
Submitted:
30 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Background: The human oral microbiome is a critical reservoir for antibiotic resistance, however, subgingival peri-implant biofilms remain underexplored in this context. We aimed to explore the prevalence and distribution of antibiotic resistance genes (ARGs) in metagenomes derived from saliva and subgingival peri-implant biofilms. Methods: A total of 100 metagenome datasets from 40 individuals were retrieved from the Sequence Read Archive (SRA) database. Of these, 20 individuals had exclusively healthy implants and 20 had both healthy and affected implants with peri-implantitis. ARGs and their taxonomic assignments were identified using the ABRicate tool, and plasmid detection was performed with PlasmidFinder. Results: Four plasmid replicons were identified in 72 metagenomes, and 55 distinct ARGs from 13 antibiotic classes were detected in 89 metagenomes. The repUS43 plasmid replicon and ARGs conferring resistance to macrolides-lincosamides-streptogramins, tetracyclines, beta-lactams, and fluoroquinolones were the most prevalent. The msr(D) and mef(A) genes showed the highest prevalence, except in saliva samples from individuals with healthy implants, where mef(A) ranked fourth. Saliva samples harbored greater ARG diversity than subgingival biofilm samples. Principal Coordinates Analysis of Jaccard distances revealed no significant differences between healthy and peri-implantitis-affected subgingival biofilm groups (pairwise PERMANOVA, p > 0.05). The taxonomic origins of the detected ARGs were also obtained to better understand their distribution and potential impact on oral microbial communities. Conclusions: Resistome profiles associated with both peri-implant health and disease showed no significant differences and higher salivary abundance of ARGs compared to subgingival biofilm samples.
Keywords:
antibiotic resistance genes (ARGs)
; peri-implantitis
; oral metagenomes
; oral resistome
; saliva
; subgingival peri-implant biofilm
1. Introduction
The oral microbiome is among the most diverse in the human body and plays a critical role in maintaining oral health. The microbial community composing the oral microbiome is typically arranged in biofilms. Oral biofilms form not only on natural teeth but also on restorative materials, dental prosthetic devices, and dental implants [1,2]. Bacteria growing within a biofilm often exhibit altered phenotypes, being more tolerant to antimicrobial agents, host immune defense mechanisms, and mechanical removal [3]. The structural properties and proximity of the bacterial cells within the biofilm facilitate horizontal gene transfer, which can lead to the spread of antibiotic resistance genes (ARGs) [1,4]. Furthermore, numerous ARGs are found on mobile genetic elements (MGEs), such as transposons and conjugative elements (e.g., plasmids), facilitating their efficient transfer between cells, including from non-pathogenic and commensal bacteria to pathogenic species [5].
Therefore, the oral microbiome is increasingly recognized as a significant reservoir of antibiotic resistance, encompassing the oral resistome, which comprises all ARGs present within this microbial community [6,7,8,9]. A comprehensive evaluation of the distribution of ARGs in specific niches in the oral cavity is a useful step toward understanding the oral resistome. Still, there is currently limited understanding of the oral resistome in its entirety [10,11]. Similar to oral microbiome, the oral resistome is highly individualized [12], limiting its general knowledge. In the era of personalized medicine, integrating oral health is paramount due to the unique and dynamic nature of the oral microbiome, which maintains a delicate balance with the host to promote health [12]. Advances in genomic technologies, particularly next-generation sequencing (NGS), are transforming our understanding of the microbiome and driving the emergence of microbiome medicine, which aims to leverage human microbiota and their derived molecules for the prevention and treatment of diseases [13]. These tools are also essential for decoding the oral resistome and its role in antimicrobial resistance (AMR) [11].
In clinical dental practice, antibiotics are frequently prescribed to treat oral infections or prophylactically to prevent systemic and local complications associated with dental procedures [6,14]. However, these prescriptions are often based on empirical clinical judgment rather than definitive diagnoses, contributing to the growing concern over inappropriate antibiotic use in dentistry and the consequent pressing need for dental antibiotic stewardship [14,15].
Here, we performed a data analysis on 100 metagenomes that we previously obtained by shotgun metagenomics and deposited on the National Centre for Biotechnology Information (NCBI) SRA database. These metagenomes were derived from saliva and subgingival peri-implant biofilm samples collected from patients with healthy implants (HI group) and those with a mix of peri-implantitis-affected implants and healthy implants (PI group). This study aimed to identify ARGs and plasmids within metagenomic datasets and attribute taxonomic origins to the detected ARGs.
2. Materials and Methods
2.1. Data Acquisition and Information
A total of 100 metagenome datasets with a read length of 150 bp that we have previously obtained and uploaded to the NCBI SRA database (https://www.ncbi.nlm.nih.gov/sra) under NCBI BioProject number PRJNA1163384, were used in this study (https://www.ncbi.nlm.nih.gov/sra/?term=PRJNA1163384). The datasets were derived from 40 individuals from Portugal with one or more dental implants. Of these, 20 individuals had exclusively healthy implants (HI group), while the remaining 20 had both healthy implants and implants affected by peri-implantitis (PI group). Each participant provided a saliva sample and one (in the HI group) or two (in the PI group) subgingival biofilm samples. Each biosample or metagenome was categorized into one of five study groups: HI_Sa, HI_HIS, PI_Sa, PI_HIS, and PI_PIS. The abbreviations are as follows: 'Sa' for saliva, 'HIS' for healthy implant site, and 'PIS' for peri-implantitis affected site. Detailed information, including accession numbers, is provided in Table S1 in the Supplementary Materials.
2.2. Data Analysis and Statistics
Sequenced reads were quality-filtered with Trimmomatic version 0.39 [16] using the following parameters: 1) sequencing adapters were removed, 2) bases with an average quality lower than Q25 in a window of 5 bases were trimmed, and 3) reads with less than 100 bases were discarded. High-quality reads were filtered against the reference human genome sequence assembly GRCh38/hg38 with Bowtie version 2.5 [17]. High-quality sequences from each sample were de novo assembled with metaSPAdes version 3.15.5 [18], using default parameters. Metagenome assembled contigs were analyzed for the presence of ARGs with ABRicate version 1.0.1 [19] with the ResFinder [20], CARD [21], NCBI [22], and ARG-ANNOT [23] databases. ARGs were identified with a sequence identity of at least 90% and coverage of at least 80%. Taxonomic classification of genes was provided for each database result. ARGs were classified into antibiotic resistance classes according to the CARD database and literature searches. The taxonomic attribution of each identified ARG was retrieved directly from the ABRicate output results or from the GenBank accession numbers also present in the ABRicate outputs. Metagenome assembled contigs were also analyzed for the presence of plasmids with PlasmidFinder within the staramr package version 0.10 [24]. Plasmids were searched against the complete PlasmidFinder database version 2.2 [25]. Plasmids were identified with a sequence identity of at least 90% and coverage of at least 80%. The plasmidome was characterized through prevalence analysis per study group.
The resistome was characterized through prevalence analysis at the gene and antibiotic class level per study group. Differences between group prevalences were analyzed by Principal Coordinates Analysis (PCoA) in phyloseq version 1.44.0 [26] using the Jaccard similarity coefficient. The indexes were tested for statistical differences with PERMANOVA, followed by pairwise PERMANOVA using the adonis function of the vegan package version 2.6-4 [27] with 1000 permutations and the Benjamini-Hochberg procedure for multiple comparison corrections. Homoscedasticity was tested with the betadisper function of the vegan package. The differential prevalence of genes between study groups was further analyzed by Firth’s logistic regression with the logistf package, version 1.26.0 [28]. Multiple comparisons were corrected using the Benjamini-Hochberg procedure.
A Circos representation of the prevalence of antibiotic classes and study groups was created on the online version of the tool [29].
Beta diversity (Jaccard distance), prevalence and Firth´s logistic regression analysis were performed using R Statistical Software version 4.4.1 [30] in RStudio version 2024.09.0 build 375 [31]. Plots were produced with ggplot2 version 3.5.1 [32]. A p-value and an adjusted p-value of < 0.05 were considered statistically significant.
3. Results
3.1. Detection and Prevalence of ARGs and Plasmids Across Study Groups
ARGs were detected in 89 out of the 100 samples, while plasmids were identified in 72. Samples with no detectable ARGs or plasmids are listed in Supplementary Materials, Table S2 and Table S3, respectively.
Only four plasmids were identified in the samples. The most prevalent was repUS43, followed by repUS38 and repUS34. The repUS37 plasmid replicon was only found in one sample of group HI_Sa (Figure S1 in the Supplementary Materials). These plasmids typically harbor multiple ARGs; for instance, the repUS43 plasmid encodes both tet(M) and erm(B), which confer resistance to tetracyclines and macrolides-lincosamides-streptogramins (MLS), respectively [33]. This genetic composition significantly contributes to multidrug resistance and its spread among bacterial populations.
Overall, 55 different ARGs conferring resistance to 13 antibiotic classes were found. The ARGs and corresponding specific antibiotic classes are listed in Table S4 in the Supplementary Materials. All 89 samples with ARGs harbored multiple distinct ARGs.
ARGs conferring resistance to MLS, macrolides, tetracyclines, beta-lactams and fluoroquinolones were most prevalent in all study groups, and in particular in saliva samples (HI_Sa and PI_Sa groups) as shown in Figure 1, Figure S2 and Figure S3 in the Supplementary Materials. ARGs conveying resistance to pleuromutilins-streptogramins-lincosamides (PSL), phenicols, nucleoside antibiotics and cephalosporins were less prevalent across all study groups. Notably, aminoglycoside resistance genes were particularly less prevalent in the PI_Sa and PI_PIS groups (Figure 1 and Figure S3 in the Supplementary Materials).
The prevalence of ARGs per study group is shown in Figure 2. Among the total of 55 ARGs identified, 19 were shared across all study groups.
The msr(D) and mef(A) genes ranked highest in prevalence across all study groups, except in the HI_Sa group, where mef(A) was the fourth most common. Notably, it has been consistently reported that in mef-carrying genetic elements, the msr(D) gene is invariably associated with and co-transcribed with the mef(A) gene, playing a key role in macrolide efflux resistance [34,35,36]. The mef(A) and msr(D) genes form a two-component ATP-binding cassette (ABC) efflux transport system, with mef(A) encoding the transmembrane channel and msr(D) encoding the two ATP-binding domains. However, in the absence of Mef(A), Msr(D) likely utilizes an alternative transmembrane channel for macrolide efflux [34].
Saliva groups (HI_Sa and PI_Sa) harbored a higher number of ARGs than the subgingival peri-implant biofilm groups (PI_HIS, PI_PIS and HI_HIS). Peri-implantitis-affected implants (PI_PIS group) generally displayed a higher prevalence of most ARGs than healthy implants (PI_HIS and HI_HIS groups). A PCoA based on the Jaccard distance was performed to evaluate differences in group prevalences (Figure S4, Supplementary Materials), and PERMANOVA analysis indicated significant overall differences (p = 0.001). However, pairwise PERMANOVA revealed no significant differences between any two subgingival biofilm groups or between saliva groups (p > 0.05) (Table S5 in the Supplementary Materials).
Ten ARGs were differentially prevalent among study groups (Figure 3).
The genes tet(W), erm(X), mrcB, and tetA(60) were detected in the HI_Sa group but were absent in the HI_HIS group. The genes tet(B) and mrcB were only detected in saliva groups.
3.2. Taxonomic Assignment of Identified ARGs
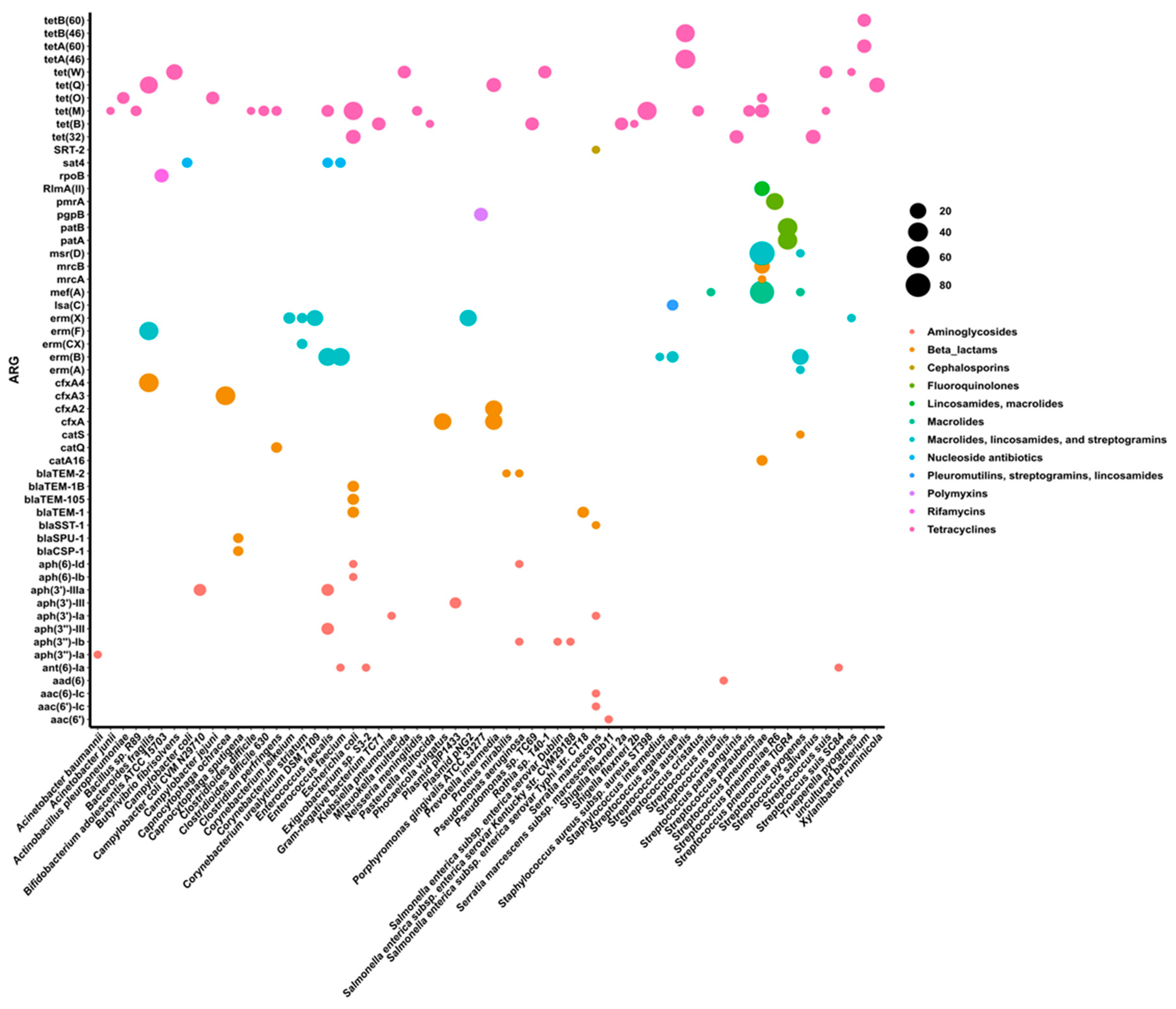
To estimate the taxonomy of ARGs, the taxonomic attribution for each identified ARG was directly retrieved from ABRicate. Figure 4 displays each ARG along with its corresponding bacterial species and its prevalence within each species across all samples, regardless of the study group.
Several distinct ARGs were associated with individual bacterial species. For example, Escherichia coli was linked to tet(M), tet(32), blaTEM-1, blaTEM-1B, blaTEM-105, aph(6)-Id, and aph(6)-Ib; Enterococcus faecalis was associated with tet(M), sat4, erm(B), aph(3’)-IIIa, and aph(3’’)-III; and Streptococcus pneumoniae with tet(O), tet(M), RlmA(II), msr(D), mef(A), mrcB, mrcA, and catA16. Some ARGs were also specifically associated with oral bacterial species, including Porphyromonas gingivalis (pgpB), Prevotella intermedia (tet(Q), cfxA, cfxA2), and Streptococcus salivarius and S. parasanguinis (tet(32)). In a previous study [9], P. gingivalis was also associated with the pgpB gene, which encodes resistance to peptide antibiotics such as polymyxins. Also consistent with other research, the production of β-lactamase in Prevotella species has been linked to the expression of the cfxA and cfxA2 genes [37,38]. Notably, the pgpB, cfxA, and cfxA2 genes were detected across all study groups, as shown in Figure 2.
Genes encoding resistance to tetracyclines stand out due to their widespread presence across a diverse range of bacterial species (Figure 4). The gene tet(M) was assigned to 13 different species.
The erm(X) gene was predominantly associated with Corynebacterium urealyticum and the Corynebacterium diphtheriae plasmid, the plasmid pNG2. This plasmid is known to mediate inducible resistance to erythromycin and other macrolide-lincosamide-streptogramin (MLS) antibiotics [39]. Additionally, the aph(3’)-III gene, encoding resistance towards aminoglycosides was also identified within a plasmid, the plasmid plP1433 [40].
4. Discussion
In this secondary study, metagenomes from saliva and subgingival peri-implant biofilm samples were explored for the presence of ARGs and plasmids. This first metagenomic sequencing study reports implants affected by peri-implantitis generally exhibited a higher prevalence of most ARGs compared to healthy implants but were not statistically significant. However, dental implants were confirmed as reservoirs for ARGs and plasmids. We also confirmed that salivary samples present higher levels of ARGs and plasmids compared to subgingival samples.
To gain insights into the ARG profiles of these samples we integrated multiple databases (including ABRicate, CARD, ResFinder, NCBI, and ARG-ANNOT), a common procedure to enhance the comprehensiveness and accuracy of ARG detection [13]. We used PlasmidFinder database to identify plasmid replicons in the metagenomes [41].
Three plasmid replicons (repUS43, repUS38, and repUS34) were found in all study groups; however, they were slightly more prevalent in salivary than in subgingival peri-implant samples. The repUS43 replicon is a conjugative plasmid frequently identified in Enterococcus spp. isolates [42] and is associated with ARGs, including tet(M) and erm(B) [28]. Conjugative plasmids are self-transmissible, enabling their transfer between bacterial cells through conjugation. Consequently, plasmids like repUS43, which carry multiple ARGs, may play a pivotal role in disseminating antibiotic resistance, particularly in these bacterial communities of the oral cavity. Their presence in this niche facilitates horizontal gene transfer among resident and pathogenic bacteria, further complicating treatment strategies and posing a significant challenge to the clinical management of oral and systemic infections.
The metagenomic analysis revealed a high prevalence of antibiotic resistance genes (ARGs) across all five study groups. In total, ARGs conferring resistance to 13 antibiotic classes were identified, with macrolides-lincosamides-streptogramins (MLS), macrolides, tetracyclines, beta-lactams, and fluoroquinolones being the most prevalent. These antibiotic classes are ranked by the World Health Organization (WHO) as critically or highly important for human medicine [43]. The most commonly detected ARGs were associated with mechanisms such as antibiotic efflux (e.g., mef(A), msr(D), patA, and patB), antibiotic inactivation via beta-lactamases (cfxA, cfxA3), and inhibition of protein synthesis (tet(M) and erm(F)).
The analyzed metagenomes were obtained from samples collected from individuals in Portugal. A recent cross-sectional web survey revealed that 64% of Portuguese dentists consistently use antibiotic prophylaxis for dental implant placement, 29% prescribe antibiotics postoperatively, and 55% use both pre- and postoperative regimens. Amoxicillin combined with clavulanic acid was the most prescribed antibiotic (57%) [44]. Similarly, a questionnaire-based study investigating European dentists' prescribing practices for dental implant treatments found that amoxicillin alone (61.0%) or combined with clavulanic acid (55.9%) were the most commonly prescribed antibiotics, followed by clindamycin (32.3%), erythromycin, and azithromycin [45]. These findings highlight a high rate of antibiotic prescriptions for dental implant procedures. Such overprescription contributes significantly to the rise of antibiotic resistance and the prevalence of antibiotic resistance genes (ARGs) in both healthy and diseased oral environments.
To our knowledge, this is the first study to report the distribution patterns of ARGs in both healthy implants and those affected by peri-implantitis based on shotgun metagenomic sequencing data. Nevertheless, similar metagenomic studies have been conducted on dental plaque from periodontal healthy and diseased sites to characterize its resistome [7,9,46]. Kang et al. [7] reported that resistance genes for bacitracin, beta-lactams, MLS, and tetracyclines were present in almost all dental plaque samples at relatively high abundances. However, periodontitis was associated with an increased number of ARGs and significant alterations in the composition of ARGs in dental plaque compared to the healthy condition [7]. In another study, Anderson et al. [9] identified mef(A), msr(D), cfxA, and erm(F) as the most prevalent ARGs in healthy oral biofilm, whereas tet(Q), pgpB, and tet(32) were more common in cases of periodontitis. Gager et al. [46] examined the prevalence of ARGs in the subgingival biofilm of a population of German patients diagnosed with periodontitis. Their study revealed that macrolide resistance genes were the most prevalent and that two-thirds of the participants carried ARGs associated with at least one class of antibiotics commonly used in periodontal treatment and research, including beta-lactams, lincosamides, macrolides, nitroimidazoles, and tetracyclines.
A total of 40 different ARGs were identified in saliva samples from both the HI_Sa and PI_Sa groups, representing a higher number than those detected in the subgingival peri-implant biofilm groups (26 in HI_HIS, 32 in PI_HIS, and 30 in PI_PIS). The genes tet(B) and mrcB were detected in saliva groups but not in the subgingival biofilm groups. In both saliva groups, the majority of ARGs, including the most prevalent ones, were shared between the two groups. Regarding antibiotic classes, the order of prevalence in the HI_Sa group was MLS, beta-lactams, tetracyclines, fluoroquinolones, and macrolides. In the PI_Sa group, the order was MLS, tetracyclines, macrolides, beta-lactams, and fluoroquinolones. Similarly, Caselli et al. [47], using a microarray approach to analyze the oral resistome in oral rinse samples (not saliva) from healthy Italians, also reported a high prevalence of resistance to MLS and tetracyclines.
The tet(M) gene was the most prevalent tetracycline resistance gene in all three subgingival peri-implant biofilm groups, while in the saliva groups, tetA(46) was predominant. Similarly, Anderson et al. [9] reported that tet(M) was the most abundant tetracycline resistance gene in healthy dental plaque samples. Kang et al. [7] also observed a high prevalence of tet(M) in dental plaque samples both with and without periodontitis. Notably, the tet(M) gene is often linked to the erm(B) gene, as both are frequently found on conjugative plasmids or transposons, such as repUS43 and Tn916 [8]. This association suggests that the abundance of tetracycline resistance genes in the dental plaque resistome may result from coselection pressures. The use of erythromycin, for example, can coselect for tetracycline resistance, which could explain the high prevalence of tet(M) in oral metagenomes [7,8]. Consistent with this, the tet genes identified in our studied metagenomes were widely distributed and taxonomically associated with various bacterial species, encompassing both Gram-positive and Gram-negative bacteria. Particularly, the tet(M) gene was assigned to 13 different bacterial species.
Based on the taxonomic attribution of each identified ARG, we identified 62 bacterial species and two plasmids as potential hosts for 55 ARGs. The taxonomic assignment of ARGs revealed that typical commensal taxa, such as Rothia spp. and Bifidobacterium spp., were associated with resistance genes for tetracyclines and rifamycins, respectively. Gram-positive bacteria, including Streptococcus spp., Enterococcus spp., Corynebacterium spp., and Clostridioides spp., carried a diverse array of ARGs conferring resistance to MLS and tetracyclines. Additionally, ARGs conferring resistance to macrolides, lincosamides, and fluoroquinolones were attributed to Streptococcus pneumoniae. Among Gram-negative bacteria, Escherichia coli and Prevotella intermedia predominantly harbored ARGs for beta-lactam and tetracycline resistance. Notably, resistance to polymyxins was exclusively associated with Porphyromonas gingivalis.
These putative ARG-carrying pathogens have the potential to compromise the efficacy of commonly used antibiotics, highlighting the need for targeted antimicrobial strategies and the judicious use of antibiotics in dental implantology. It is important to note, however, that bacteria may harbor an ARG that is not expressed, meaning it would not result in phenotypic resistance [48]. Nonetheless, Anderson et al. [9] reported a notably high concordance of nearly 60% between genotypic and phenotypic analyses. Furthermore, the same study highlighted that while metagenomic analysis has greatly advanced our understanding of the distribution and diversity of ARGs within the microbiome, it has limitations. Specifically, current bioinformatic methods are often unable to detect point mutations, which means resistance to certain antibiotics, such as vancomycin, may go undetected at the genomic level despite being phenotypically present [9]. This limitation also applies to our study, as we used the ABRicate tool, which is designed to detect acquired resistance genes but is not capable of identifying point mutations [9].
Dental implants can serve as significant reservoirs for biofilm-associated ARGs, posing potential risks not only to oral health but also to systemic health. Biofilms formed on dental implants provide a protective environment for ARG-carrying bacteria, enabling their persistence, horizontal gene transfer and potential dissemination. Similar to findings in periodontitis, where periodontal bacteria have been implicated in complications such as ventilator-associated pneumonia due to their migration to the respiratory tract, implant-associated biofilms may contribute to analogous risks [49,50,51,52]. The ability of peri-implant bacteria to enter systemic circulation or adjacent anatomical regions underscores the importance of understanding and mitigating the role of dental implants as reservoirs of ARGs. Further studies are required to elucidate the extent to which implant-associated biofilms contribute to systemic antibiotic resistance and related health complications.
5. Conclusions
Comparable levels of ARGs were observed in both healthy and peri-implantitis subgingival and salivary samples. Salivary ARG profiles were higher in abundance and prevalence of ARGs than in subgingival biofilms. The most prevalent ARGs were those conferring resistance to macrolides-lincosamides-streptogramins, tetracyclines, beta-lactams, and fluoroquinolones. Future studies should explore the functional impact of these ARGs and clinical relevance in dental and medical settings.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, L.J.B., J.B.; V.M.; G.A., R.A. and J.J.M.; methodology, L.J.B. and C.E.; formal analysis, C.E.; investigation, L.J.B.; resources, R.A. and J.J.M.; data curation, C.E.; writing—original draft preparation, L.J.B.; writing—review and editing, C.E.; J.B., V.M., G.A. and R.A.; visualization, L.J.B.; project administration, L.J.B.; funding acquisition, L.J.B. and R.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Portuguese National Funds provided by the FCT (Fundação para a Ciência e Tecnologia) under grant number 2022.01430.PTDC for the project "Metagenomic snapshot of the microbiome and resistome in peri-implantitis: pilot study” (http://doi.org/10.54499/2022.01430.PTDC).
Informed Consent Statement
Not applicable.
Data Availability Statement
The metagenomic data are available in the NCBI SRA database (https://www.ncbi.nlm.nih.gov/sra) under NCBI BioProject number PRJNA1163384 (https://www.ncbi.nlm.nih.gov/sra/?term=PRJNA1163384).
Conflicts of Interest
The author João Botelho is a Special Issue Editor for the journal.
References
- Roberts, A.P.; Mullany, P. Oral Biofilms: A Reservoir of Transferable, Bacterial, Antimicrobial Resistance. Expert Rev. Anti. Infect. Ther. 2010, 8, 1441–1450. [CrossRef]
- Sharma, N.; Bhatia, S.; Sodhi, A.S.; Batra, N. Oral Microbiome and Health. AIMS Microbiol. 2018, 4, 42–66. [CrossRef]
- Olsen, I. Biofilm-Specific Antibiotic Tolerance and Resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [CrossRef]
- Michaelis, C.; Grohmann, E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms. Antibiotics 2023, 12. [CrossRef]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for Enhanced Metagenomic Taxonomic Profiling. Nat. Methods 2015, 12, 902–903. [CrossRef]
- Bessa, L.J.; Botelho, J.; Machado, V.; Alves, R.; Mendes, J.J. Managing Oral Health in the Context of Antimicrobial Resistance. Int. J. Environ. Res. Public Health 2022, 19, 16448. [CrossRef]
- Kang, Y.; Sun, B.; Chen, Y.; Lou, Y.; Zheng, M.; Li, Z. Dental Plaque Microbial Resistomes of Periodontal Health and Disease and Their Changes after Scaling and Root Planing Therapy. mSphere 2021, 6, 1–20. [CrossRef]
- Sukumar, S.; Roberts, A.P.; Martin, F.E.; Adler, C.J. Metagenomic Insights into Transferable Antibiotic Resistance in Oral Bacteria. J. Dent. Res. 2016, 95, 969–976. [CrossRef]
- Anderson, A.C.; von Ohle, C.; Frese, C.; Boutin, S.; Bridson, C.; Schoilew, K.; Peikert, S.A.; Hellwig, E.; Pelz, K.; Wittmer, A.; et al. The Oral Microbiota Is a Reservoir for Antimicrobial Resistance: Resistome and Phenotypic Resistance Characteristics of Oral Biofilm in Health, Caries, and Periodontitis. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 1–19. [CrossRef]
- Sukumar, S.; Wang, F.; Simpson, C.A.; Willet, C.E.; Chew, T.; Hughes, T.E.; Bockmann, M.R.; Sadsad, R.; Martin, F.E.; Lydecker, H.W.; et al. Development of the Oral Resistome during the First Decade of Life. Nat. Commun. 2023, 14, 1–14. [CrossRef]
- Sukumar, S.; Rahmanyar, Z.; El Jurf, H.Q.; Akil, W.S.; Hussain, J.; Elizabeth Martin, F.; Ekanayake, K.; Martinez, E. Mapping the Oral Resistome: A Systematic Review. J. Med. Microbiol. 2024, 73, 1–10. [CrossRef]
- Belibasakis, G.N.; Bostanci, N.; Marsh, P.D.; Zaura, E. Applications of the Oral Microbiome in Personalized Dentistry. Arch. Oral Biol. 2019, 104, 7–12. [CrossRef]
- Kim, N.; Ma, J.; Kim, W.; Kim, J.; Belenky, P.; Lee, I. Genome-Resolved Metagenomics : A Game Changer for Microbiome Medicine. Exp. Mol. Med. 2024, 56, pages1501–1512. [CrossRef]
- Thompson, W.; Teoh, L.; Pulcini, C.; Williams, D.; Pitkeathley, C.; Carter, V.; Sanderson, S.; Torres, G.; Walsh, T. Dental Antibiotic Stewardship: Study Protocol for Developing International Consensus on a Core Outcome Set. Trials 2022, 23, 1–6. [CrossRef]
- Teoh, L.; Thompson, W.; Suda, K. Antimicrobial Stewardship in Dental Practice. J. Am. Dent. Assoc. 2020, 151, 589–595. [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [CrossRef]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 2017, 27, 824–834. [CrossRef]
- Seemann, T. ABRicate: mass screening of contigs for antibiotic resistance genes. 2016. Available online: https://github.com/tseemann/abricate.
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; Doshi, S.; Courtot, M.; Lo, R.; Williams, L.E.; Frye, J.G.; Elsayegh, T.; Sardar, D.; Westman, E.L.; Pawlowski, A.C.; Johnson, T.A.; Brinkman, F.S.; Wright, G.D.; McArthur, A.G. CARD 2017: Expansion and Model-Centric Curation of the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2017, 45, D566–D573. [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; Tadesse, D.A.; Morales, C.; Simmons, M.; Tillman, G.; Wasilenko, J.; Folster, J.P.; Klimke, W. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Resistance Genotype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, e00483–19 Erratum in: Antimicrob. Agents Chemother. 2020, 64, e00361–20, doi:10.1128/AAC.00361-20. [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT: A New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [CrossRef]
- Bharat, A.; Petkau, A.; Avery, B.P.; Chen, J.C.; Folster, J.P.; Carson, C.A.; Kearney, A.; Nadon, C.; Mabon, P.; Thiessen, J.; Alexander, D.C.; Allen, V.; El Bailey, S.; Bekal, S.; German, G.J.; Haldane, D.; Hoang, L.; Chui, L.; Minion, J.; Zahariadis, G.; Domselaar, V.; Reid-Smith, R.J.; Mulvey, M.R. Correlation between Phenotypic and In Silico Detection of Antimicrobial Resistance in Salmonella enterica in Canada Using Staramr. Microorganisms 2022, 10, 292. [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; Wagner, H. vegan: Community Ecology Package; R Package Version 2.6-4, 2020. Available online: https://CRAN.R-project.org/package=vegan.
- Heinze, G.; Ploner, M. logistf: Firth’s Bias-Reduced Logistic Regression. R Package Version 1.23.1, 2020. Available online: https://CRAN.R-project.org/package=logistf.
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://www.R-project.org/.
- Posit Team. RStudio: Integrated Development Environment for R; Posit Software, PBC: Boston, MA, 2023. Available online: https://posit.co/.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, 2016; ISBN 978-3-319-24277-4.
- Rofael, S.; Leboreiro Babe, C.; Davrandi, M.; Kondratiuk, A.L.; Cleaver, L.; Ahmed, N.; Atkinson, C.; McHugh, T.; Lowe, D.M. Antibiotic Resistance, Bacterial Transmission and Improved Prediction of Bacterial Infection in Patients with Antibody Deficiency. JAC Antimicrob. Resist. 2023, 5, dlad135. [CrossRef]
- Fox, V.; Santoro, F.; Pozzi, G.; Iannelli, F. Predicted Transmembrane Proteins with Homology to Mef(A) Are Not Responsible for Complementing mef(A) Deletion in the mef(A)–msr(D) Macrolide Efflux System in Streptococcus pneumoniae. BMC Res. Notes 2021, 14, 1–7. [CrossRef]
- Vitali, L.A.; Di Luca, M.C.; Prenna, M.; Petrelli, D. Correlation Between Genetic Features of the mef(A)–msr(D) Locus and Erythromycin Resistance in Streptococcus pyogenes. Diagn. Microbiol. Infect. Dis. 2016, 84, 57–62. [CrossRef]
- Iannelli, F.; Santoro, F.; Santagati, M.; Docquier, J.-D.; Lazzeri, E.; Pastore, G.; Cassone, M.; Oggioni, M.R.; Rossolini, G.M.; Stefani, S.; Pozzi, G. Type M Resistance to Macrolides Is Due to a Two-Gene Efflux Transport System of the ATP-Binding Cassette (ABC) Superfamily. Front. Microbiol. 2018, 9, 1670. [CrossRef]
- Iwahara, K.; Kuriyama, T.; Shimura, S.; Williams, D.W.; Yanagisawa, M.; Nakagawa, K.; Karasawa, T. Detection of cfxA and cfxA2, the β-Lactamase Genes of Prevotella spp., in Clinical Samples from Dentoalveolar Infection by Real-Time PCR. J. Clin. Microbiol. 2006, 44, 172–176. [CrossRef]
- Yokoyama, S.; Hayashi, M.; Goto, T.; Muto, Y.; Tanaka, K. Identification of cfxA Gene Variants and Susceptibility Patterns in β-Lactamase-Producing Prevotella Strains. Anaerobe 2023, 79, 102688. [CrossRef]
- Serwold-Davis, T.M.; Groman, N.B. Identification of a Methylase Gene for Erythromycin Resistance Within the Sequence of a Spontaneously Deleting Fragment of Corynebacterium diphtheriae Plasmid pNG2. FEMS Microbiol. Lett. 1988, 56, 7–13. [CrossRef]
- Trieu-Cuot, P.; Gerbaud, G.; Lambert, T.; Courvalin, P. In Vivo Transfer of Genetic Information Between Gram-Positive and Gram-Negative Bacteria. EMBO J. 1985, 4, 3583–3587. [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895-903. [CrossRef]
- Karlsson, P.A.; Zhang, T.; Järhult, J.D.; Joffré, E.; Wang, H. Heterogeneity and Metabolic Diversity Among Enterococcus Species During Long-Term Colonization. bioRxiv 2024. [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine: 6th Revision; World Health Organisation: Geneva, 2019.
- Correia, F.; Ribeiro-Vidal, H.; Gouveia, S.; Faria Almeida, R. Prescription of Antibiotic Prophylaxis in Implant Placement Among Portuguese Dentists: A Web Survey. Clin. Oral Implants Res. 2024, 35, 242–250. [CrossRef]
- Becker, K.; Gurzawska-Comis, K.; Klinge, B.; Lund, B.; Brunello, G. Patterns of Antibiotic Prescription in Implant Dentistry and Antibiotic Resistance Awareness Among European Dentists: A Questionnaire-Based Study. Clin. Oral Implants Res. 2024, 35, 771–780. [CrossRef]
- Gager, Y.; Koppe, J.; Vogl, I.; Gabert, J.; Jentsch, H. Antibiotic Resistance Genes in the Subgingival Microbiome and Implications for Periodontitis Therapy. J. Periodontol. 2023, 94, 1295–1301. [CrossRef]
- Caselli, E.; Fabbri, C.; Accolti, M.D.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the Oral Microbiome by Whole-Genome Sequencing and Resistome Analysis: The Complexity of the Healthy Picture. BMC Microbiol. 2020, 20, 120. [CrossRef]
- Fu, C.X.; Chen, C.; Xiang, Q.; Wang, Y.F.; Wang, L.; Qi, F.Y.; Zhu, D.; Li, H.Z.; Cui, L.; Hong, W.L.; et al. Antibiotic Resistance at Environmental Multi-Media Interfaces Through Integrated Genotype and Phenotype Analysis. J. Hazard. Mater. 2024, 480, 136160. [CrossRef]
- Scannapieco, F.A. Role of Oral Bacteria in Respiratory Infection. J. Periodontol. 1999, 70, 793–802. [CrossRef]
- Han, Y.W.; Wang, X. Mobile Microbiome: Oral Bacteria in Extra-Oral Infections and Inflammation. J. Dent. Res. 2013, 92, 485–491. [CrossRef]
- Molina, A.; Huck, O.; Herrera, D.; Montero, E. The Association Between Respiratory Diseases and Periodontitis: A Systematic Review and Meta-Analysis. J. Clin. Periodontol. 2023, 50, 842–887. [CrossRef]
- Sánchez-Peña, M.K.; Orozco-Restrepo, L.A.; Suárez-Brochero, Ó.F.; Barrios-Arroyave, F.A. Association Between Oral Health, Pneumonia and Mortality in Patients of Intensive Care. Rev. Med. Inst. Mex. Seguro Soc. 2020, 58, 468–476. [CrossRef]
Figure 1.
Circos diagram illustrating the prevalence of antibiotic classes and their associations with the respective study groups. LS: lincosamides-macrolides; MLS: macrolides-lincosamides-streptogramins; PLS: pleuromutilins-streptogramins-lincosamides.
Figure 1.
Circos diagram illustrating the prevalence of antibiotic classes and their associations with the respective study groups. LS: lincosamides-macrolides; MLS: macrolides-lincosamides-streptogramins; PLS: pleuromutilins-streptogramins-lincosamides.
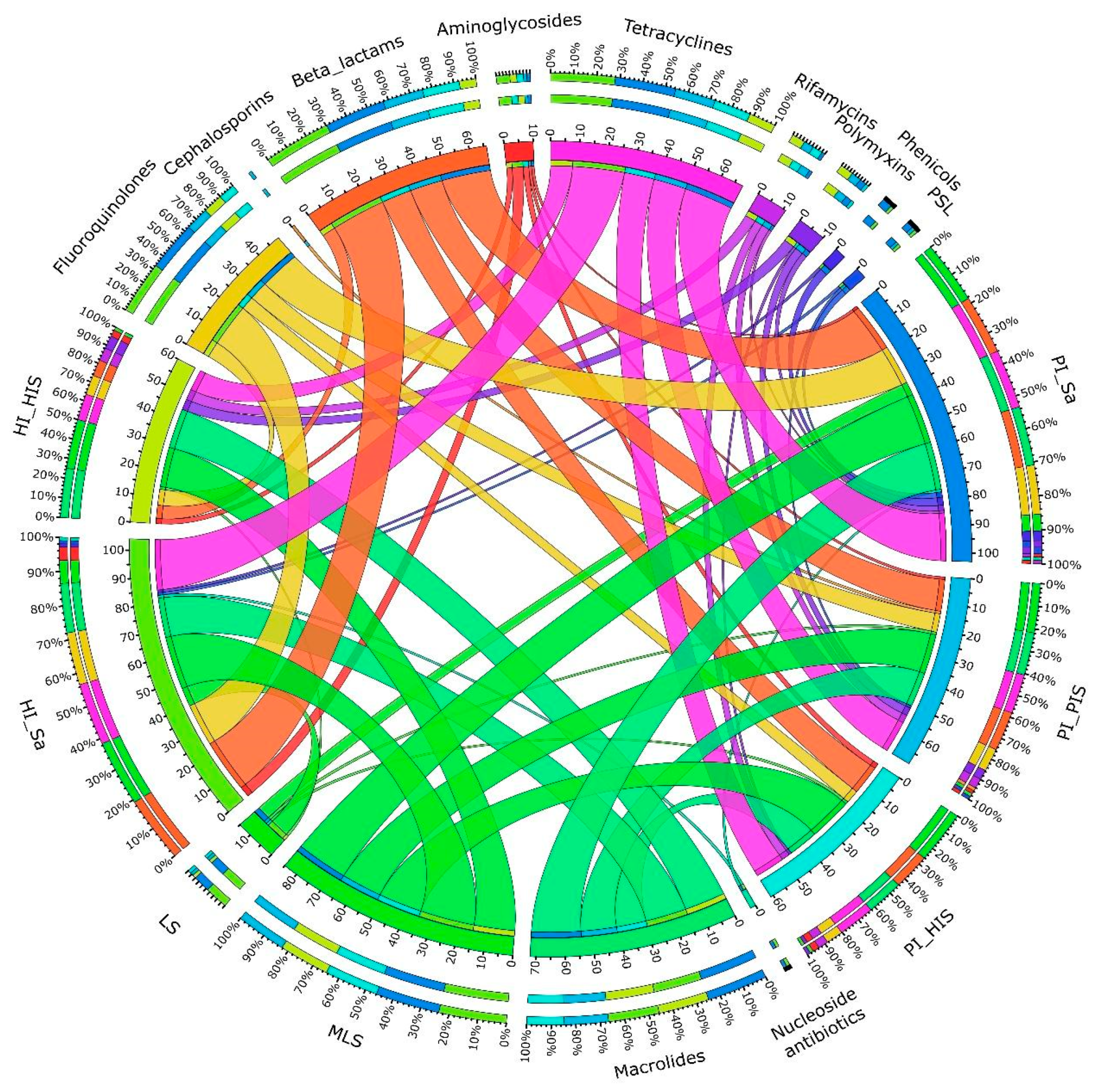
Figure 2.
Prevalence of ARGs across each of the five study groups: PI_Sa, PI_HIS, PI_PIS, HI_Sa and HI_HIS.
Figure 2.
Prevalence of ARGs across each of the five study groups: PI_Sa, PI_HIS, PI_PIS, HI_Sa and HI_HIS.
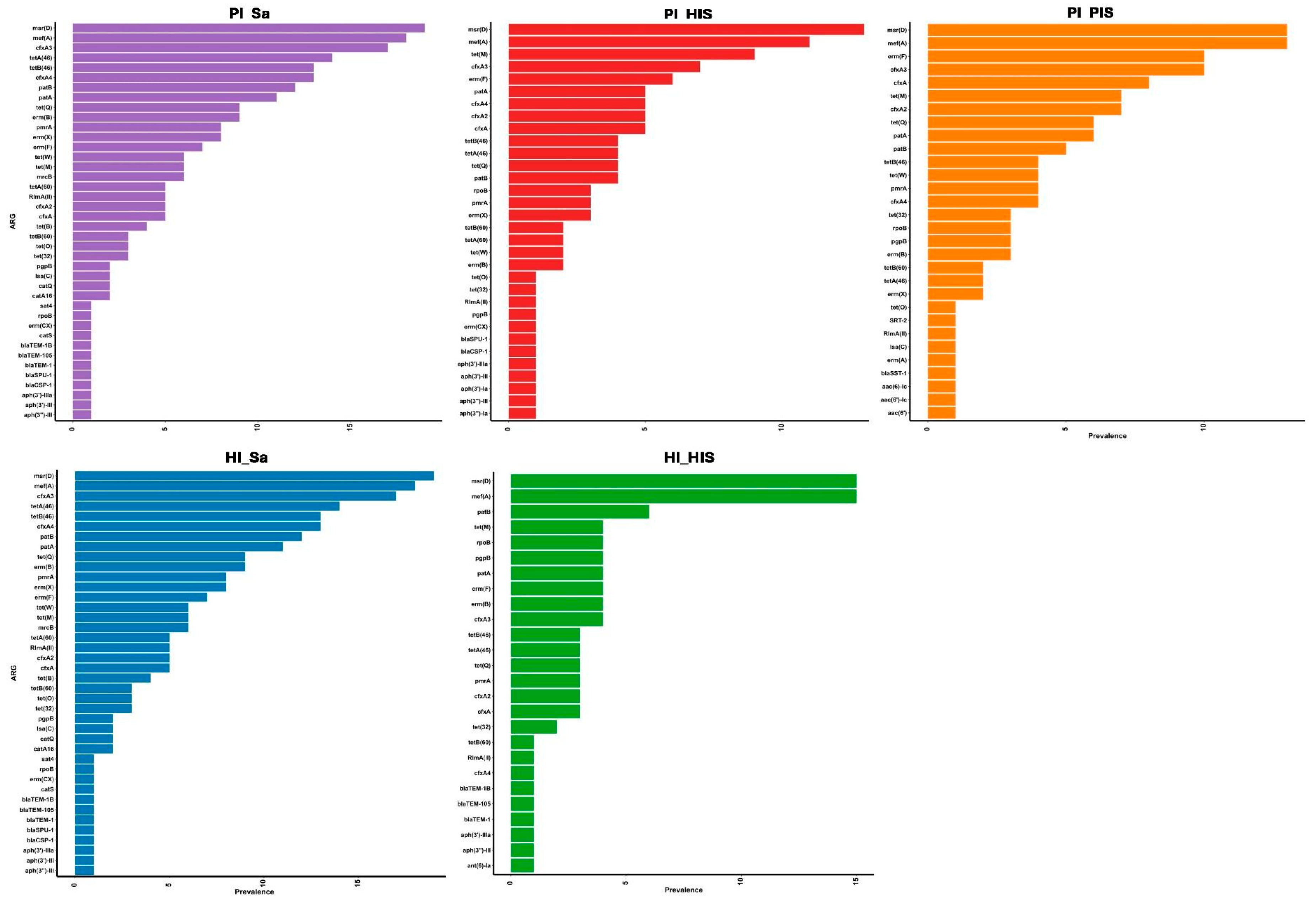
Figure 3.
Differentially prevalent ARGs between the study groups, with significance determined by Firth’s logistic regression *p ≤ 0.05, **p ≤ 0.01 and ***p ≤ 0.001.
Figure 3.
Differentially prevalent ARGs between the study groups, with significance determined by Firth’s logistic regression *p ≤ 0.05, **p ≤ 0.01 and ***p ≤ 0.001.
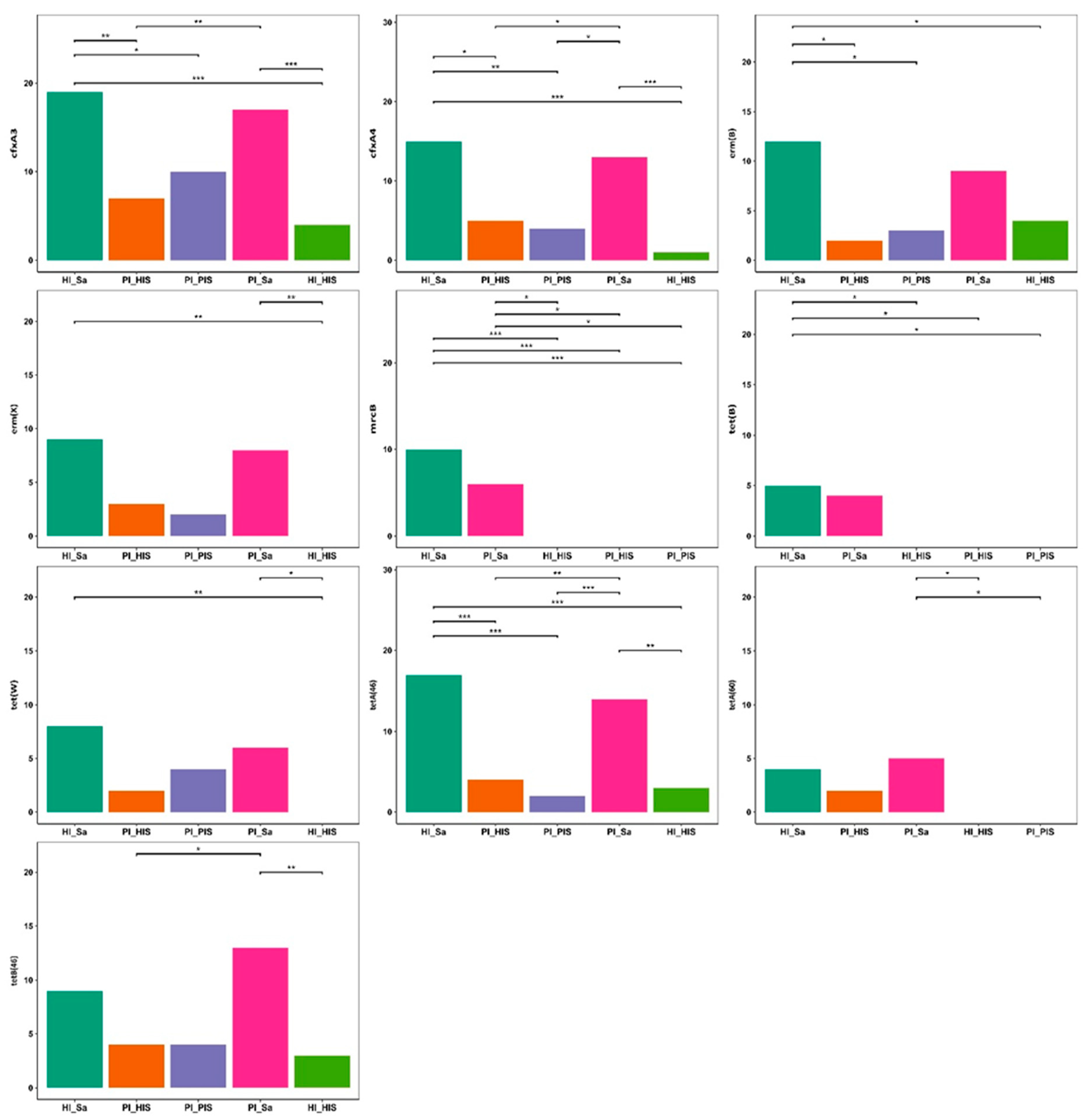
Figure 4.
Distribution of ARGs per bacterial species/plasmid. The size of the dots represents the prevalence of each gene within the respective bacterial species or plasmid. The taxonomic attribution of each identified ARG was retrieved directly from the ABRicate.
Figure 4.
Distribution of ARGs per bacterial species/plasmid. The size of the dots represents the prevalence of each gene within the respective bacterial species or plasmid. The taxonomic attribution of each identified ARG was retrieved directly from the ABRicate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



