
Submitted:
31 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Background: Fibromyalgia (FM), long-lasting pain over several months, is a global medical issue with both personal and societal implications. It is one of the hardest types of pain to heal, given the lacking objective parameters for diagnosis and progression evaluation. The main symptoms of FM are long-lasting widespread pain together with anxiety, fatigue, sleep disorder, cognitive dysfunction, and obesity. Programmed cell death 1 ligand 1 (PD-L1) has been used as target in cancer immunotherapy. PD-L1 can inhibit acute and chronic pain by suppressing nociceptive neuron activity via PD-1 receptors. Methods: The current study aimed to investigate the role of PD-L1/PD1 in a mouse FM pain model. Mice were exposed to intermittent cold stress (ICS) to produce a murine FM model characterized through von Frey and Hargraves’ tests. Results: The ICS-induced mice FM pain model showed mechanical (2.26 0.18 g) and thermal (4.36 0.31 s) hyperalgesia. Nociceptive responses could be relieved by electroacupuncture (EA), intracerebral PD-L1 injection, or Trpv1 deletion. We also identified a lower PD1 level in dorsal root ganglion, spinal cord, thalamus, and somatosensory cortex. In contrast, levels of pain-related kinases increased after FM induction. Effects which could be reversed by EA, PD-L1, or Trpv1 deletion. Conclusions: Our findings shed light on the contribution of PD-L1/PD1 to EA and FM pain, indicating its potential as a treatment target for fibromyalgia.
Keywords:
Electroacupuncture
; Fibromyalgia
; PD-L1
; TRPV1
; Spinal cord
; Somatosensory cortex
1. Introduction
Programmed cell death 1 ligand 1 (PD-L1) is reported to suppress T cell function and induce immune tolerance via its receptor, PD-1 (1). Anti-PD1 and PD-L1 treatments have been used for treating neuropathic or cancer pain [1,2]. However, it is unclear whether PD-L1/PD-1 can regulate chronic pain through immune neuronal modulation. Nociceptors appear to share characteristics with immune cells, as the brain not only responds to cytokines but can also regulate neural function through cell receptors. PD-L1/PD-1 is crucial as an immune mediator in the brain but also in chronic pain [1,2]. Current treatment approaches for fibromyalgia (FM) face multiple problems, ranging from cost efficacy to sustainability and target specificity. Furthermore, current clinical applications for general pain management include surgery, opioid receptors, non-steroidal anti-inflammatory drugs (NSAIDS), COX-1 and COX-2 inhibitors, and sodium channel blockers. Although the financial limitations, availability and accessibility are all valid concerns alongside the plethora of side effects resultant from these treatments [3]. It remains unclear whether neurons express functional PD-1 receptors in chronic pain, especially in relationship to electroacupuncture (EA).
Transient receptor potential V1 (TRPV1) has been implicated in chronic pain, inflammation, cancer, and immunity [4,5]. TRPV1 was used as drug target for various pain conditions. And target for TRPV1 structure, agonists, and mechanisms is urgent [5,6]. TRPV1 activation mediate the contribution of phosphorylated phosphoinositide 3-kinase (pPI3K), phosphorylated Akt (pAkt), and phosphorylated mammalian target of rapamycin (pmTOR) in pain modulation, indicating a crucial role in central sensitization associated with FM pain at both peripheral and central nervous systems [7,8,9,10]. Further, the epsilon isoform of protein kinase C (PKCɛ) has been considered as a transient insult that reliably delivers hyperalgesia to the nociceptors, resulting in long-lasting central sensitization, an essential factor in FM pain. Mitogen-activated protein kinase (MAPK) is reportedly involved in inflammation and pain signaling in a pathway involving extracellular signal-regulated protein kinase, protein kinase p38 (p38), and c-Jun N-terminal kinase/stress-activated protein kinase (JNK) [11,12]. All these elements increase their levels during nociception, a process associated with painful sensation, neuronal plasticity, central sensitization and function ability of certain cognitive aptitudes [13,14]. PI3K-Akt-mTOR signaling appears to be associated with changes in the nociceptive response at both peripheral and central levels for its role in central sensitization. Moreover, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) has been involved in FM pain development; suppressing its expression has been associated with therapeutic benefits in pain [15,16].
FM pain is highly related to physiological and psychological issues. FM is a chronic pain disease representative of a serious growing issue including subjects, healthcare, economic and society problem. FM is characterized by a chronic widespread pain in the body. FM pain is frequently experienced in the arms, legs, head, chest, abdomen, and back. FM patients usually describe it as painful, burning, or soreness. The incident of FM is about 2–8% of the population, occurring often in women who suffer from tension headaches, irritable bowel syndrome, anxiety and/or depression. Further, women show lower pain thresholds and more severe symptoms than men [17,18]. Currently, there is no cure for FM pain, medications can only manage symptoms. In addition, nutrition, exercise, meditation, sleep therapy, yoga, and relaxation may help. FM may result from repeated nerve stimulation central sensitization, and/or overexpression of neurotransmitters or neuromodulators in the brain level. Its pathogenesis is not clear, although peripheral (inflammatory) and central (cognitive–emotional) mechanisms appear to interact resulting in impaired pain perception [19,20]. The economic impact of FM is high and the precise cost drivers of health care spending for symptoms. On the other hand, inflammatory cytokines could disrupt neural networks during the interaction of the nervous system with immune cells, causing increased central and peripheral sensitization as well as neuroinflammation [21,22,23].
Acupuncture, a main component of traditional Chinese medicine (TCM), consists of inserting a steel needle into specific local acupoints as is reported to relieve many diseases and disorders, being especially useful in pain management. TCM describes acupuncture as a practice for balancing the chi to flow through the body meridians to improve health. In contrast, in Western medicine, acupuncture is supposed to stimulate peripheral nerves via connective tissue and muscle to manage symptoms. EA on the ST36 acupoint could activate the vagal-adrenal axis in mice via optogenetic stimulation of nerve terminals [24], while another report described a novel anti-inflammatory effect of EA in a mouse sepsis model [25]. We previously demonstrated that EA can trigger the release of adenosine triphosphate, interleukin-1, interleukin-6, glutamate, substance P, and histamine at local acupoints [26]. Further, EA has been reported to relieve Parkinson’s disease [27,28], manage body weight [29,30], decrease several pain syndromes in mice models, possibly by decreasing inflammatory cytokines such as interleukins, TNF-α, and IFN-γ in mouse plasma [16,31,32].
Based on the above, in the current study we aimed to verify the hypothesis that FM pain is associated with altered PD-L1/PD1 and related molecules in a mouse FM model. In addition, we wanted to evaluate whether EA’s analgesic effect in both mechanical and thermal hyperalgesia is mediated by PD-L1 signaling. To address this issue, we developed a mouse model of FM pain by intermittent cold stress (ICS). Consistent with our hypothesis, ICS caused significant mechanical and thermal hyperalgesia, which was attenuated by EA. PD-L1/PD1 and associated molecules reliably changed in ICS mice, being reversed by EA. Our data postulates that the analgesic effect of EA is related to PD-L1/PD1 in the dorsal root ganglion (DRG), spinal cord (SC), thalamus, and somatosensory cortex (SSC). We bring solid recommendation that EA modulate the PD-L1/PD1 signaling pathway, thus signifying new probable beneficial goals for treating FM pain.
2. Materials and Methods
Mice and FM Pain Initiation
For the experiments, we used 8–12-week-old female C57B/L6 wild type mice (18 –20 g) sourced from BioLasc Taiwan Ltd (Yilan, Taiwan) and housed in a specific pathogen-free environment. After their arrival, mice were housed in cages under a 12-h light/dark cycle (light from 6 a.m. to 6 p.m.), at a room temperature of 25ºC with 60% humidity. We performed statistical analysis to estimate the sample size; nine mice per group was considered as the minimum number required for a significance α level of 0.05 and a power of 80%. Animal experiments were approved by the Institute of Animal Care and Use Committee of China Medical University (Permit no. CMUIACUC-2023-071), Taiwan, following the Guide for the use of Laboratory Animals (National Academy Press). Mice were randomly divided into four groups: normal (Normal), cold stress-induced FM, cold stress-induced FM with EA (FM+EA), cold stress-induced FM with PD-L1 intracerebral injection (FM+PD-L1), and cold stress-induced FM in Trpv1-/- mice (FM+ Trpv1-/-). To create the FM model, mice were placed at 4ºC, while the normal group was consistently maintained at 25ºC. At 10 a.m. the following day, FM mice were moved to 25ºC for 30 min before returning to 4ºC for another 30 min. This process was performed for 6 h until 4 p.m., before they were replaced again overnight from 4 p.m. during the first 3 days.
Electroacupuncture
Mice were anesthetized with 5% isoflurane for induction and maintained at 1% isoflurane for inhalation. Dual 1” steel acupuncture needles (32G, Yu Kuang Chem. Ind. Corp., Taiwan) were implanted bilaterally into the mouse ST36 acupoint, located 3–4 mm under the patella, between the fibula and tibia, on the anterior side of the anterior tibial muscle. We delivered 1 mA strength, 2 Hz frequency, and 150μs continuous square pulses for 20 min using an electronic Trio 300 stimulator (Ito, Japan). EA produced minor observable muscle twitching around the acupoint. EA was performed thrice from day 3 to 4 after ICS.
Nociceptive Behavior Tests
Mechanical and thermal pain were measured three times at day 0, day 3, and day 4 before and after ICS. The pain threshold on the day before ICS induction was considered as baseline. Mice were first placed in plexiglass boxes directly above a steel mesh in a dark, noiseless, room temperature to keep animals calm down and to adjust to the new environment. When mice were not moving, standing, sleeping, scratching, or grooming, the von Frey filament test was performed three times per session at 10-min intervals (IITC Life Science Inc., USA). Next, the Hargreaves’ tests was performed to measure thermal latency. The test started after 30 min of acclimatation. An IITC Plantar Analgesia Meter (IITC Life, Sciences, SERIES8, Model 390G) determined the withdrawal latency of the mouse foot over radiant thermal light. The device was set to self-cut at 20 s to avoid damaging the hindpaws.
Western Blot
The DRG, SC, thalamus, and SSC were excised to extract proteins. Tissues were placed on ice and stored at −80ºC before extracting proteins. Proteins were incubated in cold radioimmunoprecipitation (RIPA) lysis buffer (50 mM Tris-HCl pH 7.4, 250 mM NaCl, 1% NP-40, 5 mM EDTA, 50 mM NaF, 1 mM Na3VO4, 0.02% NaN3, and 1× protease inhibitor cocktail [AMRESCO]). The extracted proteins were subjected to 8% SDS-Tris glycine gel electrophoresis and transmitted to a polyvinylidene difluoride (PVDF) membrane. The PVDF was incubated in 5% non-fat milk in TBS-T buffer (10 mM Tris pH 7.5, 100 mM NaCl, 0.1% Tween 20) and a primary antibody in TBS-T with 1% bovine serum albumin (BSA) for 1 h at room temperature. Then, a peroxidase-conjugated anti-rabbit antibody or anti-mouse antibody (1: 5000) was used as secondary antibody. Blot bands were imagined by a chemiluminescent substrate kit (PIERCE) and LAS-3000 Fujifilm (Fuji Photo Film Co., Ltd, Tokyo, Japan). The protein concentration of the bands was measured using NIH Image J 1.54h software (Bethesda, MD, USA). β-actin or α-tubulin were used as internal controls.
Immunofluorescence
Mice were anesthetized with 5% isoflurane and intracardially injected with 0.9% normal saline followed by 4% paraformaldehyde. Tissues were instantly excised and fixed with 4% paraformaldehyde at 4ºC for 3 days. Then, samples were incubated in 30% sucrose for post cryoprotection overnight at 4ºC, before fixating in an optimal cutting temperature complex and being quickly frozen in liquid nitrogen before storage at −80ºC. Frozen tissues were then cut into 20 mm sections using a cryostat and directly placed on glass slides. The sections were fixed with 4% paraformaldehyde, and incubated with a blocking solution consisting of 3% BSA, 0.1% Triton X-100, and 0.02% sodium azide for 1 h at room temperature. The samples were then incubated with the primary antibody (1:200, Alomone), PD-1 and TRPV1 prepared in 1% BSA solution at overnight. Then, samples were incubated with the secondary antibody (1:500) 488-conjugated AffiniPure donkey anti-rabbit IgG (H + L) and 594-conjugated AffiniPure donkey anti-goat IgG (H + L) for 2 h at room temperature before fixation with coverslips for immunofluorescence visualization.
Intracerebroventricular Injection
Mice were anesthetized with isoflurane with immobilized heads in a stereotaxic frame to get a cannula implanted at the ventricle. The stereotaxic cannula (23 gauge 2 mm stainless steel) was then placed at 0.5 mm in the anteroposterior axis, around 1 mm in the mediolateral axis, and about 2.5 mm in the dorsoventral axis under the cortical surface and fixed at the skull with dental cement. Afterwards, the cannula was inserted and connected to a Hamilton syringe through a PE tube (PE10, Portex, Kent, UK). A total of 5 μg of PD-L1 (5 μL/ventricle) was injected over a period of 5 min using a syringe pump (KD Scientific, Shanghai, China). After inoculation, the cannula was left at the ventricle for an extra 2 min to allow the PD-L1 to diffuse.
Statistical Analysis
Statistical analysis was performed using SPSS. All statistic data are presented as the mean ± standard error (SEM). Statistical differences among groups were tested using an ANOVA test, followed by a post hoc Tukey’s test. P < 0.05 was considered to indicate statistical significance.
3. Results
EA, PD-L1 Injection, or Trpv1 Deletion Care for FM Pain in a Mouse Model
We induced FM pain in WT and Trpv1−/− mice through ICS induction. In normal mice, mechanical threshold and thermal latency were normal on day 3 to 4 (Figure 1A). Due to ICS, mechanical and thermal hyperalgesia increased, being maintained for 2 days as indicated by von Frey and Hargraves’ tests (Figure 1A and B, red circles, 2.26 ± 0.18 g and 4.36 ± 0.31 s, *P < 0.05, n = 9). Treatment with 2 Hz EA significantly attenuated both mechanical and thermal hyperalgesia (Figure 1A and B, blue circles, 3.81 ± 0.28 g and 7.38 ± 0.48 s, #P < 0.05, n = 9). Intracerebral PD-L1 injections relieved mechanical and thermal hyperalgesia (Figure 1A and B, green circles, 3.61 ± 0.26 g and 6.08 ± 0.31 s, #P < 0.05, n = 9). TRPV1 loss improved both mechanical and thermal hyperalgesia on day 4 after ICS induction (Figure 1A and B, gray circles, 3.66 ± 0.22 g and 7.48 ± 0.48 s, #P < 0.05, n = 9).
EA Controlled PD1-TRPV1 Pain Signaling in the Peripheral DRG of FM Mice
Initially, we aimed to determine whether PD1-TRPV1 affect FM pain in peripheral DRG areas. We used Western blot to define the valence associated with PD1-TRPV1 modulation. PD1 was expressed in the DRG, but its level was reduced with FM (Figure 2A, 65.91 ± 6.23%, black column, *P < 0.05, n = 6). EA significantly augmented the reduction of PD1 in the mouse DRG after ICS induction (Figure 2A, 97.99 ± 5.11%, black column, #P < 0.05, n = 6). Intracerebral PD-L1 administration did not change expression of the PD1 receptor in the DRG (Figure 2A, 68.98 ± 3.46%, black column, *P < 0.05, n = 6). To establish a relationship between PD1 and TRPV1, we subjected Trpv1−/− mice to ICS induction. Our results indicate that PD1 receptors return to normal levels in FM model mice (Figure 2A, 98.82 ± 3.60%, black column, #P < 0.05, n = 6). TRPV1 was present in the DRG, increasing with FM pain (Figure 2A, 133.89 ± 4.63%, red column, *P < 0.05, n = 6), while 2 Hz EA inhibited such overexpression (Figure 2A, 102.93 ± 2.57%, red column, #P < 0.05, n = 6). Further, TRPV1 expression in the DRG was not attenuated by PD-L1 injection (Figure 2A, 133.46 ± 2.36%, red column, *P < 0.05, n = 6), being almost lost in Trpv1−/− mice (Figure 2A, 16.47 ± 2.06%, red column, #P < 0.05, n = 6). Next, we verified the expression of associated nociceptive ion channels such as Nav1.7 and Nav1.8, potential targets for drug development in pain. Their protein levels increased with ICS and could be alleviated by 2 Hz EA. PD-L1 administration similarly attenuated Nav1.7 and Nav1.8 overexpression. Similar results were observed in Trpv1−/− mice under ICS (Figure 2A, gray and yellow columns, *P < 0.05, n = 6).
We next aimed to identify TRPV1-related kinases in this model, namely components of the pPI3K-pAkt-pmTOR axis. As shown in Figure 2B, the expression of pPI3K-pAkt-pmTOR increased in the DRG of FM mice. Western blot results indicated that those protein levels were down-regulated after EA treatment. Similar results were observed with PD-L1 injection and in Trpv1−/− mice (Figure 2B, individual column, *P < 0.05, n = 6). A noticeably higher expression of MAPK (pERK, pp38, and pJNK) kinases was observed in the DRG of FM mice compared to the normal group, an effect inhibited by 2 Hz EA treatment. Similar results were obtained after PD-L1 injection and in Trpv1−/− mice. Moreover, the expression of pNKκB and pCREB increased in the DRG of FM mice compared to normal mice. 2 Hz EA, PD-L1 injection and Trpv1 deletion all reversed these changes (Figure 2B & C, individual column, *P < 0.05, n = 6).
EA at ST36 Diminished Cold Stress-Induced FM Pain Through TRPV1-CB1 in the SCDH
To gain insights into the role of PD1/PD-L1 in the SC of FM mice, we applied Western blot to samples obtained at 2 days after ICS induction, the subacute stage of pain. PD-1 expression was normal in the normal group but significantly lower in the SCDH of FM mice (Figure 3A, 78.64 ± 2.11%, black column, *P < 0.05, n = 6), a decrease deepened by 2 Hz EA treatment (Figure 3A, 104.69 ± 4.80%, black column, #P < 0.05, n = 6). This pattern was also obtained with intracerebral PD-L1 injection (Figure 3A, 104.14 ± 5.67%, black column, #P < 0.05, n = 6) and in Trpv1−/− mice (Figure 3A, 102.75 ± 5.43%, black column, #P < 0.05, n = 6). Interestingly, TRPV1 increased after FM induction compared to controls (Figure 3A, 134.21 ± 2.45%, red column, *P < 0.05, n = 6). 2 Hz EA not only relieve FM pain but decreased TRPV1 levels in the SCDH (Figure 3A, 100.13 ± 3.32%, red column, #P < 0.05, n = 6), as did PD-L1 injection (Figure 3A, 101.16 ± 2.95%, red column, #P < 0.05, n = 6) and disappeared in Trpv1 knock out (Figure 3A, 13.66 ± 1.09%, red column, #P < 0.05, n = 6). The levels of nociceptive ion channels Nav1.7 and Nav1.8 increased after FM induction, being decreased by 2 Hz EA, PD-L1 administration and in Trpv1−/− mice (Figure 3A, gray and yellow columns, *P < 0.05, n = 6).
With respect to pPI3K, pAkt, and pmTOR, their levels were increased in the SC of FM mice but decreased by 2 Hz EA treatment, PD-L1 injection and in Trpv1−/− mice (Figure 3B, *P < 0.05, n = 6). Similar results were obtained for pERK, pp38, and pJNK. pNKκB and pCREB levels were increased in the SC of FM mice but decreased with 2 Hz EA, PD-L1 injection and Trpv1 knock out (Figure 3B & C, *P < 0.05, n = 6).
EA at ST36 Altered FM Pain and Regulated PD-1 Signaling Pathway in the Thalamus
To observe the effects of PD-L1/PD1 on FM mice, we measured the expression levels of PD1 in the ascending pain pathway thalamus. After completing behavioral tests on day 4, we collected the thalamus and calculated the expression levels of previously mentioned proteins via Western blot. PD-1 levels were consistently abridged in thalamus of FM mice (Figure 4A, 79.07 ± 0.88%, black column, *P < 0.05, n = 6) and 2 Hz EA effectively increased them (Figure 4A, 100.83 ± 3.26%, black column, #P < 0.05, n = 6), as did intracerebral injection of PD-L1 (Figure 4A, 101.15 ± 3.39%, black column, #P < 0.05, n = 6). Conversely, the reduction in PD1 level was not observed in the thalamus of Trpv1−/− mice (Figure 4A, 97.55 ± 3.04%, black column, #P < 0.05, n = 6). Consistent with earlier findings, TRPV1 levels were increased at day 4 after ICS induction in the thalamus of FM mice compared with the normal group (Figure 4A, 132.78 ± 2.47%, black column, *P < 0.05, n = 6). Mice treated with 2 Hz EA showed decreased TRPV1 levels in the thalamus (Figure 4A, 100.09 ± 1.49%, red column, #P < 0.05, n = 6). Similar trends were also obtained with intracerebral injection of PD-L1 (Figure 4A, 100.21 ± 1.72%, red column, #P < 0.05, n = 6) and disappearance in Trpv1−/− (Figure 4A, 15.97 ± 1.06%, red column, #P < 0.05, n = 6) mice as well as for Nav1.7 and Nav1.8 (Figure 4A, gray and yellow columns, *P < 0.05, n = 6).
Similarly, pPI3K, pAkt, and pmTOR levels increased after ICS, but decreased with 2 Hz EA, PD-L1 injection, and Trpv1 knock out (Figure 4B, *P < 0.05, n = 6). FM pain also increased the expression of pERK, pp38, and pJNK in the thalamus suggesting the vital function in ascending pain signaling. 2Hz EA treatment demonstrated a key therapeutic effect on FM pain through regulating these kinases. The same trends were observed after intracerebral PD-L1 injection and Trpv1 knock out. Likewise, ICS resulted in of pNFκB and pCREB overexpression, an effect reverted by 2 Hz EA, PD-L1 injection, and Trpv1 knock out (Figure 4B & C, *P < 0.05, n = 6).
TRPV1 and Related Factors were Inhibited in the SSC after ICS and Recovered by 2 Hz EA and Trpv1 Deletion
Finally, to see whether there is a difference of PD1 to decide the influence of ICS-induced FM pain. We also want to identify if EA could affect FM pain signaling in the ascending pathway, we dissevered the mice SSC for protein analysis. ICS significantly abridged the expression of PD1 in the mice SSC compared with normal mice (Figure 5A, 72.70 ± 3.25%, black column, *P < 0.05, n = 6). This diminution was reverted by 2 Hz EA (Figure 5A, 102.16 ± 2.42%, black column, #P < 0.05, n = 6), PD-L1 injection (Figure 5A, 92.94 ± 5.38%, black column, #P < 0.05, n = 6) and Trpv1 deletion (Figure 5A, 100.87 ± 3.64%, black column, #P < 0.05, n = 6). Compared to controls, a significant increase in TRPV1, Nav1.7, and Nav1.8 levels was observed in FM mice. This increase was reverted by 2 Hz EA, intracerebral PD-L1 injection and Trpv1 deletion (Figure 5A, *P < 0.05, n = 6). Similarly, pPI3K, pAkt, and pmTOR expression increased in FM mice compared to normal mice, getting significantly decreased by 2 Hz EA, intracerebral injection of PD-L1, and Trpv1 deletion. Similar were obtained for pERK, pp38, pJNK, pNFκB, and pCREB (Figure 5B & C, *P < 0.05, n = 6).
EA, Intracerebral PD-L1 Injection or TRPV1 Loss Mitigated FM in the DRG or SSC
Immunofluorescence staining of the DRG for PD-1 showed lower PD-1 expression in the DRG of FM mice than in normal mice. An effect reverted by 2 Hz EA, intracerebral injection of PD-L1 or Trpv1 knock out. In contrast, TRPV1 protein levels were augmented in FM mice being attenuated by 2 Hz EA, PD-L1 injection or TRPV1 loss (Figure 6A, yellow arrows, n = 3). Finally, we confirmed that PD-1 levels were decreased after ICS in the SSC, but increased after EA treatment, PD-L1 injection or Trpv1 deletion. Conversely, TRPV1 levels increased after FM induction and were decreased by EA, PD-L1 injection or Trpv1 deletion (Figure 6B, yellow arrows, n = 3).
4. Discussion
To our knowledge, this is the first comprehensive study on the relationship between EA, FM pain, and PD-L1/PD-1 signaling. ICS-induced FM resulted in mechanical and thermal pain as indicated by von Frey and Hargraves’ tests in mice; an effect that can be reversed with 2 Hz EA, PD-L1 injection, or Trpv1 deletion. We also found that PD-L1/PD-1 signaling was attenuated in the peripheral DRG as well as in the central SCDH, thalamus, and SSC. We also demonstrated that levels of nociceptive TRPV1 and related molecules were increased after FM induction. These tendencies could be reversed by 2 Hz EA, PD-L1 injection, or Trpv1 deletion. However, intracerebral ventricular injection of PD-L1 could not attenuate TRPV1 overexpression at peripheral DRG sites suggesting its advanced and subsequent levels. The turnover of these protein alterations in signaling protein kinases through the use of EA suggests that these managements, especially its relationship with PD-L1/PD-1, can be used as an efficient beneficial intervention.
PD-L1 had been characteristically secreted by cancer cells and inhibited immune function via PD-1 expressed on T cells. Chen et al. demonstrated that in the DRG PD-L1 can effectively prevent acute and chronic pain. An intraplantar PD-L1 shot induced anti-nociception in normal mice via PD-1. In addition, blockage of PD-L1 talk into mechanical allodynia. Gene deletion of Pd1 displayed mechanical and thermal hypersensitivity. Further, PD-1 activation in DRG nociceptive neurons was reported to induce SHP-1 phosphorylation to activate TREK2 K+ channels for pain relief [32]. Recent articles reported that PD-1 is expressed in DRG neurons and that after binding to PD-L1 it can prevent inflammatory and neuropathic pain by decreasing neuronal excitability. Liu et al. demonstrated that PD-L1 can stimulate Src homology 2 domain-containing tyrosine phosphatase-1 (SHP-1) to decrease TRPV1 expression in the DRG, relieving mouse bone cancer pain. Moreover, local PD-L1 injection significantly initiated analgesia through SHP-1 phosphorylation in TRPV1 neurons, decreasing their activation and showed PD-1 colocalization with TRPV1 and SHP-1 in mouse DRG neurons. In addition, TRPV1 blockade reliably attenuated cancer pain-related hyperalgesia and PD-L1 analgesia, suggesting a crucial role of TRPV1 in PD-L1 signaling. They also reported that a conditioned SHP-1 knock out increased bone cancer pain and reduced PD-L1 inhibition on TRPV1 in peripheral DRG neurons [33].
Spinal cord injury (SCI) is a major traumatic event that can cause persistent central neuropathic pain affecting motor dysfunction and often ineffectively alleviated. Kong et al. demonstrated that PD-L1 was increased after SCI on microglia or macrophages at the injury site. Loss of PD-L1 in mice caused poorer quality of motor function and nociception with increased polarization of M1-like macrophages. They next determined that PD-L1’s effects after SCI occur by attenuating phosphorylation of p38 and pERK2 [34]. Shi et al. reported a crucial role of PD-L1/PD-1 in the trigeminal ganglia in a mouse migraine model, after showing PD-L1/PD-1 immunoreactivity in normal trigeminal ganglia neurons. The mRNA levels of PD-L1/PD-1 were meaningfully increased after migraine induction. In addition, PD-L1/PD-1 increased after migraine initiation. Inhibition of PD-1 amplified migraine-induced hyperalgesia accompanied by increased calcitonin gene-related peptide, interleukin 1β (IL-1β), Tumor necrosis factor α (TNF-α), IL-6 and IL-18 in the mice trigeminal ganglia [35].
As we know, the maternal pain threshold steadily rises throughout pregnancy, particularly in late pregnancy. Tan et al. informed the mechanical and thermal onsets were innocently increased in late pregnancy. They found that the level of PD-L1 was increased in late pregnancy. In addition, after giving soluble PD-1 can significantly decrease the pain thresholds of late pregnancy mice. Moreover, IL-10 was visibly increased, and TNF-α and IL-6 were attenuated in late pregnancy mice, suggesting the crucial role of PD-L1/PD-1 pathway in pregnancy-induced analgesia [36]. Anti-nociceptive peptides have been developed as therapeutic methods without opioid-associated side effects lacking effective and safe chronic pain medicines. Recently, PD-L1/PD-1 activation attenuated nociceptor excitability and induces important pain-relieving effects suggesting its beneficial target for pain management. Zhao et al. revealed that H-20 can bind to PD-1 with micromolar affinity and later initiate SHP-1 phosphorylation in DRG neurons. Spinal H-20 administration showed actual and longer analgesic effect in preclinical models without several side effects [37]
5. Conclusions
Our experiments provide evidence in necessary possessions of 2 Hz EA in mice FM pain, especially via PD-L1/PD-1 and TRPV1 signaling pathway. We demonstrated the analgesic effect of EA on mouse FM pain and its underlying mechanisms. We found that TRPV1 and related kinases were overexpressed in a mouse model of FM pain in the DRG, SCDH, thalamus, and SSC. These effects were abolished by 2 Hz EA, PD-L1 injection, or Trpv1 deletion. However, intracerebroventricular PD-L1 injection did not reduce TRPV1 overexpression at peripheral DRG sites suggesting an ineffective long site effect. Our results support the clinical application of EA in FM pain. A schematic drawing of our findings is offered in Figure 7.
Author Contributions
HC Lin, HC Hsu, and HY Liao: Conceptualization, Methodology, Software, Data curation, Writing - original draft, Visualization, Investigation. Arbee LPC and YW Lin: Supervision, Validation, Writing - review & editing.
Funding
This work was supported by the following grants: NSTC 113-2314-B-039-047, CMU113-ASIA-15, CMUHCH-DRM-113-025, and the "Chinese Medicine Research Center, China Medical University" from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education in Taiwan.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article.
Conflicts of Interest
There are no financial or other relationships that might lead to a conflict of interest for all authors.
References
- Wang, G.; Kang, X.; Chen, K. S.; Jehng, T.; Jones, L.; Chen, J.; Huang, X. F.; Chen, S. Y., An engineered oncolytic virus expressing PD-L1 inhibitors activates tumor neoantigen-specific T cell responses. Nat Commun 2020, 11, (1), 1395. [CrossRef]
- Chen, G.; Kim, Y. H.; Li, H.; Luo, H.; Liu, D. L.; Zhang, Z. J.; Lay, M.; Chang, W.; Zhang, Y. Q.; Ji, R. R., PD-L1 inhibits acute and chronic pain by suppressing nociceptive neuron activity via PD-1. Nat Neurosci 2017, 20, (7), 917-926. [CrossRef]
- Brown, E. N.; Pavone, K. J.; Naranjo, M., Multimodal General Anesthesia: Theory and Practice. Anesth Analg 2018, 127, (5), 1246-1258. [CrossRef]
- Benitez-Angeles, M.; Morales-Lazaro, S. L.; Juarez-Gonzalez, E.; Rosenbaum, T., TRPV1: Structure, Endogenous Agonists, and Mechanisms. Int J Mol Sci 2020, 21, (10). [CrossRef]
- Koivisto, A. P.; Voets, T.; Iadarola, M. J.; Szallasi, A., Targeting TRP channels for pain relief: A review of current evidence from bench to bedside. Curr Opin Pharmacol 2024, 75, 102447. [CrossRef]
- Fernandez-Carvajal, A.; Fernandez-Ballester, G.; Ferrer-Montiel, A., TRPV1 in chronic pruritus and pain: Soft modulation as a therapeutic strategy. Front Mol Neurosci 2022, 15, 930964. [CrossRef]
- Inprasit, C.; Lin, Y. W., TRPV1 Responses in the Cerebellum Lobules V, VIa and VII Using Electroacupuncture Treatment for Inflammatory Hyperalgesia in Murine Model. Int J Mol Sci 2020, 21, (9). [CrossRef]
- Lottering, B.; Lin, Y. W., TRPV1 Responses in the Cerebellum Lobules VI, VII, VIII Using Electroacupuncture Treatment for Chronic Pain and Depression Comorbidity in a Murine Model. Int J Mol Sci 2021, 22, (9). [CrossRef]
- Hsiao, I. H.; Lin, Y. W., Electroacupuncture Reduces Fibromyalgia Pain by Attenuating the HMGB1, S100B, and TRPV1 Signalling Pathways in the Mouse Brain. Evid Based Complement Alternat Med 2022, 2022, 2242074. [CrossRef]
- Liao, H. Y.; Lin, Y. W., Electroacupuncture reduces cold stress-induced pain through microglial inactivation and transient receptor potential V1 in mice. Chin Med 2021, 16, (1), 43. [CrossRef]
- Hsu, H. C.; Hsieh, C. L.; Wu, S. Y.; Lin, Y. W., Toll-like receptor 2 plays an essential role in electroacupuncture analgesia in a mouse model of inflammatory pain. Acupunct Med 2019, 37, (6), 356-364. [CrossRef]
- Hsu, H. C.; Hsieh, C. L.; Lee, K. T.; Lin, Y. W., Electroacupuncture reduces fibromyalgia pain by downregulating the TRPV1-pERK signalling pathway in the mouse brain. Acupunct Med 2020, 38, (2), 101-108. [CrossRef]
- Liao, H. Y.; Lin, Y. W., Electroacupuncture Attenuates Chronic Inflammatory Pain and Depression Comorbidity through Transient Receptor Potential V1 in the Brain. Am J Chin Med 2021, 49, (6), 1417-1435. [CrossRef]
- Yang, J.; Hsieh, C. L.; Lin, Y. W., Role of Transient Receptor Potential Vanilloid 1 in Electroacupuncture Analgesia on Chronic Inflammatory Pain in Mice. Biomed Res Int 2017, 2017, 5068347. [CrossRef]
- Lottering, B.; Lin, Y. W., Functional characterization of nociceptive mechanisms involved in fibromyalgia and electroacupuncture. Brain Res 2021, 1755, 147260. [CrossRef]
- Lin, Y. W.; Chou, A. I. W.; Su, H.; Su, K. P., Transient receptor potential V1 (TRPV1) modulates the therapeutic effects for comorbidity of pain and depression: The common molecular implication for electroacupuncture and omega-3 polyunsaturated fatty acids. Brain Behav Immun 2020, 89, 604-614. [CrossRef]
- Siracusa, R.; Paola, R. D.; Cuzzocrea, S.; Impellizzeri, D., Fibromyalgia: Pathogenesis, Mechanisms, Diagnosis and Treatment Options Update. Int J Mol Sci 2021, 22, (8). [CrossRef]
- Janssen, L. P.; Medeiros, L. F.; Souza, A.; Silva, J. D., Fibromyalgia: A Review of Related Polymorphisms and Clinical Relevance. An Acad Bras Cienc 2021, 93, (suppl 4), e20210618. [CrossRef]
- Assavarittirong, C.; Samborski, W.; Grygiel-Gorniak, B., Oxidative Stress in Fibromyalgia: From Pathology to Treatment. Oxid Med Cell Longev 2022, 2022, 1582432. [CrossRef]
- Farag, H. M.; Yunusa, I.; Goswami, H.; Sultan, I.; Doucette, J. A.; Eguale, T., Comparison of Amitriptyline and US Food and Drug Administration-Approved Treatments for Fibromyalgia: A Systematic Review and Network Meta-analysis. JAMA Netw Open 2022, 5, (5), e2212939. [CrossRef]
- Peck, M. M.; Maram, R.; Mohamed, A.; Ochoa Crespo, D.; Kaur, G.; Ashraf, I.; Malik, B. H., The Influence of Pro-inflammatory Cytokines and Genetic Variants in the Development of Fibromyalgia: A Traditional Review. Cureus 2020, 12, (9), e10276. [CrossRef]
- Menzies, V.; Lyon, D. E., Integrated review of the association of cytokines with fibromyalgia and fibromyalgia core symptoms. Biol Res Nurs 2010, 11, (4), 387-94. [CrossRef]
- Ghowsi, M.; Qalekhani, F.; Farzaei, M.H.; Mahmudii, F.; Yousofvand, N.; Joshi, T. Inflammation, oxidative stress, insulin resistance, and hypertension as mediators for adverse effects of obesity on the brain: A review. Biomedicine 2021, 11, 13–22. [CrossRef]
- Liu, S.; Wang, Z.; Su, Y.; Qi, L.; Yang, W.; Fu, M.; Jing, X.; Wang, Y.; Ma, Q., A neuroanatomical basis for electroacupuncture to drive the vagal-adrenal axis. Nature 2021, 598, (7882), 641-645. [CrossRef]
- Torres-Rosas, R.; Yehia, G.; Pena, G.; Mishra, P.; del Rocio Thompson-Bonilla, M.; Moreno-Eutimio, M. A.; Arriaga-Pizano, L. A.; Isibasi, A.; Ulloa, L., Dopamine mediates vagal modulation of the immune system by electroacupuncture. Nat Med 2014, 20, (3), 291-5. [CrossRef]
- Hsiao, I. H.; Liao, H. Y.; Cheng, C. M.; Yen, C. M.; Lin, Y. W., Paper-Based Detection Device for Microenvironment Examination: Measuring Neurotransmitters and Cytokines in the Mice Acupoint. Cells 2022, 11, (18). [CrossRef]
- Tsai, S. T.; Wei, T. H.; Yang, Y. W.; Lu, M. K.; San, S.; Tsai, C. H.; Lin, Y. W., Transient receptor potential V1 modulates neuroinflammation in Parkinson's disease dementia: Molecular implications for electroacupuncture and rivastigmine. Iran J Basic Med Sci 2021, 24, (10), 1336-1345.
- Lu, K. W.; Yang, J.; Hsieh, C. L.; Hsu, Y. C.; Lin, Y. W., Electroacupuncture restores spatial learning and downregulates phosphorylated N-methyl-D-aspartate receptors in a mouse model of Parkinson's disease. Acupunct Med 2017, 35, (2), 133-141. [CrossRef]
- Inprasit, C.; Huang, Y. C.; Lin, Y. W., Evidence for acupoint catgut embedding treatment and TRPV1 gene deletion increasing weight control in murine model. Int J Mol Med 2020, 45, (3), 779-792. [CrossRef]
- Choowanthanapakorn, M.; Lu, K. W.; Yang, J.; Hsieh, C. L.; Lin, Y. W., Targeting TRPV1 for Body Weight Control using TRPV1(-/-) Mice and Electroacupuncture. Sci Rep 2015, 5, 17366. [CrossRef]
- Yen, L. T.; Hsieh, C. L.; Hsu, H. C.; Lin, Y. W., Targeting ASIC3 for Relieving Mice Fibromyalgia Pain: Roles of Electroacupuncture, Opioid, and Adenosine. Sci Rep 2017, 7, 46663. [CrossRef]
- Liao, H. Y.; Hsieh, C. L.; Huang, C. P.; Lin, Y. W., Electroacupuncture Attenuates Induction of Inflammatory Pain by Regulating Opioid and Adenosine Pathways in Mice. Sci Rep 2017, 7, (1), 15679. [CrossRef]
- Liu, B. L.; Cao, Q. L.; Zhao, X.; Liu, H. Z.; Zhang, Y. Q., Inhibition of TRPV1 by SHP-1 in nociceptive primary sensory neurons is critical in PD-L1 analgesia. JCI Insight 2020, 5, (20). [CrossRef]
- Kong, F.; Sun, K.; Zhu, J.; Li, F.; Lin, F.; Sun, X.; Luo, X.; Ren, C.; Lu, L.; Zhao, S.; Sun, J.; Wang, Y.; Shi, J., PD-L1 Improves Motor Function and Alleviates Neuropathic Pain in Male Mice After Spinal Cord Injury by Inhibiting MAPK Pathway. Front Immunol 2021, 12, 670646. [CrossRef]
- Shi, S.; Han, Y.; Wang, D.; Guo, P.; Wang, J.; Ren, T.; Wang, W., PD-L1 and PD-1 expressed in trigeminal ganglia may inhibit pain in an acute migraine model. Cephalalgia 2020, 40, (3), 288-298. [CrossRef]
- Tan, H.; Ding, Z.; Zhang, C.; Yan, J.; Yang, Y.; Li, P., The Programmed Cell Death Ligand-1/Programmed Cell Death-1 Pathway Mediates Pregnancy-Induced Analgesia via Regulating Spinal Inflammatory Cytokines. Anesth Analg 2021, 133, (5), 1321-1330. [CrossRef]
- Zhao, L.; Luo, H.; Ma, Y.; Zhu, S.; Wu, Y.; Lu, M.; Yao, X.; Liu, X.; Chen, G., An analgesic peptide H-20 attenuates chronic pain via the PD-1 pathway with few adverse effects. Proc Natl Acad Sci U S A 2022, 119, (31), e2204114119. [CrossRef]
Figure 1.
Mechanical and thermal hypersensitivity of mice in all groups. (A) von Frey filament test indicating the mechanical threshold. (B) Hargreaves' test estimating thermal latency. *significant difference to normal group. # significant difference to FM group. n = 9 per group.
Figure 1.
Mechanical and thermal hypersensitivity of mice in all groups. (A) von Frey filament test indicating the mechanical threshold. (B) Hargreaves' test estimating thermal latency. *significant difference to normal group. # significant difference to FM group. n = 9 per group.
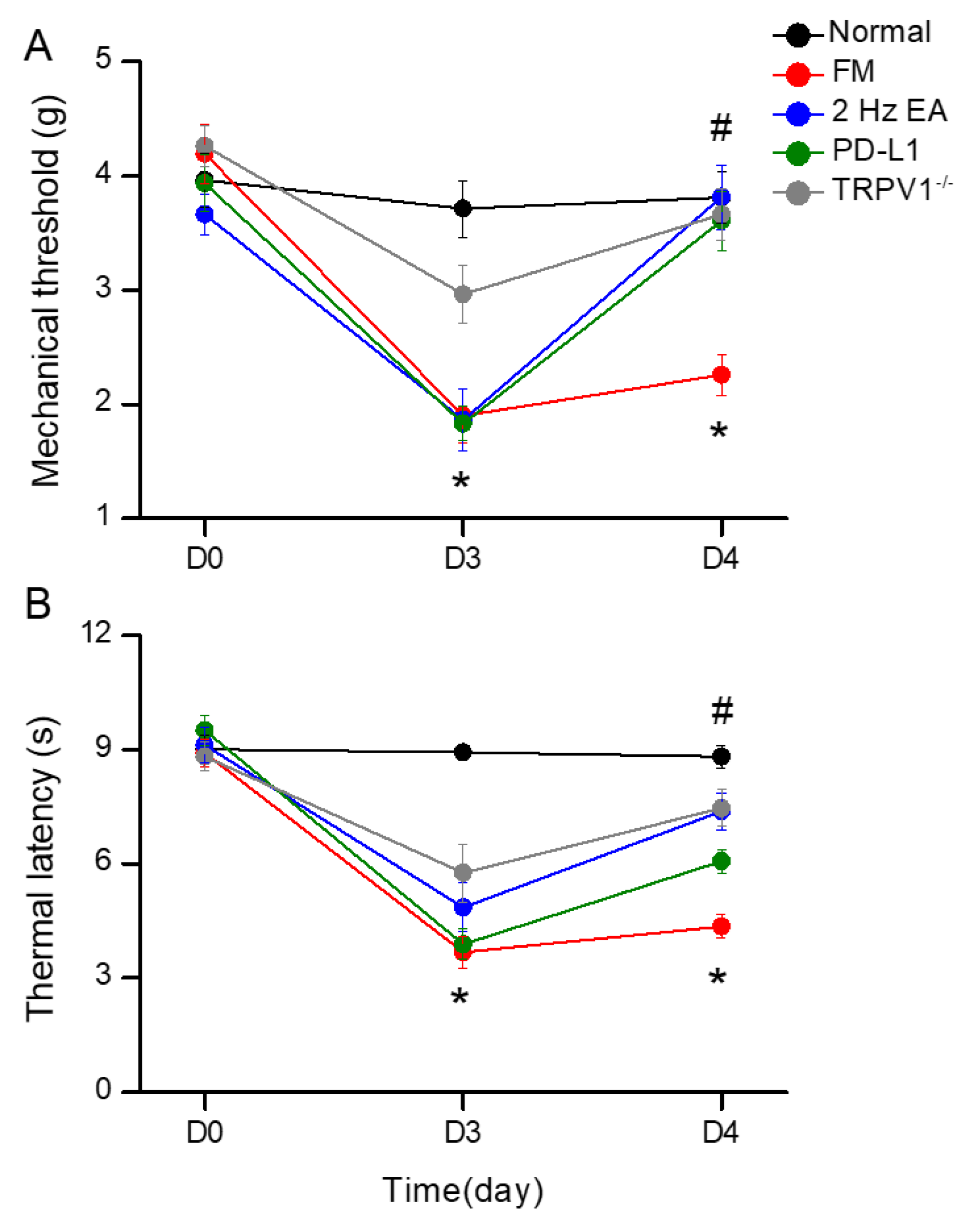
Figure 2.
Changing levels of PD-1 and related molecules in the DRG. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
Figure 2.
Changing levels of PD-1 and related molecules in the DRG. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
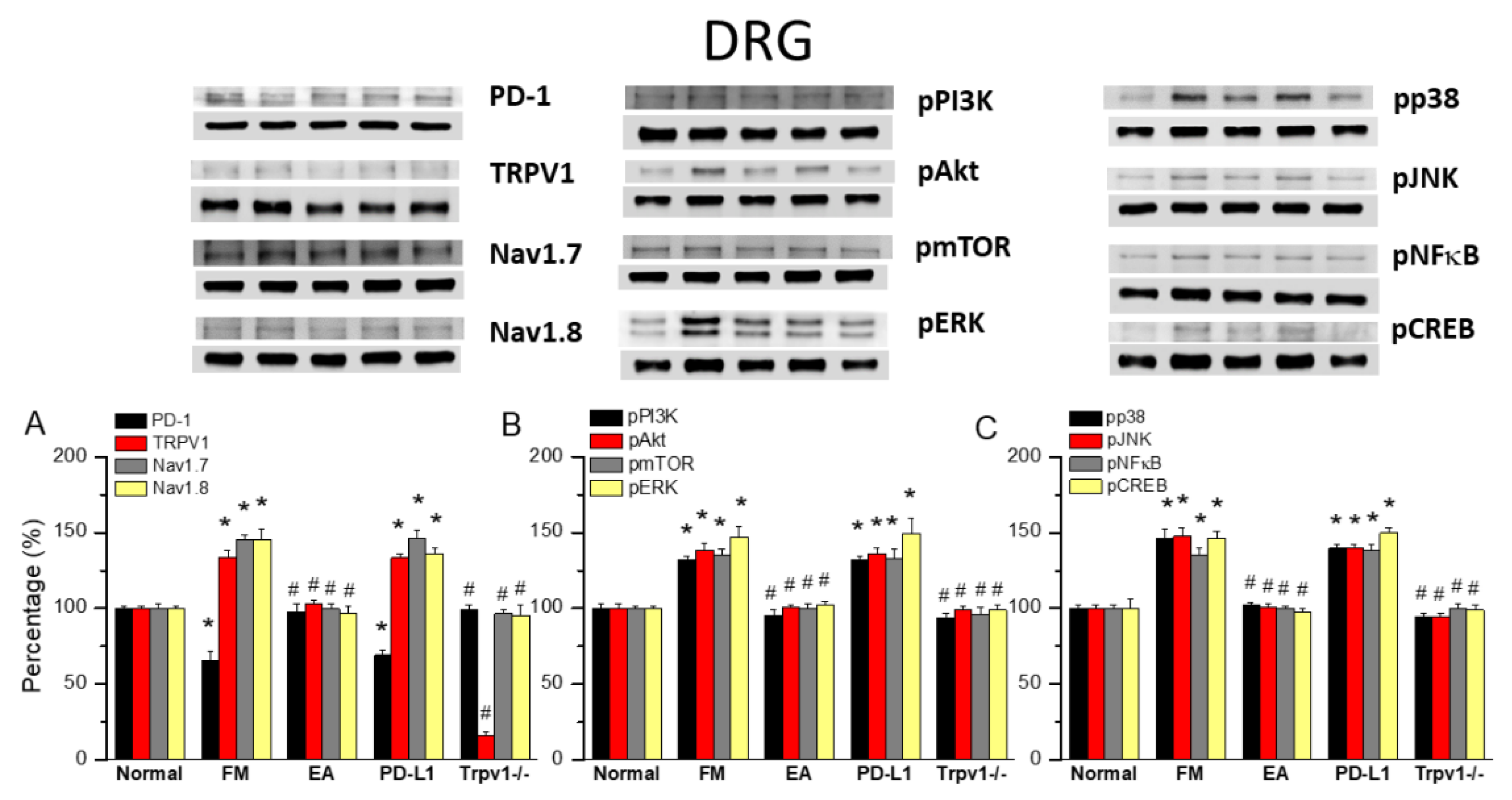
Figure 3.
Changing levels of PD-1 and related molecules in the SC. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
Figure 3.
Changing levels of PD-1 and related molecules in the SC. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
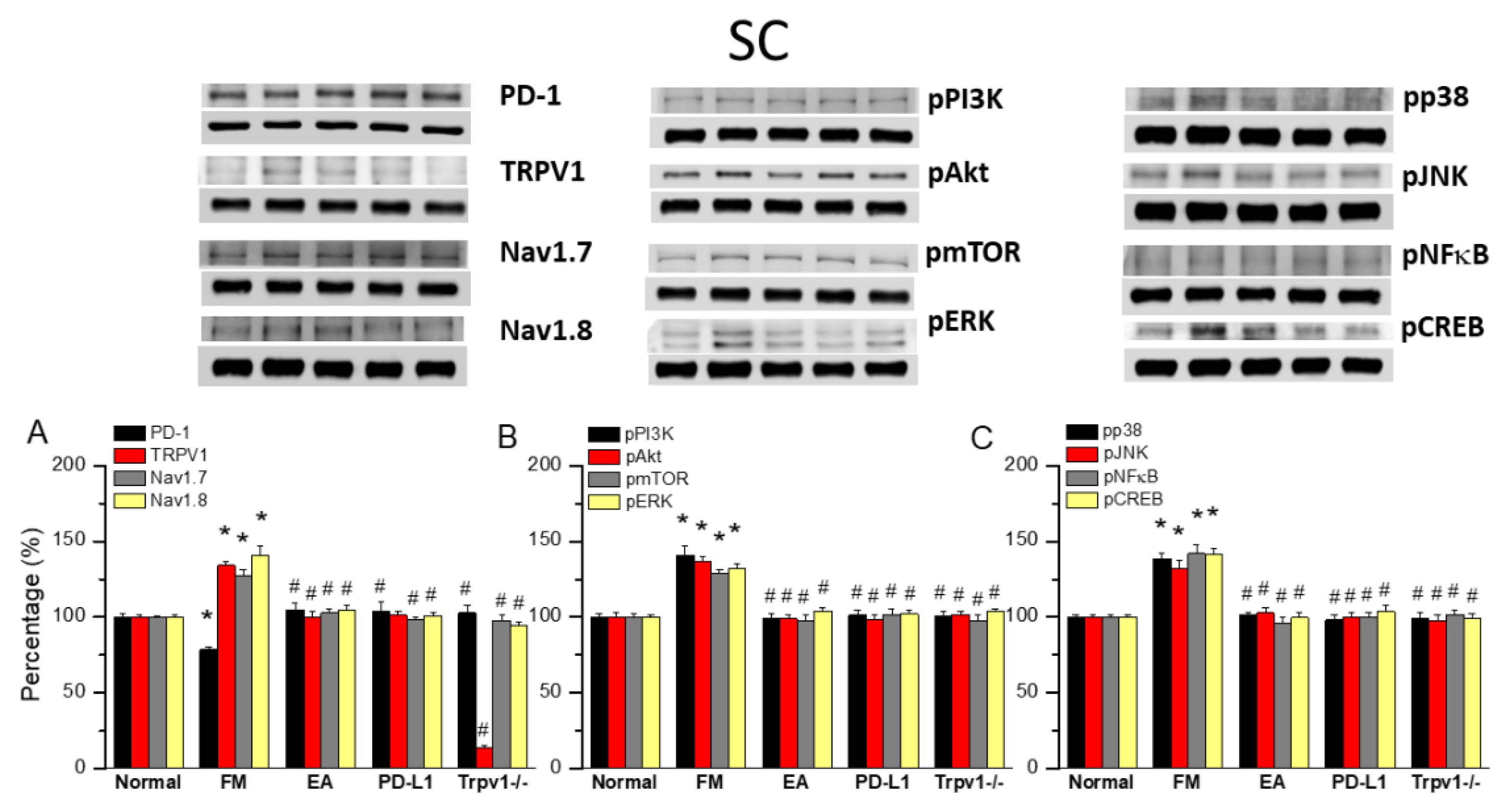
Figure 4.
Changing levels of PD-1 and related molecules in the thalamus. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
Figure 4.
Changing levels of PD-1 and related molecules in the thalamus. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
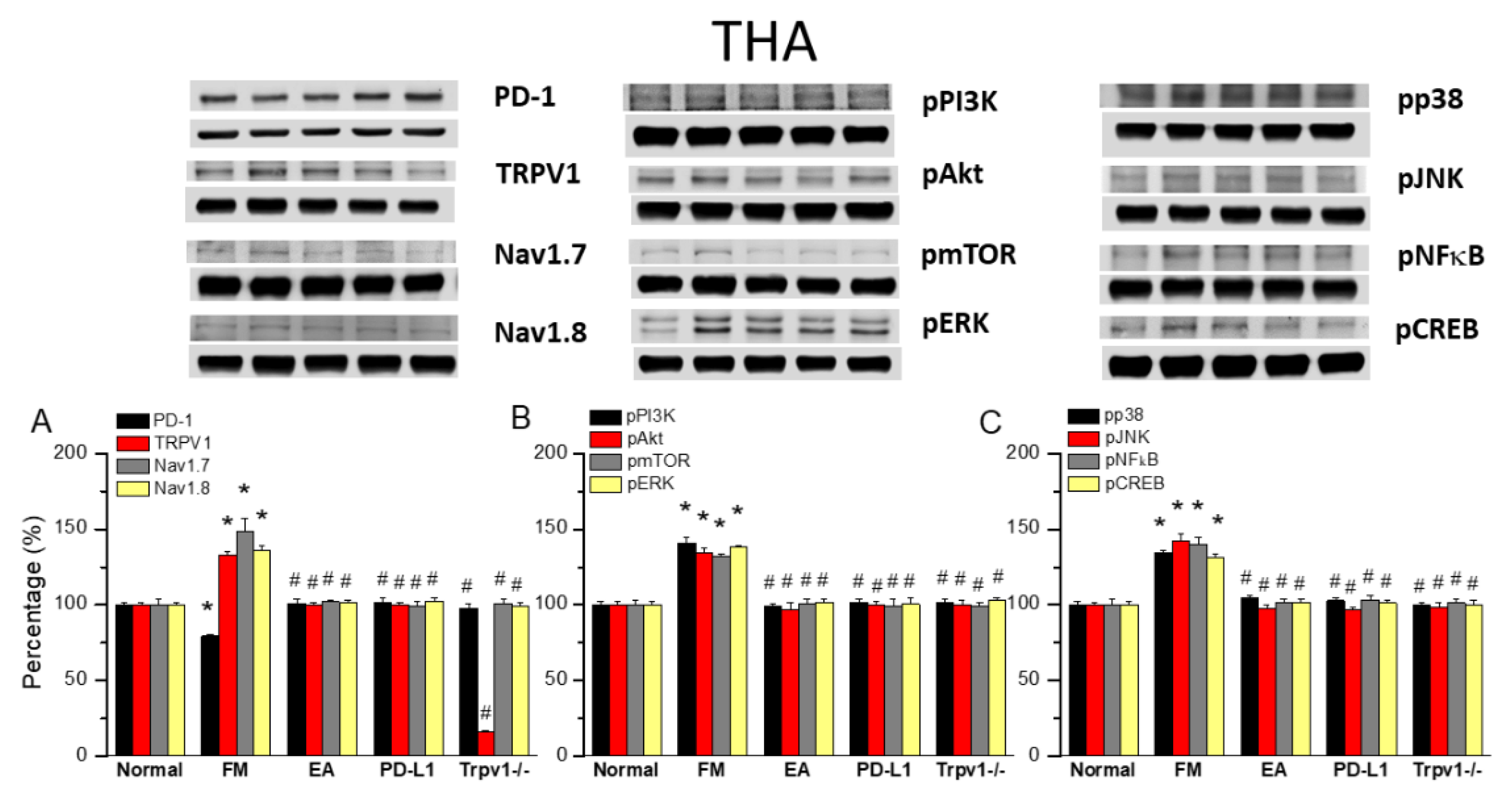
Figure 5.
Changing levels of PD-1 and related molecules in the SSC. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
Figure 5.
Changing levels of PD-1 and related molecules in the SSC. Western blot data with five lanes of proteins for Normal, FM, FM + EA, PD-L1, and Trpv1-/- groups. (A) PD-1, TRPV1, Nav1.7, and Nav1.8 (B) pPI3K, pAkt, pmTOR, pERk (C) pp38, pJNK, pNF-κB, and pCREB. *significant difference to normal. #significant difference to FM. n = 6.
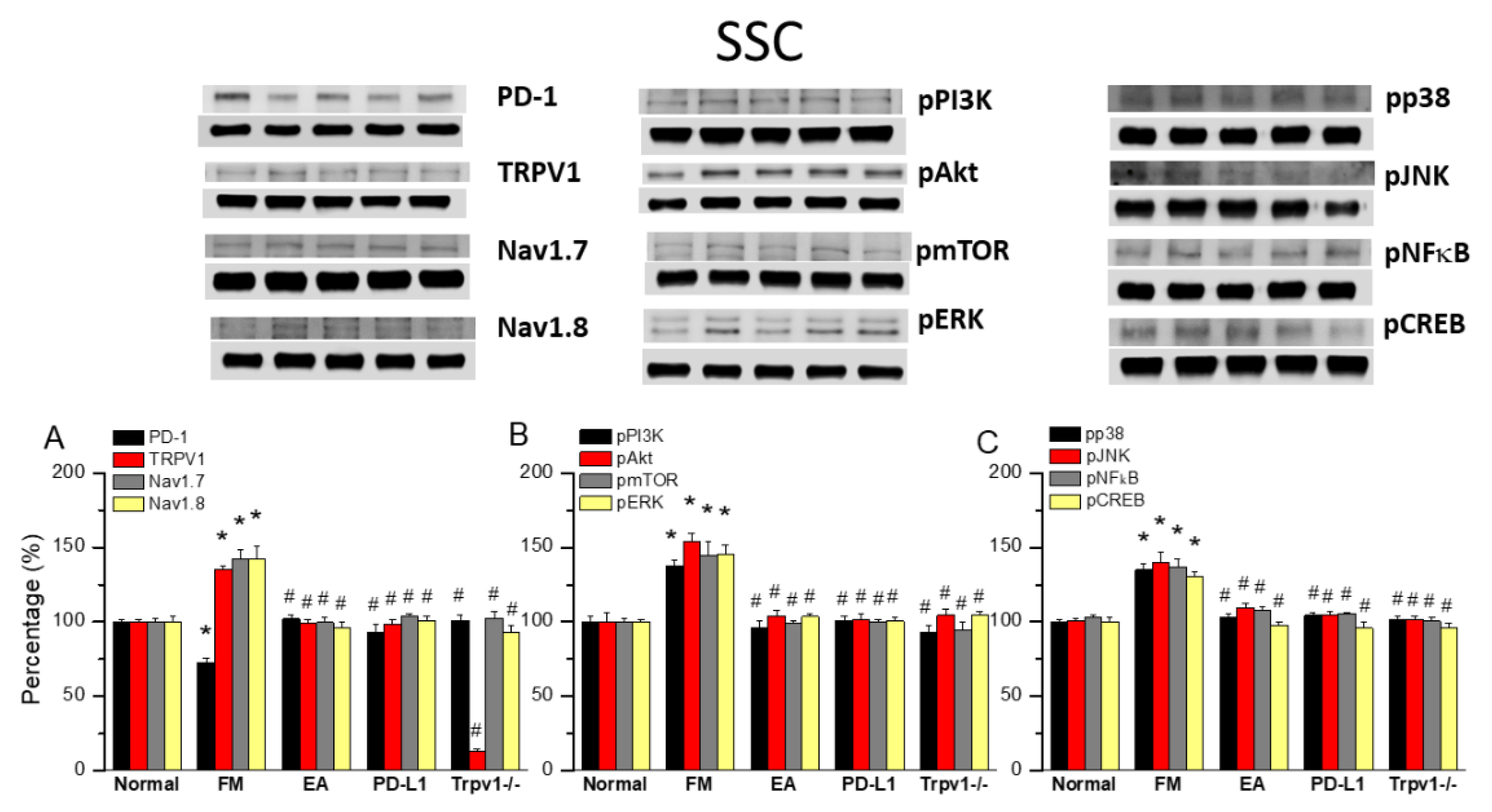
Figure 6.
Immunofluorescence staining of PD-1 and TRPV1 to evaluate protein expression in the mouse DRG and SSC. The yellow arrow indicates immune-positive signals. n = 3 for all groups.
Figure 6.
Immunofluorescence staining of PD-1 and TRPV1 to evaluate protein expression in the mouse DRG and SSC. The yellow arrow indicates immune-positive signals. n = 3 for all groups.
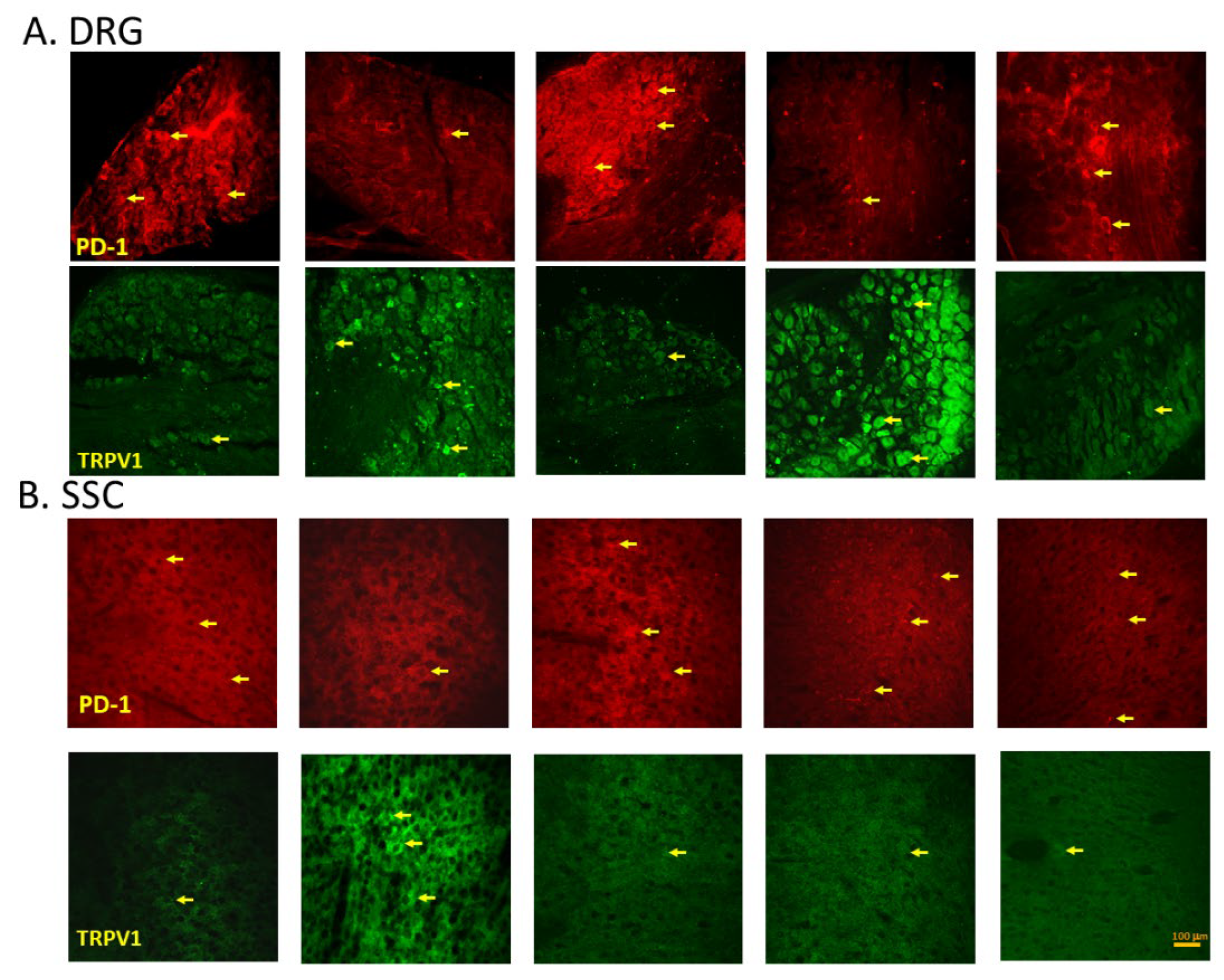
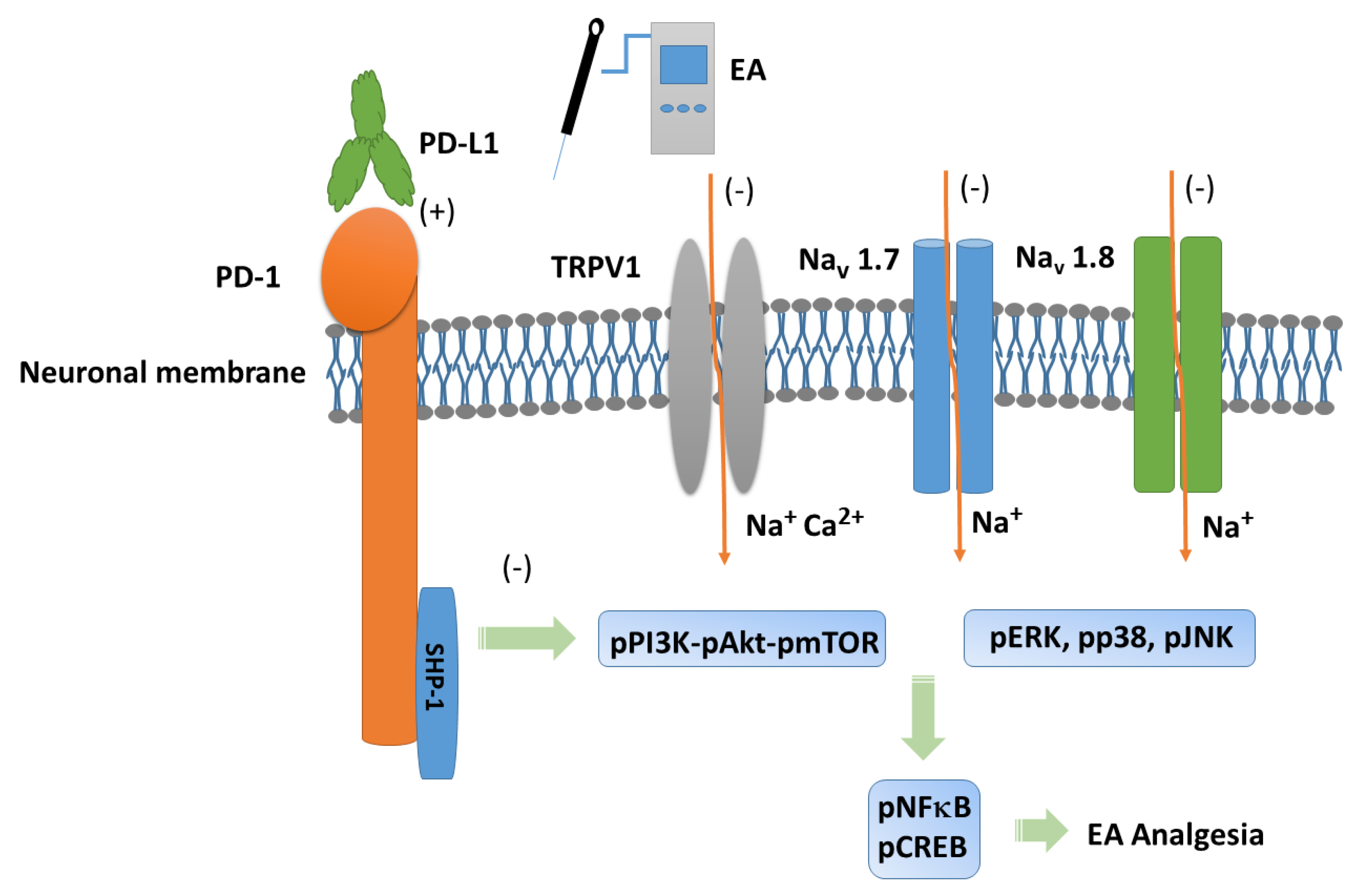
Figure 7.
Programmed cell death 1 ligand 1 (PD-L1) signaling pathway with specific effect on PD-1. Abbreviations: EA = Electroacupuncture; pERK = phosphorylated extracellular signal-regulated kinase; pp38 = phosphorylated protein kinase p38; pJNK = phosphorylated c-Jun N-terminal kinase; PD-L1 = Programmed cell death 1 ligand 1; pPI3K = phosphorylated phosphoinositide 3-kinase; pAkt = phosphorylated Akt; pmTOR = phosphorylated mammalian target of rapamycin; TRPV1 = Transient receptor potential V1; pNF-kB = phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells; pCREB = phosphorylated cAMP response element-binding protein.
Figure 7.
Programmed cell death 1 ligand 1 (PD-L1) signaling pathway with specific effect on PD-1. Abbreviations: EA = Electroacupuncture; pERK = phosphorylated extracellular signal-regulated kinase; pp38 = phosphorylated protein kinase p38; pJNK = phosphorylated c-Jun N-terminal kinase; PD-L1 = Programmed cell death 1 ligand 1; pPI3K = phosphorylated phosphoinositide 3-kinase; pAkt = phosphorylated Akt; pmTOR = phosphorylated mammalian target of rapamycin; TRPV1 = Transient receptor potential V1; pNF-kB = phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells; pCREB = phosphorylated cAMP response element-binding protein.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



