Submitted:
13 February 2025
Posted:
17 February 2025
You are already at the latest version
Abstract
Globally, skin cancer accounts for more than all other malignancies combined. Skin cancer imposes a significant financial and societal burden. UVR exposure is responsible for 90% of skin cancer and is a major source of exogenous free radicals. The free radical theory of aging and cancer led to the investigation of polyunsaturated lipids, antioxidants (including synthetic phenols, polyphenols, flavonoids) and ß-carotene as potential modulators of UVR-carcinogenesis. Experimentally, both dietary lipids and antioxidants modified the tumor latent period and multiplicity. Increasing levels of omega-6 fatty acids exacerbated UVR-carcinogenesis in a near linear fashion. However, omega-3 fatty acid inhibited UVR-carcinogenesis. Both types of lipids exhibit the same level of saturation. Thus, the inhibition of UVR-carcinogenesis by omega-3 fatty acid must rest with differential metabolic intermediates that each generate, both tumor-promoting and immune-modulating. Clinically, a low-fat dietary intervention trial has shown that reduced fat intake significantly reduces the occurrence of nonmelanoma skin cancer. Dietary supplementation of antioxidants and ß-carotene into the complex milieu of the cell with its own intricate and complex free radical defense system has resulted in detrimental responses and antioxidant dietary supplements, as well as ß-carotene, are no longer recommended for the public as a mean to prevent cancer.
Keywords:
UVR
; carcinogenesis
; nonmelanoma skin cancer
; Melanoma
; Lipids
; Omega-3/Omega-6 fatty acids
; antioxidants
; carotenoids
1. Introduction
Cancer is the second leading cause of death, both globally and in the U.S. [1,2]. The economic and societal burden of cancer is great, accounting for 1.8% of the gross domestic product (GDP) in the US and 1.07% of the European Union GDP in 2017 [3,4]. Total number of procedures for skin cancer increased by 76.9% from 1,158,298 in 1992 to 2,048,517 in 2006 [5]. In 2012, 5.4 million cases of NMSC were treated
in 3.3 million people in the US. More patients are diagnosed with skin cancer in the US than all other malignancies combined, with an annual cost of treatment of ~ 8.1 billion, about 4,444.8 billion for NMSC and 3.5 billion for melanoma. [5,6]. Furthermore, metabolic disease involving altered glucose metabolism, e.g., Type 2 Diabetes, is a known risk factor for many forms of cancer, including skin cancer [7,8]. Near epidemic increases in diabetes, with
increased risk for skin cancer, suggests that these two diseases will present
an even greater global health challenge in the future.
The supposition that diet could influence cancer
was recognized by early physicians who advocated “fames cura”, or a starvation
diet, as treatment for cancer [9]. These early
medical practitioners surmised that cancer, as well as other human diseases,
proceeded from the diet the patient consumed. Although such a clinical approach
to cancer treatment was gradually abandoned as it was generally ineffective, as
practiced, and it became obvious that several metabolic abnormalities resulted
from this treatment, e.g., severe malnutrition, cachexia, and death. These
conditions were compatible with the thesis that cancer’s lethality resulted
from depleting the host of essential nutrients [10].
Thus, the early recognition that diet was an important aspect of cancer
development narrowed the search for dietary constituents that played a
significant role in neoplastic development and posited the idea that these
constituents might be manipulated to prevent cancer development. These ideas
came further into focus with the development of the free-radical theory of
aging and cancer and the dietary recommendations posed by Harmon that (1)
dietary reduction of vulnerable free radical targets such as polyunsaturated
fats and (2) supplement the diet with one or more free radical reaction
inhibitors [11,12]. It has become clear that
these tenets must be reassessed [13]. This
review addresses both lipid and antioxidant roles in skin cancer prevention -
first their experimental basis for involvement, followed by clinical studies
and their relevance.
2. Lipids
2.1. Experimental
Whereas the first report, in 1930, that dietary fat
enhanced coal tar-induced skin tumors in mice [14],
about 90% of all human skin cancers are induced by solar ultraviolet radiation
(UVR) [15]. Nine years after this initial
finding of chemically induced skin carcinogenesis, it was reported that dietary
fat had an exacerbating effect on UVR-induced skin carcinogenesis [16]. Due to the intervention of World War II, this
thread of investigation into UVR carcinogenesis was dropped and was not resumed
until the 1980’s. In 1983 it was shown that increasing the level of dietary
omega-6 polyunsaturated fatty acids (corn oil, ~66% linoleic acid) exacerbated
UVR carcinogenic expression, with respect to both tumor latency and
multiplicity, and in a near linear fashion with regard to lipid concentration [17,18]. Importantly, when animals were fed a diet
containing 4% corn oil that was partially hydrogenated (60%), tumor latency was
markedly lengthened, and tumor multiplicity reduced when compared to the normal
4% corn oil diet. Hydrogenation reduced the level of linoleic acid (LA), an
essential fatty acid (EFA) and suggested that the effect on UVR carcinogenesis
was a function of LA concentration. Indeed, Ip et al. [19] had demonstrated that LA content of diets had a
direct relationship to mammary tumor incidence and yield, suggesting that LA
was not only a potentiator, but perhaps a requirement for effective expression
of carcinogenesis. Further, Reeve et al. [20]
reported that feeding a diet supplying totally saturated sunflower oil
(hydrogenated) completely blocked the UVR carcinogenic response, whereas those
animals receiving the polyunsaturated sunflower oil (approximately 65% LA)
exhibited 100% tumor incidence.
There are two known EFA that are required by humans
and animals, Linoleic acid and Alpha-Linolenic Acid (ALA). The latter undergoes
a series of desaturation and elongation reactions that yield omega-3 FA, long
chain polyunsaturated eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids.
Orengo et. al. [21], employing a semi-defined
diet containing menhaden oil (27% omega-3 FA) were first to demonstrate that a
dietary source of omega-3 FA, when compared to omega-6 FA diets (corn oil),
inhibited UVR carcinogenesis as manifested in significant longer tumor latent
period and lower tumor multiplicity, as depicted in Table 1. After feeding the menhaden oil diets
for two weeks, edema was markedly decreased and animals twice the level of UVR
to evoke a comparable level of erythema, compared to corn oil fed animals.

Table 1.
Effect of Low-Fat Diet on Incidence and Occurrence of NMSC. * Last eight-month period of the 2-year study. ** First vs last eight-month period of study.
Table 1.
Effect of Low-Fat Diet on Incidence and Occurrence of NMSC. * Last eight-month period of the 2-year study. ** First vs last eight-month period of study.
 |
Reeve et al. [20]
speculated that the lack of eicosanoid precursors, as occurs in EFA, i.e., LA
deficiency, might prevent the establishment UVR-induction of the
immunosuppressed state. When animals were fed saturated fat diets, no tumors
developed, but when returned to a normal diet containing LA, alleviating tumor
suppression, rapid outgrowth of skin tumors occurred. Indeed, Chung, et al. [22] had shown that suppressor T-cell function was
PGE2 dependent and had correlated the abrogation of UVR-induced
suppression of contact hypersensitivity, an immune response sharing pathways
with tumor rejection, with prostaglandin synthesis. Henderson et al., [23] examined PGE2 levels after two weeks
feeding omega-6 and omega-3 dietary sources and found that plasma PGE2 levels
in the corn oil group were approximately 6-fold greater than those in the
menhaden oil group. Cutaneous levels were approximately 2.5-fold greater in the
corn oil fed group. Free radicals are known to activate Phospholipase A2 that
results in the release of Arachidonic acid (AA) from cell membrane
phospholipids. AA, when metabolized through the cyclooxygenase pathway yields
proinflammatory prostaglandins where EPA results in less inflammatory
prostaglandins. Orengo et. al. [24], in a
randomized dose-response study, found that animals receiving Celecoxib, a
cyclooxygenase 2 inhibitor, at human dose equivalents, exhibited significantly
lengthened tumor latency and reduced tumor multiplicity. In this regard, the
levels of inflammatory biomarkers in the plasma and skin of UVR-animals
receiving high-fat diets were markedly increased, compared to control diets [25]. The biomarkers included tumor necrosis
factor-alpha, cyclooxygenase-2, PGE2, interleukin-1beta,
interleukin-6, as well as proliferating cell nuclear antigen and cell survival
signals (phosphatidylinositol-3-kinase and p-Akt-Ser. The pronounced
exacerbation of the UVR-induced inflammatory responses in the skin of animals
fed the high-fat diet provides further evidence that a high-fat diet may
increase susceptibility to inflammation associated with the risk of skin
cancer.
Utsumi, et al.,[26]
obtained ESR spectra using a surface resonator and L-band ESR spectrometer. The
surface resonator was attached to the skin of griseofulvin (GF)-induced
protoporphyria mice that had been administered a systemic light probe. Upon
irradiation with a tungsten lamp (mostly visible light) the decay rates of the
signals (an indication of Reactive Oxygen Species (ROS)) formation,
significantly increased in the GF-fed mice, but it had little effect on this
parameter in non-irradiated control animals. Conversely, when the light was
switched off, there was rapid deceleration of the decay rate to the original
level [27]. The in vivo radical
detection in the skin should be invaluable for examination of ROS and other
free radicals that initiate damaging oxidative reactions. Oxidative stress is a
consequence of excessive ROS production and manifests a link to increased risk
of skin cancer [28]. Indeed, it has been
demonstrated that levels of one marker (5-hydroxymethyl-2’-deoxyuridine) of
oxidative damage to DNA in the plasma increased as dietary fat levels (corn
oil) increased from 3% to 15% [29].
Further investigations into potential
pathophysiologic parameters that resulted after feeding animals omega-6 and
omega-3 dietary sources revealed that epidermal, plasma and Red Blood Cell
(RBC) membrane FA composition exhibited marked differences, although these
changes were not accompanied with drastic effects upon membrane fragility [30]. Epidermal capacity to metabolize Arachidonic
acid (AA) through the cyclooxygenase pathway, and thus formation of PGE2,
was closely related to omega-6 FA intake. Animals fed the omega-3 FA exhibited
markedly lower plasma PGE2 levels, even when the diet was
supplemented with omega-6 FA. Importantly, UVR exposed animals receiving the
omega-3 FA source demonstrated a reduced (~30%) response to inflammatory
stimulus and a greater (4.5-fold) delayed type hypersensitivity (DTH) to
dinitrochlorobenzene (DNCB) than animals fed the omega-6 source. The influence
of omega-3 FA on immune responsiveness was confirmed when dietary EPA was shown
to prevent systemic immunosuppression in UVR-radiated mice. [31].
Additional evidence for the role of omega-3 FA in
carcinogenesis was obtained in studies employing a transgenic mouse model
designated fat-1 {32]. The transgenic mice can produce omega-3 FA from
omega-6 FA, i.e., they have received a gene encoding an omega-3 FA desaturase
that converts omega-6 FA to omega-3 FA. This results in an abundance of omega-3
and reduced omega-6 FA. It was shown that a dramatic reduction in melanoma
formation and growth occurred when fat-1 mice were injected with B16
melanoma cells, compared to their non-transgenic littermates [33]. The levels of omega-3 FA and metabolite PGE3,
(a less inflammatory PG than PGE2) were much higher in the
transgenic animals, and the omega-6/omega-3 FA ratio was much lower
Level of dietary omega-6 FA potentiates immune
suppression in UVR treated mice [34,35].
Animals, when fed low fat or high fat, 0.75% or 12% corn oil respectively, and
sensitized to DNCB, expressed significant DTH suppression in the high fat group
– even before UVR. Although both groups exhibited UVR-induced suppression of
this response, the high fat group was totally suppressed after three weeks of
UVR, whereas the low-fat group exhibited reactivity through eight weeks.
Splenic T-lymphocyte (Thy 1.2+) population had declined by about 50%
at time of UVR termination (11 weeks) whereas dietary lipid exerted no apparent
influence upon this T-cell population. However, after six weeks of UVR, I-J+
cells (a marker shown to be acquired adaptively by suppressor T lymphocytes)
began to increase and by week 15 had increased by about 65% in the high-fat
group – twice the % increase that occurred in the low-fat group. When
UVR-induced tumors were transplanted to recipient animals receiving various
periods of UVR (0, 6, 11 weeks), no significant differences in median tumor
rejection times between the dietary groups occurred at 0 and 6 weeks. However,
after 11 weeks of UVR, the low-fat group exhibited a median tumor rejection
time that was comparable to that of non-irradiated animals. Median tumor
rejection time for the high-fat group was 63 days, significantly longer than
that of the low-fat group (~20 days). These data clearly demonstrate that the
level of dietary fat (omega-6 FA) potentiates immune suppression in UVR exposed
animals and is compatible with the thesis that immune suppression may account
for the exacerbation of carcinogenic expression elicited by high fat dietary
levels.
Earlier studies had suggested that the primary
effect of dietary lipid (66% LA) in the carcinogenic continuum occurred during
the post-initiation phase [20]. To determine
the segment along the carcinogenic continuum at which dietary lipid exerts its
principal effect, animals were placed on defined isocaloric diets containing
either 0.75%, 4%, 12% corn oil or 4% and 12% menhaden oil as sources of omega-6
or omega-3 FA [36]. The animals received an
11-week course of UVR. Upon termination of UVR, some groups were crossed-over
to either low fat, high-fat, omega-6 or omega-3 FA diets. The first tumors
appeared in week 14. When animals were fed high-fat corn oil diets during initiation
and crossed to low fat corn oil diets immediately post UVR, tumor incidence
curves were not significantly different from animals fed the low-fat diet
throughout the study. When animals were fed the low-fat diet and crossed to
high-fat, the tumor incidence curves were not significantly different from that
of animals fed the high-fat diet throughout the study. Crossing menhaden oil
diets to high corn oil post UVR resulted in exacerbation of carcinogenic
expression like that when crossing low corn oil to high corn diets. However,
when crossing from high corn oil to menhaden oil did not result in the
ameliorating effect on tumor multiplicity as seen when crossing from high to
low corn oil diets or when menhaden oil was fed throughout the study. This suggests
that menhaden oil either exerts its anti-carcinogenic effect during the UVR
initiation stage or that promotion events had progressed beyond the point where
dietary intervention could alter the course of carcinogenic expression.
Nevertheless, this study demonstrated that high dietary lipid (omega-6) level
enhances UVR-induced skin carcinogenesis, with respect to both tumor latent
period and tumor multiplicity and that this lipid-induced exacerbation of
carcinogenesis occurs at the post-initiation, or promotion stage of
carcinogenesis. Importantly modification of diet to a low lipid level (omega-6)
after UVR initiation negates the enhancement of a high level of this dietary
source. This forms the rationale and impetus for development of clinical
intervention. Diet and dietary factors are variables that must be controlled in
UVR carcinogenesis protocols [37].
Conclusions from the experimental studies may be
summarized as:
- Increasing levels of dietary omega-6 FA exacerbate UVR-carcinogenesis, both with respect to shorter tumor latent period and greater tumor multiplicity.
- Hydrogenation of the EFA, LA, decreases the level of exacerbation of UVR-carcinogenesis, apparently by lowering the level of LA.
- Dietary omega-3 FA markedly inhibit UVR-carcinogenesis.
- Plasma levels of PGE2, a cyclooxygenase intermediate and pro-inflammatory prostaglandin, increase in near linear fashion with increasing levels of omega-6 FA. Dietary omega -3 FA dramatically reduce PGE2 levels.
- High levels of omega-6 FA enhance cutaneous inflammatory response and suppress immune responses.
- Omega-6 FA exert its primary effect on UVR-carcinogenesis in the post-initiation, or promotion/progression stage of the carcinogenic continuum, whereas omega-3 FA exerts its primary effect during initiation, and across the carcinogenic continuum.
2.2. Clinical Studies
2.2.1. Dietary Fat Level
Clinical studies e.g., case-control and
prospective cohort studies, had failed to find a relationship with skin cancer
incidence with dietary fat intake [38–42]. On
the other hand, Thind [43], in an
international study, found a positive association of dietary fat with skin
cancer incidence. Analytical epidemiological studies have provided the
principal evidence associating dietary factors with cancer, but these
associations have not always proved to be clinically pertinent [43,45]. Whereas the experimental studies from
cross-over feeding studies, even after exposure to a carcinogenic dose of UVR,
has provided the rationale for low-fat dietary intervention to reduce skin
cancer incidence. The high prevalence of skin cancer and the relative risk of
skin patients developing subsequent skin cancers within two years [46], made it practical to initiate a low-fat
dietary intervention. One hundred and thirty-three skin cancer (basal and
squamous cell carcinoma) patients were recruited to the two-year study, of
which 115 successfully completed. Of those patients completing, fifty-eight
patients were assigned to the control arm of the study in which no dietary
changes were introduced. The fifty-seven patients assigned to the intervention
arm learned how to adopt low-fat eating habits to their food preferences and
were assigned a “fat gram goal” that defined the grams of fat that would
provide 20% of calories from fat. Baseline and follow-up dietary data were
compiled from 7-day food records, from which 4 days were selected for analysis.
Food records were verified for types of food, quantities, and methods of
preparation. Nutrient analyses were performed using the Minnesota Nutrition
Data System, NMSC were diagnosed by dermatologists who were blinded to the
patient’s group assignment. NMSC were confirmed by a dermatopathologist. Stability
of body weight and calorie intake were required to prevent any confounding
effects due to these variables. Patients in the intervention group consumed
higher levels of complex carbohydrates to compensate for the reduction in fat
calories. Patients in the intervention group reduced their percentage of fat
calories from 39% to 21% (47% reduction of fat calories) and maintained this
level for the remainder of the study. There were no significant differences in
total caloric intake, or body weight, between the control and intervention
groups. The potential for low-fat intervention became apparent after only 78
patients had completed the study [47]. A
significant difference in number of actinic keratosis (premalignant lesions)
between control and intervention groups occurred after 8-12 months. Patients in
the control group (high fat) were diagnosed with new keratosis four times as
often as those in the intervention group and were found to be at 4.7 times
greater risk of having one or more actinic keratoses during the two-year study
than patients in the low-fat intervention group. The effect of reduction in the
percentage of fat calories on NMSC occurrence was observed after 101 patients
had completed the study [48], and became even
stronger after all 115 patients had completed [49].
No changes between the two groups were seen until 16 months, at which time the
cumulative skin cancers per patient plateaued in the intervention group while
the numbers in the control group continued to increase. When skin cancer
occurrence was examined in 8-month intervals across the 2-year study, skin
cancer occurrence in the intervention group was significantly less in the last
8-month period than in the control group. This was reflected as a significant
improvement in the intervention group – the control and intervention group each
had 9 with NMSC in the first eight-month period whereas the control group had 9
patients with NMSC in the last 8-month period and the intervention group had
one (p< 0.05). Data are depicted in Table 2.
Table 2.
Effect of Low-Fat Diet on Incidence and Occurrence of NMSC. * Last eight-month period of the 2-year study. ** First vs last eight-month period of study.
Table 2.
Effect of Low-Fat Diet on Incidence and Occurrence of NMSC. * Last eight-month period of the 2-year study. ** First vs last eight-month period of study.
| Treatment | No. | BCC: SCC | NMSC/Patient* | Patients with MSC* | Improvement** |
|---|---|---|---|---|---|
| Control | 58 | 0.26 | 9 | 9 9 NS | |
| 52:1 | P>0.01 | P>0.02 | |||
| Intervention | 57 | 0.02 | 1 |
9 1P>0.05
|
After results from the previous intervention trial had been published, a large Randomized Controlled Dietary Modification Trial concluded that a low-fat dietary intervention had no effect on NMSC [42]. There are several limitations in the study protocol that could result in this null finding. For example:
- (1)
- The study involved only post-menopausal women as compared to both genders in the Intervention trial.
- (2)
- It relied on the Food Frequency Questionnaire (FFQ) to assess dietary intake. Participants were assessed at baseline and at one year and thereafter 33% of participants were assessed each year. On this basis, each participant completed a FFQ every three years. Dietary recall over such long assessment periods is notoriously inaccurate. The intervention trial dietary assessment in the intervention trial were compiled from 7-day food records and verified by a dietician.
- (3)
- Participants self-reported medical outcomes by completing questionnaires every six months. All participants in the intervention trial were assessed every four months by dermatologists.
- (4)
- Perhaps more importantly, pertaining to the null result, was the failure to reduce the fat level to the 20% target. The study reported a long-term difference (eight years) in the percentage of energy from total fat, versus the comparison group, of 8.1 %. The intervention trial reported a 47% reduction at month four that was maintained through month 24.
2.2.2. Type of Dietary Fat
Western diets may contain 15-20 times more LA than ALA (both EFA). Metabolic competition for shared enzymatic pathways, from whence the omega-3 FA, EPA and DHA, are derived, results in greater levels of AA than EPA and DHA. Consequently, under certain conditions, supplementation with EPA and DHA may be essential for maintenance of good health [50]. Interest in the health benefits of omega-3 FA resulted from reports that these FA were associated with low incidence of ischemic heart disease, as well as other inflammatory symptoms, in Greenlandic Eskimos [51,52]. Encouraged by animal studies in which omega-3 FA strongly inhibited inflammatory responses, i.e., erythema and edema, a short-term supplementation study of mixed omega-3 FA (EPA + DHA} was undertaken by Orengo et. al. [53]. After four weeks there was a small, but significant increase in the Minimum Erythema Dose (MED). Serum triglyceride levels had decreased by 40 mg/dl. Rhodes et al. [54] undertook a second study a second study to examine the effect of omega-3 FA supplementation on UVR-induced erythema and lipid peroxidation.
The latter study employed a 3g/day of mixed omega-3 FA that was administered over 3-6 months. The MED to UVB rose progressively with increasing time of supplementation and had more than doubled in month six. The increase in MED was accompanied by an increase in epidermal omega-3 composition and increased susceptibility to lipid peroxidation. The increased MED returned to baseline 2 ½ months after cessation of supplementation. A double-blind randomized study, in which participants received 4 g/daily EPA or oleic acid (OA) for three months, examined early genotoxic markers [55]. The skin content of EPA showed an 8-fold rise from baseline; sunburn sensitivity was reduced in the EPA supplemented group, rising from a UVR-induced erythemal threshold from 36 mJ/cm2 at baseline to 49 mJ/cm2 at 3 months; UVR-induced skin p53 fell by half at 24 hour post-exposure to UVR; and susceptibility of peripheral blood lymphocytes to ex vivo UVR exhibited a 24% reduction of tail moment (DNA strand breaks in the Comet assay) in the EPA supplemented group; No significant changes in any of the parameters were observed in the Oleic acid supplemented group. Nor were there consistent alterations in basal and UVR-exposed skin in content of the antioxidants, glutathione, vitamins E and C, or lipid peroxidation (contrary to the previous study) in the EPA supplemented group. Protection to these UVR-induced genotoxic markers by EPA suggests that longer term supplementation might reduce skin cancer in humans.
Aside from these early genotoxic markers, several studies have examined eicosanoid intermediates and the influence of omega-3 FA on inflammatory PG and cytokines. Pupe et. al. [56] examined the effect of EPA, a precursor of eicosanoid biosynthesis, on the UVR response of normal human keratinocytes, as a prerequisite to investigate the FA’s immune modulating action on the UVR response in skin. EPA treatment and UVR exposure were both shown to produce higher levels of tumor necrosis factor-α (TNF-α), and IL-1α expression. EPA treatment resulted in lipid peroxidation and in decreased PGE2 and IL-6 secretion after UVR exposure. Oleic acid treatment produced none of these responses in non-UVR-exposed keratinocytes. Shahbakhti et al, [57], in a double-blind randomized study, supplemented human participants with 4 g/daily of ethyl ester of EPA or oleic acid for three months. Skin biopsies and suction blister fluid, as well as plasma, were taken from unexposed and UVR-exposed precipitants and assayed for PGE2 and pro-inflammatory cytokines, Interleukin-1ß, TNF-α, Interleukin-6, and Interleukin-8. There was, again, a significant rise in the MED in EPA supplemented participants. At baseline, and before supplementation, 3X Med UVR increased blister fluid TNF-α, IL-6, IL-8 and PGE2 at 16 hours post-exposure. No significant changes occurred in baseline or in UVR-induced skin levels in cytokines after supplementation with either EPA or OA, although UVR-induction of PGE2 was abolished after EPA supplementation. Nor were there changes in plasma cytokine and PGE2 levels. Thus, there was no evidence that reduction in the sunburn response by EPA was mediated by the pro-inflammatory cytokines examined, although the abrogation of UVR-generated PGE2 certainly appears to play a role. Whereas there was no systemic effect of EPA on TNF-α or IL-8, when fibroblast and keratinocyte cell lines were supplemented with EPA, DHA, or OA, EPA and DHA were shown to reduce basal secretion of IL-8 in keratinocytes by 66% and 63%, respectively, with a similar pattern in occurring in fibroblasts [58]. UVR-induced levels were reduced by ~65% 48 hours after exposure. Further, secretion by keratinocytes was reduced by 54% and 42% by EPA and DHA, respectively. In summary, both EPA and DHA inhibit the production of TNF-α-induced IL-8 in skin cells and may play a role in the photoprotective, anti-inflammatory, and immunoresponsive effects manifested by these FA. In fact, a small double-blind, randomized intervention trial examined the impact of oral omega-3 FA on UVR suppression of cell-mediated immunity that was assessed through the nickel contact hypersensitivity (CHS) response [59]. The study involved 79 female volunteers who were nickel allergic (only ~15%of women in the general population exhibit CHS to this metal). Volunteers took 5 g/day of mixed omega-3 FA or consumed a control lipid (glyceryl tricoprylate, a medium-chain triglyceride that contains 4 fatty acids (caproic, caprylic, capric, and lauric acids in a 2:55:42:1 ratio), for three months. After supplementation, nickel was applied to three pre-exposed sites on three consecutive days, to UVR (solar simulated radiation, SSR) and to unexposed control sites, Nickel CHS responses were quantified 72 hours later and SSR-induced immunosuppression calculated. SSR dose-related suppression of nickel CHS response was observed in both supplemented groups. Although not statistically significant, a UVR dose-dependent increase in the percentage of photo-immune suppression of the nickel CHS response, that reached 43%, was observed at the highest SSR dose used. Immunosuppression was 50% lower after active supplementation compared with the control at an SSR dose of 3.8 J/cm2, whereas immunosuppression induced by the highest SSR dose (7.6 J/cm2) showed little influence of omega-3 FA. Because the dose-response for some protective effects of omega-3 FA are unknown, higher supplemental doses remain to be explored, but it should be noted that RBC omega-3 FA was four times higher in the supplemented group compared to control. Overall, these data provide further evidence that increased oral omega-3 FA intake could provide protection from the cutaneous immunosuppression that occurs during short exposures to solar UVR.
Observational studies have failed to provide clear evidence that dietary omega-3 FA reduce the risk for NMSC, albeit direct evidence from experimental, and promising evidence from cell culture and clinical studies make it important that the potential of omega-3 FA as a preventive nutrient to NMSC be fully explored. The direct way to address this issue is through intervention trials with populations with high, and known, risk for NMSC. It has been proposed that a study design, such as the low-fat intervention trial occurrence in NMSC patients [47,60]. Certain caveats must be considered, i.e., unlike the animal studies in which omega-6 and omega-3 FA were the sole source of dietary fat, this will not be the case in human studies. Thus, the relative omega-6/omega-3 FA ratios will determine response, and careful monitoring is essential as it has been shown that SCC risk is increased when AA level of RBC membranes is highest [61]. AA is the precursor for LA, an omega-6 FA. Diets must be carefully monitored to ensure that any potential benefit of omega-3 FA supplementation is not diminished by increased omega-6 FA intake. Dietary assessment and protocol compliance should be indexed to omega=6/omega-3 ratios that can be determined by RBC membrane composition, although the omega-3 index is recommended as a better metric [62]. One further caveat that may make such an intervention trial problematic is that experimental data indicate that the effect of omega-3 FA on UVR-carcinogenesis occurs in the initiation stage and throughout the carcinogenic continuum. Omega-3 FA suppress the UVR-mediated immune suppression that allows tumor outgrowth. Thus, although such a study has not been undertaken, an intervention may influence the time of response, suggesting that the follow-up period of such a study may have to be extended.
As noted, observational studies have failed to provide clear evidence that dietary omega-3 FA reduce the risk for NMSC. A population-based case-control study showed a consistent tendency for a lower risk of SCC with higher intakes of omega-3 FA [62]. Further, there was a tendency toward decreased risk of SCC with increased intake of diets with high omega-3/omega-6 FA ratios. A prospective study examined plasma omega-3 and omega-6 FA levels and risk of BCC and SCC [63]. Plasma EPA concentrations and omega-3/omega-6 ratios showed a significant inverse association with SCC occurrence and these associations were stronger among participants with a prior history of skin cancer. Total omega-6 FA was also inversely associated with BCC when analyzed in a multivariate model. Confoundingly, both LA and ALA FA were also inversely associated with BCC occurrence, although when these FA were analyzed as continuous variables, there was no evidence of linear or non-linear associations. Although this study provides support for reduced NMSC occurrence with high concentrations of omega-3 FA, the results depended on how the data were analytically modelled. Another population-based prospective study by this group of investigators found that intake of AA was associated with increased SCC incidence whereas total omega-6 FA was associated with reduced BCC [64]. Overall, their data did not support an association of these FA and BCC or SCC risk. On the other hand, in a hospital-based case-control study, the FA composition of RBC in newly diagnosed BCC patients found that LA and AA were significantly higher in BCC patients [61]. Omega-3 FA was significantly lower and omega-6 FA was significantly higher in BCC patients than in controls. Total PUFA and omega-6/omega-3 PUFA ratios were significantly higher in BCC patients. A mendelian Randomization analysis [66] found that an increase in genetically predicted levels of LA and ALA was associated with a reduced BCC risk – a result in agreement with that of reference [63]. Higher levels of AA and EPA were associated with an increase in BCC risk. A two-sample Mendelian carcinoma study found a direct link between the susceptibility of BCC and PUFA and their degree of unsaturation [67]. It was posited that modifying one’s diet, specifically focusing on the intake of PUFAs, and metabolic profiling for these parameters, may be an effective strategy for preventing BCC. Associations between fat intake and risk of SSC, BCC, and malignant melanoma (MM) were also examined within two prospective studies – the Nurses’ Health Study (NHS) and the Health Professionals Follow-up Study (HPFS) [68]. Higher PUFA intake was associated with the risk of SCC and BCC. Higher omega-6 FA was associated with higher risk of BCC, SCC, and MM. Omega-3 FA was associated with risk of BCC, but not SCC and MM. The mixed results of these studies illustrate the need for a direct dietary intervention trial.
Organ transplant recipients are at a very high risk of developing skin cancer, compared to the general population. The feasibility of omega-3 FA supplementation to reduce skin cancer among lung transplant recipients was investigated from the outpatient clinic of an Australian hospital [69]. Participants were randomly assigned to receive either mixed omega-3 FA or olive oil (control) for 12 months. After 12 months plasma EPA levels increased substantially in the intervention group, six patients developed NMSC compared to 11 in the control group. A second study of kidney or liver transplant recipients were evaluated via a food frequency questionnaire and the PUFA intakes were ranked as low, medium, or high [70]. Transplant recipients with a high total long chain omega-3 FA, compared to low intakes exhibited substantially lower SCC risk and those with high ALA intakes exhibited significantly fewer BCC. Thus, high intakes of long-chain omega-3 FA and ALA may reduce risks of SCC and BCC, respectively.
Finally, an experimental study with omega-3 FA is described here as it potentially has clinical relevance in PUVA therapy [71]. 8-methoxypsoralen (8-MOP) and UVA (PUVA) therapy has. been shown to be carcinogenic. Animals received dietary omega-3 FA or omega-6 FA sources. After two weeks, the animals were treated topically with 8-MOP and then exposed to UVA. Mice receiving the omega-3 FA source exhibited a marked decrease in inflammatory response and more rapid repair. The effects on PUVA tumorigenesis were examined in long-term studies in which animals were treated topically 3X/week with 8-MOP, after which they were exposed to UVA. Omega-3 FA supplementation markedly ameliorated the course of PUVA toxicity but did not impede the course of PUVA tumorigenesis. In this regard, it is known that experimental PUVA tumorigenesis does not exhibit time-dose reciprocity, i.e., cumulative dose, per se, is not an accurate index of tumorigenic risk. In fact, this has been suggested as an explanation for differences between European and American studies of the risk associated with PUVA therapy [72]. Thus, taking advantage of the effect of omega-3 FA on reduction of PUVA toxicity, omega-3 FA supplementation might be used in conjunction with PUVA therapy to allow a more aggressive PUVA therapy in the American protocol while reducing the risk skin cancer from PUVA therapy alone.
Conclusions from clinical studies (human and human cultured cells may be summarized as follows:
- A direct link exists between PUFA intake and degree of unsaturation and susceptibility to BCC.
- Omega-3 FA supplementation significantly increases the erythema threshold to UVR.
- Omega-3 FA modulate a number of cytokines (in human cells in vitro) and eicosanoids that mediate inflammatory and immune responses.
- Omega-3 FA inhibit certain genotoxic markers of UVR-induced DNA damage, e.g., UVR- induced cutaneous p53.
- Omega-3 FA abrogate UVR-induced immunosuppression of cell mediated immunity assessed as nickel CHS
- Omega-3 FA reduce the risk of SCC and BCC in organ transplant patients.
3. Antioxidants
3.1. Experimental
3.1.1 Phenolic Compounds
A dietary cocktail of antioxidants was shown to play a pivotal role in both inhibition of the primary event(s) involved in UVR mediated skin tumor induction and the subsequent development of precancerous lesions into tumors [73]. Not only was tumor occurrence dramatically reduced, but the severity of those tumors that did develop were of markedly less severity than those of non-supplemented controls. The antioxidant cocktail consisted of 2% (w/w) of the diet and was composed of 1.2% ascorbic acid, 0.5% butylated hydroxytoluene (BHT), 0.2% DL-Alpha tocopherol, and 0.1% glutathione. This antioxidant mixture was also examined in regard to their effects, with respect to unsaturated lipid level, upon lipid peroxidation and UVR carcinogenesis [74]. Antioxidant supplementation produced an inhibitory effect, both in regard to tumor latency and multiplicity, almost equal to the degree of exacerbation of carcinogenesis evoked by increasing unsaturated FA levels. The antioxidants dramatically inhibited lipid peroxidative reactions. These data strongly suggested that free radical reactions, specifically lipid peroxidation, play a part in the UVR-carcinogenic response. Subsequently, it was shown that BHT was the active principle in the antioxidant cocktail [75]. Vitamins C and E and glutathione were without effect at the levels in the cocktail [75,76]. BHT supplementation, alone, provided the level of photoprotection as that of the cocktail.
De Rios et. al., [77] reported that BHT provided marked systemic protection against UVR-mediated erythema in hairless mice. BHT also provided diminished, but statistically significant, photoprotection when applied topically. Vitamins C and E, nor glutathione, provided negligible protection when administered at the concentrations of the Cocktail. Not only does BHT supplementation result in a 2-fold increase in the level of UVR required to produce a MED, but it also markedly inhibited UVR-induction of epidermal ornithine decarboxylase (ODC) [78]. ODC induction, whereas lacking specificity for tumor promotion, is an early event closely associated with the process and was thought to be a necessary component of skin tumor formation. The latter is supported by the fact that BHT did not have a similar result in inhibition of 12-0-tetradecanolyphorbol-13-acetate (TPA)-induced ODC. Data from this study suggested that dietary BHT inhibition of ODC induction was a response related directly to the degree of UVR insult rather than a general effect upon the processes associated with carcinogenic promotion. These studies suggested that BHT might act through diminution of UVR reaching critical epidermal target sites. Thus, efforts were undertaken to understand the mechanism(s) of the BHT-mediated UVR photoprotection. It was shown that BHT supplementation resulted in increased epidermal spectral absorption [79]. As the absorption maximum of BHT lies within the VR region of the electromagnetic spectrum, its protective effect of photic energy against erythema and carcinogenesis might result from effective dose of UVR reaching respective target sites. It had been shown that BHT is rapidly metabolized in the rodent and that minimal levels occur in skin [80,81]. From forward scattering scans, obtained over a range of 250-400 nm and based upon BHT concentrations in the epidermis, a dose reduction factor (DRF) of 2 for BHT was calculated -in agreement with the protective effect of BHT on erythema and of a magnitude that could significantly prolong the onset of UVR-induced skin cancer. Nevertheless, the data indicated that the maximum levels of BHT accruing skin would only absorb about 1% of the incident UVR, suggesting that BHT produces its effect indirectly. Additional studies were undertaken to determine whether the photoprotective properties of BHT to skin might result from the alteration of epidermal parameters that could account for the DRF [82]. None of the major anabolic parameters examined, i.e., there was no indication that BHT elevated DNA synthesis; BHT did not affect radiolabeled leucine incorporation (protein synthesis0 in UV-irradiated epidermis; there were no effects of BHT on cell labeling index; and no significant change in epidermal layers. It was suggested that catabolic processes, normal oxidative events leading to keratin stabilization and altering the optical properties of the stratum corneum (SC), might be involved. To explore the contribution SC might contribute to BHT’s photoprotection, SC was examined from animals receiving the BHT supplement and non-supplemented controls [83]. Spectral transmission from both groups revealed that SC transmission, recorded between 280-320 nm, was approximately 65% greater in controls compared to that from BHT supplemented animals. Further, when the SC was stripped and the animals irradiated, BHT provided the usual inhibition of ODC induction in non-stripped animals, but ODC activity in BHT-treated, tape stripped animals was restored to levels that did not significantly differ from controls. This protective effect of SC in BHT supplemented animals could not be attributed to BHT-induced change of physical dimensions, as neither the thickness of the SC nor the number of SC layers differed from controls. BHT might alter the optical properties of SC and not alter the physical dimensions by eliciting its photoprotective properties via its antioxidant properties. It is possible that during normal SC maturation (oxidation) sulfhydryl moieties are protected from oxidation, resulting in fewer disulfide cross-links. The degree of cross-linking is known to alter x-ray diffraction patterns of keratin [84} and it would be expected that optical properties would be altered as well. Oxidation of two sulfhydryl groups to a disulfide bond in the stabilization of keratin generates a molecule of water and increased sensitivity of hydrated skin to sunlight is well documented. The degree of SC hydration markedly affects its transmission characteristics [85,86]. In summary, it has been demonstrated that a major component of BHT’s protective effect resides in the SC and that the changes affected by this antioxidant appear to be related to chemical properties of SC rather than changes in physical parameters.
BHT is but one compound of a class of organic chemicals, the phenols, that demonstrate antioxidant properties. On the basis of structural characteristics, the phenolic compounds butylated hydroxyanisole (BHA), propyl gallate, and vanillin were compared to BHT for anti-.UVR-induced carcinogenesis [87]. Of the four phenols tested, only BHT conveyed significant inhibition of both tumor latency and tumor multiplicity, increasing tumor latency 5.2 weeks and reducing multiplicity by 60%. Vanillin reduced tumor multiplicity by 48% but had no significant effect on tumor latency. BHA and propyl gallate were without effect. ODC was assayed to determine whether it might serve as a rapid screen for anti-carcinogenic phenols. Both BHT and BHA significantly inhibited ODC induction whereas propyl gallate and vanillin were without effect. The conclusion drawn from these data is that neither antioxidant or lipophilic properties, nor the tert-butyl moiety common to both BHT and BHA are sufficient determinants (although BHT has two tert-butyl moieties that may account for its distribution to skin) of anti-UVR-induced carcinogenesis. Nor is ODC induction an indicator of such activity.
BHT is widely used as a food preservative and in the cosmetic industry. The Food and Drug Agency (FDA) has classified BHT as “generally recognized as safe” (GRAS) when used in an appropriate manner. The International Agency for Research on Cancer (IARC) has evaluated the literature on BHT’s potential role in carcinogenicity and found no clear evidence that BHT affects carcinogenicity in animals and there were no data for humans [89]. Williams et al., [89] reviewed experimental studies on carcinogenesis in regard to cancer hazard assessment of exposure to humans and concluded that BHT did not pose a cancer hazard and that it may be anti-carcinogenic at current levels of food additive use. Due to the low concentration found in cosmetic formulations, it was concluded that BHT is safe as used in the US [90]. At the low concentrations used in cosmetic products BHT is poorly absorbed through the skin with most remaining on the surface, thus, producing no systemic effects. However, there were two early reports that raised concern. The first by Olsen et. al. [91] that reported on hepatocellular neoplasms in rats induced by BHT. The second reported on the enhancement of lung tumor formation in mice by butylated hydroxytoluene: dose-time relationships and cell kinetics [92]. A number of studies have reported BHT-induced carcinogenicity at different organ sites and have been evaluated in the IARC monograph [88].
When any chemical is introduced into a biological system, physiological effects occur. BHT induces hepatomegaly in the hairless mice, although there appears to be no pathology associated with this condition (e.g., fatty liver etc., but note ref. 91) [93]. Accompanying the induced hepatomegaly, induction of hepatic Phase I and II microsomal detoxification enzymes (mixed function oxidases) occurred. The enzymes influence the mutagenic frequency of a range of chemicals. Thus, a modified Ames test was employed to assess the influence of the antioxidant supplement on activation of N-2-fluorenylacetamide (2-FAA) that is a potent hepatocarcinogen [94]. Animals were maintained on their respective diet supplemented with the antioxidant cocktail (of which BHT was the active component) for 38 weeks, administered the 2-FAA and one week later a microsomal fraction (S-9) was prepared and used to test for activation in the Ames mutagenicity test. Antioxidant significantly increased the mutation frequency (3-fold). These data suggested that beneficial modification of BHT to UVR-carcinogenesis may actually predispose an individual to another carcinogenic agent. Consequently, it may be necessary to develop an algorithm for each antioxidant supplement based upon the benefit to be derived for NMSC and the potential risks to each individual to other forms of cancer [13].
Conclusions from Experimental Studies:
- BHT dramatically inhibits UVR-carcinogenesis, both in regard to tumor latency period and tumor multiplicity and reduces the severity of those tumors that do develop.
- Inhibition of carcinogenesis occurs through a mechanism of dose diminution by altering the chemical characteristics of the stratum corneum.
- BHT provides marked systemic protection against UVR-mediated erythema in hairless mice. BHT also provides statistically significant protection when administered topically.
- BHT inhibits the UVR induction of ODC
- Several structurally related phenols were evaluated for anti-UVR-carcinogenesis, but only BHT conveyed significant inhibition of both tumor latency and tumor multiplicity.
- BHT supplementation may predispose the host to chemical carcinogenesis by inducing hepatic phase I and II microsomal detoxification/activation enzymes.
- Clinical Relevance:
- Human data is scarce and assessments of BHT and human risk have found that BHT levels in food stuff pose no risk and may even lower the carcinogenic risk. There appears to be no risk involved in the levels of BHT used in cosmetics.
- However, in view of experimental studies of BHT’s promotion or induction of carcinogenesis at various organ sites and the capacity to induce activation of pro-carcinogens, and potential endocrine disrupters, the intake of BHT should be approached with caution [95].
3.1.2. Polyphenols
Polyphenols are a diverse group of approximately 8,000 phytochemicals that exhibit antioxidant properties. They are generally classified as Flavonoids, that constitute ~60% of polyphenols; phenolic acids; lignans; stilbenes; and some miscellaneous polyphenols [96]. Polyphenols are strong antioxidants, neutralizing radical reactions by donating a hydrogen atom or electron. Thus, polyphenols suppress the generation of free radicals by inhibiting the active free radical species or deactivating the precursors of free radicals. They also act as direct free radical scavengers of lipid peroxidation by acting as chair-breakers of the peroxidation chain reactions.
- (1)
- Anthocyanins
Anthocyanins are widely dispersed in the Plant Kingdom and are responsible for the red, blue, and purple colors of fruits and vegetables. They are usually glycosylated with sugars and identified as cyanidin, delphinidin, peonidin, pelargonidin and malvidin [97]. Potential antitumor effects are reported to be based on a wide variety of biological activities including antioxidant; anti-inflammation; anti-mutagenesis; induction of differentiation; inhibiting proliferation by modulating signal transduction pathways, inducing cell cycle arrest and stimulating apoptosis or autophagy of cancer cells; anti-invasion; and anti-metastasis [98]. The treatment of highly malignant melanoma B16-F-10 cells with berry extracts containing anthocyanins reduced cell proliferation by 30-38%, compared to controls [99], Mulberry anthocyanins reduced the levels of Ras, P13K, phosphor-Akt, and NF-ĸß in B16-F-1 melanoma cells and showed that the anthocyanins inhibited the metastasis of these cells in vivo [100]. The photoprotective effects of delphinidin were demonstrated in human HaCaT keratinocytes and the skin of SKH-1 mice [101]. The anthocyanin inhibited UVR-mediated lipid peroxidation; protected against a decrease in cell viability; induction of apoptosis; a decrease in proliferating cell nuclear antigen expression; an increase in 8-OHdG, an increase in poly (ADP-ribose) polymerase; and a number of other markers. Topical application to SKH-1 mouse skin inhibited UVR-mediated apoptosis and markers of DNA damage, suggesting that the anthocyanin inhibited UVR-mediated oxidative stress, reduced DNA damage, and thus protected the cells from UVR-induced apoptosis. After an extensive review of the use of flavonoids in skin cancer prevention and treatment, it has been proposed that these agents can play a beneficial role as systemic chemopreventives to UVR-mediated skin cancer and topically as adjuvants to sunscreens and cosmetic products as anti-cancer therapy [102].
- (2)
- Phenolic acids:
Epigallocatechin gallate (EGCG) is a major catechin in green tea (GT) and is a powerful antioxidant and anti-tumor agent. Several proteins have been identified as direct interactors with EGCG and that are involved in cell proliferation, survival, and metastatic spread.[103]. Inhibitory effects of green tea catechins are the result from the targeting of genes and proteins on Ras/extracellular signal regulated kinases MAPK and P13K/Akt. These are receptor tyrosine kinase-related downstream pathways that are constitutively activated in tumor cells. Studies of chemically induced carcinogenesis have shown that green tea consumption protects against a number of cancers, including skin [104]. Flavonoids are effective immunomodulators as they modulate cell growth, induce apoptosis, and reduce ROS production, thus suggesting their benefit in skin cancer prevention and treatment [105106]. Green tea, when fed in drinking water of mice, afforded significant protection against UVR-induced skin carcinogenesis [104]. GT catechins and its derivatives also inhibited P450-dependent mixed function oxidases in skin, thus exhibiting antimutagenic effects and showed substantial anti-tumorigenesis effects in bioassay systems [106].
Inhibition of UVR-carcinogenesis by GT is mediated, partly, through the rapid repair of DNA that, in turn, is mediated through the induction of IL-12 [107,108]. EGCG was found to be most effective in protecting against UVR-induced suppression of CHS. Topical treatment inhibited the induction of inflammation, probably through reduction of PGE2. On the other hand, Gensler et. al. [109] reported that systemic administration of EGCG, at concentrations employed, did not reduce occurrence of UVR-induced skin cancer, but topical application reduced the incidence significantly (62%) through a mechanism distinct from inhibition of photo-immunosuppression.
Epidemiological and clinical studies have reported varying responses of green tea and cancer incidence with some studies showing inverse relationships and some reporting increased risks, depending upon cancer site [110]. Other flavonoids and phenolic acids found in green tea extracts that show anti-cancer properties include quercetin [111], ellagic [112], and gallic acids [109].
Resveratrol is a Stilbene, with antioxidant properties that is derived primarily from red grape skin and seed concentrate, that has shown promise as an anti-cancer agent [113]. It affects all carcinogenic stages, i.e., initiation, promotion, and progression, as well as induced apoptosis [114]. Dietary supplementation of Resveratrol is target-organ specific with respect to its anti-carcinogenic effect and clinical trials have generally been ambiguous and some even detrimental [114]. Resveratrol imparts strong chemopreventive effects against UVR exposure-mediated skin carcinogenesis [115]. The inhibition of carcinogenesis occurred whether the skin was pre-or post-treated with Resveratrol. The chemopreventive effects of stilbene may, at least in part, be mediated via modulations in Survivin (an anti-apoptotic protein), and other associated events, e.g., interactions with phase I and II enzymes both in vitro and in vivo. These interactions may be beneficial or harmful. Indeed, individuals taking drugs whose efficacy is highly specific and cytochrome P450 enzyme-dependent, could be particularly affected, as well as those individuals exposed to environmental carcinogens. Therefore, caution should be taken when using supplemental resveratrol doses for health benefits, such as chemoprevention [116].
Curcumin, a polyphenol found in turmeric, has been shown to exhibit anti-cancer properties in both preclinical and clinical studies [117]. Human clinical trials found no toxicity when administered at doses to 10g/day. Curcumin exhibits anti-cancer potential by suppressing proliferation of tumor cells; down-regulation of transcription factors, e.g., NF-kappa ß; down-regulation of expression of COX- 2, LOX, NOS (and is thus, a potent anti-inflammatory); cell surface adhesion molecules; and protein tyrosine and threonine/threonine kinases. Evidence exists that curcumin suppresses the initiation, promotion, and metastatic stages of carcinogenesis. Topical application of curcumin caused delay in tumor appearance, multiplicity, and size of tumors that did develop in UVR-irradiated hairless mice [118]. The protective effect was accompanied by downregulation of cell proliferative controls, apoptosis, transcription factor NF-kß, and inflammatory responses. Similar results were obtained when curcumin was administered systemically [119]. Curcumin inhibited UVR-carcinogenesis whether administered systemically or topically.
Gallic acid, a predominant polyphenol, has been shown to inhibit two-stage chemically induced carcinogenesis in skin [120] and inhibits SCC through targeting heat shock protein HSP90AB1, the most efficient target to repress metastasis and invasion [121]. Gallic acid has also been shown to inhibit carcinogenesis in vitro in cancerous cell lines [122]. The inhibitory effect on cancer cell growth is mediated via the modulation of genes which encode for cell cycle, metastasis, angiogenesis and apoptosis. Gallic acid also inhibits the activation of NF-κB and Akt signaling pathways along with the activity of COX, ribonucleotide reductase and GSH. Moreover, gallic acid activates ATM kinase signaling pathways to prevent the processes of carcinogenesis.
Ferulic acid (FA), found in coffee and cereal grains, down regulates NF-kß, COX-2, and VEGF expression in 7,12-dimethylbenzen(a)anthracene-induced oral (hamster buccal pouch} carcinogenesis [123]. Administration of FA to two osteosarcoma cell lines resulted in inhibition of cell proliferation and induced apoptosis [124]. Importantly, FA inhibited P13K/Akt activation. When the cells were transfected with adenoviruses expressing active Pkt, the anti-proliferative and pro-apoptosis of FA were reverted, suggesting that FA inhibits cell proliferation and induces apoptosis via inhibiting the P13K/Akt pathway. Human keratinocyte HaCaT cells were exposed to UVR and FA. Ferulic acid inhibited UVR-induced cytotoxicity, apoptosis, and cyclobutene pyrimidine dimer formation.[125]. FA also attenuated the mRNA levels of apoptosis -regulatory genes (p53-p21 and c-fos), the protein levels of p53, and the secretion of cytokines IL-6 and TNF-α. Proliferating cell nuclear antigen and replication protein A genes were also blocked. When Swiss Mice were chronically exposed to UVR, after FA was intraperitoneally or topically administered, significant reduced incidence of tumor volume and weight resulted [126]. FA treatment inhibited UVR-induced oxidative damage.
It is interesting to note that Homer, in his Illiad, recorded, as early as 700 B.C., the use of a soothing, healing “Ambrosia” to warriors wounded in battle. [127]. Although the compounders of this “ambrosia” were secretive as to its composition, it is almost certain that it was constituted from polyphenol, camphor, menthol (from mint plants) and olive oil (rich in polyphenols) containing plant preparations. Indeed, the bark extract of Pinus Maritima (PBE), from that region and rich in polyphenols, phenolic acids, and a mixture of flavonoids, was examined for skin cancer preventive activity that was evaluated in two different experimental animal tumor models induced by UVR and combination of UVR with 7,12-dimethylbenz[a] anthracene [128]. Animals ingested the PBE in drinking water. PBE exhibited potent NMSC preventive, anticancer activity. The chemopreventive action was demonstrated by the late appearance of the first skin papilloma/tumors, the reduced number of skin tumors per animal, the reduced percentage of animals bearing skin tumors and the prolongation of animal viability.
- (3)
- Vitamins C and E:
Nobel Laurate, Linus Pauling, popularized vitamin C supplementation with publication of his international best-selling book, Vitamin C and the Common Cold. The Linus Pauling Institute has concluded that, thus far, there are insufficient data to suggest a link between vitamin C status and the risk of developing a given type of cancer [129].
Pauling et al. [77] examined vitamins C and E at levels found in the cocktail mix [73] and did not see significant inhibition of UVR-induced skin tumors. However, when L- ascorbic acid was supplemented at very high dose levels (up to 10% of the diet (w/w))/day), statistically significant decreases in the incidence of dermal neoplasms (papilloma and SCC) occurred and there was a delay in the onset of malignant lesions [130].
Vitamin E has been shown to reduce the incidence of UVR-induced skin cancer in mice [131]. Supplementation with 100 or 200 IU of dl-alpha-tocopheryl acetate/ kg diet led to 46% and 19% reduction in tumor incidence in UVR exposed mice, respectively. Reason for the lower rate of tumor incidence in the 200 IU supplemented animals was that 40% of the animals died after 31 weeks post the first UVR treatment. Thus, oral supplementation of vitamin E inhibited photocarcinogenesis, but at doses that were toxic. It was also shown that topical application of vitamin E (dl-α-tocopherol) reduces UVR -induced cancer formation and an immunosuppression that prevents the host from rejecting antigenic UVR-induced tumors [132]. Topical application of thermostable esters of vitamin E, i.e., alpha-tocopheryl acetate and alpha-tocopheryl succinate did not inhibit photocarcinogenesis and actually enhanced carcinogenesis [133]. Nor did these esters prevent the UVR-induction of immune susceptibility. Burke [134,135] reports that when high concentrations of the nonesterified, optimal isomer, of vitamins C and E are applied topically, significant inhibition of acute erythema and tanning occur, as well as chronic UVR photoaging and skin cancer. Such an approach could result in more effective and beneficial cosmeceutical formulations.
Although the utilization of vitamin C as a form of anti-cancer therapy has been advanced, preclinical studies have resulted in contradictory results as to the rationale for such an approach [136]. However, in a small clinical trial, intravenous infusions of 75 g vitamin C, 3X weekly increased serum Vitamin C from micromolar to millimolar levels and increased overall survival from 8.3 months to 16 months when added to gemcitabine + nab-paclitaxel chemotherapy regimen in stage IV pancreatic cancer patients [137]. Infusion of Vitamin C prolonged overall and progression free survival without detriment to quality of life or added toxicity (Clinical Trials.gov No. NCT02905578).Miscellaneous antioxidants has shown that
Glutathione is a major endogenous antioxidant. N-Acetylcysteine (NAC) is a stable form of l-cysteine that is necessary to produce glutathione. NAC has been reported to protect melanocytes against oxidative stress damage and to delay the onset of UVR-induced melanoma in mice [138]. However, NAC supplementation has been shown to increase melanoma metastasis in experimental animals [139]. a consequence of the differential role that ROS plays in the early and late stages of cancer [140]. NAC, alone, or in combination with Vitamin C, has been reported to modulate UVR-induced skin tumors in hairless mice [141].
Elevated oxidative stress (OS), and mitochondrial dysfunction, have been identified as key hallmarks of aging. Prior studies demonstrated that deficiency of the intracellular antioxidant glutathione (GSH) could play a role and reported that supplementing GlyNAC (combination of glycine and N-acetylcysteine [NAC]) in aged mice improved GSH deficiency, OS, mitochondrial fatty-acid oxidation (MFO), and insulin resistance (IR) [142]. GlyNAC supplementation in mice increased the length of life by correcting glutathione deficiency, oxidative stress, mitochondrial dysfunction, abnormalities in mitophagy and nutrient sensing, and genomic damage. To test whether GlyNAC supplementation in aged adults could improve GSH deficiency, OS, mitochondrial dysfunction, IR, physical function, and aging hallmarks, a placebo-controlled randomized clinical trial demonstrated that when GlyNAC was provided to T2DM patients, GlyNAC improved mitochondrial dysfunction, insulin resistance, improved glutathione synthesis, and lowered oxidative stress. [143]. GlyNAC approaches the requirements for the catalytic antioxidant posited by Brownlee [144]. It would endogenously regenerate glutathione and restore and maintain the cell’s redox balance.
As UVR is an accumulative carcinogenic agent, NMSC occurs later in life. About 95% of BCC occurs after the age of 40 and 75-80% of SCC occurs in men older than 60 years of age [145]. Thus, NMSC exhibits a long latent period and is generally a disorder of the aged. UVR carcinogenesis shares many of the pathophysiologic markers of aging, particularly that of OS. If GlyNAC is effective in diminishing aging markers, and if it circumvents the detrimental effects of NAC on melanoma, it should also reduce the risk of NMSC. It may be a promising avenue for future research.
Carbon 60 (C60), or Buckminsterfullerene, or sometimes referred to as a “Bucky ball” because of its resemblance to a soccer ball, exhibits high stability, high reactivity, and is known as a powerful antioxidant [146]. Using a pig skin model, it was demonstrated that C60 dispersions in transcutol/isopropyl myristate were suitable for transdermal delivery [147]. Basic cosmetic formulations with transcutol/isopropyl myristate, without harsh organic solvents, show a high potential for delivery of C60 for biopharmaceutical and cosmetics applications [148]. Polyvinylpyrrolidone entrapped C60 provided prominent cytoprotective effects against intermittently repeated UVA radiation induced injuries and suppressed intracellular superoxide radicals in human keratinocytes [149]. There was also suppression of apoptosis (does it boost metastasis?). Growth experiments with water-soluble C60 fullerenes were performed on groups of mice and was found to efficiently inhibit the growth of transplanted malignant tumors [150]. As with other antioxidants, C60 exhibits a dual ability, it can act as a radical scavenger or as an oxygen radical generator and must be administered in a manner that achieves the beneficial result. However, as more safety and therapeutic data is accrued, it has the potential to play a significant role in dermatology.
As noted earlier, a number of clinical trials, primarily observational studies of case-control designs, have reported lower antioxidant levels being inversely associated with cancer risk. These include skin, However, when intervention trials (the gold standard) reported adverse effects of antioxidant therapy with the same antioxidants, the safety and effectiveness of antioxidant supplementation was questioned. A review and meta-analysis reported a significant (16%) increase in the all-cause mortality of trial participants receiving ß-carotene and vitamins A and C treatments [151]. An adverse side effect of high vitamin C supplement intake has been associated with an increased risk of cardiovascular disease (CVD) mortality in diabetic postmenopausal women [152]. A water-soluble analog of vitamin E, Trolox, has been shown to increase melanoma metastasis in experimental animals [137]. A small clinical study, in which subjects were supplemented with vitamin E found that the antioxidant significantly reduced skin malondialdehyde levels but did not affect other measures of oxidative stress in human skin [153]. Antioxidant supplementation was reported to negate some of the health-promoting effects of physical exercise [154]. When vitamins C and E were supplemented to T2DM patients, blocking the exercise-dependent formation of ROS, the health-promoting effects of physical exercise, i.e., by promoting insulin sensitivity and the antioxidant defense, were negated. The disappointing and conflicting results of large doses of vitamins C and E and ß-carotene should not be too surprising Supplementing the highly complex and intricate natural antioxidant defense system with a high level of one antioxidant may alter the stoichiometry of the antioxidant pathways and push the pathway from an antioxidant to a pro-oxidant state or through other pro-oxidant paths [72,155] As a result of these reports on adverse effects of antioxidant supplementation on cancer risk and incidence, the World Cancer Fund/American Institute for Cancer Research withdrew recommendations for dietary antioxidant supplementation as a means for cancer prevention [156].
Reports of ambiguous and adverse effects of antioxidants should not deter efforts to seek endogenously metabolized agents that maintain the redox balance and thus have the potential to prevent cancer. It may require a reassessment of our current methods and the development of new algorithms for safety testing and overall risk/benefit analyses, weigh, , weighing the risk of one form of cancer to other forms and adverse pathophysiological responses [13]. The application of new methods must be developed to determine when antioxidant therapy must cease to avoid antioxidant/ROS inhibition, leading to cancer de-velopment and metastasis. In this regard, a recent report describes a mitochondria targeting nanoparticle that delivers an antioxidant directly to mitochondria, the major source of endogenous ROS [157]. The perfection of these targeting systems hold promise for targeted antioxidant therapy. In addition, the timing of antioxidant therapy is important to the oxidative stress/ROS link to cancer. The reduction of ROS and oxidative stress, however, remains a fertile avenue of investigation to preventing and ameliorating conditions that are linked to skin cancer.
Conclusions from Experimental Studies:
• Photoprotective effects of Anthocyanins have been demonstrated in human HaCaT keratinocyte cells and skin of SKH-1 mice.
• The treatment of highly malignant B16-F-10 melanoma cells with berry extracts (containing anthocyanins) reduced cell proliferation by one-third and inhibited the me-tastasis of these cells in vivo.
• Among the polyphenols, anthocyanins have shown several anti-tumor effects, e.g., antioxidant, anti-inflammatory, anti-mutagenic, inhibiting proliferation through modulation of signal transduction pathways, inducing cell cycle arrest and stimu-lating apoptosis, and anti-metastasis.
• Photoprotective effects have been shown for phenolic acids, including Epigallocatechin (EGCG), quercetin, ellagic, and gallic acids, – all strong antioxidants and an-ti-tumor agents and constituents of green tea (GT) catechins.
• ECGC protects against UVR-induced suppression of Contact Hypersensitivity.
• Topical application of EGCG significantly reduced (by two thirds) the incidence of UVR-induced skin cancer.
- effects of Anthocyanins have been demonstrated in human HaCaT keratinocyte cells and skin of SKH-1 mice.
- The treatment of highly malignant B16-F-10 melanoma cells with berry extracts (containing anthocyanins) reduced cell proliferation by one-third and inhibited the metastasis of these cells in vivo.
- Among the polyphenols, anthocyanins have shown several anti-tumor effects, e.g., antioxidant, anti-inflammatory, anti-mutagenic, inhibiting proliferation through modulation of signal transduction pathways, inducing cell cycle arrest and stimulating apoptosis, and anti-metastasis.
- Photoprotective effects have been shown for phenolic acids, including Epigallocatechin (EGCG), quercetin, ellagic, and gallic acids, – all strong antioxidants and anti-tumor agents and constituents of green tea (GT) catechins.
- ECGC protects against UVR-induced suppression of Contact Hypersensitivity.
- Topical application of EGCG significantly reduced (by two thirds) the incidence of UVR-induced skin cancer.
- GT, when fed in drinking water, afforded significant protection against UVR-induced skin carcinogenesis through a mechanism distinct from inhibition of photo-immunosuppression.
- Resveratrol, a stilbene with antioxidant properties, has shown promise as an anti-cancer agent. It affects all carcinogenic stages, i.e., initiation, promotion, progression and induces apoptosis.
- Resveratrol provides strong protective effects against UVR-mediated skin carcinogenesis.
- Curcumin, a polyphenol found in turmeric, inhibits UVR-carcinogenesis whether administered systemically or topically.
- Gallic acid inhibits two-stage chemically induced carcinogenesis in skin and inhibits SCC through targeting heat shock protein that represses metastasis and invasion.
- Ferulic acid inhibits UVR-induced cytotoxicity, apoptosis, and cyclobutene pyrimidine dimer formation in human HaCaT human keratinocytes.
- Systemically or topically administered Ferulic acid to chronically UVR exposed mice resulted in reduced tumor volume and weight.
- Vitamin C, at very high doses, resulted in significant decreases in papilloma and SCC in UVR-radiated mice, as well as a delay in onset of malignant lesions.
- Topical application of vitamin E inhibits UVR-induced skin cancer formation and immunosuppression.
- High concentrations of the nonesterified, optimal isomers of vitamin C and E, applied topically, resulted in significant inhibition of acute erythema and tanning as well as chronic UVR-mediated aging and skin cancer.
- N—Acetylcysteine (NAC), alone, or in combination with vitamin C, modulates UVR-induced skin tumors in mice.
- NAC supplementation increases melanoma metastasis in animals – a consequence of the differential role that OS plays in the early and late stages of cancer.
- Glycine added to NAC (GlyNAC) supplements improved glutathione synthesis and reduced OS.
- Carbon 60 (C60) is a powerful antioxidant (100 to 1000-fold greater antioxidant capacity than other antioxidants), provides protective effects against UVA radiation injuries, suppresses superoxide radical formation, and suppresses apoptosis.
- Clinical relevance:
- Epidemiological and clinical studies of GT polyphenols have reported varying responses with regard to cancer incidence, with some studies showing an inverse relationship and some reporting increased risks.
- Resveratrol has shown promise as an anti-cancer agent. It affects all carcinogenic stages.
- Resveratrol is target-site specific and clinical trials have been ambiguous. Some trials have reported detrimental effects. Caution is advised when supplements are taken for chemoprevention.
- Most observational studies examining vitamin C intake in relation to cancer incidence have found no association.
- Randomized controlled trials have reported no effect of vitamin C supplementation on cancer risk.
- High vitamin C intake has been associated with increased cardiovascular disease mortality in diabetic postmenopausal women.
- High dose, intravenous vitamin C infusions have doubled overall survival of pancreatic patients when used as co-therapy with gemcitabine + nab-paclitaxel.
- Vitamin E has been shown to significantly reduce malondialdehyde levels but does not affect other measures of OS in human skin.
- Conflicting reports of benefits and risks of antioxidant supplementation has led the World Cancer Research Fund/American Institute for Cancer Research to conclude “A general recommendation to consume supplements for cancer prevention might have unexpected adverse effects” and thus, “Dietary supplements are not recommended for cancer prevention.”
The rationale upon which the antioxidant theory of disease is based rests with the assertion that oxidants assault and degrade essential biological molecules that result in aging cancer, and other disorders that can be directly related to oxidant insult [11]. The free radicals, both endogenous and exogenous in origin, that are involved in skin cancer are primarily reactive oxygen species (ROS) that result in Oxidative Stress (OS), and lipid radicals resulting from lipid peroxidative reactions. Antioxidants are chemicals that inhibit oxidative insult. The second tenet of the free radical theory of disease was to add one or more antioxidants to the diet [11,12]. Both experimental and clinical data now suggest that reassessment of this recommendation be tempered by the observations that, under certain conditions, an antioxidant may act as a pro-oxidant that results in a carcinogenic state [28,72].
Topically applied antioxidants may play a beneficial role as adjuvants to sunscreens and cosmeceuticals but suffer from the limitations of topical efficacy.
4. Carotenoids
a. Experimental
Epstein examined the potential influence of intraperitoneally injected β-carotene on UVR-induced tumor formation in the hairless mouse [158]. The carotenoid was found to exert a limited, but protective, effect with regard to time of tumor appearance and tumor growth. In mixed chemical/UVR carcinogenic protocols, oral administration of β-carotene or canthaxanthin was found to delay the appearance of skin tumors [159]. Employing a UVR-carcinogenic protocol and the Skh-Hr-1 hairless mouse model, it was determined that canthaxanthin, but not β-carotene or phytoene, could slow the development of subsequent tumors if administered immediately after development of the primary tumor, indicating an effect upon the promotion stage of UVR carcinogenesis [160]. Nevertheless, phytoene, when administered (injected intraperitoneally) for ten weeks prior to exposure to a single, large, tumor-initiating fluence of UVR, was photoprotective. Mathews-Roth and Pathak [161] demonstrated a protective effect to sunburn with phytoene in guinea pig skin. Phytoene is the triene precursor to β-carotene and absorbs strongly in the UVB range of the electromagnetic spectrum.
Additional studies confirmed that oral (dietary) administration of 0.1% (w/w) of either β-carotene or canthaxanthin, for six weeks prior to exposure to a single, tumor-initiating UVR dose, and then continuing carotenoid administration during the progression stage of carcinogenesis, could significantly prevent the development of skin tumors [162]. β-carotene, when administered only before UV exposure, provided no significant protection. Thus, in this study, it was inferred that photoprotection resulted only when the carotenoid was administered during the progression phase of tumor development. Subsequently, in a somewhat complicated protocol (complicated from the standpoint that animals received the varying levels of carotenoids in different “run-in” feeding periods ranging from 4 days to one month and that animal ages at the time of irradiation ranged from 8 to 20 weeks), the investigators found that both canthaxanthin and β-carotene provided significant photoprotection to carcinogenesis when the pigment was administered throughout the course of the study and at a level of 0.07% (w/w) in the diet [163]. Lower levels did not provide significant protection. Employing the C3H/HeN mouse model in an UVR-carcinogenic protocol, a diet supplemented with 1% (w/w) canthaxanthin significantly reduced the average skin tumor burden, although the tumor latent period was unaffected [164,165]. At this point, there seemed to be overwhelming experimental evidence to substantiate the anti-UV-carcinogenic potential of β-carotene.
However, in 1998, it was reported that ß-carotene supplementation not only failed to provide protection against UVR-carcinogenesis but resulted in significant exacerbation. Exacerbation by astaxanthin was observed, but not by lycopene [166].
Matrix analyses experimental variables in those studies showing photo-protection or exacerbation. indicated that animal age, carotenoid dose, or type of diet were the most probable causes of the divergent responses to β-carotene. No significant differences in the tumor latent period occurred in young animals (2.5 months of age) fed the semi-defined diet, regardless of β-carotene dose (either 0.07% or 0.3% (w/w) of diet) Nevertheless, tumor multiplicity was significantly increased at each higher dose of the carotenoid. Both young and older animals were equally responsive to exacerbation of UV-carcinogenesis with respect to this tumor parameter [167]. Animal age was not an adequate determinant of carcinogenic response to β-carotene.
It seemed logical that the presence and interactions with other dietary factors in the closed-formula diet, and the lack thereof in the semi-defined diet, might be responsible for the disparate influence of β-carotene on UVR-carcinogenesis. An explanation of the two types of rations employed in the β-carotene studies is warranted. Commercial rodent rations are examples of closed-formula diets [168]. They are composed of feedstuffs such as milled grains, alfalfa, and so forth. The composition of these diets will vary with season, regional locale of source, and market prices. Thus, the specific composition of each lot will vary, making it impossible to quantitatively duplicate. It was this type of ration that was employed in earlier studies in which photo-protection was observed. Semi-defined diets are those formulated from refined foodstuffs, e.g., corn starch, refined oils, casein, and defined vitamin and mineral supplements. It is this type of diet that was employed where exacerbation of UVR-carcinogenesis was observed. Care was taken in this study to ensure that comparable levels of the carotenoid were consumed in both diets and none of the effects, or lack thereof, on carcinogenic expression could be attributed to differential carotenoid intake [169]. Here, it is worth pointing out that the 0.07% β-carotene supplementation in the mouse is equivalent to human consumption, on a body weight basis, to ~160-fold higher levels than that employed in many clinical trials (50 mg/day per ~75 kg body weight). The practical relevance of employing a higher dose of the carotenoid, even if effective as a photoprotectant, would be questionable. Nevertheless, it is apparent from these studies that dietary components, such as other carotenoids, their isomers, or some unidentified phytochemicals, are present in the closed-formula ration that are absent in the semi-defined diet, and which potentiate the carcinogenic response to β-carotene. Moreover, β-carotene has been shown to exhibit either limited antioxidant protection or to behave as a pro-oxidant under oxidative stress conditions [155]. The possible molecular mechanisms and the role of oxygen concentration have been discussed [170]. Some of these mechanisms suggest a role for the interaction of vitamin C with carotenoid radicals. There was no listed vitamin C content in the commercial closed-formula ration in those studies in which β-carotene had provided photoprotection or in those in which no effects of the carotenoid were observed [168]. In previous studies, it was proposed that either a lack of adequate levels of vitamin C in smokers or an excess of vitamin C could stoichiometrically drive the proposed repair mechanism to a pro-oxidant state that, in turn, would block the effective antioxidant activity of vitamin E.
Addressing this issue, hairless mice were fed β-carotene-supplemented semi-defined diets containing varying levels of vitamins C and E (either increasing their concentrations or reducing them to reflect levels found in closed-formula rations) and subjected to a UVR-carcinogenic protocol. The addition of high levels of vitamin E (10-fold) or vitamin C (six-fold) did not ameliorate the exacerbated effects of β-carotene on carcinogenesis, nor did the complete removal of vitamin C from the diet have an effect. However, by reducing the level of vitamin E in the semi-defined diet to that present in the closed-formula ration, there was a significant (nearly two-fold) increase of exacerbation of UVR-carcinogenesis [171]. β-carotene supplementation of the semi-defined diet, at the lower level of vitamin E, significantly (p = 0.0065) increased tumor multiplicity nearly six-fold. These data suggest that β-carotene and vitamin E interact to partially ameliorate the pro-carcinogenic influence of the former. The data with vitamins C and E supplementation also suggest that antioxidant supplementation would have a limited range of upregulation in healthy, well-nourished animals.
A schema for UVR-induced cutaneous free radical quenching, based on one electron reduction potentials, is depicted in the FIGURE. In this schema, UVR produces a radical in skin that is repaired by tocopherol to form a tocopherol radical cation. This radical cation is repaired by ß-carotene and forms the carotenoid radical cation. The1radical cation is, itself, a strong oxidant and if left unrepaired could result in cellular damage leading to exacerbation of carcinogenesis in the semi-defined diet-fed animals. Based on electron transfer potential, ascorbate (vitamin C) should repair the ß-carotene radical cation. However, as noted, the removal of vitamin C or a 6-fold increase in vitamin C level to the semi-defined diet had no effect upon the ß-carotene-induced exacerbation of UVR carcinogenesis (171). Thus, ascorbate does not repair the carotenoid radical cation. The question remains of how the carotene radical cation, itself a strong oxidant, is repaired in the closed-formula diets. It may be isomers of the carotenoid, other carotenoids, other phytochemicals, or a combination of a broad array of these ingredients. This mechanism might explain the divergent results obtained from the two diets [171,172].
Figure 1.
SCHEMA FOR THE REPAIR OF UVR-INDUCED CUTANEOUS FREE. RADICALS BASED ON ONE ELECTRON REDUCTION POTENTIALS.
Figure 1.
SCHEMA FOR THE REPAIR OF UVR-INDUCED CUTANEOUS FREE. RADICALS BASED ON ONE ELECTRON REDUCTION POTENTIALS.
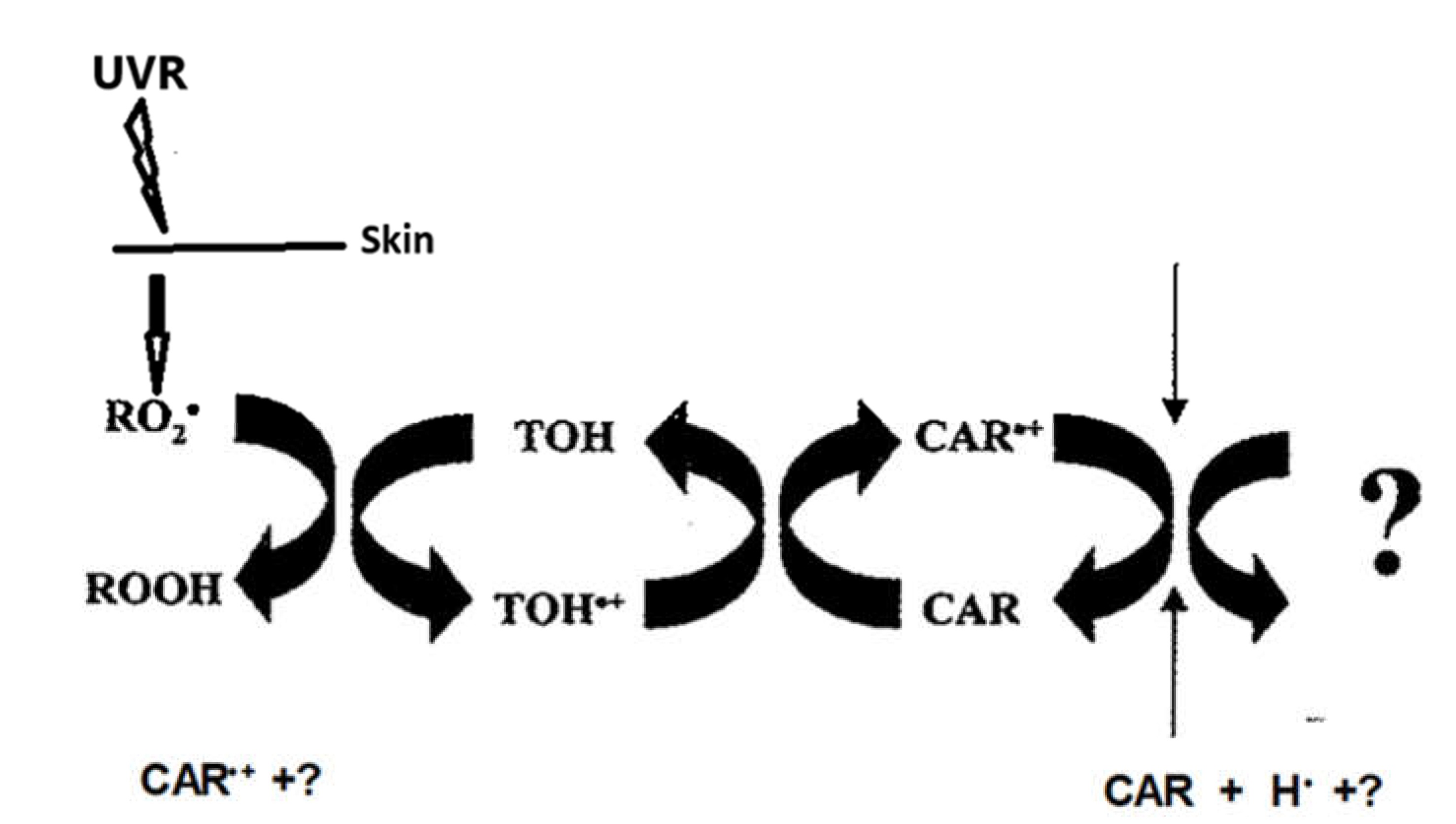
b. Clinical:
It had been assumed that NMSC risk could be ameliorated by β-carotene, based upon its known strong singlet oxygen quenching and antioxidant properties [170,173]; its effect upon human erythema [174], and its effect in photosensitivity diseases [175]. Indeed, a case-control study found that the incidence of NMSC was inversely related to the level of serum β-carotene [176]. However, in a controlled clinical trial in which 50 mg of oral β-carotene was administered daily, there were no significant differences in any of the primary endpoints after five years [177]. The amount of β-carotene administered in this study increased the plasma carotenoid levels nearly 10-fold above that of controls. A second randomized trial in Australia examined the influence of 30 mg/day supplementation of β-carotene on the incidence of basal and squamous cell carcinomas over a four-year period [178]. A small, but statistically insignificant, increase (1508 vs 1146/100,000; 1.315 (0.84–2.19) in the incidence of squamous cell carcinoma was observed with β-carotene supplementation. The investigators concluded that there were no beneficial or detrimental effects on skin cancer rates as a result of β-carotene supplementation. Further evaluations from nested case-control studies showed that β-carotene supplementation had no effect on any of the controlled patient subgroups, i.e., numbers of previous skin cancers, age, gender, smoking, skin type or baseline plasma β-carotene levels [179,180]. Interestingly, those persons who were in the highest quartile of the initial plasma β-carotene level had the lowest risk of death from all causes, but β-carotene supplementation did not affect mortality.
Few studies have examined the influence of β-carotene on the occurrence of melanoma skin cancer. Nevertheless, in three case-control studies, no association was found between blood carotenoid levels and the risk of melanoma [181,182,183]. In one of these studies the risk of melanoma among men and women in the three highest quartiles of β-carotene intake actually increased by 40–50%, albeit neither the risk for the fourth quartile nor the test for trend was statistically significant. A side note of interest is that β-carotene supplementation (50 mg/day for 36 months) has been reported to have beneficial effects in dysplastic nevus syndrome, producing a statistically significant retardation in the size of dysplastic nevi on some body sites. β-Carotene did not have a significant effect on the average number of newly developed nevi [184]. A meta-analysis of randomized controlled trials found that β-carotene supplementation had no effect on the incidence of melanoma (RR, 098; 95% CI, 0.65–1.46) nor upon NMSC risk (RR, 0.99; 95% CI, 0.93–1.05) [185].
Astaxanthin, a xanthophyll carotenoid, is a very effective free radical scavenger and quencher of reactive oxygen and nitrogen species. This carotenoid has been shown to significantly decrease tumor size in a xenograft model and has potent in vivo and in vitro inhibiting effects on melanoma tumor growth and metastasis (cell migration). Astaxanthin inhibits migration of melanoma cells through suppression of metalloproteinases (MPP) [186].
At this point, there seemed to be convincing epidemiological evidence (association) for the cancer-preventive effect of β-carotene. Although intervention clinical trials failed to show a beneficial effect, it was both surprising and disturbing when results from the eight-year intervention trial of the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study (known as the ATBC trial) reported an 18% increase in the incidence of lung cancer in male smokers who were administered 20 mg/day of a β-carotene supplement, compared to non-supplemented controls [187]. Previously, five intervention trials had been undertaken. Except for a trial in which β-carotene had been co-administered with selenium and α-tocopherol, none of these provided evidence of a protective effect of β-carotene [188]. A second lung cancer chemoprevention trial, the β-carotene and retinol efficacy trial (known as CARET), sought to determine whether the combination of β-carotene, with its presumed antioxidant effects, and the tumor-suppressing molecular actions of vitamin A could reduce the incidence of lung cancer in high-risk populations [189]. Participants in the intervention arm of the study received 30 mg/day of β-carotene plus 25,000 IU daily of retinol. β-carotene plus retinol was associated with an increased incidence of lung cancer among the entire cohort of 18,314 smokers, former smokers and asbestos-exposed participants (RR, 1.28; 95% CI, 1.04–1.57; p = 0.02). In view of the ATBC trial results and consideration of the safety of participants in the CARET trial, the intervention was halted about 21 months early. This pretty much signaled the end of β-carotene clinical trials [190]. However, very high doses (75–150 mg/day) of β-carotene continue to be used to treat skin photosensitivity. This is discussed by Bayerl [191] who points out both the criticisms of the ATBC and CARET trials and suggests that the use of β-carotene should be discussed with patients only after weighing the risk/benefit ratio.
The IARC [188] has recommended that “Until further insight is gained, beta-carotene should not be recommended for use in cancer prevention in the general population and it should not be assumed that beta-carotene is responsible for the cancer protecting effects of diets rich in carotenoid containing fruits and vegetables’”
- (a)
- Conclusions from Experimental Studies:
- ∙
- Phytoene protects against sunburn in guinea pig skin.
- ∙
- Eary studies, in which animals were fed closed-formula rations, ß-carotene, canthaxanthin and phytoene were shown to have anti-tumor activity.
- ∙
- Later studies, employing a semi-defined ration, ß-carotene was found to exacerbate UVR-carcinogenesis.
- ∙
- Differences between the two responses to UVR was found to be diet related.
- ∙
- Based upon one electron rate constants between ß-carotene, vitamins E and C, ß-carotene is highly reactive with peroxyl radicals that result with the formation of a carotenoid radical cation (this radical is a strong oxidizing agent, itself) that is repaired by vitamin E to form a vitamin E radical cation that would be repaired by vitamin C to form the vitamin C radical cation. The vitamin C radical cation would either be cleared or repaired by some other phytochemical found in closed-formular rations, but not in semi-defined diets. It also must be remembered that mice synthesize their own vitamin C, unlike humans who must have a dietary source. When vitamin C was either eliminated from the semi-defined diet or increased 6-fold, ß-carotene exacerbation of UVR carcinogenesis was not affected. Nor did 10-fold increases in vitamin E affect exacerbation. However, when vitamin E levels were reduced, there was augmentation of ß-carotene exacerbation of UVR carcinogenesis. These data do not support a role for vitamin C in ß-carotene radical cation repair but do suggest a vitamin E and ß-carotene interaction.
- ∙
- Astaxanthin has been shown to significantly decrease tumor size in a xenograft model and has potent in vivo and in vitro inhibiting effects on melanoma tumor growth and metastasis (cell migration).
- (b)
- Clinical Relevance:
- ∙
- Epidemiological studies have provided persuasive evidence for an association for cancer-preventive effects with ß-carotene.
- ∙
- ß-carotene modestly protects against UVA and UVB -induced erythema.
- ∙
- A controlled clinical trial found that ß-carotene supplementation had no effect on BCC or SCC occurrence.
- ∙
- A randomized trial in Australia found that ß-carotene did not significantly affect the incidence of BCC or SCC over a four-year period, although an increase in SCC (1508 vs 1145/100,000) was observed.
- ∙
- Three case-controlled studies found no association between blood carotenoid levels and risk of melanoma.
- ∙
- A meta-analysis of randomized controlled trials found that ß-carotene had no effect on the incidence of melanoma.
- ∙
- The ATBC trial reported that an 18% increase in the incidence of lung cancer in male smokers administered 20mg/day of ß-carotene occurred.
- ∙
- The CARET trial found that patients administered 30 mg/day of ß-carotene plus 25,000 IU of retinol was associated with an increased incidence of lung cancer among the cohort of 18, 314.
- ∙
- The IARC recommended that ß-carotene should not be used in cancer prevention until further insight is gained. Nor should it be assumed that ß-carotene is responsible for the cancer-protecting effects of diets rich in carotenoid containing fruits and vegetables.
5. Conclusions
The second tenet posited in the free radical theory of aging and cancer, i.e., “add one or more antioxidants to the diet to diminish the effects of free radicals and reduce the risk of cancer” has resulted in a monumental number of studies of systemic antioxidants. So much so, that dietary antioxidants have come to be regarded as a panacea for maintenance of good health. A large international industry has developed to meet consumer demand for these dietary supplements. Unfortunately, this industry is unregulated and commercialization and over-the-counter purchases risk introducing potential health risks. Although a number of antioxidants, including synthetic phenols, polyphenols (including flavonoids) NAC, and GlyNAC, and some carotenoids have been shown to modulate experimental UVR-carcinogenesis, supplementing a highly complex and intricate natural antioxidant defense system with a high level of one type of antioxidant may alter the stoichiometry of the antioxidant pathways – driving the reaction from an antioxidant to a pro-oxidant or pro-carcinogenic state. It must also be recognized that when any chemical is introduced into a biological system, physiological effects occur. Systemically administered antioxidants are processed in the liver and induce hepatic Phase I and II microsomal Phase I and II microsomal detoxification enzymes (mixed function oxidases). These enzymes influence the mutagenic frequency of a range of chemicals and may activate pro-carcinogens that act at other target sites. In addition, the timing of antioxidant therapy (supplementation) is important to the oxidative stress/ROS link to cancer. Early antioxidant supplementation may be effective in reducing ROS-induced malignant transformation, proliferation, and progression of carcinogenesis, but late supplementation in the carcinogenic continuum reduces ROS, suppresses apoptosis, and boosts metastasis. These factors could account for the conflicting and detrimental results from antioxidant supplementation studies. Consequently, the World Cancer Research Fund/American Institute for Cancer Research concluded “A general recommendation to consume supplements for cancer prevention might have unexpected adverse effects” and thus, “Dietary supplements are not recommended for cancer prevention.” However, topically applied antioxidants may play a beneficial role as adjuvants to sunscreens and cosmeceuticals but suffer from the limitations of topical application efficacy manifested by other photo-protectants. Caution is warranted with strong antioxidants, such as C60, that has a high permeation through skin, so that concentrations are employed that avoid the risks associated with systemically administered antioxidants. Nevertheless, the perfection of new antioxidant delivery systems hold promise for targeted antioxidant therapy and the reduction of ROS and oxidative stress remains a fertile avenue of investigation for preventing and ameliorating conditions that are linked to skin cancer.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Bray, F. ; Laversanne, M,; Sung, H.; et al.: Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin, 2: 74(3). [CrossRef]
- Siege, l R. L.; Miller, K. D., Ed.; Fuchs H.E.; Jemal, A.: Cancer Statistics, 2021. [Google Scholar] [CrossRef]
- Jemal, A, Torre, L, Soerjomataram I, Bray F (Eds). The Cancer Atlas. Third Ed. Atlanta, GA: American Cancer Society, 2019.: http://www.cancer.
- Urban, K. ; Mehrmal, S, Uppal, P.; et. al.: The global burden of skin cancer: A longitudinal analysis from the Global Burden of Disease Study, 1990-2017. JAAD Int, 2021. [Google Scholar] [CrossRef]
- Rogers HW, Weinstock MA, Harris AR, Hinckley MR, Feldman SR, Fleischer AB, Coldiron BM. Arch Dermatol. 2010, Incidence estimate of nonmelanoma skin cancer in the United States, 2006;146(3):283-287.
- Cancer Facts and Figures, 2024. Cancer.org/research/cancer/facts-statistics/all-cancer-facts-figures.
- Collins, KK. : The diabetes-cancer link. 2014, Diabetes Spectrum, 27: 276-280.
- Black, HS: Oxidative Stress and ROS Link Diabetes and Cancer. 2024, J. Mol. Pathol., 5, 96–119. https:// doi.org/10. 3390.
- Garrison, F.H. ; An Introduction to the History of Medicine, Saunders: Philadelphia, USA, 1929; p. 31.
- Warren, S. ; The immediate causes of death in cancer. AM. J. Med. Sci. 1932, 184, 610–615. [Google Scholar] [CrossRef]
- Harmon, D. ; Role of free radicals in mutation, cancer, aging, and the maintenance if life. Radiat. Res. 1962, 16. [Google Scholar] [CrossRef]
- Harmon, D. ; The free radical theory of aging. In Free Radicals in Biology, Pryor, WA. Ed.; Academic Press; NY, USA, 1982; pp. 255-275.
- Black HS: Reassessment of a free radical theory of cancer with emphasis on ultraviolet carcinogenesis. Integr. Cancer Therapies. 2004, 3:279-293.
- Watson, A.F. , Mellanby, E.: Tar cancer in mice II. The condition of the skin when modified by external treatment or diet, as a factor in influencing this cancerous reaction. Br. J. Exp. Pathol. 1930, 11: 311-322.
- Elmets, CA. , Mukhtar, H.: Ultraviolet radiation and skin cancer: progress in pathophysiologic mechanisms. Prog. Dermatol. 1996, 30: 1-16.
- Baumann, CA. , Rusch, HP.: Effect of diet on tumors induced by ultraviolet light. Am. J. Cancer. 1939, 35: 213-221.
- Black, HS. , Lenger, W., Phelps, AW., Thornby, JI.: Influence of dietary lipid upon ultraviolet light-carcinogenesis. Nutr. Cancer. 1983, 5: 59-68.
- Black, HS. : Dietary factors in ultraviolet carcinogenesis. The Cancer Bull. 1993, 45: 232- 237.
- Ip, C. , Carter, CA.; Ip, MM.: Requirement of essential fatty acid for mammary tumorigenesis in the rat. Cancer Res. 1985, 45: 1997-2001. [Google Scholar]
- Reeve, VE. , Matheson, M., Greenoak, GE., Canfield, PJ., Boehm-Wilcox, C., Gallagher, CH.: Effect of dietary lipid on UV light carcinogenesis in the hairless mouse. Photochem. Photobiol. 1988, 48: 689-696. [CrossRef]
- 21.Orengo, IF. 21.Orengo, IF., Black, HS., Kettler, AH., Wolf, JE., Jr.: Influence of dietary Menhaden oil upon carcinogenesis and various cutaneous responses to ultraviolet radiation. Photochem. Photobiol. 1989, 49: 71-77.
- Chung, HT. , Burnham, DK., Robertson, B., Roberts, LK., Daynes, RA.: Involvement of prostaglandins in the immune alterations caused the exposure of mice to ultraviolet radiation. J. Immunol. 1986, 137: 2378-2884.
- Henderson, CD. , Black, HS., Wolf, JE, Jr.: Influence of omega-3 and omega-6 fatty acid sources on prostaglandin levels in mice. Lipids. 1989, 24:502-505.
- Orengo, IF. , Gerguis, J., Phillips, R., Guevare. A., Lewis, AT., Black, HS.: Celecoxib, a cyclooxygenase 2 inhibitor as a potential chemopreventive to UV-induced skin cancer. 2002, Arch. Dermatol. 138: 751-755. www.ARCHDERMATOL.
- Meeran, SM. , Singh, Nagy, TR., Katiyar, SK.: High-fat diet exacerbates inflammation and cell survival signals in the skin of ultraviolet B-irradiated C57BL/6 mice. Toxicol Appl. Pharmacol. 2009, 15;241(3):303-310. [CrossRef]
- Utsumi, H. , Ichikawa, K., Takeshita, K.: In vivo ESR measurements of free radical reactions in living mice. Toxicol. Lett. 1995, 82: 561-565.
- Black, HS. : ROS: A step closer to elucidating their role in the etiology of light-induced skin disorders. J. Invest. Dermatol. 2004, 122: xiii-ix (Editorial) Doi. 10111/j.0022-202X.2004.22625.
- Black, HS. : Oxidative Stress and ROS Link Diabetes and Cancer. J. Mol. Pathol. 2024, 5: 96–119. https:// doi.org/10. 3390. [Google Scholar]
- Djuric, Z., Lewis, SM., Lu, MH., Mayhugh, M,, Tang, N., Hart, RW.: Effect of varying dietary fat levels on rat growth and oxidative DNA damage. Nutr Cancer 2001, 39:214–9. [CrossRef]
- Fischer, MA. , Black, HS.: Modification of membrane composition, eicosanoid metabolism, and immunoresponsiveness by dietary omega-3 and omega-6 fatty acid sources, modulators of ultraviolet-carcinogenesis. Photochem. Photobiol. 1991, 54: 381-387.
- Moison, RM. , Beijersbergen Van Henegouwen, GM.: Dietary eicosapentaenois acid prevents systemic immunosuppression in mice induced by UVB radiation. Radiat. Res. 2001, 156: 36-44.
- Kang, JX. , Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leukot. Essent. Fatty acids. 2007, 77: 263-267.
- Xia, S. , Lu, Y., Wang, J., He, C., Hong, S., Serhan, CN., Kang, JX.: Melanoma growth is reduced in fat-1 transgenic mice: Impact of omega-6/omega-3 essential fatty acids. Proc. Natl. Acad. Sci. USA 2006, 103: 12499-12504.
- Black, HS. , Okotie-Eboh, G., Gerguis, J.: Dietary fat modulates immunoresponsiveness in UV-irradiated mice. Photochem, Photobiol. 1995, 62: 964-969.
- Okotie-Eboh, G. , Gerguis, J., Black, HS.: Influence of dietary lipid on hapten-specific UV-induced immunosuppression. Photodermatol.Photoimmunol. Photomed. 1998, 14:116-118.
- Black, HS. , Thornby, JI., Gerguis, J., Lenger, W.: Influence of dietary omega-6, 3-fatty acid sources on the initiation and promotion stages of photocarcinogenesis. Photochem. Photobiol. 1992, 56: 195-199.
- Forbes, PD. , Beer, JZ., Black, H.S, Cesarini, J-P., Cole, CA., Davies, RE., Davitt, JM., deGruijl, F., Epstein, J., Fourtanier, A. et.al.: Standardized protocols for photocarcinogenesis safety testing. Frontiers in Bioscience 2003, 8: 848-854.
- Graham, S. : Results of case-controlled studies of diet and cancer in Buffalo, New York. Cancer Res. 1983, 43 (Suppl): 2409-2413.
- Hunter, DJ. , Colditz, GA., Stampler, MJ.,Rosner, B., Willett, WC., Soeizer, FE.: Diet and risk of basal cell carcinoma of the skin in a prospective cohort of women. Ann. Epidermal. 1992, 2: 231-239.
- van Dam, RM. , Huang, Z., Giovannucci, E., Rimm, EB., Hunter, DJ., Colditz, GA., Stampfer, MJ., Willett, WC.: Dit and basal cell carcinoma of the skin in a prospective cohort of men. Am. J. Clin. Nutr. 2000, 71: 135-141.
- Davies, TW. , Treasure, FP., Welch, AA., Day, NE.: Diet and basal cell skin cancer: results from the EPIC-Norfolk cohort. Br. J. Dermatol. 2002, 146: 1017-1022.
- Gamba, CS. , Stefanick, ML, Larson, J., Linos., E., Sims, ST., Marshall, J., Van Horn, L., Zeitouni, N., Tang, JY.: Low-Fat Diet and Skin Cancer Risk: The Women's Health Initiative Randomized Controlled Dietary Modification Trial. Cancer Epidemiol. Biomarkers Prev. 2013, 22 (9): 1509–1519. [CrossRef]
- Thind, IS. : Diet and cancer – an international study. Int. J. Epidemiol. 1: 1986, 15, 1986. [Google Scholar]
- Armstrong, B. , Doll, R. Environmental factors and cancer incidence and mortality in different countries, with special reference to dietary practices. Int. J. Cancer, 1975, 15: 617-631.
- Rackett, SC. , Rothe, MJ., Grant-Kels, JM.: Diet and Dermatology. J. Am. Acad. 4: Dermatol, 1993, 29, 1993. [Google Scholar]
- Karagas, MR. : Occurrence of cutaneous basal cell an squamous cell malignancies among those with a prior history of skin cancer. J. Invest. Dermatol. 1: 1994, 102 (Suppl), 1994. [Google Scholar]
- Black, HS. , Herd, JA., Goldberg, LH., Wolf, JE., Thornby, JI., Rosen, T., Bruce, S., Tschen, JA., Foreyt, JP., Scott, LW.et.al.: Jaax, S.: Effect of a low-fat diet on the incidence of actinic keratosis. N. Engl. J. Med. 1994, 330: 1272-1275.
- Black, HS. , Thornby, JI., Wolf, JE., Goldberg, LH., Herd, JA., Rosen, T., Bruce, S., Tschen, JA., Scott, LW., Jaax, S., et.al.: Evidence hat a low-fat diet reduces the occurrence of non-melanoma skin cancer. Int. J. Cancer. 1995, 62: 165-169.
- Jaax, S. , Scott, LW., Wolf, JE., Thornby, JI., Black, HS.: General guidelines for a low-fat diet effective in the management and prevention of nonmelanoma skin cancer. Nutr. Cancer, 1997, 27: 150-156.
- 49. Black, HS. 49. Black, HS.: Omega-3 fatty acids and non-melanoma skin cancer. In Handbook of diet. Nutrition and skin. Preedy, VR., Ed.: Wageningen Academic: Wagenin, The Netherlands, 2012; pp. 367-378.
- Bang, H. , Dyerberg, J.: Plasma lipids and lipoprotein pattern in Greenlandic west-coast Eskimos. Acta Medical Scandinavia, 1972, 192: 85-94.
- Dyerberg, J. , Bang, H.: Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 1978, 15: 117-119.
- Orengo, IF. , Black, HS., Wolf, JE, Jr., Influence of fish oil supplementation on the minimal erythema dose in humans. Arch. Dermatol. Res., 1992, 284: 219-221.
- Rhodes, LE. , O’Farrell, S., Jackson, MJ., Friedmann, PS.: Dietary fish-oil supplementation in humans reduces UVB-erythemal sensitivity but increases lipid peroxidation. J. Invest. Dermatol. 1994, 103: 151-154.
- Rhodes, LE. , Shahbakhti, H., Azurdia, RM., Moison, RM., Steenwinkel, M.J, Homburg, MI., Dean MP., McArdle, F., Beijersbergen van Henegouwen, GM., Epe, B., et. Al.: Effect of eicosapentaenoic acid, an omega-3 polyunsaturated fatty acid, on UVR-related cancer risk in humans. An assessment of early genotoxic markers. Carcinogenesis. 2003, 24(5):919-925. [CrossRef] [PubMed]
- Pupe, R. , Moison, R., De Haes, P., van Henegouwen, GB., Rhodes, L., Degree, H., Garmyn, M.: Eicosapentaenoic Acid, a n-3 Polyunsaturated Fatty Acid Differentially Modulates TNF-α, IL-1α, IL-6 and PGE2 Expression in UVB-Irradiated Human Keratinocytes, J. Invest. Dermatol. 2002, 118: 692-698. [CrossRef]
- Shahbakhti,H., Watson, REB., Azurdia,,RM., Ferreira, CZ., Garmyn, M., Rhodes, LE.: Influence of Eicosapentaenoic Acid, an Omega-3 Fatty Acid, on Ultraviolet-B Generation of Prostaglandin-E2 and Proinflammatory Cytokines Interleukin-1β, Tumor Necrosis Factor-α, Interleukin-6 and Interleukin-8 in Human Skin In Vivo. Photochem. Photobiol. 2004, 80: 231-235. [CrossRef]
- Storey. A., McArdle, F., Friedmann, PS., Jackson, MJ., Rhodes, LE.: Eicosapentaenoic Acid and Docosahexaenoic Acid Reduce UVB- and TNF-α-induced IL-8 Secretion in Keratinocytes and UVB-induced IL-8 in Fibroblasts. J. Invest. Dermatol. 2005, 124: 248-255. [CrossRef]
- Pilkington, SM. , Massey, KA., Bennett, SP., MI Al-Aasswad, N., Roshdy,K., Gibbs, NK. Friedmann, PS., Nicolaou, A., Rhodes, LE.: Randomized controlled trial of oral omega-3 PUFA in solar-simulated radiation-induced suppression of human cutaneous immune responses. 2013, Am. J. Clin. Nutr. 97: 646-652. [CrossRef]
- Black, HS. , Rhodes, LE.: The potential of omega-3 fatty acids in the prevention of non-melanoma skin cancer. Can. Det. Prev. 2006, 30: 224-232. Doi. 10.1016/j.cdp.2006.04.
- Rahrovani, F. , Javanbakht, MH., Ghaedi, E., Mohammadi, H., Ehsani, AH., Esrafili, A., Djalali, M.: Erythrocyte Membrane Unsaturated (Mono and Poly) Fatty Acids Profile in Newly Diagnosed Basal Cell Carcinoma Patients. Clin Nutr Res. 2018, 7: 21-30. [CrossRef]
- Harris, WS. : The Omega-6: Omega-3 ratio: A critical appraisal and possible successor. Prostaglandins, Leukotrienes and Essential Fatty Acids, 2018, 132, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hakim, I. , Harris, R., Ritenbaugh, C.: Fat intake and risk of squamous cell carcinoma of the skin. Nutr. Cancer. 2000, 36: 155-162. [CrossRef]
- Wallingford, SC. , Hughes, MC., Adèle C. Green, AC., van der Pols, JC.: Plasma Omega-3 and Omega-6 Concentrations and Risk of Cutaneous Basal and Squamous Cell Carcinomas in Australian Adults. Cancer Epidemiol. Biomarkers Prev. 2013, 22): 1900–1905. [CrossRef]
- Wallingford, SC. , van As, JA., Hughes, MC., TI., Green, AC., van der Pols, JC.: Intake of omega-3 and omega-6 fatty acids and risk of basal and squamous cell carcinomas of the skin: a longitudinal community-based study in Australian adults. Nutr. Cancer. 2012, 2012;64:982-990. [CrossRef]
- Seviiri, M. , Law, MH., Ong, JS., Gharahkhani, P., Nyholt, DR., Olsen., CM., Whiteman., DC., MacGregor, S.: Polyunsaturated Fatty Acid Levels and the Risk of Keratinocyte Cancer: A Mendelian Randomization Analysis. Cancer Epidemiol. Biomarkers Prev. 2021, 30:1591-1598. [CrossRef]
- Wu. B, Pan, F., Wang, Q., Liang Q., Qiu, H., Zhou, S., Zhou, X.: Association between blood metabolites and basal cell carcinoma risk: a two-sample Mendelian randomization study. Front. Endocrinol. 2024, 15:1413777. [CrossRef]
- Park MK, Li WQ, Qureshi AA, Cho E. Fat Intake and Risk of Skin Cancer in U.S. Adults. Cancer Epidemiol Biomarkers Prev. 2018,27:776-782. [CrossRef]
- Miura K, Way M, Jiyad Z, et al. Omega-3 fatty acid intake and decreased risk of skin cancer in organ transplant recipients. European Journal of Nutrition. 2021 Jun;60(4):1897-1905. [CrossRef]
- Miura K, Vail A, Chambers D, Hopkins PM, Ferguson L, Grant M, Rhodes LE, Green AC. Omega-3 fatty acid supplement skin cancer prophylaxis in lung transplant recipients: A randomized, controlled pilot trial. J Heart Lung Trans-plant. 2019 Jan;38(1):59-65. [CrossRef]
- Yen, A. , Black, HS., Tschen, J.: Effect of dietary omega-3 and omega-6 fatty acid sources on PUVA-induced cutaneous toxicity and tumorigenesis. Arch. Dermatol. Res. 1994, 286: 331-336.
- Gibbs, NK. , Honigsmann, H., Young, AR.: PUVA treatment strategies and cancer risk. Lancet, 1986, 1: 150-151.
- Black, HS. , Chan, JT.: Suppression of ultraviolet light-induced tumor formation by dietary antioxidants. J. Invest. Dermatol. 1975, 65: 412-414.
- Black, HS. , Lenger, WA., Gerguis, J., Thornby, JI.: Relation of antioxidants and level of dietary lipid to epidermal peroxidation and ultraviolet carcinogenesis. Cancer Res. 1985, 45: 6254-6259.
- Black, HS. , Chan, JT., Brown, GE.: Effects of dietary constituents on ultraviolet light-mediated carcinogenesis. Cancer Res. 1978, 38: 1384-1387.
- Pauling, L. , Willoughby, R., Reynolds, R., Blaisdell, BE., Lawson, S.: Incidence of squamous cell carcinoma in hairless mice irradiated with ultraviolet light on relation to intake of ascorbic acid (vitamin C) and D,L-alpha-tocopherol acetate (vitamin E). 1982, Int. J. Vitam. Nutr. Res. Suppl. 23: 53-82.
- De Rios, G. , Chan, JT., Black, HS., Rudolph, AH., Knox, JM.: Systemic protection by antioxidants against UVL-induced erythema. J Invest Dermatol. 1978, 70: 123-5. [CrossRef]
- Peterson, AO. , McCann, V., Black, HS.: Dietary modification of UV-induced epidermal ornithine decarboxylase, 1980, J. Invest. Dermatol. 75: 408-410.
- Black, HS. , Kleinhans, CM., Hudson, HT., Sayre, RM., Agin, PP.: Studies on the mechanism of protection by butylated hydroxytoluene to UV-radiation damage. Photobiochem. Photobiophys. 1980, 1: 119-123.
- Daniel, JW. , Gage, JB.: The absorption and excretion of butylated hydroxytoluene (BHT) in the rat. Food Cosmet. Toxicol. 1965, 3: 405-415. [CrossRef]
- Hathway, DE. : Metabolic fate in animals of hindered phenolic antioxidants in relation to their safety evaluation and antioxidant function. Adv. Food Res. 1966, 15: 1-56. [CrossRef]
- Black, HS. , Lenger, W., Gerguis, J., McCann, V.: Studies on the photoprotective mechanism of butylated hydroxytoluene. Photochem. Photobiol. 1984, 40: 69-75.
- Koone, MD. , Black, HS.: A mode of action for butylated hydroxytoluene-mediated photoprotection. J. Invest. Dermatol. 1986, 87: 343-347. [CrossRef]
- Astbury, WT. , Street, A.: X-ray studies of the structure of hair, wool, and related fibres. Philos. Trans. R. Soc. Lond [Biol.] 1932, A230: 75-102.
- Solan, JL. , Laden, K.: Factors affecting the penetration of light through stratum corneum. J. Soc. Cosmet. Chem. 1977, 28: 125-137.
- Owens, DW. , Knox, JM., Hudson, HT., Troll, D.: Influence of humidity on ultraviolet injury. J. Invest. Dermatol. 1975, 64: 250-252.
- Black, HS. , Tigges, J.: Evaluation of structurally related phenols for anti-photocarcinogenic and photoprotective properties. Photochem. Photobiol. 1986, 43: 403-408. [CrossRef]
- IARC Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Humans: Some Naturally Occurring and Synthetic Food Components, Furocoumarins and Ultraviolet Radiation. IARC, Lyon, France, Vol. 40, 1986, pp.161-206.
- Williams, GM. , Iatropoulos, MJ., Whysner, J.: Safety assessment of butylated hydroxyanisole and butylated hydroxytoluene as antioxidant food additives. Food. Chem. Toxicol. 1999, 37: 1027-1038. [CrossRef]
- Lanigan, RS. , Yamarik, TA.: Final report on the safety assessment of BHT (1). Int. J. Toxicol. 2002, 21 ( Suppl 2): 19-94. [CrossRef]
- Olsen, P. , Bille, N., Meyer, O.: Hepatocellular neoplasms in rats induced by butylated hydroxytoluene (BHT). Acta. Pharmacol. Toxicol. 1983, 53: 433-434.
- Witschi, HP. , Morse, CC.: Enhancement of tumor formation in mice by dietary butylated hydroxytoluene: dose-time relationships and cell kinetics. J. Natl. Cancer Inst. 1983, 71: 859-866.
- Chan, JT. , Ford, JO., Rudolph, AH., Black, HS.: Physiological changes in hairless mice maintained on an antioxidant supplemented diet. Experientia, 1977, 33: 72.
- Black, HS. , Gerguis, J.: Use of the Ames test in assessing the relation of dietary lipid and antioxidants to N-2-fluorenylacetamide activation. J. Environ. Pathol. Toxicol. 1980, 4: 131-138.
- Centers for Science in the Public Interest. Butylated Hydroxytoluene (BHT) Washington, DC 20005. https://www.cspinet.
- Zhou, Y. , Zheng, J., Li, Y., Xu, D-P., Li, S., Chem, Y0M., Li, H-B.: Natural polyphenols for prevention and treatment of cancer. 5: Nutrients 2016, 8, 2016. [Google Scholar] [CrossRef]
- Afaq, F. , Katiyar, SK.: Polyphenols: Skin Photoprotection and inhibition of photocarcinogenesis. Mini Rev. Med. Chem. 2011, 11: 1200-1215.
- Lin, BW. , Gong, CC., Song, HF., Cu,i YY.: Effects of anthocyanins on the prevention and treatment of cancer. Br J Pharmacol. 2017, 174: 1226-1243. [CrossRef]
- Wang, E. , Liu, Y., Xu, C., Liu, J.: Antiproliferative and proapoptotic activities of anthocyanin and anthocyanidin extracts from blueberry fruits on B16-F10 melanoma cells. Food Nutr. Res. 2017, 19:1325308. [CrossRef]
- Huang, H-P., Yuan-Wei Shih, Y-W., Yun-Ching, Chang, Y-C., Chi-Nan Hung, C-N., Chau-Jong Wang, C-J.: Chemoinhibitory Effect of Mulberry Anthocyanins on Melanoma Metastasis Involved in the Ras/PI3K Pathway J. Agric. Food Chem. 9: 2008, 56, 2008. [CrossRef]
- Afaq, F. , Syed, DN., Malik, A., Hadi, N., Sarfaraz, S., Kweon, MH., Khan, N., Zaid, MA., Mukhtar, H.: Delphinidin, an anthocyanidin in pigmented fruits and vegetables, protects human HaCaT keratinocytes and mouse skin against UVB-mediated oxidative stress and apoptosis. J Invest Dermatol. 2007, 127: 222-232. [CrossRef]
- Wang, LS. , Stoner, GD.: Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269: 281-290. [CrossRef]
- Negri, A. , Naponelli, V., Rizzi F., Bettuzzi, S.: Molecular Targets of Epigallocatechin-Gallate (EGCG): A Special Focus on Signal Transduction and Cancer. Nutrients. 2018 Dec 6;10(12):1936. [CrossRef]
- Gębka, N. , Gębka-Kępińska, B., Adamczyk, J., Mizgała-Izworska. E.: The use of flavonoids in skin cancer prevention and treatment. J. Pre-Clin. and Clin. Res. 2022,16:108-113. [CrossRef]
- Mukhtar, H. , Ahmad, N.: Tea polyphenols: prevention of cancer and optimizing health. Am. J. Clin. Nutr. 2000, ;71S:1698S-1702S. [CrossRef]
- Mukhtar H, Wang ZY, Katiyar SK, Agarwal R. Tea components: antimutagenic and anticarcinogenic effects. Prev Med. 1992 May;21(3):351-60. [CrossRef]
- Katiyar, SK. : Green tea prevents non-melanoma skin cancer by enhancing DNA repair. Arch Biochem Biophys. 2011, 15::152-158. [CrossRef]
- Meeran SM, Akhtar S, Katiyar SK. Inhibition of UVB-induced skin tumor development by drinking green tea polyphenols is mediated through DNA repair and subsequent inhibition of inflammation. J Invest Dermatol. 2009,129:1258-70. [CrossRef]
- Gensler, HL. , Timmermann, BN., Valcic, S., Wächter, GA., Dorr, R., Dvorakova, K., Alberts, DS.: Prevention of photocarcinogenesis by topical administration of pure epigallocatechin gallate isolated from green tea. Nutr. Cancer. 1996; 26:325-335. [CrossRef]
- Bushman, JL: Green tea and cancer in humans: A review of the literature. Nutr. Cancer 1998, 31: 151–159. [CrossRef]
- Zheng, SY. , Li, Y., Jiang, D., Zhao, J., Ge, JF.: Anticancer effect and apoptosis induction by Quercetin in the human lung cancer cell line A-549. Mo; Med. Rep. 2012, 822-826.
- Chen, HS. , Bai, MH., Zhang, T., Li, GD., Liu, M.: Ellagic acid induces cell cycle arrest and apoptosis through TGF-beta/Smad3 signaling pathway in human breast cancer MCF-7 cells. Int. J. Oncol. 2015, 46: 1730-1738.
- Berman, AY., Motechin; Wiesenfeld, MY. , Holz, MK.: The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis Oncol. 3: 2017, 1, 2017. [Google Scholar] [CrossRef]
- AL-Ishag, RK. , Abotaleb, M., Kubatka, P., Kajo, K.., Busselberg, D.: Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. 4: Biomolecules 2019, 9, 2019. [Google Scholar] [CrossRef]
- Aziz, MH. , Reagan-Shaw., S., Wu, J., Longley, J., Ahmad, N.: Chemoprevention of skin cancer by grape constituent resveratrol: relevance to human disease? FASEB J. 2005, 19: 1193-1195. [CrossRef]
- Salehi, B. , Mishra, AP., Nigam, M., Sener, B., Kilic, M., Sharifi-Rad, M., Fokou, PVT., Martins, N., Sharifi-Rad, J.: Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines. 2018, 9:91. [CrossRef]
- Aggarwal, BB. , Kumar, A., Bharti, AC.: Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Res. 2003, 23:363-98.
- Tsai, K-D., Lin, J-C., Yang,S-m., Tseng,M-J., Hsu, J-D. Lee, Y-J. Cherng, L-M.; Curcumin Protects against UVB-Induced Skin Cancers in SKH-1 Hairless Mouse: Analysis of Early Molecular Markers in Carcinogenesis Evidence-Based Complementary and Alternative Med. 2012, 1-11. [CrossRef]
- Phillips, J. , Moore-Medlin, T., Sonavane, K., Ekshyyan, O., McLarty, J., O Nathan, C-A.: Curcumin inhibits UV radiation-induced skin cancer in SKH-1 mice. Otolaryngol. Head Neck Surg. 2013, 148:797-803. [CrossRef]
- Subramanian, V. , Venkatesan, B., Tumala, A, Vellaichamy, E.: Topical application of Gallic acid suppresses the 7,12-DMBA/Croton oil induced two-step skin carcinogenesis by modulating anti-oxidants and MMP-2/MMP-9 in Swiss albino mice. Food and Chem.Toxicol.2014, 66: 44-55. [CrossRef]
- de Jesus, SF. , de Souza, MG., Queiroz, LDRP., de Paula, DPS., Tabosa, ATL., Alves, WSM., da Silveira, LH., Ferreira, ATDS., Martuscelli, OJD., Farias, LC., et.al.: Gallic acid has an inhibitory effect on skin squamous cell carcinoma and acts on the heat shock protein HSP90AB1. Gene. 2023, 30: 851:147041. [CrossRef]
- Singh, A. , Mishra, A.: Gallic acid: molecular rival of cancer. Environ. Toxicol. Pharmacol.. 2013, 35:473-485. [CrossRef]
- Shanmugam, M. , Rejitharaji, S., Prabhaka, T., Murugaraj, M.. Singh, RB.: Modulating Effect of Ferulic Acid on NF-kB, COX-2 and VEGF Expression Pattern During 7, 12-Dimethylbenz(a)anthracene Induced Oral Carcinogenesis The Open Nutraceuticals J. 2014, 7: 33-38: DOI 10. 2174. [Google Scholar]
- Eroglu, C. , Secme, M., Bagei, G., Dodurga, Y.: Assessment of the anticancer mechanism of ferulic acid via cell cycle and apoptotic pathways in human prostate cancer cell lines. Tumor Biol, 2015, 36: 9437-9446.
- Lin, X-F., Min, W., Luo, D.: Anticarcinogenic effect of ferulic acid on ultraviolet-B irradiated human keratinocyte HaCaT cells. J. 1: Medicinal Plants, 2010, 4, 2010. [CrossRef]
- Ambothi, K. , Prasad, N., Rajendra, N., Balupillai, A.: Ferulic acid inhibits UVB-radiation induced photocarcinogenesis through modulating inflammatory and apoptotic signaling in Swiss albino mice. Food and Chem. Toxicol. 2015, 82: 72-78. [CrossRef]
- Alexander, C. The Illiad, Homer, A New Translation.; HarperCollins Publishers, Sydney, NSW 2000, Australia, 2016; www.harpercollins.com.
- Kyriazia, M. , Yovaa, D., Rallis, M., Limac, A.: Cancer chemopreventive effects of Pinus Maritima bark extract on ultraviolet radiation and ultraviolet radiation-7,12,dimethylbenz(a) anthracene induced skin carcinogenesis of hairless mice. Cancer Letters. 2006, 237: 234–241. www.elsevier.
- The Linus Pauling Institute, Oregon State University. https://lpi.oregonstate.
- Dunham, WB. , Zuckerkandl, E., Reynolds, R., Willoughby, R., Marcuson, R., Barth, R., Pauling, L.: Effects of intake of L-ascorbic acid on the incidence of dermal neoplasms induced in mice by ultraviolet light. Proc. Natl. Acad. Sci. U S A. 1982, 79: :7532-7536. [CrossRef]
- Gerrish, KE. , Gensler, HL.: Prevention of photocarcinogenesis by dietary vitamin E. Nutr. Cancer. 1993, 19:125-33. [CrossRef]
- Gensler, HL, Magdaleno M. Topical vitamin E inhibition of immunosuppression and tumorigenesis induced by ultraviolet irradiation. Nutr. Cancer. 1991;15:97-106. [CrossRef]
- Gensler, HL. , Aickin, M., Peng, YM., Xu, M.: Importance of the form of topical vitamin E for prevention of photocar-cinogenesis. Nutr. Cancer. 1996, 26:183-91. [CrossRef]
- Burke, KE. : Interaction of vitamins C and E as better cosmeceuticals. Dermatologic Therapy 2007, 20: 314–32. Blackwell Publishing, Inc. [CrossRef]
- Burke, KE. : Photoprotection of the Skin with Vitamins C and E: Antioxidants and Synergies. In: Pappas, A. (eds) Nutrition and Skin. 2011, Springer, New York, NY. [CrossRef]
- Mussa, A. , Mohd Idris, RA., Ahmed, N., Ahmad, S., Murtadha, AH., Tengku Din, TADAA., Yean, CY., Wan, A., Rahman, WF., Lazim, N.,et. Al.: High-Dose Vitamin C for Cancer Therapy. Pharmaceuticals (Basel). 2022, 3: 15:711. [CrossRef]
- Bodeker, KL. , Smith, BJ., Berg, DJ., Chandrasekharan, C., Sharif, S., Fei, N., Vollstedt, S., Brown, H., Chandler, M., Lorack, A., et. al.: A randomized trial of pharmacological ascorbate, gemcitabine, and nab-paclitaxel for metastatic pancreatic cancer. Redox Biology 2024, 77: 103375, www.elsevier.
- Cotter. MA., Thomas, J., Cassidy, P., Robinette, K., Jenkins, N., Florel, SR., Leachman S, Samlowski, WE., Grossman, D.: N-acetylcysteine protects melanocytes against oxidative stress/damage and delays onset of ultraviolet-induced mela-noma in mice. Clin Cancer Res. 2007, 113(19):5952-8. [CrossRef]
- Le Gal, K.; Ibrahim, M.X.; Wiel, C.; Sayin, V.I.; Akula, M.K.; Karlsson, C.; Dalin, M.G.; Akyürek, L.M.; Lindahl, P.; Nilsson, J.; et al. Antioxidants can increase melanoma metastasis in mice. Sci. Transl. Med. 2015, 7, 308re8 wwwScienceTranslationalMedicineorg. [Google Scholar] [CrossRef]
- Assi, M. The differential role of reactive oxygen species in early and late stages of cancer. Am. J. Physiol.-Regul. In-tegr. Comp. Physiol. 2017, 313, R646–R653. [Google Scholar] [CrossRef]
- D'Agostini,F., Balansky,RM., Camoirano, A., De Flora, S. 6: Modulation of light-induced skin tumors by N -acetylcysteine and/or ascorbic acid in hairless mice Carcinogenesis, 2005, 26, 2005. [CrossRef]
- Kumar, P. , Osahon, OW. , Sekhar, RV.: GlyNAC (Glycine and N-Acetylcysteine) Supplementation in Mice Increases Length of Life by Correcting Glutathione Deficiency, Oxidative Stress, Mitochondrial Dysfunction, Abnormalities in Mitophagy and Nutrient Sensing, and Genomic Damage. Nutrients 2022, 14, 1114. [Google Scholar] [CrossRef]
- Sekhar, R.V. : GlyNAC (Glycine and N-Acetylcysteine) supplementation improves impaired mitochondrial fuel oxidation and lowers insulin resistance in patients with type 2 diabetes: Results of a pilot study. Antioxidants 2022, 11: 154. [CrossRef]
- Fernandes, NB. , Shenoy, RUK., Kajampady, MK., Dcruz, CEM., Shirodkar, RK., Kumar, L., Verma, R.: Fullerenes for the treatment of cancer: an emerging tool. Environ. Sci. Pollut. Res. Int. 2022, ;29: 58607-58627. [CrossRef]
- Martins, M. , Azoia, NG., Melle-Franco, M., Ribeiro, A., Cavaco-Paulo, A.: Permeation of skin with (C60) fullerene dispersions. Eng. Life Sci. 2017,17(7): 732-738. [CrossRef]
- Rondags, A. , Yuen, WY., Jonkman, MF., Horváth, B.: Fullerene C60 with cytoprotective and cytotoxic potential: prospects as a novel treatment agent in Dermatology? Exp Dermatol. 2017 Mar;26(3):220-224. [CrossRef]
- Saitoh, Y. , Ohta, H., Hyodo, H.: Protective effects of polyvinylpyrrolidone-wrapped fullerene against intermittent ultraviolet-A irradiation-induced cell injury in HaCaT cells. J. Photochem. Photobiol. B: Biology 2016, 163: 22-29. [CrossRef]
- Prylutska, SV. , Burlaka, AP., Klymenko, PP., Grynyuk, II., Prylutskyy, YI., Schütze, C., Ritter, U.: Using water-soluble C60 fullerenes in anticancer therapy. Cancer Nanotechnol. 2011, 2:105-110. [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: Systematic review and meta-analysis. JAMA 2007, 297, 842–857. [Google Scholar] [CrossRef]
- Lee, D.-H.; Folsom, AR.; Harnack, L.; Halliwell, B.; Jacobs, DR., Jr. Does supplemental vitamin C increase cardiovascular disease risk in women with diabetes? Am. J. Clin. Nutr. 2004, 80, 1194–2000 https://expertsumnedu/en/publications/does. [Google Scholar] [CrossRef] [PubMed]
- McArdle, F. , Rhodes, L. E., Parslew, RAG., Close, GL., Jack, CIA., Friedmann, PS., Jackson, MJ.: Effects of oral vitamin E and beta-carotene supplementation on ultraviolet radiation-induced oxidative stress in human skin. Am. J. Clin. Nutr. 2004, 80, 1270–1275. [Google Scholar] [CrossRef]
- Ristow, M.; Zarse, K.; Oberbach, A.; Kloting, N.; Birringer, M.; Kiehntopf, M.; Stumvoll, M.; Kahn, C.R.; Bluher, M. Antioxidants prevent health-promoting effects of physical exercise in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 8665–8670. [Google Scholar] [CrossRef]
- Burton, GW. , Ingold, KU. : β-carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [PubMed]
- World Cancer Research Fund/American Institute for Cancer Research. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; AICR: Washington, DC, USA, 2007. [Google Scholar]
- Hibino, M.; Maeki, M.; Tokeshi, M.; Ishitsuka, Y.; Harashima, H.; Yamada, Y. : A system that delivers an antioxidant to mitochondria for the treatment of drug-induced liver injury. Nat. Sci. Rep. 2023, 13: 6961. [CrossRef]
- Epstein, JH. : Effects of β-carotene on UV-induced skin cancer formation in the hairless mouse skin. Photochem. Photobiol. 1977, 25, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, L. , Bianchi, A. , Arnaboldi, A., Andreoni, L:. Prevention of benzo(α)-pyrene photocarcinogenic effect by β-carotene and canthaxanthin. Med. Biol. Environ. 1981, 9, 113–120. [Google Scholar]
- Mathews-Roth, MM. : Antitumor activity of β-carotene, canthaxanthin, and phytoene. Oncology 1982, 38, 33–37. [Google Scholar] [CrossRef]
- Mathews-Roth, MM. , Pathak, MA.: Phytoene as a protective agent against sunburn (>280 nm) radiation in guinea pigs. Photochem. Photobiol. 1975, 21: 261–263. [CrossRef]
- Mathews, MM. , Krinsky, NI.: Carotenoids affect development of UV-B induced skin cancer. Photochem. Photobiol. 1987, 46: 507–509. [CrossRef]
- Mathews-Roth, MM. , Krinsky, NI.: Carotenoid dose level and protection against UV-B- induced skin tumors. Photochem. Photobiol. 1985, 42: 35–38. [CrossRef]
- Gensler, HL, Aickin, M., Peng, YM.: Cumulative reduction of primary skin tumor growth in UV-irradiated mice by the combination of retinyl palmitate and canthaxanthin. Cancer Lett. 1990, 53: 27–31. [CrossRef]
- Rybski, JA. , Grogan, TM., Gensler, HL: Reduction of murine cutaneous UVB-induced tumor-inflitrating T lymphocytes by dietary canthaxanthin. J. Investig. Dermatol. 1991, 9:, 892–897. [CrossRef]
- Black, HS. : Radical interception by carotenoids and effects on UV carcinogenesis. Nutr. Cancer 1998, 31, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Black, HS. , Okotie-Eboh, G., Gerguis, J.: Aspects of β-carotene-potentiated photocarcinogenesis. Photochem. Photobiol. 1999, 69: 32S.
- Nutrient Requirements of Laboratory Animals, 3rd ed.; Research Council/National Academy of Science: Washington, DC, USA, 1978; pp.
- Black, HS. , Okotie-Eboh, G., Gerguis, J.: Diet potentiates the UV-carcinogenic response to β-carotene. Nutr. Cancer 2000, 37, 173–178. [Google Scholar] [CrossRef]
- Black, HS. , Boehm, F., Edge, R., Truscott, TG.: The benefits and risks of certain dietary carotenoids that exhibit both anti- and pro-oxidative mechanisms – A comprehensive review. Antioxidants 2020, 9: 265-295. [CrossRef]
- Black, HS. , Gerguis, J. : Modulation of dietary vitamins E and C fails to ameliorate β-carotene exacerbation of UV Carcinogenesis in mice. Nutr. Cancer 2003, 45, 36–45. [Google Scholar] [CrossRef]
- Black, HS. : Pro-carcinogenic activity of β-carotene, a putative systemic photoprotectant. Photochem. Photobiol. Sci. 203, 3, 753–758. [CrossRef]
- Edge, R. , Truscott, TG. : Singlet oxygen and free radical reactions of retinoids and carotenoids—A Review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef]
- Mathews-Roth, MM. , Pathak, MA. , Parrish, J., Fitzpatrick, TB., Kass, EH., Toda, K., Clemens, W.: A clinical trial of the effects of beta-carotene on the responses of human skin to solar radiation. J. Invest. Dermatol. 1972, 59, 349–353. [Google Scholar] [CrossRef]
- Mathews-Roth, MM. : Carotenoids in erythropoietic protoporphyria and other photosensitivity diseases. Ann. N. Y. Acad. Sci. 1964, 691, 127–138. [Google Scholar] [CrossRef]
- Kune, GA. , Bannerman, S. , Field, B., Watson, LF., Cleland, H., Merenstein, D., Vitetta, L.: Diet, alcohol, smoking, serum β-carotene, and vitamin A in male nonmelanocytic skin cancer patients and controls. Nutr. Cancer 1992, 18, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, ER. , Baron, JA., Stukel, TA., Stevens, MM., Mandel, JS., Spencer, SK., Elias, PM., Lowe, N., Nierenberg, DW., Bayrd, G.; et. al.: A clinical trial of β-carotene to prevent basal-cell and squamous-cell cancer of the skin. N. Engl. J. Med. 1990, 323: 789–795. [CrossRef]
- Green, A. , Williams, G., Neale, R., Hart, V., Leslie, D., Parsons, P., Marks, GC., Gaffney, P., Battistuta, D., Frost, C., et. al.: A daily sunscreen application and beta carotene supplementation in prevention of basal-cell and squamous-cell carcinomas of the skin: A randomized trial. Lancet 1999, 354: 723–729. [CrossRef]
- Greenberg, ER. , Baron, JA., Karagas, MR., Stukel, TA., Nierenberg, DW., Stevens, MM., Mandel, JS., Haile, RW.: Mortality associated with low plasma concentration of beta carotene and the effect of oral supplementation. JAMA 1996, 275: 699–703. [CrossRef]
- Karagas, MR. , Greenberg, ER., Nierenberg, D., Stukel, TA., Morris, JS., Stevens, MM., Baron, JA.: Risk of squamous cell carcinoma of the skin in relation to plasma selenium, α-tocopherol, β-carotene, and retinol: A nested case-control study. Cancer Epidemiol. Biomark. Prev. 1997, 6: 25–29.
- Stryker, WS. , Stampfer, MJ., Stein, EA., Kaplan, L., Louis, TA., Sober, A., Willett, WC.: Diet, plasma levels of β-carotene and alpha-tocopherol, and risk of malignant melanoma. Am. J. Epidemiol. 1990, 131: 597–611. [CrossRef]
- Kiripatrick, CS. , White, E., Lee, JAH.: Case-control study of malignant melanoma in Washington State. Am.. J. Epidemiol. 1994, 139: 869–880. [CrossRef]
- Breslow, RA. , Alberg, AJ., Helzlsouer, KJ., Bush, TL., Norkus, EP., Morris, JS., Spate, VE, Comstock, GW.: Serological precursors of cancer: Malignant melanoma, basal and squamous cell skin cancer, and diagnostic levels of retinol, β-carotene, lycopene, α-tocopherol, and selenium. Cancer Epidemiol. Biomark. Prev. 1995, 4: 837–842.
- Bayerl, C. , Schwarz, B. In , Jung, EG.: Beneficial effect of β-carotene in dysplastic nevus syndrome—A randomized trial (abstract 068). In Proceedings of the 8th Congress of the European Society for Photobiology, Granada, Spain, 3–8 September 1999; p. 89. [Google Scholar]
- Druesne-Pecollo, N. , Latino-Martel, P., Norat, T., Barradon, E., Bertrais, S., Galan, P., Hercberg, S.: Beta-carotene supplementation and cancer risk: A systematic review and metanalysis of randomized controlled trials. Int..Cancer 2010, 127: 172–184. [CrossRef]
- Chen, Y-T., Kao, C-J., Huang, H-Y. Huang, S-Y., Chen, C-Y., Lin, Y-S., Wen, Z-H., Wang, H-MD.: Astaxanthin reduces MMP expressions, suppresses cancer cell migrations, and triggers apoptotic caspases of in vitro and in vivo models in melanoma. J. 2: Functional Foods 2017,31, 2017. [CrossRef]
- The α-Tocopherol, β-Carotene Cancer Prevention Study Group.: The effect of vitamin E and β-carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1996, 330: 1029–1035. [CrossRef]
- IARC Working Group on the Evaluation of Cancer-Preventive Agents. C: IARC Handbooks of Cancer Prevention, 1998.
- Gary, E. , Goodman, MD., Thornquist, JB., Cullen, MR. Meyskens, FL. Omenn, GS., Valanis,B., Williams, JH. 1: Beta-Carotene and Retinol Efficacy Trial: Incidence of Lung Cancer and Cardiovascular Disease Mortality During 6-Year Follow-up After Stopping β-Carotene and Retinol Supplements, JNCI 2004, 96, 2004. [Google Scholar] [CrossRef]
- Omenn, GS. : Chemoprevention of lung cancer: The rise and demise of beta-carotene. Ann. Rev. Public Health 1998, 19: 73–99. [CrossRef]
- Bayerl, C. : Beta-carotene in dermatology: Does it help? Acta Dermatoven APA 2008, 17, 160–166. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



