
Submitted:
10 January 2025
Posted:
13 January 2025
You are already at the latest version
Abstract
Bovine viral diarrhea (BVD) is caused by bovine viral diarrhea virus (BVDV). The disease incurs $1.5-2.5 billion/year and $0.50 to $687.80/cow loss in beef and dairy farms respectively. This review aims to provide insights into the interaction of BVDV with host immunity, currently available vaccines, and strategies used to advance the vaccines. The virus causes immunosuppression by interfering with the innate and adaptive immune systems in a strain and biotypes-dependent manner. Interferon production, apoptosis, neutrophil activity, macrophages, and antigen-presenting cells are significantly affected during viral infection. Maternal antibodies (MatAb) are crucial to protect calves from early infection. However, the MatAb is counterproductive during vaccination against the virus. There are several types of licensed commercial inactivated or modified live vaccines, most of which are made of cytopathic BVDV 1 and 2 and the BVDV 1a subtype. Subunit and marker vaccines are made of E2, Erns, and NS3 proteins in combination with modern adjuvants, respectively. Such types of vaccines are not yet licensed and are in the experimental stage. The limitations of currently available conventional vaccines are minimal or lack of cross-protection, production costs, safety concerns, and inefficiency in provoking both humoral and cellular immune systems. To alleviate these limitations, the knowledge of developing next-generation vaccines using appropriate viral proteins and the use of modern adjuvants is promising.
Keywords:
BVDV
; BVD
; Immunity
; Maternal antibody
; Vaccines
1. Introduction
The disease Bovine viral diarrhea (BVD) globally affects over 80% of the cattle population and causes severe economic loss in cattle-producing countries [1,2]. BVD is characterized by diarrhea, respiratory disease, immunosuppression, abortion, congenital malformations, and the birth of persistently infected (PI) calves [3,4]. It is caused by bovine viral diarrhea virus (BVDV) of the genus Pestivirus, family Flaviviridae [5]. The virus is categorized into 3 species: BVDV 1, which has 21 sub-genotypes (1a-1u); BVDV 2, which has four sub-genotypes (2a-2d) [6]; and four Hobi-like (BVDV-3) species (3a-3d) [7]. Based on the cytopathic effect on cell lines, the virus is also classified into two biotypes: cytopathic (cp) and non-cytopathic (ncp) BVDV [8].
According to Hilleman[9], viruses employ two different strategies to remain allied with their hosts: 1) a 'hit-and-run' strategy characterized by the sequential spread of infection in a series of hosts; 2) a hit-stay or infect and persist strategy that enables the virus to remain in the individual host for a longer period, and occasionally or rarely transmit to subsequent hosts. During transient infection which is seen mostly in adult cattle, the virus follows the 'hit-and-run' strategy [10] whereas “hit-stay or infected and persist” is seen in persistently infected calves. Hit-and-stay viruses use strategies such as sequestration, blockade of antigen presentation, cytokine escape, evasion of natural killer cell activities, escape from apoptosis, and changing the antigenicity as means to evade the immune system of the host [9].
The innate immune system acts as the first line of defense during viral infections, aiming to prevent viral invasion or replication before the adaptive immune system generates more specific protection [11,12]. BVDV impairs both the adaptive and innate immune systems in a strain-dependent manner [8,13,14,15]. It is suggested that BVDV can maintain prolonged infection in immune-privileged sites such as the ovaries, testes, brain, and circulating white blood cells [16] .
Depending on the level of risk and livestock density, systematic control strategies like strict biosecurity, elimination of PI animals, and surveillance in combination with vaccination become useful in the prevention and control of the disease [17,18,19].
The limitations in the existing conventional vaccines include variation in vaccinal and field strain, limited cross-protection, safety, immunosuppression, reversion to virulence, induction of neonatal pancytopenia, production cost, repeated injection and neutralization by MatAb [20,21,22]. Brackenbury et al. [23]suggested that designing and developing an effective vaccine against BVDV infection requires a detailed understanding of the immune response to the virus. Therefore, this review aims to provide insight into the effect of BVDV on the host's immune system, commercially available vaccines, and existing strategies to develop next-generation vaccines
2. Pathogenesis
Smirnova et al. [24]showed that the vertical transmission of the virus to the fetus is determined by the complex interaction between the fetoplacental and maternal immune systems, as well as the developmental stage at which the fetus becomes infected. As shown in Figure 1 infection with the ncp BVDV biotype during the early stage of pregnancy (40-125 days) leads to the production of persistently infected (PI) calves [25]. This is because, at this stage of pregnancy the fetus lacks a fully developed immune system to defend the virus [25,26].
3. BVDV and the Innate Immune System
3.1. Interferon α/β
The induction of interferon α/β is the first line of defense against acute viral infection. It has a key role in linking the innate and adaptive immune systems [27]. This multifaceted cytokine has different effects such as inhibition of viral replication, induction of NK cells and Macrophages, increased expression of MHC I, and activation of T-cells [28].
A pathogen-infected cell produces an interferon type that binds to the receptors of infected and uninfected cells to induce an antiviral effect [29]. Type 1 interferons activate the synthesis of more than 100 cellular proteins [27]. The long-term co-evolution of the host and pathogens allows many viruses to evade the antiviral state of the interferon α/β system [27,30,31]. However, some viruses prevent the action of interferon by inhibiting the binding to receptors after production, hindering the signal transduction after binding to receptors, or preventing the antiviral activity of IFN-induced proteins [29,31,32].
Virus infection activates the transcription of various cellular genes [33] either directly through activation of cellular transcription factors such as IRF-3 or indirectly through prior induction of IFN-α/β. The ssRNA of both biotypes (ncp and cp) of BVDV block induction of apoptosis and alpha/beta interferon (IFN-α/β) or Mx protein synthesis [34]. Resistance to interferon (IFN-α/β) depends on the biotype of the BVDV virus infecting the host. Moreover, in persistently infected (PI) animals, the strain variation also aids the virus in escaping from the adaptive immune system, i.e. the adaptive immune system directed to one strain does not interfere with the activity of another strain "non-self" that may replicate in cells infected with ncp BVDV [29].
The success of persistent infection or immunotolerance depends on the highly selective "self" and "non-self" model of evasion of the interferon defense system by BVDV during the early stage of fetal development (40-125 days) [35,36]. Daniel et al. [37]suggested that the greater amount of ssRNA produced by cp BVDV has been postulated to stimulate IFN production via RIG-I– and TLR–7/8 mediated activation of IRF-3(Figure 2).
The N-terminal fragment protein of noncytopathic bovine viral diarrhea virus (ncpBVDV), Npro, inhibits the activity of interferon regulatory factor 3 (IRF3) and interferes with the production of IFN-α/β [32,41,42,43] (Figure 2 and Figure 3). During early infection with ncpBVDV, IRF 3 is relocated from the cytoplasm to the nucleus. However, IRF 3 is blocked from binding to interferon gene promoter in the nucleus. Subsequently, inhibiting the transcription of the IFN-α/β gene and subsequently blocking interferon synthesis [43]. The Npro results in the polyubiquitination and subsequent destruction of the regulatory factor by cellular multicatalytic proteasomes in the cytoplasm [41,43]. NCP BVDV can also induce complete mitophagy and effectively inhibit type I IFN expression, inflammatory cytokines expression, and apoptosis [44].
The Npro and Erns antagonize IFN1 production in infected and non-infected cells, respectively [42,45]. As a consequence, the innate immune system of the host is inhibited [32,46]. The interference with type I IFN signaling has been implicated as a contributing factor in the establishment of PI calves [36,45,47]. Interestingly, non-cytopathic BVDV fails to induce interferon type I in cultured bovine macrophages; whereas, cytopathic biotypes readily trigger this response [10,48].
Cells infected with non-cytopathic BVDV are also resistant to the induction of interferon by double-stranded RNA, a potent interferon inducer signaling the presence of viral replication in the cell [10]. The expression of interferon-stimulating genes (ISGs) in the endometrium is upregulated by IFN-τ, which is crucial for maintaining pregnancy in ruminants. The expression of interferon-stimulating genes (ISGs) in the endometrium is upregulated by IFN-τ, which is crucial for maintaining pregnancy in ruminants. However, the expression of ISGs is down-regulated by acute infection with ncp BVDV. As a result, the virus persists in the host without being effectively cleared by the immune system [29,49]. IFN-tau (IFN-τ) is known to have antiviral activity similar to the other type I IFNs [50], and it may well act to prevent infection of the embryo [10].
3.2. BVDV Triggers Apoptosis
Grummer et al.[51] reported that cpBVDV affects membrane potential (Dcm), translocates cytochrome c to the cytosol, increases caspase-9 activity, and leads to overexpression of apoptotic protease-activating factor 1; hence, the virus triggers the intrinsic pathway of apoptosis. Endoproteases caspase-3 and caspase-9 were significantly higher in mucosal disease (MD). In contrast, the expression of caspase-8 was not elevated in both MD cases and control groups Hilbe et al. [52]. The authors concluded that the intrinsic apoptosis pathway is more critical in the pathogenesis of MD in cattle than the extrinsic pathway. It should be noted that in cells infected with cpBVDV, dsRNA is the main inducer of apoptosis.
3.3. BVDV and Innate Immune Cells
The innate immune response is critical for the body's defense against pathogens and is induced through the interaction with different receptors, including danger-associated molecular patterns (DAMPs) [54]. The complement system, natural killer cells, acute-phase protein response, interferon-alpha, and beta (IFN-α/β) are the major responses by the innate immune system [55]. Neutrophils, among the white blood cells, play a critical role in the innate immune response [56].
Activated innate immune cell, such as neutrophils, monocytes, macrophages, or endothelial cells, expresses S100A9, a member of the Damage Associated Molecular Patterns (DAMPs) protein family, which initiates its effect through the TLR4/MyD88 pathway [54]. The Npro protein of BVDV binds to S100A9 as a result suppresses the host's innate immunity system. The replication of the virus in the infected cell is boosted due to the inhibition of the S100A9 protein by small interfering RNA (siRNA) [54]. The impaired innate immune system due to infection with BVDV predisposes animals to concurrent infection [45].
3.3.1. Neutrophils
The release of different mediators by neutrophils, such as interleukin-8 (IL-8), platelet-activating factor (PAF), leukotriene B4 (LTB4), or complement fragment 5a (C5a), activates the innate immune response during infection [57,58,59,60]. The oxidative burst and neutrophil extracellular activity, as well as the expression of CD18 and L-selectin on neutrophils, are significantly decreased in association with infection with BVDV biotypes [61,62] (Figure 4). The phagocytic activity of neutrophils is reduced according to the biotype of the BVDV; cpBVDV significantly reduces phagocytosis, while ncpBVDV does not [62,63]. Similarly, the expression of CD14 on neutrophils is enhanced by ncpBVDV compared to cpBVDV biotype. Furthermore, the cp BVDV biotype does not have any effect on the chemotactic activity of neutrophils [62].
3.3.2. Macrophages
The innate immunity is mainly triggered by Macrophages which also play a key role during viral infection [66]. The function of macrophages counteract indirectly in a biotype-dependent manner [66]. Macrophages infected with ncpBVDV fail to induce type I IFN during persistent infection, whereas in mucosal disease infected with cpBVDV do release type I IFN [67,68]. Bovine bone marrow-derived macrophages infected with BVDV decreased production of TNF-α, indicating suppression of proinflammatory cytokine production [69].
The conserved motifs of microorganisms called pathogen-associated molecular patterns (PAMPs) are recognized by pattern recognition receptors (PRRs) on the surface of macrophages [70]. Subsequently, induce pro-inflammatory and anti-viral immune responses [71]. Viral and bacterial infections are recognized by Toll-like receptors (TLRs), which are a class of PRRs conserved across mammalian species. However, disease-causing agents have developed similar mechanisms to circumvent recognition by these receptors, leading to infection by the pathogen [72,73,74].
Macrophages produce microbicidal molecules like nitric oxide (NO) and cytokines (e.g., Interleukin 1 β), which attract inflammatory cells from the bloodstream to the site of infection during the inflammatory response [75]. An in vitro study by Adler et al.[76] showed that both cp and ncpBVDV biotypes downregulate the production of tumor necrosis factor-alpha (TNFα), a highly active and pleiotropic cytokine mainly produced by macrophages [77]. It modulates the production of various types of cytokines during the immune response [78].
Monocytes/macrophages are highly susceptible to BVDV infection among different populations of leukocytes [14]. BVDV infection causes immunosuppression and alters the signaling pathway of pathogen recognition receptors (PRRs) and Toll-like receptors (TLRs) in macrophages of innate cells (Figure 5). The study by Schaut et al. [71] showed there is variation in the expression of pro-inflammatory cytokines between BVDV-2 infected and non-infected macrophages in a strain-dependent manner. Generally, BVDV-2-infected macrophages are known to impair the production of pro-inflammatory cytokines in response to TLR ligation, except for TLR7 [71]. Moreover, the author indicated that there are distinct differences in TLR pathways modulated following BVDV-2 infection, which have implications for macrophage responses to secondary infections [71]. Non-cytopathic BVDV infection stimulates cytokines from macrophages in vitro but the effect of BVDV infection in vivo on macrophages or in vitro with monocytes is not clear [36].
Bovine viral diarrhea infection increases the production of nitric oxide while decreasing the production of TNFα by macrophages [76]. Moreover, the expression of Fc and C3 receptors and phagocytic activity of alveolar macrophages become decreased due to the effect of BVDV on the immune cells [79]. Lee et al.[80] suggest that cp BVDV interferes with the antigen presentation to immune-competent T cells while monocyte activation and differentiation are enhanced. As a result, macrophage-mediated uncontrolled inflammation increases viral spread and impedes the host antiviral defense mechanism.
3.3.3. Antigen-Presenting (APC)
Professional antigen-presenting cells (APCs) like dendritic cells (DCs) and their myeloid progenitors, monocytes/macrophages- play a significant role in both the innate and adaptive immune systems of the host [81]. In bovine monocytes (professional APCs), the expression of MHC-I and MHC-II molecules is relatively higher [82]. However, the cpBVDV biotype reduces the ability of monocytes to phagocytize antigens, and stimulate allogeneic and memory CD4+ T-cell responses [36,75,80,83]. Lee et al. [80] indicated that proteins associated with immune responses, such as cell adhesion, apoptosis, antigen uptake, processing and presentation, acute-phase response proteins, MHC class I- and II-related proteins, and other molecules involved in the immune function of professional antigen presentation, are critically affected during BVDV infection (Figure 6).
The comparison made by Rajput et al. [84] on ncp and cpBVDV biotypes revealed that the expression of surface markers is crucial for immune response activation. MHC I, MHC II, and CD86 expression on monocyte-derived dendritic cells (Mo-DC) were decreased by ncpBVDV and increased in cells infected with cpBVDV biotypes. As a result, viruses that infect DCs can have a detrimental impact on the immune system [84]. In contrast to this finding, Glew et al. [75]suggested that BVDV does not affect DCs' ability to present antigens to T cells in vitro and instead induces apoptosis in these cells. Although BVDV infection does not affect DCs, the virus exploits these cells to spread from the site of infection to other parts of the body. Moreover, DC infection is crucial for triggering the immune response [85].
3.4. Adaptive Immune System
BVDV causes immunosuppression by interfering the adaptive immune response, which is the host's second line of defense against invading pathogens [36]. B- and T-cell immunotolerance retained by animals persistently infected with BVDV in a strain-specific manner [45]. In both cp and ncpBVDV biotypes, the major envelope proteins Erns and E2, as well as the non-structural protein NS2-3, are the primary activators of CD4+ T cells in inducing protective immunity against the virus [86,87,88]. However, a response from short-term T-cell lines indicated that the capsid (C) and/or the amino-terminal proteinase (Npro) were identified as T-cell determinants [89].
After seven days of infection with cpBVDV, the amount of antibody produced by B-cells decreases compared to ncpBVDV. The ncpBVDV biotype induces high levels of IgG2 antibody isotype due to immune response polarization towards the Th1 response [36,90]. This demonstrates that acute infection caused by ncpBVDV is alleviated through cell-mediated and humoral immunity [89,91]. However, infection with the virus impairs the cytotoxic T lymphocyte response [36]. The adaptive immune system for PI animals is normal except for the strain of the virus to which they are immunotolerant. The level of gamma delta T cells may be related to mucosal disease [36].
Donis [92] stated that the antibody produced against E2 is more effective than Erns in protecting the host from BVDV infection. Although Erns and NS3 elicit strong antibodies, the antibodies cannot neutralize the virus. Therefore, these proteins are good candidates for developing epitope knock-out vaccines or DIVA kits [93]. B-cells do not respond to the major structural protein (C) and E1 protein, resulting in no production of neutralizing antibodies against the virus [92]. According to Peterhans and Schweizer [45], ncpBVDV induces the production of both homologous and heterologous neutralizing antibodies more effectively than cpBVDV infection. However, cpBVDV is better at activating the cell-mediated immune response [94] . The production of IgG1 isotype antibodies is significantly suppressed during BVDV infection [90]. Depending on the viral strains and biotypes of the virus the adaptive immune system and maturity of antigen-presenting cells are critically affected [49].
3.5. The Role of Maternal Antibody(MatAb) in Protecting Calves Against BVDV
Acquired MatAb can protect calves from BVDV infection for a limited time, but interference by high titers of MatAb prevents antibody response following vaccination [95,96,97,98]. The duration of protection provided by MatAb varies depending on the level of MatAb present in the calf and the strain of BVDV infecting the host [95,96]. The decrease in the MatAb titer makes calves more prone to viral infection at an early growth stage. Passive antibodies also help to control the spread of viral infection in a group of calves and the accumulation of the virus in the herd by preventing viral shedding from infected or diseased animals the virus [96].
Downey et al.[97] indicated in their research that the MatAb titer level should reach 3.12 titers for the BVDV vaccine to provoke an immune response in vaccinated calves. However, the age at which the calf reaches this antibody threshold depends on the dam's age and the quantity of passively transferred antibodies. The antibodies may start decaying as early as 2 days after consuming colostrum [98]. Depending on the success of the passive transfer, MatAb for BVDV type 1 and 2 have been shown to persist for 210 and 180 days, respectively [99].
3.6. BVDV Vaccines and Application
3.6.1. Conventional Vaccines
Bovine respiratory disease complex (BRD) is a multifactorial infectious disease and BVDV is among the major pathogens that cause the disease. To prevent the clinical disease and dissemination of the virus, vaccinating animals in dairy and beef farms is needed [103]. A polyvalent commercial vaccine containing major pathogens including BVDV has been used for several years to prevent the disease [104,105]. Over the past 30 years, conventional vaccines for BVDV (inactivated or attenuated) have been developed [106] . Particularly, in the case of BVDV, the viral genetic variation, existence of persistent infection, viral tropism for immune cells, limited cross-protection, and safety and efficacy of vaccines present challenges to the disease prevention and control strategy [107,108,109,110,111].
BVDV vaccines have been available since the 1960s and studies conducted under controlled conditions have shown vaccines to be efficacious in reducing disease and viral transmission [112]. Various types of licensed BVDV vaccines for cattle use are currently available (Table 1). Most of these vaccines are multivalent, containing BHV-1, BVDV, PI-3V, and BRSV viral antigens in combination with multiple viral components and bacterial vaccines/bacterins/toxoids. The vaccines can be modified live vaccines (MLV), inactivated, or chemically altered live vaccines [113]. These commercially available vaccines provide efficient fetal protection, even with prolonged exposure to BVDV [19].
The efficacy of licensed vaccines in protecting the dam and fetus ranges from 91-100% under experimental conditions [110,114,115,116,117]. However, the efficacy of the vaccines is critically determined by the duration of exposure to PI animals under field conditions [118]. In a study by Grooms et al. [119], using two doses of inactivated vaccine, a 73% protection level for the fetus was reported following 98-day exposure to wild BVDV viral strains.
MLV vaccines provide higher protection and a longer-lasting immune response compared to killed vaccines [120,121]. In contrast, inactivated or killed vaccines have low efficiency in protecting the fetus, and require higher production costs, and higher doses of the antigen during vaccination [122]. Due to safety concerns associated with modified live vaccines, farmers prefer to vaccinate their animals with killed vaccines, especially during the breeding period [123].
An efficient vaccine candidate should provide cross-protection against the common circulating subtypes of BVDV-1 and 2 [124]. Most inactivated vaccines induce antibody production and MHC II-restricted helper and/or cytotoxic T lymphocytes (CTL). Nevertheless, they lack activation of CD8+ MHC I-restricted CTL which plays a significant role during viral infection [125]. The strategy to vaccinate against BVDV is to prevent the vertical transmission of the virus from the dam to the fetus during an early stage of pregnancy to prevent the production of PI calves [126].
BVDV conventional vaccines (inactivated and modified live vaccines) contain BVDV 1 and 2 strains [105]. Platt et al. [127] revealed that a pentavalent MLV containing BVDV 1 and 2 strains induces T cell responses (CD4+ and CD8+) in calves aged 1-8 weeks with maternal antibodies. MLV (modified live virus) vaccines have been found to stimulate specific T-cells even in the presence of maternal antibodies [127]. On the other hand, inactivated BVD vaccines elicit a robust humoral response and activate subsets of cell-mediated immunity, including the T helper 1 response, gd TCR+ T cell response, and CD8+ γδTCR+ T cell response. However, they do not induce a detectable CD8+γδTCR- (cytotoxic T cell) level during an immune response [128].
Recent studies indicated that modified live vaccines (MLV) offer better protection than killed vaccines against BVDV. Though MLV is easily neutralized by a higher level of MatAb [22], studies showed that certain MLV vaccines like Bovela® live double deleted vaccine could be used during high MatAb titer levels in calves [129,130].
3.6.2. Next Generation Vaccines
The latest advanced technologies are utilized to produce BVDV subunit and marker vaccines using the virus's structural and non-structural proteins. The immunodominant structures of BVDV are E2, Erns, and NS3 proteins mainly used to develop the vaccines. The E2 protein, which triggers the production of antigen-neutralizing antibodies, is commonly employed in the design of subunit and DNA vaccines [131,132,133]. Conversely, the non-neutralizing inducer Erns and NS3 are used to develop marker vaccines (Table 2).
Nelson et al. [134] developed plant-based recombinant tE2 vaccines that induce the immune systems in guinea pigs against BVDV.Gómez-Romero et al. (2023) induced a strong immune response against BVDV using a combination of rE2 glycoprotein. Carlsson et al. [136] developed an immunostimulating complex (ISCOMs) subunit vaccine based on the E2 protein that can induce high viral neutralizing titers and protect fetuses against transplacental infection with BVDV. Takahashi et al. (1990) indicated that ISCOMs vaccines can elicit both cell-mediated and humoral responses against BVDV. However, the research on ISCOMs-based subunit vaccines highlights the potential of the vaccine to prevent BVDV at the experiment level.
E2-based subunit vaccines provide significant protection against BVDV 2; therefore, this structural protein is a better candidate for developing engineered vaccines [143]. Pecora et al. [135] developed a truncated E2 (tE2) vaccine for BVDV 1b and 2a by binding the tE2 version to a single-chain fragment variable (scFV) called APCH to produce APCH-tE2-1a-1b-2a, which is safe and induces a rapid and sustained neutralizing antibody response compared to a conventional vaccine in cattle [145]. APCH-tE2, developed using alfalfa transgenic plants, provides complete protection against BVDV [146]. The efficacy of E2 subunit vaccines to protect against BVDV can be enhanced by using adjuvants in combination with the vaccine [147,148].
The E2 protein of BVDV was expressed on Lactobacillus and vaccine pPG-E2_ctxB/Lc W56 conjugated with cholera toxin B subunit (ctxB) as an adjuvant. This vaccine is immunogenic and induces effective mucosal, humoral, and cellular immune responses against BVDV in mice [149]. Sangewar et al. [21]developed a vaccine called Expi293TM expressing antigens (293F-E2123, 293F-NS2-31, and 293F-NS2-32) that elicits strong and cross-reactive IFN-γ responses and neutralizing antibodies specific to BVDV. A multivalent E2 subunit vaccine, generated from BVDV strains in sub-genotypes 1a, 1b, and 2, protects sheep from heterogenous BVDV strains [137]. However, the expression vector used and the dose of the vaccines determine the potency of the vaccines, as discussed by Thomas et al. [148].
The chimeric E2 and flag-tagged Erns, fused with the granulocyte monocyte colony-stimulating factor (GM-CSF) subunit vaccines, are expressed on a mutated BoHV-1 vaccine vector. The vaccine triggers the production of cross-reactive neutralizing antibodies and a cellular immune response [138]. The Pestivirus Erns gene originated from the pronghorn antelope used to develop a chimeric NADL/P-Erns marker vaccine and applied during the BVDV control and eradication program [139].
The potency of the DNA vaccine for BVDV (NTC E2t(co) and NTC NS3(co) in cattle is augmented by the co-expression of the Retinoic acid-inducible gene I (RIG-I) agonist with the viral antigen [140]. Koethe et al. [141] developed synthetic chimera vaccines BVDV-1b_synCP7_∆Npro_Erns and Bungoand BVDV-1b_synCP7_∆Npro_Erns Bungo_E1E2BVDV-2 CS, which are safe to use in cattle and provide efficient protection against BVDV. Moreover, these vaccines also serve as marker vaccines. Three novel mosaic polypeptide chimeras NproE2123, NS231, and NS232, were designed using Adenovirus as a vaccine vector. This induces higher IFN-γ spot-forming cells, T-cell proliferation, and higher specific antibodies against the BVDV-1 strain [22].
Despite the existence of several subunit vaccines for BVDV, limitations such as minimal or lack of activation of both humoral and cellular immune responses and toxicity associated with adjuvants hinder the production and use of the vaccines [150,151]. Although the safety of using DNA and subunit vaccines in animals is questionable, it still opens the door for the introduction of next-generation vaccines to the vaccine market [152]. The use of modern adjuvants, such as silica vesicles and hollow-type mesoporous silica nanoparticles (HMSA), as nanocarriers to load E2 protein enhances the production of cellular and humoral immunity against BVDV [151]. Furthermore, the fusion vaccine made of E2s and the complement C3d (E2s-C3d) provides sufficient protection against BVDV [152].
4. Conclusions
Bovine viral diarrhea significantly affects the financial return from cattle production by increasing morbidity and mortality in dairy and beef farms. The prevention and control of the disease depend on a combination of biosecurity, vaccines, and the detection and elimination of persistently infected calves from the herd. The virus causes immunosuppression by impeding the innate and adaptive immune responses in a strain-dependent manner. Currently, there are numerous commercially available vaccines, including monovalent or polyvalent, modified or killed, and subunit vaccines. The main drawbacks of these vaccines are the lack of or minimum cross-protection, high production costs (for killed and subunit vaccines), safety concerns (for MLV vaccines), and weak activation of humoral and cellular immune responses. Researchers are currently developing subunit vaccines and immunomodulators, such as modern adjuvants, to overcome these limitations. However, most next-generation vaccines are still in experimental trials and are not yet commercially available for BVDV prevention and control.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated and analyzed during this study are included as tables in the manuscript. However, the raw data are available from the corresponding author upon reasonable request.
Acknowledgments
This research work was made possible by the support of the VLIR-UOS Global Minds program and the members of the Host‒pathogen interactions and the staff at the National Animal Health Institute. The authors appreciate the support staff at the Animal Health Institute in terms of sample collection and laboratory analysis. We also thank Professor Teshal Sori and Dr. Hassen Chaka for reviewing this manuscript.
Conflicts of Interest
There was no competing interest.
References
- Fulton RW, Briggs RE, Ridpath JF, Saliki JT, Confer AW, Payton ME, et al. Transmission of Bovine viral diarrhea virus 1b to susceptible and vaccinated calves by exposure to persistently infected calves. Can J Vet Res. 2005, 69, 161–9.
- Khodakaram-Tafti, A.; Farjanikish, G.H. Persistent bovine viral diarrhea virus (BVDV) infection in cattle herds. Iran J Vet Res. 2017, 18, 154–63. [Google Scholar] [PubMed]
- Deregt, D.; Loewen, K.G. Bovine viral diarrhea virus: biotypes and disease. Can Vet J. 1995, 36, 371–8. [Google Scholar]
- Brock, K.V. The many faces of bovine viral diarrhea virus. Vet Clin North Am - Food Anim Pract. 2004, 20, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Smith DB, Meyers G, Bukh J, Gould EA, Monath T, Muerhoff AS, et al. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J Gen Virol. 2017, 98, 2106–12. [CrossRef] [PubMed]
- Yeşilbağ, K.; Alpay, G.; Becher, P. Variability and global distribution of subgenotypes of bovine viral diarrhea virus. Viruses. 2017;9.
- Liu, L.; Xia, H.; Wahlberg, N.; Belák, S.; Baule, C. Phylogeny, classification and evolutionary insights into pestiviruses. Virology. 2009, 385, 351–7. [Google Scholar] [CrossRef] [PubMed]
- Ammari, M.; McCarthy, F.M.; Nanduri, B.; Pinchuk, L.M. Analysis of Bovine Viral Diarrhea Viruses-infected monocytes: identification of cytopathic and non-cytopathic biotype differences. BMC Bioinformatics. 2010, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hilleman, M.R. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections. Proc Natl Acad Sci U S A. 2004;101 SUPPL. 2:14560–6.
- Peterhans E, Jungi TW, Schweizer M. BVDV and innate immunity. Biologicals. 2003, 31, 107–12.
- Koyama, S.; Ishii, K.J.; Coban, C.; Akira, S. Innate immune response to viral infection. Cytokine. 2008, 43, 336–41. [Google Scholar] [CrossRef] [PubMed]
- Bouyssi-Kobar M, De Asis-Cruz J, Murnick J, Chang T, Limperopoulos C. 乳鼠心肌提取 HHS Public Access. J Pediatr. 2019, 227, 13–2.
- Alcami, A.; Koszinowski, U.H. Viral mechanisms of immune evasion. Trends Microbiol. 2000, 8, 410–8. [Google Scholar] [CrossRef]
- Sopp, P.; Hooper, L.B.; Clarke, M.C.; Howard, C.J.; Brownlie, J. Detection of bovine viral diarrhoea virus p80 protein in subpopulations of bovine leukocytes. J Gen Virol. 1994, 75, 1189–94. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L.; Elmowalid, G.; Yousif, A.A.A. The immune response to bovine viral diarrhea virus: A constantly changing picture. Vet Clin North Am - Food Anim Pract. 2004, 20, 95–114. [Google Scholar] [CrossRef]
- Givens, M.D.; Marley, M.S. Immunology of chronic BVDV infections. Biologicals. 2013, 41, 26–30. [Google Scholar] [CrossRef]
- Baker, J.C. The clinical manifestations of bovine viral diarrhea infection. Vet Clin North Am Food Anim Pract. 1995, 11, 425–45. [Google Scholar] [CrossRef]
- Moennig V, Eicken K, Flebbe U, Frey HR, Grummer B, Haas L, et al. Implementation of two-step vaccination in the control of bovine viral diarrhoea (BVD). Prev Vet Med. 2005, 72, 109–14.
- Rodning SP, Marley MSD, Zhang Y, Eason AB, Nunley CL, Walz PH, et al. Comparison of three commercial vaccines for preventing persistent infection with bovine viral diarrhea virus. Theriogenology. 2010, 73, 1154–63.
- Falkenberg, S.M.; Dassanayake, R.P.; Terhaar, B.; Ridpath, J.F.; Neill, J.D.; Roth, J.A. Evaluation of Antigenic Comparisons Among BVDV Isolates as it Relates to Humoral and Cell Mediated Responses. Front Vet Sci. 2021;8 June:1–13.
- Sangewar N, Hassan W, Lokhandwala S, Bray J, Reith R, Markland M, et al. Mosaic Bovine Viral Diarrhea Virus Antigens Elicit Cross-Protective Immunity in Calves. Front Immunol. 2020;11 November:1–15.
- Lokhandwala S, Fang X, Waghela SD, Bray J, Njongmeta LM, Herring A, et al. Priming cross-protective bovine viral diarrhea virus-specific immunity using live-vectored mosaic antigens. PLoS One. 2017, 12, 1–23.
- Brackenbury, L.S.; Carr, B.V.; Charleston, B. Aspects of the innate and adaptive immune responses to acute infections with BVDV. Vet Microbiol. 2003, 96, 337–44. [Google Scholar] [CrossRef]
- Smirnova NP, Webb BT, Bielefeldt-Ohmann H, Van Campen H, Antoniazzi AQ, Morarie SE, et al. Development of fetal and placental innate immune responses during establishment of persistent infection with bovine viral diarrhea virus. Virus Res. 2012, 167, 329–36.
- Brock, K.V. The persistence of bovine viral diarrhea virus. Biologicals. 2003, 31, 133–5. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Zanzi, C.A.; Hietala, S.K.; Thurmond, M.C.; Johnson, W.O. Quantification, risk factors, and health impact of natural congenital infection with bovine viral diarrhea virus in dairy calves. Am J Vet Res. 2003, 64, 358–65. [Google Scholar] [CrossRef] [PubMed]
- Katze, M.G.; He, Y.; Gale, M. Viruses and interferon: A fight for supremacy. Nat Rev Immunol. 2002, 2, 675–87. [Google Scholar] [CrossRef] [PubMed]
- Bencze, D.; Fekete, T.; Pázmándi, K. Type i interferon production of plasmacytoid dendritic cells under control. Int J Mol Sci. 2021, 22, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Schweizer M, Mätzener P, Pfaffen G, Stalder H, Peterhans E. “Self” and “Nonself” Manipulation of Interferon Defense during Persistent Infection: Bovine Viral Diarrhea Virus Resists Alpha/Beta Interferon without Blocking Antiviral Activity against Unrelated Viruses Replicating in Its Host Cells. J Virol. 2006, 80, 6926–35.
- Bonjardim, C.A. Interferons (IFNs) are key cytokines in both innate and adaptive antiviral immune responses - And viruses counteract IFN action. Microbes Infect. 2005, 7, 569–78. [Google Scholar] [CrossRef] [PubMed]
- García-Sastre, A. Mechanisms of inhibition of the host interferon α/β-mediated antiviral responses by viruses. Microbes Infect. 2002, 4, 647–55. [Google Scholar] [CrossRef] [PubMed]
- Gil LHVG, Ansari IH, Vassilev V, Liang D, Lai VCH, Zhong W, et al. The Amino-Terminal Domain of Bovine Viral Diarrhea Virus N pro Protein Is Necessary for Alpha/Beta Interferon Antagonism. J Virol. 2006, 80, 900–11.
- Chang, Y.E.; Laimins, L.A. Microarray Analysis Identifies Interferon-Inducible Genes and Stat-1 as Major Transcriptional Targets of Human Papillomavirus Type 31. J Virol. 2000, 74, 4174–82. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L.; Thakur, N.; Darweesh, M.F.; Morarie-Kane, S.E.; Rajput, M.K. Immune response to bovine viral diarrhea virus - Looking at newly defined targets. Anim Heal Res Rev. 2015, 16, 4–14. [Google Scholar] [CrossRef]
- Schweizer, M.; Peterhans, E. Pestiviruses. Annu Rev Anim Biosci. 2014, 2, 141–63. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.C.L. The impact of BVDV infection on adaptive immunity. Biologicals. 2013, 41, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Daniel Givens M, Marley MSD, Jones CA, Ensley DT, Galik PK, Zhang Y, et al. Protective effects against abortion and fetal infection following exposure to bovine viral diarrhea virus and bovine herpesvirus 1 during pregnancy in beef heifers that received two doses of a multivalent modified-live virus vaccine prior to breeding. J Am Vet Med Assoc. 2012, 241, 484–95.
- Funami, K.; Matsumoto, M.; Oshiumi, H.; Akazawa, T.; Yamamoto, A.; Seya, T. The cytoplasmic “linker region” in Toll-like receptor 3 controls receptor localization and signaling. Int Immunol. 2004, 16, 1143–54. [Google Scholar] [CrossRef] [PubMed]
- Funami, K.; Sasai, M.; Ohba, Y.; Oshiumi, H.; Seya, T.; Matsumoto, M. Spatiotemporal Mobilization of Toll/IL-1 Receptor Domain-Containing Adaptor Molecule-1 in Response to dsRNA. J Immunol. 2007, 179, 6867–72. [Google Scholar] [CrossRef] [PubMed]
- Chen 2021. Chen, 2021.pdf. 2021, 22, 609–32.
- Chen Z, Rijnbrand R, Jangra RK, Devaraj SG, Qu L, Ma Y, et al. Ubiquitination and proteasomal degradation of interferon regulatory factor-3 induced by Npro from a cytopathic bovine viral diarrhea virus. Virology. 2007, 366, 277–92.
- Hong, T.; Yang, Y.; Wang, P.; Zhu, G.; Zhu, C. Pestiviruses infection: Interferon-virus mutual regulation. Front Cell Infect Microbiol. 2023;13 March:1–13.
- Hilton L, Moganeradj K, Zhang G, Chen Y-H, Randall RE, McCauley JW, et al. The NPro Product of Bovine Viral Diarrhea Virus Inhibits DNA Binding by Interferon Regulatory Factor 3 and Targets It for Proteasomal Degradation. J Virol. 2006, 80, 11723–32.
- Li Z, Zhang Y, Zhao B, Xue Q, Wang C, Wan S, et al. Non-cytopathic bovine viral diarrhea virus (BVDV) inhibits innate immune responses via induction of mitophagy. Vet Res. 2024, 55, 27.
- Peterhans E, Schweizer M. BVDV: A pestivirus inducing tolerance of the innate immune response. Biologicals. 2013, 41, 39–51.
- Ruggli, N.; Bird, B.H.; Liu, L.; Bauhofer, O.; Tratschin, J.D.; Hofmann, M.A. Npro of classical swine fever virus is an antagonist of double-stranded RNA-mediated apoptosis and IFN-α/β induction. Virology. 2005, 340, 265–76. [Google Scholar] [CrossRef]
- Smirnova NP, Bielefeldt-Ohmann H, Van Campen H, Austin KJ, Han H, Montgomery DL, et al. Acute non-cytopathic bovine viral diarrhea virus infection induces pronounced type I interferon response in pregnant cows and fetuses. Virus Res. 2008, 132, 49–58.
- Schweizer, M.; Peterhans, E. Noncytopathic Bovine Viral Diarrhea Virus Inhibits Double-Stranded RNA-Induced Apoptosis and Interferon Synthesis. J Virol. 2001, 75, 4692–8. [Google Scholar] [CrossRef] [PubMed]
- Pang, F.; Long, Q.; Wei, M. Immune evasion strategies of bovine viral diarrhea virus. Front Cell Infect Microbiol. 2023;13 October:1–8.
- Alexenko, A.P.; Ealy, A.D.; Roberts, R.M. The cross-species antiviral activities of different IFN-τ- subtypes on bovine, murine, and human cells: Contradictory evidence for therapeutic potential. J Interf Cytokine Res. 1999, 19, 1335–41. [Google Scholar] [CrossRef] [PubMed]
- Grummer, B.; Bendfeldt, S.; Wagner, B.; Greiser-Wilke, I. Induction of the intrinsic apoptotic pathway in cells infected with cytopathic bovine virus diarrhoea virus. Virus Res. 2002, 90, 143–53. [Google Scholar] [CrossRef] [PubMed]
- Hilbe, M.; Girao, V.; Bachofen, C.; Schweizer, M.; Zlinszky, K.; Ehrensperger, F. Apoptosis in Bovine Viral Diarrhea Virus (BVDV)-Induced Mucosal Disease Lesions: A Histological, Immunohistological, and Virological Investigation. Vet Pathol. 2013, 50, 46–55. [Google Scholar] [CrossRef]
- Verburg, S.G.; Lelievre, R.M.; Westerveld, M.J.; Inkol, J.M.; Sun, Y.L.; Workenhe, S.T. Viral-mediated activation and inhibition of programmed cell death. 2022.
- Darweesh MF, Rajput MKS, Braun LJ, Rohila JS, Chase CCL. BVDV Npro protein mediates the BVDV induced immunosuppression through interaction with cellular S100A9 protein. Microb Pathog. 2018;121 April:341–9.
- Flint, M.S.; Miller, D.B.; Tinkle, S.S. Restraint-induced modulation of allergic and irritant contact dermatitis in male and female B6. 129 Mice. Brain Behav Immun. 2000, 14, 256–69. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; DeLeo, F.R. Role of neutrophils in innate immunity: A systems biology-level approach. Wiley Interdiscip Rev Syst Biol Med. 2009, 1, 309–33. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol. 2011, 11, 519–31. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.B.; Albright, B.N.; Caswell, J.L. Effect of interleukin-8 and granulocyte colony-stimulating factor on priming and activation of bovine neutrophils. Infect Immun. 2003, 71, 1643–9. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Lin, K.S.; King, M.R. Fluid shear stress increases neutrophil activation via platelet-activating factor. Biophys J. 2014, 106, 2243–53. [Google Scholar] [CrossRef] [PubMed]
- Stevens MGH, Van Poucke M, Peelman LJ, Rainard P, De Spiegeleer B, Rogiers C, et al. Anaphylatoxin C5a-induced toll-like receptor 4 signaling in bovine neutrophils. J Dairy Sci. 2011, 94, 152–64.
- Brown, G.B.; Bolin, S.R.; Frank, D.E.; Roth, J.A. Defective function of leukocytes from cattle persistently infected with bovine viral diarrhea virus, and the influence or recombinant cytokines. Am J Vet Res. 1991, 52, 381–7. [Google Scholar] [CrossRef]
- Thakur N, Evans H, Abdelsalam K, Farr A, Rajput MKS, Young AJ, et al. Bovine viral diarrhea virus compromises Neutrophil’s functions in strain dependent manner. Microb Pathog. 2020;149 May:104515.
- Abdelsalam, K.; Kaushik, R.S.; Chase, C. The Involvement of Neutrophil in the Immune Dysfunction Associated with BVDV Infection. Pathogens. 2023;12.
- Yoshitake, H.; Takeda, Y.; Nitto, T.; Sendo, F. Cross-linking of GPI-80, a possible regulatory molecule of cell adhesion, induces up-regulation of CD11b/CD18 expression on neutrophil surfaces and shedding of L-selectin. J Leukoc Biol. 2002, 71, 205–11. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.A.; Kaeberle, M.L. Suppression of neutrophil and lymphocyte function induced by a vaccinal strain of bovine viral diarrhea virus with and without the administration of ACTH. Am J Vet Res. 1983, 44, 2366–72. [Google Scholar] [CrossRef] [PubMed]
- The E, Viral B, Virus D, Apoptosis L, Abdelsalam K, Rajput M, et al. viruses Strains and the Corresponding Infected-Macrophages ’. 2020.
- Charleston B, Brackenbury LS, Carr B V. , Fray MD, Hope JC, Howard CJ, et al. Alpha/Beta and Gamma Interferons Are Induced by Infection with Noncytopathic Bovine Viral Diarrhea Virus In Vivo. J Virol. 2002, 76, 923–7.
- Charleston, B.; Fray, M.D.; Baigent, S.; Carr, B.V.; Morrison, W.I. Establishment of persistent infection with non-cytopathic bovine viral diarrhoea virus in cattle is associated with a failure to induce type I interferon. J Gen Virol. 2001, 82, 1893–7. [Google Scholar] [CrossRef] [PubMed]
- Barone, L.J.; Cardoso, N.P.; Mansilla, F.C.; Castillo, M.; Capozzo, A.V. Enhanced infectivity of bovine viral diarrhoea virus (BVDV) in arginase-producing bovine monocyte-derived macrophages. Virulence. 2023. [CrossRef]
- Medzhitov, R. Approaching the Asymptote: 20 Years Later. Immunity. 2009, 30, 766–75. [Google Scholar] [CrossRef] [PubMed]
- Schaut, R.G.; Ridpath, J.F.; Sacco, R.E. Bovine viral diarrhea virus type 2 impairs macrophage responsiveness to toll-like receptor ligation with the exception of toll-like receptor 7. PLoS One. 2016, 11, 1–17. [Google Scholar] [CrossRef]
- Yonkers NL, Rodriguez B, Milkovich KA, Asaad R, Lederman MM, Heeger PS, et al. TLR Ligand-Dependent Activation of Naive CD4 T Cells by Plasmacytoid Dendritic Cells Is Impaired in Hepatitis C Virus Infection. J Immunol. 2007, 178, 4436–44.
- Biasin M, Piacentini L, Lo Caputo S, Naddeo V, Pierotti P, Borelli M, et al. TLR Activation Pathways in HIV-1–Exposed Seronegative Individuals. J Immunol. 2010, 184, 2710–7.
- Wortham BW, Eppert BL, Motz GT, Flury JL, Orozco-Levi M, Hoebe K, et al. NKG2D Mediates NK Cell Hyperresponsiveness and Influenza-Induced Pathologies in a Mouse Model of Chronic Obstructive Pulmonary Disease. J Immunol. 2012, 188, 4468–75.
- Glew, E.J.; Carr, B.V.; Brackenbury, L.S.; Hope, J.C.; Charleston, B.; Howard, C.J. Differential effects of bovine viral diarrhoea virus on monocytes and dendritic cells. J Gen Virol. 2003, 84, 1771–80. [Google Scholar] [CrossRef] [PubMed]
- Adler, H.; Jungi, T.W.; Pfister, H.; Strasser, M.; Sileghem, M.; Peterhans, E. Cytokine regulation by virus infection: bovine viral diarrhea virus, a flavivirus, downregulates production of tumor necrosis factor alpha in macrophages in vitro. J Virol. 1996, 70, 2650–3. [Google Scholar] [CrossRef]
- Beutler, B.; Cerami, A. The biology of cachectin/TNF - A primary mediator of the host response. Annu Rev Immunol. 1989, 7, 625–55. [Google Scholar] [CrossRef] [PubMed]
- Philip, R.; Epstein, L.B. Tumour necrosis factor as immunomodulator and mediator of monocyte cytotoxicity induced by itself, γ-interferon and interleukin-1. Nature. 1986, 323, 86–9. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.D.; Adair, B.M.; Foster, J.C. Effect of BVD virus infection on alveolar macrophage functions. Vet Immunol Immunopathol. 1995, 46, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Nanduri, B.; Pharr, G.T.; Stokes, J.V.; Pinchuk, L.M. Bovine Viral Diarrhea Virus infection affects the expression of proteins related to professional antigen presentation in bovine monocytes. Biochim Biophys Acta - Proteins Proteomics. 2009, 1794, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Medzhitov, R. Innate immune recognition. Annu Rev Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Kruger, E.F.; Boyd, B.L.; Pinchuk, L.M. Bovine monocytes induce immunoglobulin production in peripheral blood B lymphocytes. Dev Comp Immunol. 2003, 27, 889–97. [Google Scholar] [CrossRef]
- Boyd, B.L.; Lee, T.M.; Kruger, E.F.; Pinchuk, L.M. Cytopathic and non-cytopathic bovine viral diarrhoea virus biotypes affect fluid phase uptake and mannose receptor-mediated endocytosis in bovine monocytes. Vet Immunol Immunopathol. 2004, 102, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Rajput, M.K.; Darweesh, M.F.; Park, K.; Braun, L.J.; Mwangi, W.; Young, A.J.; et al. The effect of bovine viral diarrhea virus (BVDV) strains on bovine monocyte-derived dendritic cells (Mo-DC) phenotype capacity to produce, B. V.D.V. Virol J. 2014, 11, 1–15. [Google Scholar] [CrossRef]
- Cardoso N, Franco-Mahecha OL, Czepluch W, Quintana ME, Malacari DA, Trotta MV, et al. Bovine Viral Diarrhea Virus Infects Monocyte-Derived Bovine Dendritic Cells by an E2-Glycoprotein-Mediated Mechanism and Transiently Impairs Antigen Presentation. Viral Immunol. 2016, 29, 417–29.
- Collen, T.; Carr, V.; Parsons, K.; Charleston, B.; Morrison, W.I. Analysis of the repertoire of cattle CD4+ T cells reactive with bovine viral diarrhoea virus. Vet Immunol Immunopathol. 2002, 87, 235–8. [Google Scholar] [CrossRef]
- Van Gennip, H.G.P.; Van Rijn, P.A.; Widjojoatmodjo, M.N.; De Smit, A.J.; Moormann, R.J.M. Chimeric classical swine fever viruses containing envelope protein E(RNS) or E2 of bovine viral diarrhoea virus protect pigs against challenge with CSFV and induce a distinguishable antibody response. Vaccine. 2000, 19, 447–59. [Google Scholar] [CrossRef]
- Weiland, E.; Ahl, R.; Stark, R.; Weiland, F.; Thiel, H.J. A second envelope glycoprotein mediates neutralization of a pestivirus, hog cholera virus. J Virol. 1992, 66, 3677–82. [Google Scholar] [CrossRef]
- Collen T, Morrison WI. CD4+ T-cell responses to bovine viral diarrhoea virus in cattle. Virus Res. 2000, 67, 67–80.
- Rajput, M.K.S.; Darweesh, M.F.; Braun, L.J.; Mansour, S.M.G.; Chase, C.C.L. Comparative humoral immune response against cytopathic or non-cytopathic bovine viral diarrhea virus infection. Res Vet Sci. 2020;129 July 2019:109–16.
- Howard, C.J.; Clarke, M.C.; Sopp, P.; Brownlie, J. Immunity to bovine virus diarrhoea virus in calves: the role of different T-cell subpopulations analysed by specific depletion in vivo with monoclonal antibodies. Vet Immunol Immunopathol. 1992, 32, 303–14. [Google Scholar] [CrossRef] [PubMed]
- Donis, R.O. Molecular biology of bovine viral diarrhea virus and its interactions with the host. Elsevier Masson SAS; 1995.
- Deregt, D.; Masri, S.A.; Cho, H.J.; Ohmann, H.B. Monoclonal antibodies to the p80/125 and gp53 proteins of bovine viral diarrhea virus: Their potential use as diagnostic reagents. Can J Vet Res. 1990, 54, 343–8. [Google Scholar]
- Lambot, M.; Douart, A.; Joris, E.; Letesson, J.J.; Pastoret, P.P. Characterization of the immune response of cattle against non-cytopathic and cytopathic biotypes of bovine viral diarrhoea virus. J Gen Virol. 1997, 78, 1041–7. [Google Scholar] [CrossRef] [PubMed]
- Endsley, J.J.; Roth, J.A.; Ridpath, J.; Neill, J. Maternal antibody blocks humoral but not T cell responses to BVDV. Biologicals. 2003, 31, 123–5. [Google Scholar] [CrossRef] [PubMed]
- Howard, C.J.; Clarke, M.C.; Brownlie, J. Protection against respiratory infection with bovine virus diarrhoea virus by passively acquired antibody. Vet Microbiol. 1989, 19, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Downey ED, Tait RG, Mayes MS, Park CA, Ridpath JF, Garrick DJ, et al. An evaluation of circulating bovine viral diarrhea virus type 2 maternal antibody level and response to vaccination in Angus calves. J Anim Sci. 2013, 91, 4440–50.
- Fulton RW, Briggs RE, Payton ME, Confer AW, Saliki JT, Ridpath JF, et al. Maternally derived humoral immunity to bovine viral diarrhea virus (BVDV) 1a, BVDV1b, BVDV2, bovine herpesvirus-1, parainfluenza-3 virus bovine respiratory syncytial virus, Mannheimia haemolytica and Pasteurella multocida in beef calves, antibody decline. Vaccine. 2004, 22, 643–9.
- Muñoz-zanzi C a, Thurmond MC, Johnson WO, Hietala SK. Predicted ages of dairy calves when colostrum- derived bovine viral diarrhea virus antibodies or interfere with vaccination. J Am Vet Med Assoc. 2002, 221, 678–85.
- Reppert EJ, Chamorro MF, Robinson L, Cernicchiaro N, Wick J, Weaber RL, et al. Reppert et al 2019 vaccination of pregnant beef and antibodies in calves. 2019, 4, 313–6.
- Kim UH, Kang SS, Jang SS, Kim SW, Chung KY, Kang DH, et al. Bovine Viral Diarrhea Virus Antibody Level Variation in Newborn Calves after Vaccination of Late-Gestational Cows. Vet Sci. 2023;10.
- Menanteau-Horta, A.M.; Ames, T.R.; Johnson, D.W.; Meiske, J.C. Effect of maternal antibody upon vaccination with infectious bovine rhinotracheitis and bovine virus diarrhea vaccines. Can J Comp Med. 1985, 49, 10–4. [Google Scholar]
- Chamorro MF, Walz PH, Passler T, Santen E, Gard J, Rodning SP, et al. Efficacy of multivalent, modified- live virus (MLV) vaccines administered to early weaned beef calves subsequently challenged with virulent Bovine viral diarrhea virus type 2. BMC Vet Res. 2015, 11, 1–9.
- Fulton RW, Ridpath JF, Saliki JT, Briggs RE, Confer AW, Burge LJ, et al. Bovine viral diarrhea virus (BVDV) 1b: Predominant BVDV subtype in calves with respiratory disease. Can J Vet Res. 2002, 66, 181–90.
- Fulton RW, Purdy CW, Confer AW, Saliki JT, Loan RW, Briggs RE, et al. Bovine viral diarrhea viral infections in feeder calves with respiratory disease: Interactions with Pasteurella spp., parainfluenza-3 virus, and bovine respiratory syncytial virus. Can J Vet Res. 2000, 64, 151–9.
- Fray, M.D.; Paton, D.J.; Alenius, S. The effects of bovine viral diarrhoea virus on cattle reproduction in relation to disease control. Anim Reprod Sci. 2000;60–61:615–27.
- Fulton, R.W.; Cook, B.J.; Payton, M.E.; Burge, L.J.; Step, D.L. Immune response to bovine viral diarrhea virus (BVDV) vaccines detecting antibodies to BVDV subtypes 1a, 1b, 2a, and 2c. Vaccine. 2020, 38, 4032–7. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Chamorro, M.F.; Walz, P.H. Vaccination of cattle against bovine viral diarrhea virus. Vet Microbiol. 2017;206 December 2016:78–83.
- Griebel, PJ. BVDV vaccination in North America: Risks versus benefits. Anim Heal Res Rev. 2015, 16, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Mattick, D.; Smith, L. Protection from persistent infection with a bovine viral diarrhea virus (BVDV) type 1b strain by a modified-live vaccine containing BVDV types 1a and 2, infectious bovine rhinotracheitis virus, parainfluenza 3 virus and bovine respiratory syncytial virus. Vaccine. 2011, 29, 4657–62. [Google Scholar] [CrossRef] [PubMed]
- Bolin, S.R. Control of bovine viral diarrhea infection by use of vaccination. Vet Clin North Am Food Anim Pract. 1995, 11, 615–25. [Google Scholar] [CrossRef] [PubMed]
- Deregt, D.; Dubovi, E.J.; Jolley, M.E.; Nguyen, P.; Burton, K.M.; Gilbert, S.A. Mapping of two antigenic domains on the NS3 protein of the pestivirus bovine viral diarrhea virus. Vet Microbiol. 2005, 108, 13–22. [Google Scholar] [CrossRef]
- Fulton RW, Confer AW, Burge LJ, Perino LJ, d’Offay JM, Payton ME, et al. Antibody responses by cattle after vaccination with commercial viral vaccines containing bovine herpesvirus-1, bovine viral diarrhea virus, parainfluenza-3 virus, and bovine respiratory syncytial virus immunogens and subsequent revaccination at day 140. Vaccine. 1995, 13, 725–33.
- Dean, H.J.; Hunsaker, B.D.; Bailey, O.D.; Wasmoen, T. Prevention of persistent infection in calves by vaccination of dams with noncytopathic type-1 modified-live bovine viral diarrhea virus prior to breeding. Am J Vet Res. 2003, 64, 530–7. [Google Scholar] [CrossRef]
- Fairbanks, K.K.; Rinehart, C.L.; Ohnesorge, C.; Loughin, M.M.; Chase, C.C.L. Evaluation of fetal protection against experimental infection with type 1 and type 2 bovine viral diarrhea virus after vaccination of the dam with a bivalent modified-live virus vaccine. J Am Vet Med Assoc. 2004, 225, 1898–904. [Google Scholar] [CrossRef]
- Ficken, M.D.; Ellsworth, M.A.; Tucker, C.M.; Cortese, V.S. Effects of modified-live bovine viral diarrhea virus vaccines containing either type 1 or types 1 and 2 BVDV on heifers and their offspring after challenge with noncytopathic type 2 BVDV during gestation. J Am Vet Med Assoc. 2006, 228, 1559–64. [Google Scholar] [CrossRef]
- Frey, H.R.; Eicken, K.; Grummer, B.; Kenklies, S.; Oguzoglu, T.C.; Moennig, V. Foetal protection against bovine virus diarrhoea virus after two-step vaccination. J Vet Med Ser B. 2002, 49, 489–93. [Google Scholar] [CrossRef]
- Fulton RW, Johnson BJ, Briggs RE, Ridpath JF, Saliki JT, Confer AW, et al. Challege with Bovine viral diarrhea virus by exposure to persistently infected calves: Protection by vaccination and negative results of antigen testing in nonvaccinated acutely infected calves. Can J Vet Res. 2006, 70, 121–7.
- Grooms, D.L.; Bolin, S.R.; Coe, P.H.; Borges, R.J.; Coutu, C.E. To Bovine Viral Diarrhea Virus Following To Cattle. 2007, 68, 1417–22.
- Newcomer, B.W.; Walz, P.H.; Givens, M.D.; Wilson, A.E. Efficacy of bovine viral diarrhea virus vaccination to prevent reproductive disease: A meta-analysis. Theriogenology. 2015, 83, 360–365e1. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Givens, D. Diagnosis and Control of Viral Diseases of Reproductive Importance: Infectious Bovine Rhinotracheitis and Bovine Viral Diarrhea. Vet Clin North Am - Food Anim Pract. 2016, 32, 425–41. [Google Scholar] [CrossRef] [PubMed]
- Beer, M.; Hehnen, H.R.; Wolfmeyer, A.; Poll, G.; Kaaden, O.R.; Wolf, G. A new inactivated BVDV genotype I and II vaccine An immunisation and challenge study with BVDV genotype I. Vet Microbiol. 2000, 77, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.A.; Daly, R.F.; Chase, C.C. Influence of Modified Live Vaccines on Reproductive Performance in Beef Cattle. Range Beef Cow Symp. 2017;:75–83.
- Kalaycioglu, A.T. Bovine viral diarrhoea virus (bvdv) diversity and vaccination. a review. Vet Q. 2007, 29, 60–7. [Google Scholar] [CrossRef]
- Germain, R.N. The ins and outs of antigen processing. Nature. 1986, 322, 687–9. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.R.; Shilleto, R.W.; Williams, J.; Alexander, D.C.S. Prevention of transplacental infection of bovine foetus by bovine viral diarrhoea virus through vaccination. Arch Virol. 2002, 147, 2453–63. [Google Scholar] [CrossRef] [PubMed]
- Platt, R.; Widel, P.W.; Kesl, L.D.; Roth, J.A. Comparison of humoral and cellular immune responses to a pentavalent modified live virus vaccine in three age groups of calves with maternal antibodies, before and after BVDV type 2 challenge. Vaccine. 2009, 27, 4508–19. [Google Scholar] [CrossRef] [PubMed]
- Platt, R.; Coutu, C.; Meinert, T.; Roth, J.A. Humoral and T cell-mediated immune responses to bivalent killed bovine viral diarrhea virus vaccine in beef cattle. Vet Immunol Immunopathol. 2008, 122, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Platt, R.; Kesl, L.; Guidarini, C.; Wang, C.; Roth, J.A. Comparison of humoral and T-cell-mediated immune responses to a single dose of Bovela® live double deleted BVDV vaccine or to a field BVDV strain. Vet Immunol Immunopathol. 2017;187 December 2016:20–7.
- Zimmerman, A.D.; Boots, R.E.; Valli, J.L.; Chase, C.C.L. Evaluation of protection agianst virulent bovine viral diarrhea virus type 2 in calves that had maternal antibodies and were vaccinated with a modified-live vaccine. J Am Vet Med Assoc. 2006, 228, 1757–61. [Google Scholar] [CrossRef]
- Gómez-Romero N, Arias CF, Verdugo-Rodríguez A, López S, Valenzuela-Moreno LF, Cedillo-Peláez C, et al. Immune protection induced by E2 recombinant glycoprotein of bovine viral diarrhea virus in a murine model. Front Vet Sci. 2023;10.
- Wang S, Yang G, Nie J, Yang R, Du M, Su J, et al. Recombinant Erns-E2 protein vaccine formulated with MF59 and CPG-ODN promotes T cell immunity against bovine viral diarrhea virus infection. Vaccine. 2020, 38, 3881–91.
- Chung, Y.C.; Cheng, L.T.; Zhang, J.Y.; Wu, Y.J.; Liu, S.S.; Chu, C.Y. Recombinant E2 protein enhances protective efficacy of inactivated bovine viral diarrhea virus 2 vaccine in a goat model. BMC Vet Res. 2018, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Marconi, P.; Periolo, O.; La Torre, J.; Alvarez, M.A. Immunocompetent truncated E2 glycoprotein of bovine viral diarrhea virus (BVDV) expressed in Nicotiana tabacum plants: A candidate antigen for new generation of veterinary vaccines. Vaccine. 2012, 30, 4499–504. [Google Scholar] [CrossRef]
- Pecora A, Malacari DA, Perez Aguirreburualde MS, Bellido D, Nuñez MC, Dus Santos MJ, et al. Development of an APC-targeted multivalent E2-based vaccine against Bovine Viral Diarrhea Virus types 1 and 2. Vaccine. 2015, 33, 5163–71.
- Carlsson, U.; Alenius, S.; Sundquist, B. Protective effect of an ISCOM bovine virus diarrhoea virus (BVDV) vaccine against an experimental BVDV infection in vaccinated and non-vaccinated pregnant ewes. Vaccine. 1991, 9, 577–80. [Google Scholar] [CrossRef]
- Bruschke, C.J.M.; Van Oirschot, J.T.; Van Rijn, P.A. An experimental multivalent bovine virus diarrhea virus E2 subunit vaccine and two experimental conventionally inactivated vaccines induce partial fetal protection in sheep. Vaccine. 1999, 17, 1983–91. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury SI, Pannhorst K, Sangewar N, Pavulraj S, Wen X, Stout RW, et al. Bohv-1-vectored bvdv-2 subunit vaccine induces bvdv cross-reactive cellular immune responses and protects against bvdv-2 challenge. Vaccines. 2021, 9, 1–30.
- Luo, Y.; Yuan, Y.; Ankenbauer, R.G.; Nelson, L.D.; Witte, S.B.; Jackson, J.A.; et al. Construction of chimeric bovine viral diarrhea viruses containing glycoprotein E rns of heterologous pestiviruses evaluation of the chimeras as potential marker vaccines against, B. V.D.V. Vaccine. 2012, 30, 3843–8. [Google Scholar]
- El-Attar, L.M.R.; Thomas, C.; Luke, J.; Williams, J.A.; Brownlie, J. Enhanced neutralising antibody response to bovine viral diarrhoea virus (BVDV) induced by DNA vaccination in calves. Vaccine. 2015, 33, 4004–12. [Google Scholar] [CrossRef] [PubMed]
- Koethe S, König P, Wernike K, Pfaff F, Schulz J, Reimann I, et al. A synthetic modified live chimeric marker vaccine against bvdv-1 and bvdv-2. Vaccines. 2020, 8, 1–23.
- Takahashi, H.; Takeshita, T.; Berzofsky, J.A. Induction of CDS + cytotoxic T purified HIV -1 envelope. Nature. 1990;344 April:873–5.
- Snider M, Garg R, Brownlie R, van den Hurk J V. , Hurk S van DL van den. The bovine viral diarrhea virus E2 protein formulated with a novel adjuvant induces strong, balanced immune responses and provides protection from viral challenge in cattle. Vaccine. 2014, 32, 6758–64.
- Pecora A, Malacari DA, Pérez Aguirreburualde MS, Bellido D, Escribano JM, Dus Santos MJ, et al. Development of an enhanced bovine viral diarrhea virus subunit vaccine based on E2 glycoprotein fused to a single chain antibody which targets to antigen-presenting cells. Rev Argent Microbiol. 2015, 47, 4–8.
- Bellido D, Baztarrica J, Rocha L, Pecora A, Acosta M, Escribano JM, et al. A novel MHC-II targeted BVDV subunit vaccine induces a neutralizing immunological response in guinea pigs and cattle. Transbound Emerg Dis. 2021, 68, 3474–81.
- Aguirreburualde MSP, Gómez MC, Ostachuk A, Wolman F, Albanesi G, Pecora A, et al. Efficacy of a BVDV subunit vaccine produced in alfalfa transgenic plants. Vet Immunol Immunopathol. 2013, 151, 315–24.
- Sadat, S.M.A.; Snider, M.; Garg, R.; Brownlie, R.; van Drunen Littel-van den Hurk, S. Local innate responses and protective immunity after intradermal immunization with bovine viral diarrhea virus E2 protein formulated with a combination adjuvant in cattle. Vaccine. 2017, 35, 3466–73. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Young, N.J.; Heaney, J.; Collins, M.E.; Brownlie, J. Evaluation of efficacy of mammalian and baculovirus expressed E2 subunit vaccine candidates to bovine viral diarrhoea virus. Vaccine. 2009, 27, 2387–93. [Google Scholar] [CrossRef]
- Jia S, Huang X, Li H, Zheng D, Wang L, Qiao X, et al. Immunogenicity evaluation of recombinant Lactobacillus casei W56 expressing bovine viral diarrhea virus E2 protein in conjunction with cholera toxin B subunit as an adjuvant. Microb Cell Fact. 2020, 19, 1–20.
- Mody KT, Mahony D, Cavallaro AS, Zhang J, Zhang B, Mahony TJ, et al. Silica vesicle nanovaccine formulations stimulate long-term immune responses to the Bovine Viral Diarrhoea Virus E2 protein. PLoS One. 2015, 10, 1–16.
- Mahony D, Cavallaro AS, Mody KT, Xiong L, Mahony TJ, Qiao SZ, et al. In vivo delivery of bovine viral diahorrea virus, E2 protein using hollow mesoporous silica nanoparticles. Nanoscale. 2014, 6, 6617–26.
- Wang, L.; Sunyer, J.O.; Bello, L.J. Fusion to C3d Enhances the Immunogenicity of the E2 Glycoprotein of Type 2 Bovine Viral Diarrhea Virus. J Virol. 2004, 78, 1616–22. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The pathogenesis of persistent infection. The virus enters the host through the respiratory or oral route. Both pregnant and non-pregnant cows are susceptible to ncp and cp BVDV biotypes. BVDV may cause transient and persistent infections which differ fundamentally in the host’s antiviral immune response. Transient infection may be due to both cytopathic and non-cytopathic biotypes of BVDV and leads to a specific immune response. In contrast, only non-cytopathic BVD viruses can establish persistent infection as a result of infection of the embryo early in its developmental stage [10]. Infection with ncp biotype during the early stage of pregnancy (40-125 days) can result in embryo absorption, abortion, stillbirth, or the birth of a PI calf. The CP biotype of BVDV evolves from the same genotype through mutation in a PI calf, leading to life-threatening mucosal disease. Non-pregnant cows only seroconvert during acute infection and quickly clear the virus (adapted from www.vetres.org and figure made using Biorender).
Figure 1.
The pathogenesis of persistent infection. The virus enters the host through the respiratory or oral route. Both pregnant and non-pregnant cows are susceptible to ncp and cp BVDV biotypes. BVDV may cause transient and persistent infections which differ fundamentally in the host’s antiviral immune response. Transient infection may be due to both cytopathic and non-cytopathic biotypes of BVDV and leads to a specific immune response. In contrast, only non-cytopathic BVD viruses can establish persistent infection as a result of infection of the embryo early in its developmental stage [10]. Infection with ncp biotype during the early stage of pregnancy (40-125 days) can result in embryo absorption, abortion, stillbirth, or the birth of a PI calf. The CP biotype of BVDV evolves from the same genotype through mutation in a PI calf, leading to life-threatening mucosal disease. Non-pregnant cows only seroconvert during acute infection and quickly clear the virus (adapted from www.vetres.org and figure made using Biorender).
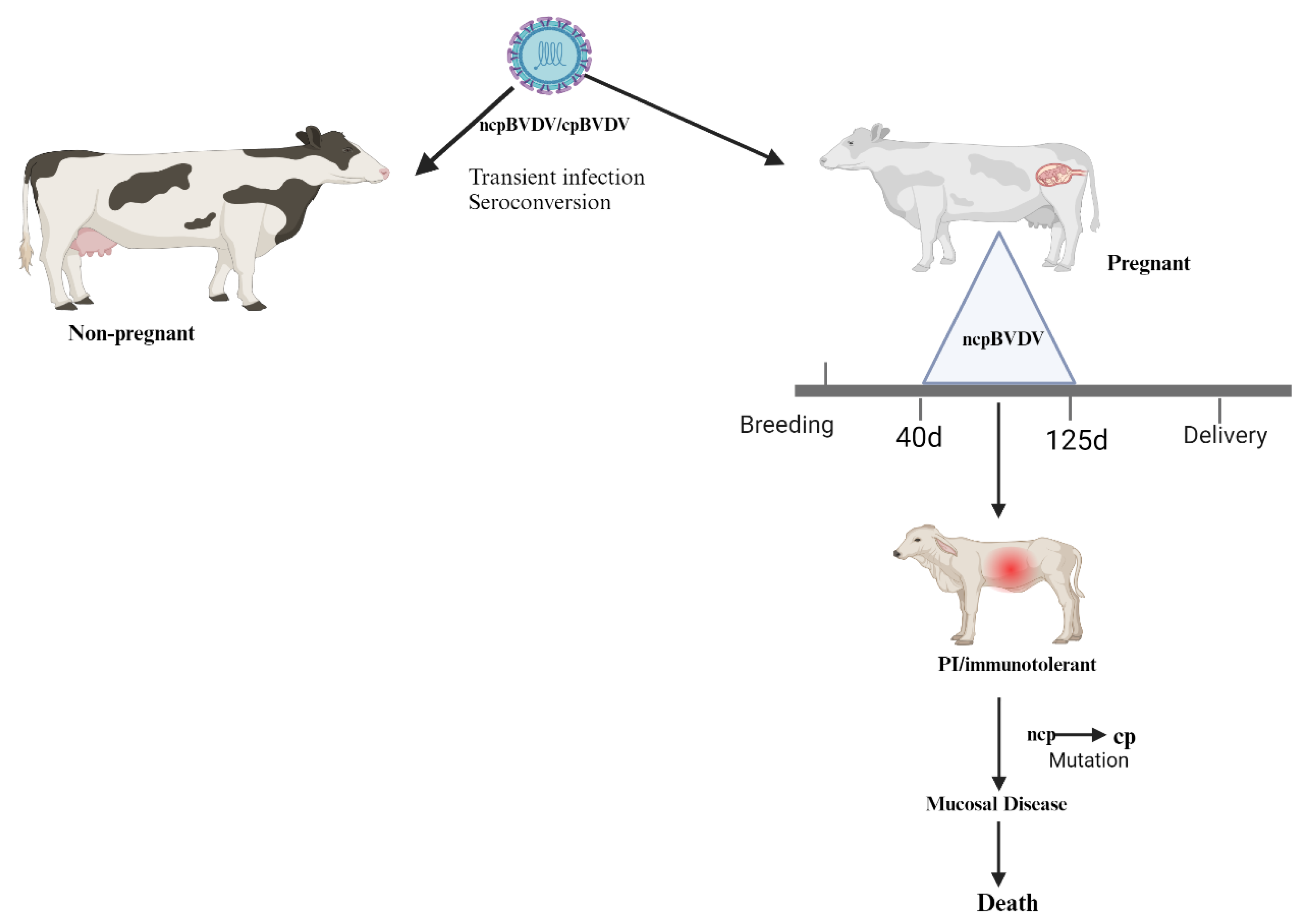
Figure 2.
TLR 7/8 activation by cpBVDV ssRNA to induce interferon-β (IFN-β). After single-stranded RNA (ssRNA) of the BVDV activates endosomal TLR7/8, TRIF transiently colocalizes with TLR 7/8; it dissociates from the receptor and forms a speckle-like structure that relocalizes with downstream-signaling molecules such as tumor necrosis factor (TNF) receptor-associated factor 2 (TRAF2), TRAF6, and receptor-interacting protein-1 (RIP-1) [38,39]. Then, with the cooperation of TRAF3, TRAF family member-associated nuclear transcription factor-κB (NF-κB) activator (TANK)-binding kinase 1 (TBK1), inhibitor of κB (IκB) kinase-related kinase- ε (IKK- ε; also called IKK-i), and NF-κB-activating kinase (NAK)-associated protein 1 (NAP1), this signaling pathway ultimately activates transcription factors, namely IFN-regulatory factor 3/7 (IRF3/7), the TRIF-dependent NFκB, and the activator protein 1 (AP-1), thus mediating the production of type I IFNs, proinflammatory cytokines, and chemokines, respectively. ZCCHC3: zinc finger CCHC-type containing 3; WDFY1: WD-repeat- and FYVE-domain-containing protein 1; TRIM23: tripartite motif-containing protein 23; LUBAC: linear ubiquitin chain assembly complex; RNF166: ring finger protein 166; HECTD3: E6-associated protein carboxyl terminus domain containing 3; Nedd4l: neural precursor cell expressed developmentally down-regulated 4-like; MIB: mind bomb; RNF128: RING finger 128; USP2b: ubiquitin-specific protease 2 isoform b; CYLD: conserved cylindromatosis; OTUD1: termed OTU domain-containing protein 1; PI3K: phosphoinositide 3-kinase; NEMO: NF-κB essential modifier; MAPK: p38-mitogen-activated protein kinase; FLIP: viral FLICE-like inhibitory protein. Adapted from [40] and Created with BioRender.com .
Figure 2.
TLR 7/8 activation by cpBVDV ssRNA to induce interferon-β (IFN-β). After single-stranded RNA (ssRNA) of the BVDV activates endosomal TLR7/8, TRIF transiently colocalizes with TLR 7/8; it dissociates from the receptor and forms a speckle-like structure that relocalizes with downstream-signaling molecules such as tumor necrosis factor (TNF) receptor-associated factor 2 (TRAF2), TRAF6, and receptor-interacting protein-1 (RIP-1) [38,39]. Then, with the cooperation of TRAF3, TRAF family member-associated nuclear transcription factor-κB (NF-κB) activator (TANK)-binding kinase 1 (TBK1), inhibitor of κB (IκB) kinase-related kinase- ε (IKK- ε; also called IKK-i), and NF-κB-activating kinase (NAK)-associated protein 1 (NAP1), this signaling pathway ultimately activates transcription factors, namely IFN-regulatory factor 3/7 (IRF3/7), the TRIF-dependent NFκB, and the activator protein 1 (AP-1), thus mediating the production of type I IFNs, proinflammatory cytokines, and chemokines, respectively. ZCCHC3: zinc finger CCHC-type containing 3; WDFY1: WD-repeat- and FYVE-domain-containing protein 1; TRIM23: tripartite motif-containing protein 23; LUBAC: linear ubiquitin chain assembly complex; RNF166: ring finger protein 166; HECTD3: E6-associated protein carboxyl terminus domain containing 3; Nedd4l: neural precursor cell expressed developmentally down-regulated 4-like; MIB: mind bomb; RNF128: RING finger 128; USP2b: ubiquitin-specific protease 2 isoform b; CYLD: conserved cylindromatosis; OTUD1: termed OTU domain-containing protein 1; PI3K: phosphoinositide 3-kinase; NEMO: NF-κB essential modifier; MAPK: p38-mitogen-activated protein kinase; FLIP: viral FLICE-like inhibitory protein. Adapted from [40] and Created with BioRender.com .
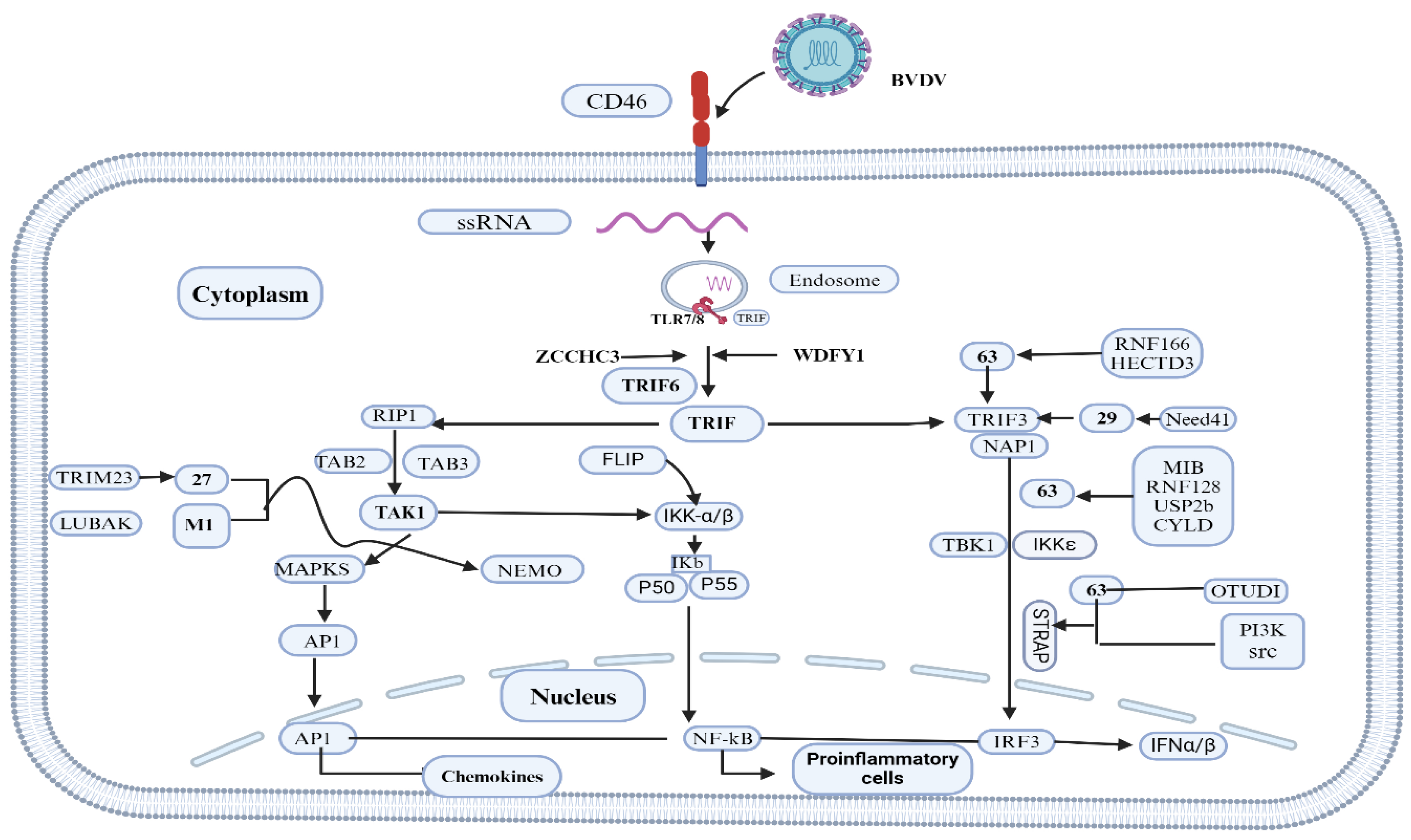
Figure 3.
The induction of intrinsic apoptosis pathways during BVDV infection. The intrinsic apoptosis pathway is induced during MD than the extrinsic pathway. The intrinsic pathway of apoptosis arises from proapoptotic stimuli, subsequently initiating mitochondrial membrane permeabilization and activation of caspase-9 through cytochrome C. Both pathways converge at the terminal activation of caspase-3 that executes apoptosis. ATF4AU=activating transcription factor 4; ATF6== activating transcription factor 6; BCL 2= B cell lymphoma-2; BAK= BCL 2-associated antagonist or killer; BAX= BCL-2-associated X protein; cGAS/STING= cyclic GMP–AMP synthetase/stimulator of IFN genes; CHOP= CCAAT-enhancer-binding protein homologous protein; DISC, death-inducing signaling complex; HPV=Human papillomavirus; HCV=hepatitis C virus; Influenza virus; endoplasmic reticulum; IRE1= inositol-requiring enzyme 1; IRF3=IFN regulatory factor 3; MVA= modified vaccinia virus; RV=Reovirus; PERK= PKR-like ER kinase; RIG-I=retinoic acid-inducible gene I; TLR3=Toll-like receptor 3; TN=tumor necrosis factor; TRIF=Toll/IL-1 receptor domain-containing adapter inducing IFN-beta Adapted from Verburg et al.(2022) and Created with BioRender.com.
Figure 3.
The induction of intrinsic apoptosis pathways during BVDV infection. The intrinsic apoptosis pathway is induced during MD than the extrinsic pathway. The intrinsic pathway of apoptosis arises from proapoptotic stimuli, subsequently initiating mitochondrial membrane permeabilization and activation of caspase-9 through cytochrome C. Both pathways converge at the terminal activation of caspase-3 that executes apoptosis. ATF4AU=activating transcription factor 4; ATF6== activating transcription factor 6; BCL 2= B cell lymphoma-2; BAK= BCL 2-associated antagonist or killer; BAX= BCL-2-associated X protein; cGAS/STING= cyclic GMP–AMP synthetase/stimulator of IFN genes; CHOP= CCAAT-enhancer-binding protein homologous protein; DISC, death-inducing signaling complex; HPV=Human papillomavirus; HCV=hepatitis C virus; Influenza virus; endoplasmic reticulum; IRE1= inositol-requiring enzyme 1; IRF3=IFN regulatory factor 3; MVA= modified vaccinia virus; RV=Reovirus; PERK= PKR-like ER kinase; RIG-I=retinoic acid-inducible gene I; TLR3=Toll-like receptor 3; TN=tumor necrosis factor; TRIF=Toll/IL-1 receptor domain-containing adapter inducing IFN-beta Adapted from Verburg et al.(2022) and Created with BioRender.com.
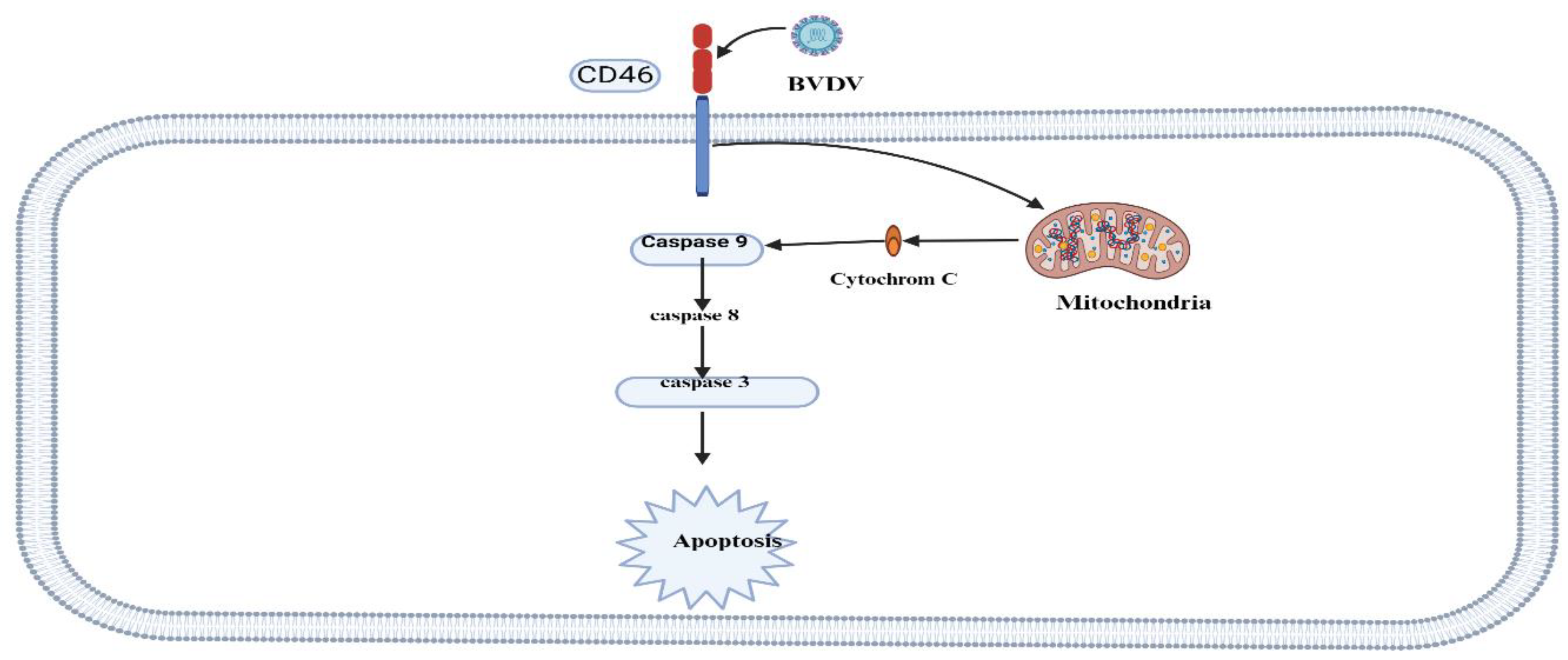
Figure 4.
Model for the interaction between the two biotypes of BVDV (cp and ncp) and cells of the innate and adaptive immune system. Infection with cp-BVDV at a mucosal or epithelial surface causes the expression of cytokines such as IFNα from various cell types, including DCs. These cytokines activate effector cells of the innate immune response, such as eosinophils, macrophages, and NK cells. The combined effect of cytokines and effector cell function reduces viral replication of the virus. The environment created by the cp infection activates DCs, leading to enhanced antigen capture, maturation, and migration. DCs are likely resistant to the lytic effects of cp infection and can migrate to the local lymph node. After migrating to the lymph node, the antigen-carrying DCs present antigens via peptide-MHC I complexes to antigen-specific lymphocytes. The resulting activated T lymphocytes (e.g., cytotoxic lymphocytes—CTLs, helper T cells) return to the site of infection to eliminate the virus or virus-infected cells. Activated B cells migrate to the germinal center (GC) or more in the lymph node, where they mature into plasma cells that produce antibodies to neutralize the virus. In comparison to cp infection, ncp infection cannot stimulate an early cytokine response. Consequently, the replication of the virus is not limited, and DCs do not become highly activated. Cell-free virus and virus-infected DCs migrate to the local lymph node. In the lymph node, the virus encounters , which produce large quantities of IFNα. The increasing quantities of IFNα in the circulation and tissue enhance the activation of DCs and limit viral replication. However, the ncp virus has already disseminated throughout the animal [23]. The activity of neutrophils to move and eliminate microorganisms entering the body is determined by the expression of surface markers such as cluster of differentiation 14 (CD14), CD-18, and L-selectin [64]. Bovine viral diarrhea virus (BVDV) has been found to compromise neutrophil functions in a biotype-dependent manner [62,63]. The cpBVDV biotype down-regulates nitric oxide production and neutrophil extracellular traps (NET) induction [63]. Roth and Kaeberle [65] indicated vaccinal strain of BVDV caused a decrease in circulating lymphocytes and neutrophils. Created with BioRender.com.
Figure 4.
Model for the interaction between the two biotypes of BVDV (cp and ncp) and cells of the innate and adaptive immune system. Infection with cp-BVDV at a mucosal or epithelial surface causes the expression of cytokines such as IFNα from various cell types, including DCs. These cytokines activate effector cells of the innate immune response, such as eosinophils, macrophages, and NK cells. The combined effect of cytokines and effector cell function reduces viral replication of the virus. The environment created by the cp infection activates DCs, leading to enhanced antigen capture, maturation, and migration. DCs are likely resistant to the lytic effects of cp infection and can migrate to the local lymph node. After migrating to the lymph node, the antigen-carrying DCs present antigens via peptide-MHC I complexes to antigen-specific lymphocytes. The resulting activated T lymphocytes (e.g., cytotoxic lymphocytes—CTLs, helper T cells) return to the site of infection to eliminate the virus or virus-infected cells. Activated B cells migrate to the germinal center (GC) or more in the lymph node, where they mature into plasma cells that produce antibodies to neutralize the virus. In comparison to cp infection, ncp infection cannot stimulate an early cytokine response. Consequently, the replication of the virus is not limited, and DCs do not become highly activated. Cell-free virus and virus-infected DCs migrate to the local lymph node. In the lymph node, the virus encounters , which produce large quantities of IFNα. The increasing quantities of IFNα in the circulation and tissue enhance the activation of DCs and limit viral replication. However, the ncp virus has already disseminated throughout the animal [23]. The activity of neutrophils to move and eliminate microorganisms entering the body is determined by the expression of surface markers such as cluster of differentiation 14 (CD14), CD-18, and L-selectin [64]. Bovine viral diarrhea virus (BVDV) has been found to compromise neutrophil functions in a biotype-dependent manner [62,63]. The cpBVDV biotype down-regulates nitric oxide production and neutrophil extracellular traps (NET) induction [63]. Roth and Kaeberle [65] indicated vaccinal strain of BVDV caused a decrease in circulating lymphocytes and neutrophils. Created with BioRender.com.
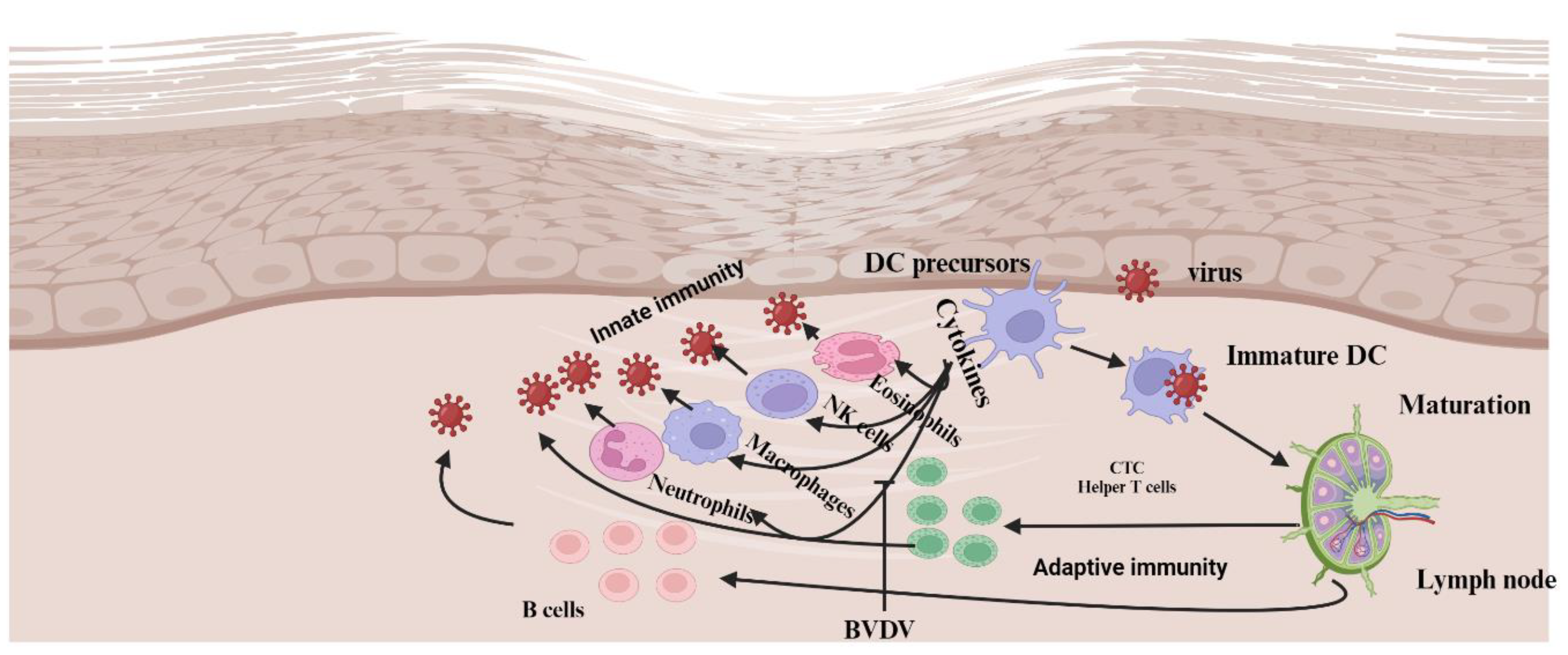
Figure 5.
Retinoic Acid-inducible Gene-I-like Receptors (RLR) and Toll-Like Receptor (TLR) signaling. The RIG-I-like receptors (RLR: RIG-I, Mda-5, and LGP2) are PRRs that detect various forms of dsRNA. The signal via the mitochondrial adapter protein MAVS activates the transcription factors AP-1 (c-Jun/ATF-2), NF-kB, and IRF-3/7 via several kinases, such as MAP kinase (MAPK), the I-kB kinase complex (IKK), and Tbk-1 or IKKε. The Toll-like receptors TLR-3, -7/8, and -9 located in endosomal compartments detect dsRNA, ssRNA, and dsDNA, respectively, and signal via different transduction pathways using the adapter protein Trif or MyD88 to activate AP-1, NF-kB, and IRF followed by expression of "IFN-stimulated genes" (ISG). In addition, viruses replicating in the cytoplasm may similarly be detected by TLRs upon autophagy and fusion of the autophagosome with the endosomal compartments [45]. Created with BioRender.com.
Figure 5.
Retinoic Acid-inducible Gene-I-like Receptors (RLR) and Toll-Like Receptor (TLR) signaling. The RIG-I-like receptors (RLR: RIG-I, Mda-5, and LGP2) are PRRs that detect various forms of dsRNA. The signal via the mitochondrial adapter protein MAVS activates the transcription factors AP-1 (c-Jun/ATF-2), NF-kB, and IRF-3/7 via several kinases, such as MAP kinase (MAPK), the I-kB kinase complex (IKK), and Tbk-1 or IKKε. The Toll-like receptors TLR-3, -7/8, and -9 located in endosomal compartments detect dsRNA, ssRNA, and dsDNA, respectively, and signal via different transduction pathways using the adapter protein Trif or MyD88 to activate AP-1, NF-kB, and IRF followed by expression of "IFN-stimulated genes" (ISG). In addition, viruses replicating in the cytoplasm may similarly be detected by TLRs upon autophagy and fusion of the autophagosome with the endosomal compartments [45]. Created with BioRender.com.
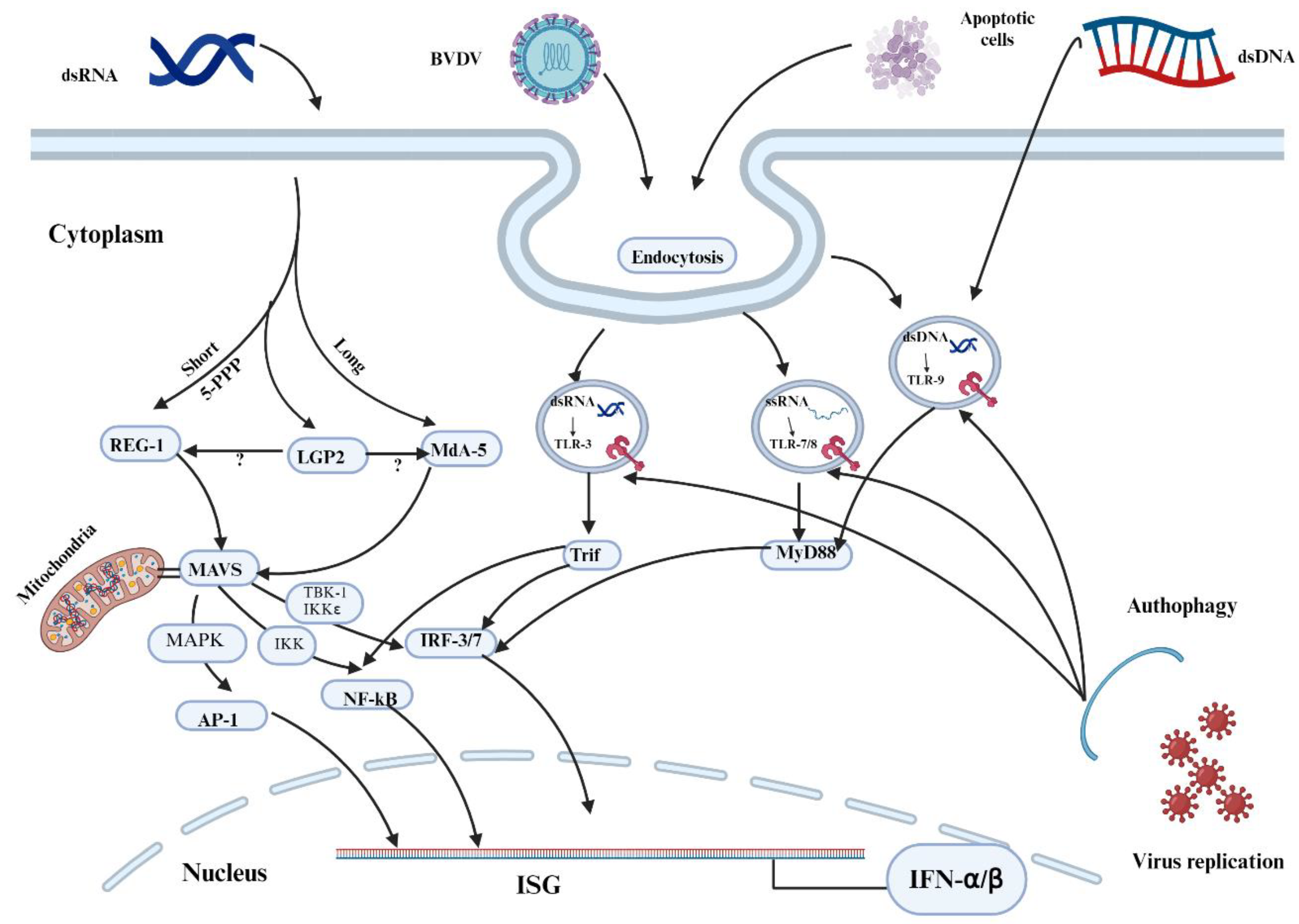
Figure 6.
Activation of antigen-presenting cells and interactions with T cells. The viral pathogen-associated molecular pattern (PAMP) binds to pathogen recognition receptors (PRRs) on antigen-presenting cells, such as dendritic cells (DCs). The activated mature DCs migrate with the antigen to lymph nodes, where they encounter naïve T lymphocytes and present the processed peptides loaded onto MHC II to CD4+ cells. These CD4+ cells then differentiate into Th1 and Th2 cells in the presence of cytokines interleukins. Th1 cells activate macrophages to phagocytize and kill the pathogen, as well as help B cells to produce antibodies against the pathogen. In the case of viral diseases like BVDV, the activation of cytotoxic T cells (CTL) by CD4+ cells is important. DCs with the processed peptides on MHC-I migrate to lymph nodes and bind to naïve CTL. This activated CTL releases effector molecules such as perforin, granzymes, and granulysin to kill virus-infected cells. PRR=pathogen recognition receptor, PAMP= pathogen-associated molecular pattern. Toll signifies Toll-like receptor, B7/CD80/CD86 represents APC co-receptor, CD28 denotes T cell co-receptor, and MHC II refers to the major histocompatibility II receptor expressed by APC to "present" antigen to the T cell. Created with BioRender.com.
Figure 6.
Activation of antigen-presenting cells and interactions with T cells. The viral pathogen-associated molecular pattern (PAMP) binds to pathogen recognition receptors (PRRs) on antigen-presenting cells, such as dendritic cells (DCs). The activated mature DCs migrate with the antigen to lymph nodes, where they encounter naïve T lymphocytes and present the processed peptides loaded onto MHC II to CD4+ cells. These CD4+ cells then differentiate into Th1 and Th2 cells in the presence of cytokines interleukins. Th1 cells activate macrophages to phagocytize and kill the pathogen, as well as help B cells to produce antibodies against the pathogen. In the case of viral diseases like BVDV, the activation of cytotoxic T cells (CTL) by CD4+ cells is important. DCs with the processed peptides on MHC-I migrate to lymph nodes and bind to naïve CTL. This activated CTL releases effector molecules such as perforin, granzymes, and granulysin to kill virus-infected cells. PRR=pathogen recognition receptor, PAMP= pathogen-associated molecular pattern. Toll signifies Toll-like receptor, B7/CD80/CD86 represents APC co-receptor, CD28 denotes T cell co-receptor, and MHC II refers to the major histocompatibility II receptor expressed by APC to "present" antigen to the T cell. Created with BioRender.com.
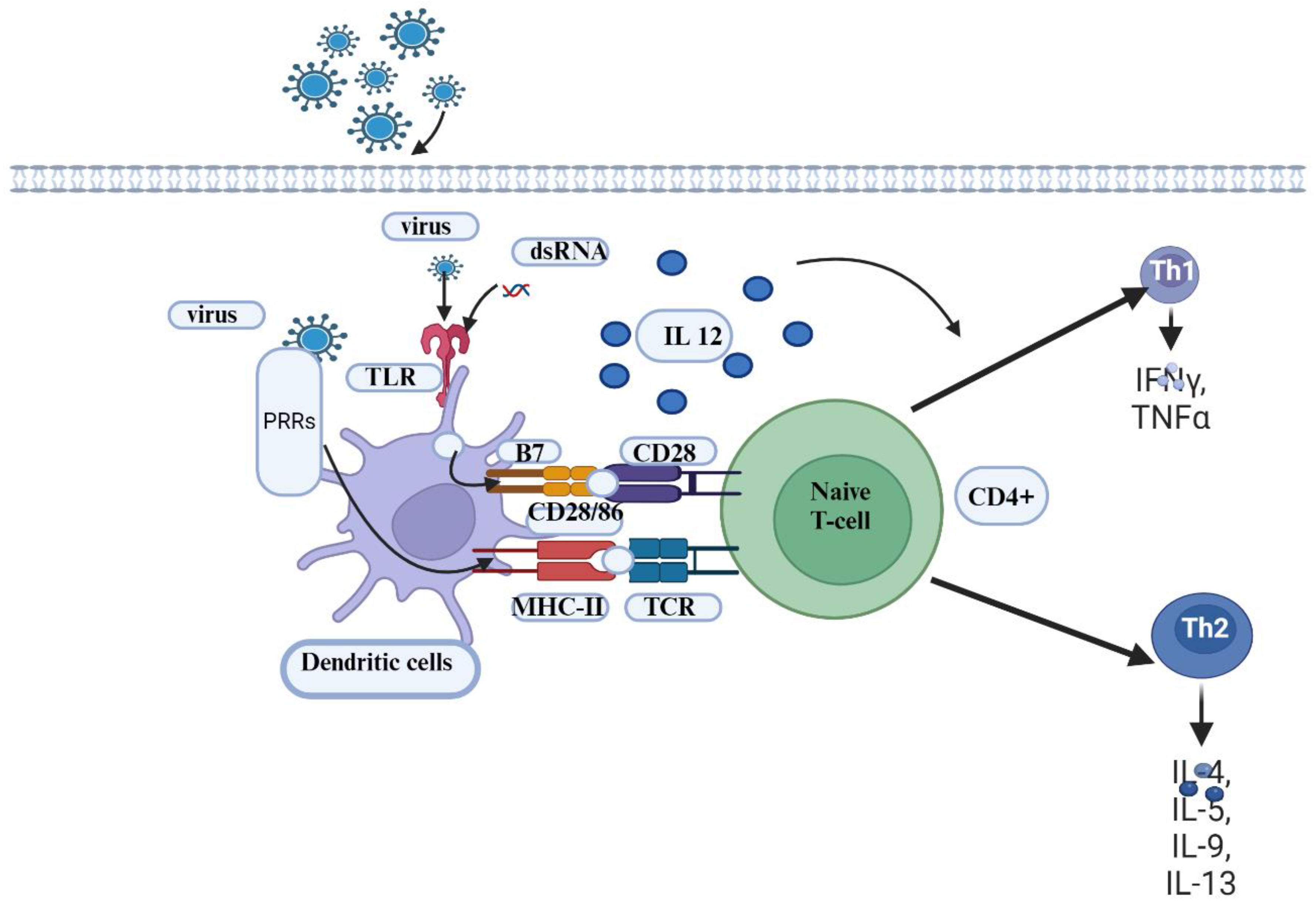

Table 1.
Various types of BVD-licensed commercial vaccines available for use in cattle.
| Vaccines name | Types | Dose(ml) | RA | Booster | Used During pregnancy | Genotypes | Strains | Subtypes | Biotypes | Manufacturers | Website |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bovi-Shield GOLD® IBR-BVD | MLV | 2ml | SC/IM | 1 year | No | BVDV1 & 2 | NADL | 1a | ncp | Zoetis | www.zoetis.com |
| Bovela | MLV | 2ml | IM | 1year | Yes | BVDV1& 2 | KE-9 & NY_93 | 1a/2a | ncp | Boehringer | www.vetmedica.de |
| Bovidec | IV | 4ml | SC | 1year | No | BVDV 1 | KY1203 | NA | ncp | Zoetis | www.zoetis.com |
| Bovilis® BVD | IV | 2ml | IM | 1 year | Yes | BVDV1 | C-86 | 1a | cp | Zoetis | www.zoetis.com |
| ONE SHOT® BVD | IV | 2ml | SC | 1year | No | BVDV1 & 2 | NA | NA | NA | Zoetis | www.zoetis.com |
| PESTIGARD® | IV | 2ml | SC | Yes | BVDV | NA | NA | NA | Zoetis | www.zoetis.com | |
| BOVI-SHIELD GOLD FP 5 | IV | 2ml | IM | 1year | No | BVDV1 &2 | NADAL/53637 | 1a/2a | cp | Zoetis | www.zoetis.com |
| CattleMaster GOLD FP 5 | IV | 2ml | SC | 1year | Yes | BVDV1 & 2 | 5960/53637 | 1a/2a | cp | Zoetis | www.zoetis.com |
| CATTLEMASTER 4 | IV | 2ml | IM | 1year | Yes | BVDV1 | 5960/6309 | 1a/1 | ncp/cp | Zoetis | www.zoetis.com |
| Elit 4 | IM | 5ml | IM | 1year | Yes | BVDV1 | Singer | 1a | cp | Boehringer | www.vetmedica.de |
| Master Guard 10 HB | MLV/IV | 3ml | IM/SC | 1year | Yes | BVD1 & 2 | C24V | 1a | cp | Elanco | elanco@elanco.com |
| Triangel 5 | IV | 2ml | IM/SC | 1 year | Yes | BVDV1& 2 | Singer/5912 | 1a/2a | cp/cp | Boehringer | www.vetmedica.de |
| Vira Shield 6 + VL5 HB | IV | 5ml | SC | 1year | Yes | BVDV1 &2 | K12/GL 760/TN131 | 1a/1a | cp/ncp/cp | Novarties | elanco@elanco.com |
| Express 5 | MLV | 2ml | 1year | Yes | BVDV 1& 2 | Singer | 1a/2c | cp/ncp/cp | Boehringer | www.vetmedica.de | |
| Pyramid 4 | MLV | 2ml | SC/IM | 1year | No | BVDV1 & 2 | Singer | 1a | cp | Boehringer | www.vetmedica.de |
| Titanium 5 | MLV | 2ml | SC | 1year | Yes | BVDV1 &2 | C24V/296 | 1a/2c | cp | Elanco | elanco@elanco.com |
| TRIANGLE® 4+PH-K | IV | 5ml | SC | 1year | Not tested | BVD1 | Singer | 1a | cp | Boehringer | www.vetmedica.de |
Table 2.
BVDV next-generation vaccines with modern adjuvants.
| Antigen | Adjuvants | Vaccine Types | References |
|---|---|---|---|
| tE2 | Montanide ISA 70 SEPPIC® and (Al(OH)3Hydrogel®) | Subunit | [134,135] |
| rE2 | ISA 61 VG | Recombinant | [131] |
| E2 | Immunostimulating complexes (ISCOMs) | Recombinant | [136] |
| E2 | cholera toxin B subunit (ctxB) | Recombinant | [132] |
| E2,NS2-3 | MONTANIDETM ISA 201 VG adjuvant | Subunit | [21] |
| E2 | NA | Subunit | [137] |
| E2 and flag-tagged Erns | NA | Subunit | [138] |
| Erns | NA | Marker vaccine | [139] |
| Erns | NA | DNA | [140] |
| Npro_Erns | NA | Chimeric vaccines | [141] |
| Npro,E2 and NS2-3 | BenchMark-Vaxliant | Mosaic vaccines | [22] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



