
Submitted:
25 December 2024
Posted:
26 December 2024
You are already at the latest version
Abstract
Background/Objectives: Vitamin D deficiency has been frequently observed in every generation worldwide. Pregnant women are no exception. The associations between maternal vitamin D status and fetal growth, immune system, or metabolism are reported. These infant characteristics are also reported to be associated with epigenetic gestational age at birth. Epigenetic gestational age acceleration at birth longitudinally relates to infant health. This study aimed to investigate the associations between maternal serum vitamin D levels and infants' epigenetic gestational age acceleration in Japan. Methods: The data were obtained from the hospital-based birth cohort study conducted at the National Center for Child Health and Development in Tokyo, Japan. Maternal blood was collected in the 2nd trimester to measure serum vitamin D concentration. Cord blood was collected at birth to measure serum vitamin D and to extract DNA. DNA methylation was assessed using Illumina methylation EPIC array. Epigenetic gestational age was calculated using “methylclock” R package. Linear regression analysis was performed to see associations. Results: Maternal serum vitamin D levels in the 2nd trimester were negatively associated with epigenetic gestational age acceleration at birth when calculated by Bohlin’s method (regression coefficient [95% CI]: -0.022 [-0.039, -0.005], n = 157), which was still significant after considering infants sex (-0.022 [-0.039, -0.005]). Cord blood serum vitamin D levels were not associated with epigenetic age acceleration. Maternal age at delivery and birth height were associated in positive and negative ways with epigenetic gestational age acceleration, respectively (0.048 [0.012, 0.085] and -0.075 [-0.146, -0.003]). Conclusions: Maternal vitamin D deficiency was related to an infant’s epigenetic gestational age acceleration at birth. These results indicate that maternal vitamin D epigenetically programs fetal development which may affect an infant's health after birth.
Keywords:
maternal serum vitamin D
; epigenetic clock
; cord blood
; DNA methylation
1. Introduction
Vitamin D deficiency has been frequently observed in every generation worldwide. Pregnant women are no exception [1]. The prevalence of low vitamin D status in pregnant women was especially high in Asia [2,3]. Maternal vitamin D intake during pregnancy or cord blood 25-hydroxyvitamin D (25[OH]D) is inversely associated with the risk of recurrent wheezing in children [4,5]. In addition, prenatal vitamin D deficiency is shown to sex-dependently program adipose tissue metabolism in offspring [6] or epigenetically program embryonic hematopoietic stem cells, resulting in adipose macrophage infiltration and type 2 diabetes in mice [7]. These results indicate that immune cells are epigenetically affected in utero by vitamin D deficiency.
Epigenetics are very informative biological markers. DNA methylation patterns in several tissues have been used to predict age accurately. This epigenetic age is recognized as a biological clock. Epigenetic age acceleration increases adult mortality risk [8,9]. Similarly, epigenetic gestational age acceleration based on DNA methylation patterns in cord blood cells is recognized as a biomarker of physiological development [10,11,12]. In addition to cord blood cells, the pediatric buccal epigenetic clock has recently been developed to non-invasively predict epigenetic age acceleration in infants [13], which showed increased epigenetic age acceleration at term equivalent age in very preterm infants with neonatal infection [14]. Long-lasting effects of neonatal environments are also detected in buccal DNA methylation. Epigenetic age acceleration based on buccal cells at age 30 to 35 years in men with extremely low birth weight was significantly more than those with normal birth weight [15].
From these backgrounds, we aimed to investigate an association between maternal vitamin D levels in gestation and epigenetic gestational age acceleration at birth to assess whether maternal vitamin D deficiency influences fetal development. Vitamin D is commonly referred to as vitamin D2 and vitamin D3. In human blood, vitamin D3 is much more abundant than vitamin D2. Vitamin D3 is formed from provitamin D3, the final intermediate in cholesterol biosynthesis in the body, which is converted in the skin to pre-vitamin D3 by exposure to ultraviolet radiation and then to vitamin D3 by body temperature. Vitamin D is hydroxylated in the liver to 25-hydroxy vitamin D (25(OH)D), which circulates stably in the blood. Eventually, the biologically active form of vitamin D is 1,25-dihydroxyvitamin D, made from 25(OH)D by further hydroxylation in the kidney. Different people have different backgrounds in these biosynthetic processes. So far, Maternal vitamin D3 supplementation slows infants’ epigenetic gestational age at birth in a randomized controlled trial [16]. We considered that the blood 25(OH)D level measurement more accurately represented the nutritional status than information on the presence or absence of supplementation intake. In the present study, we measured serum 25(OH)D concentrations in maternal 2nd trimester peripheral blood and cord blood, then assessed genome-wide DNA methylation of cord blood cells to see the effects of gestational vitamin D on the epigenetic age acceleration.
2. Materials and Methods
Participants
The participants of this study were recruited from a hospital-based birth study conducted at the National Center for Child Health and Development (NCCHD) in Tokyo, Japan, from 2010 to 2013. In the study, pregnant women were recruited during the first trimester of pregnancy. Their informed consent for participation and that of their newborns was obtained from them at the beginning of the study. Blood samples were collected from mothers in 2nd trimester between 24 to 28 gestational weeks. Cord blood samples were collected at delivery. Mother-child pairs were excluded if their mothers smoked during pregnancy, were diagnosed with a pregnancy complication, or had a pre-existing disease. In addition, we filtered out newborns who were delivered after 37 gestational weeks and those with a birth weight within the 90th percentile because stillbirth and small or large birth weight have an influence on DNA methylation in cord blood cells. Finally, we focused on maternal intrinsic vitamin D levels. Samples were randomly selected so that the population’s median blood 25(OH)D level could be 20 ng/ml, which was considered adequate by IOM 2011.
Maternal and Newborn Assessments
Pre-pregnancy BMI was defined as pre-pregnancy weight (kg) divided by height (m2), which was reported in the questionnaire. Gestational weight gain was calculated by subtracting the above pre-pregnant weight from the weight at the latest prenatal checkup. Gestational age (GA) was estimated using a combined method based on the last menstrual period, first-trimester ultrasound, and 20-week ultrasound. Birth weight SD score – namely birth weight z-score for GA according to Japanese reference data – was calculated from GA, birth weight, infants’ sex, and parity, through a program provided by the Japanese Society for Pediatric Endocrinology (downloaded from following site on 10th, December, 2024; http://jspe.umin.jp/medical/keisan.html). Birth height SD score was calculated from GA, birth height, and infant’s sex, through the same program as well. Blood samples were stored at 4°C until assayed the next day. Maternal or cord blood serum 25(OH)D concentrations were measured by an external laboratory (LSI Medience, Itabashi, Tokyo, Japan) using a competitive protein-binding assay as previously described [17,18]. The inter- and intra-assay coefficient of variation was ≤10%.
DNA Methylation Analysis Using Umbilical Cord Blood Samples
DNA was collected from whole cord blood cells. DNA Methylation data were obtained using the Infinium MethylationEPIC BeadChip array (Illumina, San Diego, CA, USA) as previously described [19,20]. Methylation data were obtained with the iScan system (Illumina) as idat files and then processed using the ENmix (v.1.30.3) package in R-4.1.3 [21]. The manifest file was annotated using “IlluminaHumanMethylationEPICanno.ilm10b4.hg19”. Beta values were calculated by “mpreprocess” of ENmix.
Calculation of Gestational Age Acceleration
Statistical Analyses
Association tests were performed with linear regression with and without adjustment for the infant’s sex. Spearman correlation tests were performed to see the relationship between characteristics and epigenetic age acceleration.
3. Results
3.1. Subsection
1. General characteristics of the study population
The median serum 25(OH)D levels at mid-gestation of 157 participants was 20.5 ng/ml (Figure 1). 76 of the 157 participants had 25(OH)D levels below 20 ng/ml, which the IOM defined as an adequate serum level. Data on gestational weeks at birth, birth weight, birth height, and cord blood 25(OH)D levels were collected for the newborn’s information. For the parents, data were collected on maternal age, paternal age, pre-pregnancy BMI, and gestational weight gain (Table 1).
2. Correlation of DNAmGA with chronological gestational age
DNAmGA were significantly correlated with chronological gestational age even though our samples were collected at limited gestational age, such as between 37 to 42 weeks. The correlation coefficient was better in Bohlin’s method (r = 0.71) than in Knight’s method (r = 0.48) (Figure 2). Bohlin’s method estimated older DNAmGA than Knight’s method.
3. Gestational age acceleration at birth was associated with maternal serum 25(OH)D levels but not with cord blood 25(OH)D
A linear regression analysis between DNAmGA age accelerations and each character regarding perinatal factors revealed significant associations with the ones estimated by only Bohlin’s method (Table 2). Maternal blood 25(OH)D levels in mid-gestation and birth height showed significant negative associations with a regression coefficient of -0.022 (-0.039 to -0.005: 95%CI). There was no significant association between DNAmGA age accelerations and cord blood 25(OH)D levels. Also, maternal age at delivery and birth height showed significant positive and negative associations with DNAmGA age accelerations with a regression coefficient of 0.049 (0.013 to 0.085: 95%CI) and -0.071 (-0.142 to -0.005: 95%CI), respectively (Table 2). These associations were still significant after adjustment for the infant’s sex (Table 3). Correlation coefficients between Maternal blood 25(OH)D levels and DNAmGA age accelerations were -0.227 and -0.170 in male and female infants, respectively (Figure 3). Although maternal blood 25(OH)D levels and birth height were both negatively associated with DNAmGA age accelerations, they had no significant correlation (p = 0.248). There was no correlation between maternal blood 25(OH)D levels and gestational age at delivery (p = 0.409).
4. Discussion
Our results showed a stronger correlation between chronological age and DNAmGA with Bohlin’s method than with Knight’s method, as other cohort studies showed [10,23]. Furthermore, when measured in Bohlin’s method, some prenatal environments were significantly associated with DNAmGA age accelerations differently. A negative association between maternal 25(OH)D and gestational age acceleration in our study suggested that maternal 25(OH)D deficiency might accelerate fetal development improperly. In this study, we examined an association between serum 25(OH)D levels and the fetal epigenetic clock because several factors could regulate serum 25(OH)D levels [24]. For instance, the variant in GC vitamin D binding Protein, cytochrome P450 Family 27 Subfamily B Member 1, or Vitamin D Receptor gene was shown to be associated with differences in serum 25(OH)D levels in pregnant women, respectively [25]. Besides, abdominal obesity and solar UV-B exposure reduced and increased serum vitamin D levels, respectively [26,27]. However, the congruence of results with the previous report, in which supplement intake of 4000 IU/day vitamin D3 during pregnancy was negatively associated with the infant’s gestational age acceleration at birth, could indicate that active intake of vitamin D3 during pregnancy affects fetal development via the upregulation of intrinsic 25(OH)D concentration [16]. A prospective birth cohort study reported the longitudinal associations between DNAmGA and long-term obesity [28,29] or child blood pressure [30]. Although a systematic review showed no consistent evidence that vitamin D supplementation during pregnancy has clinically meaningful health benefits for infants and pregnant women [31], our results indicated that, including supplement intake, the effects of adequate vitamin D supply from mother to fetus could be observed in infants in the future through regulating epigenetic development, which assessed by DNAmGA.
Various maternal metabolites are associated with gestational age acceleration at birth. Higher maternal plasma homocysteine concentrations are positively associated with gestational age acceleration [32]. Higher homocysteine levels are caused by vitamin B12 deficiency [33], which indicates that maternal inadequate “One carbon metabolism” could affect fetal gestational age acceleration. In addition, maternal plasma fatty acid pattern characterized by higher concentrations of n-3 polyunsaturated fatty acids was reported to be associated with accelerated epigenetic gestational aging [34]. These results suggest that inclusive maternal nutrition cooperatively programs DNAmGA. It is known that Vitamin D deficiency increases the risk of anemia [35,36]. Vitamin D suppresses hepcidin expression in cell levels [37,38] and is supposed to increase serum iron. In vivo, high-dose one-time oral vitamin D3 intake reduces circulating hepcidin concentration in healthy adults [39]. Regarding pregnant women, positive relationships were observed in late pregnancy between plasma 25(OH)D and hepcidin and between plasma 25(OH)D and iron status, assessed by ferritin levels. However, vitamin D3 supplementation did not affect these two levels [40]. The discrepancy between serum 25(OH)D and vitamin D intake was also reported in women of reproductive age. Hemoglobin level in women of reproductive age was associated with serum 25(OH)D but not dietary vitamin D intake [41]. Different studies also showed that plasma 25(OH)D was positively associated with hemoglobin levels in all trimesters [42,43] and inversely associated with erythropoietin at both midgestation and delivery [43]. These results suggested that assessing plasma 25(OH)D levels tells more accurate causal relationships. Further investigation is needed to elucidate how maternal plasma 25(OH)D status affects fetal DNAmGA, considering maternal anemia or iron metabolism.
Maternal age was positively associated with DNAmGA age acceleration in this study. This was partially consistent with the previous report, in which maternal age was associated with DNAmGA age acceleration of the offspring at birth but was limited to over 40 years [44]. The mean maternal age at delivery in our study is 37.5 ± 3.7. A different cohort study showed no association with maternal age at delivery, even after being classified by age as more than 35 years old. Meanwhile, the study showed a significant association between DNAmGA age acceleration and maternal parity [45]. Maternal physiological influences on the fetus cannot be derived only from maternal age. Parity and other physical characteristics must also be taken into account. Further studies are needed to elucidate the effects of maternal age. Regarding infant characteristics, several reports showed an association between DNAmGA age acceleration and birth weight. However, this study showed DNAmGA age acceleration was negatively associated with birth height. It needs further studies to reveal the associations between birth height and epigenetic clock because the SD score of birth height did not associate with DNAmGA age acceleration.
Lastly, it is reported that the first-trimester maternal serum 25(OH)D, but not one of the second-trimester, is positively associated with fetal growth patterns between 16 and 42 gestational weeks [46]. Meanwhile, second-trimester maternal 25(OH)D concentrations were positively correlated with neonates’ birth weight, body length, and head circumference in significant levels [47]. We collected maternal serum 25(OH)D levels once during gestation. Collecting the data for vitamin D levels in several time points during pregnancy could reveal which or the whole period vitamin D status is critical for DNAmGA age acceleration.
5. Conclusions
We found an association between maternal mid-pregnancy vitamin D deficiency and neonates’ gestational age acceleration estimated by DNA methylation in cord blood cells. Our result is consistent with a previous study, which showed that vitamin D supplement intake during pregnancy decreases gestational age acceleration. Our study could be limited by sample size, collection period, potential selection bias, or residual confounding. Future studies of the relationship between maternal vitamin D deficiency and neonates’ DNAmGA age acceleration could include larger sample sizes and racial/ethnically diverse samples. The longitudinal effects of maternal vitamin D and age acceleration at development stages are supposed to be considered.
Author Contributions
TK designed the study; SCJ, KO, HT, NM, and TF contributed resources; HK performed the experiments; SA contributed the analytical tools; TK analyzed the data. TK wrote the manuscript; TK, TF, and KH acquired funding; TK and KH reviewed and edited the paper. All authors read and approved the final manuscript.
Funding
This research was funded by Japan Environment and Children’s Study and the Ministry of Health, Labour and Welfare, H24-jisedai-shitei-007. JSPS KAKENHI, JP22K11793 to Tomoko Kawai. National Center for Child Health and Development (NCCHD) Valid, 2020B-21 to Tomoko Kawai: 2022A-3 to Kenichiro Hata. The Japan Foundation for Pediatric Research, 22-007 to Tomoko Kawai.
Institutional Review Board Statement
This study strictly adhered to the guidelines of the Declaration of Helsinki. All methods were performed in accordance with the following ethical guidelines in Japan: Ethics Guidelines for Human Genome/Gene Analysis Research, Ethical Guidelines for Medical and Health Research Involving Human Subjects, and Ethical Guidelines for Epidemiological Research. Ethical approval was obtained from the Ethics Committee of the National Center for Child Health and Development, Tokyo, Japan (approval ID: 234, 461). All the participants provided written informed consent for themselves and their newborns.
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, [TK], upon reasonable request.
Acknowledgments
We are deeply grateful to all of the women who participated in the present study and to the hospital staff for their cooperation.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GA | Gestational age |
| DNAmGA | DNA methylation Gestational Age |
| 25(OH)D | 25-hydroxy vitamin D |
References
- Palacios, C., and Gonzalez, L. (2014) Is vitamin D deficiency a major global public health problem? J Steroid Biochem Mol Biol 144 Pt A, 138-145.
- Palacios, C., Kostiuk, L. K., and Peña-Rosas, J. P. (2019) Vitamin D supplementation for women during pregnancy. Cochrane Database Syst Rev 7, Cd008873. [CrossRef]
- Nakanishi, K., Mutoh, M., Itoh, S., Kobayashi, S., Yamaguchi, T., Iwata, H., Tamura, N., Koishi, M., Kasai, M., Kikuchi, E., Yasuura, N., Kishi, R., and Sato, Y. (2024) Vitamin D concentration in maternal serum during pregnancy: Assessment in Hokkaido in adjunct study of the Japan Environment and Children’s Study (JECS). PLoS One 19, e0312516. [CrossRef]
- Camargo, C. A., Jr., Rifas-Shiman, S. L., Litonjua, A. A., Rich-Edwards, J. W., Weiss, S. T., Gold, D. R., Kleinman, K., and Gillman, M. W. (2007) Maternal intake of vitamin D during pregnancy and risk of recurrent wheeze in children at 3 y of age. Am J Clin Nutr 85, 788-795. [CrossRef]
- Camargo, C. A., Jr, Ingham, T., Wickens, K., Thadhani, R., Silvers, K. M., Epton, M. J., Town, G. I., Pattemore, P. K., Espinola, J. A., Crane, J., Asthma, t. N. Z., and Group, A. C. S. (2011) Cord-Blood 25-Hydroxyvitamin D Levels and Risk of Respiratory Infection, Wheezing, and Asthma. Pediatrics 127, e180-e187.
- Seipelt, E. M., Tourniaire, F., Couturier, C., Astier, J., Loriod, B., Vachon, H., Pucéat, M., Mounien, L., and Landrier, J. F. (2020) Prenatal maternal vitamin D deficiency sex-dependently programs adipose tissue metabolism and energy homeostasis in offspring. Faseb j 34, 14905-14919. [CrossRef]
- Oh, J., Riek, A. E., Bauerle, K. T., Dusso, A., McNerney, K. P., Barve, R. A., Darwech, I., Sprague, J. E., Moynihan, C., Zhang, R. M., Kutz, G., Wang, T., Xing, X., Li, D., Mrad, M., Wigge, N. M., Castelblanco, E., Collin, A., Bambouskova, M., Head, R. D., Sands, M. S., and Bernal-Mizrachi, C. (2023) Embryonic vitamin D deficiency programs hematopoietic stem cells to induce type 2 diabetes. Nat Commun 14, 3278. [CrossRef]
- Marioni, R. E., Shah, S., McRae, A. F., Chen, B. H., Colicino, E., Harris, S. E., Gibson, J., Henders, A. K., Redmond, P., Cox, S. R., Pattie, A., Corley, J., Murphy, L., Martin, N. G., Montgomery, G. W., Feinberg, A. P., Fallin, M. D., Multhaup, M. L., Jaffe, A. E., Joehanes, R., Schwartz, J., Just, A. C., Lunetta, K. L., Murabito, J. M., Starr, J. M., Horvath, S., Baccarelli, A. A., Levy, D., Visscher, P. M., Wray, N. R., and Deary, I. J. (2015) DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol 16, 25. [CrossRef]
- Horvath, S., Pirazzini, C., Bacalini, M. G., Gentilini, D., Di Blasio, A. M., Delledonne, M., Mari, D., Arosio, B., Monti, D., Passarino, G., De Rango, F., D’Aquila, P., Giuliani, C., Marasco, E., Collino, S., Descombes, P., Garagnani, P., and Franceschi, C. (2015) Decreased epigenetic age of PBMCs from Italian semi-supercentenarians and their offspring. Aging (Albany NY) 7, 1159-1170. [CrossRef]
- Bozack, A. K., Rifas-Shiman, S. L., Gold, D. R., Laubach, Z. M., Perng, W., Hivert, M. F., and Cardenas, A. (2023) DNA methylation age at birth and childhood: performance of epigenetic clocks and characteristics associated with epigenetic age acceleration in the Project Viva cohort. Clin Epigenetics 15, 62. [CrossRef]
- Bohlin, J., Håberg, S. E., Magnus, P., Reese, S. E., Gjessing, H. K., Magnus, M. C., Parr, C. L., Page, C. M., London, S. J., and Nystad, W. (2016) Prediction of gestational age based on genome-wide differentially methylated regions. Genome Biol 17, 207. [CrossRef]
- Knight, A. K., Craig, J. M., Theda, C., Bækvad-Hansen, M., Bybjerg-Grauholm, J., Hansen, C. S., Hollegaard, M. V., Hougaard, D. M., Mortensen, P. B., Weinsheimer, S. M., Werge, T. M., Brennan, P. A., Cubells, J. F., Newport, D. J., Stowe, Z. N., Cheong, J. L., Dalach, P., Doyle, L. W., Loke, Y. J., Baccarelli, A. A., Just, A. C., Wright, R. O., Téllez-Rojo, M. M., Svensson, K., Trevisi, L., Kennedy, E. M., Binder, E. B., Iurato, S., Czamara, D., Räikkönen, K., Lahti, J. M., Pesonen, A. K., Kajantie, E., Villa, P. M., Laivuori, H., Hämäläinen, E., Park, H. J., Bailey, L. B., Parets, S. E., Kilaru, V., Menon, R., Horvath, S., Bush, N. R., LeWinn, K. Z., Tylavsky, F. A., Conneely, K. N., and Smith, A. K. (2016) An epigenetic clock for gestational age at birth based on blood methylation data. Genome Biol 17, 206. [CrossRef]
- McEwen, L. M., O’Donnell, K. J., McGill, M. G., Edgar, R. D., Jones, M. J., MacIsaac, J. L., Lin, D. T. S., Ramadori, K., Morin, A., Gladish, N., Garg, E., Unternaehrer, E., Pokhvisneva, I., Karnani, N., Kee, M. Z. L., Klengel, T., Adler, N. E., Barr, R. G., Letourneau, N., Giesbrecht, G. F., Reynolds, J. N., Czamara, D., Armstrong, J. M., Essex, M. J., de Weerth, C., Beijers, R., Tollenaar, M. S., Bradley, B., Jovanovic, T., Ressler, K. J., Steiner, M., Entringer, S., Wadhwa, P. D., Buss, C., Bush, N. R., Binder, E. B., Boyce, W. T., Meaney, M. J., Horvath, S., and Kobor, M. S. (2020) The PedBE clock accurately estimates DNA methylation age in pediatric buccal cells. Proc Natl Acad Sci U S A 117, 23329-23335. [CrossRef]
- Gomaa, N., Konwar, C., Gladish, N., Au-Young, S. H., Guo, T., Sheng, M., Merrill, S. M., Kelly, E., Chau, V., Branson, H. M., Ly, L. G., Duerden, E. G., Grunau, R. E., Kobor, M. S., and Miller, S. P. (2022) Association of Pediatric Buccal Epigenetic Age Acceleration With Adverse Neonatal Brain Growth and Neurodevelopmental Outcomes Among Children Born Very Preterm With a Neonatal Infection. JAMA Netw Open 5, e2239796. [CrossRef]
- 15. Van Lieshout, R. J., McGowan, P. O., de Vega, W. C., Savoy, C. D., Morrison, K. M., Saigal, S., Mathewson, K. J., and Schmidt, L. A. (2021) Extremely Low Birth Weight and Accelerated Biological Aging. Pediatrics 147. [CrossRef]
- Chen, L., Wagner, C. L., Dong, Y., Wang, X., Shary, J. R., Huang, Y., Hollis, B. W., and Zhu, H. (2020) Effects of Maternal Vitamin D3 Supplementation on Offspring Epigenetic Clock of Gestational Age at Birth: A Post-hoc Analysis of a Randomized Controlled Trial. Epigenetics 15, 830-840. [CrossRef]
- Jwa, S. C., Ogawa, K., Kobayashi, M., Morisaki, N., Sago, H., and Fujiwara, T. (2016) Validation of a food-frequency questionnaire for assessing vitamin intake of Japanese women in early and late pregnancy with and without nausea and vomiting. J Nutr Sci 5, e27. [CrossRef]
- Kao, P. C., and Heser, D. W. (1984) Simultaneous determination of 25-hydroxy- and 1,25-dihydroxyvitamin D from a single sample by dual-cartridge extraction. Clin Chem 30, 56-61. [CrossRef]
- Kasuga, Y., Kawai, T., Miyakoshi, K., Hori, A., Tamagawa, M., Hasegawa, K., Ikenoue, S., Ochiai, D., Saisho, Y., Hida, M., Tanaka, M., and Hata, K. (2022) DNA methylation analysis of cord blood samples in neonates born to gestational diabetes mothers diagnosed before 24 gestational weeks. BMJ Open Diabetes Res Care 10. [CrossRef]
- Kasuga, Y., Kawai, T., Miyakoshi, K., Saisho, Y., Tamagawa, M., Hasegawa, K., Ikenoue, S., Ochiai, D., Hida, M., Tanaka, M., and Hata, K. (2021) Epigenetic Changes in Neonates Born to Mothers With Gestational Diabetes Mellitus May Be Associated With Neonatal Hypoglycaemia. Front Endocrinol (Lausanne) 12, 690648. [CrossRef]
- Xu, Z., Niu, L., Li, L., and Taylor, J. A. (2016) ENmix: a novel background correction method for Illumina HumanMethylation450 BeadChip. Nucleic Acids Res 44, e20. [CrossRef]
- Pelegí-Sisó, D., de Prado, P., Ronkainen, J., Bustamante, M., and González, J. R. (2021) methylclock: a Bioconductor package to estimate DNA methylation age. Bioinformatics 37, 1759-1760. [CrossRef]
- Simpkin, A. J., Suderman, M., and Howe, L. D. (2017) Epigenetic clocks for gestational age: statistical and study design considerations. Clin Epigenetics 9, 100. [CrossRef]
- McGrath, J. J., Saha, S., Burne, T. H., and Eyles, D. W. (2010) A systematic review of the association between common single nucleotide polymorphisms and 25-hydroxyvitamin D concentrations. J Steroid Biochem Mol Biol 121, 471-477. [CrossRef]
- Baca, K. M., Govil, M., Zmuda, J. M., Simhan, H. N., Marazita, M. L., and Bodnar, L. M. (2018) Vitamin D metabolic loci and vitamin D status in Black and White pregnant women. Eur J Obstet Gynecol Reprod Biol 220, 61-68. [CrossRef]
- Hajhashemy, Z., Shahdadian, F., Ziaei, R., and Saneei, P. (2021) Serum vitamin D levels in relation to abdominal obesity: A systematic review and dose-response meta-analysis of epidemiologic studies. Obes Rev 22, e13134. [CrossRef]
- Nakajima, H., Sakamoto, Y., Honda, Y., Sasaki, T., Igeta, Y., Ogishima, D., Matsuoka, S., Kim, S. G., Ishijima, M., and Miyagawa, K. (2023) Estimation of the vitamin D (VD) status of pregnant Japanese women based on food intake and VD synthesis by solar UV-B radiation using a questionnaire and UV-B observations. J Steroid Biochem Mol Biol 229, 106272. [CrossRef]
- Yaskolka Meir, A., Wang, G., Hong, X., Hu, F. B., Wang, X., and Liang, L. (2024) Newborn DNA methylation age differentiates long-term weight trajectories: the Boston Birth Cohort. BMC Med 22, 373. [CrossRef]
- Simpkin, A. J., Howe, L. D., Tilling, K., Gaunt, T. R., Lyttleton, O., McArdle, W. L., Ring, S. M., Horvath, S., Smith, G. D., and Relton, C. L. (2017) The epigenetic clock and physical development during childhood and adolescence: longitudinal analysis from a UK birth cohort. Int J Epidemiol 46, 549-558. [CrossRef]
- Hu, J., Yaskolka Meir, A., Hong, X., Wang, G., Hu, F. B., Wang, X., and Liang, L. (2024) Epigenetic Clock at Birth and Childhood Blood Pressure Trajectory: A Prospective Birth Cohort Study. Hypertension 81, e113-e124. [CrossRef]
- Yang, W. C., Chitale, R., O’Callaghan, K. M., Sudfeld, C. R., and Smith, E. R. (2024) The Effects of Vitamin D Supplementation During Pregnancy on Maternal, Neonatal, and Infant Health: A Systematic Review and Meta-analysis. Nutr Rev.
- Monasso, G. S., Küpers, L. K., Jaddoe, V. W. V., Heil, S. G., and Felix, J. F. (2021) Associations of circulating folate, vitamin B12 and homocysteine concentrations in early pregnancy and cord blood with epigenetic gestational age: the Generation R Study. Clin Epigenetics 13, 95. [CrossRef]
- Gilfix, B. M. (2005) Vitamin B12 and homocysteine. Cmaj 173, 1360.
- Monasso, G. S., Voortman, T., and Felix, J. F. (2022) Maternal plasma fatty acid patterns in mid-pregnancy and offspring epigenetic gestational age at birth. Epigenetics 17, 1562-1572. [CrossRef]
- Liu, T., Zhong, S., Liu, L., Liu, S., Li, X., Zhou, T., and Zhang, J. (2015) Vitamin D deficiency and the risk of anemia: a meta-analysis of observational studies. Ren Fail 37, 929-934. [CrossRef]
- Smith, E. M., Alvarez, J. A., Martin, G. S., Zughaier, S. M., Ziegler, T. R., and Tangpricha, V. (2015) Vitamin D deficiency is associated with anaemia among African Americans in a US cohort. Br J Nutr 113, 1732-1740. [CrossRef]
- Bacchetta, J., Zaritsky, J. J., Sea, J. L., Chun, R. F., Lisse, T. S., Zavala, K., Nayak, A., Wesseling-Perry, K., Westerman, M., Hollis, B. W., Salusky, I. B., and Hewison, M. (2014) Suppression of iron-regulatory hepcidin by vitamin D. J Am Soc Nephrol 25, 564-572. [CrossRef]
- Zughaier, S. M., Alvarez, J. A., Sloan, J. H., Konrad, R. J., and Tangpricha, V. (2014) The role of vitamin D in regulating the iron-hepcidin-ferroportin axis in monocytes. J Clin Transl Endocrinol 1, 19-25. [CrossRef]
- Smith, E. M., Alvarez, J. A., Kearns, M. D., Hao, L., Sloan, J. H., Konrad, R. J., Ziegler, T. R., Zughaier, S. M., and Tangpricha, V. (2017) High-dose vitamin D(3) reduces circulating hepcidin concentrations: A pilot, randomized, double-blind, placebo-controlled trial in healthy adults. Clin Nutr 36, 980-985.
- Braithwaite, V. S., Crozier, S. R., D’Angelo, S., Prentice, A., Cooper, C., Harvey, N. C., and Jones, K. S. (2019) The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom. Nutrients 11.
- Michalski, E. S., Nguyen, P. H., Gonzalez-Casanova, I., Nguyen, S. V., Martorell, R., Tangpricha, V., and Ramakrishnan, U. (2017) Serum 25-hydroxyvitamin D but not dietary vitamin D intake is associated with hemoglobin in women of reproductive age in rural northern Vietnam. J Clin Transl Endocrinol 8, 41-48. [CrossRef]
- Si, S., Peng, Z., Cheng, H., Zhuang, Y., Chi, P., Alifu, X., Zhou, H., Mo, M., and Yu, Y. (2022) Association of Vitamin D in Different Trimester with Hemoglobin during Pregnancy. Nutrients 14. [CrossRef]
- Thomas, C. E., Guillet, R., Queenan, R. A., Cooper, E. M., Kent, T. R., Pressman, E. K., Vermeylen, F. M., Roberson, M. S., and O’Brien, K. O. (2015) Vitamin D status is inversely associated with anemia and serum erythropoietin during pregnancy. Am J Clin Nutr 102, 1088-1095. [CrossRef]
- Girchenko, P., Lahti, J., Czamara, D., Knight, A. K., Jones, M. J., Suarez, A., Hämäläinen, E., Kajantie, E., Laivuori, H., Villa, P. M., Reynolds, R. M., Kobor, M. S., Smith, A. K., Binder, E. B., and Räikkönen, K. (2017) Associations between maternal risk factors of adverse pregnancy and birth outcomes and the offspring epigenetic clock of gestational age at birth. Clin Epigenetics 9, 49. [CrossRef]
- Daredia, S., Huen, K., Van Der Laan, L., Collender, P. A., Nwanaji-Enwerem, J. C., Harley, K., Deardorff, J., Eskenazi, B., Holland, N., and Cardenas, A. (2022) Prenatal and birth associations of epigenetic gestational age acceleration in the Center for the Health Assessment of Mothers and Children of Salinas (CHAMACOS) cohort. Epigenetics 17, 2006-2021. [CrossRef]
- Beck, C., Blue, N. R., Silver, R. M., Na, M., Grobman, W. A., Steller, J., Parry, S., Scifres, C., and Gernand, A. D. (2024) Maternal vitamin D status, fetal growth patterns, and adverse pregnancy outcomes in a multisite prospective pregnancy cohort. Am J Clin Nutr. [CrossRef]
- Mosavat, M., Arabiat, D., Smyth, A., Newnham, J., and Whitehead, L. (2021) Second-trimester maternal serum vitamin D and pregnancy outcome: The Western Australian Raine cohort study. Diabetes Res Clin Pract 175, 108779. [CrossRef]
Figure 1.
Maternal blood 25(OH)D levels in the study population.
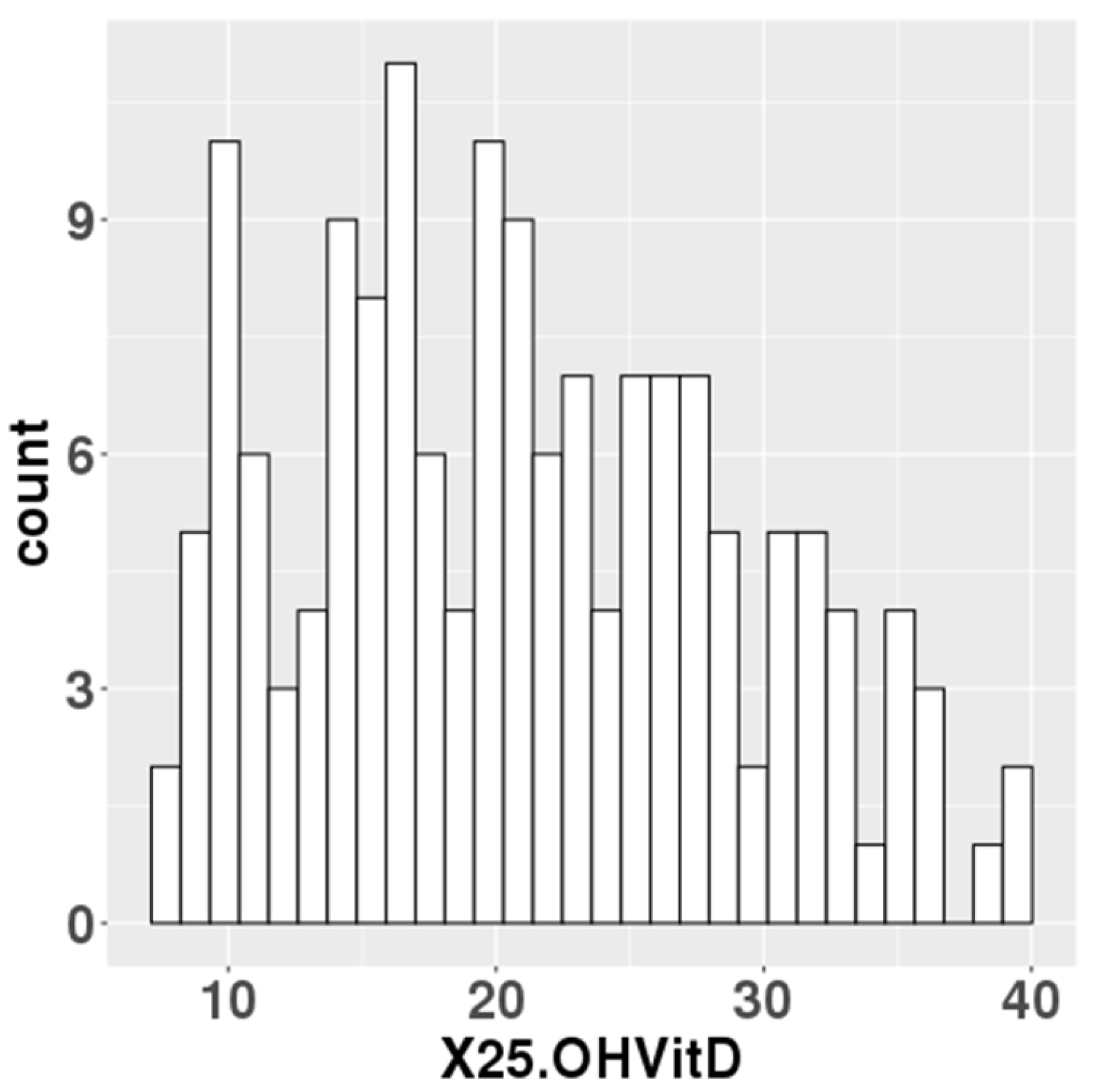
Figure 2.
Correlations between chronological gestational age and DNAmGAs.
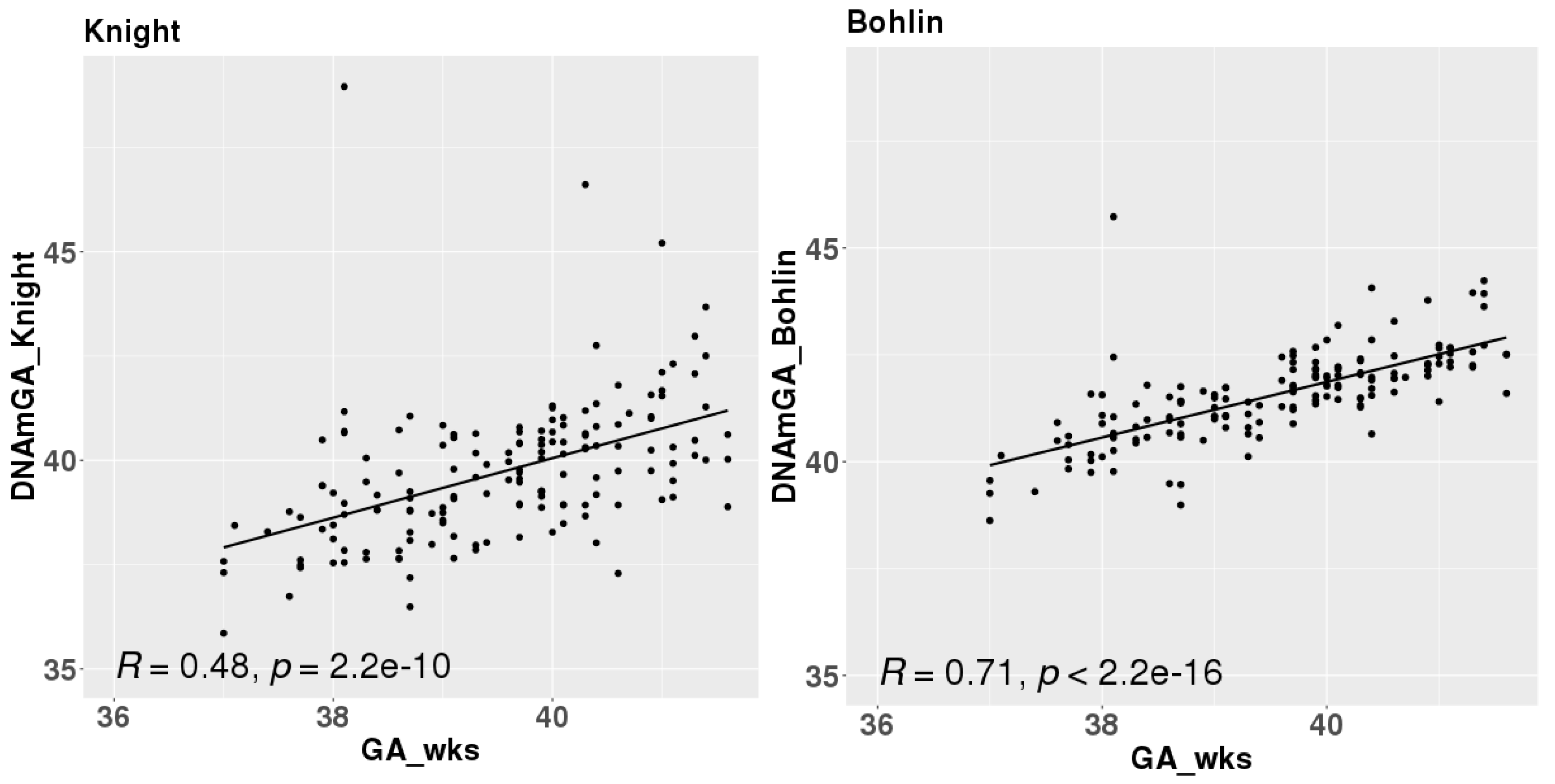
Figure 3.
Correlations between age acceleration estimated by Bohlin’s method and maternal 25(OH)D levels (a), birth height (b), and maternal age at delivery (c) adjusted for infants’ sex.
Figure 3.
Correlations between age acceleration estimated by Bohlin’s method and maternal 25(OH)D levels (a), birth height (b), and maternal age at delivery (c) adjusted for infants’ sex.
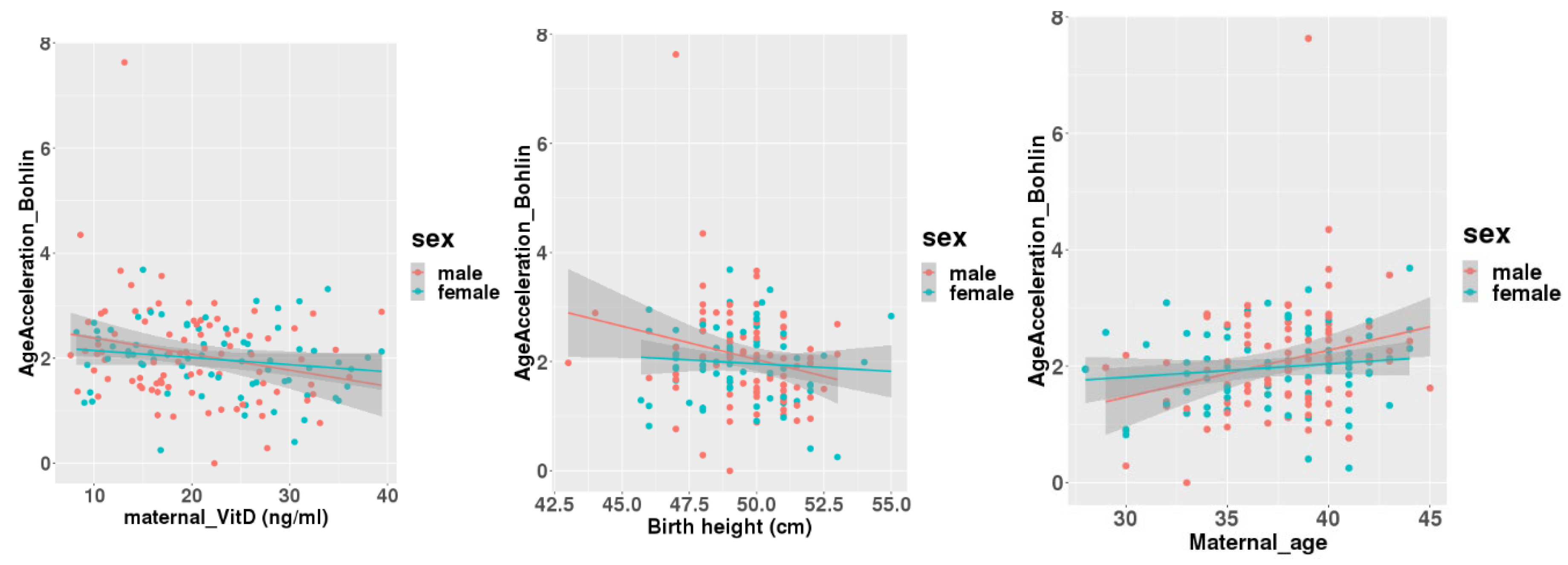

Table 1.
General characteristics of the participants (n = 157).
| Parental Characteristics | |
| Age at delivery (years) | 37.5 ± 3.7 |
| Paternal age at newborn’s birth (years) | 38.9 ± 8.2 (missing n = 15) |
| Pre-pregnant BMI (kg/m2) | 19.97 ± 2.48 |
| Gestational weight gain (kg) | 10.64 ± 4.98 |
| Maternal blood 25(OH)D (ng/ml) | 21.02 ± 7.87 |
| Newborn’s Characteristics | |
| Birth weight (g) | 3054 ± 383 |
| Birth height (cm) | 49.5 ± 1.9 |
| Gestational age at birth (weeks) | 39.52 ± 1.15 |
| Birth weight SD score | 0.10 ± 1.02 |
| Birth height SD score | 0.26 ± 0.92 |
| Cord blood 25(OH)D (ng/ml) | 13.15 ± 4.39 (missing n = 3) |
Table 2.
Associations of characteristics with AgeAcceleration of DNAmGA.
| AgeAcceleration_Bohlin | AgeAcceleration_Knight | |||
| Coefficiency (95% CI) | P value | Coefficiency (95% CI) | P value | |
| Parental Characteristics | ||||
| Age at delivery (years) | 0.049 (0.013, 0.085) | 0.009* | 0.016 (-0.051, 0.083) | 0.635 |
| Paternal age at newborn’s birth (years) | 0.003 (-0.015, 0.021) | 0.748 | -0.002 (-0.034, 0.030) | 0.917 |
| Pre-pregnant BMI (kg/m2) | -0.005 (-0.059, 0.049) | 0.856 | 0.021 (-0.078, 0.119) | 0.676 |
| Gestational weight gain (kg) | 0.002 (-0.026, 0.029) | 0.913 | 0.021 (-0.028, 0.070) | 0.398 |
| Maternal blood 25(OH)D (ng/ml) | -0.022 (-0.039, -0.005) | 0.010* | -0.015 (-0.046, 0.016) | 0.335 |
| Newborn’s Characteristics | ||||
| Birth weight (g) | -0.0002 (-0.0006, 0.0001) | 0.192 | -0.0003 (-0.0009, 0.0004) | 0.411 |
| Birth height (cm) | -0.071 (-0.142, -0.005) | 0.048* | -0.105 (-0.234, 0.024) | 0.109 |
| Birth weight SD score | 0.069 (-0.062, 0.200) | 0.302 | 0.044 (-0.195, 0.284) | 0.715 |
| Birth height SD score | 0.020 (-0.128, 0.167) | 0.792 | -0.081 (-0.348, 0.186) | 0.551 |
| Cord blood 25(OH)D (ng/ml) | -0.016 (-0.047, 0.015) | 0.299 | -0.011 (-0.068, 0.045) | 0.695 |
Table 3.
Associations of characteristics with AgeAcceleration of DNAmGA adjusted for infants sex.
| AgeAcceleration_Bohlin | AgeAcceleration_Knight | ||||
| Coefficiency (95% CI) | P value | Coefficiency (95% CI) | P value | ||
| Parental Characteristics | |||||
| Age at delivery (years) | 0.048 (0.012, 0.085) | 0.009* | 0.016 (-0.050, 0.084) | 0.630 | |
| Paternal age at newborn’s birth (years) | 0.002 (-0.016, 0.020) | 0.852 | -0.001 (-0.034, 0.031) | 0.928 | |
| Pre-pregnant BMI (kg/m2) | -0.004 (-0.0058, 0.051) | 0.895 | 0.019 (-0.080, 0.119) | 0.698 | |
| Gestational weight gain (kg) | 0.002 (-0.025, 0.029) | 0.899 | 0.021 (-0.028, 0.070) | 0.405 | |
| Maternal blood 25(OH)D (ng/ml) | -0.022 (-0.039, -0.005) | 0.013* | -0.017 (-0.048, 0.015) | 0.299 | |
| Newborn’s Characteristics | |||||
| Birth weight (g) | -0.0002 (-0.0006, 0.0001) | 0.170 | -0.0003 (-0.0009, 0.0004) | 0.433 | |
| Birth height (cm) | -0.075 (-0.146, -0.003) | 0.040* | -0.103 (-0.233, 0.027) | 0.118 | |
| Birth weight SD score | 0.072 (-0.060, 0.204) | 0.285 | 0.042 (-0.199, 0.282) | 0.733 | |
| Birth height SD score | 0.014 (-0.135, 0.162) | 0.858 | -0.075 (-0.344, 0.195) | 0.585 | |
| Cord blood 25(OH)D (ng/ml) | -0.019 (-0.050, 0.013) | 0.238 | -0.010 (-0.067, 0.048) | 0.740 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



