Submitted:
07 January 2025
Posted:
08 January 2025
You are already at the latest version
Abstract
Gastric cancer (GC) ranks as the fifth most prevalent malignant neoplasm globally, with an increased death rate despite recent advancements in research and therapeutic options. Different molecular subtypes of GC have distinct interactions with the immune system, impacting the tumor microenvironment (TME), prognosis, and reaction to immunotherapy. Tumor-infiltrating lymphocytes (TILs) in the TME are crucial for preventing tumor growth and metastasis, as evidenced by research showing that patients with GC who have a significant density of TILs have better survival rates. But cancer cells have evolved a variety of mechanisms to evade immune surveillance, both sialic acid-binding immunoglobulin-like lectin 15 (Siglec-15) and Programmed Death-Ligand 1 (PD-L1) playing a pivotal role in the development of an immunosuppressive TME. They prevent T cell activation and proliferation resulting in a decrease in the immune system’s capacity to recognize and eliminate malignant cells. These immune checkpoint molecules function via different but complementary mechanisms, the expression of Siglec-15 being mutually exclusive with PD-L1 and, therefore, providing a different therapeutic approach. The review explores how TILs affect tumor growth and patient outcomes in GC, with particular emphasis on their interactions within the TME and potential targeting of the PD-L1 and Siglec-15 pathways for immunotherapy.
Keywords:
1. Introduction
2. TILs
I. T lymphocytes
1. CD8+ T cells
1. CD4+ T cells
I. B lymphocytes
1. Regulatory B Cells (Bregs)
3. Tertiary Lymphoid Structures (TLSs)
I. NK Cells
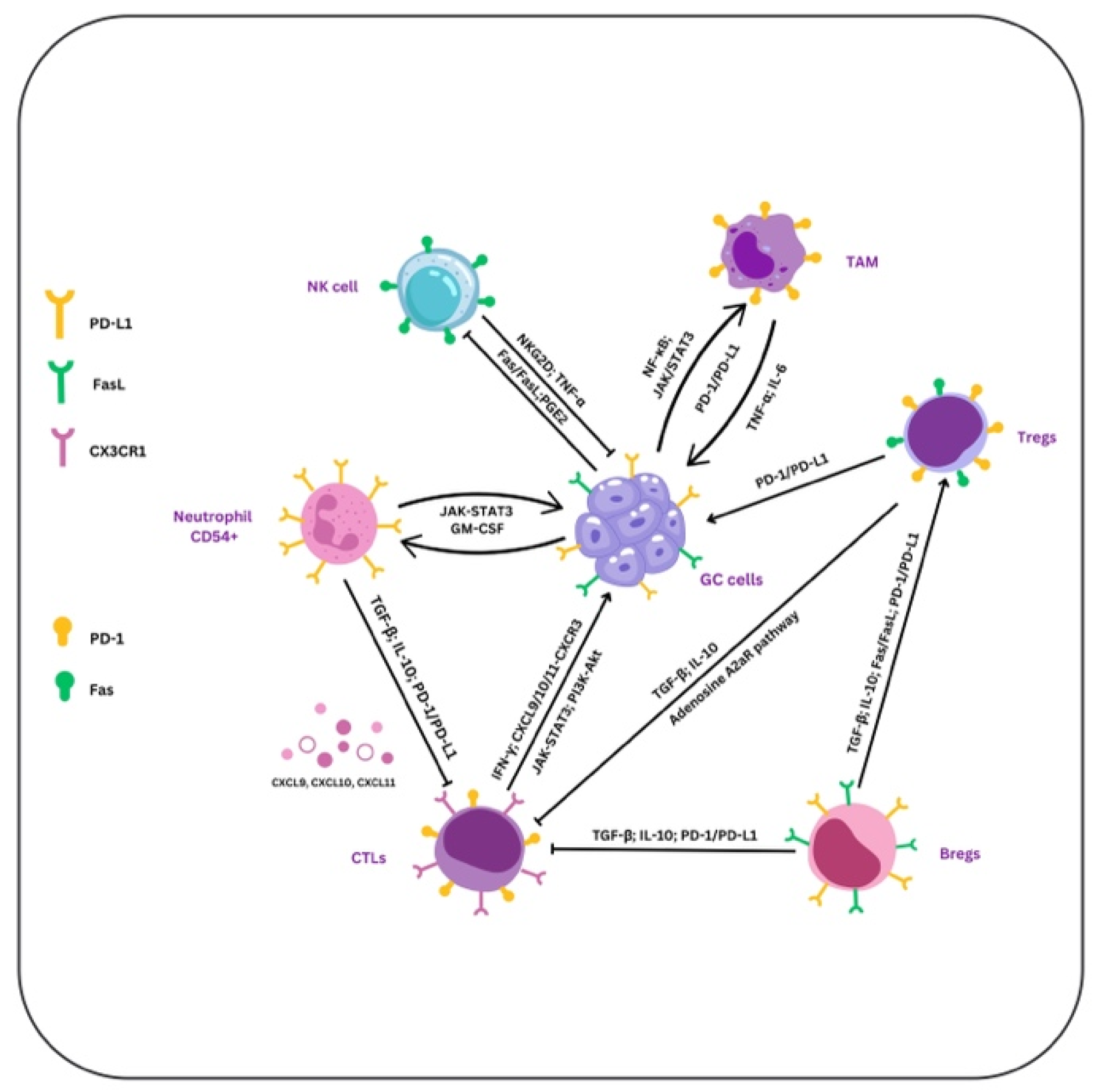
4. TAMs
5. PD-1, PD-L1 and PD-L2

| Characteristics | EBV-positive GC | MSI GC | GS GC | CIN GC |
|---|---|---|---|---|
| Molecular features |
|
|
|
|
| Immune cell infiltration |
|
|
|
|
| Immunotherapy response |
|
|
|
|
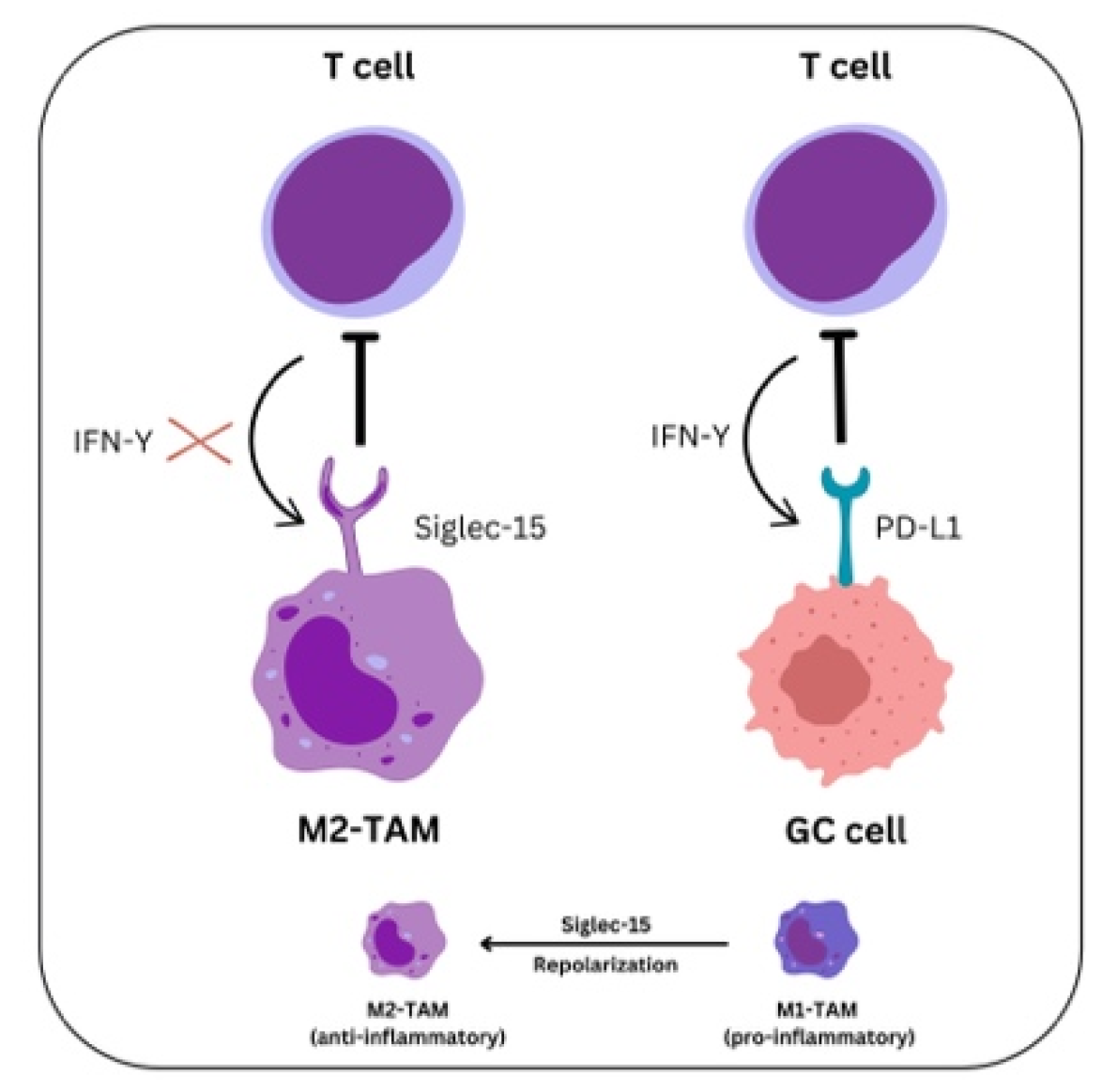
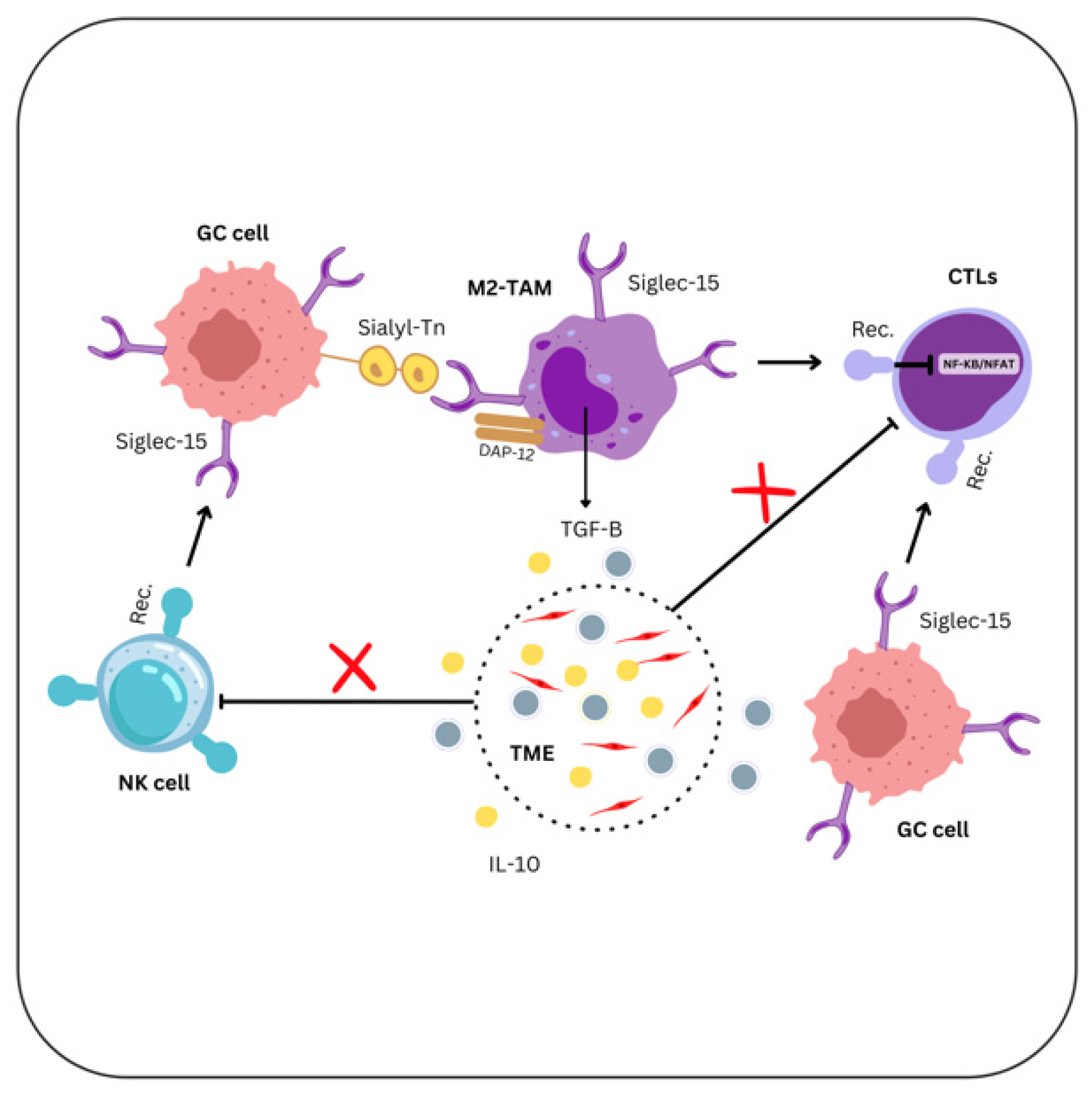
6. Siglec-15
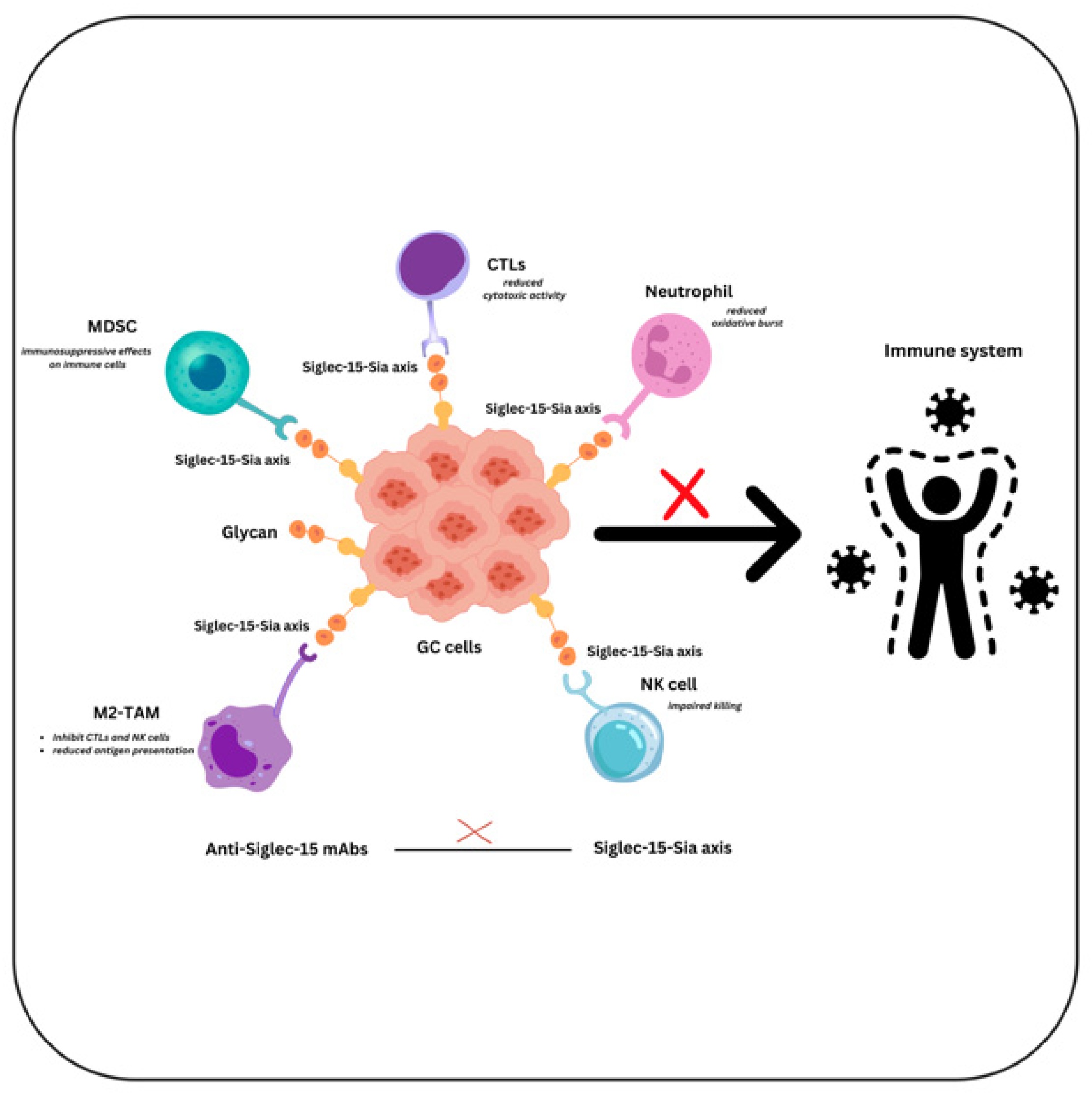
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations Used in the Main Text, Figures and Table
| A2AR | Adenosine A2A Receptor |
| ARID1A | AT-rich Interactive Domain-containing protein 1A |
| Bregs | B Regulatory Cells |
| CIN | Chromosomal Instability |
| CTLs | Cytotoxic T Lymphocytes |
| CXCL9, CXCL10, and CXCL11 | C-X-C Motif Chemokine Ligand 9, 10, and 11 |
| CXCR3 | C-X-C Motif Chemokine Receptor 3 |
| DAP12 | DNAX-Activation Protein of 12 kDa |
| DCs | Dendritic Cells |
| DNA | Deoxyribonucleic Acid |
| EBV | Epstein-Barr Virus |
| EGFR | epidermal growth factor receptor |
| Fas/FasL | Fas/Fas Ligand |
| FDC | follicular dendritic cell |
| FOXP3 | Forkhead Box Protein P3 |
| GC | Gastric Cancer |
| GS | Genomic Stable |
| H. pylori | Helicobacter pylori |
| HER-2 | Human Epidermal Growth Factor Receptor 2 |
| IFN-γ | Interferon-gamma |
| Ig | Immunoglobulin |
| IL-10 | Interleukin-10 |
| IL-17 | interleukin-17 |
| IL-21 | interleukin-21 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| JAK/STAT3 | Janus Kinase/Signal Transducer and Activator of Transcription 3 |
| mAbs | monoclonal antibodies |
| MDSCs | Myeloid-Derived Suppressor Cells |
| MHC | major histocompatibility complex |
| MMR | Mismatch Repair |
| MSI | Microsatellite Instability |
| NF-κB/NFAT | Nuclear Factor kappa-light-chain-enhancer of activated B cells/Nuclear Factor of Activated T-cells |
| NK | Natural Killer |
| NKG2D | Natural Killer Group 2 Member D |
| NSCLC | non-small cell lung cancer |
| PD-1 | Programmed Cell Death-Protein-1 receptor |
| PD-L1 | Programmed Cell Death Ligand-1 |
| PD-L2 | Programmed Cell Death Ligand-2 |
| PGE2 | Prostaglandin E2 |
| PI3K-AKT | Phosphoinositide 3-Kinase-Protein Kinase B |
| RA | Rheumatoid Arthritis |
| Sialyl-Tn | Sialylated Tn Antigen |
| Siglec-15 | Sialic Acid-Binding Immunoglobulin-Like Lectin 15 |
| TAMs | Tumor-Associated Macrophages |
| TCR | T Cell Receptor |
| TGF-β | Transforming Growth Factor-beta |
| Th | T helper |
| TILs | Tumor-Infiltrating Lymphocytes |
| TLSs | Tertiary lymphoid structures |
| TME | Tumor Microenvironment |
| TNF-α | Tumor Necrosis Factor-alpha |
| Tregs | Regulatory T Cells |
| TRM | Tissue-Resident Memory |
| VEGF | Vascular Endothelial Growth Factor |
References
- Filho, A.M.; Laversanne, M.; Ferlay, J.; Colombet, M.; Piñeros, M.; Znaor, A.; Parkin, D.M.; Soerjomataram, I.; & Bray, F. (2024). The GLOBOCAN 2022 cancer estimates: Data sources, methods, and a snapshot of the cancer burden worldwide. International journal of cancer, 10.1002/ijc.35278. Advance online publication. [CrossRef]
- Cisło, M.; Filip, A.; Anna, Arnold Offerhaus, G.; Johan, Ciseł, B.; Rawicz-Pruszyński, K.; Skierucha, M.; Polkowski, W. Piotr Distinct molecular subtypes of gastric cancer: from Laurén to molecular pathology. Oncotarget. 2018; 9: 19427-19442. Available online: https://www.oncotarget.com/article/24827/text/.
- Kim, M.; & Seo, A.N. (2022). Molecular Pathology of Gastric Cancer. Journal of gastric cancer, 22(4), 273–305. [CrossRef]
- Kim, J.Y.; Kim, W.G.; Kwon, C.H.; et al. Differences in immune contextures among different molecular subtypes of gastric cancer and their prognostic impact. Gastric Cancer 22, 1164–1175 (2019). [CrossRef]
- Wang, G.; Yang, L.; Wang, Y.; Hu, R.; Zhang, K.; Guo, T.; Chen, B.; Jiang, X.; & Cui, R. (2022). Characterization of Immune-Related Molecular Subtypes and a Prognostic Signature Correlating With the Response to Immunotherapy in Patients With Gastric Cancer. Frontiers in immunology, 13, 939836. [CrossRef]
- Ma, E.S.; Wang, Z.X.; Zhu, M.Q.; & Zhao, J. (2022). Immune evasion mechanisms and therapeutic strategies in gastric cancer. World journal of gastrointestinal oncology, 14(1), 216–229. [CrossRef]
- Wang, J.; Liu, T.; Huang, T.; Shang, M.; & Wang, X. (2022). The mechanisms on evasion of anti-tumor immune responses in gastric cancer. Frontiers in oncology, 12, 943806. [CrossRef]
- Tian, C.; Jing, H.; Wang, C.; Wang, W.; Cui, Y.; Chen, J.; & Sha, D. (2021). Prognostic role of tumour-infiltrating lymphocytes assessed by H&E-stained section in gastric cancer: a systematic review and meta-analysis. BMJ open, 11(1), e044163. [CrossRef]
- Cao, X.; Kang, Y.; Tai, P.; Zhang, P.; Lin, X.; Xu, F.; Nie, Z.; & He, B. (2024). Prognostic role of tumor-infiltrating lymphocytes in gastric cancer: A systematic review and meta-analysis. Clinics and research in hepatology and gastroenterology, 49(1), 102510. Ad-vance online publication. [CrossRef]
- Chiu, Y.M.; Tsai, C.L.; Kao, J.T.; Hsieh, C.T.; Shieh, D.C.; Lee, Y.J.; Tsay, G.J.; Cheng, K.S.; & Wu, Y.Y. (2018). PD-1 and PD-L1 Up-regulation Promotes T-cell Apoptosis in Gastric Adenocarcinoma. Anticancer research, 38(4), 2069–2078. [CrossRef]
- Sun, J.; Lu, Q.; Sanmamed, M.F.; & Wang, J. (2021). Siglec-15 as an Emerging Target for Next-generation Cancer Immunotherapy. Clinical cancer research : an official journal of the American Association for Cancer Research, 27(3), 680–688. [CrossRef]
- Huang, Z.; Guo, Y.; Li, B.; et al. Siglec-15 on macrophages suppress the immune microenvironment in patients with PD-L1 negative non-metastasis lung adenocarcinoma. Cancer Gene Ther 31, 427–438 (2024). [CrossRef]
- Ren, X. (2019). Immunosuppressive checkpoint Siglec-15: a vital new piece of the cancer immunotherapy jigsaw puzzle. Cancer biology & medicine, 16(2), 205–210. [CrossRef]
- Chen, X.; Mo, S.; Zhang, Y.; Ma, H.; Lu, Z.; Yu, S.; & Chen, J. (2022). Analysis of a novel immune checkpoint, Siglec-15, in pancreatic ductal adenocarcinoma. The journal of pathology. Clinical research, 8(3), 268–278. [CrossRef]
- Paijens, S.T.; Vledder, A.; de Bruyn, M.; et al. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell Mol Immunol 18, 842–859 (2021). [CrossRef]
- Frankowska, K.; Zarobkiewicz, M.; Dąbrowska, I.; et al. Tumor infiltrating lymphocytes and radiological picture of the tumor. Med Oncol 40, 176 (2023). [CrossRef]
- Gong, L.P.; Chen, J.N.; Xiao, L.; He, Q.; Feng, Z.Y.; Zhang, Z.G.; Liu, J.P.; Wei, H.B.; & Shao, C.K. (2019). The implication of tumor-infiltrating lymphocytes in Epstein-Barr virus-associated gastric carcinoma. Human pathology, 85, 82–91. [CrossRef]
- Ma, J.; Li, J.; Hao, Y.; Nie, Y.; Li, Z.; Qian, M.; Liang, Q.; Yu, J.; Zeng, M.; & Wu, K. (2017). Differentiated tumor immune micro-environment of Epstein–Barr virus-associated and negative gastric cancer: implication in prognosis and immunotherapy. On-cotarget, 8, 67094 - 67103.
- Knochelmann, H.M.; Dwyer, C.J.; Bailey, S.R.; et al. When worlds collide: Th17 and Treg cells in cancer and autoimmunity. Cell Mol Immunol 15, 458–469 (2018). [CrossRef]
- Wegrzyn, A.S.; Kedzierska, A.E.; & Obojski, A. (2023). Identification and classification of distinct surface markers of T regulatory cells. Frontiers in immunology, 13, 1055805. [CrossRef]
- Alcover, A.; Alarcón, B.; & Di Bartolo, V. (2018). Cell Biology of T Cell Receptor Expression and Regulation. Annual review of immunology, 36, 103–125. [CrossRef]
- Li, F.; Li, C.; Cai, X.; Xie, Z.; Zhou, L.; Cheng, B.; Zhong, R.; Xiong, S.; Li, J.; Chen, Z.; Yu, Z.; He, J.; & Liang, W. (2021). The association between CD8+ tumor-infiltrating lymphocytes and the clinical outcome of cancer immunotherapy: A systematic review and meta-analysis. EClinicalMedicine, 41, 101134. [CrossRef]
- Lieberman, J. (2010). Anatomy of a murder: how cytotoxic T cells and NK cells are activated, develop, and eliminate their targets. Immunological reviews, 235(1), 5–9. [CrossRef]
- Jameson, S.C.; & Masopust, D. (2018). Understanding Subset Diversity in T Cell Memory. Immunity, 48(2), 214–226. [CrossRef]
- Golubovskaya, V.; & Wu, L. (2016). Different Subsets of T Cells, Memory, Effector Functions, and CAR-T Immunotherapy. Cancers, 8(3), 36. [CrossRef]
- Kaech, S.M.; Wherry, E.J.; & Ahmed, R. (2002). Effector and memory T-cell differentiation: implications for vaccine development. Nature reviews. Immunology, 2(4), 251–262. [CrossRef]
- Damei, I.; Trickovic, T.; Mami-Chouaib, F.; & Corgnac, S. (2023). Tumor-resident memory T cells as a biomarker of the response to cancer immunotherapy. Frontiers in immunology, 14, 1205984. [CrossRef]
- Xu, W.; Bergsbaken, T.; & Edelblum, K.L. (2022). The multifunctional nature of CD103 (αEβ7 integrin) signaling in tissue-resident lymphocytes. American journal of physiology. Cell physiology, 323(4), C1161–C1167. [CrossRef]
- Sullivan, P.M.; Reed, S.J.; Kalia, V.; & Sarkar, S. (2021). Solid Tumor Microenvironment Can Harbor and Support Functional Properties of Memory T Cells. Frontiers in immunology, 12, 706150. [CrossRef]
- Beumer-Chuwonpad, A.; Taggenbrock, R.L.R.E.; Ngo, T.A.; & van Gisbergen, K.P.J.M. (2021). The Potential of Tissue-Resident Memory T Cells for Adoptive Immunotherapy against Cancer. Cells, 10(9), 2234. [CrossRef]
- Simoni, Y.; Becht, E.; Fehlings, M.; Loh, C.Y.; Koo, S.L.; Teng, K.W.W.; Yeong, J.P.S.; Nahar, R.; Zhang, T.; Kared, H.; Duan, K.; Ang, N.; Poidinger, M.; Lee, Y.Y.; Larbi, A.; Khng, A.J.; Tan, E.; Fu, C.; Mathew, R.; Teo, M.; … Newell, E.W. (2018). Bystander CD8+ T cells are abundant and phenotypically distinct in human tumour infiltrates. Nature, 557(7706), 575–579. [CrossRef]
- Yan, M.; Hu, J.; Ping, Y.; Xu, L.; Liao, G.; Jiang, Z.; Pang, B.; Sun, S.; Zhang, Y.; Xiao, Y.; & Li, X. (2021). Single-Cell Transcriptomic Analysis Reveals a Tumor-Reactive T Cell Signature Associated With Clinical Outcome and Immunotherapy Response In Melanoma. Frontiers in immunology, 12, 758288. [CrossRef]
- Borràs, D.M.; Verbandt, S.; Ausserhofer, M.; Sturm, G.; Lim, J.; Verge, G.A.; Vanmeerbeek, I.; Laureano, R.S.; Govaerts, J.; Sprooten, J.; Hong, Y.; Wall, R.; De Hertogh, G.; Sagaert, X.; Bislenghi, G.; D’Hoore, A.; Wolthuis, A.; Finotello, F.; Park, W.Y.; Naulaerts, S.; … Garg, A.D. (2023). Single cell dynamics of tumor specificity vs. bystander activity in CD8+ T cells define the diverse immune landscapes in colorectal cancer. Cell discovery, 9(1), 114. [CrossRef]
- Martín-Sierra, C.; Martins, R.; Laranjeira, P.; Coucelo, M.; Abrantes, A.M.; Oliveira, R.C.; Tralhão, J.G.; Botelho, M.F.; Furtado, E.; Domingues, M.R.; & Paiva, A. (2019). Functional and Phenotypic Characterization of Tumor-Infiltrating Leukocyte Subsets and Their Contribution to the Pathogenesis of Hepatocellular Carcinoma and Cholangiocarcinoma. Translational oncology, 12(11), 1468–1479. [CrossRef]
- Li, Q.; Li, Q.; Chen, J.; Liu, Y.; Zhao, X.; Tan, B.; Ai, J.; Zhang, Z.; Song, J.; & Shan, B. (2013). Prevalence of Th17 and Treg cells in gastric cancer patients and its correlation with clinical parameters. Oncology reports, 30(3), 1215–1222. [CrossRef]
- Meng, X.; Zhu, S.; Dong, Q.; Zhang, S.; Ma, J.; & Zhou, C. (2018). Expression of Th17/Treg related molecules in gastric cancer tissues. The Turkish journal of gastroenterology : the official journal of Turkish Society of Gastroenterology, 29(1), 45–51. [CrossRef]
- Kindlund, B.; Sjöling, Å.; Yakkala, C.; Adamsson, J.; Janzon, A.; Hansson, L.E.; Hermansson, M.; Janson, P.; Winqvist, O.; & Lundin, S.B. (2017). CD4+ regulatory T cells in gastric cancer mucosa are proliferating and express high levels of IL-10 but little TGF-β. Gastric cancer : official journal of the International Gastric Cancer Association and the Japanese Gastric Cancer Association, 20(1), 116–125. [CrossRef]
- Ng, T.H.; Britton, G.J.; Hill, E.V.; Verhagen, J.; Burton, B.R.; & Wraith, D.C. (2013). Regulation of adaptive immunity; the role of interleukin-10. Frontiers in immunology, 4, 129. [CrossRef]
- Wang, Y.A.; Li, X.L.; Mo, Y.Z.; Fan, C.M.; Tang, L.; Xiong, F.; Guo, C.; Xiang, B.; Zhou, M.; Ma, J.; Huang, X.; Wu, X.; Li, Y.; Li, G.Y.; Zeng, Z.Y.; & Xiong, W. (2018). Effects of tumor metabolic microenvironment on regulatory T cells. Molecular cancer, 17(1), 168. [CrossRef]
- Peng, X.; He, Y.; Huang, J.; Tao, Y.; & Liu, S. (2021). Metabolism of Dendritic Cells in Tumor Microenvironment: For Immuno-therapy. Frontiers in immunology, 12, 613492. [CrossRef]
- Wu, M.S.; Lin, J.T.; Hsu, P.N.; Lin, C.Y.; Hsieh, Y.T.; Chiu, Y.H.; Hsueh, P.R.; & Liao, K.W. (2007). Preferential induction of transforming growth factor-beta production in gastric epithelial cells and monocytes by Helicobacter pylori soluble proteins. The Journal of infectious diseases, 196(9), 1386–1393. [CrossRef]
- Kao, J.Y.; Zhang, M.; Miller, M.J.; Mills, J.C.; Wang, B.; Liu, M.; Eaton, K.A.; Zou, W.; Berndt, B.E.; Cole, T.S.; Takeuchi, T.; Owyang, S.Y.; & Luther, J. (2010). Helicobacter pylori immune escape is mediated by dendritic cell-induced Treg skewing and Th17 suppression in mice. Gastroenterology, 138(3), 1046–1054. [CrossRef]
- Larussa, T.; Leone, I.; Suraci, E.; Imeneo, M.; & Luzza, F. (2015). Helicobacter pylori and T Helper Cells: Mechanisms of Immune Escape and Tolerance. Journal of immunology research, 2015, 981328. [CrossRef]
- Rezalotfi, A.; Ahmadian, E.; Aazami, H.; Solgi, G.; & Ebrahimi, M. (2019). Gastric Cancer Stem Cells Effect on Th17/Treg Balance; A Bench to Beside Perspective. Frontiers in oncology, 9, 226. [CrossRef]
- Li, W.; Huang, X.; Han, X.; Zhang, J.; Gao, L.; & Chen, H. (2024). IL-17A in gastric carcinogenesis: good or bad?. Frontiers in immunology, 15, 1501293. [CrossRef]
- Wang, J.; Li, Q.; Cheng, X.; Zhang, B.; Lin, J.; Tang, Y.; Li, F.; Yang, C.S.; Wang, T.C.; & Tu, S. (2020). Bone Marrow-Derived Myofibroblasts Promote Gastric Cancer Metastasis by Activating TGF-β1 and IL-6/STAT3 Signalling Loop. OncoTargets and therapy, 13, 10567–10580. [CrossRef]
- Zhao, J.; Chen, X.; Herjan, T.; & Li, X. (2020). The role of interleukin-17 in tumor development and progression. The Journal of experimental medicine, 217(1), e20190297. [CrossRef]
- Maruyama, T.; Kono, K.; Mizukami, Y.; Kawaguchi, Y.; Mimura, K.; Watanabe, M.; Izawa, S.; & Fujii, H. (2010). Distribution of Th17 cells and FoxP3(+) regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer. Cancer science, 101(9), 1947–1954. [CrossRef]
- Downs-Canner, S.M.; Meier, J.; Vincent, B.G.; & Serody, J.S. (2022). B Cell Function in the Tumor Microenvironment. Annual review of immunology, 40, 169–193. [CrossRef]
- Zhang, E.; Ding, C.; Li, S.; Zhou, X.; Aikemu, B.; Fan, X.; Sun, J.; Zheng, M.; & Yang, X. (2023). Roles and mechanisms of tu-mour-infiltrating B cells in human cancer: a new force in immunotherapy. Biomarker research, 11(1), 28. [CrossRef]
- Michaud, D.; Steward, C.R.; Mirlekar, B.; & Pylayeva-Gupta, Y. (2021). Regulatory B cells in cancer. Immunological reviews, 299(1), 74–92. [CrossRef]
- Catalán, D.; Mansilla, M.A.; Ferrier, A.; Soto, L.; Oleinika, K.; Aguillón, J.C.; & Aravena, O. (2021). Immunosuppressive Mechanisms of Regulatory B Cells. Frontiers in immunology, 12, 611795. [CrossRef]
- Sarvaria, A.; Madrigal, J.A.; & Saudemont, A. (2017). B cell regulation in cancer and anti-tumor immunity. Cellular & molecular immunology, 14(8), 662–674. [CrossRef]
- Moreira, H.; Dobosz, A.; Cwynar-Zając, Ł.; Nowak, P.; Czyżewski, M.; Barg, M.; Reichert, P.; Królikowska, A.; & Barg, E. (2022). Unraveling the role of Breg cells in digestive tract cancer and infectious immunity. Frontiers in immunology, 13, 981847. [CrossRef]
- Sautès-Fridman, C.; Lawand, M.; Giraldo, N.A.; Kaplon, H.; Germain, C.; Fridman, W.H.; & Dieu-Nosjean, M.C. (2016). Tertiary Lymphoid Structures in Cancers: Prognostic Value, Regulation, and Manipulation for Therapeutic Intervention. Frontiers in immunology, 7, 407. [CrossRef]
- Zhang, Q.; & Wu, S. (2023). Tertiary lymphoid structures are critical for cancer prognosis and therapeutic response. Frontiers in immunology, 13, 1063711. [CrossRef]
- Yamakoshi, Y.; Tanaka, H.; Sakimura, C.; Deguchi, S.; Mori, T.; Tamura, T.; Toyokawa, T.; Muguruma, K.; Hirakawa, K.; & Ohira, M. (2020). Immunological potential of tertiary lymphoid structures surrounding the primary tumor in gastric cancer. Interna-tional journal of oncology, 57(1), 171–182. [CrossRef]
- Groen-van Schooten, T.S.; Franco Fernandez, R.; van Grieken, N.C.T.; Bos, E.N.; Seidel, J.; Saris, J.; Martínez-Ciarpaglini, C.; Fleitas, T.C.; Thommen, D.S.; de Gruijl, T.D.; Grootjans, J.; & Derks, S. (2024). Mapping the complexity and diversity of tertiary lymphoid structures in primary and peritoneal metastatic gastric cancer. Journal for immunotherapy of cancer, 12(7), e009243. [CrossRef]
- Peng, H.; Wu, X.; Zhang, C.; Liang, Y.; Cheng, S.; Zhang, H.; Shen, L.; & Chen, Y. (2024). Analyzing the associations between tertiary lymphoid structures and postoperative prognosis, along with immunotherapy response in gastric cancer: findings from pooled cohort studies. Journal of cancer research and clinical oncology, 150(3), 153. [CrossRef]
- Wolf, N.K.; Kissiov, D.U.; & Raulet, D.H. (2023). Roles of natural killer cells in immunity to cancer, and applications to im-munotherapy. Nature reviews. Immunology, 23(2), 90–105. [CrossRef]
- Sanseviero, E. (2019). NK Cell-Fc Receptors Advance Tumor Immunotherapy. Journal of clinical medicine, 8(10), 1667. [CrossRef]
- Du, Y.; & Wei, Y. (2019). Therapeutic Potential of Natural Killer Cells in Gastric Cancer. Frontiers in immunology, 9, 3095. [CrossRef]
- Yajima, T.; Hoshino, K.; Muranushi, R.; Mogi, A.; Onozato, R.; Yamaki, E.; Kosaka, T.; Tanaka, S.; Shirabe, K.; Yoshikai, Y.; & Kuwano, H. (2019). Fas/FasL signaling is critical for the survival of exhausted antigen-specific CD8+ T cells during tumor immune response. Molecular immunology, 107, 97–105. [CrossRef]
- Chen, Y.Y.; Feng, Y.; Mao, Q.S.; Ma, P.; Liu, J.Z.; Lu, W.; Liu, Y.F.; Chen, X.; Hu, Y.L.; & Xue, W.J. (2020). Diagnostic and prognostic value of the peripheral natural killer cell levels in gastric cancer. Experimental and therapeutic medicine, 20(4), 3816–3822. [CrossRef]
- Saito, H.; Osaki, T.; & Ikeguchi, M. (2012). Decreased NKG2D expression on NK cells correlates with impaired NK cell function in patients with gastric cancer. Gastric cancer : official journal of the International Gastric Cancer Association and the Japanese Gastric Cancer Association, 15(1), 27–33. [CrossRef]
- Li, T.; Zhang, Q.; Jiang, Y.; Yu, J.; Hu, Y.; Mou, T.; Chen, G.; & Li, G. (2015). Gastric cancer cells inhibit natural killer cell proliferation and induce apoptosis via prostaglandin E2. Oncoimmunology, 5(2), e1069936. [CrossRef]
- Rafei, H.; Daher, M.; & Rezvani, K. (2021). Chimeric antigen receptor (CAR) natural killer (NK)-cell therapy: leveraging the power of innate immunity. British journal of haematology, 193(2), 216–230. [CrossRef]
- Scheck, M.K.; Hofheinz, R.D.; & Lorenzen, S. (2024). HER2-Positive Gastric Cancer and Antibody Treatment: State of the Art and Future Developments. Cancers, 16(7), 1336. [CrossRef]
- Li, C.; Xu, X.; Wei, S.; Jiang, P.; Xue, L.; Wang, J.; Senior Correspondence. (2021). Tumor-associated macrophages: potential therapeutic strategies and future prospects in cancer. Journal for immunotherapy of cancer, 9(1), e001341. [CrossRef]
- Yerolatsite, M.; Torounidou, N.; Gogadis, A.; Kapoulitsa, F.; Ntellas, P.; Lampri, E.; Tolia, M.; Batistatou, A.; Katsanos, K.; & Mauri, D. (2023). TAMs and PD-1 Networking in Gastric Cancer: A Review of the Literature. Cancers, 16(1), 196. [CrossRef]
- Takamiya, R.; Ohtsubo, K.; Takamatsu, S.; Taniguchi, N.; & Angata, T. (2013). The interaction between Siglec-15 and tu-mor-associated sialyl-Tn antigen enhances TGF-β secretion from monocytes/macrophages through the DAP12-Syk pathway. Glycobiology, 23(2), 178–187. [CrossRef]
- Bai, R.; Li, Y.; Jian, L.; Yang, Y.; Zhao, L.; & Wei, M. (2022). The hypoxia-driven crosstalk between tumor and tumor-associated macrophages: mechanisms and clinical treatment strategies. Molecular cancer, 21(1), 177. [CrossRef]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; Han, X.; Archer, K.A.; O’Neill, T.; Herbst, R.S.; Boto, A.N.; Sanmamed, M.F.; Langermann, S.; Rimm, D.L.; & Chen, L. (2019). Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nature medicine, 25(4), 656–666. [CrossRef]
- Kang, F.B.; Chen, W.; Wang, L.; & Zhang, Y.Z. (2020). The diverse functions of Siglec-15 in bone remodeling and antitumor responses. Pharmacological research, 155, 104728. [CrossRef]
- Jiang, K.Y.; Qi, L.L.; Kang, F.B.; & Wang, L. (2022). The intriguing roles of Siglec family members in the tumor microenvi-ronment. Biomarker research, 10(1), 22. [CrossRef]
- Zhou, Z.; Yang, Z.; Wang, J.; Wang, Q.; Zhang, H.; & Ju, X. (2021). Research progress on tumour associated macrophages in gastric cancer (Review). Oncology Reports, 45, 35. [CrossRef]
- Parvez, A.; Choudhary, F.; Mudgal, P.; Khan, R.; Qureshi, K.A.; Farooqi, H.; & Aspatwar, A. (2023). PD-1 and PD-L1: architects of immune symphony and immunotherapy breakthroughs in cancer treatment. Frontiers in immunology, 14, 1296341. [CrossRef]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; & Sharpe, A.H. (2008). PD-1 and its ligands in tolerance and immunity. Annual review of immunology, 26, 677–704. [CrossRef]
- Cui, J.W.; Li, Y.; Yang, Y.; Yang, H.K.; Dong, J.M.; Xiao, Z.H.; He, X.; Guo, J.H.; Wang, R.Q.; Dai, B.; & Zhou, Z.L. (2024). Tumor immunotherapy resistance: Revealing the mechanism of PD-1/PD-L1-mediated tumor immune escape. Biomedicine & pharma-cotherapy = Biomedecine & pharmacotherapie, 171, 116203. [CrossRef]
- Chen, J.; Jiang, C.C.; Jin, L.; & Zhang, X.D. (2016). Regulation of PD-L1: a novel role of pro-survival signalling in cancer. Annals of oncology: official journal of the European Society for Medical Oncology, 27(3), 409–416. [CrossRef]
- Song, T.L.; Nairismägi, M.L.; Laurensia, Y.; Lim, J.Q.; Tan, J.; Li, Z.M.; Pang, W.L.; Kizhakeyil, A.; Wijaya, G.C.; Huang, D.C.; Nagarajan, S.; Chia, B.K.; Cheah, D.; Liu, Y.H.; Zhang, F.; Rao, H.L.; Tang, T.; Wong, E.K.; Bei, J.X.; Iqbal, J.; … Ong, C.K. (2018). Oncogenic activation of the STAT3 pathway drives PD-L1 expression in natural killer/T-cell lymphoma. Blood, 132(11), 1146–1158. [CrossRef]
- Mandai, M.; Hamanishi, J.; Abiko, K.; Matsumura, N.; Baba, T.; & Konishi, I. (2016). Dual Faces of IFNγ in Cancer Progression: A Role of PD-L1 Induction in the Determination of Pro- and Antitumor Immunity. Clinical cancer research : an official journal of the American Association for Cancer Research, 22(10), 2329–2334. [CrossRef]
- Mimura, K.; Teh, J.L.; Okayama, H.; Shiraishi, K.; Kua, L.F.; Koh, V.; Smoot, D.T.; Ashktorab, H.; Oike, T.; Suzuki, Y.; Fazreen, Z.; Asuncion, B.R.; Shabbir, A.; Yong, W.P.; So, J.; Soong, R.; & Kono, K. (2018). PD-L1 expression is mainly regulated by in-terferon gamma associated with JAK-STAT pathway in gastric cancer. Cancer science, 109(1), 43–53. [CrossRef]
- Ju, X.; Zhang, H.; Zhou, Z.; Chen, M.; & Wang, Q. (2020). Tumor-associated macrophages induce PD-L1 expression in gastric cancer cells through IL-6 and TNF-ɑ signaling. Experimental cell research, 396(2), 112315. [CrossRef]
- Tokunaga, R.; Zhang, W.; Naseem, M.; Puccini, A.; Berger, M.D.; Soni, S.; McSkane, M.; Baba, H.; & Lenz, H.J. (2018). CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation - A target for novel cancer therapy. Cancer treatment reviews, 63, 40–47.
- Zhang, C.; Li, Z.; Xu, L.; Che, X.; Wen, T.; Fan, Y.; Li, C.; Wang, S.; Cheng, Y.; Wang, X.; Qu, X.; & Liu, Y. (2018). CXCL9/10/11, a regulator of PD-L1 expression in gastric cancer. BMC cancer, 18(1), 462. [CrossRef]
- Wang, T.T.; Zhao, Y.L.; Peng, L.S.; Chen, N.; Chen, W.; Lv, Y.P.; Mao, F.Y.; Zhang, J.Y.; Cheng, P.; Teng, Y.S.; Fu, X.L.; Yu, P.W.; Guo, G.; Luo, P.; Zhuang, Y.; & Zou, Q.M. (2017). Tumour-activated neutrophils in gastric cancer foster immune suppression and disease progression through GM-CSF-PD-L1 pathway. Gut, 66(11), 1900–1911. [CrossRef]
- Shan, Z.G.; Yan, Z.B.; Peng, L.S.; Cheng, P.; Teng, Y.S.; Mao, F.Y.; Fan, K.; Zhuang, Y.; & Zhao, Y.L. (2021). Granulo-cyte-Macrophage Colony-Stimulating Factor-Activated Neutrophils Express B7-H4 That Correlates with Gastric Cancer Pro-gression and Poor Patient Survival. Journal of immunology research, 2021, 6613247. [CrossRef]
- Li, J.J.; & Lee, C.S. (2023). The Role of the AT-Rich Interaction Domain 1A Gene (ARID1A) in Human Carcinogenesis. Genes, 15(1), 5. [CrossRef]
- Kim, Y.B.; Ahn, J.M.; Bae, W.J.; Sung, C.O.; & Lee, D. (2019). Functional loss of ARID1A is tightly associated with high PD-L1 expression in gastric cancer. International journal of cancer, 145(4), 916–926. [CrossRef]
- Nakano, H.; Saito, M.; Nakajima, S.; Saito, K.; Nakayama, Y.; Kase, K.; Yamada, L.; Kanke, Y.; Hanayama, H.; Onozawa, H.; Okayama, H.; Fujita, S.; Sakamoto, W.; Saze, Z.; Momma, T.; Mimura, K.; Ohki, S.; Goto, A.; & Kono, K. (2021). PD-L1 over-expression in EBV-positive gastric cancer is caused by unique genomic or epigenomic mechanisms. Scientific reports, 11(1), 1982. [CrossRef]
- Saito, R.; Abe, H.; Kunita, A.; Yamashita, H.; Seto, Y.; & Fukayama, M. (2017). Overexpression and gene amplification of PD-L1 in cancer cells and PD-L1+ immune cells in Epstein-Barr virus-associated gastric cancer: the prognostic implications. Modern pathology : an official journal of the United States and Canadian Academy of Pathology, Inc, 30(3), 427–439. [CrossRef]
- Zhang, M.; Dong, Y.; Liu, H.; Wang, Y.; Zhao, S.; Xuan, Q.; Wang, Y.; & Zhang, Q. (2016). The clinicopathological and prognostic significance of PD-L1 expression in gastric cancer: a meta-analysis of 10 studies with 1,901 patients. Scientific reports, 6, 37933. [CrossRef]
- Sun, H.; Nered, S.; Tryakin, A.; Abgaryan, M.; Artamonova, E.; Stroganova, A.; Stilidi, I. (2024). Microsatellite instability (MSI) in patients with gastric cancer (GC) and correlation with PD-L1 expression. JCO 42, 389-389(2024). [CrossRef]
- Kim, J.Y.; Kim, W.G.; Kwon, C.H.; & Park, D.Y. (2019). Differences in immune contextures among different molecular subtypes of gastric cancer and their prognostic impact. Gastric cancer : official journal of the International Gastric Cancer Association and the Japanese Gastric Cancer Association, 22(6), 1164–1175. [CrossRef]
- Jiang, K.Y.; Qi, L.L.; Liu, X.B.; Wang, Y.; & Wang, L. (2023). Prognostic value of Siglec-15 expression in patients with solid tumors: A meta-analysis. Frontiers in oncology, 12, 1073932. [CrossRef]
- Cui, L.; Tian, X.; Yan, L.; Guan, X.; Dong, B.; Zhao, M.; Lv, A.; Liu, D.; Wu, J.; & Hao, C. (2022). Expression and function of Siglec-15 in RLPS and its correlation with PD-L1: Bioinformatics Analysis and Clinicopathological Evidence. International journal of medical sciences, 19(13), 1977–1988. [CrossRef]
- Huang, R.; Zheng, J.; Shao, Y.; Zhu, L.; & Yang, T. (2023). Siglec-15 as multifunctional molecule involved in osteoclast differen-tiation, cancer immunity and microbial infection. Progress in biophysics and molecular biology, 177, 34–41. [CrossRef]
- Li, B.; Zhang, B.; Wang, X.; Zeng, Z.; Huang, Z.; Zhang, L.; Wei, F.; Ren, X.; & Yang, L. (2020). Expression signature, prognosis value, and immune characteristics of Siglec-15 identified by pan-cancer analysis. Oncoimmunology, 9(1), 1807291. [CrossRef]
- Shafi, S.; Aung, T.N.; Robbins, C.; Zugazagoitia, J.; Vathiotis, I.; Gavrielatou, N.; Yaghoobi, V.; Fernandez, A.; Niu, S.; Liu, L.N.; Cusumano, Z.T.; Leelatian, N.; Cole, K.; Wang, H.; Homer, R.; Herbst, R.S.; Langermann, S.; & Rimm, D.L. (2022). Development of an immunohistochemical assay for Siglec-15. Laboratory investigation; a journal of technical methods and pathology, 102(7), 771–778. [CrossRef]
- Hao, J.Q.; Nong, J.Y.; Zhao, D.; Li, H.Y.; Su, D.; Zhou, L.J.; Dong, Y.J.; Zhang, C.; Che, N.Y.; Zhang, S.C.; Lin, J.Z.; Yang, J.B.; Zhang, H.T.; & Wang, J.H. (2020). The significance of Siglec-15 expression in resectable non-small cell lung cancer. Neoplasma, 67(6), 1214–1222. [CrossRef]
- ClinicalTrials.gov. (2025). The Study of NC318 Alone or in Combination With Pembrolizumab in Patients With Advanced Non-small Cell Lung Cancer. ClinicalTrials.gov Identifier: NCT04699123. Available online: https://clinicaltrials.gov/study/NCT04699123?cond=NC318&rank=2 (accessed on 4 January 2025).
- Tolcher, A.; Hamid, O.; Weber, J.; LoRusso, P.; Shantz, K.; Heller, K.; & Gutierrez, M. (2019, November). Single agent anti-tumor activity in PD-1 refractory NSCLC: phase 1 data from the first-in-human trial of NC318, a Siglec-15-targeted antibody. In JOURNAL FOR IMMUNOTHERAPY OF CANCER (Vol. 7). CAMPUS, 4 CRINAN ST, LONDON N1 9XW, ENGLAND: BMC.
- ClinicalTrials.gov. (2025). Study of PYX-106 in Solid Tumors.
- ClinicalTrials.gov Identifier: NCT05718557. Available online: https://clinicaltrials.gov/study/NCT05718557 (accessed on 4 January 2025).
- Quirino, M.W.L.; Pereira, M.C.; Deodato de Souza, M.F.; Pitta, I.D.R.; Da Silva Filho, A.F.; Albuquerque, M.S.S.; Albuquerque, A.P.B.; Martins, M.R.; Pitta, M.G.D.R.; & Rêgo, M.J.B.M. (2021). Immunopositivity for Siglec-15 in gastric cancer and its association with clinical and pathological parameters. European journal of histochemistry : EJH, 65(1), 3174. [CrossRef]
- Moreira, R.S.; da Silva, M.M.; de Melo Vasconcelos, C.F.; da Silva, T.D.; Cordeiro, G.G.; Mattos-Jr, L.A.R.; da Rocha Pitta, M.G.; de Melo Rêgo, M.J.B.; & Pereira, M.C. (2023). Siglec 15 as a biomarker or a druggable molecule for non-small cell lung cancer. Journal of cancer research and clinical oncology, 149(19), 17651–17661. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



