
Submitted:
08 May 2025
Posted:
09 May 2025
You are already at the latest version
Abstract
Fibromyalgia is a disease difficult to cure that causes chronic long-term pain, with symptoms lasting for months to years. Given the lack of evidence-based diagnosis and assessment methods for fibromyalgia, it ranks among the most difficult chronic pain conditions to treat. Programmed cell death ligand 1 (PD-L1) can inhibit acute and chronic pain transmission by inhibiting neuronal ion channels. Here, we aimed to explore the analgesic efficacy and mechanism of PD-L1/PD1 in a mouse fibromyalgia pain model. We used intermit-tent cold stress (ICS) to induce fibromyalgia in mice, this model was characterized by von Frey and Har-greaves’ tests. ICS-induced fibromyalgia pain mouse model exhibited mechanical and thermal hyperalgesia. Electroacupuncture (EA) or intraventricular PD-L1 injection effectively alleviated the nociceptive response and exhibited low PD-1 levels in the mouse dorsal root ganglia, spinal cord, thalamus, somatosensory cortex, and cerebellum. In contrast, pain-related kinase levels increased after fibromyalgia induction, and these effects was reversed by EA or PD-L1 via inhibition of microglia/astrocytes and toll-like receptor 4. Our results show that EA can treat fibromyalgia pain in mice through the PD-L1/PD1 pathway, indicating its therapeutic po-tential as a target in fibromyalgia.
Keywords:
electroacupuncture
; fibromyalgia
; PD-L1
; TLR4
; microglia
; astrocyte
1. Introduction
Fibromyalgia pain had been predicted to increase owing to lack of medicines for the symptoms and the illness. Fibromyalgia is challenging particularly because it is difficult to ascertain the cause of pain due to deficient diagnosis, symptomatology, and lack of meaningful biomarkers. So far, there is no standard therapy and no other alternatives to help fibromyalgia patients [1]. A recent study suggested that fibromyalgia patients had predominantly higher salivary cortisol concentrations than healthy subjects, which were risk factors for insomnia, stress, anxiety, and depression [2]. Neuroinflammation has also been involved in fibromyalgia progression [3]. Further, Cordón et al. identified that fibromyalgia patients had decreasing inner retinal neurons, suggesting neuroinflammation and neurodegeneration associated with fibromyalgia severity and a worsened quality of life [4]. In addition, fibromyalgia patients were found to have central sensitization in the central nervous system, including the brain and spinal cord, with increased pain sensation. Despite medications such as pregabalin, duloxetine, and milnacipran being approved by the US FDA for fibromyalgia pain relief, they have low patient satisfaction and side effects [5]. Furthermore, a healthy lifestyle including practice of Tai chi, meditation, exercise, or yoga can help fibromyalgia patients to relax muscles, lower stress, and reduce pain syndromes [6,7,8].
Programmed cell death ligand 1 (PD-L1), also known as CD274, regulates cellular immune responses in dendritic cells, lymphocytes, and endothelial cells. PD-L1 is present in tumor cell membranes and can bind to the programmed cell death protein 1 (PD-1) expressed in T cells. Accordingly, PD-L1/PD-1 signaling modulates immune function within the tumor microenvironment [9,10]. A recent article reported that interferon gamma (IFN-γ) significantly induced PD-L1 expression in tumor cells responding to cancer progression [11]. Furthermore, interleukin (IL)-17 and TNF-α can independently control PD-L1 by activating AKT, nuclear factor-κB (NFκB), and extracellular regulated protein kinases (ERK) signaling in cancer cells [12]. In addition, IFN-γ inhibition can attenuate PD-L1 expression via the myeloid differentiation primary response 88 (MyD88)/ (TNF receptor associated factor 6) TRAF6 pathway [13]. Thus, several drugs including atezolizumab, avelumab, nivolumab, and metformin aiming to inhibit the PD-L1/PD-1 pathway have been developed for cancer treatment. The latter can attenuate PD-L1 and exhibit antitumoral effects by stimulating endoplasmic-reticulum-linked deprivation [14].
The communication between neurons and glial cells, mainly microglia (Iba1-positive) and astrocytes (GFAP-positive), in pain signaling is crucial for the development of chronic pain. In case of injury or pathophysiological situations, allodynia or hyperalgesia co-occur with irritable microglia or astrocyte activation. Overactivated microglia are considered proinflammatory M1 that can further increase the levels of inflammatory cytokines such as ILs, TNF-α, IFN-γ, and chemokines to initiate allodynia and hyperalgesia and cause chronic pain [15,16]. High mobility group protein B1 (HMGB1) can be released increase by microglia or astrocyte in response to inflammation, and bind to inflammatory receptors such as the receptor for advanced glycation end products and toll-like receptors, further activating the mitogen-activated protein kinase and NFκB signaling pathways [17,18]. Astrocytic marker S100 calcium binding protein B (S100B) was also indicated to be activated in the brain areas, resulting in several chronic pain models. Toll-like receptor 4 (TLR4) can participate in innate and adaptive immune responses. It has an extracellular domain to sense extracellular inflammatory mediators such as S100B, HMGB1, and lipopolysaccharide, which can trigger a series of downstream pathways responding to the induction, transduction, and maintenance of chronic pain. Activation could further trigger MyD88 to increase TRAF6 and mitogen-activated protein kinase (MAPS)/NKκB and the production of proinflammatory cytokines such as IL-1, IL-2, IL-6, TNF-α, and IFN-γ [19,20].
Acupuncture, an ancient element of Chinese medicine, was first used in Asia over 3 thousand years for disease relief, especially in pain management. Given its well-known therapeutic effect as well as low side effects, acupuncture is practiced worldwide to treat back, low back, and dental pain, headaches, tennis elbow, and fibromyalgia. Modern acupuncture involves inserting a very fine steel needle into a specific point called acupoint. Manual acupuncture involves inserting a fine needle into the acupoint with lifting and rotating to ensure the occurrence of the de-qi, suggesting successful manipulation. Electroacupuncture (EA) is reported to be therapeutically effective and consistent. A recent double-blinded, randomized controlled trial demonstrated that acupuncture could relieve chronic pain and major depression disorder [21]. Further, EA can initiate antinociceptive efficacy in various mouse models of inflammatory and neuropathic pain, as well as fibromyalgia [22,23,24,25,26]. The evidenced EA efficacy is considered to occur through increased opioid, dopamine, cannabinoid, and adenosine receptors. EA can also attenuate the release of proinflammatory IL-1β, IL-6, TNFα, and IFN-γ. In addition, we previously demonstrated that EA could reduce mechanical and thermal hyperalgesia by suppressing TRPV1 signaling [25].
In the present study, we found the association of PD-L1/PD-1 present on the microglia/astrocytes with TLR4 and its allied signaling pathways in a mouse model of fibromyalgia pain induced by intermittent cold stress (ICS). Our results indicate that microglia or astrocyte numbers were increased in the dorsal root ganglion (DRG) of fibromyalgia mice. Simultaneous results were observed for the neurotransmitters HMGB1 and S100B. Next, we observed reduced expression of the PD-1 receptor in the DRG of fibromyalgia mice. Levels of TLR4 and downstream molecules such as MyD88, TRAF6, and pNFκB, all increased in the DRG of fibromyalgia mice. These phenomena were all reversed in mice receiving EA treatment but not in i.c.v. PD-L1 injected mice, suggesting a central effect. Similar results were obtained in the spinal cord (SC), thalamus, somatosensory cortex (SSC), and cerebellum (CB). EA treatment or i.c.v. PD-L1 injection significantly reversed those effects. Our results provide novel insights into the relationship between EA analgesia and the PD-L1/ PD-1 pathway.
2. Materials and Methods
2.1. Mice and Fibromyalgia Pain Model
Female C57B/L6 mice aged 8–12 weeks obtained from BioLasco Taiwan Ltd (Yilan, Taiwan) were used in the current study. Mouse weight was 18–20 g and they were kept in a specific pathogen-free environment before the experiments. After arrival, the mice were placed in a home cage under a 12-h light/dark cycle (light 6 a.m. to 6 p.m.). The temperature was maintained at 25ºC with 60% moisture. The experiments were approved by the Institute of Animal Care and Use Committee of China Medical University (Permit no. CMUIACUC-2024-076), Taiwan according to the Guide for the usage of Laboratory Animals (National Academy Press). Mice were randomly divided into four groups: normal (Normal); cold stress-induced fibromyalgia pain (FM); cold stress-induced fibromyalgia pain with EA (FM+EA); and cold stress-induced fibromyalgia pain with i.c.v. injection with PD-L1 (FM+PD-L1). To develop the mouse FM model, mice were kept in a 4ºC environment, while normal mice remained at 25ºC. At 10 a.m. the next day, FM mice were transferred to 25°C for 30 min, before being taken back at 4ºC for 30 min. This procedure was performed until 4 p.m., when they were moved again overnight from 4 p.m. during the first 3 days.
2.2. Nociceptive Behavior Examinations
Mice were placed in separate Plexiglas boxes perforated overhead, placed on an elevated horizontal wire mesh stand, covered with a dark cloth, maintained in a silent environment, at room temperature (25ºC), and allowed to habituate for 30 min before starting the behavioral test. The experiments were only conducted when the mice were calm, all feet were placed on the surface, and without grooming or sleeping. The von Frey filament measuring instrument (IITC Life Science Inc., USA) was used to increase the pain pressure in the center of the right plantar hind paw of the mice. The maximum pressure was achieved when the right hind paw was lifted using the plastic tip and the mouse reflexively withdrew the hind paw. We allowed for 3-min breaks between stimuli. The results were recorded as mechanical sensitivity. The Hargreaves’ test was used for measuring mice thermal sensitivity and preparations were similar to those as the von Frey test. The subjects were placed in an animal enclosure that separated the mice to limit interaction and covered using a dark cloth. After allowing 30 min of habituation, the experiments were initiated. The IITC Plantar Analgesia Meter (IITC Life, Sciences, SERIES8, Model 390G) was used to measure the withdrawal latency time of mice subjected to radiant heat applied on the surface, targeting the center of the right hind paw.
2.3. Western Blot Analysis
The DRG, SC, thalamus, SSC, and CB of the mice were fleshy excise to extract proteins. Protein extracts were prepared, then, 10% radioimmunoprecipitation (RIPA) lysis buffer (Fivephoton Biochemicals, RIPA-50), 100 μL of a protease inhibitor (Bionovas, FC0070-0001), and 100 μL of phosphatase inhibitor (Bionovas, FC0050-0001) were added to the lysate. After homogenization, the mixture was centrifuged at 10000 rpm for 10 min at 4ºC in an adjustable centrifuge. A total of 10 μL of extracted tissue sample was subjected to 8% or 12% SDS Tris-glycine gel electrophoresis, according to the protein size. A current electrophoresis power supply (PowerPac, Singapore) was used to run the gels in two sections: section 1 for 40 min at 50 V and section 2 for 1 h and 50 min at 100 V. A semidry transfer machine (Trans-Blot SD Cell, USA) transferred the gels onto PVDF membranes at 15 V for 45 min. The transferred membranes were washed with phosphate-buffered saline Tween (PBST with 0.05% Tween20) and blocked with bovine serum albumin for 30 min at 4ºC. Thereafter, the membranes were cultured with primary antibodies in PBST with 1% BSA and incubated overnight at 4ºC. Then, the membranes were incubated with secondary antibodies at 1:5000, peroxidase-conjugated goat anti-rabbit antibody (Jackson Immuno Research Laboratory, goat anti-mouse antibody (Jackson Immuno Research Laboratory)), for 2 h at 25ºC. Finally, we used an enhanced chemiluminescence substrate kit (PIERCE) to visualize the protein bands on the membranes with LAS-3000 Fujifilm (Fuji Photo Film Co., Ltd, Tokyo, Japan). The image density levels of specific protein bands were quantified using NIH Image J 1.54h software (Bethesda, MD, USA). α-Tubulin was used as internal controller.
2.4. Statistical Analysis
Statistical analysis was performed using SPSS, a statistical program. All statistical data are presented as the mean ± standard error (SEM). Differences among groups were analyzed using an ANOVA test, followed by a post hoc Tukey’s test. P < 0.05 was considered statistically significant.
3. Results
3.1. EA or PD-L1 Injection Relieved Mechanical and Thermal Hyperalgesia in an Intermittent Cold Stress-induced FM Mouse Model
To explore the role of EA and PD-L1 in fibromyalgia pain, we first generated a fibromyalgia pain model via ICS induction. Compared to normal mice (Figure 1A, black column, day 4: 4.12 ± 0.16 g, n = 9), FM mice showed lower mechanical threshold (Figure 1A, red column, day 4: 2.08 ± 0.13 g, n = 9), indicating mechanical hyperalgesia in this FM model. To confirm that EA had therapeutic effects in FM, we applied 2 Hz EA to FM mice. Our results demonstrated that EA treatment successfully diminished mechanical hyperalgesia (Figure 1A, blue column, day 4: 3.53 ± 0.24 g, n = 9). In addition, i.c.v. PD-L1 injection showed an analgesic effect in FM mice (Figure 1A, green column, day 4: 3.61 ± 0.25 g, n = 9). As expected, in FM mice undergoing ICS, the Hargraves’ test showed a significant thermal hyperalgesia (Figure 1B, red column, day 4: 3.93 ± 0.45 g, n = 9); however, 2 Hz EA treatment alleviated it (Figure 1B, blue column, day 4: 9.75 ± 0.23 g, n = 9). Furthermore, mice subjected to an i.c.v. injection of PD-L1 presented a longer thermal latency, suggesting its analgesic effect (Figure 1B, green column, day 4: 8.56 ± 0.45 g, n = 9).
3.2. Beneficial Effects of EA or PD-L1 on Fibromyalgia Pain were Mediated via Microglia, Astrocytes, and PD-1 Pathway in the Mouse DRG
The effects of EA or PD-L1 on microglia, astrocytes, and PD-1 receptor expression were evaluated 4 days after FM induction. On day 4, microglia (Iba1) and astrocytes (GFAP) were significantly increased in the L3–L5 DRG compared with normal mice (Figure 2A & B, *P < 0.05, n = 6). Iba1 or GFAP potentiation was alleviated by EA treatment but not PD-L1 i.c.v. injection. To determine whether HMGB1 and S100B were involved in this FM model and the effects of EA or PD-L1. Our results indicated that either HMGB1 or S100B were increased after FM induction and be mitigated by EA (Figure 2C & D, *P < 0.05, n = 6) but not PD-L1 injection. We next evaluated PD-1 expression in the mouse DRG. PD-1 expression in the DRG decreased in FM mice (Figure 2E, *P < 0.05, n = 6), 2 Hz EA but not PD-L1 injection increased it. We further measured TLR4 expression among groups. We determined TLR4 band intensity without ICS induction in mice. Once, TLR4 was amplified in the DRG of FM pain mice (Figure 2F, *P < 0.05, n = 6). However, the inflammatory protein pattern was relieved upon EA administration on day 4 but not in the PD-L1-treated group. Comparable trends were detected for MyD88. MyD88 increased in the DRG of FM mice compared with the normal group (Figure 2G, *P < 0.05, n = 6). 2 Hz EA, but not PD-L1 injection, significantly attenuated MyD88 overexpression. Next, we observed expression of downstream molecules of TLR4 TRAF6in the mouse DRG. In FM mice, TRAF6 was more strongly expressed in the DRG (Figure 2H, *P < 0.05, n = 6): this increased expression was inhibited by EA but not by PD-L1. Finally, detected pNFκB in the DRG of normal mice, which was overexpressed in the DRG of FM mice (Figure 2I, *P < 0.05, n = 6). This increase was markedly reduced in the EA group but not in PD-L1 mice.
3.3. EA and PD-L1 Treatment Reduced FM Pain in the Spinal Cord
Since microglia, astrocytes, and PD-1 receptors are crucial in FM pain and stress-related inflammation in the central nervous system of mice, we examined those in the lumbar SC. The ICS-initiated FM model exhibited an increase in the number of microglia and astrocytes labeled with Iba1 and GFAP than normal mice (Figure 3A & B, *P < 0.05, n = 6). In addition, 2 Hz EA treatment significantly decreased the numbers of those cells. Similar results were also observed in mice receiving i.c.v. PD-L1 injection (Figure 3A & B, #P < 0.05, n = 6). HMGB1 and S100B their expression was markedly increased in the lumbar SC of FM mice than in that of normal mice (Figure 3C & D, *P < 0.05, n = 6). In addition, this increase can be inhibited by 2 Hz EA treatment or PD-L1 injection, aligning with the behavioral results (Figure 3C & D, #P < 0.05, n = 6). We next examined the expression of PD-1. We found lower PD-1 expression in FM mice (Figure 3E, *P < 0.05, n = 6) which could be increased using 2 Hz EA or PD-L1 (Figure 3E, #P < 0.05, n = 6). Levels of TLR4 and MyD88 increased upon FM induction (Figure 3F & G, *P < 0.05, n = 6), an increase attenuated by EA treatment and PD-L1 injection (Figure 3F & G, #P < 0.05, n = 6). TRAF6 expression in the lumbar SC of normal mice increased in FM mice (Figure 3H, *P < 0.05, n = 6). This potentiation was alleviated by either EA treatment or PD-L1 injection (Figure 3H, #P < 0.05, n = 6). Furthermore, we detected increased pNFκB levels (Figure 3I, *P < 0.05, n = 6) in the SC and can be advanced declined by EA management and PD-L1 injection (Figure 3I, #P < 0.05, n = 6).
3.4. Hz EA or PD-L1 Treatment Can Reverse Microglial/Astrocytic Activation and TLR4 Accumulation in the Thalamus of FM Mice
No noticeable microglial or astrocyte accumulation was observed in the thalamus of normal mice at day 4. Accumulation of Iba1-positive microglia and GFAP-positive astrocytes were found in the thalamus of mice after ICS-induced fibromyalgia pain (Figure 4A & B, *P < 0.05, n = 6). However, after 4 days of ICS, Iba1 and GFAP levels were attenuated by 2 Hz EA management or PD-L1 injection (Figure 4A & B, #P < 0.05, n = 6). In the mouse thalamus, HMGB1 and S100B accumulation started at day 4 after ICS (Figure 4C & D, *P < 0.05, n = 6), however, these levels decreased in mice undergoing 2-Hz EA or receiving PD-L1 injection (Figure 4C & D, #P < 0.05, n = 6). Significantly, a reduction in PD-1 expression was observed in the thalamus of FM mice (Figure 4E, *P < 0.05, n = 6). Likewise, 2-Hz EA and PD-L1 resulted in increased PD-1 expression (Figure 4A & B, #P < 0.05, n = 6). Furthermore, augmented levels of TLR4-MyD88-TRAF6 were detected in the thalamus of FM mice (Figure 4F-H, *P < 0.05, n = 6), which decreased with 2 Hz EA or PD-L1 (Figure 4F-H, #P < 0.05, n = 6). pNFκB expression in the thalamus of FM mice was higher than in normal mice (Figure 4I, *P < 0.05, n = 6). 2-Hz EA treatment and PD-L1 injection reliably reversed this pattern of expression (Figure 4I, #P < 0.05, n = 6).
3.5. EA or PD-L1 Treatment Reduced FM Pain by Inhibiting the Microglia/Astrocyte-TLR4 Pathway in the Somatosensory Cortex
Iba1- and GFAP-positive immunoreactivity was next assessed in the SSC, which indicated that the SSC was documented as a region for pain establishment. Iba1 and GFAP expression was observed in the SSC of normal mice. In FM mice, the levels of Iba1 and GFAP were increased (Figure 5A & B, *P < 0.05, n = 6) but 2-Hz EA treatment significantly attenuated this effect (Figure 5A & B, #P < 0.05, n = 6). A similar tendency was observed with PD-L1 injection (Figure 5A & B, #P < 0.05, n = 6). In addition, there were increased HMGB1 and S100B immunopositive signals in the SSC of FM mice than normal mice (Figure 5C & D, *P < 0.05, n = 6). A significant decrease in HMGB1 and S100B immunolabeling was observed with 2-Hz EA or PD-L1 (Figure 5C & D, #P < 0.05, n = 6). Next, we detected PD-1 in the SSC of mice, which increased in FM mice (Figure 5E, *P < 0.05, n = 6); an increase inhibited by 2 Hz EA or PD-L1 administration (Figure 5E, #P < 0.05, n = 6). In addition, the increased TLR4 and MyD88 concentration in the SSC of FM mice (Figure 5F & G, *P < 0.05, n = 6) were inhibited by 2-Hz EA or PD-L1 injection (Figure 5F & G, #P < 0.05, n = 6). Similarly, TRAF6 levels increased after ICS induction (Figure 5H, *P < 0.05, n = 6), but decreased in the EA and PD-L1 groups (Figure 5H, #P < 0.05, n = 6). Augmented pNFκB were found in fibromyalgia mice (Figure 5I, *P < 0.05, n = 6) and further aggravated by 2-Hz EA or PD-L1 usage (Figure 5I, #P < 0.05, n = 6).
3.6. Cold Stress Initiated Fibromyalgia Pain by Increasing the Microglia/Astrocyte-TLR4 Pathway in the Cerebellum 5-7, the Effect Reversed by EA and PD-L1 Treatment
Lastly, to define the effect of ICS-induced fibromyalgia pain and EA or PD-L1 on fibromyalgia pain signaling in the mice CB5-7, we analyzed aforementioned molecules using the CB5-7 regions. We perceived a noteworthy increase in GFAP and Iba1 expression in ICS-initiated fibromyalgia pain mice (Figure 6-8A & B, *P < 0.05, n = 6). The potentiated effect was reversed by 2 Hz EA or PD-L1 injection (Figure 6-8A & B, #P < 0.05, n = 6). As well, ICS also augmented HMGB1 and S100B concentrations in the mice CB5-7 (Figure 6, Figure 7 and Figure 8C & D, *P < 0.05, n = 6). Such intensification was then abridged by 2 Hz EA or PD-L1 injection (Figure 6-8C & D, #P < 0.05, n = 6). In addition, PD-1 receptor was found attenuated in mice CB5-7 after receiving ICS induction (Figure 6-8E, *P < 0.05, n = 6). Series phenomena were also abrogated by using 2 Hz EA or PD-L1 treatment (Figure 6-8E, #P < 0.05, n = 6). Moving to TLR4 cascades, our results indicated a comparable tendency in TLR4, MyD88, and TRAF6 protein concentrations in fibromyalgia mice compared to normal mice (Figure 6-8F-H, *P < 0.05, n = 6). The potentiated protein levels was considerably alleviated by 2 Hz EA and PD-L1 injection. Similar propensities were also detected for pNFκB (Figure 6-8I, n = 6).
4. Discussion
Growing scientific evidence indicates that the immune checkpoint PD-L1 could be stimulated in cancer cells to inhibit T cell function through activation of the PD-1 receptor. Less is known about how the PD-1 signaling pathway can regulate pain signaling, especially in neuromodulation. A recent article reported that healthy tissue DRGs could release PD-L1 to notably inhibit either acute or chronic pain. Local PD-L1 injection can produce reliable antinociception in healthy mice through activation of PD-1. In addition, PD-L1 inhibition via neutralization or PD-1 receptor considerably initiated mechanical allodynia, suggesting its role in pain control. Genetic deletion of Pd1 resulted in thermal and mechanical hyperalgesia. The released PD-L1 then binds to the PD-1 receptor of DRG nociceptive neurons, resulting in the phosphorylation of Src homology region 2 domain-containing phosphatase 1 (SHP-1). Phosphorylated SHP-1 next suppresses sodium channels and induces neuronal hyperpolarization over triggering potassium channels [27]. TLRs expressed in primary nociceptive neurons, are important modulators of the immune system to regulate pain sensation via regulating ion channels; however, its relationship with PD-L1 is unelucidated.
Wang et al. reported that the immune therapy nivolumab, a monoclonal antibody drug against PD-1, has a therapeutic effect in tumor suppression. They then utilized the genetic knockout of the PD-1 receptor (Pd1−/−) to demonstrate the defense of lung cancer cell-induced bone destruction. Mice lacking the PD-1 receptor showed higher nociceptive responses than healthy mice. In addition, this study determined that PD-L1 and chemokine ligand 2 (CCL2) were simultaneously increased in the tumor microenvironment. Cancer cells also exhibited increased the expression of chemokine receptor 2 (CCR2) in the mouse primary sensory neurons. The antagonism of CCR2 next produced significant attenuation of bone cancer pain [28]. Spinal cord injury (SCI) causes motor disability and neuropathic pain, that is difficult to treat. PD-L1 increased after SCI in the microglia present at the epicenter of the injury site. Mice without PD-L1 showed more serious neuropathic pain due to increased polarization of M1-like microglia compared with normal mice. Furthermore, PD-L1 attenuated neuropathic pain after SCI by inhibiting pp38 and pERK1/2 [29]. Shi et al. observed PD-L1 and PD-1 immuno-positive signals in normal trigeminal ganglia neurons. They then determined that the mRNA and protein levels of PD-L1 and PD-1 were considerably increased after the acute nitroglycerin-induced mouse migraine model. Furthermore, they indicated that the inhibition of the PD-1 receptor potentiated acute nitroglycerin-induced migraine. Likewise, this phenomenon was simultaneously go along with amplified calcitonin gene-related peptide (CGRP), IL-1β, IL-6, IL-18, and TNF-α expression in the trigeminal ganglia of neuropathic pain mice [30]. Our findings are consistent in that EA or PD-L1 could reliably alleviate FM pain through the PD-1 receptor to attenuate the microglia/astrocyte signaling pathway.t
Clinically, after stroke, patents often experience thalamic pain, which is a neuropathic pain syndrome; an increase in cases of this type of neuralgia has been observed. A recent study declared that collagenase IV was injected into the right thalamus of the mice to induce thalamic pain, and dexmetatomidine (a selective α2 adrenergic receptor agonist) relieved the thalamic pain. The increased levels of Iba1, GFAP, and TLR4/NF-κB signaling in thalamic pain mice were reverted by intraperitoneal injection of dexmetatomidine [31]. In neuropathic pain, targeting ion channels are on the rise for new drug development through the modulation of pain sensation, signal transduction, and management. Transient receptor potential channel melastatin 2 (TRPM2) was stated to adjust the Ca2+ concentration and had a crucial effect in pain signaling, osmosis, and temperature sensing. A recent study indicated that TLR4 is one of the main receptors responding to inflammatory conditions. TLR4 activation resulted in higher oxidative stress and expression of inflammatory cytokines such as IL-1β, IL-6, and TNF-α, etc. The potentiated molecules further activate TRPM2, which results in over influx of Ca2+ to induce neuronal damage [32]. Converting M1, increased proinflammatory factors, or M2, neuroprotective effects, and the microglial phenotype was reported as an encouraging healing approach for acute and chronic pain. A recent article declared that the antiinflammatory dietary flavonoid kaempferol had an analgesic effect in a mouse chronic constriction injury-induced neuropathic pain model. Their data indicated that kaempferol reliably relieved neuropathic pain accompanied by reduced proinflammatory cytokine production. They showed that kaempferol reliably reduced the neuropathic pain-induced overexpression ofTLR4/NF-κB in the SC of rats. Kaempferol can attenuate neuropathic pain by promoting microglial polarization from the M1 to M2 phenotype [33].
5. Conclusions
Our study is the first systematical study to investigate the effects of EA on PD-L1 to TLR4 signaling for the treatment of FM pain. Mechanical and thermal pain in FM mice via ICS induction was reverted by 2-Hz EA or PD-L1. Further, levels of microglia/astrocytes as well as HMGB1 and S100B were augmented whereas PD-1 was decreased in the DRG, SC, thalamus, SSC, and cerebellum of FM mice. Further, expression of nociceptive TLR4 and related downstream elements increased after FM induction, which could be repressed by EA or PD-L1 in all tested regions except for the DRG, where PD-L1 injection cannot reverse expression. Our results indicate that PD-L1/PD1 pathway might be the therapeutic potential as a target in fibromyalgia (Figure 9).
Author Contributions
IH Hsiao, WH Chen, and MC Lin: Conceptualization, Methodology, Software, Data curation, Writing-original draft, Visualization, Investigation. HC Hsu, Y Chae and YW Lin: Supervision, validation, writing, review and editing.
Funding
This work was supported by the following grants: NSTC 113-2314-B-039-047, CMUHCH-DMR-114-027, EDAHP114025, and the "Chinese Medicine Research Center, China Medical University" from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan.
Institutional Review Board Statement
The animal study protocol was approved by the Institute of Animal Care and Use Committee of China Medical University (Permit no. CMUI-ACUC-2021-336).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
There are no financial or other relationships that might lead to a conflict of interest for any of the authors.
References
- Barati, M.; Mirzavi, F.; Atabaki, M.; Bibak, B.; Mohammadi, M.; Jaafari, M.R. A review of PD-1/PD-L1 siRNA delivery systems in immune T cells and cancer cells. Int Immunopharmacol 2022, 111, 109022. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Li, X.; Zheng, Y.; Shi, H.; Zhang, D.; Jing, B.; Chen, Z.; Qian, G.; Zhao, G. Kaempferol exerts a neuroprotective effect to reduce neuropathic pain through TLR4/NF-kB signaling pathway. Phytother Res 2022, 36, 1678–1691. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Kim, Y.H.; Li, H.; Luo, H.; Liu, D.L.; Zhang, Z.J.; Lay, M.; Chang, W.; Zhang, Y.Q.; Ji, R.R. PD-L1 inhibits acute and chronic pain by suppressing nociceptive neuron activity via PD-1. Nat Neurosci 2017, 20, 917–926. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, Z.; Ma, X. The role of astrocytes in neuropathic pain. Front Mol Neurosci 2022, 15, 1007889. [Google Scholar] [CrossRef] [PubMed]
- Cordon, B.; Orduna, E.; Vilades, E.; Garcia-Martin, E.; Garcia-Campayo, J.; Puebla-Guedea, M.; Polo, V.; Larrosa, J.M.; Pablo, L.E.; Vicente, M.J.; et al. Analysis of Retinal Layers in Fibromyalgia Patients with Premium Protocol in Optical Tomography Coherence and Quality of Life. Curr Eye Res 2022, 47, 143–153. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, J.; Cai, Y.; Fu, S.; Zhang, N.; Fu, X.; Li, L. IFN-gamma-mediated inhibition of lung cancer correlates with PD-L1 expression and is regulated by PI3K-AKT signaling. Int J Cancer 2018, 143, 931–943. [Google Scholar] [CrossRef]
- Hsiao, I.H.; Liao, H.Y.; Lin, Y.W. Optogenetic modulation of electroacupuncture analgesia in a mouse inflammatory pain model. Sci Rep 2022, 12, 9067. [Google Scholar] [CrossRef]
- Hsiao, I.H.; Yen, C.M.; Hsu, H.C.; Liao, H.Y.; Lin, Y.W. Chemogenetics Modulation of Electroacupuncture Analgesia in Mice Spared Nerve Injury-Induced Neuropathic Pain through TRPV1 Signaling Pathway. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Illescas-Montes, R.; Costela-Ruiz, V.J.; Melguizo-Rodriguez, L.; De Luna-Bertos, E.; Ruiz, C.; Ramos-Torrecillas, J. Application of Salivary Biomarkers in the Diagnosis of Fibromyalgia. Diagnostics (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Khan, S.; Thalib, H.I.; Kauther, D.; Jamal, A.; Moallem, A.A.; Aldera, S.S.; Abou Touk, A.; Hassan, F.E. Genetic and immunological implications in fibromyalgia: A Literature Review. Egypt J Immunol 2024, 31, 13–26. [Google Scholar] [CrossRef]
- Kong, F.; Sun, K.; Zhu, J.; Li, F.; Lin, F.; Sun, X.; Luo, X.; Ren, C.; Lu, L.; Zhao, S.; et al. PD-L1 Improves Motor Function and Alleviates Neuropathic Pain in Male Mice After Spinal Cord Injury by Inhibiting MAPK Pathway. Front Immunol 2021, 12, 670646. [Google Scholar] [CrossRef] [PubMed]
- Langhorst, J.; Hauser, W.; Lauche, R.; Perrot, S.; Alegre, C.; Sarzi Puttini, P.C. Complementary and alternative medicine for the treatment of fibromyalgia. Evid Based Complement Alternat Med 2014, 2014, 408436. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, W. The Role of Satellite Glial Cells, Astrocytes, and Microglia in Oxaliplatin-Induced Neuropathic Pain. Biomedicines 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, K.; Liu, Y.; Wu, H.; He, Y.; Li, C.; Wang, Q.; Su, X.; Yan, S.; Su, W.; et al. A Novel Drug Combination of Mangiferin and Cinnamic Acid Alleviates Rheumatoid Arthritis by Inhibiting TLR4/NFkappaB/NLRP3 Activation-Induced Pyroptosis. Front Immunol 2022, 13, 912933. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.Y.; Hsieh, C.L.; Huang, C.P.; Lin, Y.W. Electroacupuncture Attenuates CFA-induced Inflammatory Pain by suppressing Nav1.8 through S100B, TRPV1, Opioid, and Adenosine Pathways in Mice. Sci Rep 2017, 7, 42531. [Google Scholar] [CrossRef]
- Liao, H.Y.; Satyanarayanan, S.K.; Lin, Y.W.; Su, K.P. Clinical efficacy and immune effects of acupuncture in patients with comorbid chronic pain and major depression disorder: A double-blinded, randomized controlled crossover study. Brain Behav Immun 2023, 110, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Chou, A.I.W.; Su, H.; Su, K.P. Transient receptor potential V1 (TRPV1) modulates the therapeutic effects for comorbidity of pain and depression: The common molecular implication for electroacupuncture and omega-3 polyunsaturated fatty acids. Brain Behav Immun 2020, 89, 604–614. [Google Scholar] [CrossRef]
- Liu, J.; Hamrouni, A.; Wolowiec, D.; Coiteux, V.; Kuliczkowski, K.; Hetuin, D.; Saudemont, A.; Quesnel, B. Plasma cells from multiple myeloma patients express B7-H1 (PD-L1) and increase expression after stimulation with IFN-gamma and TLR ligands via a MyD88-, TRAF6-, and MEK-dependent pathway. Blood 2007, 110, 296–304. [Google Scholar] [CrossRef]
- Mandlem, V.K.K.; Rivera, A.; Khan, Z.; Quazi, S.H.; Deba, F. TLR4 induced TRPM2 mediated neuropathic pain. Front Pharmacol 2024, 15, 1472771. [Google Scholar] [CrossRef]
- Perry, R.; Leach, V.; Davies, P.; Penfold, C.; Ness, A.; Churchill, R. An overview of systematic reviews of complementary and alternative therapies for fibromyalgia using both AMSTAR and ROBIS as quality assessment tools. Syst Rev 2017, 6, 97. [Google Scholar] [CrossRef]
- Shadbad, M.A.; Asadzadeh, Z.; Hosseinkhani, N.; Derakhshani, A.; Alizadeh, N.; Brunetti, O.; Silvestris, N.; Baradaran, B. A Systematic Review of the Tumor-Infiltrating CD8(+) T-Cells/PD-L1 Axis in High-Grade Glial Tumors: Toward Personalized Immuno-Oncology. Front Immunol 2021, 12, 734956. [Google Scholar] [CrossRef]
- Shi, S.; Han, Y.; Wang, D.; Guo, P.; Wang, J.; Ren, T.; Wang, W. PD-L1 and PD-1 expressed in trigeminal ganglia may inhibit pain in an acute migraine model. Cephalalgia 2020, 40, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Ali, S.A. Multifunctional Role of S100 Protein Family in the Immune System: An Update. Cells 2022, 11. [Google Scholar] [CrossRef]
- Tsujita, R.; Tsubota, M.; Sekiguchi, F.; Kawabata, A. Role of high-mobility group box 1 and its modulation by thrombomodulin/thrombin axis in neuropathic and inflammatory pain. Br J Pharmacol 2021, 178, 798–812. [Google Scholar] [CrossRef]
- Vega-Ramirez, M.T.; Becerril-Villanueva, E.; Maldonado-Garcia, J.L.; Pavon, L.; Perez-Sanchez, G. S100 proteins: a new frontier in fibromyalgia research. Mol Brain 2024, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gu, Y.; Liao, Y.; Bang, S.; Donnelly, C.R.; Chen, O.; Tao, X.; Mirando, A.J.; Hilton, M.J.; Ji, R.R. PD-1 blockade inhibits osteoclast formation and murine bone cancer pain. J Clin Invest 2020, 130, 3603–3620. [Google Scholar] [CrossRef]
- Wang, X.; Yang, L.; Huang, F.; Zhang, Q.; Liu, S.; Ma, L.; You, Z. Inflammatory cytokines IL-17 and TNF-alpha up-regulate PD-L1 expression in human prostate and colon cancer cells. Immunol Lett 2017, 184, 7–14. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Kwong, W.J.; Armstrong, H.; Behling, M.; Niemira, J.; Lang, K. Analysis of Real-World Dosing Patterns for the 3 FDA-Approved Medications in the Treatment of Fibromyalgia. Am Health Drug Benefits 2018, 11, 293–301. [Google Scholar]
- Wingenfeld, K.; Nutzinger, D.; Kauth, J.; Hellhammer, D.H.; Lautenbacher, S. Salivary cortisol release and hypothalamic pituitary adrenal axis feedback sensitivity in fibromyalgia is associated with depression but not with pain. J Pain 2010, 11, 1195–1202. [Google Scholar] [CrossRef]
- Ye, G.; Miao, R.; Chen, J.; Huang, J.; Jiang, M. Effectiveness of Complementary and Alternative Medicine in Fibromyalgia Syndrome: A Network Meta-Analysis. J Pain Res 2024, 17, 305–319. [Google Scholar] [CrossRef]
- Ye, S.; Ma, F.; Mahmood, D.F.D.; Vera, P.L. Modulation of persistent bladder pain in mice: The role of macrophage migration inhibitory factor, high mobility group box-1, and downstream signaling pathways. Bladder (San Franc) 2024, 11, e21200011. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.A.; Hsu, H.C.; Lin, M.C.; Chen, T.S.; Lin, W.C.; Huang, H.M.; Lin, Y.W. Electroacupuncture Regulates Cannabinoid Receptor 1 Expression in a Mouse Fibromyalgia Model: Pharmacological and Chemogenetic Modulation. Life (Basel) 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Li, G.; Wang, J.; Liu, M.; Wang, Z.; Song, Y.; Zhang, X.; Wang, X. PD-L1: expression regulation. Blood Sci 2023, 5, 77–91. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Electroacupuncture or PD-L1 i.c.v. inoculation reduced the fibromyalgia pain after ICS. (A) Mechanical hyperalgesia verified using the von Frey test. (B) Thermal hyperalgesia determined using the Hargraves’ analysis. Normal: normal mice; FM: fibromyalgia mice; FM+EA: fibromyalgia mice treated with EA; FM+PD-L1: fibromyalgia mice treated with PD-L1. *P < 0.05 vs Normal. #P < 0.05 vs FM group.
Figure 1.
Electroacupuncture or PD-L1 i.c.v. inoculation reduced the fibromyalgia pain after ICS. (A) Mechanical hyperalgesia verified using the von Frey test. (B) Thermal hyperalgesia determined using the Hargraves’ analysis. Normal: normal mice; FM: fibromyalgia mice; FM+EA: fibromyalgia mice treated with EA; FM+PD-L1: fibromyalgia mice treated with PD-L1. *P < 0.05 vs Normal. #P < 0.05 vs FM group.
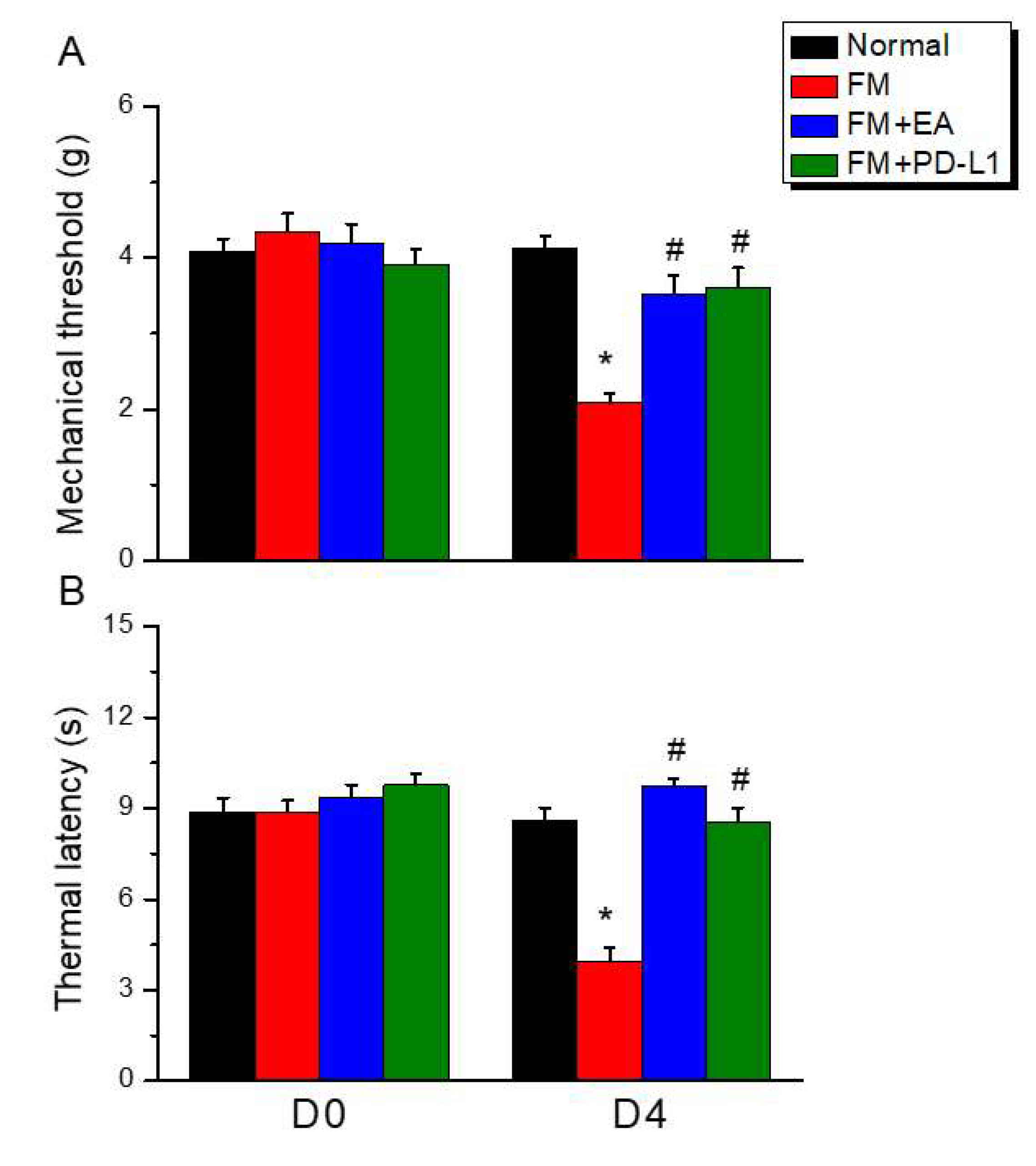
Figure 2.
Changing protein levels of PD-1 on pain-related molecules in the DRG of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 2.
Changing protein levels of PD-1 on pain-related molecules in the DRG of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
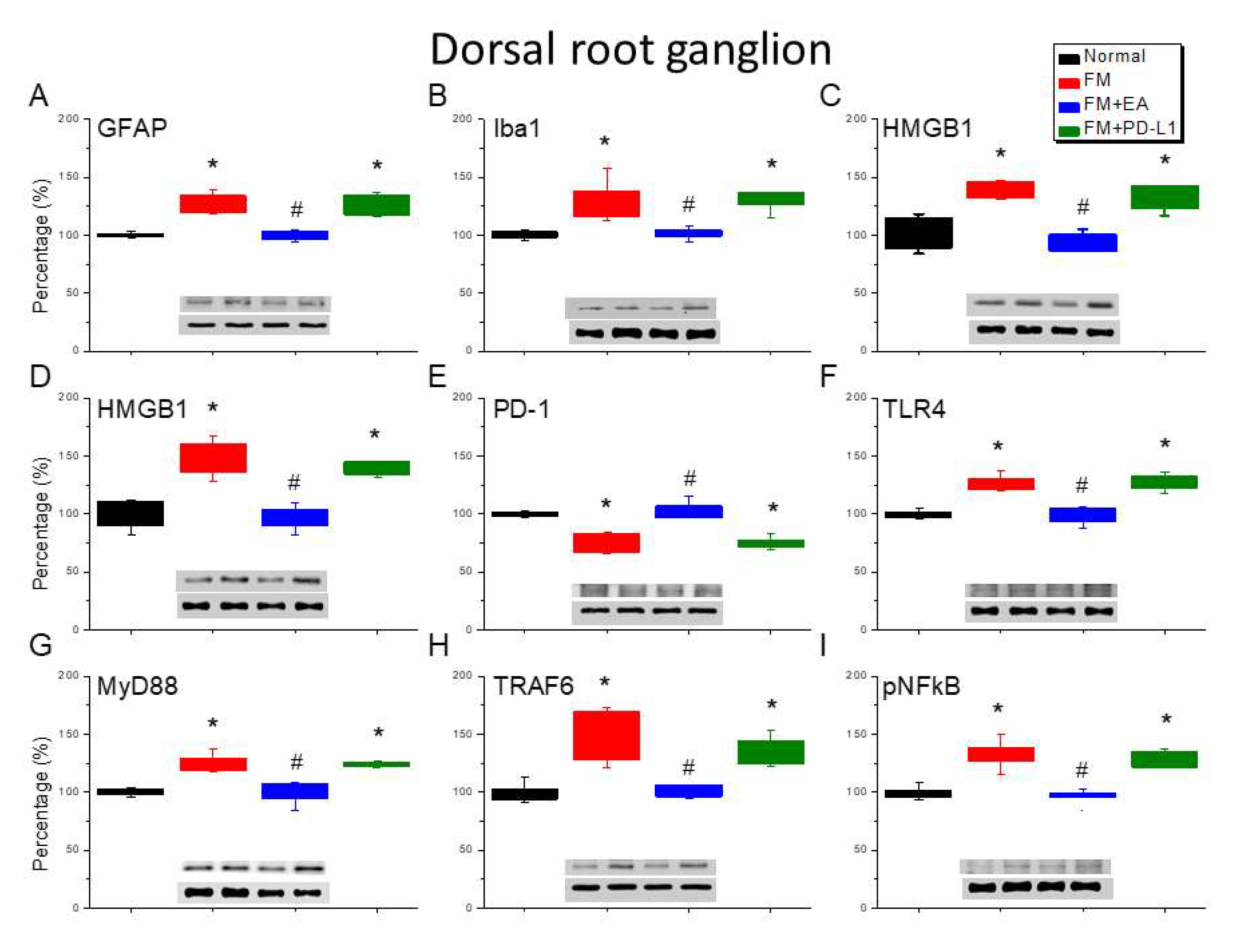
Figure 3.
Changing protein levels of PD-1 on pain-related molecules in the spinsl cord (SC) of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 3.
Changing protein levels of PD-1 on pain-related molecules in the spinsl cord (SC) of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
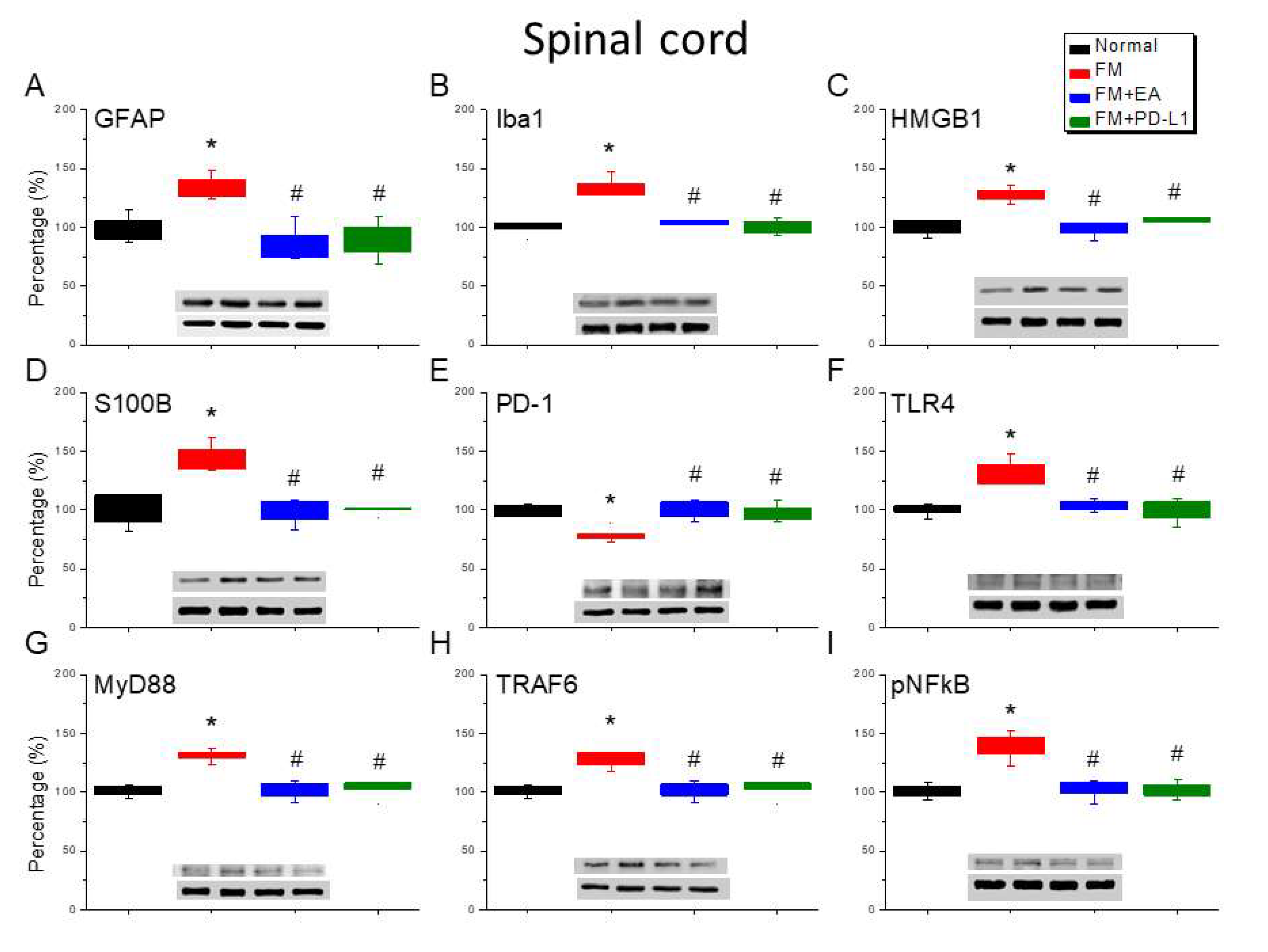
Figure 4.
Changing protein levels of PD-1 on pain-related molecules in the thalamus of mice in all groups including four lanes of A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 4.
Changing protein levels of PD-1 on pain-related molecules in the thalamus of mice in all groups including four lanes of A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 5.
Changing protein levels of PD-1 on pain-related molecules in the somatosensory cortex of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 5.
Changing protein levels of PD-1 on pain-related molecules in the somatosensory cortex of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
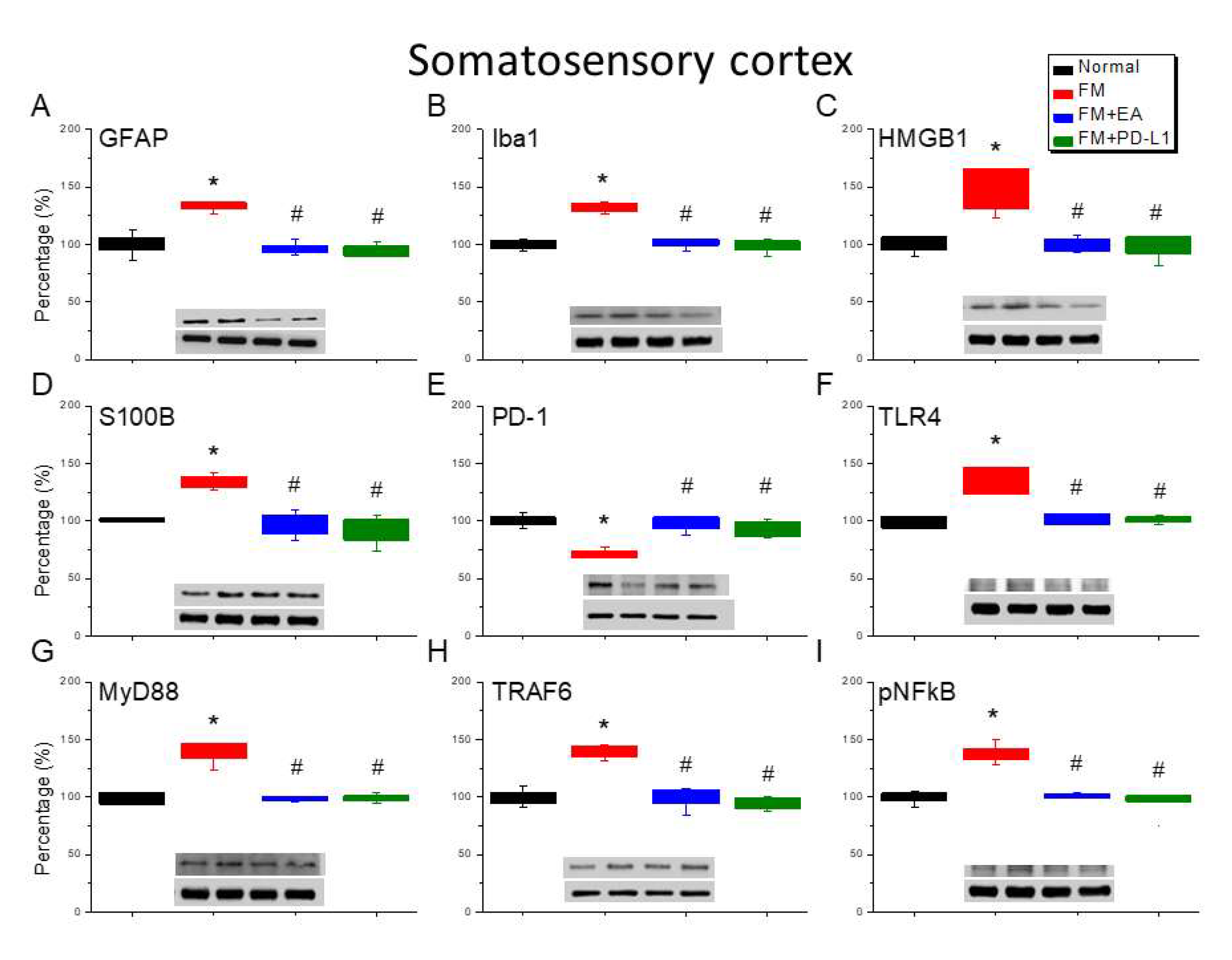
Figure 6.
Altering protein levels of PD-1 on pain-linked proteins in the CB5 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 6.
Altering protein levels of PD-1 on pain-linked proteins in the CB5 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
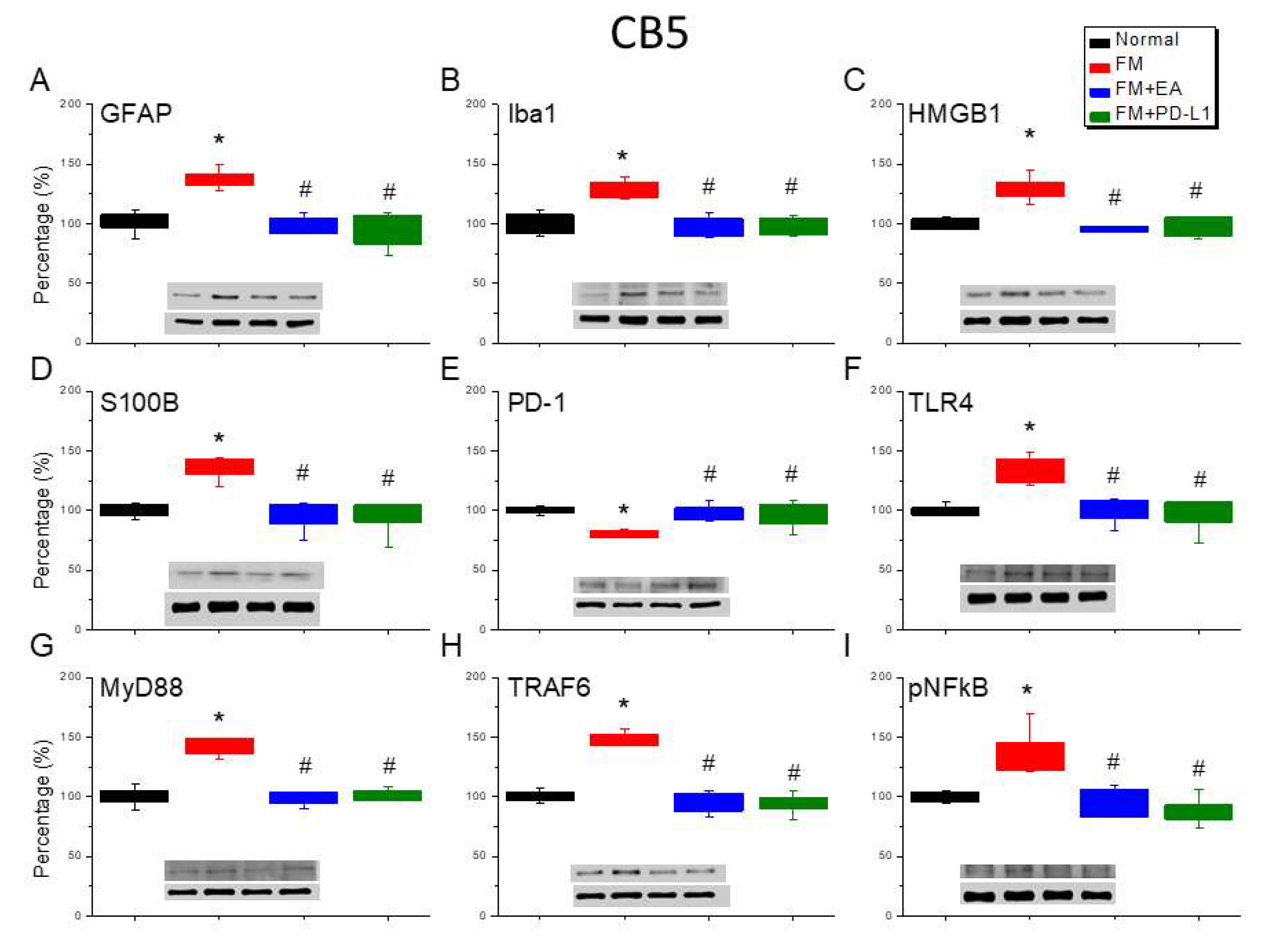
Figure 7.
Varying protein concentrations of PD-1 on pain-associated proteins in the CB6 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 7.
Varying protein concentrations of PD-1 on pain-associated proteins in the CB6 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
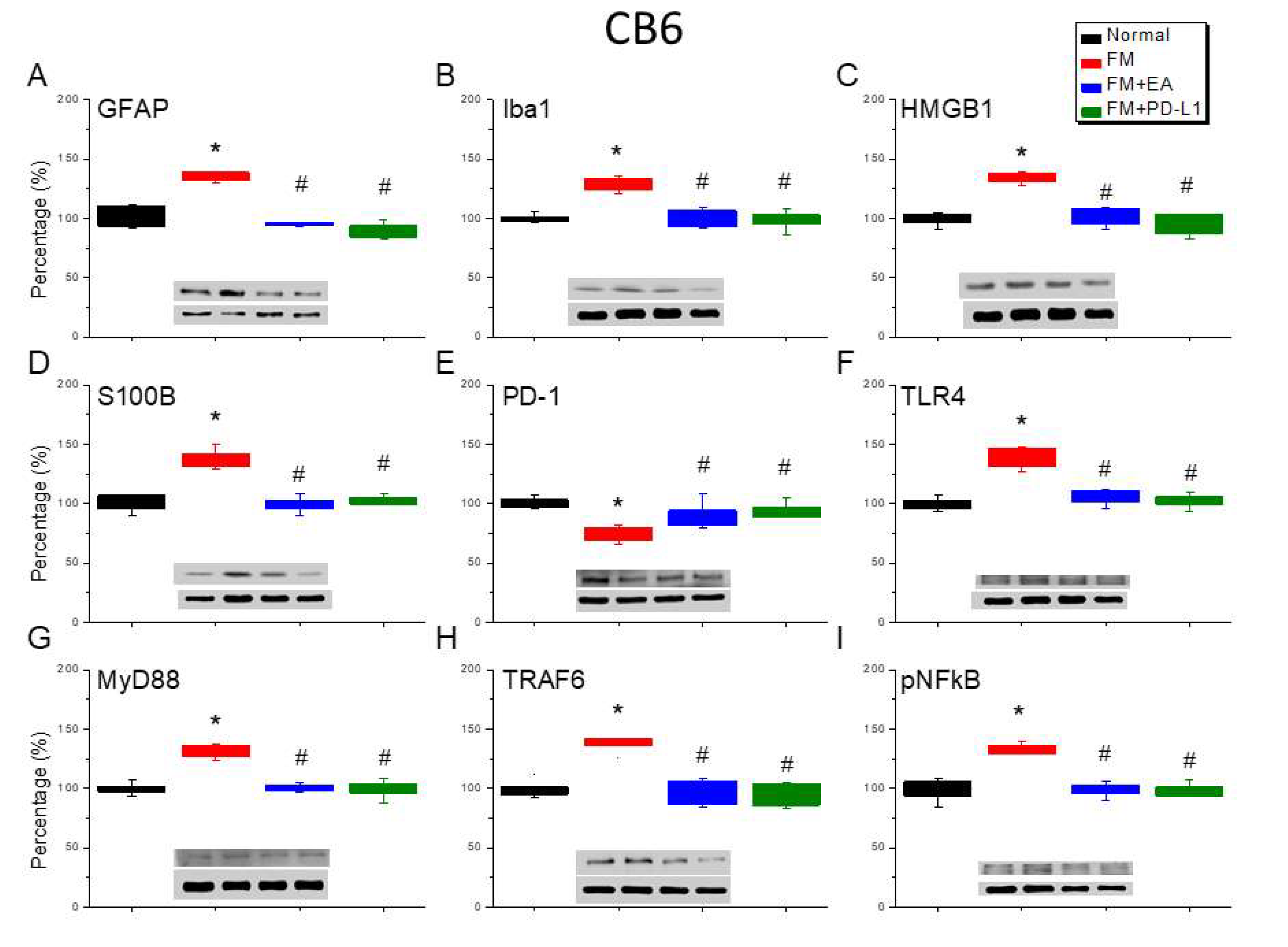
Figure 8.
Modifying protein levels of PD-1 on pain-associated proteins in the CB7 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
Figure 8.
Modifying protein levels of PD-1 on pain-associated proteins in the CB7 of mice in all groups including four lanes of (A) GFAP, (B) Iba1, (C) S100B, (D) HMGB1, (E) PD-1, (F) TLR4, (G) MyD88, (H) TRAF6, (I) pNF-κB protein levels in Normal, FM, FM + EA, and FM + PD-L1. *P < 0.05 vs the normal group, #P < 0.05 vs the FM group.
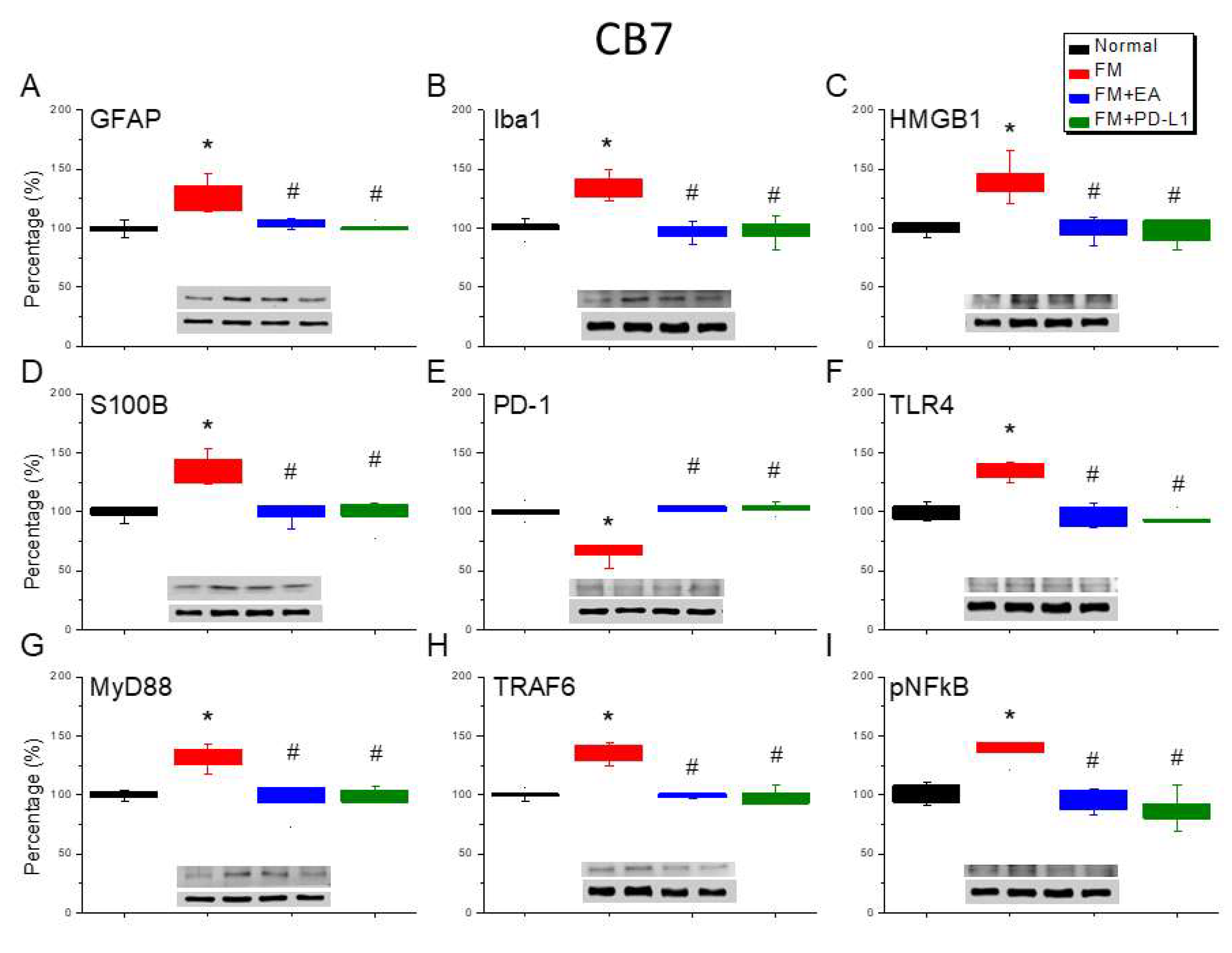
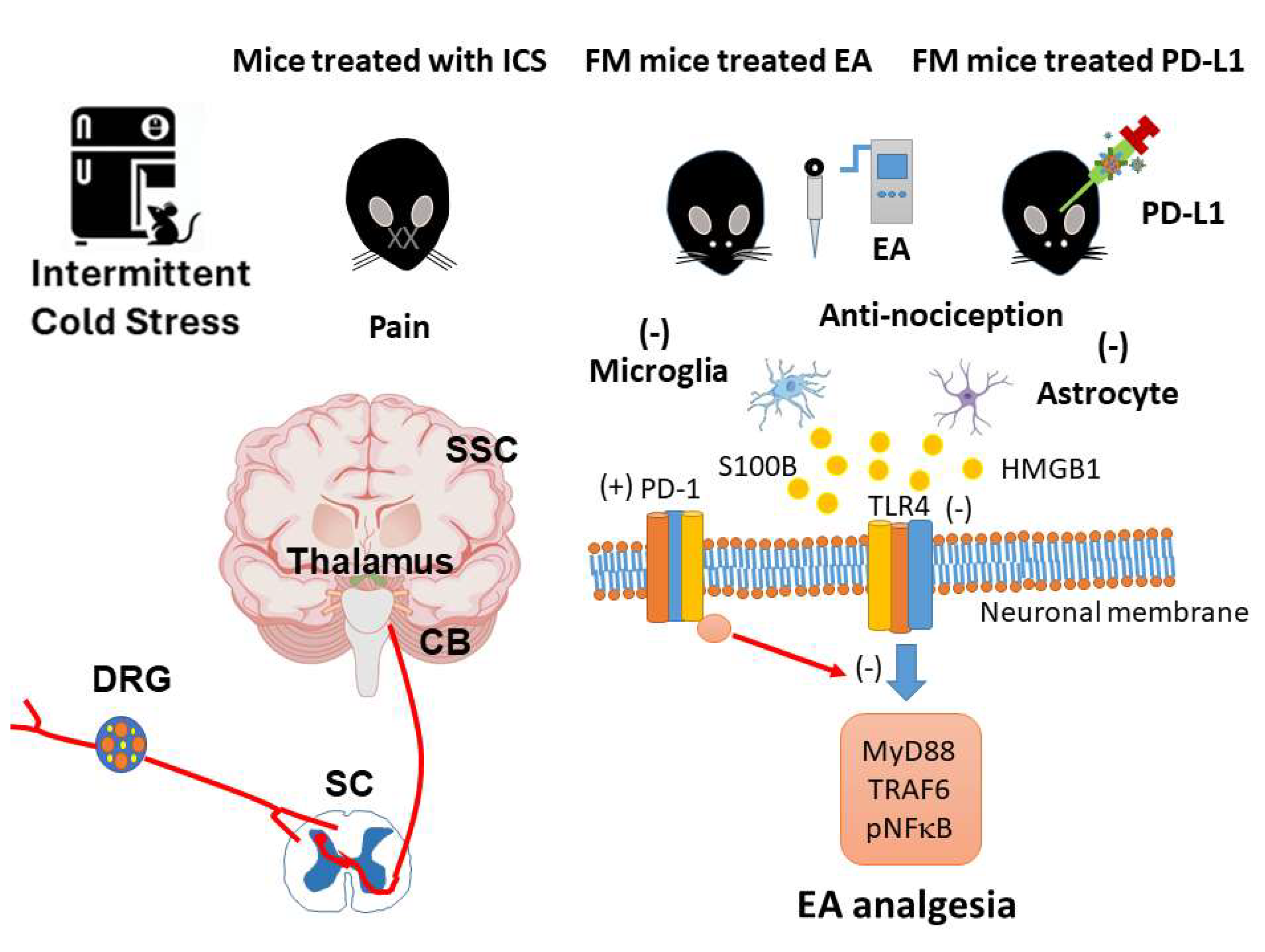
Figure 9.
The role of EA on PD-1 signaling pathway in mice fibromyalgia model. Abbreviations: EA = Electroacupuncture; TLR4 = Toll-like receptor 4; PD-L1 = Programmed cell death ligand 1; PD-1 = Programmed cell death protein 1; MyD88 = Myeloid differentiation primary response 88; TRAF6 = TNF Receptor Associated Factor 6; pNF-kB = phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells.
Figure 9.
The role of EA on PD-1 signaling pathway in mice fibromyalgia model. Abbreviations: EA = Electroacupuncture; TLR4 = Toll-like receptor 4; PD-L1 = Programmed cell death ligand 1; PD-1 = Programmed cell death protein 1; MyD88 = Myeloid differentiation primary response 88; TRAF6 = TNF Receptor Associated Factor 6; pNF-kB = phosphorylated nuclear factor kappa-light-chain-enhancer of activated B cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



