac on the Oxidative Stress Parameters of Freshwater Fish Oreochromis niloticus. J. Appl. Life Sci. Int. 2021, 44–51. [CrossRef]
- Pandey, L.K.; Bergey, E.A.; Lyu, J.; Park, J.; Choi, S.; Lee, H.; Depuydt, S.; Oh, Y.-T.; Lee, S.-M.; Han, T. The use of diatoms in ecotoxicology and bioassessment: Insights, advances and challenges. Water Res. 2017, 118, 39–58. [CrossRef]
- Guiloski, I.C.; Piancini, L.D.S.; Dagostim, A.C.; Calado, S.L.d.M.; Fávaro, L.F.; Boschen, S.L.; Cestari, M.M.; da Cunha, C.; de Assis, H.C.S. Effects of environmentally relevant concentrations of the anti-inflammatory drug diclofenac in freshwater fish Rhamdia quelen. Ecotoxicol. Environ. Saf. 2017, 139, 291–300. [CrossRef]
- Ghelfi, A.; Ribas, J.L.C.; Guiloski, I.C.; Bettim, F.L.; Piancini, L.D.S.; Cestari, M.M.; Pereira, A.J.; Sassaki, G.L.; de Assis, H.C.S. Evaluation of Biochemical, Genetic and Hematological Biomarkers in a Commercial Catfish Rhamdia quelen Exposed to Diclofenac. Bull. Environ. Contam. Toxicol. 2015, 96, 49–54. [CrossRef]
- Petersen, A.; Zetterberg, M.; Sjöstrand, J.; Pålsson, A.; Karlsson, J.-O. Potential Protective Effects of NSAIDs/ASA in Oxidatively Stressed Human Lens Epithelial Cells and Intact Mouse Lenses in Culture. Ophthalmic Res. 2005, 37, 318–327. [CrossRef]
- Feito, R., Valcárcel, Y., & Catalá, M. Biomarker assessment of toxicity with miniaturized bioassays: diclofenac as a case study. Ecotoxicology, 2012, 21, 289-296.
- Praskova, E.; Plhalova, L.; Chromcova, L.; Stepanova, S.; Bedanova, I.; Blahova, J.; Hostovsky, M.; Skoric, M.; Maršálek, P.; Voslarova, E.; et al. Effects of Subchronic Exposure of Diclofenac on Growth, Histopathological Changes, and Oxidative Stress in Zebrafish (Danio rerio). Sci. World J. 2014, 2014, 1–5. [CrossRef]
- Cahill, J.D.; Furlong E.T.; Burkhardt, M.R.; Kolpin, D.; Anderson, L.G.; Determination of pharmaceutical compounds in surface- and ground-water samples by solid-phase extraction and high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Chromatogr., 2004, 1041(1-2), 171–180.
- Pereira, C.D.S.; Maranho, L.A.; Cortez, F.S.; Pusceddu, F.H.; Santos, A.R.; Ribeiro, D.A.; Cesar, A.; Guimarães, L.L. Occurrence of pharmaceuticals and cocaine in a Brazilian coastal zone. Sci. Total. Environ. 2016, 548-549, 148–154. [CrossRef]
- Hodkovicova, N.; Hollerova, A.; Blahova, J.; Mikula, P.; Crhanova, M.; Karasova, D.; Franc, S. Pavlokova, Mares, Postulkova, E.; Tichy, F.; Marsalek, P.; Lanikova, J.; Faldyna, M.; Svobodova, Z. Non-steroidal anti-inflammatory drugs caused an outbreak of inflammation and oxidative stress with changes in the gut microbiota in rainbow trout (Oncorhynchus mykiss). Sci. Total Environ., 2022, 849, 157921.
- Ragugnetti, M.; Adams, M.L.; Guimarães, A.T.B.; Sponchiado, G.; de Vasconcelos, E.C.; de Oliveira, C.M.R. Ibuprofen Genotoxicity in Aquatic Environment: An Experimental Model Using Oreochromis niloticus. Water, Air, Soil Pollut. 2010, 218, 361–364. [CrossRef]
- Stancova, V.; Plhalova, L.; Blahova, J.; Zivna, D.; Bartoskova, M.; Siroka, Z.; Marsalek, P.; Svobodova, Z. cc. Veterinární medicína, 2017, 62(2), 90-97.
- Mathias, F.T.; Fockink, D.H.; Disner, G.R.; Prodocimo, V.; Ribas, J.L.C.; Ramos, L.P.; Cestari, M.M.; de Assis, H.C.S. Effects of low concentrations of ibuprofen on freshwater fish Rhamdia quelen. Environ. Toxicol. Pharmacol. 2018, 59, 105–113. [CrossRef]
- Zhang, N.; Liu, X.; Pan, L.; Zhou, X.; Zhao, L.; Mou, X.; Zhou, H.; Liu, J.; Wang, X. Evaluation of ibuprofen contamination in local urban rivers and its effects on immune parameters of juvenile grass carp. Fish Physiol. Biochem. 2021, 47, 1405–1413. [CrossRef]
- Sánchez-Aceves, L.; Pérez-Alvarez, I.; Gómez-Oliván, L.M.; Islas-Flores, H.; Barceló, D. Long-term exposure to environmentally relevant concentrations of ibuprofen and aluminum alters oxidative stress status on Danio rerio. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2021, 248, 109071. [CrossRef]
- Wadhah Hassan, A.E. Occurrence of paracetamol in aquatic environments and transformation by microorgan-isms: a review. Chron. Pharm. Sci., 2017, 1, 341-355.
- Satayeva, A.; Kerim, T.; Kamal, A.; Issayev, J.; Inglezakis, V.; Kim, J.; Arkhangelsky, E. Determination of aspirin in municipal wastewaters of Nur-Sultan City, Kazakhstan. IOP Conf. Series: Earth Environ. Sci. 2022, 1123, 012067. [CrossRef]
- Wille, K.; Noppe, H.; Verheyden, K.; Bussche, J.V.; De Wulf, E.; Van Caeter, P.; Janssen, C.R.; De Brabander, H.F.; Vanhaecke, L. Validation and application of an LC-MS/MS method for the simultaneous quantification of 13 pharmaceuticals in seawater. Anal. Bioanal. Chem. 2010, 397, 1797–1808. [CrossRef]
- Paíga, P.; Santos, L.H.; Ramos, S.; Jorge, S.; Silva, J.G.; Delerue-Matos, C. Presence of pharmaceuticals in the Lis river (Portugal): Sources, fate and seasonal variation. Sci. Total. Environ. 2016, 573, 164–177. [CrossRef]
- Ilie, M.; Deák, G.; Marinescu, F.; Ghita, G.; Tociu, C.; Matei, M.; Covaliu, C.I.; Raischi, M.; Yusof, S.Y. Detection of Emerging Pollutants Oxytetracycline and Paracetamol and the Potential Aquatic Ecological Risk Associated with their Presence in Surface Waters of the Arges-Vedea, Buzau-Ialomita, Dobrogea-Litoral River Basins in Romania. IOP Conf. Series: Earth Environ. Sci. 2020, 616, 012016. [CrossRef]
- Koagouw, W.; Arifin, Z.; Olivier, G.W.; Ciocan, C. High concentrations of paracetamol in effluent dominated waters of Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2021, 169, 112558. [CrossRef]
- Al-Kaf, G.; Naji, M.K.; Abdullah Q.Y.M.A.; Wadhah Hassan, A.E. Occurrence of paracetamol in aquatic environments and transformation by microorganisms: a review. Chron. Pharm. Sci, 2007, 1, 341-355.
- Gayen, T.; Tripathi, A.; Kumari, U.; Mittal, S.; Mittal, A.K. Ecotoxicological impacts of environmentally relevant concentrations of aspirin in the liver of Labeo rohita: Biochemical and histopathological investigation. Chemosphere 2023, 333, 138921. [CrossRef]
- Wang, Y., Wang, C., Bao, S., & Nie, X. Responses of the Nrf2/Keap1 signaling pathway in Mugilogobius abei (M. abei) exposed to environmentally relevant concentration aspirin. Environ Sci Pollut Res Int, 2020, 27, 15663-15673.
- Erhunmwunse, N.O.; Tongo, I.; Ezemonye, L.I. Acute effects of acetaminophen on the developmental, swimming performance and cardiovascular activities of the African catfish embryos/larvae (Clarias gariepinus). Ecotoxicol. Environ. Saf. 2021, 208, 111482. [CrossRef]
- Choi, E.; Alsop, D.; Wilson, J.Y. The effects of chronic acetaminophen exposure on the kidney, gill and liver in rainbow trout ( Oncorhynchus mykiss ). Aquat. Toxicol. 2018, 198, 20–29. [CrossRef]
- Yen, F.-L.; Wu, T.-H.; Lin, L.-T.; Lin, C.-C. Hepatoprotective and antioxidant effects of Cuscuta chinensis against acetaminophen-induced hepatotoxicity in rats. J. Ethnopharmacol. 2006, 111, 123–128. [CrossRef]
- Guiloski, I.C.; Ribas, J.L.C.; Piancini, L.D.S.; Dagostim, A.C.; Cirio, S.M.; Fávaro, L.F.; Boschen, S.L.; Cestari, M.M.; da Cunha, C.; de Assis, H.C.S. Paracetamol causes endocrine disruption and hepatotoxicity in male fish Rhamdia quelen after subchronic exposure. Environ. Toxicol. Pharmacol. 2017, 53, 111–120. [CrossRef]
- Rosas-Ramírez, J.R.; Orozco-Hernández, J.M.; Elizalde-Velázquez, G.A.; Raldúa, D.; Islas-Flores, H.; Gómez-Oliván, L.M. Teratogenic effects induced by paracetamol, ciprofloxacin, and their mixture on Danio rerio embryos: Oxidative stress implications. Sci. Total. Environ. 2022, 806, 150541. [CrossRef]
- Nunes, B.; Verde, M.F.; Soares, A.M.V.M. Biochemical effects of the pharmaceutical drug paracetamol on Anguilla anguilla. Environ. Sci. Pollut. Res. 2015, 22, 11574–11584. [CrossRef]
- Wan, J.; Guo, P.; Peng, X.; Wen, K. Effect of erythromycin exposure on the growth, antioxidant system and photosynthesis of Microcystis flosaquae. J. Hazard Mater., 2015, 283, 778–786.
- Zhao, X.-L.; Li, P.; Zhang, S.-Q.; He, S.-W.; Xing, S.-Y.; Cao, Z.-H.; Lu, R.; Li, Z.-H. Effects of environmental norfloxacin concentrations on the intestinal health and function of juvenile common carp and potential risk to humans. Environ. Pollut. 2021, 287, 117612. [CrossRef]
- Abu-Zahra, N.I.S.; Atia, A.A.; Elseify, M.M.; Soliman, S. Biological and histological changes and DNA damage in Oreochromis niloticus exposed to oxytetracycline: a potential amelioratory role of ascorbic acid. Aquac. Int. 2023, 32, 1–28. [CrossRef]
- Matthee, C.; Brown, A.R.; Lange, A.; Tyler, C.R. Factors Determining the Susceptibility of Fish to Effects of Human Pharmaceuticals. Environ. Sci. Technol. 2023, 57, 8845–8862. [CrossRef]
- Bojarski, B.; Kot, B.; Witeska, M. Antibacterials in Aquatic Environment and Their Toxicity to Fish. Pharmaceuticals 2020, 13, 189. [CrossRef]
- Almeida, A.R.; Tacão, M.; Machado, A.L.; Golovko, O.; Zlabek, V.; Domingues, I.; Henriques, I. LongTerm Effects of Oxytetracy1cline Exposure in Zebrafish: A Multi-Level Perspective. Chemosphere, 2019, 222, 333–344.
- Massarsky, A.; Kozal, J.S.; Di Giulio, R.T. Glutathione and zebrafish: Old assays to address a current issue. Chemosphere 2017, 168, 707–715. [CrossRef]
- Hu, F.; Dong, F.; Yin, L.; Wang, H.; Zheng, M.; Fu, S.; Zhang, W. Effects of sulfamethoxazole on the growth, oxidative stress and inflammatory response in the liver of juvenile Nile tilapia (Oreochromis niloticus). Aquaculture 2021, 543, 736935. [CrossRef]
- Plhalova, L.; Zivna, D.; Bartoskova, M.; Blahova, J.; Sevcikova, M.; Skoric, M.; Marsalek, P.; Stancova, V.; Svobodova, Z. The effects of subchronic exposure to ciprofloxacin on zebrafish (Danio rerio). Neuroendocrinol Lett 2014, 64–70.
- Salahinejad, A.; Meuthen, D.; Attaran, A.; Chivers, D.P.; Ferrari, M.C. Effects of common antiepileptic drugs on teleost fishes. Sci. Total. Environ. 2023, 866, 161324. [CrossRef]
- Tiehm, A.; Schmidt, N.; Stieber, M.; Sacher, F.; Wolf, L.; Hoetzl, H. Biodegradation of Pharmaceutical Compounds and their Occurrence in the Jordan Valley. Water Resour. Manag. 2010, 25, 1195–1203. [CrossRef]
- Chiţescu, C.; Ene, A.; Bahrim, G.E.; Enachi, E. Pharmaceutical residues monitoring in surface water in Romania. Status and concerns. Environ. Toxic. Freshwater Marine Ecosys. Black Sea Basin, 2020, 55-56.
- Kim, Y.; Choi, K.; Jung, J.; Park, S.; Kim, P.-G.; Park, J. Aquatic toxicity of acetaminophen, carbamazepine, cimetidine, diltiazem and six major sulfonamides, and their potential ecological risks in Korea. Environ. Int. 2007, 33, 370–375. [CrossRef]
- Da Silva Santos, N.; Oliveira, R.; Lisboa, C.A.; Mona e Pinto, J.; Sousa-Moura, D.; Camargo, N.S.; Perillo, V.; Oliveira, M.; Grisolia, C.K.; Domingues, I. Chronic effects of carbamazepine on zebrafish: Behavioral, reproductive and biochemical endpoints. Ecotoxicol. Environ. Saf. 2018, 164, 297–304. [CrossRef]
- Liang, X.; Csenki, Z.; Ivánovics, B.; Bock, I.; Csorbai, B.; Molnár, J.; Vásárhelyi, E.; Griffitts, J.; Ferincz, Á.; Urbányi, B.; et al. Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio). Antioxidants 2022, 11, 1136. [CrossRef]
- Li, Z.; H.; Li, P.; Rodina, M.; Randak, T. Effect of human pharmaceutical Carbamazepine on the quality parameters and oxidative stress in common carp (Cyprinus carpio L.) spermatozoa. Chemosphere, 2010, 80(5), 530-534.
- Gasca-Pérez, E.; Galar-Martínez, M.; García-Medina, S.; Pérez-Coyotl, I.A.; Ruiz-Lara, K.; Cano-Viveros, S.; Borja, R.P.-P.; Gómez-Oliván, L.M. Short-term exposure to carbamazepine causes oxidative stress on common carp (Cyprinus carpio). Environ. Toxicol. Pharmacol. 2018, 66, 96–103. [CrossRef]
- Moreira, D. G.; Aires, A.; de Lourdes Pereira, M.; Oliveira, M. Levels and effects of antidepressant drugs to aquatic organisms. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2022, 256, 109322.
- Singh, A.; Saidulu, D.; Gupta, A.K.; Kubsad, V. Occurrence and fate of antidepressants in the aquatic environment: Insights into toxicological effects on the aquatic life, analytical methods, and removal techniques. J. Environ. Chem. Eng. 2022, 10. [CrossRef]
- Orozco-Hernández, J.M.; Elizalde-Velázquez, G.A.; Gómez-Oliván, L.M.; Santamaría-González, G.O.; Rosales-Pérez, K.E.; García-Medina, S.; Galar-Martínez, M.; Juan-Reyes, N.S. Acute exposure to fluoxetine leads to oxidative stress and hematological disorder in Danio rerio adults. Sci. Total. Environ. 2023, 905, 167391. [CrossRef]
- Kosma, C.I.; Nannou, C.I.; Boti, V.I.; Albanis, T.A. Psychiatrists and selected metabolites in hospital and urban wastewaters: occurrence, removal, mass loading, seasonal influence and risk assessment. Sci. Total Environ. 2019, 659, 1473–1483.
- Christensen, A.M.; Markussen, B.; Baun, A.; Halling-Sørensen, B. Probabilistic environmental risk characterization of pharmaceuticals in sewage treatment plant discharges. Chemosphere 2009, 77, 351–358. [CrossRef]
- Weinberger, J.; Klaper, R. Environmental concentrations of the selective serotonin reuptake inhibitor fluoxetine impact specific behaviors involved in reproduction, feeding and predator avoidance in the fish Pimephales promelas (fathead minnow). Aquat. Toxicol. 2014, 151, 77–83. [CrossRef]
- Duarte, I.A.; Reis-Santos, P.; Novais, S.C.; Rato, L.D.; Lemos, M.F.; Freitas, A.; Pouca, A.S.V.; Barbosa, J.; Cabral, H.N.; Fonseca, V.F. Depressed, hypertense and sore: Long-term effects of fluoxetine, propranolol and diclofenac exposure in a top predator fish. Sci. Total. Environ. 2020, 712, 136564. [CrossRef]
- Pan, C.; Yang, M.; Xu, H.; Xu, B.; Jiang, L.; Wu, M. Tissue bioconcentration and effects of fluoxetine in zebrafish (Danio rerio) and red crucian cap (Carassius auratus) after short-term and long-term exposure. Chemosphere 2018, 205, 8–14. [CrossRef]
- Ding, J.; Lu, G.; Li, Y. Interactive effects of selected pharmaceutical mixtures on bioaccumulation and biochemical status in crucian carp (Carassius auratus). Chemosphere 2016, 148, 21–31. [CrossRef]
- Cunha, V.; Rodrigues, P.; Santos, M.; Moradas-Ferreira, P.; Ferreira, M. Danio rerio embryos on Prozac ⿿ Effects on the detoxification mechanism and embryo development. Aquat. Toxicol. 2016, 178, 182–189. [CrossRef]
- Duarte, I.A.; Reis-Santos, P.; Novais, S.C.; Rato, L.D.; Lemos, M.F.; Freitas, A.; Pouca, A.S.V.; Barbosa, J.; Cabral, H.N.; Fonseca, V.F. Depressed, hypertense and sore: Long-term effects of fluoxetine, propranolol and diclofenac exposure in a top predator fish. Sci. Total. Environ. 2020, 712, 136564. [CrossRef]
- de Oliveira, M.R. Fluoxetine and the mitochondria: a review of the toxicological aspects. Toxicol. Lett., 2016, 258, 185–191.
- Sathishkumar, P.; Meena, R.A.A.; Palanisami, T.; Ashokkumar, V.; Palvannan, T.; Gu, F.L. Occurrence, interactive effects and ecological risk of diclofenac in environmental compartments and biota - a review. Sci. Total. Environ. 2020, 698, 134057. [CrossRef]
- Jurado, A.; Labad, F.; Scheiber, L.; Criollo, R.; Nikolenko, O.; Pérez, S.; Ginebreda, A. Occurrence of pharmaceuticals and risk assessment in urban groundwater. Adv. Geosci. 2022, 59, 1–7. [CrossRef]
- Sharma, J.; Joshi, M.; Bhatnagar, A.; Chaurasia, A.K.; Nigam, S. Pharmaceutical residues: One of the significant problems in achieving ‘clean water for all’and its solution, Environ. Res., 2022, 215, 114219.
- Lushchak, V.I. Contaminant-induced oxidative stress in fish: a mechanistic approach. Fish Physiol. Biochem. 2015, 42, 711–747. [CrossRef]
- Stoliar, O.B.; Lushchak, V.I. Environmental pollution and oxidative stress in fish. Oxidative stress-environmental induction and dietary antioxidants, Environmental Induction and Dietary Antioxidants, 2012, 131-166.
- Nava-Álvarez, R.; Razo-Estrada, A.C.; García-Medina, S.; Gómez-Olivan, L.M.; Galar-Martínez, M. Oxidative Stress Induced by Mixture of Diclofenac and Acetaminophen on Common Carp (Cyprinus carpio). Water, Air, Soil Pollut. 2014, 225, 1–9. [CrossRef]
- Beghin, M.; Schmitz, M.; Betoulle, S.; Palluel, O.; Baekelandt, S.; Mandiki, S.N.; Gillet, E.; Nott, K.; Porcher, J.-M.; Robert, C.; et al. Integrated multi-biomarker responses of juvenile rainbow trout (Oncorhynchus mykiss) to an environmentally relevant pharmaceutical mixture. 2021, 221, 112454. [CrossRef]
Figure 1.
Meta-analysis of areas of interest in the literature on pharmaceutical impact on fish oxidative stress.
Figure 1.
Meta-analysis of areas of interest in the literature on pharmaceutical impact on fish oxidative stress.
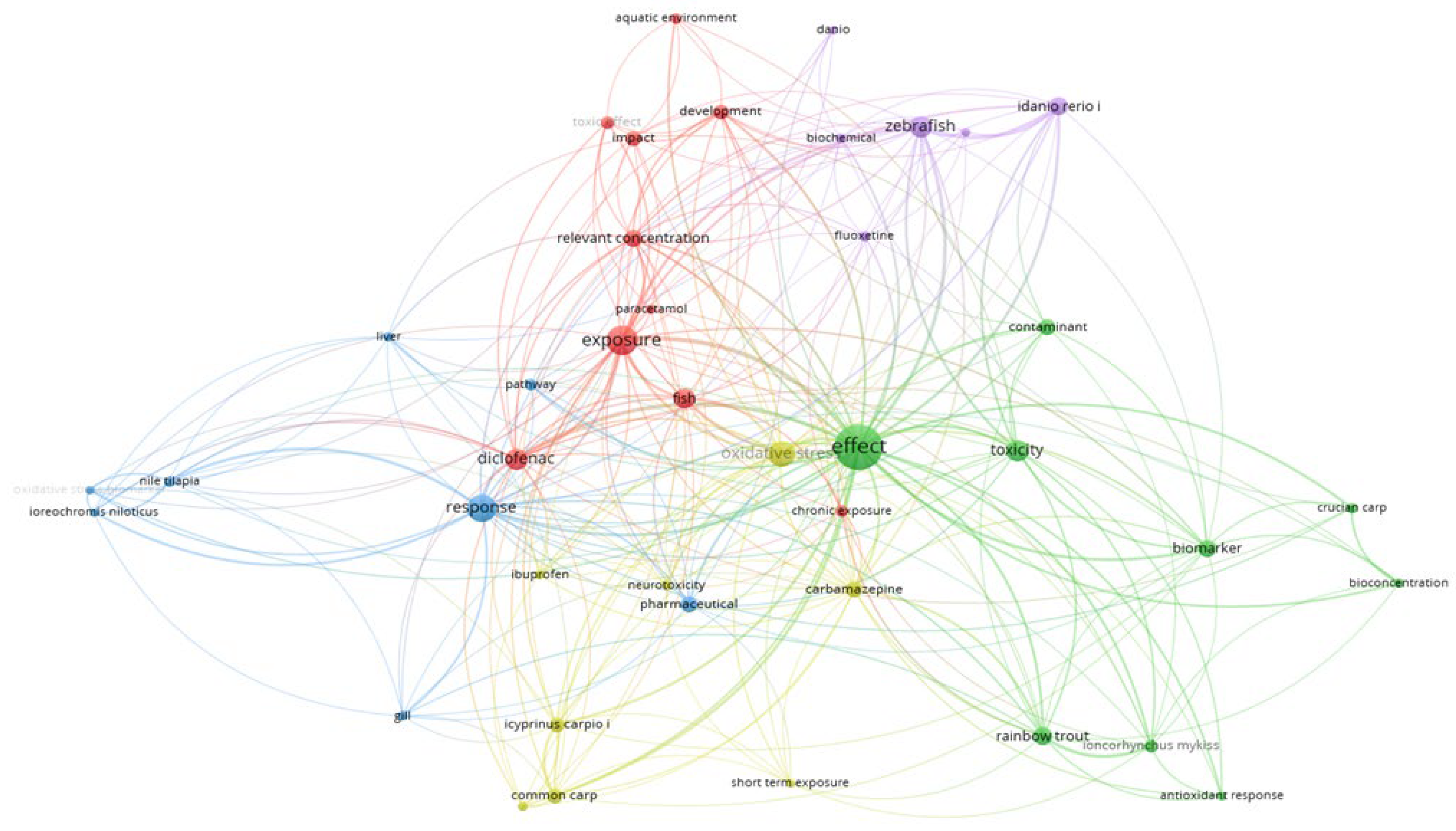
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



