
Submitted:
06 January 2025
Posted:
07 January 2025
You are already at the latest version
Abstract
Mini-puberty refers to the transient activation of the hypothalamic-pituitary-gonadal (HPG) axis during early infancy, lasting up to six months in boys and 12-24 months in girls. This phase represents the second activation of the HPG axis, following its initial activation during the second half of fetal life. At birth, the removal of the suppressive effect of placental estrogens on the HPG axis prompts a rise in both gonadotropins and sex steroid hormones, resulting in distinct clinical and laboratory markers of mini-puberty. While the clinical significance of mini-puberty remains partially understood, emerging evidence underscores its essential role in several aspects of human growth and development. In boys, testosterone influences penile growth, increases Sertoli cell numbers in the testes, and lays the foundation for future spermatogenesis. In girls, the increase in estradiol levels promote follicular maturation and stimulate breast and uterine growth. Beyond the gonadal effects, mini-puberty appears to impact body composition, affecting body weight and promoting accelerated growth. Additionally, it may affect early psychosomatic and neural maturation, playing a role in several key aspects of the infantile brain. This narrative review examines recent findings on the physiology of the activation of HPG axis before and after birth along with its significance in various aspects of human growth and development. In addition, mini-puberty unique features in specific groups, such as preterm and small-for-gestational-age infants are presented.
Keywords:
mini-puberty
; hypothalamic-pituitary-gonadal axis
; infantcy
; preterms
; small-for-gestational-age infants
1. Introduction
Throughout the human lifespan, the hypothalamic-pituitary-gonadal (HPG) axis is activated on three distinct occasions: first during fetal development, again in the early postnatal months, during the so-called mini-puberty, and finally at the onset of puberty. The significance of fetal HPG axis activation is partially understood but seems to play a role in genital development especially in males, as evidenced by the genital underdevelopment seen at birth in patients with congenital hypogonadism [1,2].
The second activation of the HPG axis, occurring in the first few months after birth, has attracted increasing attention in recent years as a focus of scientific research [3]. In males, mini-puberty involves a surge in luteinizing hormone (LH) and follicle-stimulating hormone (FSH), leading to a peak in testosterone levels between 1 and 3 months of age. This testosterone surge contributes to testicular growth, penile development, and the establishment of the Sertoli and Leydig cell populations, which are vital for future spermatogenesis and fertility. In females, mini-puberty is marked by fluctuating levels mainly of estradiol, which stimulate transient ovarian activity and the formation of antral follicles. However, the exact role of these hormonal changes in female infants remains unclear [4].
Emerging research suggests that the postnatal HPG axis activation may have broader implications beyond reproductive development. For example, it has been implicated as an important factor affecting linear growth and body composition [5]. In both sexes, the hormonal surges during mini-puberty influence neurodevelopment by shaping sex-specific brain structures and neural circuits that affect cognitive and behavioral outcomes [6]. Additionally, the mini-puberty phase may provide a unique window for diagnosing and addressing disorders of sexual development or hormonal dysfunction, while small-for-gestational age (SGA) and premature infants show some unique hormonal patterns during their mini-puberty [4].
This review examines the activation of the HPG axis during fetal life, delves into its postnatal activation, and summarizes the current knowledge of hormonal changes in the pituitary and gonads. It also highlights the physiological effects of these hormones on target tissues, emphasizing their importance in early development and potential clinical applications and delineates the unique features of mini-puberty in SGA and premature infants.
2. Hypothalamic-Pituitary-Gonadal Axis Activation During Fetal Life
As early as 40 days after conception, the 1000-1500 neurons that will be responsible for Gonadotropin Releasing Hormone (GnRH) production, appear in the olfactory placode of the developing nose and gradually migrate, along the olfactory nerve, to the hypothalamus. GnRH production in the fetal hypothalamus is detected at around 15 weeks of gestation [7,8]. By the end of the first trimester, GnRH neurons have established a complex interconnection both with the gonadotropin producing neurons of the pituitary, and with a wide range of local neuronal networks. This enables them to receive various central and peripheral inputs [9]. In addition, recent evidence suggests that GnRH pulsatile secretion as well as other important functions of GnRH neurons are under the control of the so-called Kisspeptin-Neurokinin B-Dynorphin (KNDy) neuronal network [10,11]. The exact time at which the interconnection between GnRH neurons and the KNDy system is established, remains to be elucidated [12]. It is noteworthy that GnRH neurons themselves lack estrogen receptors alpha (ER-alpha). Τhe placenta-produced estrogen suppression of GnRH and gonadotropin production that occurs near birth has been shown to be mediated through the KNDy system [13].
At the pituitary level, the gonadotropins LH, and FSH can already be detected at the fetal developing pituitary at 9 weeks [14] and in the fetal circulation by 12-14 weeks of gestation [15]. At some point during the intrauterine life, gonadotropin secretion becomes fully regulated by the hypothalamic GnRH, but when exactly this happens, it is unknown. The portal vascular system is present by the end of the first trimester but seems to fully mature much later. A study on anencephalic fetuses that have a functional pituitary but no hypothalamus, showed that gonadotropic cells are present up to 18 weeks of gestation, and then gradually involute to disappear by the 32nd week [16]. Another study showed that LH and FSH production is independent of GnRH and kisspeptin levels until midgestation and become GnRH-induced after the 30th-31st week [17]. These observations suggest that GnRH is essential for gonadotropic cell function from almost midgestation onwards. Gonadotropin levels in fetal circulation peak around the 30th week of gestation and gradually decrease until birth, most probably due to the increasing levels of estrogen produced by the placenta which suppresses the fetal HPG axis [18].
Gonadotropin levels differ between the two sexes. During the first half of intrauterine life, female fetuses have higher levels of LH and FSH, both if measured as pituitary content or as serum levels. This difference is mainly attributed to the higher levels of testicular hormones in the male fetal circulation exerting negative feedback on the hypothalamus and pituitary [19]. Another difference is the higher concentration of LH compared to FSH in male fetuses, while the opposite is true for female fetuses [20]. Human chorionic gonadotropin (hCG) which is secreted by the placenta, is also involved in the fetal HPG axis regulation. hCG is structurally and functionally similar to LH, with which it shares a common cellular receptor [21]. hCG levels in the fetus rise during the first trimester, peaking at 8-12 weeks of gestation, and then decrease at a slow rate, remaining quite high until the end of gestation [19].
At the gonads level, gonadal sex of the fetus is undifferentiated up to the 6th week of gestation. In 46,XY fetuses, SRY gene on the Y chromosome interacts with a complex network of other genes to activate the testis pathway [22,23]. In this case, placental hCG acts on the testicular mesenchymal cells transforming them into Leydig cells, which secrete testosterone and anti-Mullerian hormone (AMH) as early as the 8th week of gestation. Testosterone is transformed to its more potent metabolite dihydrotestosterone via the action of the enzyme 5-α reductase, leading to the formation of prostate, penis, and scrotum. AMH on the other hand, is important for the regression of mullerian ducts, thus averting uterus and fallopian tubes formation. Between the 10th and 20th week of gestation, testosterone levels peak in the male fetal circulation, reaching almost adult values (40–580 ng/dL) [14]. It should be noted that testosterone production in the fetal testes is regulated initially by the placental hCG and only after the 9th week of gestation does it come under the control of the fetal pituitary LH production. After the 20th week of gestation, testosterone gradually decreases toward term [24]. Regarding pituitary FSH, it seems that it plays a role in Sertoli cell proliferation, since fetal testes have been shown to express FSH receptors, but relevant literature is scarce [24,25].
In 46,XX fetuses, the absence of the SRY gene, together with the activation of some recently discovered genes, direct toward female gonadal sex [22,23]. The absence of AMH leads to the persistence of mullerian ducts and the formation of the uterus, the fallopian tubes, and the upper one third of the vagina. Primordial follicles in the fetal ovaries first appear around the 14th week of gestation [26]. Around the 20th week of gestation, the first primary follicles appear which then gradually progress to the antral stage during the third trimester [26,27]. The number of follicles starts to increase around the 14th-15th up until the 34th week of gestation and then it remains stable [28]. Concerning folliculogenesis during fetal life, the role of fetal estrogens and gonadotropins is not completely understood. Fetal estrogen production seems to play little or no role in follicle maturation, since the largest amount of estrogen during fetal life comes from the placenta. In addition, studies in anencephalic female fetuses have shown that fetal gonadotropins are important in the development of growing antral follicles only after the 34th week of gestation and not before [29]. More studies are needed to identify the importance of fetal LH, FSH, and estrogen production in developing fetal ovaries.
3. Mini Puberty in Males
Mini puberty refers to the second activation of the hypothalamic-pituitary-gonadal (HPG) axis that in males, occurs during the first six months of life. It is characterized by increased secretion of gonadotropins leading to increased sex hormone production [30]. Mini-puberty was initially described in boys by Forest et al. in 1973 [31]. It begins immediately after birth with the cessation of placental estrogens’ inhibitory effect on fetal pituitary secretion of FSH and LH (Figure 1). This triggers a rapid rise in testosterone within minutes, peaking at 2 to 3 hours after delivery, followed by a decrease within the next 6 to 12 hours [32]. Subsequently, all hormones in the HPG axis decline in both sexes for approximately one week and then they start increasing again [33,34]. More specifically, LH levels rise during the second week of life, peaking between weeks 2 and 10, leading to increased testosterone production by Leydig cells [35]. Testosterone levels reach concentrations similar to those in adults at 2 to 3 months, then decrease, returning to low levels by 6 months of age [36]. Notably, testosterone levels during mini-puberty exhibit significant variability [34] but, in any case, seem to play an important role in the continued development of the male external genitalia, particularly of the penis [37]. Furthermore, it has been shown that androgen synthesis during this phase is mainly the result of the backdoor pathway, as metabolites of the Δ4 rather than the Δ5 lyase pathway of steroidogenesis are mainly found in the urine of male infants [38]. As a response to LH surge, Leydig cells produce Insulin-like factor 3 (INSL3) with concentrations peaking around one month of age. It has been shown that this peptide is important for testicular descent and for the final placement of the testicles in the scrotum [39].
On the other hand, FSH starts rising after the first week of life, peaks at 10 to 15 days, and gradually declines after three months of life variability [34]. This FSH surge acts upon immature Sertoli cells and germ cells driving their mitotic proliferation and the development of seminiferous tubules [40,41]. In addition to increasing in number, Sertoli cells support the maturation of gonocytes into spermatogonial stem cells. This increase in cellular proliferation leads to a transient increase of testicular volume, peaking around 4 to 6 months of age, and reaching up to 40% increase compared to birth size [34]. This whole activation of the testes seems to be important for future reproductive capacity of the individuals since Sertoli and germ cells play a crucial role in postpubertal spermatogenesis [34]. Nevertheless, androgen receptors in Sertoli cells start being expressed after one year of age with a gradual increase up to 4 to 5 years of age [42]. This precludes meiotic activity and therefore spermatogenesis in Sertoli cells during mini-puberty, despite the intratesticular testosterone rise [43]. Sertoli cell stimulation by FSH leads to the production of increasing amounts of AMH and inhibin B, with serum concentrations of these peptides peaking around four months and remaining elevated during the early childhood years [33,44].
4. Mini Puberty in Females
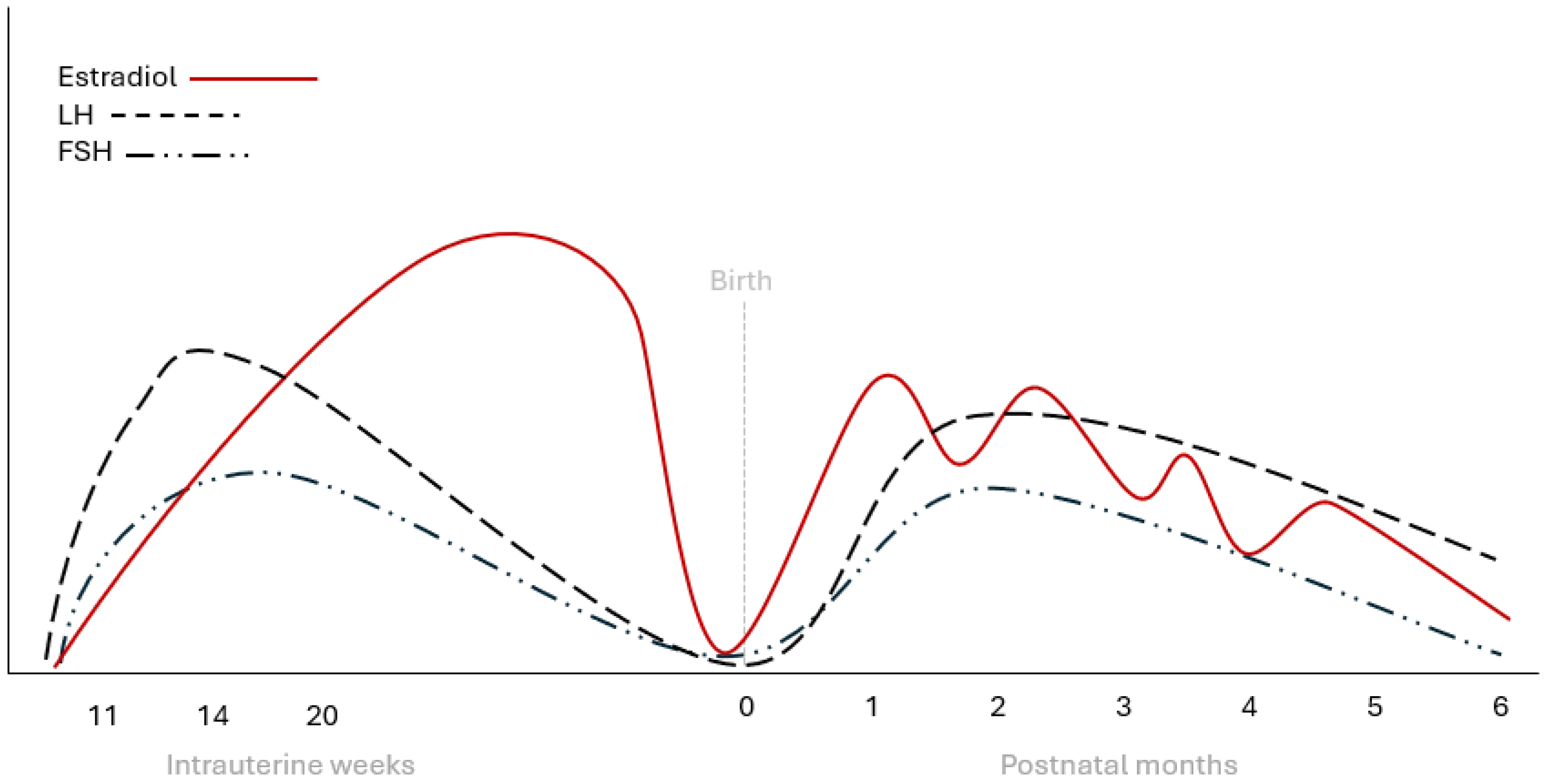
Similarly to males, in females, estradiol levels are elevated in the cord blood at birth but drop rapidly during the first days of life [45]. After the first week of life, estradiol levels begin to rise in girls and remain higher than in boys over the following months (46). This increase is the result of ovarian stimulation by pituitary gonadotropins and coincides with ovarian follicular development [46,47]. Indeed, it has been shown that estradiol intraovarian concentration is higher during the first six months of life [48], and large ovarian follicles are more common during the first 6 to 12 months of life [49]. Parallel to estrogen increase, peptides produced by the granulosa cells, such as AMH and inhibin B, also increase during the mini puberty gonadotropin surge [36,47]. In contrast to testosterone levels in boys which peak between 1 and 3 months and gradually decline by six months, estradiol levels in girls fluctuate, likely due to the cyclic maturation and regression of ovarian follicles [47] (Figure 2). After the first year, estradiol levels start to decline but they never reach the nadir that is observed in boys during childhood, when measured with ultrasensitive methods [50].
Regarding estrogen-sensitive tissues, placental estrogens stimulate breast tissue leading to its presence in most full-term infants [51]. At birth, mammary gland size does not differ between the two sexes but during infancy, girls display larger mammary glands than boys, probably due to endogenous estrogen stimulation of the breast tissue [51,52]. Regarding uterus, its growth is stimulated prenatally, with rapid size reduction occurring in the months following birth due to the withdrawal of high placental estrogen effect [47,53]. As explained in more detail in the following chapter, premature girls show reduced size of mammary glands and uterus at birth compared to full-term girls because they miss the peak late intrauterine estrogen exposure [47]. After birth, endogenous estrogen production leads to a more pronounced effect on breast and uterine growth in such girls [47].
The significance of increased reproductive hormones during mini-puberty is well established for male genital development and future fertility. In females, the primordial follicular pool formation occurs before birth and is not affected throughout much of mini-puberty [28]. Estrogen-sensitive organs such as the mammary glands and the uterus increase in size during mini-puberty in female infants as a response to the activation of the HPG axis and estrogen production by the developing ovarian follicles [47,54]. Further, studies on female mouse models suggest that during mini-puberty, gonadotropins are critical for early follicular development and may play additional roles, such as influencing sexual and maternal behaviors, and establishing coordinated GnRH pulsatility [54]. Despite these data, mini-puberty impact on female gonadal function and fertility remains incompletely understood.
5. Mini Puberty in Premature and Small-for-Gestational Age Infants
Mini puberty is slightly different in preterm infants compared to the one in full-term infants. Several studies have shown that premature boys and girls born between 28 and 42 weeks of gestation experience a higher and more prolonged postnatal LH and FSH increase [55,56]. More specifically, in preterm boys LH, FSH, and testosterone levels reach significantly higher concentrations than those observed in full-term boys at 1 to 3 months of age [55,57]. This exaggerated mini-puberty results in faster testicular and penile growth in preterm boys compared to their full-term counterparts. Regarding preterm girls, they also display an accentuated HPG axis activity, with FSH levels rising more sharply, peaking at approximately 32 weeks of corrected age and persisting for a longer duration compared to full-term girls [46,55]. Early FSH increase leads to a transient ovarian stimulation, with several follicles progressing to the antral phase and producing increased amounts of AMH [46]. Similar patterns of exaggerated mini-puberty have also been identified in infants born SGA, but relevant literature is scarce [58,59]. Overall, data on the long-term complications of being born SGA or premature on gonadal function and fertility are scarce but they mostly show no association [60,61,62].
When the infant is extremely premature, the normal prenatal and postnatal regulation of the HPG axis can be significantly affected and the normal negative feedback loops are very immature. Several cases of extremely premature girls born between 25 and 30 weeks of gestation have been described with the so-called preterm ovarian hyperstimulation syndrome. First described in 1985 by Sedin et al. [63], this condition is characterized by breast enlargement, swelling of the external genitalia (clitoris, and both labia majora and minora), and vaginal bleeding along with increased levels of gonadotropins and estradiol and the presence of ovarian cysts [64,65,66]. Interestingly, this syndrome usually self-resolves while data on its long-term effects on reproductive function are scarce and inconsistent [67].
6. Mini-Puberty, Linear Growth and Body Composition
Linear growth has been shown to differ between boys and girls during the first year of life. Indeed, male infants grow in length more rapidly than girls, especially during the first 6 months of life, leading to an average difference of 2,6 cm per year. A study by Kiviranta et al [5], compared the linear growth of 18,570 healthy boys and girls during their first year of life. In 84 of them, growth velocity was compared with repeated measurements of urinary testosterone, estradiol, and serum IGF-I during the first 6 months. Growth velocity showed a strong positive correlation with testosterone levels in both sexes. Testosterone was higher in boys compared to girls between 7 days and 6 months of age, coinciding with the higher growth velocity in boys observed from birth to 6 months. Further, the highest difference in growth velocity between boys and girls was observed during the first month of life (corresponding to 4.1 cm per year), coinciding with the postnatal testosterone surge in boys. Interestingly, no significant differences in IGF-I levels between the two sexes and no association between IGF-I levels and growth velocity were identified.
Regarding body composition, females typically exhibit a lower proportion of lean mass (muscle) and a higher proportion of fat mass compared to males, with gonadal steroids believed to play a central role in this difference. It is not totally clear if this sexual dimorphism in body composition is already present at birth, since relevant data are conflicting. Some studies suggest that such differences between sexes are already observable at birth with variations in HPG axis activation during mid-gestation being responsible [68,69]. On the contrary, other studies have identified no such differences [70,71]. During the first months after birth, it is possible that testosterone level variations play a role in further shaping sex-related body composition differences. For example, in a recent study by Davis et al [72], total mass, fat-free mass, and percent fat mass were measured by air displacement plethysmography in a large, diverse group of healthy term infants from various ethnic backgrounds. Measurements were performed once between the first and the third days of life and again at five months, and differences between the two sexes were recorded at both time points. Males were found to have greater total mass and fat free mass, but lower percent fat mass compared to females. Between the two time points, fat free mass increased more in males, with a resultant average difference of 410 grams at 5 months of age. On the opposite, females showed a relatively higher increase in percent fat mass.
7. Mini-Puberty and Neurodevelopment
Mini-puberty has been shown to influence the organization and function of the brain in a sex-specific manner. First, testosterone surge in male infants is critical for the masculinization of the brain. More specifically, it shapes the hypothalamic structures involved in regulating reproductive behaviors, including the establishment of GnRH pulsatility [3,73]. In addition, it contributes to differences in gray and white matter distribution, and cortical thickness which are later reflected in specifically male or female cognitive and behavioral patterns [6,74]. Further, sex steroid hormones during mini-puberty influence cognitive and behavioral outcomes by shaping neural circuits involved in learning, memory, and social behaviors. For example, testosterone levels have been linked to social cognitive abilities, such as face processing and empathy, particularly in boys [75]. In girls, estradiol fluctuations during mini-puberty may influence the early development of emotional regulation and social bonding circuits [76]. Another important role of sex hormones during mini-puberty is the facilitation of neural networks organization in a period coinciding of heightened brain plasticity. Indeed, it has been shown that testosterone and estradiol enhance synaptogenesis, dendritic growth, and myelination during mini-puberty, processes critical for efficient neural communication [54,73].
8. Conclusions
It is well established that during mid-gestation, the HPG axis of the fetus becomes increasingly active but is gradually suppressed as birth approaches due to the inhibitory effects of placental hormones on the fetal hypothalamus. At birth, with the removal of this suppression, the HPG axis is reactivated. This reactivation leads to a testosterone surge in males, peaking between 1 and 3 months of age, while in females, estradiol levels show fluctuations until around 6 months, with gradual decrease afterwards. Mini-puberty plays a vital role in the maturation of the genital organs and seems to create the conditions needed for future fertility. However, its precise significance, particularly in females, remains unclear and requires further investigation. Similarly, the mechanisms through which the HPG axis becomes quiescent after mini-puberty and remains inactive until puberty, are not yet fully understood. Finally, mini-puberty offers a critical window to explore potential clinical implications, particularly for specific patient groups such as preterm or small-for-gestational-age infants. Further studies are needed to clarify the clinical significance and the potential implications of mini-puberty alterations in such specific populations.
Author Contributions
Conceptualization, A.S. and V.G.; methodology, A.S. and C.K; investigation, A.L., N.A. and LE.G; resources, A.L., N.A. and LE.G; writing—original draft preparation, A.S. and C.K.; writing—review and editing, V.G.; supervision, A.S. and V.G.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Not available
Acknowledgments
None
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AMH | anti-Mullerian hormone |
| HPG | hypothalamic-pituitary-gonadal |
| FSH | follicle-stimulating hormone |
| LH | luteinizing hormone |
| SGA | small-for-gestational age |
| GnRH | gonadotropin releasing hormone |
| KNDy | Kisspeptin-Neurokinin B-Dynorphin |
| hCG | Human chorionic gonadotropin |
References
- Grinspon, R.P.; Bergadá, I.; Rey, R.A. Male Hypogonadism and Disorders of Sex Development. Front Endocrinol (Lausanne) 2020, 11, 211. [CrossRef]
- Sheng, J.A.; Bales, N.J.; Myers, S.A.; Bautista, A.I.; Roueinfar, M.; Hale, T.M.; Handa, R.J. The Hypothalamic-Pituitary-Adrenal Axis: Development, Programming Actions of Hormones, and Maternal-Fetal Interactions. Front Behav Neurosci 2020, 14, 601939. [CrossRef]
- Kuiri-Hänninen, T.; Sankilampi, U.; Dunkel, L. Activation of the hypothalamic-pituitary-gonadal axis in infancy: minipuberty. Horm Res Paediatr 2014, 82, 73-80. [CrossRef]
- Lanciotti, L.; Cofini, M.; Leonardi, A.; Penta, L.; Esposito, S. Up-To-Date Review About Minipuberty and Overview on Hypothalamic-Pituitary-Gonadal Axis Activation in Fetal and Neonatal Life. Front Endocrinol (Lausanne) 2018, 9, 410. [CrossRef]
- Kiviranta, P.; Kuiri-Hänninen, T.; Saari, A.; Lamidi, M.L.; Dunkel, L.; Sankilampi, U. Transient Postnatal Gonadal Activation and Growth Velocity in Infancy. Pediatrics 2016, 138. [CrossRef]
- Hines, M.; Spencer, D.; Kung, K.T.; Browne, W.V.; Constantinescu, M.; Noorderhaven, R.M. The early postnatal period, mini-puberty, provides a window on the role of testosterone in human neurobehavioural development. Curr Opin Neurobiol 2016, 38, 69-73. [CrossRef]
- Balasubramanian, R.; Dwyer, A.; Seminara, S.B.; Pitteloud, N.; Kaiser, U.B.; Crowley, W.F., Jr. Human GnRH deficiency: a unique disease model to unravel the ontogeny of GnRH neurons. Neuroendocrinology 2010, 92, 81-99. [CrossRef]
- Schwanzel-Fukuda, M.; Crossin, K.L.; Pfaff, D.W.; Bouloux, P.M.; Hardelin, J.P.; Petit, C. Migration of luteinizing hormone-releasing hormone (LHRH) neurons in early human embryos. J Comp Neurol 1996, 366, 547-557. [CrossRef]
- Maeda, K.; Ohkura, S.; Uenoyama, Y.; Wakabayashi, Y.; Oka, Y.; Tsukamura, H.; Okamura, H. Neurobiological mechanisms underlying GnRH pulse generation by the hypothalamus. Brain Res 2010, 1364, 103-115. [CrossRef]
- Uenoyama, Y.; Nagae, M.; Tsuchida, H.; Inoue, N.; Tsukamura, H. Role of KNDy Neurons Expressing Kisspeptin, Neurokinin B, and Dynorphin A as a GnRH Pulse Generator Controlling Mammalian Reproduction. Front Endocrinol (Lausanne) 2021, 12, 724632. [CrossRef]
- Velasco, I.; Franssen, D.; Daza-Dueñas, S.; Skrapits, K.; Takács, S.; Torres, E.; Rodríguez-Vazquez, E.; Ruiz-Cruz, M.; León, S.; Kukoricza, K.; et al. Dissecting the KNDy hypothesis: KNDy neuron-derived kisspeptins are dispensable for puberty but essential for preserved female fertility and gonadotropin pulsatility. Metabolism 2023, 144, 155556. [CrossRef]
- Beltramo, M.; Dardente, H.; Cayla, X.; Caraty, A. Cellular mechanisms and integrative timing of neuroendocrine control of GnRH secretion by kisspeptin. Mol Cell Endocrinol 2014, 382, 387-399. [CrossRef]
- Tena-Sempere, M. Hypothalamic KiSS-1: the missing link in gonadotropin feedback control? Endocrinology 2005, 146, 3683-3685. [CrossRef]
- Clements, J.A.; Reyes, F.I.; Winter, J.S.; Faiman, C. Studies on human sexual development. III. Fetal pituitary and serum, and amniotic fluid concentrations of LH, CG, and FSH. J Clin Endocrinol Metab 1976, 42, 9-19. [CrossRef]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Origin of luteinizing hormone-releasing hormone neurons. Nature 1989, 338, 161-164. [CrossRef]
- Pilavdzic, D.; Kovacs, K.; Asa, S.L. Pituitary morphology in anencephalic human fetuses. Neuroendocrinology 1997, 65, 164-172. [CrossRef]
- Guimiot, F.; Chevrier, L.; Dreux, S.; Chevenne, D.; Caraty, A.; Delezoide, A.L.; de Roux, N. Negative fetal FSH/LH regulation in late pregnancy is associated with declined kisspeptin/KISS1R expression in the tuberal hypothalamus. J Clin Endocrinol Metab 2012, 97, E2221-2229. [CrossRef]
- Massa, G.; de Zegher, F.; Vanderschueren-Lodeweyckx, M. Serum levels of immunoreactive inhibin, FSH, and LH in human infants at preterm and term birth. Biol Neonate 1992, 61, 150-155. [CrossRef]
- Takagi, S.; Yoshida, T.; Tsubata, K.; Ozaki, H.; Fujii, T.K.; Nomura, Y.; Sawada, M. Sex differences in fetal gonadotropins and androgens. J Steroid Biochem 1977, 8, 609-620. [CrossRef]
- Debieve, F.; Beerlandt, S.; Hubinont, C.; Thomas, K. Gonadotropins, prolactin, inhibin A, inhibin B, and activin A in human fetal serum from midpregnancy and term pregnancy. J Clin Endocrinol Metab 2000, 85, 270-274. [CrossRef]
- Casarini, L.; Santi, D.; Brigante, G.; Simoni, M. Two Hormones for One Receptor: Evolution, Biochemistry, Actions, and Pathophysiology of LH and hCG. Endocr Rev 2018, 39, 549-592. [CrossRef]
- Pitetti, J.L.; Calvel, P.; Romero, Y.; Conne, B.; Truong, V.; Papaioannou, M.D.; Schaad, O.; Docquier, M.; Herrera, P.L.; Wilhelm, D.; et al. Insulin and IGF1 receptors are essential for XX and XY gonadal differentiation and adrenal development in mice. PLoS Genet 2013, 9, e1003160. [CrossRef]
- O'Rahilly, R. The timing and sequence of events in the development of the human reproductive system during the embryonic period proper. Anat Embryol (Berl) 1983, 166, 247-261. [CrossRef]
- Reyes, F.I.; Boroditsky, R.S.; Winter, J.S.; Faiman, C. Studies on human sexual development. II. Fetal and maternal serum gonadotropin and sex steroid concentrations. J Clin Endocrinol Metab 1974, 38, 612-617. [CrossRef]
- O'Shaughnessy, P.J.; Baker, P.J.; Monteiro, A.; Cassie, S.; Bhattacharya, S.; Fowler, P.A. Developmental changes in human fetal testicular cell numbers and messenger ribonucleic acid levels during the second trimester. J Clin Endocrinol Metab 2007, 92, 4792-4801. [CrossRef]
- Kurilo, L.F. Oogenesis in antenatal development in man. Hum Genet 1981, 57, 86-92. [CrossRef]
- Cole, B.; Hensinger, K.; Maciel, G.A.; Chang, R.J.; Erickson, G.F. Human fetal ovary development involves the spatiotemporal expression of p450c17 protein. J Clin Endocrinol Metab 2006, 91, 3654-3661. [CrossRef]
- Forabosco, A.; Sforza, C. Establishment of ovarian reserve: a quantitative morphometric study of the developing human ovary. Fertil Steril 2007, 88, 675-683. [CrossRef]
- Baker, T.G.; Scrimgeour, J.B. Development of the gonad in normal and anencephalic human fetuses. J Reprod Fertil 1980, 60, 193-199. [CrossRef]
- Rohayem, J.; Alexander, E.C.; Heger, S.; Nordenström, A.; Howard, S.R. Mini-Puberty, Physiological and Disordered: Consequences, and Potential for Therapeutic Replacement. Endocr Rev 2024, 45, 460-492. [CrossRef]
- Forest, M.G.; Cathiard, A.M.; Bertrand, J.A. Evidence of testicular activity in early infancy. J Clin Endocrinol Metab 1973, 37, 148-151. [CrossRef]
- Corbier, P.; Dehennin, L.; Castanier, M.; Mebazaa, A.; Edwards, D.A.; Roffi, J. Sex differences in serum luteinizing hormone and testosterone in the human neonate during the first few hours after birth. J Clin Endocrinol Metab 1990, 71, 1344-1348. [CrossRef]
- Bergadá, I.; Milani, C.; Bedecarrás, P.; Andreone, L.; Ropelato, M.G.; Gottlieb, S.; Bergadá, C.; Campo, S.; Rey, R.A. Time course of the serum gonadotropin surge, inhibins, and anti-Müllerian hormone in normal newborn males during the first month of life. J Clin Endocrinol Metab 2006, 91, 4092-4098. [CrossRef]
- Busch, A.S.; Ljubicic, M.L.; Upners, E.N.; Fischer, M.B.; Raket, L.L.; Frederiksen, H.; Albrethsen, J.; Johannsen, T.H.; Hagen, C.P.; Juul, A. Dynamic Changes of Reproductive Hormones in Male Minipuberty: Temporal Dissociation of Leydig and Sertoli Cell Activity. J Clin Endocrinol Metab 2022, 107, 1560-1568. [CrossRef]
- Schmidt, H.; Schwarz, H.P. Serum concentrations of LH and FSH in the healthy newborn. Eur J Endocrinol 2000, 143, 213-215. [CrossRef]
- Andersson, A.M.; Toppari, J.; Haavisto, A.M.; Petersen, J.H.; Simell, T.; Simell, O.; Skakkebaek, N.E. Longitudinal reproductive hormone profiles in infants: peak of inhibin B levels in infant boys exceeds levels in adult men. J Clin Endocrinol Metab 1998, 83, 675-681. [CrossRef]
- Boas, M.; Boisen, K.A.; Virtanen, H.E.; Kaleva, M.; Suomi, A.M.; Schmidt, I.M.; Damgaard, I.N.; Kai, C.M.; Chellakooty, M.; Skakkebaek, N.E.; et al. Postnatal penile length and growth rate correlate to serum testosterone levels: a longitudinal study of 1962 normal boys. Eur J Endocrinol 2006, 154, 125-129. [CrossRef]
- Dhayat, N.A.; Dick, B.; Frey, B.M.; d'Uscio, C.H.; Vogt, B.; Flück, C.E. Androgen biosynthesis during minipuberty favors the backdoor pathway over the classic pathway: Insights into enzyme activities and steroid fluxes in healthy infants during the first year of life from the urinary steroid metabolome. J Steroid Biochem Mol Biol 2017, 165, 312-322. [CrossRef]
- Bay, K.; Virtanen, H.E.; Hartung, S.; Ivell, R.; Main, K.M.; Skakkebaek, N.E.; Andersson, A.M.; Toppari, J. Insulin-like factor 3 levels in cord blood and serum from children: effects of age, postnatal hypothalamic-pituitary-gonadal axis activation, and cryptorchidism. J Clin Endocrinol Metab 2007, 92, 4020-4027. [CrossRef]
- Burton, E.; Abeydeera, S.A.; Sarila, G.; Cho, H.J.; Wu, S.; Tien, M.Y.; Hutson, J.; Li, R. The role of gonadotrophins in gonocyte transformation during minipuberty. Pediatr Surg Int 2020, 36, 1379-1385. [CrossRef]
- Simorangkir, D.R.; Marshall, G.R.; Plant, T.M. Sertoli cell proliferation during prepubertal development in the rhesus monkey (Macaca mulatta) is maximal during infancy when gonadotropin secretion is robust. J Clin Endocrinol Metab 2003, 88, 4984-4989. [CrossRef]
- Chemes, H.E.; Rey, R.A.; Nistal, M.; Regadera, J.; Musse, M.; González-Peramato, P.; Serrano, A. Physiological androgen insensitivity of the fetal, neonatal, and early infantile testis is explained by the ontogeny of the androgen receptor expression in Sertoli cells. J Clin Endocrinol Metab 2008, 93, 4408-4412. [CrossRef]
- Rey, R.A. Mini-puberty and true puberty: differences in testicular function. Ann Endocrinol (Paris) 2014, 75, 58-63. [CrossRef]
- Aksglaede, L.; Sørensen, K.; Boas, M.; Mouritsen, A.; Hagen, C.P.; Jensen, R.B.; Petersen, J.H.; Linneberg, A.; Andersson, A.M.; Main, K.M.; et al. Changes in anti-Müllerian hormone (AMH) throughout the life span: a population-based study of 1027 healthy males from birth (cord blood) to the age of 69 years. J Clin Endocrinol Metab 2010, 95, 5357-5364. [CrossRef]
- Winter, J.S.; Hughes, I.A.; Reyes, F.I.; Faiman, C. Pituitary-gonadal relations in infancy: 2. Patterns of serum gonadal steroid concentrations in man from birth to two years of age. J Clin Endocrinol Metab 1976, 42, 679-686. [CrossRef]
- Kuiri-Hänninen, T.; Kallio, S.; Seuri, R.; Tyrväinen, E.; Liakka, A.; Tapanainen, J.; Sankilampi, U.; Dunkel, L. Postnatal developmental changes in the pituitary-ovarian axis in preterm and term infant girls. J Clin Endocrinol Metab 2011, 96, 3432-3439. [CrossRef]
- Kuiri-Hänninen, T.; Haanpää, M.; Turpeinen, U.; Hämäläinen, E.; Seuri, R.; Tyrväinen, E.; Sankilampi, U.; Dunkel, L. Postnatal ovarian activation has effects in estrogen target tissues in infant girls. J Clin Endocrinol Metab 2013, 98, 4709-4716. [CrossRef]
- Bidlingmaier, F.; Strom, T.M.; Dörr, H.G.; Eisenmenger, W.; Knorr, D. Estrone and estradiol concentrations in human ovaries, testes, and adrenals during the first two years of life. J Clin Endocrinol Metab 1987, 65, 862-867. [CrossRef]
- Cohen, H.L.; Shapiro, M.A.; Mandel, F.S.; Shapiro, M.L. Normal ovaries in neonates and infants: a sonographic study of 77 patients 1 day to 24 months old. AJR Am J Roentgenol 1993, 160, 583-586. [CrossRef]
- Courant, F.; Aksglaede, L.; Antignac, J.P.; Monteau, F.; Sorensen, K.; Andersson, A.M.; Skakkebaek, N.E.; Juul, A.; Bizec, B.L. Assessment of circulating sex steroid levels in prepubertal and pubertal boys and girls by a novel ultrasensitive gas chromatography-tandem mass spectrometry method. J Clin Endocrinol Metab 2010, 95, 82-92. [CrossRef]
- Jayasinghe, Y.; Cha, R.; Horn-Ommen, J.; O'Brien, P.; Simmons, P.S. Establishment of normative data for the amount of breast tissue present in healthy children up to two years of age. J Pediatr Adolesc Gynecol 2010, 23, 305-311. [CrossRef]
- Schmidt, I.M.; Chellakooty, M.; Haavisto, A.M.; Boisen, K.A.; Damgaard, I.N.; Steendahl, U.; Toppari, J.; Skakkebaek, N.E.; Main, K.M. Gender difference in breast tissue size in infancy: correlation with serum estradiol. Pediatr Res 2002, 52, 682-686. [CrossRef]
- Nguyen, R.H.; Umbach, D.M.; Parad, R.B.; Stroehla, B.; Rogan, W.J.; Estroff, J.A. US assessment of estrogen-responsive organ growth among healthy term infants: piloting methods for assessing estrogenic activity. Pediatr Radiol 2011, 41, 633-642. [CrossRef]
- Devillers, M.M.; Mhaouty-Kodja, S.; Guigon, C.J. Deciphering the Roles & Regulation of Estradiol Signaling during Female Mini-Puberty: Insights from Mouse Models. Int J Mol Sci 2022, 23. [CrossRef]
- Tapanainen, J.; Koivisto, M.; Vihko, R.; Huhtaniemi, I. Enhanced activity of the pituitary-gonadal axis in premature human infants. J Clin Endocrinol Metab 1981, 52, 235-238. [CrossRef]
- Shinkawa, O.; Furuhashi, N.; Fukaya, T.; Suzuki, M.; Kono, H.; Tachibana, Y. Changes of serum gonadotropin levels and sex differences in premature and mature infant during neonatal life. J Clin Endocrinol Metab 1983, 56, 1327-1331. [CrossRef]
- Kuiri-Hänninen, T.; Seuri, R.; Tyrväinen, E.; Turpeinen, U.; Hämäläinen, E.; Stenman, U.H.; Dunkel, L.; Sankilampi, U. Increased activity of the hypothalamic-pituitary-testicular axis in infancy results in increased androgen action in premature boys. J Clin Endocrinol Metab 2011, 96, 98-105. [CrossRef]
- Nagai, S.; Kawai, M.; Myowa-Yamakoshi, M.; Morimoto, T.; Matsukura, T.; Heike, T. Gonadotropin levels in urine during early postnatal period in small for gestational age preterm male infants with fetal growth restriction. J Perinatol 2017, 37, 843-847. [CrossRef]
- Pepe, G.; Calafiore, M.; Velletri, M.R.; Corica, D.; Valenzise, M.; Mondello, I.; Alibrandi, A.; Wasniewska, M.; Aversa, T. Minipuberty in born small for gestational age infants: A case control prospective pilot study. Endocrine 2022, 76, 465-473. [CrossRef]
- Kerkhof, G.F.; Leunissen, R.W.; Willemsen, R.H.; de Jong, F.H.; Stijnen, T.; Hokken-Koelega, A.C. Influence of preterm birth and birth size on gonadal function in young men. J Clin Endocrinol Metab 2009, 94, 4243-4250. [CrossRef]
- Boonstra, V.H.; Weber, R.F.; de Jong, F.H.; Hokken-Koelega, A.C. Testis function in prepubertal boys and young men born small for gestational age. Horm Res 2008, 70, 357-363. [CrossRef]
- Jensen, R.B.; Vielwerth, S.; Larsen, T.; Greisen, G.; Veldhuis, J.; Juul, A. Pituitary-gonadal function in adolescent males born appropriate or small for gestational age with or without intrauterine growth restriction. J Clin Endocrinol Metab 2007, 92, 1353-1357. [CrossRef]
- Sedin, G.; Bergquist, C.; Lindgren, P.G. Ovarian hyperstimulation syndrome in preterm infants. Pediatr Res 1985, 19, 548-552. [CrossRef]
- Lee, Y.L.; Jamli, F.M. Preterm ovarian hyperstimulation syndrome presenting as clitoromegaly in a premature female infant. Arch Dis Child 2022, 107, 166-167. [CrossRef]
- Mosallanejad, A.; Tabatabai, S.; Shakiba, M.; Alaei, M.R.; Saneifard, H. A Rare Case of Ovarian Hyperstimulation Syndrome in a Preterm Infant. J Clin Diagn Res 2016, 10, Sd07-sd08. [CrossRef]
- Altuntas, N.; Turkyilmaz, C.; Yuce, O.; Kulali, F.; Hirfanoglu, I.M.; Onal, E.; Ergenekon, E.; Koç, E.; Bideci, A.; Atalay, Y. Preterm ovarian hyperstimulation syndrome presented with vaginal bleeding: a case report. J Pediatr Endocrinol Metab 2014, 27, 355-358. [CrossRef]
- Sun, Y.; Chen, C.; Di, T.; Zhu, Y.; Zhu, R.; Chen, S.; Qian, Y. Clinical characteristics of preterm ovarian hyperstimulation syndrome: seven cases from China and 14 cases from the literature. Gynecol Endocrinol 2019, 35, 819-824. [CrossRef]
- Fields, D.A.; Gilchrist, J.M.; Catalano, P.M.; Giannì, M.L.; Roggero, P.M.; Mosca, F. Longitudinal body composition data in exclusively breast-fed infants: a multicenter study. Obesity (Silver Spring) 2011, 19, 1887-1891. [CrossRef]
- Fomon, S.J.; Nelson, S.E. Body composition of the male and female reference infants. Annu Rev Nutr 2002, 22, 1-17. [CrossRef]
- Andersen, G.S.; Girma, T.; Wells, J.C.; Kæstel, P.; Leventi, M.; Hother, A.L.; Michaelsen, K.F.; Friis, H. Body composition from birth to 6 mo of age in Ethiopian infants: reference data obtained by air-displacement plethysmography. Am J Clin Nutr 2013, 98, 885-894. [CrossRef]
- Jain, V.; Kurpad, A.V.; Kumar, B.; Devi, S.; Sreenivas, V.; Paul, V.K. Body composition of term healthy Indian newborns. Eur J Clin Nutr 2016, 70, 488-493. [CrossRef]
- Davis, S.M.; Kaar, J.L.; Ringham, B.M.; Hockett, C.W.; Glueck, D.H.; Dabelea, D. Sex differences in infant body composition emerge in the first 5 months of life. J Pediatr Endocrinol Metab 2019, 32, 1235-1239. [CrossRef]
- Chachlaki, K.; Le Duc, K.; Storme, L.; Prevot, V. Novel insights into minipuberty and GnRH: Implications on neurodevelopment, cognition, and COVID-19 therapeutics. J Neuroendocrinol 2024, 36, e13387. [CrossRef]
- Kung, K.T.; Browne, W.V.; Constantinescu, M.; Noorderhaven, R.M.; Hines, M. Early postnatal testosterone predicts sex-related differences in early expressive vocabulary. Psychoneuroendocrinology 2016, 68, 111-116. [CrossRef]
- Lombardo, M.V.; Ashwin, E.; Auyeung, B.; Chakrabarti, B.; Taylor, K.; Hackett, G.; Bullmore, E.T.; Baron-Cohen, S. Fetal testosterone influences sexually dimorphic gray matter in the human brain. J Neurosci 2012, 32, 674-680. [CrossRef]
- Hines, M.; Constantinescu, M.; Spencer, D. Early androgen exposure and human gender development. Biol Sex Differ 2015, 6, 3. [CrossRef]
Figure 1.
Schematic representation of LH, FSH, and testosterone changes during fetal and early infantile life in healthy boys. FSH: follicle-stimulating hormone, LH: luteinizing hormone.
Figure 1.
Schematic representation of LH, FSH, and testosterone changes during fetal and early infantile life in healthy boys. FSH: follicle-stimulating hormone, LH: luteinizing hormone.
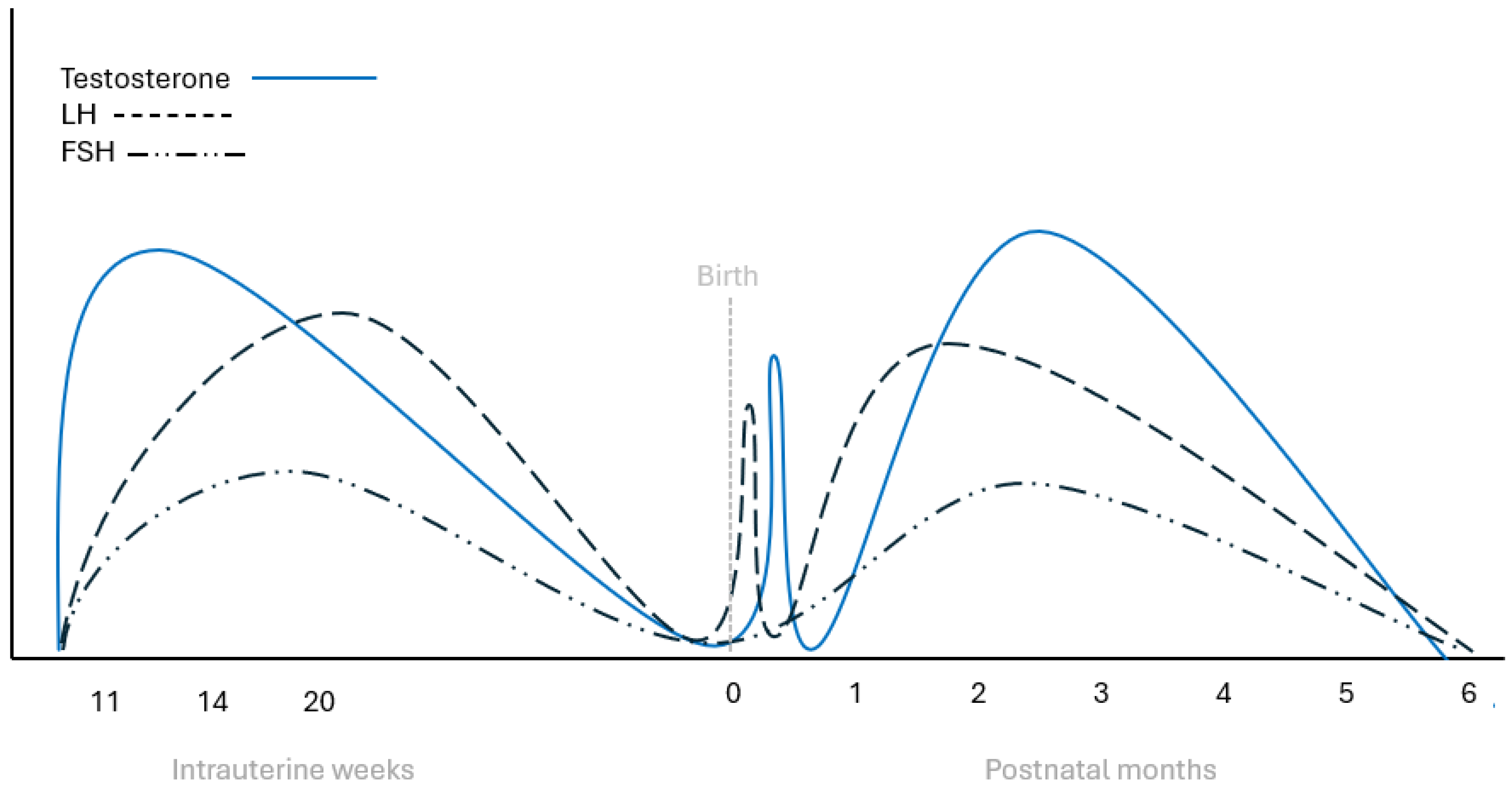
Figure 2.
Schematic representation of LH, FSH, and estradiol changes during fetal and early infantile life in healthy girls. FSH: follicle-stimulating hormone, LH: luteinizing hormone.
Figure 2.
Schematic representation of LH, FSH, and estradiol changes during fetal and early infantile life in healthy girls. FSH: follicle-stimulating hormone, LH: luteinizing hormone.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



