Submitted:
08 January 2025
Posted:
09 January 2025
You are already at the latest version
Abstract
Biofilms are the predominant lifestyle for microorganisms, contributing to over 80% of infections, including those caused by Candida species like C. albicans, C. auris, and C. parapsilosis. These species form biofilms on medical devices, making infections challenging to treat, especially with the rise of drug-resistant strains. Candida infections, particularly hospital-acquired ones, are a significant health threat due to their resistance to antifungals and the risk of developing systemic infections (i.e. sepsis). C14R, a classical pore-forming antimicrobial peptide (AMP), shows promising activities against both planktonic and biofilm forms of Candida. The 16-amino acid peptide disrupts cell membranes by integrating into lipid bilayers and forming pore-like structures. Studies demonstrate that C14R reduces the viability of C. albicans, C. auris, and C. parapsilosis, including clinical isolates. Here, we show that C14R not only inhibits biofilm growth, but also halts biofilm maturation and significantly reduces the biomass of preformed biofilms by over 70%. These findings suggest C14R could be an effective option for treating severe fungal infections, offering a potential new treatment approach for biofilm-related diseases. Further research is needed to fully understand its biofilm dispersal potential and to optimize its use for future applications as an antifungal in clinical settings.
Keywords:
novel antifungals
; infectious disease
; next-generation antimycotic
1. Introduction
Biofilms formed on various materials as substrates are by far the most relevant and the "normal" lifestyle for microorganisms in general, despite generations of microbiologists considering the traditional microbiological cultivation in Erlenmeyer flasks to be the most favorable condition for their research organisms [1]. In theory, this also holds true for all pertinent pathogenic bacteria and (fungal) eukaryotes. According to estimates, biofilms could be responsible for over 80% of all microbial infections and 65% of hospital-acquired infections [2,3,4]. The ability of microbial biofilms to significantly increase resistance to physical, chemical, and pharmacological stresses is a property that is almost universally acknowledged, making them a major threat in the clinical context, thereby posing considerable socio-economic challenges to health systems not only in underdeveloped countries [5,6]. The commensal yeast Candida albicans is supposedly the most prevalent and important species in clinical fungal infections, making fungi of this genus a class of extremely significant pathogens [7]. Up to 75% of women will face candidiasis at some point in their lives, and C. albicans is responsible for 85–95% of these infections in this cohort of patients [8]. Over the past few decades, other Candida species have become more prevalent in causing severe infections, but the dominant involvement of C. albicans in invasive infections appears to diminish [9,10]. C. parapsilosis is also of high significance among pathogenic yeasts, as it can establish persistent biofilms on catheters, other implanted medical equipment, and generally on any biotic or abiotic surface in a medical facility, posing serious risks to patients, particularly upon invasive surgery. Furthermore, as C. parapsilosis is metabolically sustained perfectly by the complete parenteral nutrition of patients (e.g. in intensive care units), it represents a significant risk to undernourished infants and newborns with low birth weights [9,10,11,12,13]. Additionally, it has been documented that several clinical isolates of this species become less susceptible even to “gold-standard” types of antifungal medications, severely restricting the effectiveness of established treatment regimens [14,15,16,17]. However, Candida auris, which was initially isolated from a Japanese patient in 2009, is a more recent pathogenic yeast and is, so far, still among the rarest types of clinically relevant Candida species [18]. In hospitalized patients, C. auris can result in serious bloodstream infections with a strikingly high mortality rate of 35% to 60% [19,20]. The fact that strains of C. auris with diverse drug-resistances (likely due to activation of ABC-type efflux pumps [21]) against common antifungals have independently emerged in various nations/continents across the world [22] poses a unique challenge to global pharmacology. As a result, C. auris has been identified as an emergent "superbug" by both the European Center for Disease Control (ECDC) and the U.S. Center for Disease Control (CDC), who have issued clinical advisories and started a widespread public conversation [23]. In 2022, the World Health Organization (WHO) published a new and alarming ranking of “fungal priority pathogens”, being C. albicans, C. auris, and C. parapsilosis at the top of this “hit list” [24]. ABC-type MFS (major facilitator superfamily) efflux pumps have been implicated in multidrug resistance of C. auris to traditional antifungal medications such as fluconazole or amphotericin B [21], and they are especially overexpressed in C. auris when it resides in biofilms [25]. These reports suggest that – in addition to resistance to traditional antifungal drugs – biofilm formation and the ensuing increase in physiological robustness are important aspects of virulence that help both, common and uncommon Candida species, to successfully establish their full pathogenic potential. Treatment of biofilm-based infections is particularly challenging, as currently, there are no medications that specifically target biofilms of Candida species or any other microorganism [26]. We believe that although systematic research to isolate dedicated anti-biofilm medication compounds against Candida species is still underestimated in its importance by the scientific community, it is urgently needed. Basically, the development of complex microbial biofilms can be divided into four stages: the attachment of planktonic cells to the biotic or abiotic biofilm substratum (adherence), cell assembly or aggregation and subsequent growth into structures resembling microcolonies (growth initiation), the production of species-specific mature biofilm architectures (maturation), and the release of the former biofilm cells into the planktonic phase (dispersal) (fig. 1). Antimicrobial peptides (AMPs) are a promising class of novel pharmaceutical compounds that can be used to combat both planktonic Candida cells and their biofilms. The majority of AMPs, including the peptide C14R reduce the functional integrity of microbial cell membranes by interacting with negatively charged phospholipids, thereby integrating into the lipid bilayer, forming pore-like structures and thus disintegrating the functional architecture of the cell envelope [27,28,29]. C14R is a derivative of the previously published antibacterial peptide BP100. It consists of an amphiphilic sequence of 16 amino acid residues with a molecular weight of 2 kDa and a predicted secondary structure of a single α-helix (Fig. 1A) [27,29,30,31,32]. In our previous studies, C14R has proved its potential to reduce cell viability of the Gram-negative human pathogen Pseudomonas aeruginosa and C. albicans as well as C. auris, including clinical isolates of these highly relevant species [27,29]. The principal activity of cell killing does not necessarily coincide with the ability to inhibit biofilm growth and/or maturation, especially not with biofilm dispersal. The latter process represents a decay of these elaborate structures by releasing viable cells into the environment, which can colonize new habitats, a physiological event with enormous relevance for virulence [33]. In this short communication, we want to share initial but, in our opinion, promising results demonstrating, for the first time, that C14R can inhibit the growth of C. albicans, C. auris, and C. parapsilosis biofilms. More important is the finding that not only ab initio growth of biofilms was inhibited in the presence of C14R, but the maturation of C. albicans biofilms could be arrested, and preformed biofilms of C. auris and C. parapsilosis could even be reduced in their biomass to less than 30%. Although in-depth characterization of the C14R biofilm dispersal potential is required, including kinetic studies of the dose-dependent induction of biofilm decay and the release of cells including a large ensemble of clinical isolates, we already believe that C14R alone or in combination with other (peptide) drugs may open a valuable option for the development of new treatment regimens for these severe fungal infections.
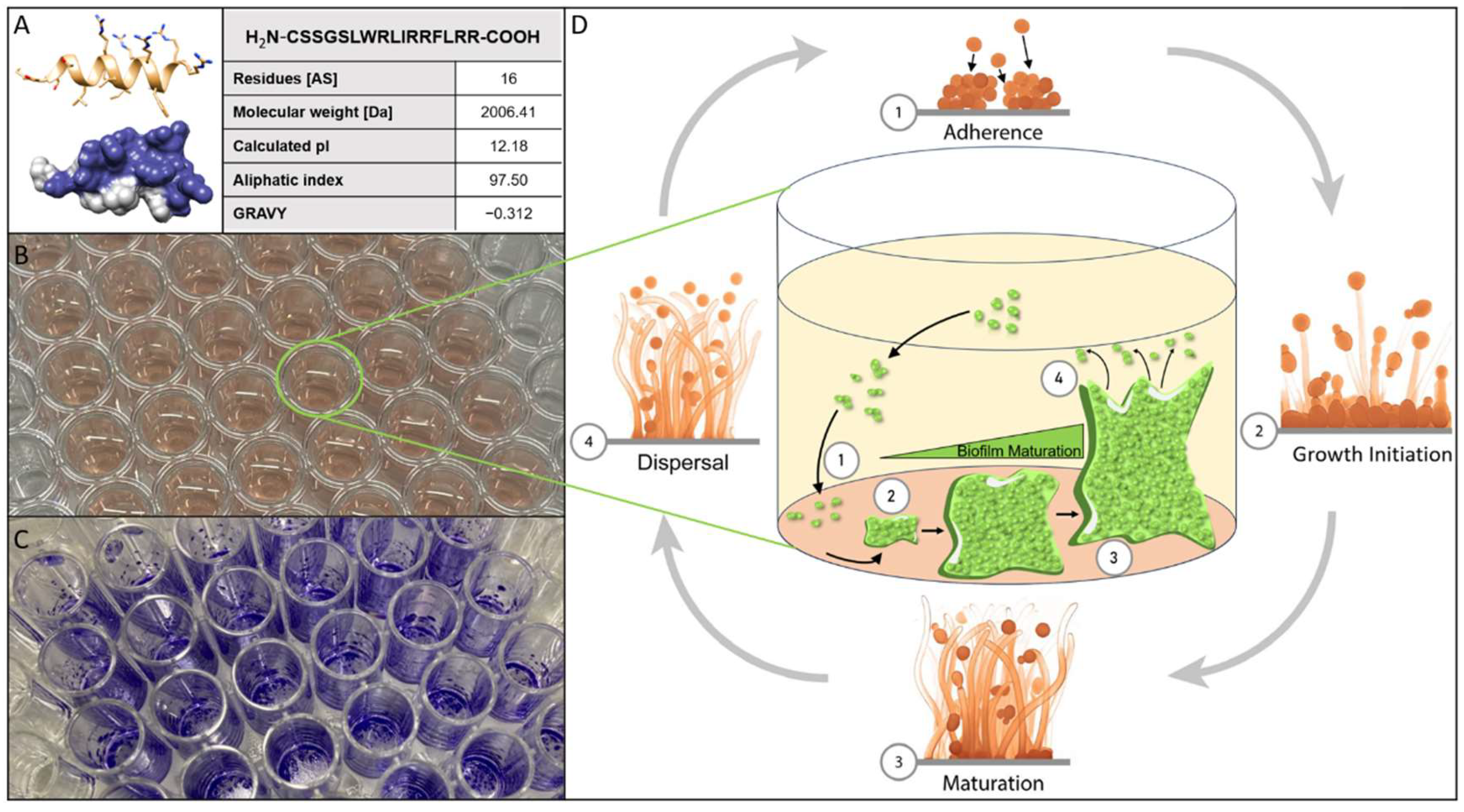
Figure 1.
(A) 3D prediction gained via an ab initio method using AlphaFold2 from Google Colabs (https://colab.research.google.com/github/sokrypton/ColabFold/) of the peptide C14R with the sequence NH2-CSSGSLWRLIRRFLRR, as well as the following properties: amino acid sequence, length, theoretical isoelectric point (pI), the aliphatic index and the grand average hydropathy index (GRAVY) of C14R calculated with ExPASy ProtParam. (B) Example of a 96-well polystyrene microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) filled with 200 µL of RPMI-1640 media and inoculated with cells of C. albicans. (C) Example of 96- well microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) after performing the staining of C. albicans biofilm cells with crystal violett. (D) Schematic overview of the lifecycle of Candida biofilm in the four steps adherence, growth initiation, maturation and dispersal.
Figure 1.
(A) 3D prediction gained via an ab initio method using AlphaFold2 from Google Colabs (https://colab.research.google.com/github/sokrypton/ColabFold/) of the peptide C14R with the sequence NH2-CSSGSLWRLIRRFLRR, as well as the following properties: amino acid sequence, length, theoretical isoelectric point (pI), the aliphatic index and the grand average hydropathy index (GRAVY) of C14R calculated with ExPASy ProtParam. (B) Example of a 96-well polystyrene microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) filled with 200 µL of RPMI-1640 media and inoculated with cells of C. albicans. (C) Example of 96- well microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) after performing the staining of C. albicans biofilm cells with crystal violett. (D) Schematic overview of the lifecycle of Candida biofilm in the four steps adherence, growth initiation, maturation and dispersal.
2. Materials and Methods
2.1. Materials
Acetic acid, crystal violet, 3-(N-morpholino) propanesulfonic acid (MOPS), and paraformaldehyde were obtained from Carl Roth GmbH (Karlsruhe, Germany). The fluorescent dye fluorescein isothiocyanate (FITC) and RPMI-1640 medium containing L-glutamine were acquired from Thermo Fisher Scientific (Waltham, MA, USA). Resazurin sodium salt was procured from Sigma-Aldrich Chemie GmbH (Steinheim, Germany). Phosphate-buffered saline (PBS) was purchased from Life Technologies (Carlsbad, CA, USA). The peptide C14R was chemically synthesized by the Core Facility Functional Peptidomics (University of Ulm, Germany).
2.2. Candida Cultivation
For each experiment, the strain of interest C. albicans (ATCC90028), C. auris (DSMZ-No. 21092) or C. parapsilosis (ATCC22019) was cultivated by inoculating 100 µL from a cryoprotectant glycerol culture in 5 mL RPMI-1640 media at 37 °C for 18 h and orbital shaking at 150 rpm.
2.3. Biofilm Kinetic Assay/ Crystal-Violet-Assay
To analyze the kinetics of biofilm formation, 2.5 × 10³ cells of C. albicans, C. auris and C. parapsilosis were inoculated in 200 µL of RPMI-1640 medium in a flat-bottomed 96-well polystyrene microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) and incubated at 37 °C without agitation. Biofilm formation was quantified at 2, 4, 6, 8, 24, 48, 72 and 96 hours of incubation in triplicates, using the crystal violet assay initially developed for bacterial biofilms by George O’Toole [34] and widely applied to Candida biofilms [35,36,37]. In brief the planktonic phase was removed and biofilms were washed twice with 200 µL of water. The biofilms were then stained for 15 minutes with 200 µL of a 0.1% (w/v) crystal violet solution. After removing the supernatant, the biofilms were washed twice with 200 µL of water. The stained biofilms were air-dried for 24 hours at 25 °C. The remaining crystal violet was dissolved in 200 µL of 30% acetic acid for 15 minutes and transferred into a new 96-well plate. Subsequently, the absorbance was measured at 560 nm using a Tecan Infinite F200 microplate reader (Tecan Group Ltd., Männedorf, Switzerland) to quantify the biofilm biomass. At the given time points, the planktonic phase, as well as the biofilms, were examined by phase-contrast microscopy at 400× magnification with the Leica DMi8 inverted fluorescent microscope (Leica Microsystems CMS GmbH, Wetzlar, Germany). Additionally, the OD600 value of the planktonic phase was measured at the time points 0, 2, 8, 24, 48, 72, and 96 hours of incubation using a Tecan Infinite F200 microplate reader (Tecan Group Ltd., Männedorf, Switzerland).
2.4. Resazurin-Reduction-Assay/Viability-Assay
The viability of C. parapsilosis cells in the presence of various concentrations of C14R (0–200 µg/mL) was assessed similarly to the “Clinical and Laboratory Standards Institute” guidelines M27-A3 broth microdilution assay[38]. by incubating 2.5 × 10³ yeast cells in triplicates in 200 µL of C14R supplemented RPMI-1640 media in flat-bottomed 96-well polystyrene microtiter plates (Sarstedt AG & Co. KG, Nümbrecht, Germany) at 37°C for 24 h and orbital shaking at 900 rpm on an Eppendorf shaker. Viability was quantified using the Resazurin reduction assay [39,40]. In short, the cells were incubated for 2h at 37°C with 20 µL of 0.15mg/mL resazurin solution. Viable cells convert resazurin to resorufin, which fluorescence can be measured at an excitation wavelength of 535 nm and an emission of 595 nm with a Tecan infinite F200 microplate reader (Tecan Group Ltd., Männedorf, Switzerland).
2.5. Permeabilization Assay
A permeabilization assay to asses the ability of C14R to form pores in the membranes of C. auris and C. parapsilosis was performed as described previously [27,29]. For this purpose, the fluorescent dye Fluorescein isothiocyanate (FITC), with a molecular weight of 389 Da, was utilized. 107 cells of C. auris and C. parapsilosis were incubated in triplicates at 37°C in 200 µL RPMI-1640 media supplemented with 12 µg/mL of C14R. Non-supplemented RPMI was used as a control. After 2 h of incubation, the cells were centrifuged at 11 000 x g, and the resulting pellet was washed once with PBS. The cells were then incubated at room temperature with 200 µL of FITC at a final concentration of 125 µg/mL in 1x PBS for 20 min, followed by centrifugation at 11 000 x g for 2 min. After discarding the supernatant, the yeast cells were fixed by incubation with 4% (w/v) paraformaldehyde in PBS for 10 min and washed three times with PBS. Eventually, the in 200 µL PBS resuspended cells were transferred in a flat-bottomed polystyrene 96 wells microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) and fluorescence was measured using a Tecan SPARK microplate reader (Tecan Group Ltd., Männedorf, Switzerland) at an excitation wavelength of 498 nm and emission wavelength of 517 nm.
2.6. Inhibition of biofilm formation Assay
The analysis of the peptides' antifungal effect on biofilm formation was evaluated under similar conditions as previously outlined [34,37,40]. In summary, triplicates of 2.5 × 10³ cells were incubated in 200 µL of RPMI-1640 media, supplemented with the corresponding peptide concentrations (0- 200 µg), in flat-bottomed 96-well polystyrene microtiter plates (Sarstedt AG & Co. KG, Nümbrecht, Germany) at 37°C for 24 hours. Afterward, the crystal violet assay was performed as described earlier.
2.7. Decay of preformed biofilm Assay
An experiment was conducted to assess the ability of C14R to decay preformed Candida biofilms. In the first phase, 2.5 × 10³ Candida cells were incubated in 200 µL of RPMI-1640 media in a flat-bottomed 96-well polystyrene microtiter plate (Sarstedt AG & Co. KG, Nümbrecht, Germany) at 37°C for 24 hours without agitation. The biomass of the formed biofilms was quantified using the Crystal Violet assay. Biofilm formation was quantified in triplicates for each strain of Candida and served as a reference for biofilm formation within 24h.
In the second phase, the supernatants in the remaining wells of the first phase were carefully replaced with 200 µL of RPMI-1640 medium containing Amphotericin B (2 µg/mL) or C14R at concentrations ranging from 0 to 400 µg/mL (also in triplicates). These wells were then incubated at 37°C for an additional 24 hours to allow C14R treatment. After this second incubation phase, the Crystal Violet assay was repeated to assess any reduction of the biofilm mass, comparing it to the reference biofilm mass before C14R treatment.
3. Results
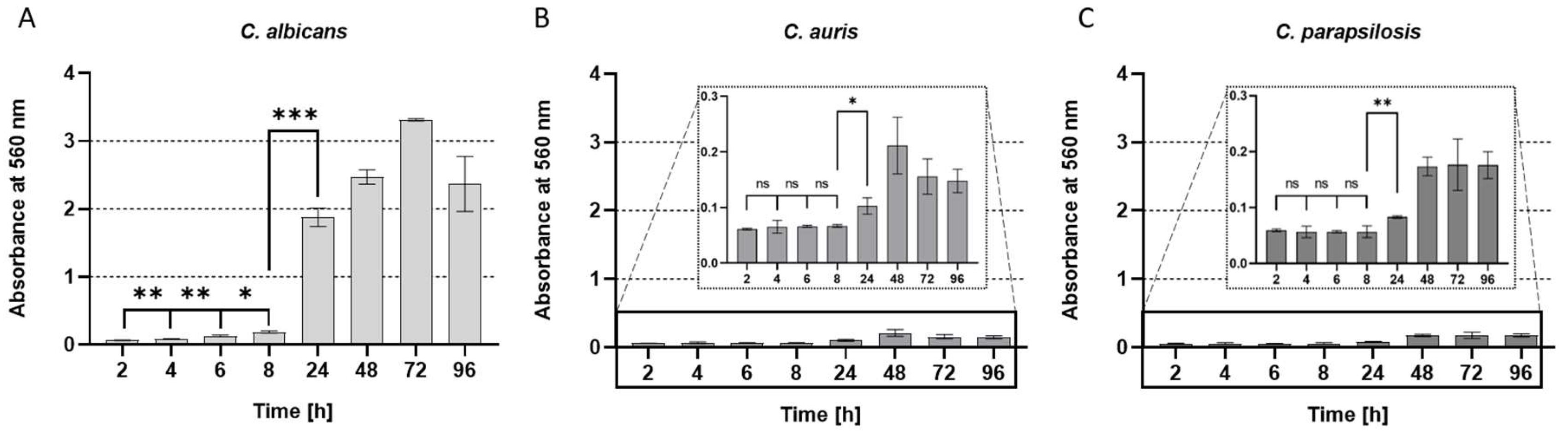
Formation of (elaborate) biofilms is the physiological process involving four distinct phases: adherence, growth initiation, maturation, and dispersal as the final phase, initiating the repetition of this process on new surfaces. The time needed for microbes to build up the final stages of the respective biofilm is individually different, and it depends on the species that have intrinsic abilities to form biofilms, the surface involved, the medium, and the incubation regime (i.e. aeration). The reference strains of C. albicans, C. auris, and C. parapsilosis were allowed to form biofilms on the polystyrene surfaces of commercial microtiter-plates in Roswell Park Memorial Institute (RPMI) 1640 Medium [38] which was originally developed for human cell culture techniques and thus represents a reasonable approximation of nutritional conditions the pathogenic yeasts will normally face in the human body (e.g. on freshly infected wounds). The kinetics of biofilm development and thus the increase of biomass as a measure for the fungal growth on the abiotic surface polystyrene was recorded for 96 hours with sampling intervals of two hours in the adherence phase and 24 hours in later stages of the biofilm formation process as proved reasonable in our previous studies [40,41,42,43,44](alle Biofilm Paper Rosenau). C. auris and C. parapsilosis formed clearly detectable biofilms at comparable intensities for each of these species (fig. 2 B, C). However, the C. auris and C. parapsilosis biofilms produced drastically lower amounts as compared to C. albicans (fig. 2 A), which is one of the most prominent biofilm-forming yeasts in literature [45]. As expected, C. albicans showed the most pronounced biofilm formation capability under these conditions, with a kinetic following the scheme that a “lag-phase” occurred between inoculation (t = 0 hours and 8 hours), which in this case represented the adherence and early growth initiation phases at the onset of biofilm development, then proceeding into the maturation phase. This important switching point was accordingly observable after a growth period of 24 hours, thereby qualifying the period 24-48 hours as the most relevant and critical stage [33]. Accordingly, the cell numbers (measured as the optical density [O.D.600] spectrophotometrically at a wavelength of 600 nm) in the liquid part of the cultures (i.e. the planktonic phase) were significantly higher in relation to the respective biofilm masses for C. auris and C. parapsilosis, but rather limited for the best biofilm performer C. albicans (fig. S1). The optical inspection by classical light microscopy of samples from the planktonic phases at each time point of the growth curves as well as for the biofilms, were in agreement with the quantitative measurements and supported these results adequately (fig. S2).
Figure 2.
Kinetic of biofilm formation at the time points 2, 4, 6, 8, 24, 48, 72 and 96 hours of (A) C. albicans, (B) C. auris and (C) C. parapsilosis, all determined using the crystal violet assay. All experiments were conducted in triplicate with error bars representing standard deviations. All experiments were conducted in triplicate with error bars representing standard deviations. p values < 0.05 were considered significant. ns denotes not significant. * p denotes < 0.05, ** denotes p < 0.01, and *** denotes p < 0.001.
Figure 2.
Kinetic of biofilm formation at the time points 2, 4, 6, 8, 24, 48, 72 and 96 hours of (A) C. albicans, (B) C. auris and (C) C. parapsilosis, all determined using the crystal violet assay. All experiments were conducted in triplicate with error bars representing standard deviations. All experiments were conducted in triplicate with error bars representing standard deviations. p values < 0.05 were considered significant. ns denotes not significant. * p denotes < 0.05, ** denotes p < 0.01, and *** denotes p < 0.001.
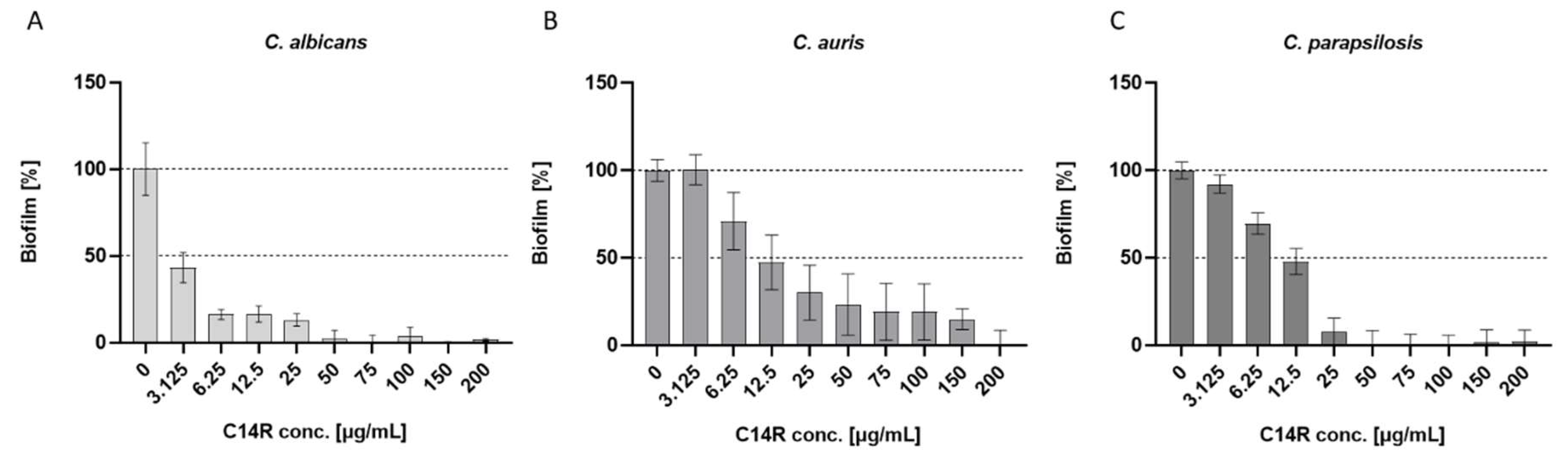
The peptide C14R was shown to be functional as a classical pore-forming antimicrobial compound with the ability to kill pathogens like the bacterium P. aeruginosa and the yeasts C. albicans and C. auris, whereas this remained open so far for C. parapsilosis. Therefore, we performed the respective measurement with cells grown in the planktonic phase also with this yeast and could determine C14R to be also active against it with a peptide concentration of 175 µg/mL sufficient to completely kill C. parapsilosis (fig. S3). The pore-forming ability in bacterial and yeast cells has previously been demonstrated using the cell (membrane) permeation assay using the otherwise not membrane penetrating fluorescent dye fluorescein isothiocyanate (FITC) [27,29]. This assay was also fully functional for planktonic C. auris and C. parapsilosis and allowed to judge C14R as a pore forming antimicrobial peptide also for these yeasts (fig. S4). The ability to reduce the viability of planktonic cells does not necessarily allow the conclusion that a respective compound also has anti-biofilm activities. This is especially the case if one intention is to support or induce the dispersal of preformed biofilms (i.e. destroy premature or matured elaborate architectures) with maturation times playing potentially an important role for this process. The principal ability to inhibit biofilm formation per se was shown by growing biofilms of C. albicans, C. auris and C. parapsilosis completely in the presence of C14R, which was added simultaneously with the inoculation of the cultures and determining the biofilm mass using again the crystal violet staining assay. The MBIC (minimal biofilm inhibitory concentrations), i.e. the lowest concentration resulting in complete inhibition of biofilm formation, was found to be 50 µg/mL of C14R for C. albicans and C. parapsilosis (fig. 3 A,C), whereas C. auris required four times higher concentrations of 200 µg/mL (fig. 3B). The total inhibition of biofilm growth in the presence of inhibitory compounds includes the very early phases of adherence to the respective surface, the subsequent establishment of productive cell-cell and/or cell-substratum interactions, which can be influenced by the respective drug.
Figure 3.
Inhibition of biofilm formation by incubation of Candida cells with the antimicrobial peptide C14R simultaneously to inoculation. Graphs show the formed biofilms after an incubation time of 24 hours. Biofilm mass was analyzed using the crystal violet assay. The resulting effective MBIC dose was 50 µg/mL for C. albicans (A), 200 µg/mL for C. auris (B) and 50 µg/mL for C. parapsilosis (C).
Figure 3.
Inhibition of biofilm formation by incubation of Candida cells with the antimicrobial peptide C14R simultaneously to inoculation. Graphs show the formed biofilms after an incubation time of 24 hours. Biofilm mass was analyzed using the crystal violet assay. The resulting effective MBIC dose was 50 µg/mL for C. albicans (A), 200 µg/mL for C. auris (B) and 50 µg/mL for C. parapsilosis (C).
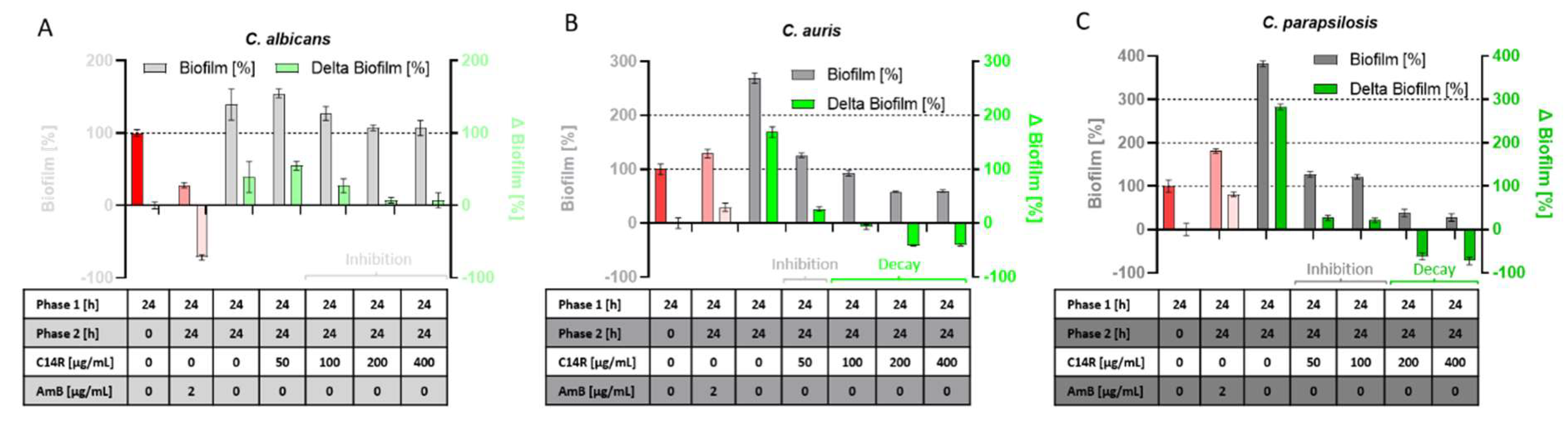
In contrast, the undisturbed early development of biofilms towards the switching point (24-48 hours) results in the onset of a maturation process leading to the final architecture and thus represents an essential phase providing stability and robustness to the cell community, which is afterwards only elaborated and developed to perfection. Thus, growing biofilms in this phase (after 24 hours) represent a challenging target for anti-biofilm compounds, which in principle can influence further growth or, in extreme cases, even induce biofilm decay or at least help to dissolve the preformed initial biofilm. To determine the effect of C14R on C. albicans, C. auris, and C. parapsilosis preformed biofilms, we divided the analysis into two distinct phases with “phase 1” including the period from inoculation (t = 0 hours) to 24 hours without C14R (red bars in fig. 4) and “phase 2” including additional 24 hours (to 48 hours finally) as a growth surplus in the presence of C14R (fig. 4 A, B, C). The classical antifungal drug Amphotericin B served as a control since its mode of action, like for C14R, is also the disintegration of the fungal cell membrane. However, the activity of this polyene results from binding to ergosterol as an essential membrane phospholipid, indirectly leading to membrane instabilities and finally cell death. Although Amphotericin B was functional in reducing biofilm mass in phase 2 in cultures of C. albicans, it completely failed for C. auris and C. parapsilosis, respectively (fig. 4). In contrast, C14R was not able to reduce biofilm mass for C. albicans, but it managed to control and significantly inhibit further growth and intensifying biomass development in phase 2, in comparison to the untreated control, which as expected gained additional biofilm mass (fig. 4A). This effect was dose dependent and appeared visible for concentrations ≥ 100 µg/mL. More important was the finding that C14R was not only active in inhibiting the gain of a surplus of C. auris and C. parapsilosis biofilm mass by further growth after the addition of the peptide, but also showed to be functional in reducing biofilm mass in phase 2, which had been pre-produced in phase 1 at concentrations of ≥ 100 µg/mL for C. auris and ≥ 200 µg/mL for C. parapsilosis (fig. 4 B and C, labeled with “decay”). These biofilm reductions in phase 2 were as drastic as 42% for C. auris and up to 73% for C. parapsilosis, as compared to the respective untreated controls.
Figure 4.
Inhibition of further biofilm growth in the incubation phase with C14R, phase 2 (24-48 hours) compared to the biofilm mass of phase 1 (24 hours of incubation without C14R, red bar). For C. albicans (A) an inhibition of gain in biofilm-biomass could be detected at concentrations ≥ 100 µg/mL. For C. auris (B) concentration of 50 µg/mL led to an inhibition of further biofilm growth while concentrations ≥ 100 µg/mL could even decay the biomass compared to the phase 1 biofilm mass. The concentration for inhibition of further growth of C. parapsilosis (C) were 50-100 µg/mL while ≥ 200 µg/mL led to a biofilm decay of C. parapsilosis.
Figure 4.
Inhibition of further biofilm growth in the incubation phase with C14R, phase 2 (24-48 hours) compared to the biofilm mass of phase 1 (24 hours of incubation without C14R, red bar). For C. albicans (A) an inhibition of gain in biofilm-biomass could be detected at concentrations ≥ 100 µg/mL. For C. auris (B) concentration of 50 µg/mL led to an inhibition of further biofilm growth while concentrations ≥ 100 µg/mL could even decay the biomass compared to the phase 1 biofilm mass. The concentration for inhibition of further growth of C. parapsilosis (C) were 50-100 µg/mL while ≥ 200 µg/mL led to a biofilm decay of C. parapsilosis.
4. Discussion
Among clinically relevant Candida species, C. albicans undoubtedly remains the major pathogen and the most commonly isolated etiological agent of candidiasis [46,47]. However, its dominance is gradually being overtaken by non-albicans species, particularly C. parapsilosis, which exhibits a notable ability to form robust biofilms on both biotic and abiotic surfaces, such as medical devices like catheters and prostheses [9,10,13]. The WHO “alarmed” set of today’s high-priority fungal pathogens is completed by C. auris having initiated striking terror into human societies predominantly in tropic and subtropic regions with the potential to spread into other regions of the world as the first multidrug-resistant fungal zoonosis emerging from climate change [48]. Consequently, it suits humanity well to redirect research to finding novel antimicrobial drugs dedicated not only to Candida species per se but also to the development of biofilm dedicated future medications including also the more than considerable virulence factor of forming biofilms reducing susceptibility towards antimicrobial agents and effective control of the pathogens by the immune system of the host.
Two different sets of experiments were performed to characterize the biofilm control activity of C14R on the three model Candida species. One set of experiments was performed to quantify biofilm growth and development when the peptide was present in the cultures from the beginning (added at the inoculation). In the second set, biofilms were allowed to preform for one day, with the subsequent addition of the drug and measurement of further growth for one additional day. Inhibition of ab initio growing biofilms was effectively possible for the three species with C14R concentrations leading to a complete embellishment of biofilm mass after 24 hours of 50 µg/mL for C. albicans and C. parapsilosis and 200 µg/mL for C. auris. Growing in biofilms provides (pathogenic) microbes with an increase of resistance towards antimicrobial compounds, which can be 10-1000-fold as compared to their planktonically growing counterparts, a fact that perfectly describes the challenge in clinical microbiology to fight those microbial encapsulated communities (nicely reviewed in [49]). The minimal inhibitory concentrations (MIC) of C14R have been determined in previous studies for sets of clinical isolates of C. albicans and C. auris being in the range of 5 µg/mL [29], qualifying the biofilm inhibitory concentrations observed here (50-200 µg/mL) as perfectly convincing with respect to this general description. A challenging task widely accepted as such in literature is the eradication of preformed biofilms as the next level of biofilm inhibition and control. In this regards, we performed the assays for the inhibition of further growth and the contingent (and desired) induction of biofilm decay. C14R can prevent further growth of the preformed biofilms of C. albicans with high efficiency at concentrations above 100 µg/mL, as indicated by a remarkable reduction of detectable biomass. The preventive effect of C14R on biofilms gaining a surplus of mass was also present for C. auris and C. parapsilosis at concentrations above 50 µg/mL. However, the crucial difference became obvious at higher concentrations of the peptide when growth inhibition switched to a real reduction (i.e. a net decay) of the initial biofilm mass at the beginning of the experiment measured prior to the addition of C14R. Interestingly, Amphotericin B, as a non-peptide antifungal which served as a control in these experiments, was able to reduce biofilms significantly in the case of C. albicans but failed completely in biofilm decay for C. auris and C. parapsilosis. This advantage of the polyene macrocyclic lactone Amphotericin B as a smaller and less charged molecule over C14R solely in the case of C. albicans may be primarily the result of an impaired penetration of antifungal agents within the biofilm structure, which has been discussed as a general property of microbial biofilms [50] with enhanced levels of extracellular chemical compounds (i.e. biopolymers) in the biofilm matrix also designated the “matrixome” and functional as a molecular sieve [51]. Surprisingly, in contrast to the results for C. albicans Amphotericin B was not functional to abolish biofilm on-growth and maturation for the less massive biofilms of C. auris and C. parapsilosis, although the strains used were also non-resistant laboratory reference strains. Regardless of the fact that the results presented here originate from experiments with so far not optimized parameters and taking into account that follow-up studies are required using large ensembles of clinical (and multi-drug resistant) isolates, measuring detailed dose-dependencies and biofilm decay kinetics, the undoubtably observed destruction of preformed biofilms qualifies C14R as a probable upcoming drug for the control of biofilms formed by C. auris and C. parapsilosis. Previously we have demonstrated that a group of noval neutralizing peptides can effectively inhibit the formation of C. albicans biofilms [40,41,43,52]. We believe that C14R action can be further improved by molecular modifications and also by combining with these neutralizing peptides. It may open new avenues to eradicate C. albicans from biofilms with high efficiency, including multi-resistant variants of the common and highly pathogenic yeast.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Kinetic of planktonic phase, measured OD600 values of the time points 0, 2, 8, 24, 48, 72 and 96 hours are shown of (A) C. albicans, (B) C. auris and (C) C. parapsilosis All experiments were conducted in triplicate with error bars representing standard deviations.; Figure S2: Light microscopy of the planktonic phase as well as the biofilms of (A) C. albicans, (B) C. auris and (C) C. parapsilosis at the time points 2, 4, 6, 8, 24, 48, 72 and 96 hours and at 400× magnification; Figure S3: Viability assay by incubation of C. parapsilosis cells with the antimicrobial peptide C14R simultaneously to inoculation. Graph shows the number of viable cells in percent compared an untreated control after a total incubation time of 24 hours. Viable cells were analyzed using the resazurin assay; Figure S4: Permeation assay of cells of C. auris and C. parapsilosis. Graph shows the fluorescence of Candida cells after the uptake of the fluorescent dye Fluorescein isothiocyanate (FITC). Fluorescence was sufficiently higher after an incubation time of 2 h with C14R compared to an untreated control. All experiments were conducted in triplicate with error bars representing standard deviations. p values < 0.05 were considered significant. * p denotes < 0.05.
Author Contributions
Conceptualization, F.R. and A.-K.K.; methodology, J.-C.W., A.-K.K., D.G., D.A.-P., E.M.M.-H., validation, J.-C.W., A.-K.K., D.G., D.A.-P.; investigation, J.-C.W., A.-K.K., D.G., D.A.-P. E.M.M.-H., N.P., A.R.-A. and A.-K.K.; data curation, D.A.-P., E.M.M.-H., M.K. and V.V.; writing—original draft preparation, J.-C.W., A.-K.K., and F.R. and A.-K.K.; writing—review and editing, J.-C.W., A.-K.K., D.G., A.R.-A., L.S., C.K., W.K., S.S., C.F., and F.R.; supervision, F.R.; project administration, F.R.; funding acquisition, L.S., C.K., W.K. and F.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the German Research Society (DFG) project 465229237 and project CRC1279 (number 316249678) (exploiting the Human Peptidome for Novel Antimicrobial and Anticancer Agents), the Alexander von Humboldt-Stiftung, Research Group Linkage Program (No. 1160914-COL-IP). We are also grateful to DAAD-Germany for facilitating the financial support of the German Ministry for Foreign Affairs via the program Global Health and Pandemic Prevention Centers (project 57592717-GLACIER). This work was also supported by the Gesellschaft für Forschungsförderung (GFF) of Lower Austria as part of the project “Aptamers and Odorant Binding Proteins – Innovative Receptors for Electronic Small Ligand Sensing” (FTI22-G-012). This work was also supported by the Austrian Research Promotion Agency (FFG) within the COMET Project “PI-SENS” (Project No 915477) as well as by the Federal Provinces of Lower Austria and Tirol.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and Supplementary Material.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nadell, C.D.; Bassler, B.L.; Levin, S.A. Observing Bacteria through the Lens of Social Evolution. J Biol 2008, 7, 27. [Google Scholar] [CrossRef]
- Sharma, S.; Mohler, J.; Mahajan, S.D.; Schwartz, S.A.; Bruggemann, L.; Aalinkeel, R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms 2023, 11, 1614. [Google Scholar] [CrossRef]
- Davies, D. Understanding Biofilm Resistance to Antibacterial Agents. Nat Rev Drug Discov 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Fox, E.P.; Nobile, C.J. A Sticky Situation: Untangling the Transcriptional Network Controlling Biofilm Development in Candida Albicans. Transcription 2012, 3, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.; Nobile, C. The Role of Candida Albicans Biofilms in Human Disease; Nova Science Publishers, Inc., 2013; ISBN 9781628088823.
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial Biofilms: From the Natural Environment to Infectious Diseases. Nat Rev Microbiol 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and Emerged Pathogenic Candida Species: Beyond the Candida Albicans Paradigm. PLoS Pathog 2013, 9, e1003550. [Google Scholar] [CrossRef]
- Sobel, J.D. Vulvovaginal Candidosis. The Lancet 2007, 369, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Paramythiotou, E.; Frantzeskaki, F.; Flevari, A.; Armaganidis, A.; Dimopoulos, G. Invasive Fungal Infections in the ICU: How to Approach, How to Treat. Molecules 2014, 19, 1085–1119. [Google Scholar] [CrossRef] [PubMed]
- Tóth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vágvölgyi, C.; Gácser, A. Candida Parapsilosis: From Genes to the Bedside. Clin Microbiol Rev 2019, 32. [Google Scholar] [CrossRef]
- Pammi, M.; Holland, L.; Butler, G.; Gacser, A.; Bliss, J.M. Candida Parapsilosis Is a Significant Neonatal Pathogen. Pediatric Infectious Disease Journal 2013, 32, e206–e216. [Google Scholar] [CrossRef] [PubMed]
- Weems, J.J.; Chamberland, M.E.; Ward, J.; Willy, M.; Padhye, A.A.; Solomon, S.L. Candida Parapsilosis Fungemia Associated with Parenteral Nutrition and Contaminated Blood Pressure Transducers. J Clin Microbiol 1987, 25, 1029–1032. [Google Scholar] [CrossRef] [PubMed]
- Harrington, R.; Kindermann, S.L.; Hou, Q.; Taylor, R.J.; Azie, N.; Horn, D.L. Candidemia and Invasive Candidiasis among Hospitalized Neonates and Pediatric Patients. Curr Med Res Opin 2017, 33, 1803–1812. [Google Scholar] [CrossRef]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of Azole-Resistant Candida Parapsilosis Causing Bloodstream Infection: Results from Laboratory-Based Sentinel Surveillance in South Africa. Journal of Antimicrobial Chemotherapy 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [PubMed]
- Cantón, E.; Pemán, J.; Quindós, G.; Eraso, E.; Miranda-Zapico, I.; Álvarez, M.; Merino, P.; Campos-Herrero, I.; Marco, F.; de la Pedrosa, E.G.G.; et al. Prospective Multicenter Study of the Epidemiology, Molecular Identification, and Antifungal Susceptibility of Candida Parapsilosis, Candida Orthopsilosis, and Candida Metapsilosis Isolated from Patients with Candidemia. Antimicrob Agents Chemother 2011, 55, 5590–5596. [Google Scholar] [CrossRef] [PubMed]
- Meletiadis, J.; Curfs-Breuker, I.; Meis, J.F.; Mouton, J.W. In Vitro Antifungal Susceptibility Testing of Candida Isolates with the EUCAST Methodology, a New Method for ECOFF Determination. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Moet, G.J.; Messer, S.A.; Jones, R.N.; Castanheira, M. Candida Bloodstream Infections: Comparison of Species Distributions and Antifungal Resistance Patterns in Community-Onset and Nosocomial Isolates in the SENTRY Antimicrobial Surveillance Program, 2008-2009. Antimicrob Agents Chemother 2011, 55, 561–566. [Google Scholar] [CrossRef]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida Auris Sp. Nov., a Novel Ascomycetous Yeast Isolated from the External Ear Canal of an Inpatient in a Japanese Hospital. Microbiol Immunol 2009, 53, 41–44. [Google Scholar] [CrossRef]
- Morales-López, S.E.; Parra-Giraldo, C.M.; Ceballos-Garzón, A.; Martínez, H.P.; Rodríguez, G.J.; Álvarez-Moreno, C.A.; Rodríguez, J.Y. Invasive Infections with Multidrug-Resistant Yeast Candida Auris, Colombia. Emerg Infect Dis 2017, 23, 162–164. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clinical Infectious Diseases 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-Resistant Candida Haemulonii and C. Auris, Tel Aviv, Israel. Emerg Infect Dis 2017, 23. [Google Scholar] [CrossRef] [PubMed]
- Osei Sekyere, J. Candida Auris: A Systematic Review and Meta-analysis of Current Updates on an Emerging Multidrug-resistant Pathogen. Microbiologyopen 2018, 7. [Google Scholar] [CrossRef]
- A New Fungal Superbug Emerging as a Global Threat. Available online: Https://Www.Forbes.Com/Sites/Judystone/2017/08/24/Candida-Auris-a-New-Fungal-Superbug-Emerging-as-a-Global-Threat/ (accessed on day month year).
- WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action. Geneva: World Health Organization; 2022. Licence: CC BY-NC-SA 3.0 IGO.;
- Kean, R.; Delaney, C.; Sherry, L.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R.; Williams, C.; Ramage, G. Transcriptome Assembly and Profiling of Candida Auris Reveals Novel Insights into Biofilm-Mediated Resistance. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Johnson, A.D. Candida Albicans Biofilms and Human Disease. Annu Rev Microbiol 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Mildenberger, V.; Alpízar-Pedraza, D.; Martell-Huguet, E.M.; Krämer, M.; Bolotnikov, G.; Otero-Gonzalez, A.J.; Weil, T.; Rodriguez-Alfonso, A.; Preising, N.; Ständker, L.; et al. The Designed Pore-Forming Antimicrobial Peptide C14R Combines Excellent Activity against the Major Opportunistic Human Pathogen Pseudomonas Aeruginosa with Low Cytotoxicity. Pharmaceuticals 2024, 17, 83. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil Med Res 2021, 8, 48. [Google Scholar] [CrossRef]
- Vélez, N.; Argel, A.; Kissmann, A.-K.; Alpízar-Pedraza, D.; Escandón, P.; Rosenau, F.; Ständker, L.; Firacative, C. Pore-Forming Peptide C14R Exhibits Potent Antifungal Activity against Clinical Isolates of Candida Albicans and Candida Auris. Front Cell Infect Microbiol 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Bodenberger, N.; Kubiczek, D.; Halbgebauer, D.; Rimola, V.; Wiese, S.; Mayer, D.; Rodriguez Alfonso, A.A.; Ständker, L.; Stenger, S.; Rosenau, F. Lectin-Functionalized Composite Hydrogels for “Capture-and-Killing” of Carbapenem-Resistant Pseudomonas Aeruginosa. Biomacromolecules 2018, 19, 2472–2482. [Google Scholar] [CrossRef]
- Bagherabadi, M.; Fleckenstein, M.; Moskalyk, O.; Belluati, A.; Avrutina, O.; Andrieu-Brunsen, A. Grafting and Controlled Release of Antimicrobial Peptides from Mesoporous Silica. J Mater Chem B 2024, 12, 8167–8180. [Google Scholar] [CrossRef]
- Kraemer, M.; Bellion, M.; Kissmann, A.-K.; Herberger, T.; Synatschke, C. V.; Bozdogan, A.; Andersson, J.; Rodriguez, A.; Ständker, L.; Wiese, S.; et al. Aptamers as Novel Binding Molecules on an Antimicrobial Peptide-Armored Composite Hydrogel Wound Dressing for Specific Removal and Efficient Eradication of Pseudomonas Aeruginosa. Int J Mol Sci 2023, 24, 4800. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida Albicans Biofilm Growth and Dispersal: Contributions to Pathogenesis. Curr Opin Microbiol 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. Journal of Visualized Experiments 2011. [Google Scholar] [CrossRef]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms Formed by Candida Albicans Bloodstream Isolates Display Phenotypic and Transcriptional Heterogeneity That Are Associated with Resistance and Pathogenicity. BMC Microbiol 2014, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida Auris. Emerg Infect Dis 2017, 23, 328–331. [Google Scholar] [CrossRef]
- Negri, M.; Gonçalves, V.; Silva, S.; Henriques, M.; Azeredo, J.; Oliveira, R. Crystal Violet Staining to Quantify Candida Adhesion to Epithelial Cells. Br J Biomed Sci 2010, 67, 120–125. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—Third Edition. CLSI Document M27-A3. Wayne, PA: Clinical and Laboratory Standards Institute; 2008.
- González García, M.; Rodríguez, A.; Alba, A.; Vázquez, A.A.; Morales Vicente, F.E.; Pérez-Erviti, J.; Spellerberg, B.; Stenger, S.; Grieshober, M.; Conzelmann, C.; et al. New Antibacterial Peptides from the Freshwater Mollusk Pomacea Poeyana (Pilsbry, 1927). Biomolecules 2020, 10, 1473. [Google Scholar] [CrossRef]
- Amann, V.; Kissmann, A.-K.; Krämer, M.; Krebs, I.; Perez-Erviti, J.A.; Otero-Gonzalez, A.J.; Morales-Vicente, F.; Rodríguez, A.; Ständker, L.; Weil, T.; et al. Increased Activities against Biofilms of the Pathogenic Yeast Candida Albicans of Optimized Pom-1 Derivatives. Pharmaceutics 2022, 14, 318. [Google Scholar] [CrossRef]
- Häring, M.; Amann, V.; Kissmann, A.-K.; Herberger, T.; Synatschke, C.; Kirsch-Pietz, N.; Perez-Erviti, J.A.; Otero-Gonzalez, A.J.; Morales-Vicente, F.; Andersson, J.; et al. Combination of Six Individual Derivatives of the Pom-1 Antibiofilm Peptide Doubles Their Efficacy against Invasive and Multi-Resistant Clinical Isolates of the Pathogenic Yeast Candida Albicans. Pharmaceutics 2022, 14, 1332. [Google Scholar] [CrossRef]
- Kubiczek, D.; Raber, H.; Gonzalez-García, M.; Morales-Vicente, F.; Staendker, L.; Otero-Gonzalez, A.J.; Rosenau, F. Derivates of the Antifungal Peptide Cm-P5 Inhibit Development of Candida Auris Biofilms In Vitro. Antibiotics 2020, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Raber, H.F.; Sejfijaj, J.; Kissmann, A.-K.; Wittgens, A.; Gonzalez-Garcia, M.; Alba, A.; Vázquez, A.A.; Morales Vicente, F.E.; Erviti, J.P.; Kubiczek, D.; et al. Antimicrobial Peptides Pom-1 and Pom-2 from Pomacea Poeyana Are Active against Candidaauris, C. Parapsilosis and C. Albicans Biofilms. Pathogens 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Amann, V.; Kissmann, A.-K.; Mildenberger, V.; Krebs, I.; Perez-Erviti, J.A.; Martell-Huguet, E.M.; Otero-Gonzalez, A.J.; Morales-Vicente, F.; Rodríguez-Castaño, G.P.; Firacative, C.; et al. Cm-P5 Peptide Dimers Inhibit Biofilms of Candida Albicans Clinical Isolates, C. Parapsilosis and Fluconazole-Resistant Mutants of C. Auris. Int J Mol Sci 2023, 24, 9788. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Santos Fontenelle, R.O.; Brito, E.H.S.; Morais, S.M. Biofilm of Candida Albicans: Formation, Regulation and Resistance. J Appl Microbiol 2021, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Quindós, G.; Marcos-Arias, C.; San-Millán, R.; Mateo, E.; Eraso, E. The Continuous Changes in the Aetiology and Epidemiology of Invasive Candidiasis: From Familiar Candida Albicans to Multiresistant Candida Auris. International Microbiology 2018, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. In; 2017; pp. 17–65.
- Garcia-Bustos, V. Is Candida Auris the First Multidrug-Resistant Fungal Zoonosis Emerging from Climate Change? mBio 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for Combating Bacterial Biofilm Infections. Int J Oral Sci 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Taff, H.T.; Mitchell, K.F.; Edward, J.A.; Andes, D.R. Mechanisms of Candida Biofilm Drug Resistance. Future Microbiol 2013, 8, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Nadar, S.; Khan, T.; Patching, S.G.; Omri, A. Development of Antibiofilm Therapeutics Strategies to Overcome Antimicrobial Drug Resistance. Microorganisms 2022, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Kissmann, A.-K.; Mildenberger, V.; Krämer, M.; Alpízar-Pedraza, D.; Martell-Huguet, E.M.; Perez-Erviti, J.A.; Cetinkaya, A.; Pietrasik, J.; Otero-Gonzalez, A.J.; Firacative, C.; et al. Anti-Biofilm Peptides Can Rescue Fluconazole and Amphotericin B Efficacies against Candida Albicans 2024.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



