Submitted:
28 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Background/Objectives: We generated a novel recombinant Vibrio cholerae ghost (rVCG)-based subunit vaccine incorporating the A1 subunit of cholera toxin (CTA1) and a multiepitope Chlamydia trachomatis (CT) antigen (MECA) derived from five chlamydial outer membrane proteins (rVCG-MECA). The ability of this vaccine to protect against a CT transcervical challenge was evaluated. Methods: Female C57BL/6J mice were immunized and boosted twice two weeks apart with rVCG-MECA or rVCG-gD2 (antigen control) via the intramuscular (IM) or intranasal (IN) route. Naïve mice or mice immunized IN with live CT served as negative and positive controls, respectively. Results: Vaccine delivery stimulated robust humoral and cell-mediated immune effectors, characterized by local mucosal and systemic antigen-specific IgG, IgG2c and IgA antibody and IFN-γ (Th1 cytokine) responses. The IgG2c and IgA antibody responses elicited in both serum and vaginal secretions persisted for 112 days postimmunization. Immunization with rVCG-MECA afforded levels of protective immunity comparable to that provided by IN immunization with live CT EBs without any side effects, irrespective of route of vaccine delivery. Conclusions: These results highlight the potential of a multiepitope vaccine as a promising resource for protecting against CT genital infection and the potential of CTA1 on the VCG platform as a mucosal and systemic adjuvant for developing CT vaccines.
Keywords:
chlamydia trachomatis
; rVCG-MECA
; vaccine delivery
; route
; protective immunity
1. Introduction
Chlamydia trachomatis (CT) is a gram-negative, obligate intracellular bacterium and the most frequently reported sexually transmitted infection (STI) worldwide. In the United States alone, the CDC reported over 1.6 million cases of CT in 2022, highlighting its persistent prevalence [1]. One of the major challenges in controlling the spread of CT is its asymptomatic nature, with more than 70% of infections going undiagnosed and untreated [2,3]. In women, untreated CT can lead to severe complications, including pelvic inflammatory disease (PID), ectopic pregnancy, and infertility, conditions that significantly affect reproductive health [4,5]. It is estimated that PID develops in 10-25% of infected women, with complications such as chronic pelvic pain, infertility, and ectopic pregnancies occurring in 10-30% of cases [6]. Additionally, C. trachomatis enhances the acquisition of other sexually transmitted infections and has been identified as an independent risk factor for cervical cancer [7]. Prior CT infection also increases the risk of fallopian tube carcinomas and epithelial ovarian cancer [8]. In men, common symptoms include urethritis, epididymitis, and orchitis, although long-term sequelae are less frequently observed [9].
Given the substantial public health burden and the severe long-term health impacts, including infertility and an increased risk of certain cancers, the development of prevention strategies is crucial [10]. While antibiotic treatment is effective, the asymptomatic nature of the infection allows it to spread widely, underscoring the need for additional public health interventions. Vaccine development is a vital strategy to combat the ongoing challenge of CT infection and its associated complications [11,12,13]. Despite its prevalence and significant burden on public health, there is currently no licensed Chlamydia vaccine. Chlamydia vaccine development has faced several challenges, including the bacterium’s unique intracellular lifecycle and its ability to evade the host immune response [14,15].
Historically, following the discovery that vaccines based on inactivated Chlamydia elementary bodies (EBs) induced pathology in recipients [16,17], vaccine development efforts have focused on identifying antigens that induce robust immune responses capable of providing long-lasting protection [15]. Development of subunit vaccines imposes the requirement for an effective delivery method capable of inducing appropriate immune responses in mucosal and systemic tissues. Recent advances in vaccine technology have introduced novel vaccine delivery strategies, including the Vibrio cholerae ghost (VCG) vaccine delivery platform, for targeting specific Chlamydia antigens to the immune system to elicit protective immune responses [18]. VCG are empty cell envelopes derived from V. cholerae by a genetic inactivation procedure that causes the release of cellular cytoplasmic contents, including cholera toxin while preserving the immunogenic surface components [19]. The VCG platform has been shown to enhance the protective immunity afforded by delivered vaccine antigens even in the absence of external adjuvants [20,21]. Another critical factor influencing vaccine efficacy is the route of administration, as different routes can lead to varying immune outcomes. For pathogens like CT, which primarily infect mucosal surfaces, both systemic and mucosal immunity are crucial for effective protection [22]. Systemic vaccination is traditionally associated with strong systemic immune responses, characterized by high levels of circulating antibodies and activated T cells, while mucosal vaccination is expected to induce mucosal immunity, which is particularly important for preventing infections at mucosal sites [21]. The enzymatically active CTA1-subunit of cholera toxin (CT) has been shown to be an effective mucosal and systemic adjuvant, stimulating strong cellular and humoral immune responses [23,24,25] and is non-toxic and safe following intranasal delivery [26].
In this study, we compared the efficacy afforded by a VCG-based multiepitope CT antigen derived from CT outer membrane proteins fused to CTA1 (rVCG-MECA) following intramuscular (IM) systemic and intranasal (IN) mucosal administration. These proteins, polymorphic membrane protein D (PmpD), PmpC, PmpG, PorB, and outer membrane complex B (OmcB) protein, play critical roles in chlamydial pathogenesis and are prime targets for vaccine development [13,18]. The study revealed that vaccine delivery induced robust protective immunity comparable to that provided by IN immunization with live CT EBs, indicating the ability of VCG-based epitope vaccines to induce protective immunity following both systemic and mucosal administration.
2. Materials and Methods
2.1. Chlamydia Stocks and Antigens
Stock preparations of C. trachomatis serovar D used in this study was previously titrated on Mccoy cell monolayers followed by purification of elementary bodies (EBs) over renografin gradients and stored at -80°C. CT antigen was prepared by UV-inactivation of EBs for 3 h and stored at 80°C until used.
2.2. Mice
The study utilized six-week-old female C57BL6/J mice obtained from The Jackson Laboratory (Bar Harbor, ME). The mice underwent a three-day period of acclimatization at the MSM Center for Laboratory Animal Resources facility prior to experimentation. The research adhered strictly to the guidelines outlined in the NIH Guide for the Care and Use of Laboratory Animals. All immunizations and challenge were performed under isoflurane anesthesia.
2.3. Construction of the Vaccine Vector, pCT-MECA and Expression of MECA by Immunoblotting Analysis
Immunogenic epitopes from five outer membrane proteins, PmpD, PmpC, PmpG, PorB, and OmcB previously shown to be immunogenic [27,28,29,30] were predicted. Selected epitopes were fused together using GPGPG linkers together with the A1 subunit of the cholera toxin (CTA1) at the N-terminal region and termed the multiepitope CT antigen (MECA). For construction of the vaccine vector, the 660 amino acids of the MECA antigen were reverse translated into a DNA coding sequence. The sequence was codon optimized to enhance expression in V. cholerae and synthesized to contain Xho I and Kpn I restriction sites at the N- and C-terminal ends (GenScript, Piscataway, NJ).
The synthesized gene (1,980 bp) was then inserted into the periplasmic targeting expression vector, pFLAG-CTS using Xho I and Kpn I restriction sites incorporated into the gene. The resultant plasmid was designated pCT-MECA and was used to transform by electroporation V. cholerae O139 strain V912 harboring the pDKLO1 plasmid, which expresses the lysis protein E. Cells grown in brain heart infusion broth at 37oC were induced for protein expression by addition of isopropyl β-D-thiogalactopyranoside (IPTG) (Roche Diagnostics, IN, USA) followed by gene E-mediated lysis as previously described [18]. Following expression, recombinant MECA is released and localized in the periplasmic space of V. cholerae where it is sequestered upon lysis. The lysed cells were harvested, washed, freeze-dried, and MECA protein expression was confirmed by western immunoblotting analysis using an anti-FLAG monoclonal antibody (Invitrogen, Waltham, MA) and imaged using the ChemiDoc MP Imagining System.
2.4. Production of rVCG-MECA Vaccine
V. cholerae strain V912 cells harboring pCT-MECA and the lysis plasmid pDKLO1 were cultured in Brain Heart Infusion (BHI) broth at 37°C to mid-log phase and rVCG-MECA was produced by genetic inactivation of the cells involving addition of 3-methyl benzoate to a growing culture as previously described [31]. The cells were harvested at the end of lysis, washed with a low ionic buffer, and lyophilized. The lyophilized rVCG-MECA were stored in cryovials at room temperature until used.
2.5. Immunization and Challenge of Immunized Mice
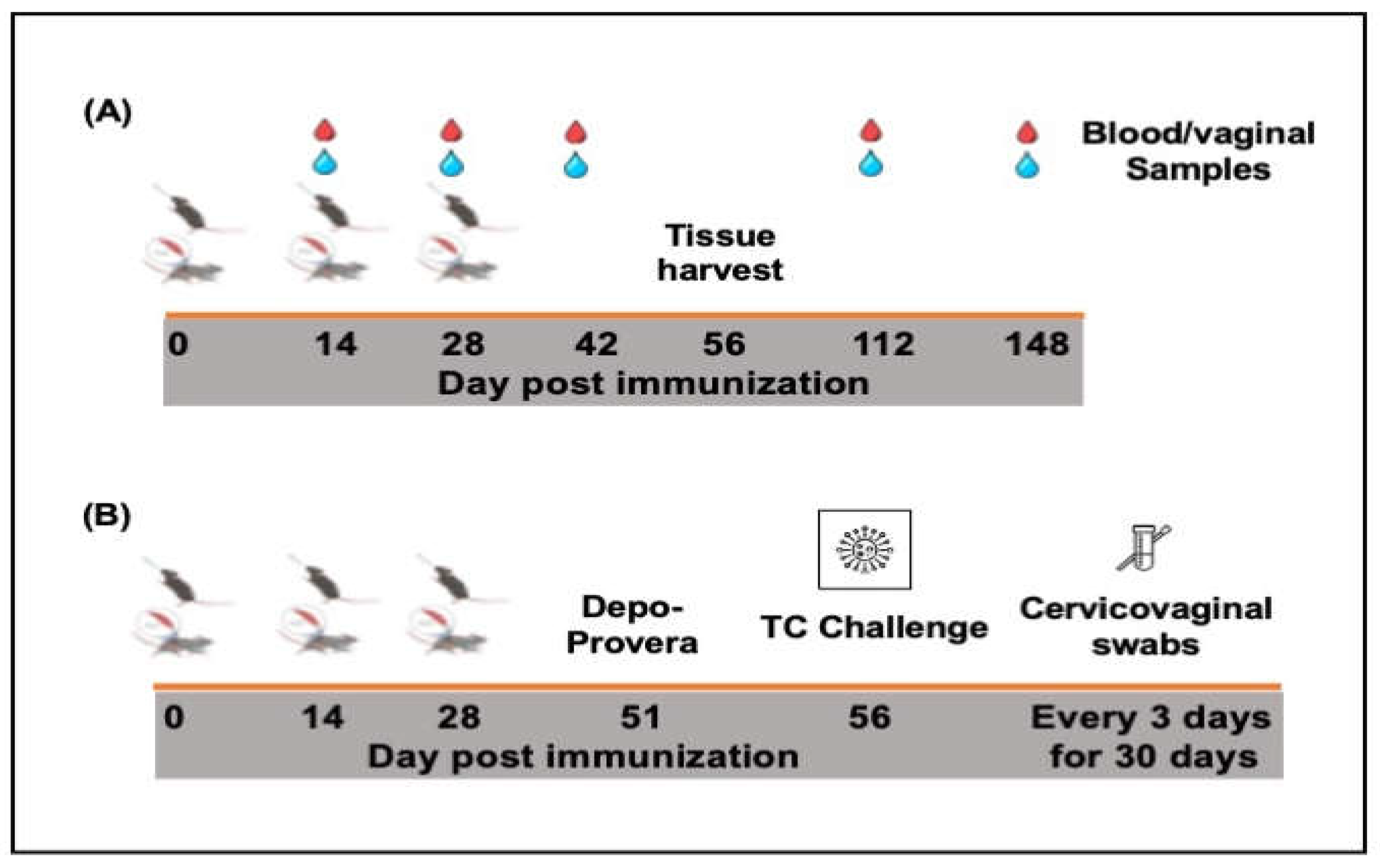
For immunogenicity studies (Figure 2A), groups of female C57BL/6 mice (16/group) were immunized either intramuscularly, IM (50 ml) or intranasally, IN (20 ml) and boosted two weeks apart with PBS containing 1.5 mg of lyophilized rVCG-MECA or rVCG-gD2 (VCG expressing the glycoprotein D2 from HSV-2, a chlamydial irrelevant antigen) (vaccine antigen control). Naïve mice or mice immunized IN with 1x106 IFU of live C. trachomatis (CT) EBs served as negative and positive controls, respectively. Blood and vaginal washes were collected on days 14, 28 and 42 as well as on days 112 (12 weeks) and 148 (16 weeks) postimmunization. Four weeks after the last immunization, mice designated for cell-mediated immune response studies (8/group) were sacrificed and immune T cells were purified from harvested spleens (SPL) and iliac lymph nodes (ILN) of immunized mice using the gentleMACS Dissociator.
In a separate experiment (Figure 2B), groups of mice were immunized as described above. Three weeks after the last immunization, mice designated for challenge studies (8/group) were treated subcutaneously with Depo-Provera (2.5 mg/mouse; UpJohn Co., Kalamazoo, MI) to synchronize the estrous cycle and enhance susceptibility to infection and challenged transcervically one week later (four weeks postimmunization) with live C. trachomatis serovar D (1 x 106 IFU/mouse). To monitor bacterial load and time to clearance of infection, cervicovaginal swabs were collected every three days for 30 days. Mice were observed daily to assess their health status and infection levels were determined by quantifying chlamydial inclusion-forming units (IFUs) from cervicovaginal swabs using indirect immunofluorescence [32]. The mean number of IFUs was then calculated at each time point.
2.6. Determination of Antigen-Specific Humoral Immune Responses
CT-specific antibodies (IgG, IgG2c and IgA) in serum and vaginal wash samples collected at designated time points after immunization and IgG1 from serum obtained 2 weeks after the last immunization were measured using a standard ELISA technique described previously [33]. Briefly, Maxisorb 96-well plates (Costar) were coated with CT antigen (UV inactivated CT Serovar D EBs) at a concentration of 10 mg protein/ml in sodium carbonate. To create a standard calibration curve, wells were coated in triplicate with varying concentrations of IgA or IgG2c standards (ranging from 0 to 1,000 ng/ml). The plates were then blocked with a solution of 1% bovine serum albumin and 5% goat serum in PBST. This was followed by the addition of serum samples (diluted 1:20) and vaginal wash samples (diluted 1:100), with each sample being incubated in the wells. After this, 50 ml of horseradish peroxidase-conjugated goat anti-mouse IgA or IgG2c (from Southern Biotechnology Associates, Birmingham, AL) was added and incubated for 1 hour. The reaction was developed using 2,2'-azino-bis(3-ethylbenzthiazoline-6-sulfonicacid) (ABTS), and the optical density of each well was measured at 490 nm using a Spectra Max 250 Microplate Auto reader (Molecular Devices Corp., Sunnyvale, CA). The results were presented alongside the standard curve, showing data sets that correlate absorbance values to mean concentrations (ng/ml) ± standard deviations, based on averages from three separate experiments.
2.7. Determination of Serum Antibody Avidity
The avidity of CT-specific serum IgG and IgG2c antibodies was evaluated using a modified ELISA protocol (Almanzar et al., 2013) incorporating antibody standards (for generation of a calibration curve) and ammonium thiocyanate (NH4SCN) as a chaotropic agent to disrupt low-affinity antigen-antibody interactions, allowing for the determination of binding strength. A 96-well plate was coated with 10 µg/ml of CT antigen in sodium carbonate buffer, incubated overnight at 4°C. After washing with PBS-T (0.05% Tween 20 in PBS), the wells were blocked for 1 hour with a solution containing 1% BSA and 5% goat serum in PBS-T. Serum samples, diluted 1:20, were added to the wells and incubated at 37°C for 1 h. The plates were then washed and 2 M NH4SCN was added to disrupt low-affinity interactions during a 20-minute incubation. Following additional washes, HRP-conjugated goat anti-mouse secondary antibody (diluted 1:5,000) was added followed by ABTS substrate and incubated for 1 h at 37°C. The reaction was stopped with 1% SDS, and the absorbance was measured at 450 nm with a reference at 650 nm. The relative avidity index was calculated by dividing the mean antibody concentration with NH4SCN by the concentration without NH4SCN. Avidity was classified as high (above 50%), intermediate (30-49%), or low (below 29%) based on the index values [34].
2.8. Assessment of Antigen-Specific Cellular Immune Responses
CT-specific Th1/Th2 cytokine production by SPL and ILN T cells was assessed as described previously [35] using the Bio-Plex cytokine assay kit in combination with the Bio-Plex Manager software (Bio-Rad, Hercules, CA). Briefly, single cell suspensions were obtained from the SPL and ILN of immunized and control mice 4 weeks after the last immunization using the gentleMACS Dissociator. Immune T cells were purified by positive selection using the Midi magnetic bead-activated cell sorting (MidiMACS) purification system in combination with Pan T cell-specific MACS microbeads (Miltenyi Biotech, Auburn, CA). Equal numbers (2×105 cells/well) of purified T cells and mitomycin C-treated splenocytes (APCs) from naïve mice were plated in triplicates in the wells of 96-well tissue culture plates and cultured for 5 days with CT antigen (10 mg/ml). T cells and APCs cultured in the absence of antigen were included as controls. The concentration of the cytokines in each sample was obtained by extrapolation from a standard calibration curve generated simultaneously. Data were calculated as mean values (± S.D.) for triplicate cultures for each experiment.
Statistical analysis
Statistical analyses were performed with the GraphPad Prism 10 package (GraphPad Software, Inc. La Jolla, CA, USA) on a Mac computer. Statistical differences between groups was evaluated by one-way ANOVA with Tukey’s post multiple comparison test. Statistical significance was determined at probability (p) values ≤ 0.05, 0.01, 0.001 or 0.0001.
3. Results
3.1. Construction of Plasmid pCT-MECA and VCG Expression of rMECA Protein
A total of 20 predicted T and B cell epitopes from PmpD (10), PmpC (5), PmpG (1), PorB (3) and OmcB (1) made up of 366 amino acids were selected on the basis of high scoring physicochemical properties that included surface accessibility, antigenicity, and hydrophilicity. The 194 amino acid CTA1 was fused to the N-terminal end of MECA via an EAAAK linker. The pCT-MECA vaccine vector was then constructed by inserting the 1,980 bp synthesized coding sequence (Figure 1A), which was codon optimized to enhance expression in V. cholerae into the periplasmic targeting vector, pFLAG-CTS in frame with the Flag Tag sequence under the transcriptional control of the Ptac promoter (Figure 1B). This vector is specifically designed to target expressed proteins in the periplasmic space of gram-negative bacteria. Sequencing results by GenScript (Piscataway, NJ) confirmed that the inserted gene was in frame with the Flag Tag fusion sequence. The pCT-MECA plasmid was transformed into V. cholerae V912 harboring the pDKLO1 lysis plasmid. After gene E-mediated lysis of the cells, the expression of recombinant MECA (~78.88 kDa) was confirmed by Western immunoblotting analysis using anti-Flag monoclonal antibodies (Figure 1C).
3.2. IM and IN Immunization with rVCG-MECA Induced Robust Antigen-Specific Antibodies in Serum and Vaginal Secretions
Mice were immunized three times, two weeks apart via the IM or IN route and challenged transcervically as outlined in the experimental protocol (Figure 2).
Figure 2.
Schematic diagram of the experimental protocol outlining the immunization, sample collection (A) and challenge (B) schedules.
Figure 2.
Schematic diagram of the experimental protocol outlining the immunization, sample collection (A) and challenge (B) schedules.
CT-specific antibody responses in serum and vaginal wash samples were analyzed two weeks after the first, second and third immunizations using a standard antibody ELISA assay. Compared to the rVCG-gD2 and naïve controls, immunization with rVCG-MECA induced significantly higher (p < 0.0001) levels of IgG, IgG2c and IgA antibodies in both mucosal and systemic compartments, which were robust even after a single vaccine dose, irrespective of route of immunization (Figure 3). Antibody responses increased with the number of immunization doses with substantial amounts elicited after the third dose. While antibody responses elicited in serum after IM and IN immunization were comparable (Figure 3A, C and E), IM immunization induced significantly higher (p < 0.0001) levels of IgG and the IgG2c isotype antibodies in virginal secretions compared to the IN route (Figure 3B and D). In contrast, IgA antibodies were significantly higher (p < 0.001) in vaginal secretions following IN immunization (Figure 3F). Antibody responses elicited by immunization with rVCG-MECA were either slightly lower or comparable to levels elicited by IN immunization with live CT EBs (positive control).
The antigen-specific Th1-associated IgG2c and Th2-associated IgG1 antibodies produced in serum two weeks after the third immunization were also compared. The results showed that IM and IN immunization with rVCG-MECA induced significantly higher (p < 0.0001) serum IgG2c compared to IgG1 antibodies (Figure 4A). Further evaluation showed that the IgG2c/IgG1 ratios > 1, irrespective of the immunization route indicating immunization with rVCG-MECA elicited a Th1-type humoral immune response two weeks post-immunization (Figure 4B).
3.3. Vaccine-Induced CT-Specific IgG2c and IgA Antibodies Persisted in Serum and Vaginal Secretions
To evaluate the ability of the vaccine to elicit long-term antibody responses, samples collected 12 weeks (day 84) and 16 weeks (day 112) post-immunization were also analyzed. The results showed that by week 12 post-immunization, high levels of antigen-specific IgG2c and IgA antibodies were detected in the serum and vaginal secretions of rVCG-MECA-immunized mice that persisted for up to 16 weeks, irrespective of the route of vaccine delivery (Figure 5A–D). While IgG2c levels in serum were significantly higher (p < 0.0001) after IM delivery, comparable levels were elicited in vaginal secretions at both time points (Figure 5A,B). Also, while serum and vaginal IgA levels in both IM- and IN-immunized mice were comparable 12 weeks post-immunization, levels were significantly lower (p > 0.0001) in IN-immunized mice 16 weeks post-immunization (the longest time point evaluated in this study) (Figure 5C,D). The results indicate that antigen-specific IgG2c and IgA antibody responses elicited in both mucosal and systemic tissues by IM and IN immunization with rVCG-MECA are long lasting.
3.4. Avidity of Antigen-Specific Serum IgG and IgG2c Antibodies
The functionality of CT-specific IgG and IgG2c antibodies elicited in serum following immunization with rVCG-MECA was evaluated by an ammonium thiocyanate (NH4SCN)-based antibody avidity ELISA assay. The data was expressed as relative avidity index, defined as the ratio between the mean antibody concentration in the presence and absence of NH4SCN. The data shows that the relative avidity index of serum IgG antibodies induced by IM and IN immunization was moderate at 2 weeks post-immunization (Figure 6A). While the avidity of serum IgG antibodies induced by IM immunization remained moderate 4 weeks post-immunization, antibodies with high avidity (> 50%) were elicited in IN-immunized mice at this time point (Figure 6A).
On the other hand, the relative avidity index of the Th1-associated serum IgG2c antibodies induced 2 weeks post-immunization was significantly higher (p < 0.001) after IN mucosal (65%) compared to IM systemic (46%) immunization (Figure 6B). However, by week 4 post-immunization, the serum IgG2c antibody avidity induced by IM (58%) and IN (62%) immunization were comparable (Figure 6B). In contrast, the relative avidity index of IgG and IgG2c antibodies induced by immunization with PBS (negative control) remained at baseline levels (20%) at both time points (Figure 6A,B). These findings indicate that the Th1-associated IgG2c antibody isotype elicited by rVCG-MECA immunization show higher functional activity compared to total IgG antibodies, irrespective of route of vaccine delivery.
3.5. High Levels of Antigen-Specific IFN-γ Were Induced in Mucosal and Systemic Tissues Following Immunization with rVCG-MECA
Four weeks postimmunization, the amount of antigen-specific Th1/Th2 cytokines secreted by immune T cells purified from the spleens and iliac lymph nodes (ILN) draining the genital tract of immunized mice was analyzed after restimulation with chlamydial antigen in the presence of antigen-presenting cells (APCs). The results show that significantly higher (p < 0.0001) levels of the Th1-type cytokine, IFN-γ were secreted by splenic and ILN immune T cells following both IM and IN immunization compared to T cells from rVCG-gD2 and naïve controls (Fig 7A). While the amount of IFN-γ produced by splenic immune T cells following IM immunization was significantly higher (p < 0.01) compared to the IN route, comparable levels were secreted by both splenic and ILN immune T cells following IN immunization with rVCG-MECA and live CT EBs (Fig 7A). As expected, immune T cells from the spleens and ILN of naïve mice or mice immunized with rVCG-gD2 did not secrete significant levels of IFN-γ. Also, significantly higher (p < 0.0001) amounts of IL-4 were secreted by splenic (p < 0.0001) and ILN (p < 0.01) immune T cells following IM and IN immunization with rVCG-MECA and live CT EBs compared to amounts stimulated by T cells from rVCG-gD2 controls (Fig 7B). While comparable IL-4 levels were produced by ILN immune T cells following IM and IN immunization with rVCG-MECA, IN immunization with rVCG-MECA produced significantly higher (p < 0.05) levels than those produced by live CT EBs. However, compared to IFN-γ, considerably lower levels of the Th2 cytokine IL-4 were produced by immune T cells obtained from both spleen and ILN, irrespective of route of vaccine delivery (Figure 7A,B). These results demonstrate that both IM and IN immunizations with rVCG-MECA induce robust Th1-type cytokine responses similar to that induced by immunization with live CT EBs (positive control).
3.6. Immune Effectors Stimulated by Immunization with rVCG-MECA Protected Mice Against Transcervical Challenge with Live CT EBs
The protective efficacy of the rVCG-MECA vaccine was evaluated after transcervical CT challenge of immunized mice and controls four weeks postimmunization. Genital chlamydial loads were evaluated by cervicovaginal swabbing of each mouse every three days, and chlamydial inclusions were enumerated following culture in HeLa cell monolayers. As early as three days postchallenge, there was a significant (p < 0.0001) reduction in bacterial load in all immunized mice compared to naïve control mice (Figure 8). A further reduction in chlamydial load was observed in rVCG-MECA vaccine-immunized mice by day 9 postchallenge. Infection loads at this time point were comparable to those in live CT-immunized mice (positive control), irrespective of route of vaccine delivery, but were significantly (p < 0.0001) lower than those in naïve mice. All rVCG-MECA vaccine- and CT-immunized mice had started to clear the infection on day 15 and by day 21, all vaccine-immunized groups had completely cleared the infection. Naïve control mice were still shedding large numbers of CT EBs at this time point.
4. Discussion
V. cholerae ghosts (VCG) have emerged as an efficient vaccine delivery platform due to their inherent adjuvant properties as they retain all the major immune stimulating constituents of the bacterial envelope, including lipopolysaccharide, peptidoglycan, toxin co-regulated pili and certain outer membrane proteins [19], which trigger innate immune responses [36]. The rVCG platform has previously been shown to enhance the protective immunity of vaccine antigens by activating dendritic cells and enhancing antigen presentation [37]. We had also reported that a rVCG-based multisubunit chlamydial vaccine induced robust antigen-specific humoral and cellular immune responses and afforded comparable protective immunity against genital CT infection following IM systemic and rectal mucosal immunization with a vaccine based on the polymorphic outer membrane proteins [21]. This study demonstrated that the route of vaccine administration plays a critical role in shaping elicited immune effectors and directing these effectors from one inductive site to different effector sites.
Although VCG-based vaccines induce robust protective immunity in the absence of external adjuvants [20,38], we previously demonstrated that co-delivery of a VCG-based subunit Chlamydia vaccine with the Fms-like tyrosine kinase 3-ligand (FL; a dendritic cell targeting adjuvant) enhanced the protective immunity of the vaccine [21,36], indicating that the use of appropriate adjuvants could further improve the efficacy of VCG-based vaccines. Although intranasal immunization with cholera toxin (Ctx) can target immunity to the genital tract, its potential to cause neurological side effects in experimental mice has precluded its use as a mucosal adjuvant [39]. We previously showed that codelivery of an rVCG vaccine co-expressing the CT major outer membrane protein (MOMP) and CTA2B, the nontoxic derivative of Ctx significantly enhanced clearance of a C. muridarum vaginal infection following intravaginal immunization [40]. Previous studies have shown that CTA1-DD, a nontoxic derivative of Ctx is an effective mucosal and systemic adjuvant, stimulating robust CD4+ T-cell and antibody responses [24,41]. Also, IN delivery of C. muridarum MOMP with CTA1-DD induced neutralizing systemic and mucosal antibodies, and reduced genital chlamydial shedding following intravaginal challenge [42]. The immunoenhancing capacity of CTA1 is said to be due its ability to bind to and directly cause the maturation of immature follicular dendritic cells (FDCs) leading to germinal center (GC) B-cell and follicular helper T cell (Tfh) development [43]. In this study, we generated a novel subunit vaccine by molecularly fusing CTA1 with a multiepitope CT antigen expressed as a single polycistronic unit (MECA) and expressed in VCG (rVCG-MECA). We evaluated the ability of rVCG-MECA to elicit immune effectors capable of protecting against a transcervical CT infection following the IM and IN routes of vaccine administration. Vaccine evaluation demonstrated the ability of both immunization routes to stimulate robust local mucosal and systemic antigen-specific IgG, IgG2c and IgA antibody responses in serum and vaginal secretions. This is likely due to the combined effect of VCG and CTA1 adjuvant that are known to enhance humoral immune responses in both mucosal and systemic tissues following mucosal and systemic immunization [21,43]. The finding that the IM route elicited significantly higher levels of IgG and IgG2c antibodies in vaginal secretions compared to the IN route indicates that these antibodies likely originate from the systemic circulation. The significantly higher levels of the Th1-associated IgG2c compared to the Th2-associated IgG1 isotype in both mucosal and systemic tissues indicates that immunization with rVCG-MECA skews elicited IgG antibodies toward a Th1 phenotype, which enhances Th1 cellular immunity essential for the clearance of intracellular pathogens such as C. trachomatis [44,45].
Notably, the IN route more effectively stimulated mucosal immunity following immunization with rVCG-MECA, as demonstrated by elevated IgA levels in vaginal secretions. The elevated IgA concentrations in vaginal washes of IN-immunized mice underscore the ability of this route to elicit mucosal immunity, which is critical for defending against pathogens, such as CT that establish infections at mucosal surfaces (Nogueira et al., 2017). This result is consistent with other studies where IN immunization has been shown to stimulate effective mucosal immune responses to subunit antigens [46,47,48], although concerns about potential side effects, such as impacts on the central nervous system have been raised when co-delivered with certain adjuvants [49]. Mucosal routes such as oral and colonic immunization have also demonstrated effectiveness in animal models but face challenges in delivering subunit vaccines due to the harsh gastrointestinal environment [50,51,52]. Secretory IgA serves as a frontline barrier at mucosal surfaces, preventing pathogen entry, a critical aspect for defense against sexually transmitted infections [49], underscoring the distinct impact of immunization route on humoral mucosal immunity to rVCG-MECA vaccine. Although the role of IgA in protection against CT is still unfolding, previous reports from both mice and human studies indicate a protective role for IgA in the genital mucosa [53,54,55]. A recent report showed that local secretory IgA elicited by oral immunization with either inactivated or live C. muridarum neutralized chlamydial infectivity in the female genital tract enhancing chlamydial clearance and reducing systemic spread [56]. The robust antibody responses persisted for up to 16 weeks (112 days) postimmunization, the longest time point assessed, establishing the longevity of mucosal and systemic IgG2c and IgA antibody responses following IM and IN immunization. The ability of this vaccine to provide long-term protection following a genital challenge infection will be evaluated in a future study. The finding that IN immunized mice produced antibodies with higher binding strength (RAI > 50%) suggests that serum antibodies, especially the IgG2c isotype, induced by mucosal immunization have higher functional capacity, which is crucial for effective pathogen clearance.
The results also showed that high levels of the Th1-associated IFN-g and low levels of the Th2-associated IL-4 cytokines were secreted by splenic and ILN cells following IM and IN immunization indicating that both routes of vaccine delivery effectively stimulated a Th1-type cellular immune response in both mucosal and systemic tissues. The ability of the IM route to induce significantly higher levels of the Th1 cytokine, IFN-γ compared to the IN route underscores the effectiveness of systemic immunization in stimulating cell-mediated immunity, which is critical for clearing intracellular pathogens [45]. Reports from human clinical and animal model studies established the primary role of IFN-γ-secreting CD4+ Th1 cells in protection against CT genital infection [44,57,58,59]. Although CD8+ T cells have been reported to play a role in protection against the respiratory and genital mucosae [60], its protective role in the female reproductive tract is yet to be completely understood. IFN-γ is crucial for macrophage activation, which inhibits the growth of CT enhancing pathogen clearance [61,62]. In addition, some studies have indicated the likely presence of IFN-γ-independent innate mechanisms with the ability to control genital CT infection [63,64]. Although some reports have suggested that a balanced Th1/Th2 response may be beneficial for long-term chlamydial immunity [65], our study demonstrated a predominant Th1 immune response, which most studies have indicated is essential for protection against Chlamydia infection.
A number of immunization routes have been used to evaluate the efficacy of subunit chlamydial vaccines. IN immunization with different chlamydial antigens have provided varying degrees of bacterial clearance in the mouse genital tract [66,67]. A previous study demonstrated that mice immunized by a combination of mucosal and systemic routes, involving two mucosal (vaginal + colonic or intranasal + sublingual) and two systemic (IM + SC) routes, afforded the best protection compared those immunized only via the mucosal route [50]. A recent report indicating that a combination of colonic (mucosal) prime followed by IM + SC (systemic) boosts compared to mucosal or systemic only routes provided the best protection confirms this report [46]. Analysis of the protective efficacy afforded by immunization with rVCG-MECA showed that both IM and IN routes provided substantial protection against CT genital infection, as indicated by significant reduction in bacterial load and a shortening of the time to clearance of infection compared to controls. This finding aligns with previous reports demonstrating the ability of VCG-based chlamydial vaccines to induce varying levels of protection following either mucosal or systemic routes of immunization [20,21,38]. These outcomes provide a broader understanding of how the route of administration and the vaccine delivery vehicle impact the protective immunity of CT vaccines. The rVCG-MECA vaccine incorporates immunogenic regions from several key CT antigens, including PmpD, PmpC, PmpG, PorB and OmcB, which will likely offer broader protection compared to single-antigen vaccines like MOMP-based vaccines. MOMP-based vaccines have been shown to offer serovar-specific protection [31] but are often less effective across multiple serovars [68]. In contrast, our multiepitope approach has the potential to overcome this limitation by targeting multiple regions of the pathogen, offering broader immunity. Other multiepitope vaccines, such as those targeting C. psittaci, have also demonstrated the ability to induce robust immune responses and cross-serovar protection [69]. Additionally, a subunit vaccine targeting multiple stages of Mycobacterium tuberculosis showed high protective efficacy in mice, reinforcing the potential of multi-antigen strategies to provide broad protection [69,70]. The inclusion of multiple epitopes in our vaccine construct offers a focused yet comprehensive immune response, avoiding some of the issues associated with whole-antigen vaccines, which may elicit responses to non-protective or pathogenic antigens [48,71].
One of the limitations of this study is the non-inclusion of rVCG-MECA construct that excludes CTA1 to enable a direct evaluation of the effect of CTA1 on vaccine-induced immune responses and protection. Also, the purified rMECA vaccine antigen (without VCG) was not included in this study that would have enabled evaluation of the effect of the VCG component on vaccine immunity. However, in a preliminary study, we showed that IN immunization with different concentrations of purified rMECA induced substantial serum and vaginal IgG2c and secretory IgA antibodies after two vaccine doses [72]. Studies evaluating the ability of purified rMECA to protect against genital CT infection are ongoing.
5. Conclusions
Our findings underscore the importance of considering both the route of immunization and the design of the vaccine when developing strategies to prevent C. trachomatis infections. The IM route was highly effective in generating strong systemic Th1 responses, which are crucial for pathogen clearance. The IN route, on the other hand was highly effective in producing high-avidity IgG2c antibodies and eliciting mucosal immunity through production of high levels of secretory IgA antibodies, which are critical in defending against CT infection in the vaginal mucosa. Additionally, the ability of rVCG-MECA vaccine to induce robust antibody responses that persisted for up to 112 days postimmunization, indicates that it is likely to provide long-lasting protection against CT infection, making it a promising candidate for future vaccine development. Future studies will utilize this immunization strategy to evaluate the ability of this multiepitope CT vaccine to protect against heterologous challenge with other chlamydial serovars.
Author Contributions
Conceptualization, F.E. and F.M.; Methodology, F.M., SL; Software, F.E.; Validation, T.T. and S.L.; Formal Analysis, F.M.; Investigation, F.M.; Resources, F.E., Y.O.; Data Curation, F.M.; Writing – Original Draft Preparation, F.M.; Writing – Review & Editing, F.E.; Visualization, F.E.; Supervision, F.E., Y.O.; Project Administration, F.E.; Funding Acquisition, F.E.
Funding
This study was supported in part by a grant (#R01AI26897) from the National Institutes of Health (NIH) and Morehouse School of Medicine institutional support through the Graduate Education in Biomedical Sciences (GEBS).
Institutional Review Board Statement
This project was approved by the Institutional Animal Care and Use Committee (IACUC) of Morehouse School of Medicine (MSM) (IACUC Approval # 19-09).
Data Availability Statement
The data presented in this study are included in the article; further inquiries can be directed to the corresponding author.
Acknowledgements
We gratefully acknowledge members of the animal care staff and veterinarians at the MSM Center for Laboratory Animal Resources (CLAR), for their care, advice and guidance throughout the course of the study. We are also grateful to the staff at the Georgia Institute of Technology Cellular Analysis and Cytometry Core for technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- CDC, C.f.D.C.a.P. National Overview of STIs, 2022; 2022.
- Farley, T.A.; Cohen, D.A.; Elkins, W. Asymptomatic sexually transmitted diseases: the case for screening. Prev Med 2003, 36, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Stamm, W.E. Chlamydia trachomatis infections: progress and problems. J Infect Dis 1999, 179 Suppl 2, S380–383. [Google Scholar] [CrossRef]
- Yonke, N.; Aragon, M.; Phillips, J.K. Chlamydial and Gonococcal Infections: Screening, Diagnosis, and Treatment. Am Fam Physician 2022, 105, 388–396. [Google Scholar]
- Curry, A.; Williams, T.; Penny, M.L. Pelvic Inflammatory Disease: Diagnosis, Management, and Prevention. Am Fam Physician 2019, 100, 357–364. [Google Scholar] [PubMed]
- Haggerty, C.L.; Gottlieb, S.L.; Taylor, B.D.; Low, N.; Xu, F.; Ness, R.B. Risk of sequelae after Chlamydia trachomatis genital infection in women. J Infect Dis 2010, 201 Suppl 2, S134–155. [Google Scholar] [CrossRef]
- Chan, P.A.; Robinette, A.; Montgomery, M.; Almonte, A.; Cu-Uvin, S.; Lonks, J.R.; Chapin, K.C.; Kojic, E.M.; Hardy, E.J. Extragenital Infections Caused by Chlamydia trachomatis and Neisseria gonorrhoeae: A Review of the Literature. Infect Dis Obstet Gynecol 2016, 2016, 5758387. [Google Scholar] [CrossRef] [PubMed]
- Perrott, S.L.; Kar, S.P. Appraisal of the causal effect of Chlamydia trachomatis infection on epithelial ovarian cancer risk: a two-sample Mendelian randomisation study. medRxiv, 2024. [Google Scholar] [CrossRef]
- Bryan, E.R.; Redgrove, K.A.; Mooney, A.R.; Mihalas, B.P.; Sutherland, J.M.; Carey, A.J.; Armitage, C.W.; Trim, L.K.; Kollipara, A.; Mulvey, P.B.M.; et al. Chronic testicular Chlamydia muridarum infection impairs mouse fertility and offspring developmentdagger. Biol Reprod 2020, 102, 888–901. [Google Scholar] [CrossRef]
- Borges, A.H.; Follmann, F.; Dietrich, J. Chlamydia trachomatis vaccine development - a view on the current challenges and how to move forward. Expert Rev Vaccines 2022, 21, 1555–1567. [Google Scholar] [CrossRef]
- Murthy, A.K.; Li, W.; Ramsey, K.H. Immunopathogenesis of Chlamydial Infections. Curr Top Microbiol Immunol 2018, 412, 183–215. [Google Scholar] [CrossRef]
- Brunham, R.C. The genome, microbiome and evolutionary medicine. CMAJ 2018, 190, E162–E166. [Google Scholar] [CrossRef]
- Brunham, R.C.; Rey-Ladino, J. Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nat Rev Immunol 2005, 5, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Poston, T.B.; Gottlieb, S.L.; Darville, T. Status of vaccine research and development of vaccines for Chlamydia trachomatis infection. Vaccine 2019, 37, 7289–7294. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; McKay, P.F. Chlamydia trachomatis: Cell biology, immunology and vaccination. Vaccine 2021, 39, 2965–2975. [Google Scholar] [CrossRef]
- Grayston, J.T.; Wang, S.-P.; Yeh, L.J.; Kuo C, C. Importance of reinfection in the pathogenesis of trachoma. Rev.Infect.Dis. 1985, 7, 717–725. [Google Scholar] [CrossRef]
- Schachter, J.; Dawson, C.R. In Human chlamydial infections, PSG Publishing Company: Littleton, MA, 1978.
- Eko, F.O.; Lubitz, W.; McMillan, L.; Ramey, K.; Moore, T.T.; Ananaba, G.A.; Lyn, D.; Black, C.M.; Igietseme, J.U. Recombinant Vibrio cholerae ghosts as a delivery vehicle for vaccinating against Chlamydia trachomatis. Vaccine 2003, 21, 1694–1703. [Google Scholar] [CrossRef]
- Eko, F.O.; Mayr, U.B.; Attridge, S.R.; Lubitz, W. Characterization and immunogenicity of Vibrio cholerae ghosts expressing toxin-coregulated pili. J Biotechnol 2000, 83, 115–123. [Google Scholar] [CrossRef]
- Eko, F.; Ekong, E.; Okenu, D.; He, Q.; Ananaba, G.; Black, C.; Igietseme, J. Induction of immune memory by a multisubunit chlamydial. Vaccine 2011, 2011, 2011–2011. [Google Scholar] [CrossRef]
- Pais, R.; Omosun, Y.; Igietseme, J.U.; Fujihashi, K.; Eko, F.O. Route of Vaccine Administration Influences the Impact of Fms-Like Tyrosine Kinase 3 Ligand (Flt3L) on Chlamydial-Specific Protective Immune Responses. Frontiers in immunology 2019, 10, 1577. [Google Scholar] [CrossRef]
- Redgrove, K.A.; McLaughlin, E.A. The Role of the Immune Response in Chlamydia trachomatis Infection of the Male Genital Tract: A Double-Edged Sword. Front Immunol 2014, 5, 534. [Google Scholar] [CrossRef]
- Helgeby, A.; Robson, N.C.; Donachie, A.M.; Beackock-Sharp, H.; Lövgren, K.; Schön, K.; Mowat, A.; Lycke, N.Y. The combined CTA1-DD/ISCOM adjuvant vector promotes priming of mucosal and systemic immunity to incorporated antigens by specific targeting of B cells. J Immunol 2006, 176, 3697–3706. [Google Scholar] [CrossRef]
- Agren, L.; Sverremark, E.; Ekman, L.; Schön, K.; Löwenadler, B.; Fernandez, C.; Lycke, N. The ADP-ribosylating CTA1-DD adjuvant enhances T cell-dependent and independent responses by direct action on B cells involving anti-apoptotic Bcl-2- and germinal center-promoting effects. J Immunol 2000, 164, 6276–6286. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Choi, J.A.; Park, H.; Yang, E.; Noh, S.; Kim, J.S.; Kim, M.J.; Song, M.; Park, J.H. Pharmaceutical and Immunological Evaluation of Cholera Toxin A1 Subunit as an Adjuvant of Hepatitis B Vaccine Microneedles. Pharm Res 2023, 40, 3059–3071. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, A.M.; Schön, K.M.; Lycke, N.Y. The cholera toxin-derived CTA1-DD vaccine adjuvant administered intranasally does not cause inflammation or accumulate in the nervous tissues. J Immunol 2004, 173, 3310–3319. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Stephens, R.S. Characterization and functional analysis of PorB, a Chlamydia porin and neutralizing target. Mol Microbiol 2000, 38, 772–780. [Google Scholar] [CrossRef]
- Brunham, R.C.; Peeling, R.W. Chlamydia trachomatis antigens: role in immunity and pathogenesis. Infect Agents Dis 1994, 3, 218–233. [Google Scholar]
- Vasilevsky, S.; Stojanov, M.; Greub, G.; Baud, D. Chlamydial polymorphic membrane proteins: regulation, function and potential vaccine candidates. Virulence 2016, 7, 11–22. [Google Scholar] [CrossRef]
- Yu, H.; Karunakaran, K.P.; Jiang, X.; Brunham, R.C. Evaluation of a multisubunit recombinant polymorphic membrane protein and major outer membrane protein T cell vaccine against Chlamydia muridarum genital infection in three strains of mice. Vaccine 2014, 32, 4672–4680. [Google Scholar] [CrossRef]
- Eko, F.; Lubitz, W.; McMillan, L.; Ramey, K.; Moore, T.; Ananaba, G.A.; Lyn, D.; Black, C.M.; Igietseme, J.U. Recombinant Vibrio cholerae ghosts as a delivery vehicle for vaccinating against Chlamydia trachomatis. Vaccine 2003, 21, 1694–1703. [Google Scholar] [CrossRef]
- Su, H.; Parnell, M.; Caldwell, H.D. Protective efficacy of a parenterally administered MOMP-derived synthetic oligopeptide vaccine in a murine model of Chlamydia trachomatis genital tract infection: serum neutralizing IgG antibodies do not protect against chlamydial genital tract infection. Vaccine 1995, 13, 1023–1032. [Google Scholar] [CrossRef]
- Macmillan, L.; Ifere, G.O.; He, Q.; Igietseme, J.U.; Kellar, K.L.; Okenu, D.M.; Eko, F.O. A recombinant multivalent combination vaccine protects against Chlamydia and genital herpes. FEMS Immunol Med Microbiol 2007, 49, 46–55. [Google Scholar] [CrossRef]
- Trzewikoswki de Lima, G.; Rodrigues, T.S.; Portilho, A.I.; Correa, V.A.; Gaspar, E.B.; De Gaspari, E. Immune responses of meningococcal B outer membrane vesicles in middle-aged mice. Pathog Dis 2020, 78. [Google Scholar] [CrossRef] [PubMed]
- Eko, F.O.; Ekong, E.; He, Q.; Black, C.M.; Igietseme, J.U. Induction of immune memory by a multisubunit chlamydial vaccine. Vaccine 2011, 29, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Pais, R.; Ohandjo, A.; He, C.; He, Q.; Omosun, Y.; Igietseme, J.U.; Eko, F.O. Comparative evaluation of the protective efficacy of two formulations of a recombinant Chlamydia abortus subunit candidate vaccine in a mouse model. Vaccine 2015, 33, 1865–1872. [Google Scholar] [CrossRef]
- Eko, F.O.; Mania-Pramanik, J.; Pais, R.; Pan, Q.; Okenu, D.M.; Johnson, A.; Ibegbu, C.; He, C.; He, Q.; Russell, R.; et al. Vibrio cholerae ghosts (VCG) exert immunomodulatory effect on dendritic cells for enhanced antigen presentation and induction of protective immunity. BMC Immunol 2014, 15, 584. [Google Scholar] [CrossRef]
- Pais, R.; Omosun, Y.; He, Q.; Blas-Machado, U.; Black, C.; Igietseme, J.U.; Fujihashi, K.; Eko, F.O. Rectal administration of a chlamydial subunit vaccine protects against genital infection and upper reproductive tract pathology in mice. PLoS One 2017, 12, e0178537. [Google Scholar] [CrossRef]
- Fujihashi, K.; Koga, T.; van Ginkel, F.W.; Hagiwara, Y.; McGhee, J.R. A dilemma for mucosal vaccination: efficacy versus toxicity using enterotoxin-based adjuvants. Vaccine 2002, 20, 2431–2438. [Google Scholar] [CrossRef]
- Ekong, E.E.; Okenu, D.; Mania-Pramanik, J.; He, Q.; Igietseme, J.; Ananaba, G.; Lyn, D.; Black, C.; Eko, F. A Vibrio cholerae ghost-based subunit vaccine induces cross-protective chlamydial immunity that is enhanced by CTA2B, the nontoxic derivative of cholera toxin. FEMS Immunology & Medical Microbiology 2009, 55, 280–291. [Google Scholar]
- Bemark, M.; Bergqvist, P.; Stensson, A.; Holmberg, A.; Mattsson, J.; Lycke, N.Y. A unique role of the cholera toxin A1-DD adjuvant for long-term plasma and memory B cell development. J Immunol 2011, 186, 1399–1410. [Google Scholar] [CrossRef]
- Cunningham, K.A.; Carey, A.J.; Lycke, N.; Timms, P.; Beagley, K.W. CTA1-DD is an effective adjuvant for targeting anti-chlamydial immunity to the murine genital mucosa. J Reprod Immunol 2009, 81, 34–38. [Google Scholar] [CrossRef]
- Schussek, S.; Bernasconi, V.; Mattsson, J.; Wenzel, U.A.; Strömberg, A.; Gribonika, I.; Schön, K.; Lycke, N.Y. The CTA1-DD adjuvant strongly potentiates follicular dendritic cell function and germinal center formation, which results in improved neonatal immunization. Mucosal Immunol 2020, 13, 545–557. [Google Scholar] [CrossRef]
- Vasilevsky, S.; Greub, G.; Nardelli-Haefliger, D.; Baud, D. Genital Chlamydia trachomatis: understanding the roles of innate and adaptive immunity in vaccine research. Clin Microbiol Rev 2014, 27, 346–370. [Google Scholar] [CrossRef] [PubMed]
- Rixon, J.A.; Depew, C.E.; McSorley, S.J. Th1 cells are dispensable for primary clearance of Chlamydia from the female reproductive tract of mice. PLoS Pathog 2022, 18, e1010333. [Google Scholar] [CrossRef] [PubMed]
- Tifrea, D.F.; Pal, S.; de la Maza, L.M. A Recombinant Chlamydia trachomatis MOMP Vaccine Elicits Cross-serogroup Protection in Mice Against Vaginal Shedding and Infertility. J Infect Dis 2020, 221, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Murthy, A.K.; Chambers, J.P.; Meier, P.A.; Zhong, G.; Arulanandam, B.P. Intranasal Vaccination with a Secreted Chlamydial Protein Enhances Resolution of Genital Chlamydia muridarum Infection, Protects against Oviduct Pathology, and Is Highly Dependent upon Endogenous Gamma Interferon Production. Infect. Immun. 2007, 75, 666–676. [Google Scholar] [CrossRef]
- O’Meara, C.P.; Armitage, C.W.; Harvie, M.C.G.; Timms, P.; Lycke, N.Y.; Beagley, K.W. Immunization with a MOMP-Based Vaccine Protects Mice against a Pulmonary Chlamydia Challenge and Identifies a Disconnection between Infection and Pathology. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Lewis, D.J.; Huo, Z.; Barnett, S.; Kromann, I.; Giemza, R.; Galiza, E.; Woodrow, M.; Thierry-Carstensen, B.; Andersen, P.; Novicki, D.; et al. Transient facial nerve paralysis (Bell's palsy) following intranasal delivery of a genetically detoxified mutant of Escherichia coli heat labile toxin. PLoS One 2009, 4, e6999. [Google Scholar] [CrossRef]
- Carmichael, J.R.; Pal, S.; Tifrea, D.; de la Maza, L.M. Induction of protection against vaginal shedding and infertility by a recombinant Chlamydia vaccine. Vaccine 2011, 29, 5276–5283. [Google Scholar] [CrossRef]
- Zhu, C.; Lin, H.; Tang, L.; Chen, J.; Wu, Y.; Zhong, G. Oral Chlamydia vaccination induces transmucosal protection in the airway. Vaccine 2018, 36, 2061–2068. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, C.; Zhang, T.; Tian, Q.; Zhang, N.; Morrison, S.; Morrison, R.; Xue, M.; Zhong, G. Nonpathogenic Colonization with Chlamydia in the Gastrointestinal Tract as Oral Vaccination for Inducing Transmucosal Protection. Infect Immun 2018, 86. [Google Scholar] [CrossRef]
- Morrison, S.G.; Morrison, R.P. The protective effect of antibody in immunity to murine chlamydial genital tract reinfection is independent of immunoglobulin A. Infect Immun 2005, 73, 6183–6186. [Google Scholar] [CrossRef]
- Brunham, R.C.; Kuo, C.-C.; Cles, L.; Holmes, K.K. Correlation of host immune response with quantitative recovery of Chlamydia trachomatis from the human endocervix. Infect.Immun. 1983, 39, 1491–1494. [Google Scholar] [CrossRef]
- Morrison, R.P.; Feilzer, K.; Tumas, D.B. Gene knockout mice establish a primary protective role for major histocompatibility complex class II-restricted responses in Chlamydia trachomatis genital tract infection. Infect. Immun 1995, 63, 4661–4668. [Google Scholar] [CrossRef]
- Shillova, N.; Howe, S.E.; Hyseni, B.; Ridgell, D.; Fisher, D.J.; Konjufca, V. Chlamydia-Specific IgA Secretion in the Female Reproductive Tract Induced via Per-Oral Immunization Confers Protection against Primary Chlamydia Challenge. Infect Immun 2020, 89. [Google Scholar] [CrossRef] [PubMed]
- Igietseme, J.U.; Eko, F.O.; Black, C.M. Contemporary approaches to designing and evaluating vaccines against Chlamydia. Expert Rev Vaccines 2003, 2, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.; Caldwell, H. Immunity to murine chlamydial genital infection. Infect Immun 2002, 70, 2741–2751. [Google Scholar] [CrossRef]
- Li, W.; Murthy, A.K.; Guentzel, M.N.; Seshu, J.; Forsthuber, T.G.; Zhong, G.; Arulanandam, B.P. Antigen-Specific CD4+ T Cells Produce Sufficient IFN-{gamma} to Mediate Robust Protective Immunity against Genital Chlamydia muridarum Infection. J Immunol 2008, 180, 3375–3382. [Google Scholar] [CrossRef]
- Nogueira, C.V.; Zhang, X.; Giovannone, N.; Sennott, E.L.; Starnbach, M.N. Protective immunity against Chlamydia trachomatis can engage both CD4+ and CD8+ T cells and bridge the respiratory and genital mucosae. J Immunol 2015, 194, 2319–2329. [Google Scholar] [CrossRef]
- Zhong, G.; de la Maza, L.M. Activation of mouse peritoneal macrophages in vitro or in vivo by recombinant murine gamma interferon inhibits the growth of Chlamydia trachomatis serovar L1. Infect.Immun. 1988, 56, 3322–3325. [Google Scholar] [CrossRef]
- Rank, R.G.; Whittum-Hudson, J.A. Protective immunity to chlamydial genital infection: evidence from animal studies. J Infect Dis 2010, 201 Suppl 2, S168–177. [Google Scholar] [CrossRef]
- Vicetti Miguel, R.D.; Quispe Calla, N.E.; Pavelko, S.D.; Cherpes, T.L. Intravaginal Chlamydia trachomatis Challenge Infection Elicits TH1 and TH17 Immune Responses in Mice That Promote Pathogen Clearance and Genital Tract Damage. PLoS One 2016, 11, e0162445. [Google Scholar] [CrossRef]
- Sturdevant, g.; Caldwell, H. Innate immunity is sufficient for the clearance of Chlamydia trachomatis from the female mouse genital tract. Pathog Dis. 2014, 72, 72–72. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.T.; Braun, K.M.; Carabeo, R.A. Characterization of the Growth of Chlamydia trachomatis in In Vitro-Generated Stratified Epithelium. Front Cell Infect Microbiol 2017, 7, 438. [Google Scholar] [CrossRef] [PubMed]
- Murthy, A.; Chaganty, B.; Li, W.; Guentzel, M.; Chambers, J.; Seshu, J.; G, Z.; Arulanandam, B. A limited role for antibody in protective immunity induced by rCPAF and CpG vaccination against primary genital Chlamydia muridarum challenge. FEMS Immunol Med Microbiol. 2009, 2009, 2009–2009. [Google Scholar]
- O'Meara, C.P.; Armitage, C.W.; Harvie, M.C.; Andrew, D.W.; Timms, P.; Lycke, N.Y.; Beagley, K.W. Immunity against a Chlamydia infection and disease may be determined by a balance of IL-17 signaling. Immunol Cell Biol 2014, 92, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Peterson, E.M.; Rappuoli, R.; Ratti, G.; de la Maza, L.M. Immunization with the Chlamydia trachomatis major outer membrane protein, using adjuvants developed for human vaccines, can induce partial protection in a mouse model against a genital challenge. Vaccine 2006, 24, 766–775. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, K.; Tan, Y.; Wen, Y.; Wang, C.; Chen, Q.; Yu, J.; Xu, M.; Tan, M.; Wu, Y. A recombinant multi-epitope peptide vaccine based on MOMP and CPSIT_p6 protein protects against Chlamydia psittaci lung infection. Appl Microbiol Biotechnol 2019, 103, 941–952. [Google Scholar] [CrossRef]
- Xin, Q.; Niu, H.; Li, Z.; Zhang, G.; Hu, L.; Wang, B.; Li, J.; Yu, H.; Liu, W.; Wang, Y.; et al. Subunit vaccine consisting of multi-stage antigens has high protective efficacy against Mycobacterium tuberculosis infection in mice. PLoS One 2013, 8, e72745. [Google Scholar] [CrossRef]
- de la Maza, L.M.; Zhong, G.; Brunham, R.C. Update on Chlamydia trachomatis Vaccinology. Clin Vaccine Immunol 2017, 24. [Google Scholar] [CrossRef]
- Tanner, T.; Medhavi, F.N.U.; Richardson, S.; Omosun, Y.O.; Eko, F.O. In silico design and analysis of a multiepitope vaccine against Chlamydia. Pathogens and disease 2024, 82. [Google Scholar] [CrossRef]
Figure 1.
Design and construction of the vaccine vector, pCT-MECA and expression of rMECA. (A) Twenty (20) immunogenic T and B cell epitopes from five CT outer membrane proteins were selected and fused with CTA1 using linkers. (B) The 1,980 bp synthesized coding sequence was inserted into the periplasmic targeting expression vector, pFLAG-CTS in frame with the Flag Tag sequence to generate plasmid pCT-MECA. (C) Following transformation of V. cholerae V912 harboring the pDKLO1 lysis plasmid with plasmid pCT-MECA and production of rVCG-MECA, the expression of rMECA was confirmed by Western immunoblotting analysis of lyophilized rVCG-MECA samples using anti-Flag monoclonal antibodies. Lane 1, uninduced pCT-MECA control, and lane 2, rMECA 4 h post IPTG induction. MW, Molecular weight marker in kilodaltons (kDa).
Figure 1.
Design and construction of the vaccine vector, pCT-MECA and expression of rMECA. (A) Twenty (20) immunogenic T and B cell epitopes from five CT outer membrane proteins were selected and fused with CTA1 using linkers. (B) The 1,980 bp synthesized coding sequence was inserted into the periplasmic targeting expression vector, pFLAG-CTS in frame with the Flag Tag sequence to generate plasmid pCT-MECA. (C) Following transformation of V. cholerae V912 harboring the pDKLO1 lysis plasmid with plasmid pCT-MECA and production of rVCG-MECA, the expression of rMECA was confirmed by Western immunoblotting analysis of lyophilized rVCG-MECA samples using anti-Flag monoclonal antibodies. Lane 1, uninduced pCT-MECA control, and lane 2, rMECA 4 h post IPTG induction. MW, Molecular weight marker in kilodaltons (kDa).
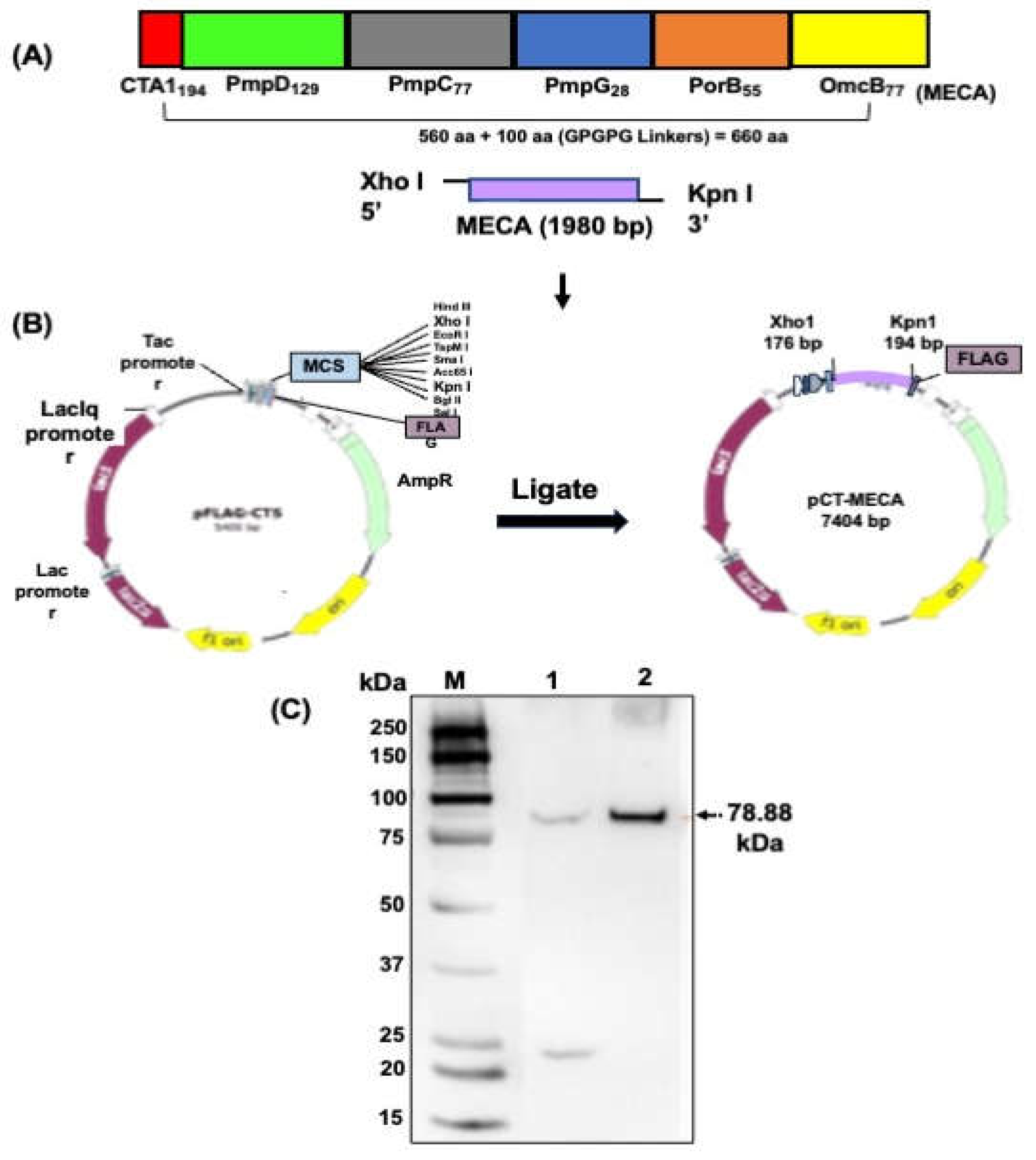
Figure 3.
CT-specific systemic and mucosal antibody responses elicited following immunization. Groups of mice were immunized three times, two weeks apart via the IM or IN route as described in the materials and methods section. Serum obtained from blood and vaginal lavage samples were obtained two weeks after the first, second and third immunizations. A standard antibody ELISA procedure was used to assess the IgG, IgG2c and IgA concentrations in serum and vaginal wash samples. The results, from three independent ELISA assays, were generated simultaneously with a standard curve and display data sets corresponding to absorbance values as mean concentrations (ng/ml) ± SD of triplicate cultures for each experiment. The data show the mean antibody concentrations elicited in serum (A, C and E) and vaginal wash (B, D and F) samples. Significant differences between groups were evaluated by one-way ANOVA with Tukey’s post multiple comparison test at (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Figure 3.
CT-specific systemic and mucosal antibody responses elicited following immunization. Groups of mice were immunized three times, two weeks apart via the IM or IN route as described in the materials and methods section. Serum obtained from blood and vaginal lavage samples were obtained two weeks after the first, second and third immunizations. A standard antibody ELISA procedure was used to assess the IgG, IgG2c and IgA concentrations in serum and vaginal wash samples. The results, from three independent ELISA assays, were generated simultaneously with a standard curve and display data sets corresponding to absorbance values as mean concentrations (ng/ml) ± SD of triplicate cultures for each experiment. The data show the mean antibody concentrations elicited in serum (A, C and E) and vaginal wash (B, D and F) samples. Significant differences between groups were evaluated by one-way ANOVA with Tukey’s post multiple comparison test at (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
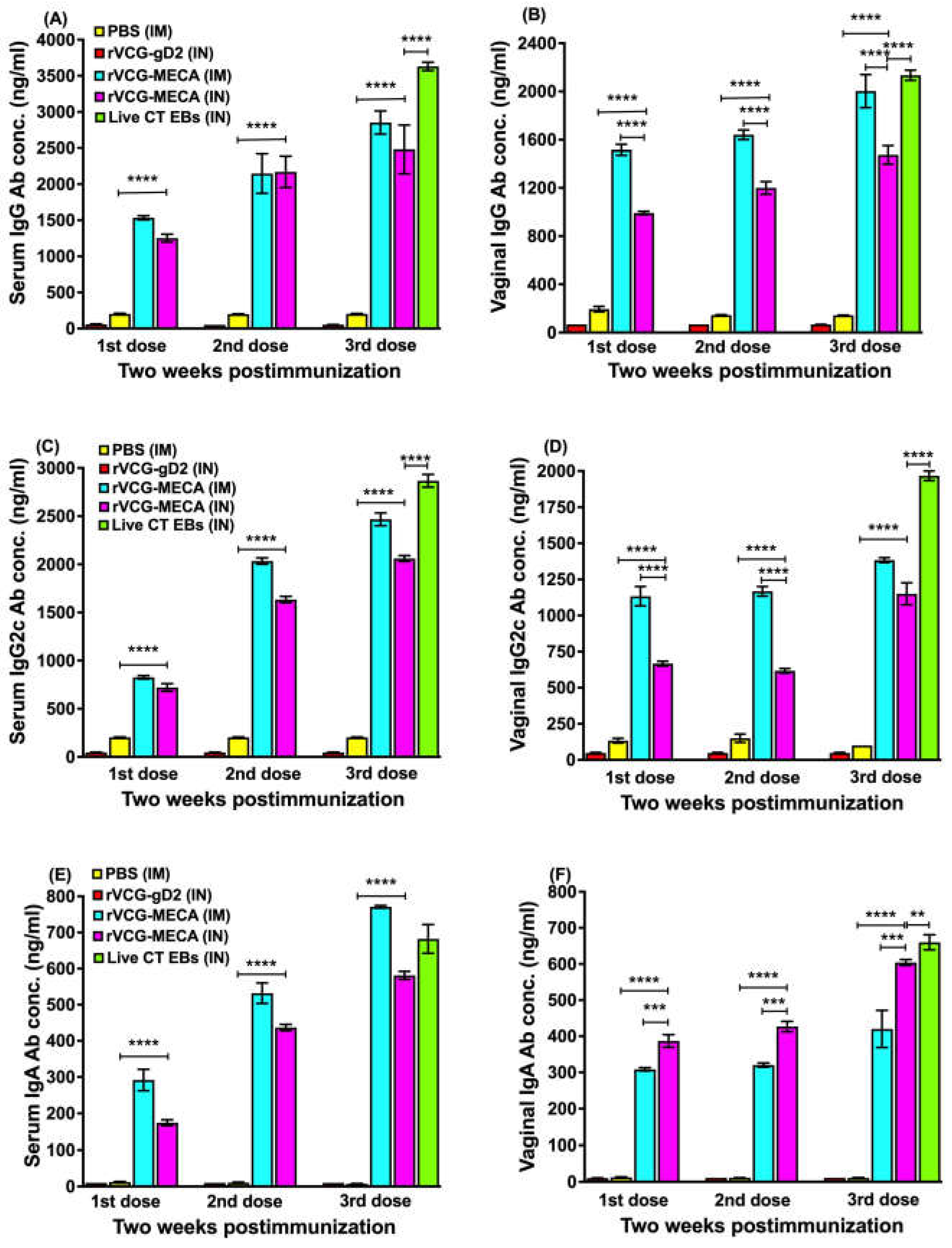
Figure 4.
Comparison of the Th1-associated IgG2c and Th2-associated IgG1 antibodies elicited in serum. Groups of mice were immunized three times, 2 weeks apart via the IM or IN route as described above. Serum samples were obtained from blood 2 weeks after the last immunization and pooled for each group. CT-specific IgG2a and IgG1 antibody concentrations were measured by a standard antibody ELISA procedure. Results generated simultaneously with antibody standards display data sets as mean concentrations (ng/ml) + SD of triplicate cultures for each experiment. The data is from one of two independent assays with similar results and show (A) the mean IgG2a and IgG1 antibody concentrations and (B) the IgG2a/IgG1 ratios. Significant differences between experimental groups were evaluated by One-way ANOVA with Tukey’s post multiple comparison test at (**p < 0.01; ****p < 0.0001).
Figure 4.
Comparison of the Th1-associated IgG2c and Th2-associated IgG1 antibodies elicited in serum. Groups of mice were immunized three times, 2 weeks apart via the IM or IN route as described above. Serum samples were obtained from blood 2 weeks after the last immunization and pooled for each group. CT-specific IgG2a and IgG1 antibody concentrations were measured by a standard antibody ELISA procedure. Results generated simultaneously with antibody standards display data sets as mean concentrations (ng/ml) + SD of triplicate cultures for each experiment. The data is from one of two independent assays with similar results and show (A) the mean IgG2a and IgG1 antibody concentrations and (B) the IgG2a/IgG1 ratios. Significant differences between experimental groups were evaluated by One-way ANOVA with Tukey’s post multiple comparison test at (**p < 0.01; ****p < 0.0001).
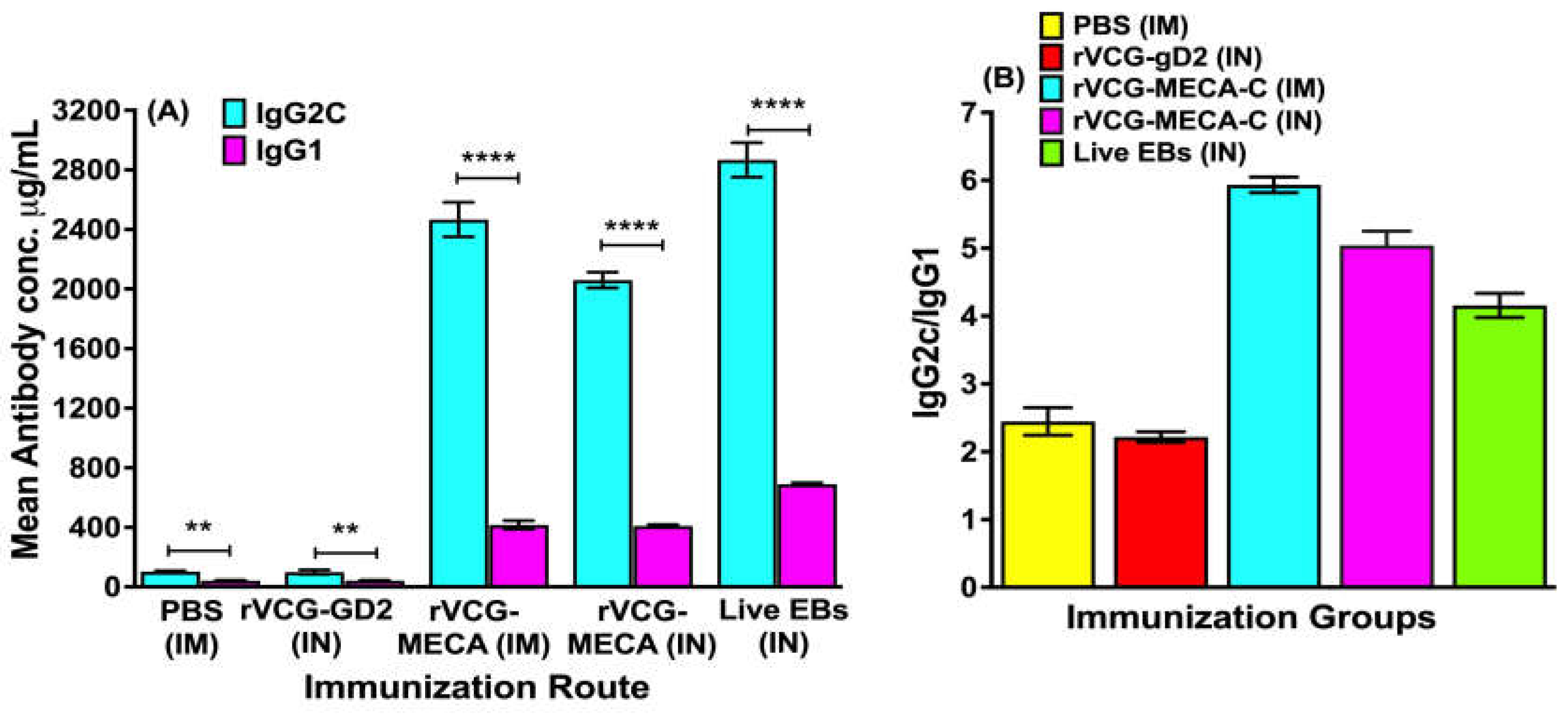
Figure 5.
Long-term CT-specific antibody responses elicited following IM and IN immunization. Groups of mice were immunized three times, 2 weeks apart via the IM or IN route as described above. Serum and vaginal wash samples were obtained from each mouse per group at 12 and 16 weeks after the last immunization. Antibody concentrations were measured by a standard antibody ELISA procedure. Results generated simultaneously with antibody standards display data sets as mean concentrations (ng/ml) + SD of triplicate cultures for each experiment. The data show the mean IgG2c and IgA antibody concentrations elicited in serum (A and C) and vaginal wash (B and D) samples. Significant differences between experimental groups were evaluated by One-way ANOVA with Tukey’s post multiple comparison test at (****p < 0.0001).
Figure 5.
Long-term CT-specific antibody responses elicited following IM and IN immunization. Groups of mice were immunized three times, 2 weeks apart via the IM or IN route as described above. Serum and vaginal wash samples were obtained from each mouse per group at 12 and 16 weeks after the last immunization. Antibody concentrations were measured by a standard antibody ELISA procedure. Results generated simultaneously with antibody standards display data sets as mean concentrations (ng/ml) + SD of triplicate cultures for each experiment. The data show the mean IgG2c and IgA antibody concentrations elicited in serum (A and C) and vaginal wash (B and D) samples. Significant differences between experimental groups were evaluated by One-way ANOVA with Tukey’s post multiple comparison test at (****p < 0.0001).
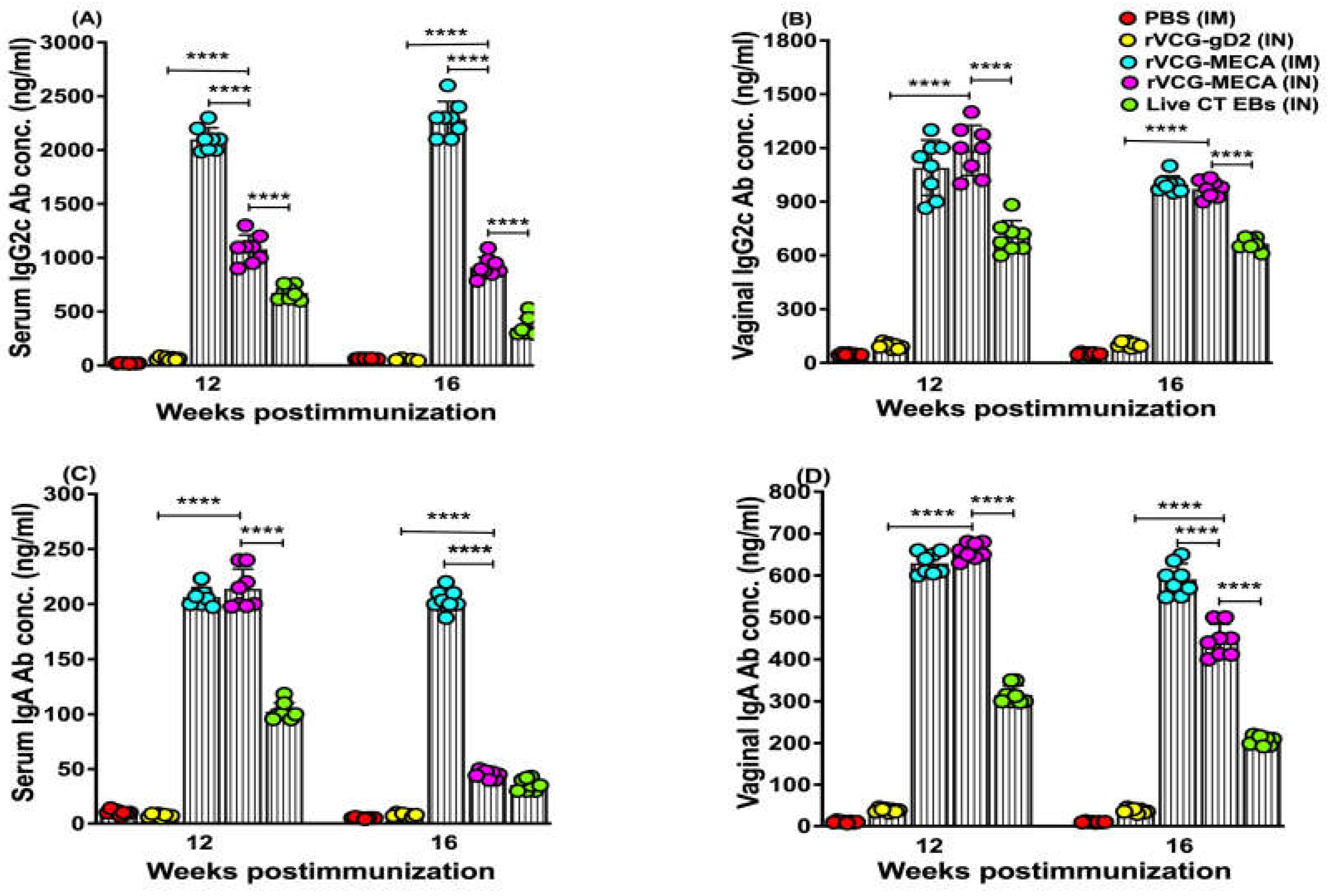
Figure 6.
Relative Avidity Index of CT-specific serum IgG and IgG2c antibodies after IM and IN immunization. The CT-specific IgG and IgG2c antibody avidity of serum samples collected from each immunization group at 2 and 4 weeks postimmunization was determined using a modified antibody ELISA assay in combination with the chaotropic agent, ammonium thiocyanate (NH4SCN). Results were generated simultaneously with a standard curve and data sets corresponding to absorbance values were calculated as mean concentrations(ng/ml) ± SD of triplicate cultures for each experiment. This experiment was repeated with similar results. The relative avidity index was calculated and displayed as a percentage of the ratio of the antibody concentration of samples treated with NH4SCN and the antibody concentration of untreated samples. The data shows the Percent Relative Avidity Index of serum IgG (A) and IgG2c (B) antibodies using 2 M NH4SCN. Significant differences between experimental groups was compared by One-way ANOVA with Tukey’s post multiple comparison test at (***p < 0.001, ****p < 0.0001).
Figure 6.
Relative Avidity Index of CT-specific serum IgG and IgG2c antibodies after IM and IN immunization. The CT-specific IgG and IgG2c antibody avidity of serum samples collected from each immunization group at 2 and 4 weeks postimmunization was determined using a modified antibody ELISA assay in combination with the chaotropic agent, ammonium thiocyanate (NH4SCN). Results were generated simultaneously with a standard curve and data sets corresponding to absorbance values were calculated as mean concentrations(ng/ml) ± SD of triplicate cultures for each experiment. This experiment was repeated with similar results. The relative avidity index was calculated and displayed as a percentage of the ratio of the antibody concentration of samples treated with NH4SCN and the antibody concentration of untreated samples. The data shows the Percent Relative Avidity Index of serum IgG (A) and IgG2c (B) antibodies using 2 M NH4SCN. Significant differences between experimental groups was compared by One-way ANOVA with Tukey’s post multiple comparison test at (***p < 0.001, ****p < 0.0001).
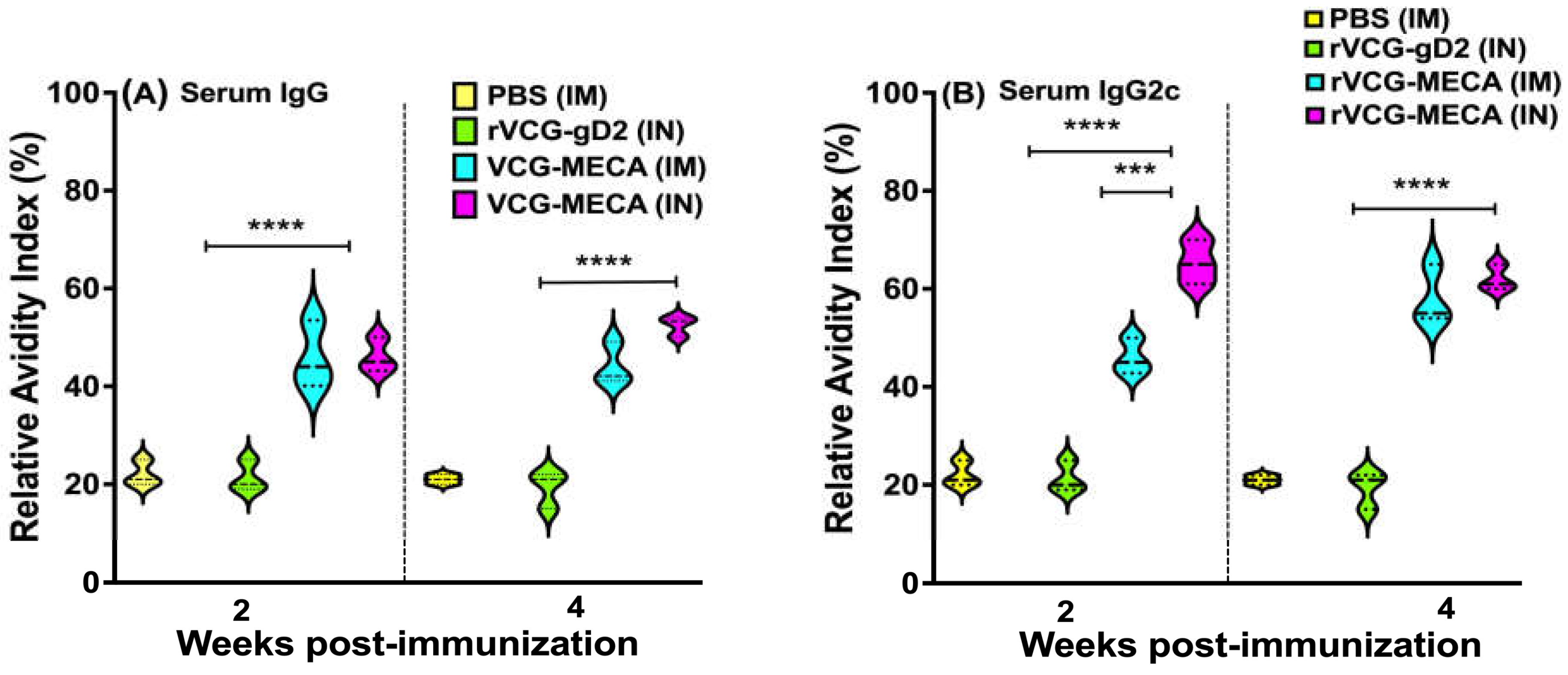
Figure 7.
CT-specific mucosal and systemic Th1/Th2 cytokine responses. Immune T cells purified from the spleens and ILNs of immunized mice and controls 4 weeks postimmunization were restimulated in vitro with CT serovar D antigen (UV-irradiated EBs; 10 microgram/ml). The amount of CT-specific IFN-γ (Th1) and IL-4 (Th2) cytokines contained in supernatants of culture-stimulated CD4+ T cells was measured using Bio-Plex cytokine assay kit in combination with the Bio-Plex Manager software. The concentration of the cytokines in each sample was obtained by extrapolation from a standard calibration curve generated simultaneously. Data were calculated as the mean values (± S.D.) for quadruplicate cultures for each experiment. The results are from two independent experiments and are shown as mean IFN-γ (A) and IL-4 (B) cytokine concentrations (pg/ml) ± SD. Significant differences between experimental groups were evaluated by one-way ANOVA with Tukey’s post multiple comparison test at (*p < 0.05, **p < 0.01, ****p < 0.0001).
Figure 7.
CT-specific mucosal and systemic Th1/Th2 cytokine responses. Immune T cells purified from the spleens and ILNs of immunized mice and controls 4 weeks postimmunization were restimulated in vitro with CT serovar D antigen (UV-irradiated EBs; 10 microgram/ml). The amount of CT-specific IFN-γ (Th1) and IL-4 (Th2) cytokines contained in supernatants of culture-stimulated CD4+ T cells was measured using Bio-Plex cytokine assay kit in combination with the Bio-Plex Manager software. The concentration of the cytokines in each sample was obtained by extrapolation from a standard calibration curve generated simultaneously. Data were calculated as the mean values (± S.D.) for quadruplicate cultures for each experiment. The results are from two independent experiments and are shown as mean IFN-γ (A) and IL-4 (B) cytokine concentrations (pg/ml) ± SD. Significant differences between experimental groups were evaluated by one-way ANOVA with Tukey’s post multiple comparison test at (*p < 0.05, **p < 0.01, ****p < 0.0001).
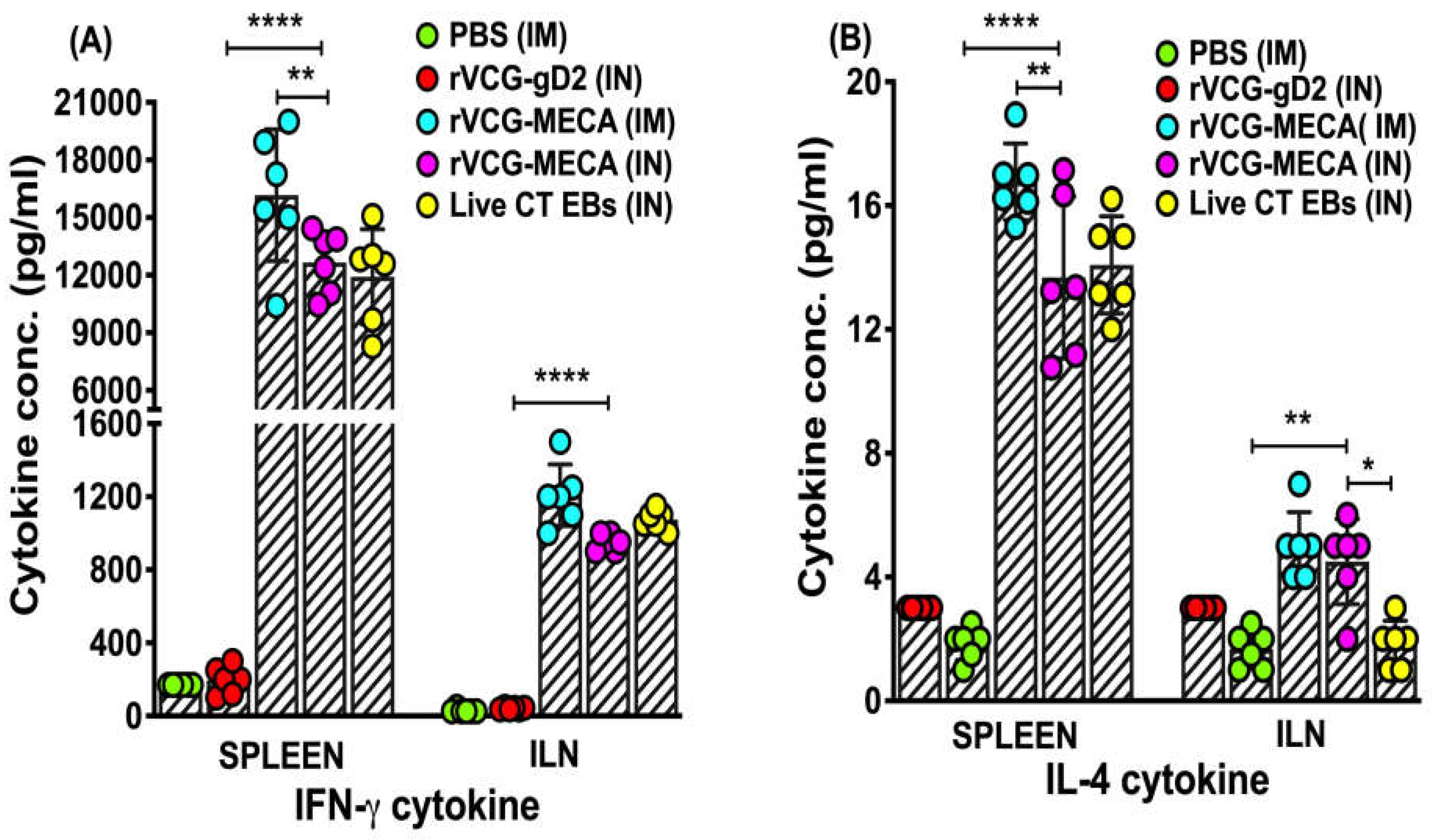
Figure 8.
Protection against transcervical challenge with CT serovar D. Groups of mice immunized IM or IN as described above were challenged transcervically with 1x106 IFU of live CT 4 weeks after the last immunization. One week prior to challenge, mice were administered Depo Provera to stabilize the estrous cycle and facilitate a productive infection. Infections were monitored by cervicovaginal swabbing of individual animals every three days for 30 days, and Chlamydia was isolated from swabs in tissue culture and enumerated. The data show the individual recoverable IFU/ml from each mouse and the mean IFUs per group. Differences between experimental groups were compared by one-way ANOVA with Tukey’s post multiple comparison test at ****p < 0.0001.
Figure 8.
Protection against transcervical challenge with CT serovar D. Groups of mice immunized IM or IN as described above were challenged transcervically with 1x106 IFU of live CT 4 weeks after the last immunization. One week prior to challenge, mice were administered Depo Provera to stabilize the estrous cycle and facilitate a productive infection. Infections were monitored by cervicovaginal swabbing of individual animals every three days for 30 days, and Chlamydia was isolated from swabs in tissue culture and enumerated. The data show the individual recoverable IFU/ml from each mouse and the mean IFUs per group. Differences between experimental groups were compared by one-way ANOVA with Tukey’s post multiple comparison test at ****p < 0.0001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



